Embed Size (px)
Citation preview

Ž .Brain Research 749 1997 320–328
Research report
Effects of extensor muscle afferents on the timing of locomotor activityduring walking in adult rats
Karim Fouad ), Keir G. PearsonDepartment of Physiology, UniÕersity of Alberta, Edmonton, Alberta T6G 2H7, Canada
Accepted 29 October 1996
Abstract
The influence of hind leg extensor muscle afferents on the timing of locomotor phase transitions was examined in adult, decerebraterats, walking on a treadwheel. Walking occurred either spontaneously or was induced by stimulation of the mesencephalic locomotorregion. Large diameter muscle afferents innervating the lateral or medial gastrocnemius were electrically stimulated during walking. Astimulus was delivered either at the onset of extensor muscle activity, or randomly during the step cycle. Stimulation with a train durationof 300 ms at the onset of extension increased the duration of the extensor bursts. The subsequent flexion phase was delayed. Stimulation
Ž .with a shorter stimulus train 150 ms early in extension had little effect on the extension phase duration. However when delivered at theend of extension the same stimulus significantly increased the duration of the extension phase and decreased the duration of the followingflexion phase. Stimulating near the end of the flexion phase delayed onset and decreased duration of the subsequent extension phase. Theeffects of stimulating extensor afferents during the extension phase were weaker but qualitatively similar, to those in cats, suggestingsimilar mechanisms. The results of this study also show major differences in the integration of extensor muscle afferents between adultand neonatal rats.
Keywords: Rat; Locomotion; Walking; Muscle afferent; Decerebrate
1. Introduction
Recently there has been increasing interest in the inves-tigation of locomotion in rats. Motor patterns during vari-ous types of rhythmic behavior have been described in
w xwalking and paralyzed animals 3,10,19,33 , supraspinalstructures involved in the initiation and regulation of loco-
w xmotion have been identified 3,6,18 and preparations fromneonatal animals have proven useful for the analysis of theorganization and function of the central pattern generator
w xfor locomotion 5,8,26,29,32 . In addition, the rat is cur-rently used extensively for examining recovery of locomo-
w xtion following spinal cord injury 4,24,43 . Despite thisattention on rat locomotion, very little is known about theinfluence of afferent input on the walking pattern. The
w xonly information comes from studies in neonates 23,27 .In these studies it has been shown that stimulation ofextensor afferents during the flexion phase with high inten-sity prolongs the flexion phase. Stimulation during theextension phase with low or high intensity resulted either
) Ž .Corresponding author. Fax: q1 403 492-8915.
in no effect or in a truncation of the extension phasew xrespectively 23 . One important observation has been that
the integration of afferent input changes early in postnatalw xdevelopment 23 . Stimulation of extensor afferents with a
low intensity stimulus delivered during the flexion phaserevealed this developmental change. This stimulation pro-
Ž .longed the flexion phase in post-natal days 1–3 P1–3animals but truncated it in the P4–6 animals.
Knowledge about sensory control of mammalian loco-motion has come largely from studies in cats. Afferent
w xcontrol of walking has been examined in intact 11,13 ,w x w xdecerebrate 12,15,22,40 and spinal cats 35,36 , as well
as during fictive locomotion in paralyzed animalsw x21,31,37 . Two groups of afferents are critical for theinitiation of flexion in a hindlimb; one group is signaling
w xhip extension 1,20,22,28 and the other is signaling thatw xthe extensor muscles are unloaded 14,40 . Preventing the
hip extension andror loading the extensor muscles in justone hindlimb can stop stepping in this limb, while the
w xremaining legs keep on stepping. Whelan et al. 40demonstrated that the effect of loading a hindlimb isprobably produced by feedback from group I afferents of
0006-8993r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 96 01328-5

( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328 321
extensor muscles. They found that electrical stimulation ofgroup I afferents in extensor nerves prolonged the stancephase during walking in decerebrate cats. It is likely thatactivity in group Ib afferents contributes significantly tothat effect, since their activation has been shown to pro-long extensor burst duration in reduced spinal preparationsw x7,36 . However, recently it has been demonstrated thatactivity in group Ia afferents is also capable of prolongingthe extension phase duration during fictive locomotionw x21 .
The aim of this study was to examine whether themechanisms of regulating the transition from the extensionto the flexion phase during walking in adult rats are similarto those found in cats. In addition, we wanted to comparethese mechanisms to those described in neonatal rats.Preliminary results of this study have been published in
w xabstract form 17 .
2. Materials and methods
Experiments were carried out on adult male Sprague-Ž .Dawley rats 300–500 g . Animals were initially anes-
thetized with halothane in 95% O r5% CO . A tracheo-2 2
tomy was performed and the anesthetic maintained via thetracheal cannula. The carotid arteries were ligated, and onejugular vein was cannulated for i.v. injections of drugs and
Ž .fluids. Atropine 0.5–1 mg was administered i.v. rou-tinely. Following this procedure the sciatic nerve of the leftleg was exposed and placed in a bipolar recording cuffelectrode. The cuff was approximately 4 mm long, with aninner diameter of about 1 mm. The bared ends of two
ŽTeflon-coated stainless steel wires Cooner Wire Co.,.AS631 were looped within the cuff and acted as recording
electrodes to monitor the strength of electrical stimuliapplied to nerves supplying the ankle extensor muscles.The nerve innervating either the lateral gastrocnemius-
Ž . Ž .soleus LGS muscle or medial gastrocnemius MG mus-cle was exposed and transected close to the muscle. About4 mm of this nerve was freed from the tibial nerve andthreaded into a small stimulation cuff. The end of the
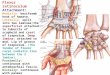
Ž .nerve was anchored to a pin fixed on the cuff Fig. 1A .The stimulation cuff was about 1 mm long with an insidediameter of roughly 0.4 mm. The stimulating electrodesconsisted of bared sections of two Teflon-coated stainless
Ž .steel wires Cooner Wire Co., AS631 divided into twostrands inside the cuff. To avoid movement of the cuffduring walking, the stimulation cuff was anchored to a
Ž .larger cuff placed on the adjacent tibial nerve Fig. 1A .This was essential because it minimized movement of thenerve in the cuff and helped to maintain constant stimulusstrength during walking. It also prevented damage of thenerve by the cuff moving within the leg.
Ž .Electromyographic EMG activity was recorded withbipolar Teflon-coated stainless steel recording electrodesŽ .Cooner Wire Co., AS631 implanted into the following
Fig. 1. A: construction of the stimulation electrode. The major parts of theelectrode were two silicone cuffs, the larger served as an anchor aroundthe distal tibial nerve, the smaller as the stimulation cuff. Bared sectionsof Teflon-coated wire were looped in the inside of the stimulation cuff.Either the LGS or MG nerve was thread through the stimulation cuff andfixed by tying the distal end to a minuten-pin attached to the cuffs. B:experimental setup. A decerebrated rat was placed in a stereotaxic holderand mounted above a treadwheel. Walking was initiated by stimulation of
Ž .the mesencephalic locomotor region MLR and electromyograms wererecorded from muscles of the hindlimbs.
Ž .hind leg muscles: ipsilateral leg – vastus lateralis VL ,Ž .semitendinosus ST , LG or MG; contralateral leg – MG
and ST. The wires from the EMG electrodes and the cuffelectrodes were led subcutaneously to a multipolar connec-tor near the middle of the animal’s back.
After placement of the electrodes, the animal wasmounted into a stereotaxic head holder and positioned on a
Ž .treadwheel diameter about 60 cm, width 8.7 cm; Fig. 1B .The treadwheel, with a negligible resistance was driven
w xonly by the animal 2 which caused a momentum of thespinning wheel, thereby stabilizing the walking speed.Both the right and the left parietal bones were removed.The sagittal and transverse sinuses were usually left intact.Cortical tissue was aspirated until the superior colliculiwere visible. A vertical decerebration was performed about2 mm rostral to the superior colliculi with a sharpened

( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328322
spatula and the remaining forebrain removed. The anes-thetic was discontinued immediately after decerebration.
Ž .In three animals, short bouts of walking up to 15 soccurred spontaneously about 40 min. after decerebration.
Ž .Longer periods of walking up to 53 s were induced in allanimals by stimulating the mesencephalic locomotor re-
w xgion 2,6,33 . The stimulus was a continuous train of 1-mspulses at 50 Hz of 5–30 mA. Usually the intensity wasslowly increased until the animal started running, thenslowly decreased to attain slow walking.
During periods of walking, the cuffed extensor nervewas stimulated at different intensities, relative to the low-est threshold volley recorded by the sciatic cuff. Thestimuli were either triggered 20 ms after the onset of
Žextensor activity in the left leg train: 300 ms duration, 200. ŽHz or delivered at various times during the step cycle 0.8
.Hz; train: 150 ms duration, 200 Hz . The onset of extensoractivity was detected by the on-line monitoring of a recti-fied and filtered EMG from the left VL muscle, controlledby an interactive computer program. All data were recordedon magnetic tape with a Vetter 4000A PCM recorder.
ŽSelected sequences were later digitized sampling rate.1000rs and stored on a computer disk using the Axotape
Ž .Axon Instruments data acquisition program. Analysiswas carried out using custom programs.
When triggering at the onset of extensor activity, theeffect of the stimulation was quantified by comparing the
Žduration of the perturbed step cycle onset of flexor burst.to onset following flexor burst to that of the preceding
step cycle. When stimulating at various times during thestep cycle, the duration of the extension and flexion phaseof the perturbed step cycle were compared with the dura-tion of this phase of the preceding cycle. The extensionphase duration was defined as the duration from the onsetof the VL burst to the onset of the subsequent ST burst,and the flexion phase duration as the duration between theonset of the ST burst and the onset of the subsequent VLburst. The results were divided into two groups, basedupon whether the stimulus occurred during the extensionor the flexion phase. These groups were further dividedinto five bins, depending on the timing of stimulus onsetŽ .1r5 of the duration of the phase of the preceding step .Step cycle, extension phase, and flexion phase duration ofthe perturbed step were compared with those of the preced-
Žing step with the use of a two-tailed, paired t-test P-.0.05 .
3. Results
3.1. General obserÕations
In three out of 10 animals, walking occurred eitherspontaneously or in response to a non-noxious cutaneousstimulus to the tail. The walking sequences lasted up to 16steps with a stepping frequency from 2–3 Hz. In these and
the other seven animals, locomotion was also initiated byelectrical stimulation of the mesencephalic locomotor re-
Ž .gion MLR . MLR stimulation induced long walking se-Žquences up to 34 bouts per animal, sometimes with
.)100 steps with a stepping frequency of 2–5 Hz. MLRinduced locomotion normally started with fast running orgalloping with full weight support. Reduction of the stimu-lus intensity after the beginning of locomotion decreasedwalking speed. Full weight support was maintained forperiods up to 50 s. Most bouts of walking ended with slow
Ž .stepping 1–2 Hz and dragging of the abdomen. Theseobservations correspond to those from studies by Bedford
w xet al. 2 .In spinal and decerebrate cats, it is well known that the
rate of stepping adapts to the speed of the motor drivenw xtreadmill upon which the animal is walking 16 . It was not
possible to establish whether this phenomenon occurs indecerebrate rats because the treadwheel was propelled bythe animal and not by a motor. However, we observed thatthe rate of stepping slowed when the turning of the tread-
Ž .wheel was resisted Fig. 2A and B which is similar toobservations in cats walking on a friction free treadmillw x38 . During these periods the magnitude of EMG activityin extensor muscles increased, and the burst duration was
Ž .prolonged Fig. 2A . When the treadwheel was preventedfrom turning, walking ceased. Another similarity to thefindings in cats was that stepping in an individual limbcould be interrupted by preventing completion of extensionŽw x .14,20 ; Fig. 2C . During these periods extensor activityremained high and the other three legs continued to step,usually at a lower rate.
3.2. Stimulating extensor afferents during extension
The similarities in the effects of resisting or preventingleg extension in the rat and the cat suggest that the afferentmechanisms controlling extensor activity are similar. Incats, feedback from group Ib afferents is one mechanismregulating the relation between load and activity of exten-sor muscles. Activity in these afferents increases the activ-ity of extensor muscles and prolongs the stance phasew x14,40 . To test whether this occurs in rats, we stimulated
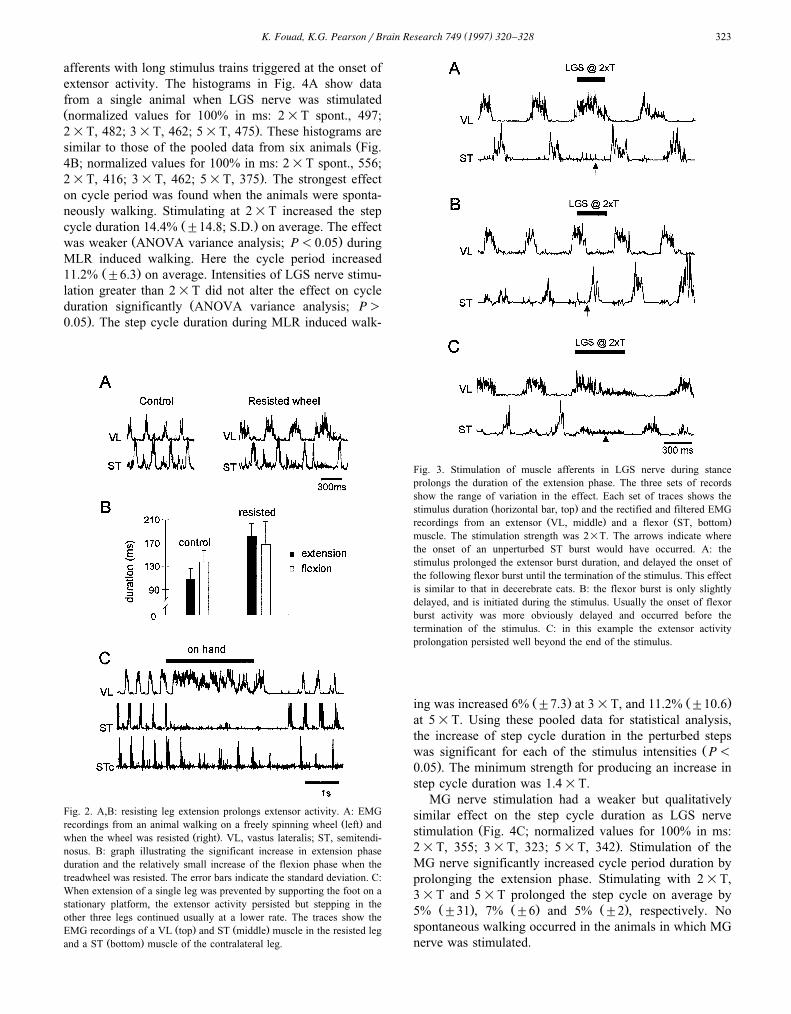
Ž . Ž .afferents in either the LGS ns6 or the MG ns4nerve. The stimulus train was triggered near the onset ofextensor activity with a delay of 20 ms. The train durationŽ .300 ms was set to outlast an undisturbed extension phase.This stimulation resulted in a prolongation of the durationof extensor EMG activity as well as in a delay of the
Ž .following flexion phase Fig. 3A . Although consistentlyprolonging extension phase duration there was consider-able variability between the trials. Generally, the effect ofthe stimulus was weak and flexion was initiated during the
Ž .period of stimulation Fig. 3B . Sometimes, however, thegeneration of the flexor burst was delayed far beyond the
Ž .stimulus duration Fig. 3C .Fig. 4 summarizes the results of stimulating extensor
( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328 323
afferents with long stimulus trains triggered at the onset ofextensor activity. The histograms in Fig. 4A show datafrom a single animal when LGS nerve was stimulatedŽnormalized values for 100% in ms: 2=T spont., 497;
.2=T, 482; 3=T, 462; 5=T, 475 . These histograms areŽsimilar to those of the pooled data from six animals Fig.
4B; normalized values for 100% in ms: 2=T spont., 556;.2=T, 416; 3=T, 462; 5=T, 375 . The strongest effect
on cycle period was found when the animals were sponta-neously walking. Stimulating at 2=T increased the step
Ž .cycle duration 14.4% "14.8; S.D. on average. The effectŽ .was weaker ANOVA variance analysis; P-0.05 during
MLR induced walking. Here the cycle period increasedŽ .11.2% "6.3 on average. Intensities of LGS nerve stimu-
lation greater than 2=T did not alter the effect on cycleŽduration significantly ANOVA variance analysis; P)
.0.05 . The step cycle duration during MLR induced walk-
Fig. 2. A,B: resisting leg extension prolongs extensor activity. A: EMGŽ .recordings from an animal walking on a freely spinning wheel left and
Ž .when the wheel was resisted right . VL, vastus lateralis; ST, semitendi-nosus. B: graph illustrating the significant increase in extension phaseduration and the relatively small increase of the flexion phase when thetreadwheel was resisted. The error bars indicate the standard deviation. C:When extension of a single leg was prevented by supporting the foot on astationary platform, the extensor activity persisted but stepping in theother three legs continued usually at a lower rate. The traces show the
Ž . Ž .EMG recordings of a VL top and ST middle muscle in the resisted legŽ .and a ST bottom muscle of the contralateral leg.
Fig. 3. Stimulation of muscle afferents in LGS nerve during stanceprolongs the duration of the extension phase. The three sets of recordsshow the range of variation in the effect. Each set of traces shows the
Ž .stimulus duration horizontal bar, top and the rectified and filtered EMGŽ . Ž .recordings from an extensor VL, middle and a flexor ST, bottom
muscle. The stimulation strength was 2=T. The arrows indicate wherethe onset of an unperturbed ST burst would have occurred. A: thestimulus prolonged the extensor burst duration, and delayed the onset ofthe following flexor burst until the termination of the stimulus. This effectis similar to that in decerebrate cats. B: the flexor burst is only slightlydelayed, and is initiated during the stimulus. Usually the onset of flexorburst activity was more obviously delayed and occurred before thetermination of the stimulus. C: in this example the extensor activityprolongation persisted well beyond the end of the stimulus.
Ž . Ž .ing was increased 6% "7.3 at 3=T, and 11.2% "10.6at 5=T. Using these pooled data for statistical analysis,the increase of step cycle duration in the perturbed steps
Žwas significant for each of the stimulus intensities P-.0.05 . The minimum strength for producing an increase in
step cycle duration was 1.4=T.MG nerve stimulation had a weaker but qualitatively
similar effect on the step cycle duration as LGS nerveŽstimulation Fig. 4C; normalized values for 100% in ms:
.2=T, 355; 3=T, 323; 5=T, 342 . Stimulation of theMG nerve significantly increased cycle period duration byprolonging the extension phase. Stimulating with 2=T,3=T and 5=T prolonged the step cycle on average by
Ž . Ž . Ž .5% "31 , 7% "6 and 5% "2 , respectively. Nospontaneous walking occurred in the animals in which MGnerve was stimulated.
( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328324
3.3. Stimulating extensor afferents at Õarious times duringthe step cycle
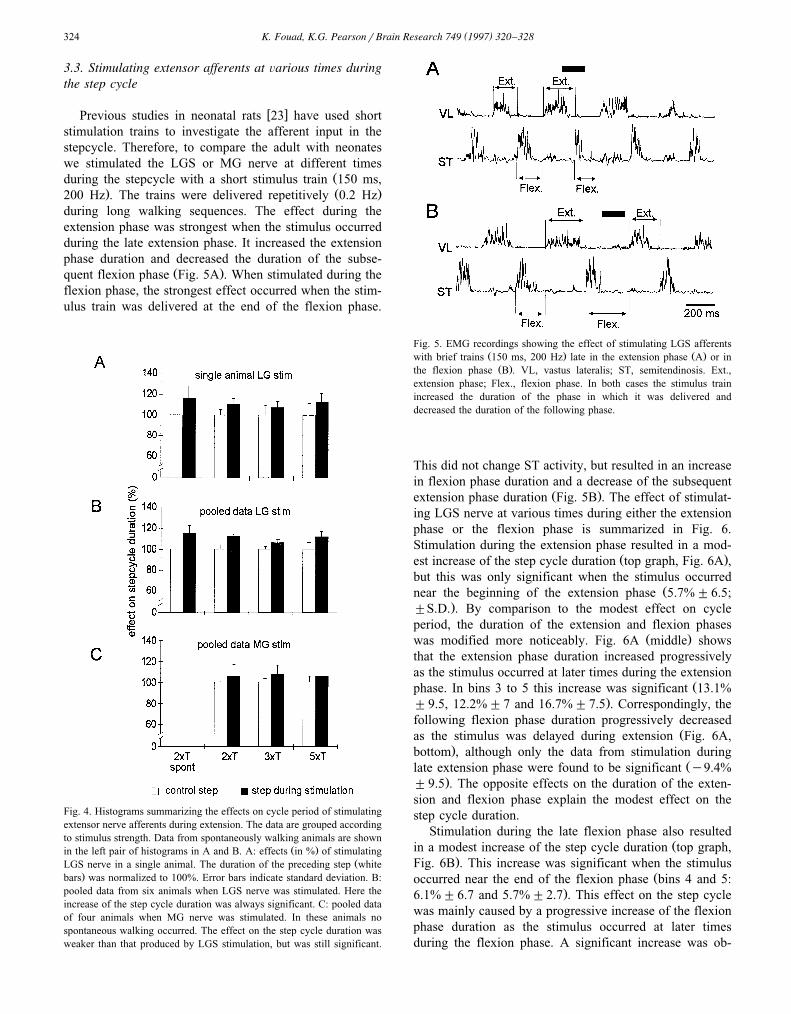
w xPrevious studies in neonatal rats 23 have used shortstimulation trains to investigate the afferent input in thestepcycle. Therefore, to compare the adult with neonateswe stimulated the LGS or MG nerve at different times
Žduring the stepcycle with a short stimulus train 150 ms,. Ž .200 Hz . The trains were delivered repetitively 0.2 Hz
during long walking sequences. The effect during theextension phase was strongest when the stimulus occurredduring the late extension phase. It increased the extensionphase duration and decreased the duration of the subse-
Ž .quent flexion phase Fig. 5A . When stimulated during theflexion phase, the strongest effect occurred when the stim-ulus train was delivered at the end of the flexion phase.
Fig. 4. Histograms summarizing the effects on cycle period of stimulatingextensor nerve afferents during extension. The data are grouped accordingto stimulus strength. Data from spontaneously walking animals are shown
Ž .in the left pair of histograms in A and B. A: effects in % of stimulatingŽLGS nerve in a single animal. The duration of the preceding step white
.bars was normalized to 100%. Error bars indicate standard deviation. B:pooled data from six animals when LGS nerve was stimulated. Here theincrease of the step cycle duration was always significant. C: pooled dataof four animals when MG nerve was stimulated. In these animals nospontaneous walking occurred. The effect on the step cycle duration wasweaker than that produced by LGS stimulation, but was still significant.
Fig. 5. EMG recordings showing the effect of stimulating LGS afferentsŽ . Ž .with brief trains 150 ms, 200 Hz late in the extension phase A or inŽ .the flexion phase B . VL, vastus lateralis; ST, semitendinosis. Ext.,
extension phase; Flex., flexion phase. In both cases the stimulus trainincreased the duration of the phase in which it was delivered anddecreased the duration of the following phase.
This did not change ST activity, but resulted in an increasein flexion phase duration and a decrease of the subsequent
Ž .extension phase duration Fig. 5B . The effect of stimulat-ing LGS nerve at various times during either the extensionphase or the flexion phase is summarized in Fig. 6.Stimulation during the extension phase resulted in a mod-
Ž .est increase of the step cycle duration top graph, Fig. 6A ,but this was only significant when the stimulus occurred
Žnear the beginning of the extension phase 5.7%"6.5;."S.D. . By comparison to the modest effect on cycle
period, the duration of the extension and flexion phasesŽ .was modified more noticeably. Fig. 6A middle shows
that the extension phase duration increased progressivelyas the stimulus occurred at later times during the extension
Žphase. In bins 3 to 5 this increase was significant 13.1%."9.5, 12.2%"7 and 16.7%"7.5 . Correspondingly, the
following flexion phase duration progressively decreasedŽas the stimulus was delayed during extension Fig. 6A,
.bottom , although only the data from stimulation duringŽlate extension phase were found to be significant y9.4%
."9.5 . The opposite effects on the duration of the exten-sion and flexion phase explain the modest effect on thestep cycle duration.
Stimulation during the late flexion phase also resultedŽin a modest increase of the step cycle duration top graph,
.Fig. 6B . This increase was significant when the stimulusŽoccurred near the end of the flexion phase bins 4 and 5:
.6.1%"6.7 and 5.7%"2.7 . This effect on the step cyclewas mainly caused by a progressive increase of the flexionphase duration as the stimulus occurred at later timesduring the flexion phase. A significant increase was ob-
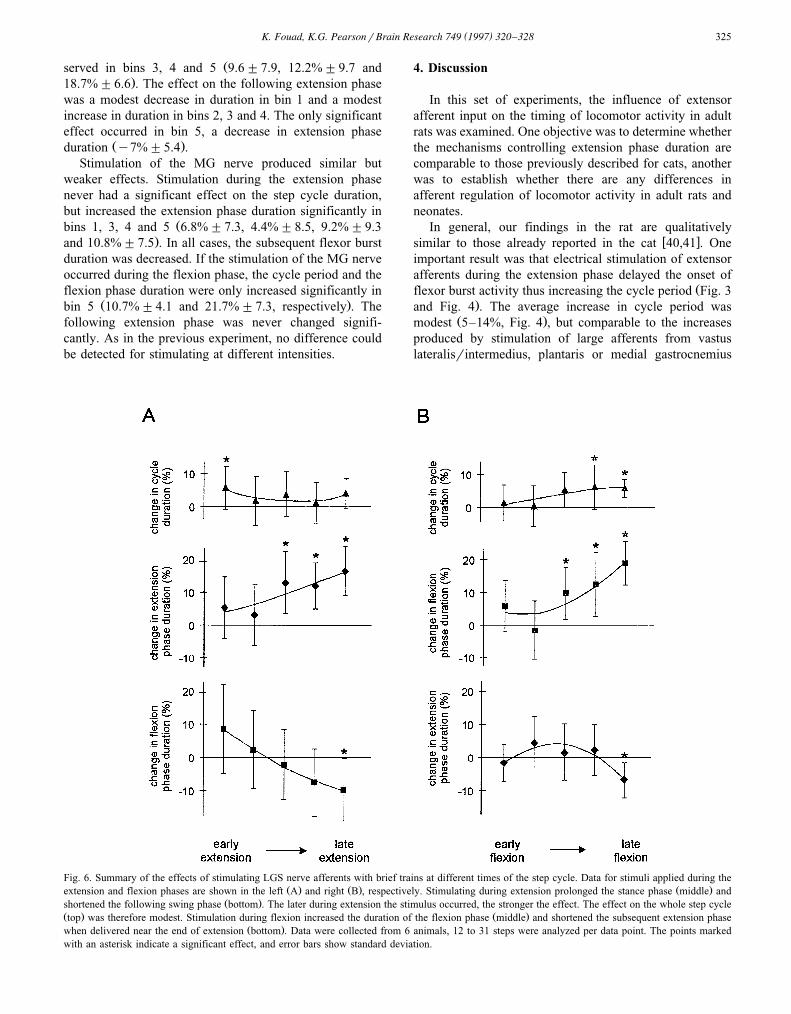
( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328 325
Žserved in bins 3, 4 and 5 9.6"7.9, 12.2%"9.7 and.18.7%"6.6 . The effect on the following extension phase
was a modest decrease in duration in bin 1 and a modestincrease in duration in bins 2, 3 and 4. The only significanteffect occurred in bin 5, a decrease in extension phase
Ž .duration y7%"5.4 .Stimulation of the MG nerve produced similar but
weaker effects. Stimulation during the extension phasenever had a significant effect on the step cycle duration,but increased the extension phase duration significantly in
Žbins 1, 3, 4 and 5 6.8%"7.3, 4.4%"8.5, 9.2%"9.3.and 10.8%"7.5 . In all cases, the subsequent flexor burst
duration was decreased. If the stimulation of the MG nerveoccurred during the flexion phase, the cycle period and theflexion phase duration were only increased significantly in
Ž .bin 5 10.7%"4.1 and 21.7%"7.3, respectively . Thefollowing extension phase was never changed signifi-cantly. As in the previous experiment, no difference couldbe detected for stimulating at different intensities.
4. Discussion
In this set of experiments, the influence of extensorafferent input on the timing of locomotor activity in adultrats was examined. One objective was to determine whetherthe mechanisms controlling extension phase duration arecomparable to those previously described for cats, anotherwas to establish whether there are any differences inafferent regulation of locomotor activity in adult rats andneonates.
In general, our findings in the rat are qualitativelyw xsimilar to those already reported in the cat 40,41 . One
important result was that electrical stimulation of extensorafferents during the extension phase delayed the onset of
Žflexor burst activity thus increasing the cycle period Fig. 3.and Fig. 4 . The average increase in cycle period was
Ž .modest 5–14%, Fig. 4 , but comparable to the increasesproduced by stimulation of large afferents from vastuslateralisrintermedius, plantaris or medial gastrocnemius
Fig. 6. Summary of the effects of stimulating LGS nerve afferents with brief trains at different times of the step cycle. Data for stimuli applied during theŽ . Ž . Ž .extension and flexion phases are shown in the left A and right B , respectively. Stimulating during extension prolonged the stance phase middle and
Ž .shortened the following swing phase bottom . The later during extension the stimulus occurred, the stronger the effect. The effect on the whole step cycleŽ . Ž .top was therefore modest. Stimulation during flexion increased the duration of the flexion phase middle and shortened the subsequent extension phase
Ž .when delivered near the end of extension bottom . Data were collected from 6 animals, 12 to 31 steps were analyzed per data point. The points markedwith an asterisk indicate a significant effect, and error bars show standard deviation.

( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328326
Ž . w xmuscles 14–20% in decerebrate walking cats 40,41 .One clear quantitative difference with cats was the effectof stimulating the LGS nerve. In cats, stimulation of largeafferents in the LGS nerve powerfully influences extensorbursts, usually prolonging these bursts for the duration of
w xthe stimulus train 40 . This strong effect was observedŽ .only rarely in rats an example is shown in Fig. 3A . Even
though the effect of LGS nerve stimulation was weak inthe rat compared to the cat, we did observe that LGS nervestimulation produced more powerful effects than MG nerve
Žstimulation average increase in cycle period 11.2% for.LGS compared to 5% for MG .
On most trials stimulation of either the LGS or MGnerve during extension had a proportionally larger effect
Žon extensor burst duration than on the cycle period Fig..6 . Usually the prolongation of the extensor burst duration
was compensated to some extent by a shortening of thefollowing flexion phase. The relatively weak effect oncycle period is probably due to the overall timing of cycleperiod being dominated by the rate of stepping in the otherthree unperturbed legs.
Although the recruitment order of different groups ofafferents in hind leg nerves of the rat have not beendefined for electrical stimuli, we attribute the effects wehave observed to stimulation of afferents arising frommuscle spindles and Golgi tendon organs. The effects onthe timing of the locomotor activity were apparent at astimulus strength of 1.4=T and were usually maximal at
Ž .2=T Fig. 4 . This range of stimulus strengths wouldhave preferentially activated large muscle afferents. Thelargest afferents from the gastrocnemius muscles in the ratare considered to arise from muscle spindles and Golgi
w xtendon organs 30 . There was no possibility of effectsbeing produced indirectly by evoked muscle contractionsbecause the nerves were cut distally. A comparison withdata from the cat also supports our conclusion that theeffects of stimulating the LGS and MG nerves are due toactivation of spindle and tendon organ afferents. The mini-mal stimulus strength for influencing the duration of exten-sor bursts in decerebrate walking cats is about 1.5=T and
w xmaximal effects are produced at 2=T 40 . Over thisrange of stimulus strengths most of the afferents fromprimary spindle endings and Golgi tendon organs, andsome afferents from secondary spindle endings, are re-
w xcruited 25 .Further similarities with the cat were found by loading
extensor muscles. Resisting the turning of the treadwheelon which the rats were walking preferentially increased the
Ž .duration of extensor muscle activity Fig. 2A,B . Qualita-tively similar observations have been made in decerebrate
w xcats walking on a friction free treadmill 38 . Furthermore,preventing the extension of a hind leg in a walking rat
Ž .prevented stepping in just this limb Fig. 3C . This iscomparable to the observation in the cat, where it has beenshown that supporting a leg on a stationary object inhibits
w xthe initiation of the flexion phase 20 . The functional
interpretation of these observations in the cat is that a legmust be unloaded and extended for the swing phase to be
w xinitiated 34 . The corresponding observations in the ratindicate that this may also be true for the rat. This issupported by our finding that extensor burst duration canbe prolonged by electrical stimulation of extensor group I
Ž .afferents Fig. 3, Fig. 4 . In the cat there is now consider-able evidence that this effect is due primarily to activation
w xof Ib afferents from the Golgi tendon organs 34 . If this istrue in the rat, then a general rule for mammalian locomo-
Ž .tion may be that stance extension is maintained byfeedback from tendon organs. Unloading of the extensor
Žmuscles near the end of stance due to the animal’s weightbeing borne by other legs and to shortening of extensor
. Ž .muscles is required for the initiation of swing flexion .One further similarity to the cat was that the effects of
stimulating the extensor nerves during the extension phasewere greater during spontaneous locomotion, compared to
Ž .those produced during MLR-induced locomotion Fig. 4 .Recently, Whelan observed that increasing the stimulusstrength to the MLR in decerebrate walking cats reducesthe effectiveness of group I afferents on extensor burst
w xduration 42 . In both rats and cats this difference is notdue to differences in walking speed, because it is observedat the same cycle periods during spontaneous and MLRinduced walking.
Although the numerous similarities in the data from catsand rats imply similar mechanisms for afferent regulationof stepping, we did find one major difference. In the adultrat, stimulating the extensor afferents during the flexionphase prolonged the flexion phase, whereas in cats the step
w xcycle is reset by termination of the flexion phase 7,40 .Our observations in adult rats differ in some important
aspects from those in neonates. In neonates of age lessthan P6, low and high intensity stimulation of extensorafferents delivered during the extension phase has eitherno effect or causes a resetting of the step cycle to flexion
w xby truncating the extensor burst, respectively 23 . In adultswe observed that stimulation of the extensor afferents with
Ž .different intensities 2–5=T always increased the dura-tion of the extension phase. Another difference is that inadults stimulation during the flexion phase at all intensitiesalways prolonged the flexion phase. The same effect wasobserved in neonatal rats only when stimulated with high
Ž . Ž .intensities )5=T . Low intensity stimulation -3=Tof the extensor afferents prolonged the flexion phase justin P1–3 animals, but truncated it in P4–6 animals, suggest-ing a developmental change in the integration of the
w xextensor afferents 23 . The observed differences betweenadult rats and neonates might be explained by the occur-rence of a second developmental change in the integrationof afferent information. The idea of such a change isstrengthened by the fact that P6 neonates have not started
w xto support their body weight 39 and by the finding thatdirect cortico-motoneuronal synaptic contacts are not es-
w xtablished until about P7 9 . Another possible explanation

( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328 327
for differences in the effects between neonatal rats andadults could be the different preparation. In the neonate,afferents from the quadriceps muscle, a knee extensor,were stimulated, whereas in this study ankle extensorafferents were examined. Another difference was thatneonatal rats were paralyzed and fictive locomotion wasexamined, whereas adult animals were walking on a tread-wheel. This explanation, however, appears unlikely, be-cause in cats these differences have not been found toqualitatively influence the results. Similar effects are pro-duced when extensor group I afferents from knee or ankle
w xextensors are stimulated 21,40 , and there are no majordifference between the results obtained in walking animalsw x w x40 and in paralyzed preparations 21 .
In conclusion, our results suggest that similar sensorymechanisms regulate the transition from the extension tothe flexion phase in adult rats and adult cats. Further,differences between older neonates and adult rats indicatedevelopmental changes of these mechanisms sometimeafter neonatal day 6.
Acknowledgements
The authors thank R. Gramlich for excellent technicalassistance. We would also like to thank D. Bennet, G.Hiebert and J. Misiaszek for reading the manuscript. Thiswork was supported by grants of the Deutsche Forschungs-gemeinschaft and the Medical Research Council of Canada.
References
w x1 Andersson, O. and Grillner, S., Peripheral control of the cat’s stepcycle. II. Entrainment of the central pattern generators for locomo-tion by sinusoidal hip movements during fictive locomotion, Acta
Ž .Physiol. Scand., 118 1983 229–239.w x2 Bedford, T.G., Loi, P.K. and Crandall, C.C., A model of dynamic
exercise: the decerebrate rat locomotor preparation, J. Appl. Physiol.,Ž .72 1992 121–127.
w x3 Bem, T., Orsal, D. and Cabelguen, J.-M., Fictive locomotion in theŽ .adult thalamic rat, Exp. Brain Res., 97 1993 301–304.
w x4 Bregman, B.S., Kunkel-Badgen, E., Schnell, E., Dai, L., Gao, H.N.and Schwab, M.E., Recovery from spinal cord injury mediated by
Ž .antibodies to neurite growth inhibitors, Nature, 378 1995 498–501.w x5 Cazalets, J.-R., Borde, M. and Clarac, F., The synaptic drive from
the spinal locomotor network to motoneurons in the newborn rat, J.Ž .Neurosci., 16 1996 298–306.
w x6 Coles, S.K., Iles, J.F. and Nicolopoulos-Stournaras, S., The mesen-cephalic centre controlling locomotion in the rat, Neuroscience, 28Ž .1989 149–157.
w x7 Conway, B.A., Hultborn, H. and Kiehn, O., Proprioceptive inputresets central locomotor rhythm in the spinal cat, Exp. Brain Res.,
Ž .68 1987 643–656.w x8 Cowley, K.C. and Schmidt, B.J., Some limitations of ventral root
recordings for monitoring locomotion in the in vitro neonatal ratŽ .spinal cord, Neurosci. Lett., 171 1994 142–146.
w x9 Curfs, M.H.J.M., Gribnau, A.A.M. and Dederen, P.J.W.C., Directcortico-motoneuronal synaptic contacts are present in the adult ratcervical spinal cord and are first established at postnatal day 7,
Ž .Neurosci. Lett., 205 1996 123–126.
w x10 de Leon, R., Hodgson, J.A., Roy, R.R. and Edgerton, R.V., Exten-sor- and flexor-like modulation within motor pools of the rat hindlimb
Ž .during treadmill locomotion and swimming, Brain Res., 654 1994241–250.
w x11 Drew, T. and Rossignol, S., A kinematic and electromyographicstudy of cutaneous reflexes evoked from the forelimbs of unre-
Ž .strained walking cats, J. Neurophysiol., 57 1987 1160–1184.w x12 Duysens, J., Reflex control of locomotion as revealed by stimulation
of cutaneous afferents in spontaneously walking premammillaryŽ .cats, J. Neurophysiol., 40 1977 737–751.
w x13 Duysens, J. and Loeb, G.E., Modulation of ipsi- and contralateralŽ .reflex responses in unrestrained walking cats, Brain Res., 44 1980
1024–1037.w x14 Duysens, J. and Pearson, K.G., Inhibition of flexor burst generation
by loading ankle extensor muscles in walking cats, Brain Res., 187Ž .1980 321–332.
w x15 Duysens, J. and Stein, R.B., Reflex induced by nerve stimulation inwalking cats with implanted cuff electrodes, Exp. Brain Res., 32Ž .1978 213–224.
w x16 Forssberg, H. and Grillner, S., The locomotion of the acute spinalŽ .cat injected with clonidine i.v., Brain Res., 50 1973 184–186.
w x17 Fouad, K. and Pearson, K.G., Effects of stimulation of extensormuscle afferents during walking in adult rats, Soc. Neurosci. Abstr.,
Ž .22 1996 p. 1097.w x18 Garcia-Rill, E., The basal ganglia and the locomotor regions, Brain
Ž .Res. ReÕ., 11 1986 47–63.w x19 Goudard, I., Orsal, D. and Cabelguen, J.-M., An electromyographic
study of the hindlimb locomotor movements in the acute thalamicŽ .rat, Eur. J. Neurosci., 4 1992 1130–1139.
w x20 Grillner, S. and Rossignol, S., On the initation of the swing phase ofŽ .locomotion in chronic spinal cats, Brain Res., 146 1978 269–277.
w x21 Guertin, P., Angel, M.J., Perreault, M.C. and McCrea, D.A., Ankleextensor group I afferents excite extensors throughout the hindlimbduring MLR evoked fictive locomotion in the cat, J. Physiol., 487Ž .1995 197–209.
w x22 Hiebert, G.W., Whelan, P.J., Prochazka, A. and Pearson, K.G.,Contribution of hind limb flexor muscle afferents to the timing of
Ž .phase transitions in the cat step cycle, J. Neurophysiol., 75 19961126–1137.
w x23 Iizuka, M., Kiehn, O. and Kudo, N., Development in neonatal rats ofthe sensory resetting of the locomotor rhythm induced by NMDA
Ž .and 5-HT, Exp. Brain Res., 1996 in press.w x24 Iwashita, Y., Kawaguchi, S. and Murata, M., Restoration of function
by replacement of spinal cord segments in the rat, Nature, 367Ž .1994 167–170.
w x25 Jack, J.J.B., Some methods for selective activation of muscle affer-Ž .ent fibres. In R. Porter Ed. , Studies in Neurophysiology, Cam-
bridge University Press, Cambridge, UK, 1978, pp. 155–176.w x26 Kiehn, O. and Kjaerulff, O., Spatiotemporal characteristics of 5-HT
and Dopamine-induced rhythmic hindlimb activity in the in vitroŽ .neonatal rat, J. Neurophysiol., 75 1996 1472–1482.
w x27 Kiehn, O., Iizuka, M. and Kudo, N., Resetting from low thresholdafferents of N-methyl-D-aspartate-induced locomotor rhythm in theisolated spinal cord–hindlimb preparation from newborn rats, Neu-
Ž .rosci. Lett., 148 1992 43–46.w x28 Kriellaars, E.I., Brownstone, R.M., Noga, B. and Jordan, L.M.,
Mechanical entrainment of fictive locomotion in the decerebrate cat,Ž .J. Neurophysiol., 71 1994 2074–2086.
w x29 Kudo, N. and Yamada, T., Development of the monosynaptic stretchŽ .reflex in the rat: in vitro study, Neurosci. Lett., 148 1985 43–46.
w x30 Lewin, G.R. and McMahon, S.B., Physiological properties of pri-mary sensory neurons appropriately and inappropriately innervating
Ž .skeletal muscle in adult rats, J. Neurophysiol., 66 1991 1218–1231.w x31 McCrea, D.A., Shefchyk, S.J., Stephens, M.J. and Pearson, K.G.,
Disynaptic group I excitation of synergist ankle extensor motoneu-Ž .rones during fictive locomotion, J. Physiol., 487 1995 527–539.
w x32 MacLean, J.N., Hochman, S. and Magnuson, D.S.K., Lamina VII

( )K. Fouad, K.G. PearsonrBrain Research 749 1997 320–328328
neurons are rhythmically active during locomotor-like-activity in theŽ .neonatal rat spinal cord, Neurosci. Lett., 197 1995 9–12.
w x33 Nicolopoulos-Stournaras, S. and Iles, J.F., Hindlimb muscle activityŽ . Žduring locomotion in the rat Rattus norÕegicus Rodentia: Muri-
. Ž .dae , J. Zool. Lond., 203 1984 427–440.w x34 Pearson, K.G., Proprioceptive regulation of locomotion, Curr. Opin.
Ž .Neurobiol., 5 1995 786–791.w x35 Pearson, K.G. and Collins, D.F., Reversal of the influence of the
group Ib afferents from plantaris on activity in medial gastrocnemiusŽ .muscle during locomotor activity, J. Neurophysiol., 70 1993
1009–1017.w x36 Pearson, K.G., Ramirez, J.M. and Jiang, W., Entrainment of the
locomotor rhythm by group Ib afferents from ankle extensor musclesŽ .in spinal cats, Exp. Brain Res., 90 1992 557–566.
w x37 Perreault, M.-C., Drew, T. and Rossignol, S., Activity of medullaryreticulospinal neurons during fictive locomotion, J. Physiol., 487Ž .1993 211–220.
w x38 Shik, M.L., Severin, F.V. and Orlovskii, G.N., Control of walking
and running by means of electrical stimulation of the mid-brain,Ž .Biofizyka, 11 1966 756–765.
w x39 Westerga, J. and Gramsbergen, A., Changes in the electromyogramof two major hindlimb muscles during locomotor development in the
Ž .rat, DeÕ. Brain Res., 57 1990 163–174.w x40 Whelan, P.J., Hiebert, G.W. and Pearson, K.G., Stimulation of the
group I extensor afferents prolongs the stance phase in walking cats,Ž .Exp. Brain Res., 103 1995 20–30.
w x41 Whelan, P.J., Hiebert, G.W. and Pearson, K.G., Plasticity of exten-sor group I pathway controlling the stance to swing transition in the
Ž .cat, J. Neurophysiol., 74 1995 2782–2787.w x42 Whelan, P.J., Function of group I extensor feedback in the control
of locomotion, Ph.D. Thesis, University of Alberta, 1996.w x43 Yakovleff, A., Cabelguen, J.-M., Orsal, D., Gimenez y Ribotta, M.,
Rajaofetra, N., Drian, M.-J., Bussel, B. and Privat, A., Fictive motoractivities in adult chronic spinal rats transplanted with embryonic
Ž .brainstem neurons, Exp. Brain Res., 106 1995 69–78.