Embed Size (px)
Citation preview

DEVELOPMENTAL TOXICOLOGY
Christian FoÈ rster á Marcus RuÈ cker á Mehdi ShakibaeiIrmela Baumann-Wilschke á JuÈ rgen VormannRalf Stahlmann
Effects of ¯uoroquinolones and magnesium de®ciencyin murine limb bud cultures
Received: 7 October 1997 /Accepted: 25 February 1998
Abstract Quinolone-induced arthropathy is probablycaused by a lack of functionally available magnesium inimmature joint cartilage. We used an in vitro assay tostudy the e�ects of ¯uoroquinolones on cartilage for-mation in mouse limb buds from 12-day-old mouseembryos in regular and in magnesium-de®cient medium.Omission of magnesium from the medium had no ad-verse e�ect on the outcome of the culture: limb budsgrew and di�erentiated well in regular and in magne-sium-de®cient Bigger's medium. Lack of calcium, how-ever, severely impaired the development of the explants;this result was even more enhanced when both minerals(magnesium and calcium) were omitted. Electron mi-croscopy revealed cell necrosis and deposition of elec-tron-dense material in the vicinity of chondrocytes fromlimb buds after 6 days in a magnesium-free medium. Aseries of seven ¯uoroquinolones was tested at 30, 60, and100 mg/l medium. At a concentration of 30 mg/l spar-¯oxacin only had a slight e�ect on limb development. Atconcentrations of 60 and 100 mg/l spar¯oxacin, tema-¯oxacin and cipro¯oxacin impaired limb developmentin vitro concentration-dependently. The e�ects wereenhanced in a magnesium-de®cient medium (concen-tration of magnesium <10 lmol/l). Fleroxacin, lo-me¯oxacin and o¯oxacin impaired limb developmentonly slightly; no signi®cant di�erences were recognizablebetween the outcome in regular and in magnesium-de-®cient medium. Pe¯oxacin did not show any e�ect on
limb development in both media. Using electron mi-croscopy, very similar alterations as described above forthe limbs cultured in magnesium-de®cient medium wereobserved with o¯oxacin at a concentration of 30 mg/l,which had no e�ect on the growth of the explants whenevaluated macroscopically. The a�nity of six ¯uoro-quinolones to magnesium was determined by the use of a¯uorescence assay. The a�nity to magnesium correlatedwith the activity of the drugs in the limb bud assay. Weconclude that ¯uoroquinolones have no e�ect on murinelimb development in vitro at concentrations that areachieved under therapeutic conditions (peak concentra-tions approx. 1±5 mg/l in plasma). E�ects at higherconcentrations (60 and 100 mg/l) are slightly enhanced(factor 2) if the magnesium concentration in the mediumis low. Macroscopically, limbs develop regularly in amagnesium-free medium, but ultrastructurally typicalalterations are exhibited (e.g. cell necrosis and pericel-lular deposition of electron-dense material).
Key words Fluoroquinolones á Chondrotoxicity áMagnesium á Calcium á Limb bud culture
Introduction
Fluoroquinolones are contraindicated in paediatric pa-tients because all compounds tested to date induce jointcartilage lesions in juvenile animals such as dogs (Ing-ham et al. 1977; Tatsumi et al. 1978; Burkhardt et al.1990), rats (Kato and Onodera 1988 a, b), non-humanprimates (Stahlmann et al. 1990) and others (for reviewsee Gough et al. 1992; Stahlmann et al. 1993a). Thechondrotoxic potential of the drugs is probably relatedto their ability to form stable chelate complexes withmagnesium ions. We showed that by feeding a magne-sium-de®cient diet, joint cartilage lesions could be in-duced in juvenile rats which were indistinguishable byhistology, electron microscopy and immunohistochem-istry from cartilage lesions observed after quinolonetreatment. From these results it was concluded that the
Arch Toxicol (1998) 72: 411±419 Ó Springer-Verlag 1998
C. FoÈ rster á M. RuÈ cker á I. Baumann-Wilschke áR. Stahlmann (&)Institut fuÈ r Klinische Pharmakologie und Toxikologie,Freie UniversitaÈ t Berlin, Garystrasse 5,D-14195 Berlin, Germany
M. ShakibaeiInstitut fuÈ r Anatomie, Freie UniversitaÈ t Berlin,KoÈ nigin-Luise-Strasse 15, D-14195 Berlin, Germany
J. VormannInstitut fuÈ r Molekularbiologie und Biochemie,Freie UniversitaÈ t Berlin, Arnimallee 22,D-14195 Berlin, Germany

initial step in quinolone-induced arthropathy is a dis-turbance of the electrolyte balance in joint cartilage bychelate formation with magnesium ions (Stahlmann et al.1995; FoÈ rster et al. 1996a, b, c; Shakibaei et al. 1996;Vormann et al. 1997).
The experiments presented in this paper were designedto investigate the e�ects of a lack of magnesium alone orin combination with a series of seven ¯uoroquinoloneson cartilaginous tissue. We used limb buds preparedfrom 12-day-old mouse embryos for our studies. Thisassay has been used before in our laboratory to charac-terize the e�ects of o¯oxacin and cipro¯oxacin in com-parison to the non-¯uorinated derivative nalidixic acid.E�ects were only seen at concentrations which are con-siderably higher than those obtained under therapeuticconditions (Stahlmann et al. 1988). Our hypothesis nowwas that, similar to the in vivo ®ndings in juvenile rats,the rather small e�ects observed earlier might be signi®-cantly enhanced in a medium with low magnesium con-centration and that a lack of magnesium alone mightseverely impair limb bud growth in vitro. In addition, wemeasured the a�nity of the ¯uoroquinolones to divalentcations (magnesium and calcium) and compared the re-sults with the ®ndings of the limb bud assay.
Materials and methods
Animal keeping
NMRI mice were kept in Macrolon cages at room temperature(23 � 1 °C) under speci®c pathogen free (spf) conditions (lights onfrom 9.00 a.m. to 9.00 p.m., relative humidity, 50%). Altrominpellet food (Altromin 1324, Lage, Germany) and tap water weregiven ad libitum. For mating the animals were caged together for aperiod of 2±3 h. Day 0 of pregnancy was the 24 h period followingthe detection of vaginal plugs.
Fluoroquinolones
Fluoroquinolones used in limb bud cultures
O¯oxacin (Tarivid i.v. 200; Hoechst, Frankfurt/Main, Germany)and cipro¯oxacin (Ciprobay 200, Bayer, Leverkusen, Germany)were added to the culture medium by using the commerciallyavailable i.v. preparations. Lome¯oxacin (Searle, Neu Isenburg,Germany), ¯eroxacin (Roche, Grenzach-Wyhlen, Germany), te-ma¯oxacin (Abott, Queensborough, UK), pe¯oxacin and spar-¯oxacin (Rhoà ne-Poulenc, KoÈ ln, Germany) were generous giftsfrom the pharmaceutical companies.
A�nity of ¯uoroquinolones to magnesium and calcium(¯uoroscence assay; cf below)
O¯oxacin and cipro¯oxacin (pure compounds) were generous giftsfrom the pharmaceutical companies (Hoechst AG, Frankfurt,Germany and Bayer AG, Leverkusen, Germany). The ¯uorescenceassay was performed using the compounds described above.
Limb bud culture
Limb buds from mouse embryos were cultured as described indetail by Blankenburg (1981). Brie¯y, embryos (somite stage, 48±
52) were removed from the uterus of pregnant mice (day 12 ofgestation) and collected in sterile Hanks' balanced salt solution(HBSS; without calcium and magnesium). The forelimbs wereprepared under a dissection microscope and transferred into asterile 50 ml culture bottle, which was sealed airtight with asterile rubber stopper and a metal clamp. Magnesium-containingor magnesium-free culture medium (see below) supplementedwith 0, 30, 60 and 100 mg quinolone/l medium was added byinserting a sterile needle through the rubber stopper. The bottleswere subsequently ¯ushed with a gas mixture of 47% O2, 5%CO2 and 48% N2 and incubated at 37 °C on a roller device(25 rpm).
After 3 days, the medium was replaced by new medium withthe same concentrations of electrolytes and/or quinolones. Afteranother 3 days of culture, limbs were removed from the bottles,washed with phosphate-bu�ered saline (PBS), ®xed in formalinand stained with a solution of alcian blue in acetic acid over-night. The stained limbs were cleared in 30% acetic acid, dehy-drated in the alcohol series, cleared in xylol and kept under cedaroil.
Media for limb bud cultures
We used a serum-free medium which roughly corresponds to Big-ger's medium and has been described in detail elsewhere (Blan-kenburg 1981). The standard medium contained 1.66 mmol Mg2+
(MgSO4.7 H2O) and 3.63 mmol Ca2+ (C6H10CaO6.5 H2O; calcium
lactate). In calcium- and/or magnesium-free culture media, calciumlactate was replaced by sodium lactate and magnesium sulphatewas replaced by sodium sulphate or by sodium chloride (prelimi-nary experiments showed no di�erence of limb development withthese two kinds of magnesium-free media). The osmolarity of themedia was adjusted to 340 mosmol by addition of saline solution.For magnesium quanti®cation, culture medium samples werebrought to appropriate dilutions with 10% trichloroacetic acid/0.175% LaCl3 and measured by atomic absorption spectrophoto-metry (Philips SP9).
Electron microscopy
At the end of the 6-day culture period we investigated limbswhich had been grown in regular medium, limbs grown in mag-nesium-de®cient medium as well as limbs grown in regular me-dium with 30 mg o¯oxacin per litre. In both groups (Mg2+-de®cient and with o¯oxacin) no alterations of limb developmentwere recognizable by macroscopic evaluation. Limbs were ®xed in1% glutaraldehyde plus 1% tannic acid in 0.1 M phosphate bu�er(pH 7.4). These samples were post®xed in 2% OsO4 in 0.1 Mphosphate bu�er. After rinsing and dehydration in ethanol, thesamples were embedded in Epon (Plano, Marburg, Germany), cutwith an Ultracut E (Reichert), contrasted with 2% uranyl acetate/lead citrate and examined by transmission electron microscopy(Zeiss EM 10).
Fluorescence assay of quinolone-cation chelate complexes
The chelation of cations by ¯uoroquinolones was investigated witha ¯uorescence assay. Fluoroquinolones were dissolved in 50 mMTRIS acetate (pH 7.4) to give a concentration of 0.1 mg/l (spar-¯oxacin, 1.0 mg/l). The bu�er was supplemented with 0, 2, 5, 10 or20 mmol Mg2+ or Ca2+ per litre by addition of magnesium orcalcium chloride. The ¯uorescence intensity of each quinolone wasdetermined in the presence and in the absence of magnesium orcalcium ions at the appropriate excitation and emission wave-lengths (Table 1) using a Shimadzu RF 5000 spectro¯uorophoto-meter. The increase of the ¯uorescence intensity (arbitrary units) asobserved in the presence of divalent cations was expressed in termsof the percentage change compared to the ¯uorescence of the¯uoroquinolone alone.
412

Results
E�ects of magnesium or calcium de®ciencyon limb bud cultures
Figure 1A shows characteristic examples of normal limbbuds grown in the magnesium- and calcium-containingstandard medium. After a culture period of 6 days, thecartilaginous anlage has di�erentiated and scapula, hu-merus, ulna, radius and the hand skeleton can clearly bedistinguished. Pronounced impairment particularly ofthe di�erentiation of the hand skeleton was observed inthe calcium-de®cient (but magnesium-containing) cul-ture medium (Fig. 1B).
Surprisingly, growth and morphogenetic di�erentia-tion of the explants were not impaired when a magne-sium-free (but calcium-containing) culture medium wasused (Fig. 1C). The explants grew even slightly betterthan those limb buds that had been cultured in thepresence of both minerals. Magnesium concentrations in
the standard medium and in the magnesium-de®cientmedium were analysed by atomic absorption spectro-photometry at the end of the culture period. The meanconcentration (�SD) of 15 aliquots was 1.48 �0.05 mmol/l in the magnesium-containing standardmedium and <10 lmol/l in the magnesium-de®cientculture medium. Complete inhibition of growth wasobserved when the limb buds were grown in the absenceof both magnesium and calcium (Fig. 1D).
E�ects of ¯uoroquinolones on limb bud cultures
The e�ects of ¯uoroquinolones on limb bud develop-ment were subsequently studied in a culture mediumcontaining both calcium and magnesium (standard me-dium) and in a magnesium-de®cient medium. The e�ectsobserved with seven ¯uoroquinolones on limb budgrowth and morphogenetic di�erentiation are summa-rized in Table 2. An overall evaluation shows that themost pronounced e�ects on limb bud di�erentiationwere observed in explants cultured in the presence ofspar¯oxacin. However, at a concentration of 30 mg/lnone of the compounds tested had a pronounced e�ectwith both media used. Slight alterations at 30 mg/l wereonly seen with spar¯oxacin (both media) and with te-ma¯oxacin or cipro¯oxacin in magnesium-de®cient me-dium. At higher concentrations clear-cut e�ects on limbbud development were observed with spar¯oxacin, ci-pro¯oxacin and tema¯oxacin. These e�ects were morepronounced in limb buds grown in the magnesium-freeculture medium compared to explants cultured in thepresence of magnesium.
Characteristic examples of limb buds grown with 30,60 or 100 mg spar¯oxacin/l culture medium are shownin Figs. 2 and 3. In the standard culture medium(Fig. 2), limb bud development was slightly a�ected at
Table 1 Excitation (Exmax) and emission (Emmax) wavelengths ofseveral ¯uoroquinolones (0.1 mg/l) in 50 mmol TRIS-acetate,pH 7.4
Compound Exmax (nm) Emmax (nm)
O¯oxacin 296 453Cipro¯oxacin 276 419Nor¯oxacin 276 415Lome¯oxacin 281 423Tema¯oxacin 275 425Fleroxacin 280 420Pe¯oxacin 276 421Spar¯oxacina a a
a Spar¯oxacin did not ¯uoresce in the absence of Mg2+ or Ca2+
Fig. 1A±D E�ects of magne-sium and calcium de®ciency inlimb bud cultures (mouse, day12 of gestation). A Normal limbbud development in magne-sium- and calcium-containingstandard medium. B Calcium-de®cient (but magnesium-con-taining) medium: retardation ofgrowth, impaired di�erentiationof hand skeleton.CMagnesium-de®cient (but calcium-contain-ing) medium: limb-buddevelopment is not impairedcompared to explants grown inthe magnesium-containingstandard medium. D Magne-sium- and calcium-de®cientmedium: complete inhibition oflimb bud growth
413

concentrations of 30 mg spar¯oxacin/l (Fig. 2B). At 60and 100 mg/l, a clear-cut retardation of limb bud de-velopment was observed: decreased size of the explants,poorly developed scapula, shortening of humerus, ulnaand radius, and impaired di�erentiation of the handskeleton (Fig. 2C, D).
In the magnesium-de®cient culture medium (Fig. 3),the size of the scapula decreased slightly in limb budscultured in the presence of 30 mg spar¯oxacin, but noe�ects on the size of the explants and only a slight al-teration of the di�erentiation of the hand skeleton wereobserved. Limb bud growth was completely inhibited byspar¯oxacin at a concentration of ³60 mg/l. The e�ectwas comparable to those of tema¯oxacin and cipro-¯oxacin at 100 mg/l (Fig. 3, Table 2).
The e�ects of o¯oxacin, ¯eroxacin and lome¯oxacinon limb bud di�erentiation were less pronounced thanthe e�ects observed with the three ¯uoroquinolones
mentioned above. Development of the scapula was re-tarded in the magnesium-containing culture medium,but no e�ects on the di�erentiation of the hand skeletonor on the growth of humerus, ulna and radius wereobserved. No clear-cut di�erences were observed be-tween explants grown in magnesium-containing ormagnesium-free culture media. Pe¯oxacin did not in-terfere with limb bud development in magnesium-con-taining and magnesium-de®cient culture media(Table 2).
Electron microscopy
Cells from limb buds grown in magnesium-de®cientmedium showed characteristic ultrastructural alterations(Fig. 4). Many chondrocytes were necrotic and electron-dense material was deposited near the cells. A typical
Table 2 E�ects of ¯uoroquinolones on limb bud development.Limb buds were prepared from mouse embryos (day 12 of gesta-tion) and grown in magnesium-containing (+Mg2+) and magne-sium-de®cient ()Mg2+) culture media in the presence of the
indicated quinolone concentrations. (SPX Spar¯oxacin, TFX te-ma¯oxacin, CPX cipro¯oxacin, FLX ¯eroxacin, OFX o¯oxacin,LFX lome¯oxacin, PFX pe¯oxacin)
Medium E�ects of ¯uoroquinolones on limb bud cultures (mouse, day 12)
c (mg/l)a SPX TFX CPX FLX OFX LFX PFX
+Mg2+ 30 + ) ) n.d. ) n.d. n.d.60 ++ ++ ++ + ) + )100 +++ +++ ++ + + + )
)Mg2+ 30 + + + n.d. ) ) n.d.60 ++++ +++ ++ + n.d. + )100 ++++ ++++ ++++ + + + )
a Concentration of quinolones), No e�ect; +, minor impairment of limb bud development (re-tarded growth of the scapula, no e�ect on di�erentiation of thehand skeleton); ++, pronounced impairment of limb-bud di�er-
entiation (retarded growth of scapula and humerus, impaired dif-ferentiation of the hand skeleton; +++, pronounced reduction insize, essential parts of the limb buds are missing; ++++, com-plete inhibition of growth; n.d., not determined
Fig. 2A±D E�ects of spar-¯oxacin on limb buds (mouse,day 12 of gestation) cultured inthe magnesium-containing stan-dard culture medium. A Un-treated control. B 30 mgspar¯oxacin/l: slightly reducedsize of the scapula, no e�ects ondi�erentiation of the hand skel-eton. C 60 mg spar¯oxacin/l:pronounced retardation ofgrowth and di�erentiation.D 100 mg spar¯oxacin/l: pro-nounced retardation of growthand di�erentiation, essentialparts of the limbs (e.g. scapula)are missing
414

®nding was loosening and reduction of matrix withmatrix-free pericellular areas. A similar morphologicaloutcome was seen in limbs cultured in the presence ofo¯oxacin at 30 mg/l, a concentration that did not inducemacroscopically recognizable alterations.
A�nity of ¯uoroquinolones to magnesium and calcium
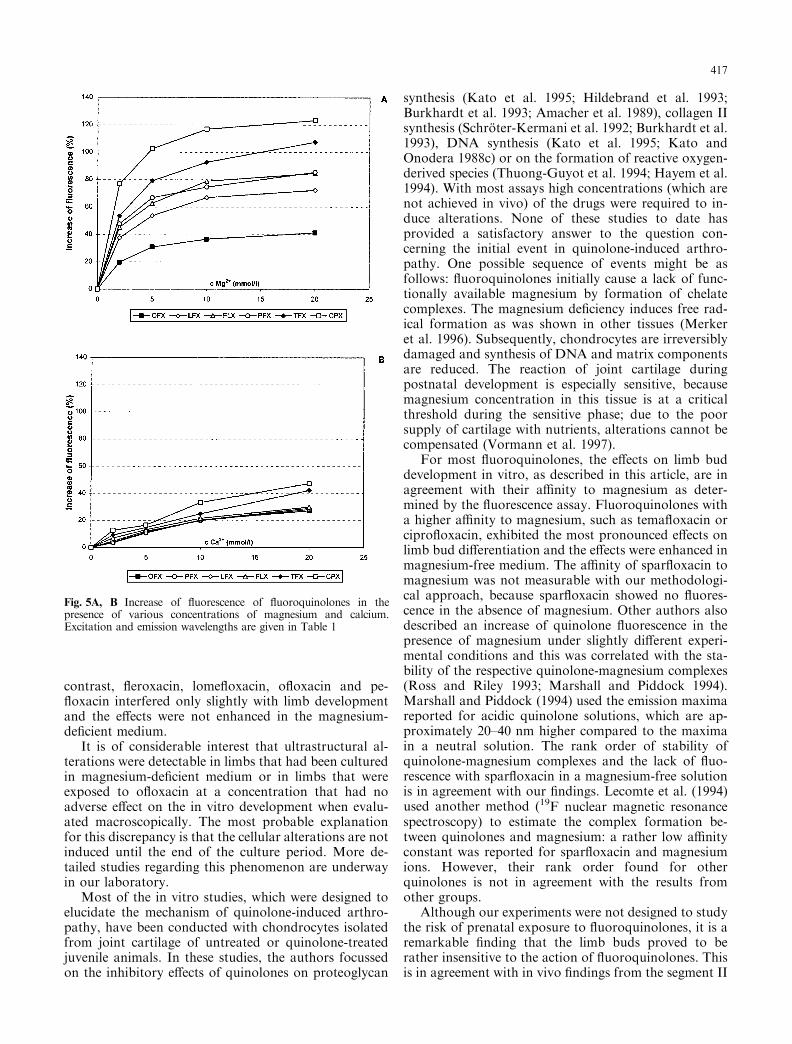
The ¯uorescence of quinolones was determined inmagnesium and calcium-free TRIS-bu�er (pH 7.4) andin bu�er supplemented with magnesium or with calcium.In magnesium-containing bu�er the ¯uorescence of thequinolones increased with increasing concentrations ofthe cation (a magnesium chloride solution did not ¯u-oresce at the excitation wavelengths indicated in Ta-ble 1). This e�ect was most pronounced withcipro¯oxacin and tema¯oxacin (+123 and +108% ofthe baseline ¯uorescence in the presence of 20 mmolMg2+) and less pronounced with pe¯oxacin (+86%),¯eroxacin (+85%), lome¯oxacin (+72%) and o¯oxacin(+41%). The a�nity of spar¯oxacin to magnesiumcould not be tested with this method because spar-¯oxacin showed no ¯uorescence in the absence of mag-nesium.
The ¯uorescence of quinolone-calcium complexeswas lower than the respective magnesium complexes.Cipro¯oxacin and tema¯oxacin had the highest a�nityto Ca2+ as re¯ected by the increase of ¯uorescence in thepresence of 20 mmol Ca2+/l TRIS-bu�er (+47 and+42%, respectively). No signi®cant di�erences wereobserved in the a�nity of o¯oxacin, pe¯oxacin, lo-me¯oxacin and ¯eroxacin to Ca2+ (+27, +28, +29 and+30% of the baseline ¯uorescence in the presence of20 mmol Ca2+/l, TRIS-bu�er). The results of the ¯uo-rescence assay are summarized in Fig. 5.
Discussion
In contrast to our ®ndings with joint cartilage from ju-venile rats (Stahlmann et al. 1995), magnesium de®-ciency had no adverse e�ect on the growth anddi�erentiation of limb buds from 12-day-old mouseembryos although the magnesium concentration in themedium was extremely low (<10 lmol/l). This di�erenceto the in vivo results with joint cartilage might be relatedto the fact that the limb buds were dissected in toto andthe magnesium content of the explants might have beensu�cient to sustain limb bud growth during the 6 daysof culture in a magnesium-de®cient medium.
Growth and di�erentiation of the explants were im-paired in the calcium-de®cient (but magnesium-con-taining) medium. Interestingly, by using a medium thatwas de®cient in both cations (magnesium and calcium)complete inhibition of growth was induced (cf. Fig. 1D),whereas magnesium de®ciency had no negative e�ect onlimb development in calcium-containing culture medium(Fig. 1C). These results suggest that calcium might atleast partly replace magnesium ions but that magnesium,on the other hand, cannot replace calcium in this system.Magnesium and calcium are bound to proteoglycans inthe extracellular matrix and the distribution of the cat-ions is in¯uenced by a Donnan e�ect. A�nity forchondroitin sulphate is higher for calcium than formagnesium (GuÈ nther et al. 1997).
Limb bud cultures have been used in a large numberof studies to investigate the in¯uences of xenobiotics onthe morphogenetic di�erentiation of cartilaginousstructures in vitro (for review see Neubert 1982, 1985;Stahlmann et al. 1993b). To our knowledge, however,no data on the in¯uence of the magnesium or calciumconcentrations on limb bud development have beenpublished before. Some of the ¯uoroquinolones used in
Fig. 3A±D E�ects of spar-¯oxacin on limb buds (mouse,day 12 of gestation) cultured inthe magnesium-de®cient culturemedium. A Untreated control.B 30 mg spar¯oxacin/l medium:slightly reduced size of thescapula. C 60 mg spar¯oxacin/lmedium: complete inhibition ofgrowth. D 100 mg spar¯oxacin/l medium: complete inhibitionof growth
415

this study have been investigated previously with thelimb bud assay in our laboratory: reproducibility ofthe e�ects was good. Results with o¯oxacin and ci-pro¯oxacin observed in regular medium are in agree-ment with the results observed earlier (Stahlmann et al.1988).
Clear-cut e�ects of the seven ¯uoroquinolones testedwere only seen at rather high concentrations (60 and100 mg/l), which are considerably higher than those
achieved under therapeutic conditions after oral ad-ministration of the drugs (1±5 mg/l). The e�ects ofspar¯oxacin, tema¯oxacin and cipro¯oxacin were morepronounced in magnesium-de®cient than in standardculture medium. These compounds clearly interferedwith limb bud growth and di�erentiation in magnesium-containing culture medium at concentrations of 60 mg/l,but at the same concentration limb bud growth was in-hibited completely in the absence of magnesium. In
Fig. 4a±c Electron micro-graphs of ultrathin sections oflimb buds from mouse fetuses(day 12). a Control; limbs werecultured in standard medium.Part of a typical chondrocyte(C ) is seen with numerouscavities of rough endoplasmicreticulum and a well developedmatrix (stars). b Cultured inmagnesium-de®cient mediumfor 6 days. Electron-dense ma-terial (arrows) is located on thesurface of the chondrocyte withloosening and reduction of thematrix. c Cultured in the pres-ence of 30 mg o¯oxacin per litremedium. Similar alterationsoccur as observed in limb budscultured in magnesium-de®cientmedium: electron densematerial in the vicinity of thechondrocyte and a reducedmatrix. a±c ´50 000
416
contrast, ¯eroxacin, lome¯oxacin, o¯oxacin and pe-¯oxacin interfered only slightly with limb developmentand the e�ects were not enhanced in the magnesium-de®cient medium.
It is of considerable interest that ultrastructural al-terations were detectable in limbs that had been culturedin magnesium-de®cient medium or in limbs that wereexposed to o¯oxacin at a concentration that had noadverse e�ect on the in vitro development when evalu-ated macroscopically. The most probable explanationfor this discrepancy is that the cellular alterations are notinduced until the end of the culture period. More de-tailed studies regarding this phenomenon are underwayin our laboratory.
Most of the in vitro studies, which were designed toelucidate the mechanism of quinolone-induced arthro-pathy, have been conducted with chondrocytes isolatedfrom joint cartilage of untreated or quinolone-treatedjuvenile animals. In these studies, the authors focussedon the inhibitory e�ects of quinolones on proteoglycan
synthesis (Kato et al. 1995; Hildebrand et al. 1993;Burkhardt et al. 1993; Amacher et al. 1989), collagen IIsynthesis (SchroÈ ter-Kermani et al. 1992; Burkhardt et al.1993), DNA synthesis (Kato et al. 1995; Kato andOnodera 1988c) or on the formation of reactive oxygen-derived species (Thuong-Guyot et al. 1994; Hayem et al.1994). With most assays high concentrations (which arenot achieved in vivo) of the drugs were required to in-duce alterations. None of these studies to date hasprovided a satisfactory answer to the question con-cerning the initial event in quinolone-induced arthro-pathy. One possible sequence of events might be asfollows: ¯uoroquinolones initially cause a lack of func-tionally available magnesium by formation of chelatecomplexes. The magnesium de®ciency induces free rad-ical formation as was shown in other tissues (Merkeret al. 1996). Subsequently, chondrocytes are irreversiblydamaged and synthesis of DNA and matrix componentsare reduced. The reaction of joint cartilage duringpostnatal development is especially sensitive, becausemagnesium concentration in this tissue is at a criticalthreshold during the sensitive phase; due to the poorsupply of cartilage with nutrients, alterations cannot becompensated (Vormann et al. 1997).
For most ¯uoroquinolones, the e�ects on limb buddevelopment in vitro, as described in this article, are inagreement with their a�nity to magnesium as deter-mined by the ¯uorescence assay. Fluoroquinolones witha higher a�nity to magnesium, such as tema¯oxacin orcipro¯oxacin, exhibited the most pronounced e�ects onlimb bud di�erentiation and the e�ects were enhanced inmagnesium-free medium. The a�nity of spar¯oxacin tomagnesium was not measurable with our methodologi-cal approach, because spar¯oxacin showed no ¯uores-cence in the absence of magnesium. Other authors alsodescribed an increase of quinolone ¯uorescence in thepresence of magnesium under slightly di�erent experi-mental conditions and this was correlated with the sta-bility of the respective quinolone-magnesium complexes(Ross and Riley 1993; Marshall and Piddock 1994).Marshall and Piddock (1994) used the emission maximareported for acidic quinolone solutions, which are ap-proximately 20±40 nm higher compared to the maximain a neutral solution. The rank order of stability ofquinolone-magnesium complexes and the lack of ¯uo-rescence with spar¯oxacin in a magnesium-free solutionis in agreement with our ®ndings. Lecomte et al. (1994)used another method (19F nuclear magnetic resonancespectroscopy) to estimate the complex formation be-tween quinolones and magnesium: a rather low a�nityconstant was reported for spar¯oxacin and magnesiumions. However, their rank order found for otherquinolones is not in agreement with the results fromother groups.
Although our experiments were not designed to studythe risk of prenatal exposure to ¯uoroquinolones, it is aremarkable ®nding that the limb buds proved to berather insensitive to the action of ¯uoroquinolones. Thisis in agreement with in vivo ®ndings from the segment II
Fig. 5A, B Increase of ¯uorescence of ¯uoroquinolones in thepresence of various concentrations of magnesium and calcium.Excitation and emission wavelengths are given in Table 1
417

type studies performed with ¯uoroquinolones: limb re-duction defects or other gross-structural defects have notbeen observed thus far in animals (for review seeStahlmann 1990; Takayama et al. 1995) and limitedhuman data also give no indication of a risk for limbdefects in association with ¯uoroquinolone use duringpregnancy (Berkovitch et al. 1994).
One possible explanation for the relative insensi-tivity of prenatally formed cartilaginous tissue is therather high supply of this tissue with magnesium. Inserum from rat fetuses magnesium concentrationswere signi®cantly higher than in serum from adult ratsshowing a constant decrease during prenatal devel-opment (GuÈ nther 1966; Vormann and GuÈ nther 1986;Husain and Sibley 1993). Also, due to the small sizeof the prenatally formed structures and the supplywith nutrients by amniotic ¯uid as well as by fetalblood, it seems to be reasonable to assume that al-teration in the concentration of functionally availablemagnesium by the chelating action of quinolones isreadily balanced.
Acknowledgements This study was supported by a grant fromDeutsche Forschungsgemeinschaft. Some of the data presented inthis paper are part of the doctoral thesis of Marcus RuÈ cker to besubmitted to the Fachbereich Humanmedizin, Freie UniversitaÈ tBerlin. The authors thank Mrs Barbara Steyn for her help in pre-paring the manuscript. The authors would like to dedicate thispaper to Prof Dr Hans-Joachim Merker and to Prof Dr TheodorGuÈ nther (Fachbereich Humanmedizin, Freie UniversitaÈ t Berlin)and thereby acknowledge the longstanding excellent cooperationwith both scientists. Their broad range of experience, continuousinterest in our work and many fruitful discussions have providedthe basis for a series of experiments which have been extraordi-narily successful in shedding new light on the chondrotoxicity ofquinolones.
References
Amacher DE, Schomaker SJ, Gootz TD, McGuirk PR (1989)Proteoglycan and procollagen synthesis in rat embryo limb budcultures treated with quinolone antibacterials. Alt MethodsToxicol 7: 307±312
Berkovitch M, Pastuszak A, Gazarian M, Lewis M, Koren G(1994) Safety of the new quinolones in pregnancy. ObstetGynecol 84: 535±538
Blankenburg G (1981) Limb bud organ cultures. In: Neubert D,Merker HJ (eds) Culture techniques. Walter de Gruyter, Berlin,pp 590±594
Burkhardt JE, Hill MA, Carlton WW, Kesterson JW (1990) His-tologic and histochemical changes in articular cartilage of im-mature beagle dogs dosed with di¯oxacin, a ¯uoroquinolone.Vet Pathol 27: 162±170
Burkhardt JE, Hill MA, Lamar CH, Smith GN, Carlton WW(1993) E�ects of di¯oxacin on the metabolism of glycosami-noglycans and collagen in organ cultures of articular cartilage.Fundam Appl Toxicol 20: 257±263
FoÈ rster C, Kociok K, Shakibaei M, Merker H-J, Vormann J,GuÈ nther T, Stahlmann R (1996a) Integrins on joint cartilagechondrocytes and alterations by o¯oxacin or magnesium de®-ciency in immature rats. Arch Toxicol 70: 261±270
FoÈ rster C, Kociok K, Shakibaei M, Merker HJ, Stahlmann R(1996b) Quinolone-induced cartilage lesions are not reversiblein rats. Arch Toxicol 70: 474±481
FoÈ rster C, Zippel U, Lozo E, Vormann J, Merker HJ, StahlmannR (1996c) E�ects of quinolones on joint cartilage in juvenile ratsand other animals: focus on mechanistic aspects. Chemother J5 (Suppl 13): 6±13
Gough AW, Kasali OB, Siegler RE, Baragi V (1992) Quinolonearthropathy ± acute toxicity to immature cartilage. ToxicolPathol 20: 436±450
GuÈ nther T (1966) Die embryonale Entwicklung des aktiven Mag-nesium-Transportes. Naturwissenschaften 53: 554±555
GuÈ nther T, RuÈ cker M, FoÈ rster C, Vormann J, Stahlmann R (1997)In vitro evidence for a Donnan distribution of Mg2+ and Ca2+
by chondroitin sulphate in cartilage. Arch Toxicol 71: 471±475Hayem G, Petit PX, Levacher M, Gaudin C, Kahn MF, Pocidalo
JJ (1994) Cyto¯uorometric analysis of chondrotoxicity of¯uoroquinolone antimicrobial agents. Antimicrob AgentsChemother 38: 243±247
Hildebrand H, Kempka G, SchluÈ ter G, Schmidt M (1993)Chondrotoxicity of quinolones in vivo and in vitro. ArchToxicol 67: 411±415
Husain SM, Sibley CP (1993) Magnesium and pregnancy. MinerElectrolyte Metab 19: 296±307
Ingham B, Brentnall DW, Dale EA, MacFadzean JA (1977)Arthropathy induced by antibacterial fused N-alkyl-3-pyri-done-carboxylic acids. Toxicol Lett 1: 21±26
Kato M, Onodera T (1988a) Morphological investigation of cavityformation in articular cartilage induced by o¯oxacin in rats.Fundam Appl Toxicol 11: 110±119
Kato M, Onodera T (1988b) Morphological investigation ofosteochondrosis induced by o¯oxacin in rats. Fundam ApplToxicol 11: 120±131
Kato M, Onodera T (1988c) E�ect of o¯oxacin on the uptake of[3H] thymidine by articular cartilage cells in the rat. ToxicolLett 44: 131±142
Kato M, Takada S, Ogawara S, Takayama S (1995) E�ect of lev-o¯oxacin on glycosaminoglycan and DNA synthesis of culturedrabbit chondrocytes at concentrations inducing cartilage lesionsin vivo. Antimicrob Agents Chemother 39: 1979±1983
Lecomte S, Baron MH, Chenon MT, Coupry C, Moreau NJ (1994)E�ect of magnesium complexation by ¯uoroquinolones on thierantibacterial properties. Antimicrob Agents Chemother 38:2810±2816
Marshall AJH, Piddock LJV (1994) Interaction of divalent cations,quinolones and bacteria. J Antimicrob Chemother 34: 465±483
Merker HJ, GuÈ nther T, HoÈ llriegl V, Vormann J, SchuÈ mann K(1996) Lipid peroxidation and morphology of rat testis inmagnesium de®ciency. Andrologia 28: 43±51
Neubert D (1982) The use of culture techniques in studies onprenatal toxicity. Pharmacol Ther 18: 397±434
Neubert D (1985) Toxicity studies with cellular models of di�er-entiation. Xenobiotica 15: 649±660
Ross D, Riley R (1993) Physicochemical properties of the ¯uoro-quinolone antimicrobials. V. E�ect of ¯uoroquinolone struc-ture and pH on the complexation of various ¯uoroquinoloneswith magnesium and calcium ions. Int J Pharmacol 93: 121±129
SchroÈ ter-Kermani C, Hinz N, Risse P, Stahlmann R, Merker HJ(1992) E�ects of o¯oxacin on chondrogenesis in murine carti-lage organoid culture. Toxicol In Vitro 6: 465±474
Shakibaei M, Merker HJ, Kociok K, FoÈ rster C, Vormann J,GuÈ nther T, Stahlmann R (1996) Comparative evaluation ofultrastructural changes in articular cartilage of juvenile ratsafter treatment with o¯oxacin and in magnesium-de®cient rats.Toxicol Pathol 24: 580±587
Stahlmann R (1990) Safety pro®le of the quinolones. J AntimicrobChemother 26 (Suppl D): 31±44
Stahlmann R, Blankenburg G, Neubert D (1988) Studies on car-tilage formation and di�erentiation in limb-bud culture in thepresence of nalidixic acid, o¯oxacin and cipro¯oxacin. RevInfect Dis 10 (Suppl 1): S147
Stahlmann R, Merker HJ, Hinz N, Webb J, Heger W, Neubert D(1990) O¯oxacin in juvenile non-human primates and rats.Arthropathia and drug plasma concentrations. Arch Toxicol64: 193±204
418

Stahlmann R, FoÈ rster C, van Sickle D (1993a) Quinolones inchildren. Are concerns over arthropathy justi®ed? Drug Safety9: 397±403
Stahlmann R, Klug S, Foerster M, Neubert D (1993b) Signi®canceof embryo culture methods for studying the prenatal toxicity ofvirustatic agents. Reprod Toxicol 7: 129±143
Stahlmann R, FoÈ rster C, Shakibaei M, Vormann J, GuÈ nther T,Merker HJ (1995) Magnesium de®ciency induces joint cartilagelesions in juvenile rats which are identical with quinolone-in-duced arthropathy. Antimicrob Agents Chemother 39: 2013±2018
Takayama S, Hirohashi M, Kato M, Shimada H (1995) Toxicity ofquinolone antimicrobial agents. J Toxicol Environ Health 45:1±45
Tatsumi H, Senda H, Yatera S, Takemoto Y, Yamayoshi M,Ohnishi K (1978) Toxicological studies on pipemidic acid V.
E�ect on diarthrodial joints of experimental animals. J ToxicolSci 3: 357±367
Thuong-Guyot M, Domarle O, Pocidalo JJ, Hayem G (1994) Ef-fects of ¯uoroquinolones on cultured articular chondrocytes.Flow cytometric analysis of free radical production. J Phar-macol Exp Ther 271: 1544±1549
Vormann J, GuÈ nther T (1986) Development of fetal mineral andtrace element metabolism in rats with normal as well asmagnesium- and zinc-de®cient diets. Biol Trace Elem Res 9:37±53
Vormann J, FoÈ rster C, Zippel U, Lozo E, GuÈ nther T, Merker HJ,Stahlmann R (1997) Joint cartilage lesions in magnesium-de®-cient rats at di�erent developmental stages. Calcif Tissue Int 61:230±238
419





























