Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIO~.~JYSICS Vol. 229, No. 1, February 15, pp. 359-36’7, 1934
Effects of Enzymatic Deglycosylation on the Biological Activities of Human Thrombin and Antithrombin’
LOUIS ROSENFELD AND ISIDORE DANISHEFSKY’
Department of Biochemistry, New York Medical College, Valhalla, New York 10595
Received September 16, 1933
Sequential digestion of human thrombin and antithrombin with neuraminidase, /3- galactosidase, /3-N-acetylglucosaminidase, and endo+N-acetylglucosaminidase D re- sulted in the successive removal of sialic acid, galactose, N-acetylglucosamine, and mannose and more N-acetylglucosamine residues. The products obtained after each stage of deglycosylation had electrophoretic. mobilites that were consistent with the calculated change in mass expected from the cleavage of the sugar moieties. The modified thrombins did not lose fibrinogen-clotting activity, amidolytic activity, nor the ability to form complexes with antithrombin. In addition, asialothrombin and asialoagalac- tothrombin caused the same extent of platelet release as did control thrombin. The products obtained after removal of sugars from antithrombin retained thrombin-neu- tralizing activity. In the presence of heparin the inhibition of thrombin as well as factor Xa was enhanced. Thus, the sugar residues of thrombin and antithrombin are not required for the formation of enzyme-inhibitor complexes or for the other activities that were measured.
Thrombin is a plasma serine protease which plays a central role in blood coag- ulation (2). It is a glycoprotein that is gen- erated from prothrombin as a result of limited proteolysis by factor Xa. The latter, in turn, is activated in earlier steps of the blood coagulation cascade (3). Recent elu- cidations of the structures of the carbo- hydrate chains of human prothrombin and thrombin have shown that both have bian- tennary sugar chains covalently attached via N-glycosidic bonds to asparagine res- idues (4, 5). The sugar sequence for pro- thrombin was found to be (NeuNAca2 - 6Gal/?l - 4GlcNAcPl - 2Manal -‘)g, 6Man/31 - 4GlcNAcfll - 4GlcNAc -
1 These studies were supported by Grant HL-16955 from the National Institutes of Health. A preliminary presentation of this work was made at the 74th Annual Meeting of the American Society of Biological Chem- ists (1).
a To whom inquires should be addressed.
Asn.3 Prothrombin has three such chains, two of which are lost upon conversion to thrombin (3,4). A recent report on the car- bohydrate structure of thrombin shows that an additional fucose moiety may be attached to the N-acetylglucosamine res- idue linked to asparagine (5).
Antithrombin is a circulating glycopro- tein that inhibits thrombin, factor Xa, and various other components of the blood co- agulation system (6). It contains four sugar chains having structures that are identical to those of prothrombin (7, 8).
Some aspects of the contribution of the carbohydrate moieties to the biological ac- tivities of thrombin and antithrombin have
’ Abbreviations used: endo D, endo+N-acetylglu- cosaminidase D; S-2238, H-D-phenylalanyl-L-pipe- colyl-L-argenine-pnitroanilide; S-2222, N-benzoyl-L- isoleucyl-L-glutamyl-L-argenine-pnitroanilide; SDS, sodium dodecyl sulfate; Neu, neuraminic acid; NAc, N-acetyl-.
359 0003-9861/34 $3.00 Copyright 0 1984 by Academic Press. Inc. All righta of reproduction in any form resenred.

360 ROSENFELD AND DANISHEFSKY
been studied previously. Removal of sugars from bovine thrombin by a mixture of en- zymes had no effect on clotting activity (9). Cleavage of sialic acid from bovine pro- thrombin dramatically decreased its cir- culatory half-life in rats (lo), indicating that the sugars may have a role in the regulation and turnover of prothrombin in the circulation. Enzymatic removal of sialic acid from human antithrombin did not re- sult in any loss of thrombin inhibitory ac- tivity both in the absence or presence of heparin (11). Also, the ability of anti- thrombin to form a complex with thrombin did not change after neuraminidase treat- ment. Labeling of the sialic acid moieties of antithrombin by means of periodate and tritiated sodium borohydride yielded fully active antithrombin (12).
This report describes a systematic study on the biological activities of both throm- bin and antithrombin after sequential re- moval of sialic acid, galactose, and N-ace- tylglucosamine residues. These partially deglycosylated glycoproteins were further treated with endo-@-N-acetylglucosamin- idase D (13) to cleave all the remaining sugar residues except for the asparagine- linked N-acetylglucosamine. After removal of each sugar unit in thrombin, the prod- ucts were assayed for fibrinogen-clotting and amidolytic activities, as well as sus- ceptibility to inhibition by antithrombin. With the modified antithrombins, mea- surements were made of their capability to neutralize thrombin and factor Xa. Ad- ditionally, the platelet release reaction was measured for asialo- and asialoagalac- tothrombin. The results demonstrate con- clusively that removal of sugar residues does not alter the biological activities of human thrombin and antithrombin.
MATERIALS AND METHODS
Materials Human thrombin (2967 units/mg) was kindly provided by Dr. John W. Fenton, New York State Department of Health, Albany, New York. Hu- man antithrombin was supplied by the American Red Cross Fractionation Center, Bethesda, Maryland. Neuraminidase (Type VI, Clostridium perftingens), (5 galactosidase (jack bean), and @-N-acetylglucosami-
nidase (jack bean) were purchased from Sigma Chemical Company, St. Louis, Missouri. Endo-P-N- acetylglucosaminidase D (Streptococcw‘ pneummziae) was obtained from Miles Laboratories, Elkhart, In- diana. Fibrinogen, purchased from Calbiochem, La Jolla, California, was further purified according to the method of Laki (14). Beef lung sodium heparin was generously supplied by the Upjohn Company, Kalamazoo, Michigan. Factor Xa, thrombin substrate S-2238 (H-D-phenylalanyl-L-pipecolyl-L-argenine-p- nitroanilide) and factor Xa substrate S-2222 (N-ben- z0yl-L-isoleucyl-L-glutamyl-L-argenine-pnitroani- lide) were the products of Kabi Diagnostica, Stock- holm, Sweden. Sugar standards were obtained from Pfanstiehl. Waukegan, Illinois.
Thrombin activities. Fibrinogen-clotting activity was determined after adding 100 ~1 of appropriately diluted solutions of thrombin in 0.15 M NaCI, 0.01 M Tris-HCl, 0.825% polyethylene glycol-6000, pH 7.5, to a mixture of 200 ~1 0.5% fibrinogen and 200 ~1 of the buffer at 3’7°C. Units were based on the thrombin provided by Dr. Fenton (2). Amidolyic activity was measured by the ability of thrombin to cleave the chromogenic substrate S-2238 (15). The ability to cause platelet activation was measured by quantitating re- leased ATP with the firefly luciferin-luciferase assay system of Detwiler (16). The formation of complexes with untreated antithrombin were studied by mixing 69.1 c(g antithrombin with 43.5 pg of thrombin (I:1 molar ratio) in a total volume of 100 ~1. After 20 s, and 3 and 10 min, 30-p] aliquots were removed and added to 15 ~1 of a buffer containing 10% sodium dodecyl sulfate (SDS) and 9% sodium mercaptoeth- anol. These aliquots were heated in boiling water for 3 min prior to slab gel electrophoresis.
Antithrvmbin activity assays. Progressive thrombin neutralization was measured by the method of Ger- endas (17). Enchancement of thrombin inhibition by heparin (heparin cofactor activity) was measured as described by Rosenberg and Damus (6). Inhibition of thrombin and factor Xa amidolytic activity in the presence of heparin was measured by a modification of the microtest plate procedure of Radoff and Dan- ishefsky (manuscript submitted for publication). Heparin and antithrombin samples were placed in the microtest wells in triplicate and the protease was added at fixed time intervals. For thrombin inhibition, 2.5 pg/ml heparin, and 0 to 0.250 unit/ml antithrombin were used; for factor Xa, 5 pg/ml heparin and 0 to 1.0 unit/ml antithrombin were used. After exactly 1 min, an aliquot of S-2238 (for thrombin) or S-2222 (for factor Xa) was placed in the wells and the plates were incubated at room temperature. The reaction was stopped after exactly 15 min with acetic acid. Residual amidolytic activity was measured by the absorbance of released p-nitroanilide, using a Bio- Tek (Burlington, Vt.) EIA reader, Model EL307, with

EFFECTS OF DEGLYCOSLYATION OF HUMAN THROMBIN AND ANTITHROMBIN 361
a 405-nm filter. Absorbances were compared with a standard curve.
The ability of antithrombin samples to form com- plexes with untreated thrombin was determined by incubating 23.7 pg thrombin with 75.4 pg antithrombin (1:2 molar ratio) in a total volume of 75 ~1 at 37°C. Thirty-microliter aliquots were removed after 20 s and at 3 min, and treated with SDS as described above.
Slab gel electrophoresis. Untreated and glycosidase- treated thrombin and antithrombin as well as the complex-formation experiments were run on SDS slab gel electrophoresis using 12.5% polyacrylamide for the running gel as described by Laemmli (18).
Carbohydrate analyses. Samples were dialyzed against 0.1 M NaCl to remove Tris buffer prior to analyses. Neutral and amino sugar hydrolysis, and conversion to alditol acetates was accomplished by the method of Reading et al. (19). Gas chromato- graphic conditions were those of Niedermeier (20), using arabinose as the internal standard. For released sugars, alditol acetates were prepared from samples that were not subjected to dialysis nor hydrolysis.
Protein estimatierk Protein concentrations were determined by corrected absorbances at 280 nm, using E’% i em = 18.3 for thrombin (2) and E:%,,, = 6.5 for an- tithrombin (22). kl, was assumed to be 36,500 for thrombin (2) and 58,000 for antithrombin (22).
Glycosidese digestions. All digestions were carried out in solutions of 0.05 M sodium acetate, 0.15 M sodium chloride, 0.02% sodium azide, pH 6.0, except for the B-galactosidase digestion of thrombin, which was performed in the same buffer adjusted to pH 5.2. Units for the exoglycosidases were measured with appro- priate substrates at the pH used in the experiments. Units for endo D were those provided by the supplier. Neuraminidase digestions were carried out for 2 days at 5°C using 0.058 and 0.044 units/mg for thrombin and antithrombin, respectively (11). /3-Galactosidase digestion of thrombin was conducted at 22°C for 42 h using 0.041 units/mg substrate. /3-Galactosidase digestion of antithrombin was done at 23°C for 22 h, then at 5°C for 119 h, using 0.0093 units/mg substrate. The rest of the incubations were carried out at 23°C for 20 to 22 h. B-N-Acetylglucosaminidase digestions were conducted using 0.88 or 0.44 units/mg substrate for thrombin or antithrombin, respectively; endo D digestions employed 0.022 or 0.0068 units/ma sub- strate for thrombin or antithrombin, respectively.
In the experiments, 20-25 mg of thrombin or an- tithrombin were generally employed. Following each digestion, an aliquot containing 3 to 4 mg was removed for activity assays and sugar analysis. After removal of a small sample that served as a parallel control, the remainder was treated with the next enzyme. All samples and controls were stored at -25°C until as- sayed. In all measurements, the samples were com-
pared both to their parallel control and to untreated thrombin or antithrombin.
RESULTS AND DISCUSSION
Glycosidic digestions. The results pre- sented describe a typical experiment for thrombin and for antithrombin. Most gly- cosidic digestions were conducted at pH 6.0 which is near the optimum for these enzymes. Digestion of thrombin with jack bean P-galactosidase was performed at pH 5.2, since the optimum for this enzyme is pH 3.5 (23). This pH was sufficient for ga- lactosidase activity and high enough to preserve thrombin biological activity. An- tithrombin was incubated with the enzyme at pH 6.0 since it is not stable below that pH; however, the incubation time was ex- tended (see Materials and Methods).
The sugar compositions of the products obtained after treatment of thrombin and antithrombin with glycosidases are shown in Table I. The analyses for galactose were consistently higher than expected. This may be due to minute amounts of glucose, whose alditol acetate was not well resolved from the galactose derivative on the gas chromatogram. In other analyses, the ratio of galactose to mannose in untreated thrombin was not as high as shown in Ta- ble I, but it was still above the expected value of 2.0. The relative values for N-ace- tylglucosamine are not as accurate as those for the other components, presumably be- cause the hydrolysis conditions are not ideal for that sugar (19).
The values presented in Table I dem- onstrate the efficiency of the enzymes to remove the appropriate sugars. Neur- aminidase treatment of thrombin and an- tithrombin resulted in complete release of sialic acid. More than half of the galactose residues from asialothrombin, and over 80% of the galactose moieties from asialoantithrombin, were removed after digestion with P-galactosidase.
Based on the known carbohydrate struc- tures, two of the four residues of N-ace- tylglucosamine should be released by digestion with /3-N-acetylglucosaminidase. For thrombin, about half of these amino

362 ROSENFELD AND DANISHEFSKY
TABLE I
SUGAR ANALYSES OF DIALYZED SAMPLES
NeuNAc Man Gal GlcNAc
Digestion of thrombin” 0 I I + II I + II + III I + II + III + IV
Digestion of antithrombin” 0 I I + II I + II + III I + II + III + IV
2.0 0.0 NDb ND ND
3.0 3.0 3.0 3.0 0.5”
2.7 4.6 2.3 3.3 1.0 3.1 1.0 1.5 0.5c 0.7C
2.1 3.0 2.5 3.5 0.0 3.0 2.4 4.6 ND 3.0 0.4 3.7 ND 3.0 0.2 1.3 ND 8.4’ 0” 4.8”
Note. Molar ratios in man = 3.0. n 0 = no treatment, I = neuraminidase, II = P-galactosidase, III = @-N-acetylglucosaminidase, IV = endo-
P-N-acetylglucosaminidase D. *ND = not determined. ‘Residues per mole protein.
sugars were released by this digestion when compared to the N-acetylglucosa- mine content of the substrate. Less than half of the N-acetylglucosamine residues remained after P-N-acetylglucosaminidase treatment of the antithrombin substrate; this result may have been anomolous, pos- sibly due to incomplete hydrolysis.
Endo D is an endoglycosidase known to hydrolyze the ,f31 - 4 linkage between the N-acetylglucosamine residues of the core region of complex carbohydrate chains. The enzyme acts only after the outer-chain sugars have been removed (13). It was thus necessary to remove sialic acid, galactose, and outer-chain N-acetylglucosamine res- idues before digestion. The product of the endo D incubation would be expected to have no mannose and one N-acetylglucos- amine residue per chain. The measure- ments in Table I show that, after digestion of partially deglycosylated thrombin with endo D, about 17% of the mannose and almost one glucosamine moiety remained bound to the protein. Digestion of partially deglycosylated antithrombin with endo D resulted in very little digestion; however, in another experiment using an anti-
thrombin preparation that had lost activ- ity, about 80% of the mannose residues were removed by this digestion. Conceiv- ably, the interior region of some of the sugar chains of native antithrombin may be inaccessible to endo D (13), or too little enzyme may have been used (see Materials and Methods).
The compositions of the bound sugars, listed in Table I, are in good agreement with measurements of the sugars released after each digestion. A recent report (5) states that thrombin contains a partial residue of fucose. Under our conditions, no fucose was detected.
The products of the glycosidic digestions as well as the starting materials were elec- trophoresed in order to determine the effect of deglycosylation on migration in SDS gels (Fig. 1). The migration distance of the ma- jor band for each sample was measured and the apparent iV, values were estimated by comparison with standards (low-mo- lecular-weight standards kit, Bio-Rad). The observed masses were found to be close to those expected after removal of the ap- propriate sugars (Table II). Plots of the log mobility (relative to the dye front) ver-

EFFECTS OF DEGLYCOSLYATION OF HUMAN THROMBIN AND ANTITHRCMBIN 363
92.5 K-
45.OK-
Aniithrombm
Thrombin C
31.OK-
21.5K-
14.4K-
MW 0 I TREATMENT: sfds
II In m ;+;, 0 I II III m
Thrombin Antlthrombm
FIG. 1. Electrophoretic mobilities of glycosidase-treated thrombin and antithrombin. SDS-poly- acrylamide gel electrophoresis, as described under Materials and Methods. Treatment: 0, No treat- ment; I, neuraminidase digestion; II, I + P-galactosidase digestion; III, I + II + @-N-acetylglucos- aminidase digestion; IV, I + II + III + Endo D digestion. MW Stds, Bio-Rad low-molecular-weight standard kit, which is a mixture of proteins with the M, values indicated on the left side of the
TABLE II
CHANGES IN APPARENT M, CAUSED
BY DEGLYCOSYLATION
Digestion’ Observed Expected
Kb WC
Thrombin 0 I I + II I + II + III I + II + III + IV
36,500* 36,500 36,100 35,884 35,700 35,560 35,000 35,154 33,700 34,465
Antithrombin 0 I I + II I + II + III I + II + III + IV
58,000* 58,000 56,200 55,536 54,400 54,240 53,300 52,616 50,500 49,860
a See Footnote (a) of Table I. *Average of three determinations. Observed dif-
ferences in M, were averaged and subtracted from untreated M, to result in the reported values.
’ Calculated by subtracting appropriate masses from untreated M,.
* Assumed Mr.
sus the number of sugars removed gave a straight line, similar to that reported for the electrophoresis of partially deglyco- sylated fetuin glycopeptides (24).
Activities of deglycosylated throw&ins. The data presented in Table III show the fibrinogen-clotting, amidolytic, and plate- let-release activities of thrombin that re- mained after each digestion, compared to parallel controls. These values were also compared to untreated thrombin, and sim- ilar results were obtained. After each suc- cessive glycosidic digestion, there was no significant loss in the activities measured. The deglycosylated thrombins could also be completely inhibited by antithrombin. The ability of asialo- and asialoagalac- tothrombin to cause the platelet release reaction indicates that the thrombin de- rivatives can cause platelets to aggre- gate (26).
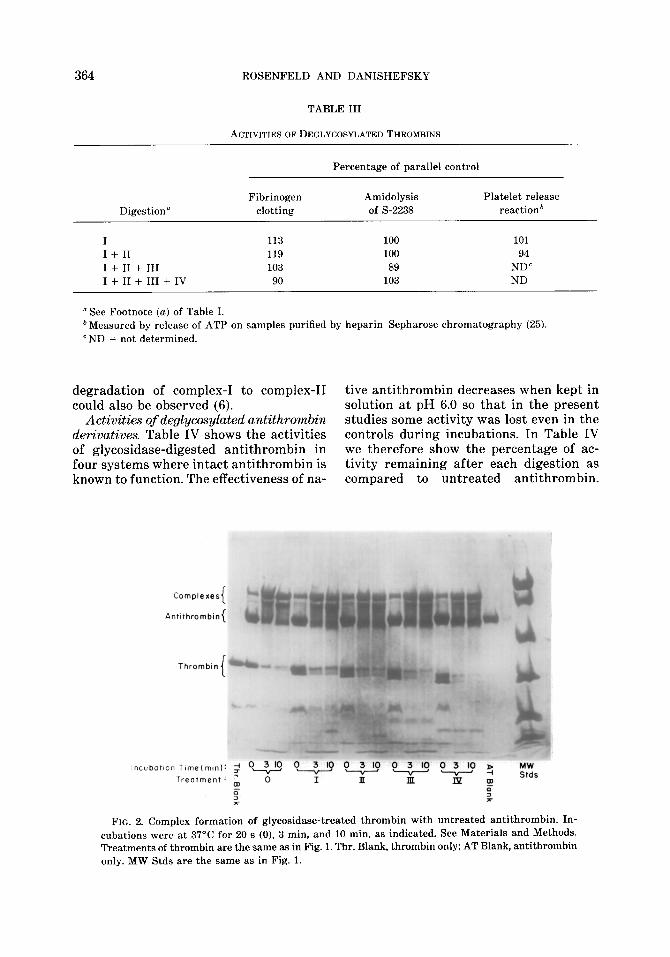
The result of incubating the thrombin derivatives with antithrombin is shown in Fig. 2. It is seen that, like thrombin, the deglycosylated thrombins form complexes with antithrombin. A 1:l molar ratio of thrombin to antithrombin was used so that
364 ROSENFELD AND DANISHEFSKY
TABLE III
ACTIVITIESOFDEGLYCOSYLATEDTHROMBINS
Percentage of parallel control
Digestion’ Fibrinogen
clotting Amidolysis of S-2238
Platelet release reaction*
I 113 100 101 I + II 119 100 94 I + II + III 103 89 ND” I + II + III + IV 90 103 ND
‘See Footnote (a) of Table I. b Measured by release of ATP on samples purified by heparin-Sepharose chromatography (25). “ND = not determined.
degradation of complex-I to complex-II tive antithrombin decreases when kept in could also be observed (6). solution at pH 6.0 so that in the present
Activities of deglycosylated antithrowzbin studies some activity was lost even in the derivatives. Table IV shows the activities controls during incubations. In Table IV of glycosidase-digested antithrombin in we therefore show the percentage of ac- four systems where intact antithrombin is tivity remaining after each digestion as known to function. The effectiveness of na- compared to untreated antithrombin.
A
!ncubotlo
Complexes
ntlthromban {
Thrombin
n Tlme(mln): ; ‘a ?-“- 3 IO 03 0 3 IO 0 3 IO 2
Treatment: m
0 I II i T m 0
2 % *
FIG. 2. Complex formation of glycosidase-treated thrombin with untreated antithromhin. In- cubations were at 37’C for 20 s (0), 3 min, and 10 min. as indicated. See Materials and Methods. Treatments of thromhin are the same as in Fig. 1. Thr. Blank, thrombin only; AT Blank, antithromhin only. MW Stds are the same as in Fig. 1.

EFFECTS OF DEGLYCOSLYATION OF HUMAN THROMBIN AND ANTITHROMBIN 365
TABLE IV
EFFECTS OF DEGLYCOSYLATION ON ANTITHROMBIN ACTIVITIES
Digestion”
Heparin activation of Inhibition of thromhin thrombin inhibition
(progressive) (heparin cofactor) Plate assay: thromhin- Plate assay:
Percentage of Percentage of S-2238 ’ Xa-S-2222’ parallel Percentage of parallel Percentage of (percentage of (percentage of control untreated control untreated untreated) untreated)
I 117 110 108 104 97 89 I + II 112 68 100 52 56 46 I + II + III 108 63 92 32 52 32 I + II + III + IV 122 40 1046 34 63 33
(L See Footnote (a) of Table I. bThe activity was compared to the sample from the previous step, rather than the parallel control, which became inactivated
during incubation. ‘These assays contained heparin (see Materials and Methods).
Comparisons to parallel controls are pre- of amidolytic activity of thrombin and Xa sented in the table only for the first two closely mimicked the loss found for the systems to show that the decrease in ac- parallel controls in the other activity as- tivity in these controls closely followed the says; it was concluded that this decrease loss in activity for the samples. The loss was not caused by the removal of sugars. in activity for heparin-mediated inhibition The electrophoretic pattern in Fig. 3
Complexes
Antithrombin 1
Thrombin I
lncubationTlme(min)~O - 3 0 3 03 03 0 3 5 MW
Treotment g 0 T‘if’TTg Stds
0 0 s :
FIG. 3. Complex formation of glycosidase-treated antithrombin with control thrombin. See Materials and Methods. Labels are the same as in Fig. 1 and Fig. 2. Incubations at 37°C were carried out for 20 s (0) and 3 min.

366 ROSENFELD AND DANISHEFSKY
shows that thrombin formed complexes with the enzyme-treated samples just as well as with untreated antithrombin. It should be noted that the electrophoretic mobilities of the complexes increased with increasing loss of sugar, as did the degly- cosylated antithrombin derivatives alone.
We conclude that removal of sugars from human antithrombin does not diminish its ability to inhibit thrombin or factor Xa activities. These findings agree with the previous observations for asialoanti- thrombin (11).
Cimclusim. In certain instances, cova- lently bound sugar moieties of other blood coagulation factors have been found to have a role in their biological functions. For example, desialylation of fibrinogen caused the resultant fibrin monomers to polymerize more rapidly (27). Removal or oxidation of penultimate galactose resulted in loss of activity of bovine factor V (28). No direct function for the carbohydrate of bovine factor VIII could be detected (29), whereas alterations of the sugar moieties of human factor VIII altered ristocetin co- factor activity (29-31). Modification of the carbohydrate of factor X to make it ra- dioactive did not affect its activation by the extrinsic system (32).
The present study represents an attempt to measure the effect of removing all but one of the sugars on each carbohydrate chain of thrombin and antithrombin. Al- though deglycosylation was not always complete, enough of the sugar chains were removed to have affected the activity if there was such a requirement. It is POS-
sible, but doubtful, that the remaining N- acetylglucosamine may have a role in these activities, or that other activities we have not measured are dependent on these sug- ars. The assays employed are represen- tative, however; they include proteolytic, amidolytic, and binding activities of thrombin. Similarly, with antithrombin, its inhibition of more than one protease was measured. Formation of complexes of antithrombin and thrombin was not af- fected by these digestions. Thus the car- bohydrate chains of these glycoproteins do
not have a direct role in the biological functions of thrombin and antithrombin.
ACKNOWLEDGMENTS
The authors thank Dr. John W. Fenton II, New York State Department of Health, Albany, N. Y., and Dr. M. Wickerhauser, American National Red Cross, Bethesda, Md., for generously supplying the thrombin and antithrombin, respectively. We also thank Dr. Thomas C. Detwiler and Dr. Richard J. Alexander, SUNY Downstate Medical Center, Brooklyn, N. Y. for platelet ATP release assays.
REFERENCES
1. ROSENFELD, L., AND DANISHEFSKY. I. (1983) Fed Proc. 42, 1860 (Abstract No. 600).
2. FENTON, J. W., FASCO, M. J., STACKROW, A. B., ARONSON, D. L., YOUNG, A. M., AND FINLAYSON,
J. S. (1977) J. BioL Chem 252,3587-3598.
3. OWEN, W. G., ESMON, C. T., AND JACKSON, C. M. (1974) J. BioL Chem. 249, 594-605.
4. MIZUOCHI, T., FUJII. J., KISIEL, W., AND KOBATA, A. (1981) J. B&hem 90. 1023-1031.
5. NILSSON, B., HORNE, M. K., AND GRALNICK, H. R. (1983) Arch. B&hem. Biophys. 224, 127-133.
6. ROSENBERG, R. D., AND DAMUS, P. S. (1973) J. BioL Chem. 248, 6490-6505.
7. MIZUOCHI, T., FUJII, J., KURACHI, K., AND KOBATA, A. (1980) Arch. B&hem. Biophys. 203,458-465.
8. FRANZEN, L.-E., SVENSSON, S., AND LARM, 0. (1980) J. BioL Chem 255,5090-5093.
9. SKAUG, K., AND CHRISTENSEN, T. B. (1971) B&him Biophys. Acta 230, 627-629.
10. NELSESTUEN, G. L., AND SUTTIE, J. W. (1971) B&hem. Biophys. Res. Commun. 45, 198-203.
11. DANISHEFSKY, I., ZWEBEN, A., AND SLOMIANY, B. (1978). J. BioL Chem. 253, 32-37.
12. DANISHEFSKY, I., KUHN, L., AND VILLANUEVA, G. B. (1981) Ann N. Y. Acud Sci 370,695-699.
13. KOIDE, N., AND MURAMATSU, T. (1974) J. BioL Chem. 249, 4897-4904.
14. LAKI, K. (1951) Arch. B&hem. Biophys. 32, 317- 321.
15. RADOFF, S., AND DANISHEFSKY, I. (1981) Throw&. Res. 22, 353-365.
16. ALEXANDER, R. J., FENTON, J. W., ANDDETWILER, T. C. (1983) Arch. B&hem. Biophys. 222,266- 275.
17. GERENDAS, M. (1959) Thromb. Lkzth. Huemorrh. 4, 56-70.
18. LAEMMLI, U. K. (1970) Nature (LolLdrnL) 227,680- 685.

EFFECTS OF DEGLYCOSLYATION OF HUMAN THROMBIN AND ANTITHROMBIN 367
19. READING, C. L., PENHOET, E. E., AND BALLOU,
C. E. (1978) J. Biol Chem. 253,5600-5612. 20. NIEDERMEIER, W. (1971) Anal. Biochem. 40,465-
475. 21. WARREN, L. (1959) J. Biol Chem. 234,19’71. 22. NORDENMAN, B., NYSTROM, C., AND BJORK, I. (1977)
Eur. J. Biochxm. 78,195-203. 23. LI, S.-C. MAZZOTTA, M. Y., CHIEN, S.-F., AND LI,
Y.-T. (1975) J. Biol. Chem. 250,6786-6791. 24. PORETZ, R. D., AND PIECZENIC, G. (1981) And
Biochem. 115, 170-176. 25. DANISHEFSKY, I., TZENG, F., AHRENS, M., AND
KLEIN, S. (1976) Thromb. Res. 8, 131-140.
26. CHARO, I. F., FEINMAN, R. D., AND DETWILER, T. C. (1977) J. Clin Invest. 60, 866-873.
27. MARTINEZ, J., PALASCAK, J., AND PETERS, C. (1977)
J. Lab. Clin. Med 89, 367-377.
28. SARASWATHI, S., AND COLEMAN, R. W. (1975) J.
Biol. Chem 250, 8111-8118.
29. ROSENFELD, L., AND KIRBY, E. P. (1979) Thrwmb. Res. 15, 255-261.
30. KAO, K.-J., PIZZO, S. V., AND MCKEE, P. A. (1980)
J. Biol Chem. 255, 1013410139.
31. GRALNICK, H. R., WILLIAMS, S. B., AND RICK,
M. E. (1983) Proc NC& Ad Sci USA 80,2771- 2774.
32. SILVERBERG, S. A., NEMERSON, Y., AND ZUR, M.
(1977) J. BioL Chem. 252, 8481-8488.





























