Embed Size (px)
Citation preview

Effects of Elevated CO2 and Pb on Phytoextractionand Enzyme Activity
Sunghyun Kim & Hojeong Kang
Received: 1 September 2010 /Accepted: 1 December 2010 /Published online: 22 December 2010# Springer Science+Business Media B.V. 2010
Abstract The wide-ranging impacts of globallyincreasing carbon dioxide (CO2) concentration andrising metal-contaminated soils are serious problemsin terrestrial ecosystems. In this study, we investigatedthe effects of elevated CO2 on the lead (Pb) uptake ofpine seedlings and the microbial activity in Pb-contaminated soil. Three-year-old pine seedlings wereexposed to ambient, as well as elevated levels of CO2
(380 and 760 ppmv, respectively) in 500 mg/kg Pb-contaminated soil. Growth rates, C/N ratios and Pbuptake of the pine seedlings were determined.Dissolved organic carbon (DOC) content and micro-bial activity were also measured in the rhizospheresoil. Elevated CO2 significantly increased the totalbiomass and accumulation of Pb in roots and shoots.In addition, the accumulation of Pb in the roots underelevated CO2 concentration was four times higherthan those in the roots under ambient CO2 concentra-tion. Elevated CO2 levels also affected C/N ratios inthe pine seedlings and soil enzyme activities. Declinein the overall nitrogen content and increases in the C/N ratios of pine needles were observed. Soil enzymeactivity increased in the rhizosphere soils, includingthose of β-glucosidases, N-acetylglucosaminidases,and phosphatases. Quality of the DOC was affected
by elevated CO2, while the quantity of DOC wasaffected by Pb additions under elevated CO2 con-ditions. Two major conclusions can be drawn fromthis study: (1) elevated CO2 significantly increasedbiomass and metal uptake of pine seedlings and (2)chemical metabolism on pine tissue and processes oforganic decomposition were more affected by elevat-ed CO2 levels than by Pb contamination.
Keywords Elevated CO2.Pinus densiflora . C/N
ratios . Enzyme activities . Lead contamination
1 Introduction
Ongoing combustion of fossil fuels leads to anincrease in the atmospheric carbon dioxide (CO2)concentration, which is contributing to global warm-ing substantially (IPCC 2007). It is well known thatthe elevation of CO2 in the atmosphere significantlyaffects global ecosystems (Woodward 2002). Numer-ous studies have investigated the effects of elevatedCO2 on a wide variety of plants and ecosystems(Bazzaz 1990; Ceulemans and Mosseau 1994;Ellsworth 1999; Freeman et al. 2004; Medlyn et al.2001; Gunderson et al. 2002; Naumburg et al. 2003;Bunce 2004; Marchi et al. 2004).
Some of those studies have shown that elevatedCO2 increases the growth rate of plants (Gifford1994; Naidu et al. 1998; King et al. 2001; Hamilton etal. 2002) and significantly affects the plant’s physio-
Water Air Soil Pollut (2011) 219:365–375DOI 10.1007/s11270-010-0713-5
S. Kim :H. Kang (*)School of Civil and Environmental Engineering,Yonsei University,134, Sinchondong, Seodaemun-gu,Seoul, South Koreae-mail: [email protected]

logical functions (Melillo et al. 1990). In addition, theelevation of CO2 in the atmosphere also causes anincrease in the C/N ratios of plant litters, eventuallyreducing N concentration (Berntson and Bazzaz 1996;Hartwig et al. 1996). Such results were due tochanges in the level of Rubisco or respiratoryproteins, or to the dilution of nitrogen resulting fromthe accumulation of nonstructural carbohydrates.Changes in primary production caused by elevatedCO2 levels could alter the microbial demand for Nand the flow of N between soil microorganisms andplant roots (Zak et al. 2000). The production andchemical constituents of plant litter could alsoinfluence the types of organic substrates availablefor microbial metabolism in the soil (Finzi et al.2006). This, in turn, may increase organic matterdecomposition and release inorganic nutrients forplant growth. These feedback loops would play animportant role in keeping the balance in ecosystemswith elevated CO2 levels (Rouhier and Read 1998).
Elevated CO2 can increase dissolved organiccarbon (DOC) in soils by increasing root exudates(Freeman et al. 2004). Increasing DOC can alsoincrease bioavailability of metals for plants andmicroorganisms in metal-contaminated soils (Davisand Leckie 1978). A survey of the literatureindicates that few studies have investigated theeffect of elevated CO2 on plant uptake of pollutantsfrom contaminated environments (Dowdall et al.2008). Among the heavy metal-contaminated soils,lead (Pb) is one of the most toxic ones (Chaney et al.1999). Exposure to Pb can result in seizures, mentalretardation or behavioral disorders (Wierzbicka1999). The toxicity of Pb in crops varies accordingto crop type (Greszta 1982). In addition, Pb mobilityis generally low because of its low solubility. Thelow solubility may be further reduced as a result ofits interactions with the soil’s solid phase forsorption and ion exchange (Glazovskaya 1994). Ina previous study, Indian mustard and sunflowerswere reported to absorb higher amounts of Cu underelevated CO2 levels (Tang et al. 2003). However,Guo et al. (2006) and Jia et al. (2007) reporteddecreasing accumulation of metals in rice grown oncontaminated soil under free-air CO2 enrichmentconditions. Therefore, the response of plant uptakeof metals with elevated CO2 levels is stronglydependent on the plant species itself.
While a few studies have reported on the effects ofelevated CO2 and metal contamination on plants,nothing has been stated about the responses of pineseedlings, even though Pinus sylvestris has beenwidely studied as a bioindicator to measure bothheavy metal and air pollution (Grodzinska andKazmierczakowa 1977; Yilmaz 2002). Because pinetrees are one of the dominant tree species in easternAsia, it is believed that information on the effects ofelevated CO2 on those trees is highly valuable.
We examined the effect of Pb concentration in thesoil and increases of CO2 in the air on root and shootgrowth, fresh biomass and C/N ratios of pine seed-lings (Pinus densiflroa). We also studied the effect onmicrobial activity in the soil from elevated CO2 andPb concentrations. The objectives of this study wereto (1) investigate the effect of elevated CO2 on Pbuptake by pine seedlings and (2) assess the impact ofincreased CO2 and Pb on the biomass (dry weight)and C/N ratios of pine seedlings, emphasizing theinterdependency of soil chemical and microbialproperties. This is believed to be the first report onthe effects of elevated CO2 levels on Pb uptake bypine trees grown in Pb-contaminated soils.
2 Materials and Methods
2.1 Experimental Design
Natural soil was sampled at depths of 5–15 cm fromthe campus pine forests of Ewha Womans Universityin Korea. The air-dried soil sample was mixed withsand (1:1 w/w), and then passed through a 2-mmsieve. For each experiment, aliquots (1 kg each in1.5-L plastic cylinders) of soil were artificiallycontaminated with 500 ppm Pb. Test pots containing1 kg of contaminated soil were planted with 3-year-oldpine seedlings, which were obtained from the Koreanforest service, then placed in a growth chamber(Dasol Scientific Co., Korea). The growth chamberwas controlled at CO2 concentrations of 380 or760 ppmv, and at 25 C, 60% humidity and subjectedto a 16-h light/8-h dark cycle for 3 months. The plantgrowth and microbial activities were comparativelyanalyzed in four types of soil samples (Fig. 1; Cont,CO2 380 ppmv+Pb 0 mg kg-1; Pb, CO2 380 ppmv+Pb 500 mg kg−1; Ele. CO2, CO2 760 ppmv+Pb
366 Water Air Soil Pollut (2011) 219:365–375

0 mg kg−1; Pb+Ele. CO2, CO2 760 ppmv+Pb500 mg kg−1). All tests were performed in triplicate.Also, 30-ml water and 20-ml 1/2 nutrient solution(KNO3, 606.66 mg L−1 ; Ca(NO3)2 ·4H2O,944.60 mg L−1; NH4H2PO4, 115.02 mg L−1; MgSO4,492.94 mg L−1; FeCl2·7H2O, 492.94 mg L−1;MnCl2·4H2O, 1.78 mg L−1; H3BO3, 2.84 mg L−1;ZnSO4·7H2O, 0.23 mg L−1; CuSO4·5H2O,0.075 mg L−1) were added to the soil once per week.Shoot and root dry weight, as well as C and Ncontent, of the plants were measured. After incuba-tion for 3 months, DOC and enzyme activities weremeasured after dividing into bulk and rhizospheresoil samples. The bulk soil was the soil remainingafter picking out the roots from the pot. Therhizosphere soil was the soil still adhered to theroots after gentle shaking.
2.2 Soil Characteristics
Soil pH was determined by adding soil to water at aratio of 1:5 (w/v). Soil moisture content was deter-mined gravimetrically by drying for 24 h at 105°C,and organic matter content was determined by loss onignition at 700°C in a furnace (MAS 7000, CEM,USA). Soil cation-exchange capacity (CEC) wasdetermined by Environmental Protection Agency9081 methods (EPA 1986). Soil nitrate was deter-mined by extraction with deionized water andmeasured with a nitrate electrode (Gelderman and
Beegle 1998). Also, NH4+ was determined by
indophenol blue methods (Dorich and Nelson 1983).
2.3 Pine Growth and C/N Ratios
The pine seedlings were planted and placed in agrowth chamber that was controlled to 25°C, 60%humidity, and 16-h light/8-h dark cycles. Every4 weeks, the plants were measured for shoot lengthfor a total of 3 months (data not shown). After3 months, plants were carefully harvested by gentlyremoving them from the soil and measurements forshoot and root lengths, and biomass (dry weight) weretaken. Percent dry weight nitrogen and carbon wereestimated from leaf and root powder on a Frash EA1112 model analyzer (Thermo Electron Corporation,USA).
2.4 Absorption and Uptake of Pb by Pine Seedlings
To test whether pine seedlings remove Pb from soil,we measured the soil Pb content after 3 months.The concentrations of total, exchangeable andsoluble Pb in the soil samples were measured. Soilsamples were dried at room temperature andanalyzed for water-soluble Pb by equilibrating 1 gof soil with 20 ml of 0.01 M KNO3 for 2 h. Theamount of exchangeable Pb in the soil was estimatedby extracting 1 g soil with 20 ml 1 N NH4COOH for1 h. For total Pb in the soil, 0.5-g soil was extractedwith 2.4-ml aqua regia (35% HCl 1.8 ml+65%HNO3·0.6 ml) in an MDS-2000 automatic micro-wave digester (CEM). The extracted Pb of each soilsample was analyzed by an atomic absorptionspectrophotometer (AAS Analysis 100, PerkinElmer, Germany), which was calibrated using certi-fied reference materials (MESS-2 Marine Sediment,National Research Council of Canada).
To determine the amount of Pb in the plants, roots,and shoots were further separated with scissors andthen dried in an oven at 70°C for 24 h. Prior toanalysis, plants were washed with water to removesoil particles. Plant samples were digested in conc.HNO3 in an MDS-2000 automatic microwave digest-er (CEM). Pb content was then determined with anatomic absorption spectrophotometer (AAS Analysis100, Perkin Elmer, Germany), which was calibratedusing certified reference materials (SRM 1575,
Fig. 1 Experimental design and measurements
Water Air Soil Pollut (2011) 219:365–375 367

National Institute of Standard Technology, USA). Thepercentage recovery for the metal of interest was92.6%.
2.5 Analysis of DOC
DOC concentrations were measured using varioustechniques. Soil DOC was determined by adding soilto a slurry of water at a ratio of 1:10 w/v and analyzedusing a TOC-5000 (Shimadzu). In addition, specificUV absorbance (SUVA) was determined, which canindicate the nature or quality of DOC in a givensample and has been used as a surrogate measurementof DOC aromaticity (Chin et al. 1994). SUVA valuesat 254 nm (SUVA254) are used for specific UVabsorbance, because natural organic matter absorbsstrongly at this wavelength, increasing sensitivity andcreating a strong correlation with the aromatic carboncontent of organic matter.
2.6 Analysis of Soil Microbial Activities
Four extracellular enzyme activities were measuredby the MUF-substrate method (Freeman et al.1996), and β-glucosidase, N-acetylglucosaminidase,phosphatase, and arylsulfatase activity was analyzed.The concentration of each substrate solution was400 μM (SIGMA; MUF-β-gludoside, MUF-N-ace-tylglucosamine, MUF-arylsulfate), except for phos-phate, which was 800 μM (SIGMA; MUF-phosphate). Enzyme activities were measured fromthe slurry of soil and substrate solutions (1:5) by afluorometer.
Dehydrogenase activity and intracellular activitywere measured by an INT assay (Tabatabai 1982).The substrate used for these assays was 2-[4-iodophenyl]-3-[4-nitrophenyl]-5-phenyltetrazolium
chloride. The mixtures (3 g of fresh soil) wereincubated for 24 h at 37°C. Reaction products weredetected using a spectrophotometer (DR/3000 Spec-trophotometer, HACH) at 485 nm.
2.7 Statistical Analysis
Data were analyzed using SPSS 9.0 software forTukey’s test after two-way ANOVA. The test wascarried out on the soil’s physical and chemicalparameters, including root and shoot length, C/Nratios, soil enzyme activity, DOC, and SUVA data todetermine the effects of CO2 levels and Pb contam-ination. The standard deviations are shown withnumerals in Tables 1, 2, 3, 4, and 5 and with errorbars in Figs. 2 and 3.
3 Results
3.1 Comparison of Physical and Chemical SoilCharacteristics
The physicochemical characteristics of the soils withelevated CO2 and Pb contamination are listed inTable 1. Under elevated CO2 levels, soil pHdecreased, while CEC increased. However, soilmoisture content and organic matter did not exhibitany significant changes with elevated CO2 and Pbcontamination. The concentration of NO3
− increasedwith elevated CO2 in the uncontaminated soils, butwas not different with elevated CO2 in the contami-nated soils. While the amount of NH4
+, the source ofthe microbial activity, slightly decreased after3 months, there was not a significant difference inconcentration between elevated CO2 soils and Pb-contaminated soils.
Table 1 The physicochemical characteristics of the soils
Treatment pH (1:5) Moisture content (%) OM (%) CEC NO3− (mg/kg) NH4
+ (mg/g)
Control 6.7±0.1 a 10.0±2.5 2.6±0.2 9.0±0.4 a 50±0.08 a 0.04±0.03
Pb 6.6±0.1 a 10.2±2.3 2.8±0.2 8.8±0.4 a 40±0.02 b 0.05±0.03
Ele. CO2 5.6±0.2 b 8.7±1.7 2.7±0.2 10.2±0.5 b 60±0.03 c 0.05±0.03
Pb±Ele. CO2 5.9±0.1 b 13.1±4.7 3.2±0.9 10.2±0.5 b 40±0.01b 0.07±0.01
Values are the mean±SD. Values represent the mean of three replicates. Values between each column followed by the same letter donot differ significantly (two-way ANOVA followed by Tukey’s test)
368 Water Air Soil Pollut (2011) 219:365–375
3.2 The Effects on Biomass and C/N Ratiosof the Pine Seedlings
The results showed that root elongation and biomassof pine seedlings increased under elevated CO2
(Table 2). Although the shoot elongation of the pineseedling was not significantly affected by elevatedCO2 in Pb-contaminated soil, the total dry weights inboth contaminated soils under elevated CO2 weresignificantly greater than those grown under ambientCO2 (p<0.05).
Table 3 shows the C/N ratios of the roots andleaves of pine seedlings after 3 months, whichmeasured chemical changes in pine seedlings causedby elevated CO2 and Pb contamination. C/N ratios inleaves slightly increased with elevated CO2, whichshows that carbon concentrations increased, althoughnitrogen concentration decreased with elevated CO2.However, C/N ratios in the roots were not affected byelevated CO2 and Pb contamination.
3.3 Absorption and Uptake of Pb from Soil by a PineSeedling
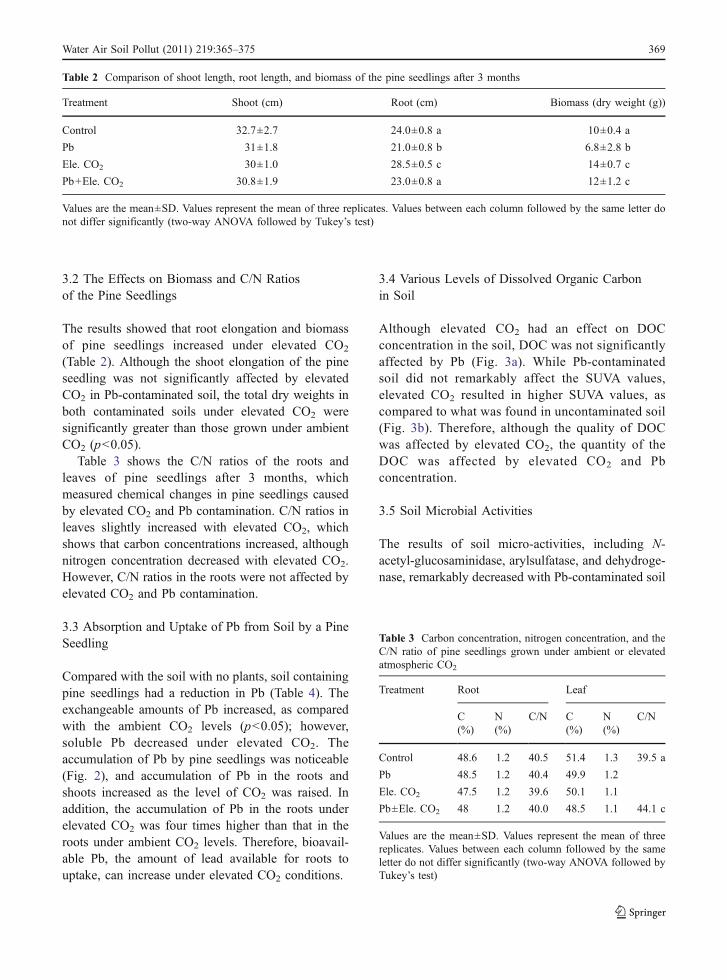
Compared with the soil with no plants, soil containingpine seedlings had a reduction in Pb (Table 4). Theexchangeable amounts of Pb increased, as comparedwith the ambient CO2 levels (p<0.05); however,soluble Pb decreased under elevated CO2. Theaccumulation of Pb by pine seedlings was noticeable(Fig. 2), and accumulation of Pb in the roots andshoots increased as the level of CO2 was raised. Inaddition, the accumulation of Pb in the roots underelevated CO2 was four times higher than that in theroots under ambient CO2 levels. Therefore, bioavail-able Pb, the amount of lead available for roots touptake, can increase under elevated CO2 conditions.
3.4 Various Levels of Dissolved Organic Carbonin Soil
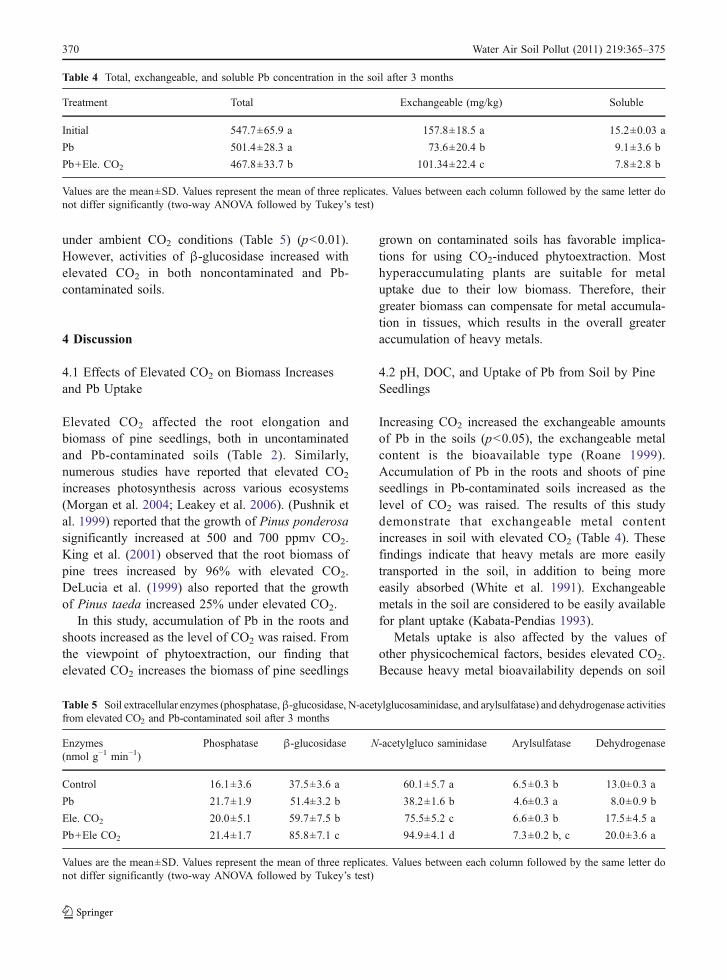
Although elevated CO2 had an effect on DOCconcentration in the soil, DOC was not significantlyaffected by Pb (Fig. 3a). While Pb-contaminatedsoil did not remarkably affect the SUVA values,elevated CO2 resulted in higher SUVA values, ascompared to what was found in uncontaminated soil(Fig. 3b). Therefore, although the quality of DOCwas affected by elevated CO2, the quantity of theDOC was affected by elevated CO2 and Pbconcentration.
3.5 Soil Microbial Activities
The results of soil micro-activities, including N-acetyl-glucosaminidase, arylsulfatase, and dehydroge-nase, remarkably decreased with Pb-contaminated soil
Table 2 Comparison of shoot length, root length, and biomass of the pine seedlings after 3 months
Treatment Shoot (cm) Root (cm) Biomass (dry weight (g))
Control 32.7±2.7 24.0±0.8 a 10±0.4 a
Pb 31±1.8 21.0±0.8 b 6.8±2.8 b
Ele. CO2 30±1.0 28.5±0.5 c 14±0.7 c
Pb+Ele. CO2 30.8±1.9 23.0±0.8 a 12±1.2 c
Values are the mean±SD. Values represent the mean of three replicates. Values between each column followed by the same letter donot differ significantly (two-way ANOVA followed by Tukey’s test)
Table 3 Carbon concentration, nitrogen concentration, and theC/N ratio of pine seedlings grown under ambient or elevatedatmospheric CO2
Treatment Root Leaf
C N C/N C N C/N(%) (%) (%) (%)
Control 48.6 1.2 40.5 51.4 1.3 39.5 a
Pb 48.5 1.2 40.4 49.9 1.2
Ele. CO2 47.5 1.2 39.6 50.1 1.1
Pb±Ele. CO2 48 1.2 40.0 48.5 1.1 44.1 c
Values are the mean±SD. Values represent the mean of threereplicates. Values between each column followed by the sameletter do not differ significantly (two-way ANOVA followed byTukey’s test)
Water Air Soil Pollut (2011) 219:365–375 369
under ambient CO2 conditions (Table 5) (p<0.01).However, activities of β-glucosidase increased withelevated CO2 in both noncontaminated and Pb-contaminated soils.
4 Discussion
4.1 Effects of Elevated CO2 on Biomass Increasesand Pb Uptake
Elevated CO2 affected the root elongation andbiomass of pine seedlings, both in uncontaminatedand Pb-contaminated soils (Table 2). Similarly,numerous studies have reported that elevated CO2
increases photosynthesis across various ecosystems(Morgan et al. 2004; Leakey et al. 2006). (Pushnik etal. 1999) reported that the growth of Pinus ponderosasignificantly increased at 500 and 700 ppmv CO2.King et al. (2001) observed that the root biomass ofpine trees increased by 96% with elevated CO2.DeLucia et al. (1999) also reported that the growthof Pinus taeda increased 25% under elevated CO2.
In this study, accumulation of Pb in the roots andshoots increased as the level of CO2 was raised. Fromthe viewpoint of phytoextraction, our finding thatelevated CO2 increases the biomass of pine seedlings
grown on contaminated soils has favorable implica-tions for using CO2-induced phytoextraction. Mosthyperaccumulating plants are suitable for metaluptake due to their low biomass. Therefore, theirgreater biomass can compensate for metal accumula-tion in tissues, which results in the overall greateraccumulation of heavy metals.
4.2 pH, DOC, and Uptake of Pb from Soil by PineSeedlings
Increasing CO2 increased the exchangeable amountsof Pb in the soils (p<0.05), the exchangeable metalcontent is the bioavailable type (Roane 1999).Accumulation of Pb in the roots and shoots of pineseedlings in Pb-contaminated soils increased as thelevel of CO2 was raised. The results of this studydemonstrate that exchangeable metal contentincreases in soil with elevated CO2 (Table 4). Thesefindings indicate that heavy metals are more easilytransported in the soil, in addition to being moreeasily absorbed (White et al. 1991). Exchangeablemetals in the soil are considered to be easily availablefor plant uptake (Kabata-Pendias 1993).
Metals uptake is also affected by the values ofother physicochemical factors, besides elevated CO2.Because heavy metal bioavailability depends on soil
Table 4 Total, exchangeable, and soluble Pb concentration in the soil after 3 months
Treatment Total Exchangeable (mg/kg) Soluble
Initial 547.7±65.9 a 157.8±18.5 a 15.2±0.03 a
Pb 501.4±28.3 a 73.6±20.4 b 9.1±3.6 b
Pb+Ele. CO2 467.8±33.7 b 101.34±22.4 c 7.8±2.8 b
Values are the mean±SD. Values represent the mean of three replicates. Values between each column followed by the same letter donot differ significantly (two-way ANOVA followed by Tukey’s test)
Table 5 Soil extracellular enzymes (phosphatase,β-glucosidase, N-acetylglucosaminidase, and arylsulfatase) and dehydrogenase activitiesfrom elevated CO2 and Pb-contaminated soil after 3 months
Enzymes(nmol g−1 min−1)
Phosphatase β-glucosidase N-acetylgluco saminidase Arylsulfatase Dehydrogenase
Control 16.1±3.6 37.5±3.6 a 60.1±5.7 a 6.5±0.3 b 13.0±0.3 a
Pb 21.7±1.9 51.4±3.2 b 38.2±1.6 b 4.6±0.3 a 8.0±0.9 b
Ele. CO2 20.0±5.1 59.7±7.5 b 75.5±5.2 c 6.6±0.3 b 17.5±4.5 a
Pb+Ele CO2 21.4±1.7 85.8±7.1 c 94.9±4.1 d 7.3±0.2 b, c 20.0±3.6 a
Values are the mean±SD. Values represent the mean of three replicates. Values between each column followed by the same letter donot differ significantly (two-way ANOVA followed by Tukey’s test)
370 Water Air Soil Pollut (2011) 219:365–375
properties, such as pH, organic matter, and CEC(Cornelis 2008), it is very important to investigate thechemical properties of each soil sample. ElevatedCO2 might lower soil pH, favoring the release ofmetals into the soil, and, as a result, helps the plant totake up more metals (Tang et al. 2003; Wu et al.2009). In this study, soil pH decreased significantlywith elevated CO2 (Table 1), which appeared toincrease bioavailability of Pb and, as a result, uptakeby the plants. In other words, elevated CO2 mightincrease root exudation, resulting in changes inrhizosphere pH and metal availability. These findingsare in accordance with the results of Tang et al.(2003), who reported that elevated CO2 triggeredhyperaccumulation of Cu by Brassica juncea andHelianthus annuus when grown on Cu-contaminatedsoil. Lieffering et al. (2004) reported that Zn and Mnshowed a strong tendency to increase in concentrationin rice grains with elevated CO2.
Under elevated CO2 conditions, DOC levelsincreased (Fig. 3a). DOC is, to a large extent,responsible for enhanced leaching of already adsorbedmetals from sorbents, sediments, and soil, since DOCis poorly adsorbed by solid materials and, thus, moveswith water flow (Herngren et al. 2005; Liu et al.2008). DOC metal complexes increase the mobility ofheavy metals and their availability for uptake by aplant’s roots (Antoniadis and Alloway 2002; Chenand Zhu 2006). DOC released from plant rootsincludes various molecules with acidic groups, suchas carboxyl and phenolic OH functional groups(Hofrichter and Fakoussa 2001), and hence providesorganic macromolecules with important roles in thetransport, bioavailability and solubility of heavy
metals (Lagier et al. 2000). It has been demonstratedthat such phenolic materials contribute to reducing thephysical mobility of various metal species in the soil.
SUVA values increased under elevated CO2 levelsin uncontaminated soil, but SUVA values decreasedunder elevated CO2 in Pb-contaminated soils(Fig. 3b). These results may be related to the factthat higher phenolics under elevated CO2 may bind toPb and be contained in plant biomass. When a plant isexposed to elevated CO2, extra-phenolics producedmay be released into the soil. However, when soil isexposed both to elevated CO2 and Pb contamination,extra-phenolics are bound to Pb and may not bereleased into the soil, resulting in lower SUVA values.
However, the limited data derived from plantsgrowing in pots are not broad enough to understandthe mechanisms involved in responses to elevatedCO2 among plant varieties. Therefore, more researchneeds to be done to investigate the relationshipbetween elevated CO2 and the increased bioavailabil-ity of metals under elevated CO2.
DO
C (
mg
g-1)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18(A)
aa
bb
SU
VA
0.000
0.002
0.004
0.006
0.008
0.010
0.012
0.014 (B)
a a
b
a
Control Pb Ele CO Pb + Ele CO22
Control Pb Ele CO Pb + Ele CO22
Fig. 3 DOC concentration (a) and SUVA (b) of rhizosphericsoils after incubation for 3 months
Pb
(µg/
g)
0
100
200
300
400
500
rootshoot leaf
Control Pb Ele CO Pb+Ele CO22
Fig. 2 Pb accumulation in a leaf, shoot, and root of pineseedlings
Water Air Soil Pollut (2011) 219:365–375 371

4.3 Effect of Soil Characteristics on C/N Ratiosin Pine Seedlings
Elevated CO2 affected the C/N ratios of pine seed-lings. In the leaf litter, the total nitrogen decreasedwith increasing CO2 but was not significantly affectedby Pb. These results may be related to the nitrogencontent of the soils (Table 1). The NO3
- concentrationincreased with elevated CO2 in the uncontaminatedsoils, but elevated CO2 had a different effect on thecontaminated soils. Elevated CO2 could likely drive Ninto the soil (Table 1; NO3
−), both of carbon andnitrogen is necessary for plant and microbial decom-position functions. Thus, when the amount of Navailable for plant uptake was decreased, C/N ratiosin the pine needles increased. Gifford et al. (2000)also reported similar results, that C/N ratios in pineseedlings increased 15%, because N content in theneedles decreased 21% when CO2 was doubled.
4.4 Effects of Elevated CO2 and Pb on MicrobialActivities
The microbial activities, including β-glucosidase, N-acetylglucosaminidase, arylsulfatase, and dehydroge-nase, increased under elevated CO2 (Table 5). How-ever, these activities decreased with Pb contaminationunder ambient CO2 levels. Several rhizosphere bacte-rial species are known to produce compounds, such asphytohormones, antifungal molecules, or sidero-phores, or to help the plant with atmospheric nitrogenfixation (Matthias et al. 1997). Increases in theactivity of N-acetylglucosaminidase often occur inresponse to nitrogen additions (Gifford et al. 2000).N-acetylglucosaminidase is a soil enzyme involved inchitin degradation, and a significant increase wasobserved in the activity of this enzyme under elevatedCO2. Similar results were found in alpine grassland,in which N-acetylglucosaminidase activity underelevated CO2 increased by nearly 30% (Mayr et al.1999). However, Kang et al. (2005) reported thatresponses of soil enzymes to elevated CO2 aredependent on the nutrient availability of soils andcan vary in different wetland ecosystems.
Enhanced enzyme activities under elevated CO2 werealso reported by Ross et al. (1995) in a short-termchamber experiment exposed to elevated CO2. This wasexplained by a greater production of microbes inresponse to increased C input. It has been shown that,
under elevated CO2, β-glucosidase releases more Cfrom organic matter into the soil (Henry et al. 2005;Larson et al. 2002). In this study, β-glucosidaseincreased with increases in CO2, with both theuncontaminated soil and the contaminated soil.
The effects of soil pollution on enzyme activitiesare complex. The response of different enzymes to thesame pollutant may vary greatly, and the sameenzyme may respond differently to different pollu-tants He et al. (2003). In this study, Pb inhibited N-acquiring enzyme activity (N-acetylglucoamidase),while Pb enhanced organic C-acquiring enzymeactivity (β-glucosidase) under ambient CO2. It is wellknown that different metal ions show differentbehaviors in their ability to act as inhibitors of β-glucosidase (Scigelova and Crout 1999). The effectsof metals on enzyme activities remain unclear. In thisstudy, β-glucosidase and N-acetylglucoamidase in-creased with elevated CO2 in Pb-contaminated soils.The adverse effect was explained by Ekenler andTabatabai (2002): metal ions may inactivate enzymesby reacting with sulfydryl groups of enzymes to formmetal sulfides Scigelova and Crout (1999) Sulfydrylgroups in enzymes may serve as integral parts of thecatalytic active sites or as groups involved inmaintaining correct structural relationships of enzymeproteins. Microbial hydrolases are important fororganic cycles, and inhibition and activation effectsmay influence organic C and N decomposition.Therefore, under elevated CO2, C and N contents,which are important for microbial metabolism, wouldbe released into the soil environment.
Our results suggest that a CO2 increase in thefuture may interfere with efficiency of phytoremedia-tion by influencing DOC quantity and quality. Thismay has a further implication for the current technol-ogy in a way that efficiency of phytoremediationcould be enhanced by artificial CO2 fertilization.Previous studies have reported that performance ofphytoremediation could be enhanced by nutrientfertilizations (Liao et al. 2007; Hutchinson et al.2001), and CO2 fertilization could be another way toachieve higher efficiency of phytoremediation.
5 Conclusions
Elevated CO2 increased Pb uptake and biomass ofpine trees in Pb-contaminated soils by increasing the
372 Water Air Soil Pollut (2011) 219:365–375

bioavailability of Pb, due to decreasing pH andincreasing concentration of DOC. In addition, chem-ical metabolism on pine tissue and processes oforganic decomposition were more affected by elevat-ed CO2 than by Pb contamination. The elevation ofCO2 in the atmosphere increased the C/N ratios of thepine litter, due to reduced soil N availability. Theactivities of soil enzymatic activities in the rhizo-sphere increased under elevated CO2. In this study,nitrogen changes in the soil explained the elevatedCO2 effects on microbial activities and pine seedlings.However, this work was limited to a growth chamber,and further investigation is needed in field conditions.In addition, more research should be conducted toclarify mechanism of phytoextraction under elevatedCO2. The results of this study demonstrated thatelevated CO2 and Pb had significant effects on boththe growth of pine seedlings and soil microbialactivity.
Acknowledgments H. Kang is grateful to NRF (2009-0079549), EcoRiver, and EcoSTAR for their financial support.
References
Antoniadis, V., & Alloway, B. J. (2002). The role of dissolvedorganic carbon in the mobility of Cd, Ni, and Zn insewage sludge-amended soils. Environment Pollution,117, 515–521.
Bazzaz, F. A. (1990). The response of natural ecosystems to therising global CO2, levels. Annual Review of Ecology andSystematics, 21, 167–196.
Berntson, G. M., & Bazzaz, F. A. (1996). Belowgroundpositive and negative feedbacks on CO2 growth enhance-ment. Plant and Soil, 187, 110–131.
Bunce, J. A. (2004). Carbon dioxide effects on stomatalresponses to the environment and water use by cropsunder field conditions. Oecologia, 140, 1–10.
Ceulemans, R. M., & Mosseau, M. (1994). Effects of elevatedatmospheric CO2 on woody plants Tansley Review no. 71.New Phytology, 127, 425–446.
Chaney, R. L., Li, Y.M., Angle, J. S., Baker, A. J.M., Reeves, R. D.,Brown, S. L., et al. (1999). Phytoremediation of ContaminatedSoil and Water. In Improving metal-hyperaccumulators wildplants to develop commercial phytoremediation systems:approaches and progress. Boca Raton: CRC.
Chen, B., & Zhu, Y. G. (2006). Humic acids increase thephytoavailability of Cd and Pb to wheat plants cultivatedin freshly spiked, contaminated soil. Journal of SoilsSediment, 6, 236–242.
Chin, Y., Aiken, G., & O’Loughlin, E. (1994). Molecularweight, polydispersity, and spectroscopic properties ofaquatic humic substances. Environmental Science andTechnology, 28, 1853–1858.
Cornelis, A. M. G. (2008). Physico-chemical and biologicalparameters determine metal bioavailability in soils. TheScience of the Total Environment, 406, 385–395.
Davis, J. A., & Leckie, J. O. (1978). Effect of absorbedcomplexing ligands on trace metal uptake by hydrousoxides. Environmental Science and Technology, 12, 1309–1315.
DeLucia, E. H., Hamilton, J. G., Naidu, S. L., Thomas, R. B.,Andrews, J. A., Finzi, A., et al. (1999). Net primaryproduction of a forest ecosystem with experimental CO2
enrichment. Science, 14(184), 1177–1179.Dorich, R. A., & Nelson, D. W. (1983). Direct measurement of
ammonium in potassium chloride extracts of soils. SoilScience Society of America Journal, 47, 833–836.
Dowdall, M., Standring, W., Shaw, G., & Strand, P. (2008).Will global warming affect soil-to-plant transfer of radio-nuclides? Journal of Environmental Radioactive, 99,1736–1745.
Ekenler, M., & Tabatabai, M. A. (2002). Effects of traceelements on β-glucosamidase activity in soils. SoilBiology and Biochemistry, 34, 1829–1832.
Ellsworth, D. S. (1999). CO2 enrichment in a maturing pineforest: are CO2 exchange and water status in the canopyaffected? Plant, Cell & Environent, 22, 461–472.
EPA, U. S. (1986). Test methods for evaluating solid waste.SW-846, method 9081. Washington DC: EnvironmentalProtection Agency.
Finzi, A. C., Moore, D. J. P., DeLucia, E. H., Lichter, J.,Hofmockel, K. S., Jackson, R. B., et al. (2006). Progres-sive nitrogen limitation of ecosystem processes underelevated CO2 in a warm-temperate forest. Ecology, 87, 15–25.
Freeman, C., Liska, G., Ostle, N. J., Lock, M. A., Reynolds, B.,& Hudson, J. (1996). Microbial activity and enzymicdecomposition processes following peatland water tabledrawdown. Plant and Soil, 180, 121–127.
Freeman, C., Fenner, N., Ostle, N. J., Kang, H., Dowrick, D. J.,& Reynolds, B. (2004). Dissolved organic carbon exportfrom peatlands under elevated carbon dioxide levels.Nature, 430, 195–198.
Gelderman, R. H., & Beegle, D. (1998). Nitrate–nitrogen.Recommended chemical soil test procedures for the NorthCentral Region. North Central Regional Research Publi-cation No. 221 (Revised). Columbia: Missouri AgriculturalExperiment Station.
Gifford, R. M. (1994). The global carbon cycle: a viewpoint onthe missing sink. Australian journal of Plant Physiology,21, 1–15.
Gifford, R. M., Barrett, D. J., & Lutze, J. L. (2000). The effectsof elevated [CO2] on the C/N and C: P mass ratios of planttissues. Plant and Soil, 224, 1–14.
Glazovskaya, M. A. (1994). Criteria for classification of soilsaccording to lead-pollution risk. Eurasian Soil Science, 26,58–74.
Greszta, J. (1982). Correlation between content of copper, zinc,lead and cadmium in the soil and the content of thesemetals in the seedlings of selected forest tree species.Fragment Florist Geo-botany, 28, 29–52.
Grodzinska, L., & Kazmierczakowa, R. (1977). Heavy metalcontent in the plants of cracow parks. Bulletin AcademyPolicy Science Ser Biology, 25, 227–234.
Water Air Soil Pollut (2011) 219:365–375 373

Gunderson, C. A., Sholtis, J. D., Wullshleger, S. D., Tissue, D.T., Hanson, P. J., & Norby, R. J. (2002). Environmentaland stomatal control of photosynthetic enhancement in thecanopy of a sweetgum (Liquidambar styraciflua L.)plantation during 3 years of CO2 enrichment. Plant, Cell& Environment, 25, 379–393.
Guo, H. Y., Jia, H. X., Zhu, J. G., & Wang, X. R. (2006).Influence of the environmental behavior and ecologicaleffect of cropland heavy metal contaminants by CO2
enrichment in atmosphere. Chinese Journal of Geochem-istry, 25, 212.
Hamilton, J. G., DeLucia, D. H., George, K., Naidu, S. L.,Finzi, A. C., & Schlesinger, W. H. (2002). Forest carbonbalance under elevated CO2. Oecologia, 131, 250–260.
Hartwig, U. A., Zanetti, S., Hebelson, T., Lüscher, A., Frehner,M., Fischer, B., et al. (1996). Symbiotic nitrogen fixation:one key to understanding the response of temperategrassland ecosystems to elevated CO2? In C. Kömer &F. Bazzaz (Eds.), Carbon dioxide, populations, communi-ties (pp. 253–264). San Diego: Academic.
He, Z. L., Yang, X. E., Baligar, V. C., & Calvert, D. V. (2003).Microbiological and biochemical indexing systems forassessing acid soil quality. Advances in Agronomy, 78, 89–138.
Henry, H., Juarez, J. D., Field, C. B., & Vitousek, P. M. (2005).Interactive effects of elevated CO2, N deposition andclimate change on extracellular enzyme activity and soildensity fractionation in a California annual grassland.Global Change Biology, 11, 1808–1815.
Herngren, L., Goonetilleke, A., & Ayoko, G. A. (2005).Understanding heavy metal and suspended solidsrelationships in urban stormwater using simulatedrainfall. Journal of Environmental Management, 76,149–158.
Hofrichter, M., & Fakoussa, R. (2001). Microbial degradationand modify cation of coal. In A. Steinbüchel & M.Hofrichter (Eds.), Lignin, humic substances and coal vol 1(pp. 393–427). Weinheim: Wiley.
Hutchinson, S. L., Banks, M. K., & Schwab, A. P. (2001).Phytoremediation of aged petroleum sludge: effect ofinorganic fertilizer. Journal of Environmental Quality, 30,395–403.
IPCC. (2007). Climate Change 2007: the physical sciencebasis: Contribution of Working Group I to the FourthAssessment Report of the Intergovernmental Panel onClimate Change. Intergovernmental Panel on ClimateChange. New York: Cambridge University Press.
Jia, H. X., Guo, H. Y., Yin, Y., Wang, Q., Sun, Q., Wang, X. R.,et al. (2007). Responses of rice growth to copper stressunder free-air CO2 enrichment (FACE). Chinese ScienceBulletin, 52, 2636–2641.
Kabata-Pendias, A. (1993). Behavioural properties of tracemetals in soils. Applied Ecochemistry, 2, 3–9.
Kang, H. J., Kim, S. Y., Fenner, N., & Freeman, C. (2005).Shift of soil enzyme activities in wetlands exposed toelevated CO2. The Science of the Total Environment, 337,207–212.
King, J. W., Mohamed, A., Taylor, S. L., Mebrahtu, T., & Paul,C. (2001). Supercritical fluid extraction of Vernoniagalmensis seeds. Industrial Crops and Products, 14,241–249.
Lagier, T., Feuillade, G., & Matejka, G. (2000). Interactionsbetween copper and organic macromolecules: determina-tion of conditional complexation constants. Agronomie,20, 537–546.
Larson, J. L., Zak, D. R., & Sinsabaugh, R. L. (2002).Extracellular enzyme activity beneath temperate treesgrowing under elevated carbon dioxide and ozone. SoilBiology and Biochemistry, 66, 1848–1856.
Leakey, A. D. B., Bernacchi, C. J., Ort, D. R., & Long, S. P.(2006). Longterm growth of soybean at elevated [CO2]does not cause acclimation of stomatal conductance underfully open-air conditions. Plant, Cell & Environment, 29,1794–1800.
Liao, X. Y., Chen, T. B., Yan, X. L., Xie, H., Yan, X. L., Zhai,L. M., et al. (2007). Selecting appropriate forms ofnitrogen fertilizer to enhance soil arsenic removal byPteris vittata: a new approach in phytoremediation.International Journal of Phytoremediation, 9, 269–280.
Lieffering, M., Kim, H. Y., Kobayashi, K., & Okada, M.(2004). The impact of elevated CO2 on the elementalconcentrations of field-grown rice grains. Field CropsResearch, 88, 279–286.
Liu, Y., Li, Y., Li, X., & Jiang, Y. (2008). Leaching behavior ofheavy metals and PAHs from MSWI bottom ash in a long-term static immersing experiment. Waste Management, 28,1126–1136.
Marchi, S., Tognetti, R., Vaccari, F. P., Lanini, M., Kaligaric,M., Miglietta, F., et al. (2004). Physiological and morpho-logical responses of grassland species to elevated atmo-spheric CO2 concentrations in FACE-systems and naturalCO2 springs. Functional Plant Biology, 31, 181–194.
Matthias, C. R., Kate, M. S., Jhon, N. K., & Michael, F. A.(1997). Microbial carbon-substrate utilization in therhizosphere of Gutierrezia Sarothrae grown in elevatedatmospheric carbon dioxide. Soil Biology and Biochemis-try, 29, 1387–1394.
Mayr, C., Miller, M., & Insam, H. (1999). Elevated CO2 alterscommunity-level physiological profiles and enzyme activ-ities in alpine grasslands. Journal of Microbiology Meth-ods, 36, 35–43.
Medlyn, B. E., Barton, C. V. M., & Broadmeadow, M. S. J.(2001). Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: a synthesis.New Phytology, 149, 247–264.
Melillo, J. M., Callaghan, T. V., Woodward, F. I., Salati, E., &Sinha, S. K. (1990). Effects on ecosystems. In J. T.Houghton, G. J. Jenkins, & J. J. Ephraums (Eds.), Climatechange: the IPCC scientific assessment, Cambridge.Cambridge: Cambridge University Press.
Morgan, J. A., Pataki, D. E., & Körner, C. (2004). Waterrelations in grassland and desert ecosystems exposed toelevated atmospheric CO2. Oecologia, 140, 11–25.
Naidu, S. L., DeLucia, E. H., & Thomas, R. B. (1998).Contrasting patterns of biomass allocation in dominant andsuppressed loblolly pine. Canadian Journal of ForestResearch, 28, 1116–1124.
Naumburg, E., Housman, D. C., Huxman, T. E., Charlet, T. N.,Loik, M. E., & Smith, S. D. (2003). Photosyntheticresponses of Mojave Desert shrubs to free air CO2
enrichment are greatest during wet years. Global ChangeBiology, 9, 276–285.
374 Water Air Soil Pollut (2011) 219:365–375

Pushnik, J. C., Garcoa-Ibilcieta, D., Bauer, S., Anderson, P. D.,& Houpis, J. (1999). Biochemical reponses and alteredgenetic expression patterns in ponderosa Pine (Pinusponderosa Doug ex P.Laws) grown under elevated CO2.Water, Air, and Soil Pollution, 116, 413–422.
Roane, T. M. (1999). Lead resistance in two bacterial isolatesfrom heavy metal-contaminated soils. Microbial Ecology,37, 218–224.
Ross, D. J., Tate, K. R., & Newton, P. C. D. (1995). ElevatedCO2 and temperature effects on soil carbon and nitrogencycling in ryegrass/white clover turves of an EndoaqueptSoil. Plant and Soil, 176, 37–49.
Rouhier, H., & Read, D. J. (1998). Plant and fungal responsesto elevated atmospheric carbon dioxide in mycorrhizalseedling of Pinus sylvestris. Environmental and Experi-ment Botany, 40, 237–246.
Scigelova, M., & Crout, D. H. G. (1999). Microbial β-N-acetylhexosaminidase and their biotechnological applica-tions. Enzyme and Microbial Technology, 25, 3–14.
Tabatabai, M. A. (1982). Soil enzymes. In A. L. Page(Ed.), Methods of soil analysis. Part 2. Agronomymonograph 9 (pp. 903–904). Madison: American Societyof Agronomy.
Tang, S. R., Xi, L., Zheng, J. M., & Li, H. Y. (2003). Responseto elevated CO2 of Indian mustard and sunflower growing
on copper contaminated soil. Bulletin EnvironmentalContaminant Toxicology, 71, 988–997.
White, M. C., Decker, A. M., & Chaney, R. L. (1991). Metalcomplexation in xylem fluid I: Chemical composition oftomato and soybean stem exudates. Plant Physiology, 67,292–300.
Wierzbicka, M. (1999). Comparison of lead tolerance in Allumcepa with other plant species. Environmental Pollution,104, 41–52.
Woodward, F. I. (2002). Potentional impacts of global elevatedCO2 concentrations on plants. Physiology and Metabo-lism, 5, 207–211.
Wu, H. B., Tang, S. R., Zhang, X. M., Guo, J. K., Song, Z. G.,Tian, S., et al. (2009). Using elevated CO2 to increase thebiomass of a Sorghum vulgare×Sorghum vulgare var.sudanense hybrid and Trifolium pratense L. and to triggerhyperaccumulation of cesium. Journal of Hazard Material,170, 861–870.
Yilmaz, S. (2002). Determination of optimal land use ofErzurum plain, Atatűrk Űniversity. Agriculture Faculty,32(4), 485–498.
Zak, D. R., Pregitzer, K. S., King, J. S., & Holmes, W. E.(2000). Elevated atmospheric CO2, fine roots and theresponse of soil microorganisms: a review and hypothesis.New Phytology, 147, 201–222.
Water Air Soil Pollut (2011) 219:365–375 375





























