Embed Size (px)
Citation preview

J. r&~¢~ Bid. VoL 5. PP. 179 to 183 0306-4565/g0/O701-017910Z00/0 O p e r p m o n Press Ltd 191;0. Printed in Great Britain
EFFECTS OF COLD ACCLIMATION AND 6-OHDA SYMPATHECTOMY ON COLD TOLERANCE OF BILATERALLY ADRENO-DEMEDULLATED RATS
MA~LYN K[ANG-ULalCH and STEVEN M. HORVATH Institute of Environmental Stress, University of California. Santa Barbara CA 93106, U.S.A.
(Received 9 September 1979; accepted in revised form 8 November 1979)
Abstract--l. Cold tolerance (-10°C) was evaluated in cold-acclimated (5°C) and non-cold acclimated (23°C), bilaterally adreno-demedullated rats with and without 6-hydroxydopamine (6-OHDA) sympath- ectomy.
2. Cold acclimation enhanced the cold tolerance of demedullated rats similar to that observed in intact rats.
3. Demedullated. c-a 6-OHDA-treated rats were able to maintain their colonic temperature better than saline-treated rats while the non-c-a 6-OHDA-treated rats were unable to maintain their colonic temperature to the same extent as the saline rats. These observations indicate that intact and adreno- demedullated animals have similar responses.
INTRODUCTION
PltEV,OUSLY we have shown that while 6-hydroxydo- pamine (6-OHDA) sympathectomized non-cold- acclimated rats had a reduced tolerance to a severe cold exposure, cold-acclimated rats subjected to 6 -OHDA sympathectomy had enhanced tolerance to this stress (Kiang-Uirich & Horvath, 1979ag Bilater- ally adreno-demeduilated rats were successfully cold- acclimated and tolerated an additional cold stress as effectively as intact cold-acclimated rats. However, demedullated rats living in 23°C did not tolerate the cold stress as well as intact rats (Kiang-Ulrich & Hor- vath, 1979bg
These observations did not help to clarify the role of sympathetic nervous system other than possible suggestion of compensatory hyperactivity of one com- ponent of sympathetic nervous system while the other component was inactivated. The present study was designed to evaluate the cold tolerance of cold- and non-cold-acclimated rats under a condition in which both components of the sympathetic nervous system would be inactivated by using the combination of 6 -OHDA sympathectomy and bilateral adreno- demedullation. 6 -OHDA specifically degenerates the sympathetic nerve terminals (Tranzer & Thocnen, 1967) with tim sympatbectomy being reversible (De Champlain. 1971) after a few months.
MATERIALS AND METHODS
Twenty-four male Sprague-Dawley rats*, each initially weighing 15011, were used in this study. Animah were indi- vidually caged and maintained at a room temperature of 23°C (± I°C]~ Purina Laboratory Chow and water were
* In conducting this research, the investigators adhered to the "Guide for Laboratory Animal Facilities and Care" as promulgated by the Committee for Laboratory Animal Facilite$ and Care, of the Institute of Animal Laboratory Resources, National Academy of Sciences--National Research Council.
provided ad libitur~ and a 12-h light and dark cycle was maintained. Bilateral adreno-demedullation was performed under sodium pentobarbital anesthesia {Evans, 1935). Three days following surgery the animals were randomly divided into two equal groups: 12 animals, cold-acclimated {c-a), were placed in a cold room maintained at 5°C (±I°C) for the next 5weeks and the remaining 12 rats, non-cold-acclimated (non-c-a), were maintained at 23°C (± I°C). At the end of this time interval 6 rats from each group were pre-treated with 6-OHDA-HBr (100mg/kg body weight, i.p.} and the remaining 6 were pre-treated with acidified saline, the carrying vehicle for 6-OHDA. The calorigenic response to an injection of tyramine was evalu. ated the next day. Tyramine was used in pcefefence to norepihephrine (NE) because tyramine liberates endoge- nous NE, is less toxic to rats than NE, and there is a linear relationship between metabolic response to tyramine and metabolic response to NE (Heroux et aL, 1975). Four or five days later these same animah were again given 6-OHDA or acidified saline and on the following day a cold tolerance tesL exposure to -10°C, was performed. The 6-OHDA was wepared immediately before each injec- tion by dissolving 6-OHDA-HBr in I ml of acidified saline (I tug ascorbate/ml of 0.9% saline). Oxygen uptake was measured in a constant-volume, closed system (Volume Meter, Med Science. model 160) with the rat in a sealed Plexiglas chamber and the expired CO, absorbed by soda lime. Rats were not free-moving since their tails were taped down with the thermistor probes. Two constant- temperature baths were used to maintain chamber tem- perature of either 28 or - 10°C. Colonic and chamber tern- peratures were measured continuously with thermistor probes and recorded on a dual-channel recorder.
Before evaluating the animal's response to tyramine, .oxygen uptakes and colonic temperatures were first
measured at 28°C for 30 min. Each animal was then tem- porarily removed from the chamber and tyramine-HCL 20 mg/kg body weight, wag given i.p. The animal was then returned to the chamber and measurements resumed for another hour. ~?old tolerance was determined by measur- ing oxygen uptakes and colonic temperatures first at 28°C for 30rain and then transferring the animal to a -10"C environment Meamrements of oxygen uptake and colonic temperature were continued for either 3 h or until the ani- mal's colonic temperature fell to 17°C.
T.IL 3/~b'--D 179
180 MARILYN KIANG.-ULRICH and S ~ v E N M. HORVA'rH f v o , o . . , ,:- ] . . o, . - , o - ~mamine inl*~m~ I - d W ~n~an~n, i ~ t i ~ 5 0 0 t e l -]E~
T I I l I
~ , I , ) T ' - I " l , , ,
i I ! , I [ * " ' " " "
, 3 0 0 , i , i l ~
mSm el~qUA term, eUHOt tetra* e~4Ot ,elm* No~-cold-occllmoted C~ld-(gclknfted Non-cokl-4cclimoted C o l d - ~ e d
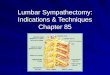
Fig. I. Demedullated rats. Oxygen uptakes and colonic temperatures in response to tyramine: Oz uptake (left) and colonic temperature (right} measured at 28°C of non-c-a and c-a rats treated with acidified saline or 6-OHDA. before and after injection of tyramine. Each bar represents the mean + SE
of values from 6 rats.
Data were analyzed utilizing a three-factor factorial analysis of variance-with repeated measures across time. Where significant interaction effects were found, tests of simple main effects were made. The 0.05 level of signifi- cance was selected for rejection of null hypothesis (Wirier. 1971). The analysis of the cold tolerance test data was per- formed using the first 100rain of cold exposure since not all the animals complete the 180-min test time. Decre~__~s_ of colonic temperature in -10~C across time were also analyzed by a series of linear regressions to determine the best-fitting curve. The significance of the differences between regremon coefficients (slopes) was determined by using Student's t-test.
RESULTS
Oxygen uptakes measured at 28°C prior to the ad- ministration of tyramine were 231 mL kg- t. 10 rain- t
for the c-a animals and 191 mL kg- t. 10 min- , for the non-c-a rat~ Although this difference was not statisti- cally significant at the 0.05 level, the absolute differ- ence of oxygen uptake between the two groups of animals followed the same trend as we have observed previously in the intact or demedullated rats (Kiang- Ulrich & Horvath, 1979a, b); that is, the c-a rats had higher oxygen uptake than the non-c-a rats. No stat- istically significant difference between the saline- and 6-OHDA-treated animals was obtained. Colonic tem- perature averaged 0.8°C higher in c-a rats (36.8°C) than in non-c-a ra ts (36°C) (P < 0.01) (Fig. 1).
All animals had significant (P < 0.01) increases in their oxygen uptakes and colonic temperatures fol- lowing tyramine administration. Saline-treated c-a animals had a 81.7% increase (from 235 to 427 mL kg- t. 10 rain- ') in oxygen uptake and 3.4°C rise ( f rom 36.6°C) in colonic temperature and 6-OHDA-treated c-a animals had a 52.6~ (from 228 to 348 mL kg- i. I0 rain- t) in oxygm uptake and a 2.6°C rise (from 37.1cC3 in colonic temperature (Fig. I), Non-c-a saline-treated animals had a 68.3% increase (from 199 to 335 ml. k8 - 1 - 1 0 r a i n - ' ) in oxy-
uptake and 2A°C elevatioo (from 36.4"C) of colo- nic temperature and 6-OHDA-trmtad non-ca ani- mals had a 17.5% increase (from 183 to 2 1 5 m L k g - l . l O m i n - I ) in oxygen uptake and 2.1°C rise (from 35.7°C) in colonic temperatur~ Cold accli- marion significantly (P < 0.05) augmented the re- sponse to tyramine while pre-treaunent with
6-OHDA significantly (P < 0.05) attenuated the re- sponse (Fig. 1).
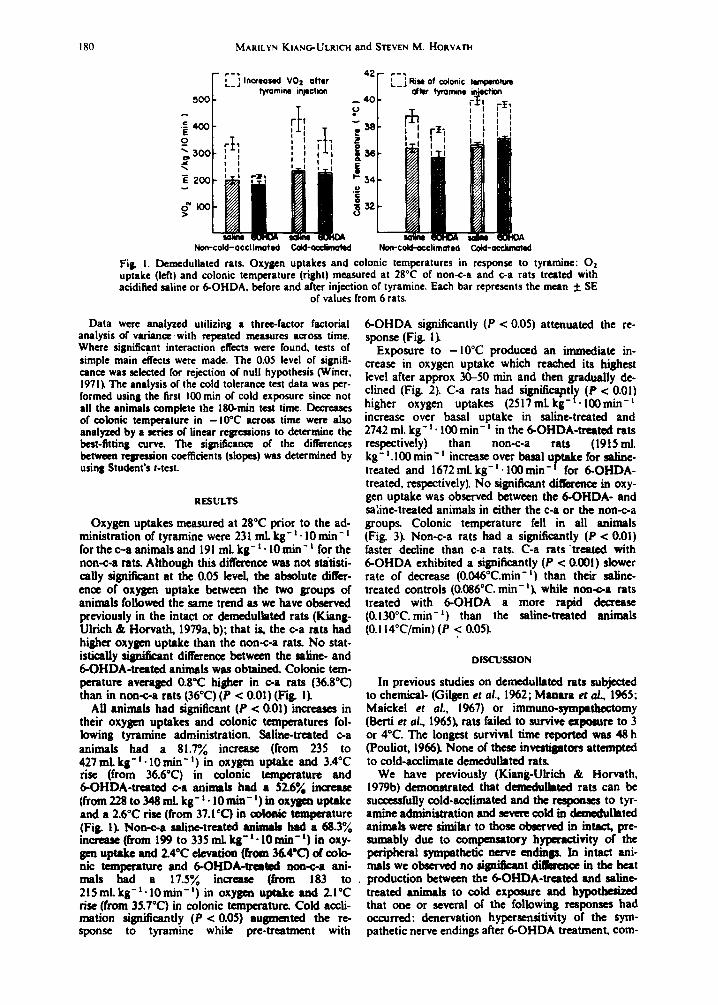
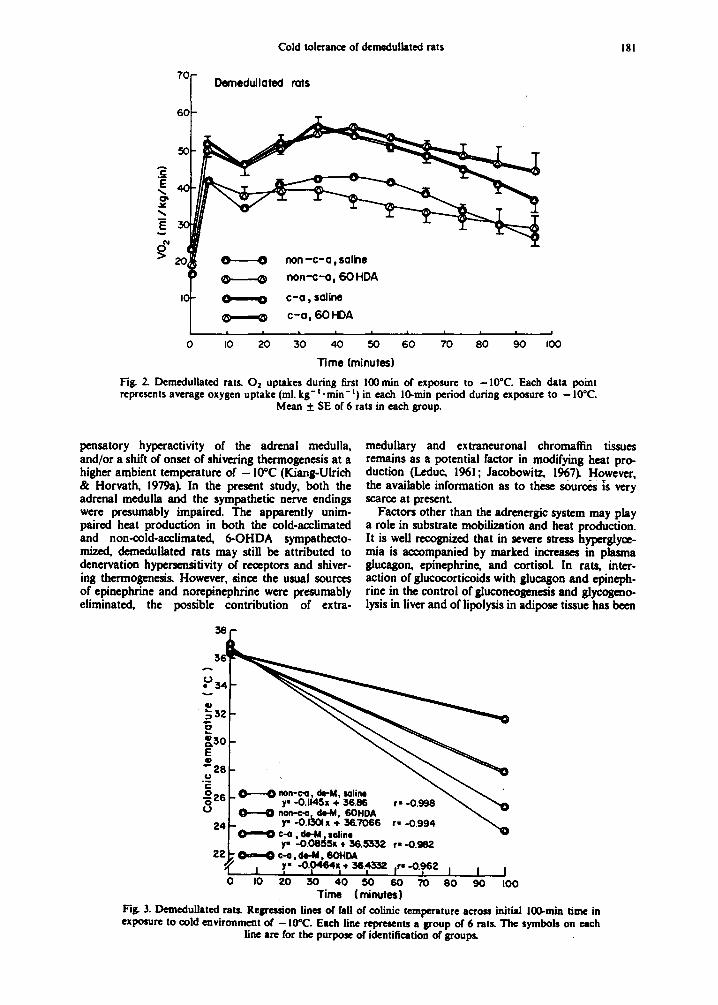
Exposure to -10°C produced an immediate in- crease in oxygen uptake which reached its highest level after approx 30-50 rain and then gradually de- clined (Fig. 2). C-a rats had signitica~ntly (P < 0.01) higher oxygen uptakes (2517mLkg-t . 100min - t increase over basal uptake in saline-treated and 2742 ml. kg- 1. 100 min- i in the 6-OHDA-treated rats respectively) than non-c-a rats (1915 mL kg-1.100 min- t increase over basal uptake for saline- treated and 1672 mL kg- t. 100 rain- t for 6-OHDA- treated, respectively). No significant difference in oxy- gen uptake was observed between the 6-OHDA- and sa~ine-treated animals in either the c-a or the non-c-a groups, Colonic temperature fell in all animals (Fig. 3). Non-c-a rats had a significantly (P < 0.01) faster decline than c-a rats. C-a rats "treated with 6-OHDA exhibited a significantly (P < 0.001) slower rate of decrease (0.046°C.min - I ) than their saline- treated controls (0.086°C. min-l) , while non-c-a rats treated with 6-OHDA a more rapid decrease (0.130°C. min - I ) than the saline-treated animals (0.114°C/min) (P < 0.05).
D I S C U S S I O N
In previous studies on demeduilated rats subjected to chemical- (Giigen et ai., 1%2; Mamma et aL 1965; Maickel et al., 1967) or immano-sympathectomy (Berti et aL, 1965), rats failed to survive eapomre to 3 or 4°C. The longest survival time reported was 48 h (Pouliot, 1966). None of these investigators attempted to cold-acclimate demedullated rats.
We have previously (Kiang-Ulrich & Horvath, 1979b) demonstrated that demedullated rats can be su__eo~__qfully told-acclimated and the responses to tyr- amine administration and severe cold in demedullated animals were similar to those obeerved in intact, pre- sunuthly due to compensatory h y l ~ v i t y of the peripheral sympathetic nerve endinl~ In intact ani- mals we observed no s i g n i ~ t d i ~ in the heat production between the 6-OHDA-treated and saline- treated animals to cold exposure and hypothesized that one or several of the following responses had occurred: denervation hypersensitivity of the sym- pathetic nerve endings after 6-OHDA treatment, corn-
Cold tolerance of demgdullated rats 181
701 Demedulloted rots
60
¢ @ @ non-c-o, 60HDA
I0~- ~ c - o , saline c-o, 60HDA
0 iO 20 30 40 5o 60 ?o 80 90 IO0
Time (minutes)
Fig. 2. Demedullated rats. O= uptakes during first 100rain of exposure to -10°C. Each data point represents average oxygen uptake (ml. kg- I. min- ~) in each 10-rain period during exposure to - 10°C.
Mean + SE of 6 rats in each group.
pensatory hyperactivity of the adrenal medulla, and/or a shift of onset of shivering thermogenesis at a higher ambient temperature of - 10°C (Kiang-Ulrich & Horvath, 1979a). In the present study, both the adrenal medulla and the sympathetic nerve endings were presumably impaired The apparently unim- paired heat production in both the cold-acclimated and non-cold-acclimated, 6-OHDA sympathe~o- mized, demedullated rats may still he attributed to denervation hypersensitivity of receptors and shiver- ing thermogenesis. However, since the usual sources of epinephrine and norepinephrine were presumably eliminated, the possible contribution of extra-
medullary and extraneuronal chrornaffin tissues remains as a potential factor in modifying heat pro- duction (Leduc, 1961; Jacobowitz, 1%7~ However, the available information as to these sourc~ ;s very scarce at presenL
Factors other than the adrenergic system may play a role in substrate mobilization and heat production. It is well recognized that in severe stress hyperglyce- mia is a__eo3mpanied by marked increases in plasma glucagnn, epinephrine, and cortisoL In rats, inter- action of glucocorticoids with glucagon and epineph- rine in the control of g l u c o ~ s and glycogmo- lysis in fiver and of lipolysis in adipose tissue has been
• E 3:, I" ~ ' ~ .
, , 3 0 I- ~
g .
22 ~" O .m- -O c-o, de-M. 60HOA / ),- -0 .0464x ,~ 36.4552 f- -0.962
o Io 20 3o 4 0 so so 70 eo 9o too Time (minutes)
Fig. 3. Demedullated rats. Regres~on lines of fall of colinic temperature across initial lO0-min time in exposure to cold environment of - 10°C. Each fine represents a group of 6 rats. The symbols on each
line are for the purpose of identification of groups.

182 MARILY,'q KIANG"ULRICH and STEVt'N M. HORVATH
observed (Exton etal. . 1972). There is the possibility that the adrenal cortex liberates glucocorticoids in re- sponse to the alerting reaction which could be trig- gered by the 6-OHDA sympathectomy and severe cold exposure. Glucocorticoids appear to be impor- tant in mobilizing energy resources and allowing increased thermogenesis through glycogenesis, gluco- neogenesis, and lipolysis in rats (Deavers & Musac- chia, 1979). Marked increase in blood glucose (Ira- bach, 1976: Bennett & Gardiner, 1978), serum free fatty acid. and liver glycogen levels (Imbach, 1976) in 6-OHDA-treated animals has been observed. 6-OHDA treatment depleted the NE contents in brown adipose tissue (BAT) (Thurenson-Klein etal . , 1976): however, the trophic response of BAT to cold was not prevented (Mory e ta l . , 1976). These data suggest the existence of some other factors indepen- dent of NE that have a role in substrate mobilization and heat production.
The heat production and body heat content for the various demedullated groups before and after tyr- amine injection were calculated based on body weights, oxygen uptakes, and colonic temperatures. We found that the changes were similar to those we observed in intact animals (Kiang-Ulrich & Horvath, 1979a). After tyramine injection non-c-a, saline- treated rats produced 55~ more heat but had slightly (0.8To) greater heat content than non-c-a, 6-OHDA rats. C-a, saline-treated rats produced 24~ more heat with slightly (0.8~) greater heat content than the 6-OHDA-treated rats. Enhanced vasoconstriction apparently occurred in the 6-OHDA treated rats. This was more evident in the c-a, 6-OHDA rats`
The variations in the maintenance of colonic tem- perature among the various demeduilated groups dur- ing the severe cold exposure observed in this study were similar to those reported in intact rats (Kiang- Ulrich & Horvath, 1979ag The rate of fall in colonic temperature in c-a rats was slower than in non-c-a rats. In the c-a groups the fall in colonic temperature was slower in 6-OHDA animals than saline rats, but in the non-c-a, 6-OHDA rats colonic temperature de- creased faster than in saline rats. Since there was no significant difference in heat production between 6-OHDA and saline animals (c-a or non-c-a), the dif- ference in the maintenance of colonic temperature could only be attributed to different environmental heat losses. It is possible that as a result of chemical sympathectomy 6-OHDA-treated non-c-a rats could not vasoconstrict to preserve heat in a severe cold environment and therefore could not maintain their colonic temperature as well as the saline-treated ani- mals. However, chemical sympathectomy did not seem to increase heat loss in c-a rats, In fact, c-a, 6-OHDA rats maintained colonic temperature better than saline-treated rats. Denervation hypersensitivity developed in the vasoconstricting smooth muscle after 6-OHDA administration could potentiate heat con- servation in the c-a, 6-OHDA animals, enabling the c-a, 6-OHDA rats to maintain colonic temperature much better than the saline-treated, c-a rats` How- ever, this explanation is not applicable to non-c-a rats.
It is apparent that 6-OHDA treatment of demedul- lated rats elicited different responses in c-a and non- c-a rats. These responses were similar to those we had
observed in the intact rats. This re-emphasizes our previous suggestion that some additional unknown factor(s) participates in the development of cold accli- mation.
SUMMARY
Cold acclimation enhanced the demedullated rat's tolerance to cold similar to the response of intact rats. This was evidenced by the greater increase in calori- genie response to tyramine administration in c-a rats than in non-c-a rats measured at 28°C and better maintenance of heat production and colonic tempera- ture in the c-a rats to a - 10°C exposure.
Administration of 6-OHDA reduced the tyramine- induced calorigenic response in c-a and non-c-a rats. 6-OHDA reduced cold tolerance (to -10°C) in the non-c-a demedullated rats, but enhanced cold toler- ance in the c-a demedullated rats` This observation was similar to that we had observed in intact rats.
Acknowledoements--This work was supported in part by the National Institutes of Health, under Grant NIH AG00021.
We wish to especially acknowledge the assistance of Dr Barbara Drinkwater in the statistical analysis of this data.
REFERENCES
BENNETT T. & GARDINER S. M. (1978) Reversal of 6-hydroxydopaminc-induced hypotension in the rat without activation of the renin-angiotemfif~ system. J. Physiol. ~]9, 1-16.
BENT1 F., LENTATI R. & USAaOI M. M. (1965) Effect of cold exposure on heart function in immunosympathecto- :nized rats. Med. Pharmac. Exp. 13, 227--232.
DEAVELS D. R. & MUSACCHtA X. J. (1979) The function of glucocorticoids in thermoigenesis, Fedn Proc. Fedn Am, Sots exp. Biol. 38, 2177-2181.
DE CI~UI'LAIN J. (1971) Degeneration and regrowth of adrenerglc ner~te fibers in the rat lx'riphm'al tissues after 6-hydroxydopamine. Can. J. Physiol. Pharmac. 49, 345-355.
Ev~'~s J. (1935)The adrenal cortex and endogenous carbo- hydrate formation. Am. J. Physiol. !!4, 297-308.
ExTot~ J. H., FRI£DMAN.'q N., WONG E. H.. PARKE BmNF.~UX J.. CoItSIN J. D. & PAgg C. R. (1972) Interaction of glucocorticoids with glucagon and epinephrine in the control of gluconeogenesis and glycogenolysis in liver and of lipolysis in adipose tissue. J. biol. Chem. 247(I). 3579-3588.
GILGEN A., M^tCKEL R. P., NIKODIZEVIC O. & BRODIE B. B. (1962) Essential role of catecholamines in the mobil- ization of FFA and glucose after exposure to cold. Lzfe Sci. 12, 709-715.
HEaoux O., PAOt+ E., I,E BLAP~C J.. ~ u c J., GILamaT R. V l ~ l l t £ A. & RlVES'r P. (1975) Nonshivering thermo- genesis and cold resistance in rats under severe cold con- ditions. J. appl. Physiol. 3~3), 436-442.
IMaAOa A. (1976) Metabolic alterations following chemical symplthectomy with 6-hydroxydopamine in the rat. In Recent AdmRces in Studies on Cardiac Structure and Metabolism. (Edited by RoY P. E. & DI, ht, l,l.~ N. S.) VoL 9, The Sarcolemma pp. 259-267. University Park Press. Baltimore.
JAccxg~vr~ D. (1967) Histocbemicai studies of the re- lationship ofchromaflin cells and adrenergic nerve fibers
• to the cardiac ganglia of several species. J. Pharmac. exp. Ther. ! ~ , 227-240.
Kt^No-ULmcH M. & HORVATH S. M. (1979a) Effects of cold acclimation and 6-OHDA sympatbectomy on cold tolerance of rats. J. therm. Biol. 4, 223-237.

Cold tolerance of demeduilated rats 183
KIANG,-ULRICH IV[. R, HORVATH S. M. (t979b) Successful cold acclimation following bilateral adreno- demedullation in rats. Proc. Soc. exp. Biol. Med. In press.
LEt~uC J. (1961) Catecholamine production and release in exposure and acclimation to cold. Acra Physiol. scand. 53 (SuppL 183), 5-101.
MAICKEL R. P., MATSU~EK N., STEgN D. N. & Baoote B. B. (1967) The sympathetic nervous system as a homeo- static mechanism. I. Absolute need for sympathetic her- vous function in body temperature maintenance of cold- exposed rats. J. Pharmac. exp. Ther. 157. 103-110.
MANARA L., COSTA E., STERN D. N. & MAICKEL R. P. (1965) Effect of chemical sympathectomy on oxygen con- sumption by the cold-exposed rat. Int. J. Neuropharm. 4, 301-307,
MORY G., NECHAD M., RICQUIER D. & HERMON PH. (1978) 6-OH-dopamine and the development of brown adipose tissue in the cold. Experientia (Suppl) 32, 113-117.
POULIOT M. (1966) Catecholamine excretion in adreno- demedullated rats exposed to cold after chronic guaneth- idine treatment. Acta Physiol. stand, 68, 164-168,
THUREN~ON-KLEIN A~ LAGEIgCRANTZ H. & BARNARD T. (1976) Chemical sympathectomy of interscapula brown adipose tissue. Acta Physiol. scand. 98, 8-18.
TXANZ~ J. P. & TttOCNEN H. (1967) An electron micro- scopic study of selective, acute degeneraton of sympathe- tic nerve terminals after administration of 6-hydroxydo. pamine. Experientia 24(2), 155-156.
WINER B. J. (1971) Statistical Principles in Experimental Design, 2nd edn. p. 539. McGraw-HilL New York.
Key Word Index--Rats; bilaterally adreno.demedullated rats, cold acclimation; cold tolerance, 6-hydroxydopamine (6-OHDA): 6-OHDA chemical sympathectomy: oxygen uptake and colonic temperature.







![Peripheral Sympathectomy for Raynaud's Phenomenon: A ... · sympathectomy [1–3,6,7]. Conventional cervical sym-pathectomy has been used, but the results are disap-pointing. It has](https://img.pdfslide.us/doc/110x75/602abfd7d9da841d876f1382/peripheral-sympathectomy-for-raynauds-phenomenon-a-sympathectomy-1a367.jpg)