Embed Size (px)
Citation preview

This article was downloaded by: [The University of Manchester Library]On: 10 October 2014, At: 11:07Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number:1072954 Registered office: Mortimer House, 37-41 Mortimer Street,London W1T 3JH, UK
Archiv für TierernaehrungPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/gaan19
Effects of cell walldegrading enzymes oncarbohydrate fractions andmetabolites in stomach andileum of pigs fed wheatbran based dietsJ. Van der Meulen c , J. Inborr b & J. G. M.Barker aa Institute for Animal Science and Health(ID‐Lelystad) , Lelystad, The Netherlandsb Finnfeeds International Ltd. , Marlborough,Wiltshire, United Kingdomc Institute for Animal Science and Health(ID‐Lelystad) , P.O. Box 65, 8200 AB, Lelystad,The NetherlandsPublished online: 10 Jan 2009.
To cite this article: J. Van der Meulen , J. Inborr & J. G. M. Barker (2001)Effects of cell wall degrading enzymes on carbohydrate fractions andmetabolites in stomach and ileum of pigs fed wheat bran based diets, Archiv fürTierernaehrung, 54:2, 101-115, DOI: 10.1080/17450390109381970
To link to this article: http://dx.doi.org/10.1080/17450390109381970
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of allthe information (the “Content”) contained in the publications on ourplatform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy,

completeness, or suitability for any purpose of the Content. Anyopinions and views expressed in this publication are the opinions andviews of the authors, and are not the views of or endorsed by Taylor& Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information.Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilitieswhatsoever or howsoever caused arising directly or indirectly inconnection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private studypurposes. Any substantial or systematic reproduction, redistribution,reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of accessand use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

Arch. Aninu Nutz, 2001, Vol. 54, pp. 101-115Reprints available directly from the publisherPhotocopying permitted by license only
© 2001 OPA (Overseas Publishers Association) N.V.Published by license under
the Harwood Academic Publishers imprint,part of Gordon and Breach Publishing,
a member of the Taylor & Francis Group.
EFFECTS OF CELL WALL DEGRADINGENZYMES ON CARBOHYDRATEFRACTIONS AND METABOLITES
IN STOMACH AND ILEUM OF PIGSFED WHEAT BRAN BASED DIETS
J. VAN DER MEULENa, J. INBORRb and J. G. M. BARKERa
aInstitute for Animal Science and Health (ID-Lelystad),Lelystad, The Netherlands; bFinnfeeds International Ltd., Marlborough,
Wiltshire, United Kingdom
(Received 28 July 2000)
Pigs were fed diets containing 40% wheat bran incubated with a water.-acetic acid mixture(control, C) and a cellulase (Cel-i) or xylanase (Xyl-i) preparation or with addition of thecellulase (Cel-a) or xylanase (Xyl-a) preparation immediately before feeding. Stomach andileal samples were analysed for pH, osmolality, soluble saccharides, volatile fatty acids (VFA)and lactic acid. Incubation of wheat bran resulted in a small reduction of NDF and an increasein the amount of soluble starch, ß-glucans and saccharides (glucose, xylose and arabinose),especially after incubation with the cellulase preparation. Two hours after feeding, significantlyhigher arabinose and xylose concentrations were present in the stomach for diets Cel-i, Cel-aand Xyl-i. In the ileum xylose and arabinose concentrations were higher 2 to 4 and 6 to 8 hoursafter feeding the enzyme-treated diets. In stomach and ileum there were no differences betweenthe diets in pH, osmolality, VFA and lactic acid concentrations, but ileal VFA concentrationfrom 4 h after feeding tended to be higher for diets Cel-i and Xyl-i. It can be concluded that theamount of soluble saccharides in stomach and small intestine and the ileal VFA concentrationmay be increased by cell wall degrading enzyme preparations.
Keywords: Pig; Wheat bran; Enzymes; Soluble saccharides; Volatile fatty acids
Address for correspondence: Dr. Jan van der Meulen, Institute for Animal Science andHealth (ID-Lelystad), P.O. Box 65, 8200 AB Lelystad, The Netherlands.
101
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

102 J. VAN DER MEULEN et al.
1. INTRODUCTION
Dietary fibre, which may be defined as non-starch polysaccharides (NSP)and lignin, influences many processes in the digestive tract of the pig (Low,1985). NSP are resistant to hydrolysis by the digestive enzymes of mono-gastrics but can be degraded by microbes. Microbial fermentation occurs inthe large intestine, but fermentation also takes place in the upper part of thegastrointestinal tract (Bach Knudsen et al, 1991). The major end productsof fermentation in stomach and small intestine are lactic acid and volatilefatty acids (VFA) (Argenzio and Southworth, 1975; Clemens et al, 1975).
The main NSP components of wheat and wheat by-products are theso-called pentosans or arabinoxylans (Annison et al., 1992). In wheat bran,the content of arabinoxylans (the sum of arabinose and xylose) varies from20.6 to 26.2% (Graham et al, 1986; Bach Knudsen and Hansen, 1991).Wheat bran is commonly used as an ingredient in pig feeds, but due toknown detrimental effects on digestion and utilisation of nutrients, its rateof inclusion has to be restricted. Treating wheat bran with cell wall degradingenzymes before feeding or supplementation of the feeds with such enzymesmay increase its digestibility and hence utilisation in rats (Neudoerffer andSmith, 1969) and broiler chickens (Nahm and Carlson, 1985; Annison,1992). Although the effect of cell wall degrading enzymes in pigs is vari-able (Dierick and Decuypere, 1994), there appears to be an opportunity forcell wall degrading enzymes to improve the nutritional value of wheat bran(Schutte et al, 1990).
ß-glucanase and cellulase activities in stomach and ileum of pigs fed dietswith 40% wheat bran increase when crude cellulase or xylanase preparationsare used (Inborr et al, 1999). The effect of these enzyme preparations onthe degradation of NSP in the upper part of the gastrointestinal tract andon physical-chemical conditions (pH and osmolality) and VFA and lacticacid concentrations are reported in this paper. Two ways of enzyme appli-cation were employed, either pre-treatment of wheat bran with the enzymepreparations or adding the enzyme preparations just before feeding.
2. MATERIALS AND METHODS
2.1. Animals, Diets and Experimental Design
Animals, diets, enzymes, experimental design and collection procedureswere described in detail by Inborr etal. (1999).
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 103
Five crossbred barrows fitted with stomach and post valve T-caecumcannulas were randomly assigned to one of five dietary treatments in fiveconsecutive periods of 14 d in a 5 x 5 Latin Square design. At the start of theexperiment the pigs weighed 38 kg and at the end of the experiment 57 kg.
All diets had the same composition with wheat bran as the only source ofNSP (Tab. I). The wheat bran was incubated with water and acetic acid at aratio of 1:6.25 and 1:0.02 (w/v), respectively, at 39°C and pH 5.0 for 3.5 hto give treatment C (control). For the preparation of diets Cel-i and Xyl-f,wheat bran was incubated in the same conditions as the control with eithera cellulase (10g/kg) or a xylanase preparation (0.1 g/kg). Before feeding,wheat bran and incubation water were mixed with an appropriate amountof the balance of the diets, resulting in a slurry with a water to feed ratio of2.5:1 (v/w). Immediately before feeding, 10 g of the cellulase or 0.1 g of thexylanase preparation per kg wheat bran were added to diet C to form dietsCel-a and Xyl-a, respectively.
The enzymes used were a crude cellulase and a crude xylanase prepara-tion containing cellulolytic, hemicellulolytic and xylanolytic activities. Thepreparations were added to provide the same amount of xylanase activity,which obviously resulted in a higher ß-glucanase activity in the diets supple-mented with the cellulase preparation compared with those supplementedwith the xylanase preparation (Tab. II).
The pigs were fed twice daily (at 08.30 and 20.30 h) at 2.3 times mainte-nance requirement (= 418 kJ ME/kg075) and had no access to water betweenthe feedings. Each treatment period lasted 14 d. The animals were switcheddirectly from one diet to the next one at the end of each period. Within a
TABLE I Composition of the diets [g/kg]
Ingredient
Wheat bran1 400.00Maize starch 497.05Casein 82.00Limestone 14.00Salt 2.50DL-Methionine 0.80Choline chloride (50%) 0.25Mineral/vitamin premix2 1.40
-starch preraix 2.00
1 Incubated with or without enzymes.2 Provided the following ingredients (mg/kg diet): retinol 3.9, cholecalciferol 0.04,DL-or-tocopherol 8, vitamin B2 4, vitamin B ) 2 0.02, nicotinic acid 20, pantothenicacid 7.84, antioxidant 125, FeSO4 • 7H2O 430, MnO 50, ZnSO4 • H2O 155, KI2,Na2SeO3-5H2O7.
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

104 J. VAN DER MEULEN et al.
TABLE II Calculated activities of the crude enzyme preparations in the diet [units/kg diet]
Substrate Cellulose treatment Xylanase treatment
Barley ß-glucan 103,900 844Oat spelt xylan 14,450 14,205Carboxymethylcellulose 73,700 505
treatment period gastric digesta was collected once daily on days 7,8,9, 10and 14 after the pigs had finished their meal (0 h) and 2,4,6 and 8 h after themorning feeding. The pH was measured and a sample was frozen and storedat —20°C. Heal digesta was collected quantitatively on the 4th and 1 lth dayin two-hour intervals for 12 h, beginning at 08.30 h. Each two-hourly samplewas weighed and immediately after pH had been measured, frozen and storedat —20°C. On days 6 and 13 faeces and urine were collected quantitativelyfor 24 h, frozen and stored at -20°C.
2.2. Analytical Methods
The (incubated) wheat bran was analysed according the NSP protocol ofvan Lonkhuijsen et al. (1997) for water soluble carbohydrates (mono- anddisaccharides and (after hydrolysis) oligosaccharides, starch, pectins andß-glucans), non-structural carbohydrates (total starch and total ß-glucansafter autoclaving in water) and structural carbohydrates (NDF and its saccha-ride composition). After sampling pH was determined immediately and aftercentrifugation osmolality was determined in the liquid parts of the stomachand ileal samples, using an osmometer based on freezing point depression(Gonotec Osmomat 030). VFA and lactic acid were measured in the liquidpart by gas-liquid chromatography (Robinson etal., 1986). Soluble mono-and disaccharides and oligosaccharides were analysed in stomach (2 and 6 hafter feeding) and ileal (2 to 4 and 6 to 8 h after feeding) samples.
2.3. Statistical Analysis
Data were subjected to analysis of variance using the GENSTAT 5 statisticalpackage (Payne et al., 1987) according to the Latin Square design and thefollowing model:
YiJk =ß + Tt + Pj + Ak+ eijk,
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4
ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 105
where ß is overall mean, 7} is effect of treatment, Pj is effect of period, Atis effect of animal and e^ is residual error with / = 1 . . . 5, j= 1 . . . 5, k=1.. .5.
The results are given as estimated means corrected for period and animaleffects.
3. RESULTS
3.1. Saccharides
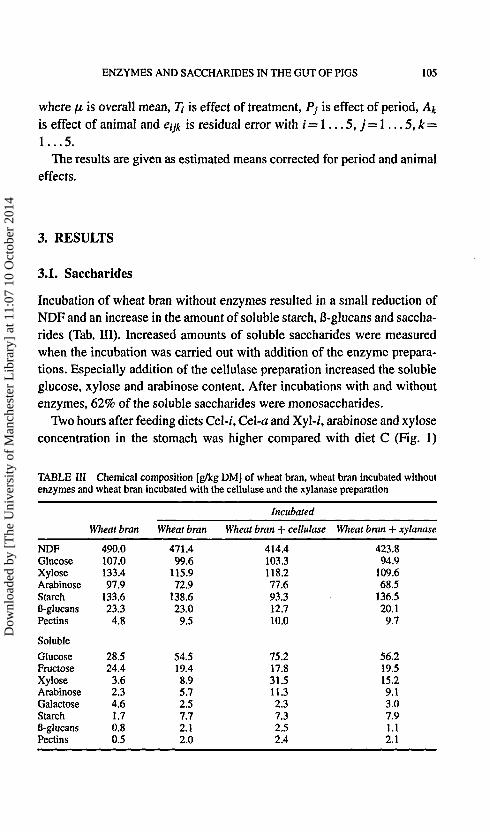
Incubation of wheat bran without enzymes resulted in a small reduction ofNDF and an increase in the amount of soluble starch, ß-glucans and saccha-rides (Tab. III). Increased amounts of soluble saccharides were measuredwhen the incubation was carried out with addition of the enzyme prepara-tions. Especially addition of the cellulase preparation increased the solubleglucose, xylose and arabinose content. After incubations with and withoutenzymes, 62% of the soluble saccharides were monosaccharides.
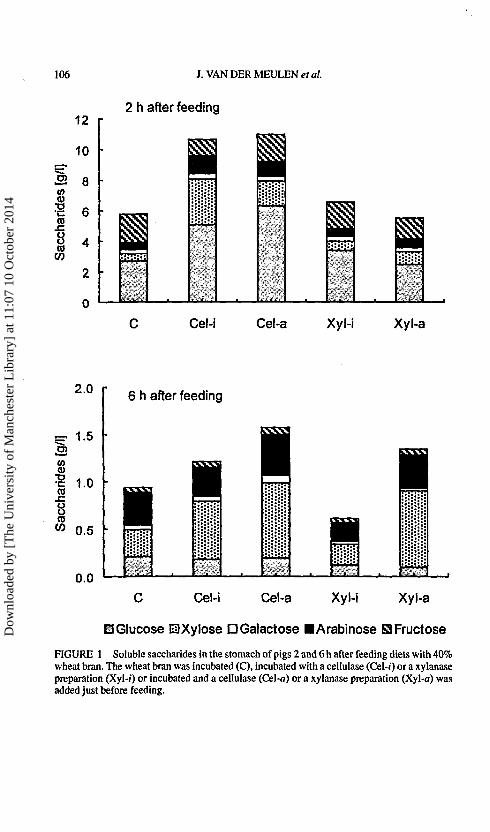
Two hours after feeding diets Cel-i, Cel-a and Xyl-/, arabinose and xyloseconcentration in the stomach was higher compared with diet C (Fig. 1)
TABLE III Chemical composition [g/kg DM] of wheat bran, wheat bran incubated withoutenzymes and wheat bran incubated with the cellulase and the xylanase preparation
NDFGlucoseXyloseArabinoseStarchß-glucansPectins
Soluble
GlucoseFructoseXyloseArabinoseGalactoseStarchß-glucansPectins
Wheat bran
490.0107.0133.497.9
133.623.34.8
28.524.4
3.62.34.61.70.80.5
Wheat bran
471.499.6
115.972.9
138.623.0
9.5
54.519.48.95.72.57.72.12.0
Incubated
Wheat bran + cellulase
414.4103.3118.277.693.312.710.0
75.217.831.511.32.37.32.52.4
Wheat bran + xylanase
423.894.9
109.668.5
136.520.1
9.7
56.219.515.29.13.07.91.12.1
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4
106
12
10
S 8CO
I 68 4
to n
2
0
J. VAN DER MEULEN et al.
2 h after feeding
Cel-i Cel-a Xyl-i Xyl-a
2.0
ç. 1.5
co
CO
CD
1.0
0.5
0.0
6 h after feeding
Cel-i Cel-a Xyl-i Xyl-a
D Glucose ED Xylose D Galactose BArabinose S3 Fructose
FIGURE 1 Soluble saccharides in the stomach of pigs 2 and 6 h after feeding diets with 40%wheat bran. The wheat bran was incubated (C), incubated with a cellulase (Cel-i) or a xylanasepreparation (Xyl-i) or incubated and a cellulase (Cel-a) or a xylanase preparation (Xyl-a) wasadded just before feeding.
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4
•g»
12
10
8m
•"g 6
(0
ENZYMES AND SACCHARIDES IN THE GUT OF PIGS
2 to 4 h after feeding
107
Cel-i Cel-a Xyl-i Xyl-a
I6
m
f 4CD•5oroco 2
6 to 8 h after feeding
Cel-i Cel-a Xyl-i Xyl-a
0 Glucose E3 Xylose D Galactose HArabinose • Fructose
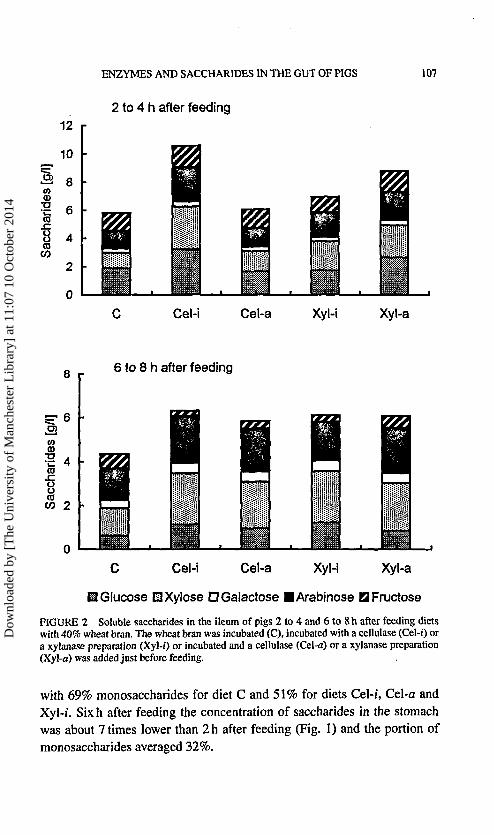
FIGURE 2 Soluble saccharides in the ileum of pigs 2 to 4 and 6 to 8 h after feeding dietswith 40% wheat bran. The wheat bran was incubated (C), incubated with a cellulase (Cel-i) ora xylanase preparation (Xyl-/) or incubated and a cellulase (Cel-a) or a xylanase preparation(Xyl-a) was added just before feeding.
with 69% monosaccharides for diet C and 51% for diets Cel-i, Cel-a andXyl-i. Six h after feeding the concentration of saccharides in the stomachwas about 7 times lower than 2 h after feeding (Fig. 1) and the portion ofmonosaccharides averaged 32%.
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

108 J.VANDERMEULENeraf.
TABLE IV Urinal excretion of saccharides [g/d] after feeding diets with 40% wheat bran
GlucoseXyloseGalactoseArabinoseFructoseTotal
C1
0.280.08a
0.030.370.080.84a
Cel-i1
0.250.74b
0.060.960.072.09c
CW-Ö3
0.210.20a
0.120.580.071.18b
Xyl-i*
0.280.18a
0.030.580.061.13ab
Xyl-a*
0.180.13a
0.020.450.070.85a
SED6
0.080.100.060.220.020.34
1C = control.2 Cel-i = incubation with cellulase.3 Cel-a = addition of cellulase.4 Xyl-i = incubation with xylanase.3 Xyl-a = addition of xylanase.6 SED = standard error of difference.Values within a line with no common superscript are significantly different (P<0.05).
Glucose, xylose, arabinose, fructose and traces of galactose were presentin the ileum 2 to 4 h after feeding (Fig. 2). Xylose and arabinose concentra-tions were higher after feeding the enzyme-treated diets, but this was onlysignificant for diet Cel-i. On average 50% of the saccharides were monosac-charides. In the period of 6 to 8 h after feeding the saccharide concentrationwas on average 0.7 times the concentration 2 to 4 h after feeding (Fig. 2).The xylose and arabinose concentrations were higher for the enzyme-treateddiets, and the portion of monosaccharides averaged 30%.
After feeding diet C, small traces of glucose, xylose, galactose and fruc-tose were found in the urine, while slightly more arabinose was found. Theexcretion of monosaccharides increased after feeding the enzyme-treateddiets, with the exception of diet Xyl-a (Tab. IV). The higher urinary excre-tion of monosaccharides for the cellulase-treated diets and for diet Xyl-i wasmainly caused by a higher excretion of xylose and arabinose.
3.2. VFA and Lactic Acid
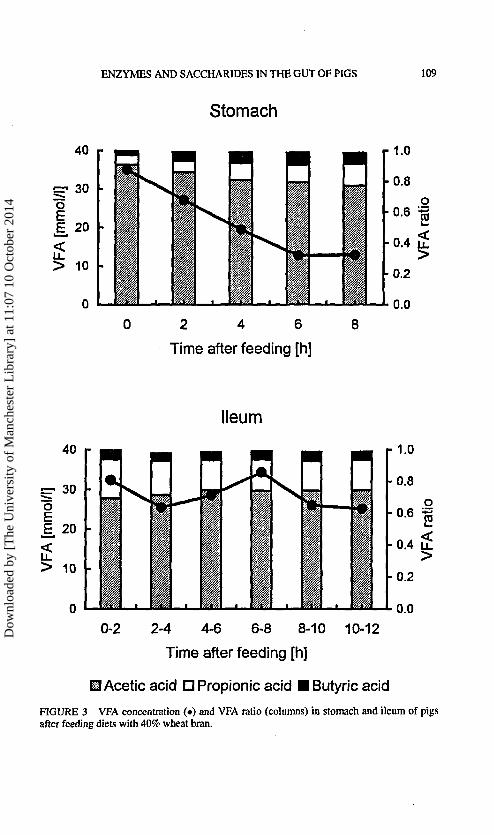
In the stomach, highest VFA concentration was measured shortly after feed-ing and thereafter the VFA concentration declined with time (Fig. 3). Therelative contributions of acetic and propionic acids were inversely related.Shortly after feeding, acetic acid accounted for 0.88 of VFA and this propor-tion declined to 0.76 at 8 h after feeding, while shortly after feeding propionicacid accounted for 0.06 and this proportion increased to 0.14 at 8 h after feed-ing. Butyric acid accounted for 0.03 of VFA shortly after feeding and thisproportion increased to 0.07 at 8 h after feeding. From 2 h after feeding also
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4
ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 109
Stomach
«tu
= • 3 0oE 20
n
-
II
m ni
m
2 4 6
Time after feeding [h]
1.0
0.6 •
0.4
0.2
0.0
CD
Ileum
40
=7 30
0-2 2-4 4-6 6-8 8-10 10-12
Time after feeding [h]
Ü Acetic acid D Propionic acid • Butyric acid
FIGURE 3 VFA concentration (•) and VFA ratio (columns) in stomach and ileum of pigsafter feeding diets with 40% wheat bran.
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4
110 S. VAN DER MEULEN et al.
Stomach
Q .
2 3 4
Time after feeding (h)
lleum
360
240
320
o
O
- 280
oinO
240
0-2 2-4 4-6 6-8 8-10
Time after feeding (h)
10-12
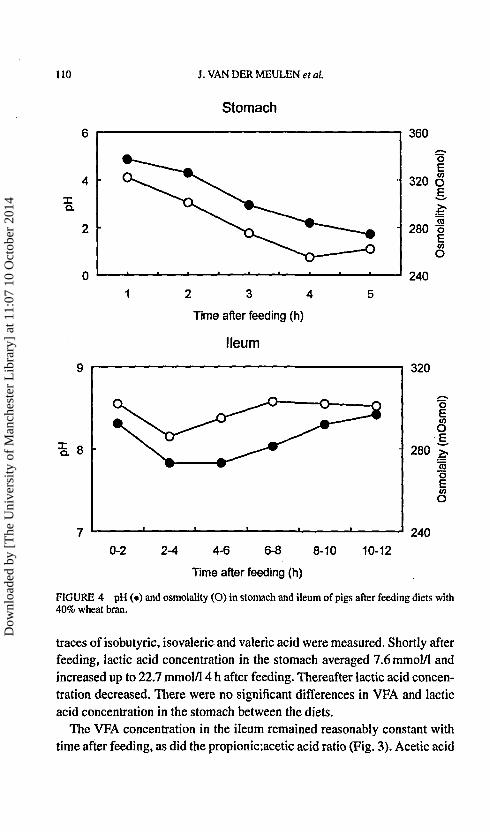
FIGURE 4 pH (•) and osmolality (O) in stomach and ileum of pigs after feeding diets with40% wheat bran.
traces of isobutyric, isovaleric and valeric acid were measured. Shortly afterfeeding, lactic acid concentration in the stomach averaged 7.6 mmol/1 andincreased up to 22.7 mmol/14 h after feeding. Thereafter lactic acid concen-tration decreased. There were no significant differences in VFA and lacticacid concentration in the stomach between the diets.
The VFA concentration in the ileum remained reasonably constant withtime after feeding, as did the propionic:acetic acid ratio (Fig. 3). Acetic acid
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 111
accounted for 0.73, propionic acid for 0.20 and butyric acid for 0.05 ofVFA, with small traces of isobutyric, isovaleric and valeric acid. From 4 hafter feeding the VFA concentration tended to be higher for diets Cel-/ andXyl-i. In the first 2 h after feeding lactic acid concentration in the ileumaveraged 2.4 mmol/1. Highest concentrations were measured 4 to 6 h afterfeeding, after which the concentration declined to 3.5mmol/I 10 h afterfeeding. There were no differences in lactic acid concentration betweenthe diets.
3.3. Stomach and Intestinal Conditions
There were no significant differences in pH in stomach and ileum between thediets. Shortly after feeding pH in the stomach averaged 4.9 and decreasedthereafter with time to 1.7 at 8 h after feeding (Fig. 4). In the ileum pHaveraged 8.3 in the first 2 h after feeding and declined to 7.8 in the next4 h (Fig. 4). Thereafter, pH returned to 8.4 in the period of 10 to 12 h afterfeeding.
There were no significant differences in osmolality in stomach and ileumbetween diets. In the stomach, osmolality ranged from 311 to 335 mOsmolimmediately after feeding and declined to 237 to 269 mOsmol at 6 h afterfeeding and was not changed 2 h later (Fig. 4). In the ileum osmolality didnot change in time and averaged 298 mOsmol (Fig. 4).
4. DISCUSSION
Besides increasing the amount of soluble glucose, incubation of wheat branwith the cellulase preparation increased the amount of soluble arabinoxylans.Since about two-thirds of all soluble saccharides were monosaccharides, theaction of the cellulase preparation was not limited to a primary attack on •the cell wall (polysaccharidase and glycan hydrolase; Chesson, 1993), butalso reduced oligosaccharides to their monomeric compounds (glycosidase;Chesson, 1993).
The cellulase preparation is not only active at pH 5 (during the incubations)but at a wider range of pH as found in the stomach (Inborr et al., 1999).This resulted in an increase in the concentration of soluble saccharides inthe stomach, both when the wheat bran was pre-treated with the cellulase
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

112 J. VAN DER MEULEN et al.
preparation as when the cellulase preparation was added to the diet. However,reductions of released oligosaccharides in the stomach seemed to be lessthan during incubation, since in the stomach only about half of all solublesaccharides were monosaccharides.
When the wheat bran was pre-treated with the xylanase preparation theamount of soluble saccharides and the concentration of soluble saccharidesin the stomach was less compared with pre-treatment of wheat bran with thecellulase preparation. This may be expected, since not only the ß-glucanaseactivity but also the xylanase activity of cellulase-treated wheat bran washigher than of xylanase-treated wheat bran, notwithstanding the aim to sup-ply same amounts of xylanase activity (Inborr et ah, 1999).
The ratio xyloserarabinose in the ileum was lower than in the stomach (seeFigs. 1 and 2). This may be caused by a difference in absorption. Xylosedisappears almost completely from the small intestine (Schutte et ah, 1991 ),while 30% of ingested arabinose enters the large intestine (Schutte et al.,1992). Moreover not only xylose and arabinose but a proportion of all sac-charides including glucose entered the large intestine, as was demonstratedbefore for wheat bran (Graham et al., 1986). The slightly higher arabinoseconcentration found in the urine after feeding diet C, may reflect an effectof incubation of the wheat bran. The higher urinary excretion of monosac-charides for the cellulase-treated diets and for diet Xyl-i reflect the effectof these cell wall degrading enzymes, since in pigs both for xylose (Schutteet al, 1991) and arabinose (Schutte et al., 1992) a dose dependent urinaryexcretion exists.
The higher concentration of soluble saccharides in the stomach for thecellulase-treated diets did not affect fermentation in the stomach as measuredby VFA and lactic acid concentrations. VFA and lactic acid concentrationsin the stomach approximated the concentrations measured by Argenzio andSouthworth (1975) and Clemens etal. (1975). A change in the ratio of aceticand propionic acid with time has not been reported before, and it is not knownwhether this is characteristic for fermentation in the stomach or specific forwheat bran.
Pre-treatment of wheat bran with the enzyme preparations lowered theNDF content with up to 12% but did not affect pH in the stomach, althoughit has been reported that pH in the stomach is higher for low-fibre than forhigh-fibre diets (Lawrence, 1972; Argenzio and Southworth, 1975; BachKnudsen et al., 1991; van der Meulen et ah, 1991). A gradual decrease inpH of the stomach contents with time after feeding was reported before
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 113
(Lawrence, 1972; Argenzio and Southworth, 1975; Clemens et al, 1975),but the rapid increase of pH just before and during feeding (Lawrence,1972) was not measured in our study as a consequence of the samplingtimes.
Although in vitro incubations of ileal contents indicate that mono- andoligosaccharides stimulate lactic acid fermentation but not VFA fermentation(Dierick and Decuypere, 1995), significant higher ileal VFA concentrationsand ileal VFA flows were measured for diets containing large amounts ofL-arabinose (100 g/kg diet; Schutte et al, 1992) or D-xylose (200 g/kgdiet, Schutte et al, 1991). In our study the amount of saccharides releasedfrom NSP was substantially lower than the amount of xylose or arabinoseincluded in the diet by Schutte et al (1991, 1992) and there was not morethan a tendency to increased VFA concentrations for the diets incubated withthe cellulase or the xylanase preparation, while Yin et al (1997) reportedno significant effect of xylanase addition on ileal VFA production in 20 kgpigs. The relatively constant VFA concentration in the ileum measured withtime after feeding is in agreement with VFA concentrations measured byArgenzio and Southworth (1975) and Clemens et al (1975).
The higher microbial activity on diets with large amounts of D-xylosenot only significantly increased ileal VFA flow but also decreased ileal pH(Schutte et al, 1991). In the present study no significant change in pH wasmeasured, like in piglets fed diets with and without ß-galactosidase (Veldmanetal, 1993). This may be caused by the relatively low amount of saccharidesreleased from the wheat bran, as mentioned before. The pH in the ileum incourse of time after feeding was similar to that reported for weanling pigs(Gabert and Sauer, 1995). The change in pH with time after feeding in theileum is limited, since post-feeding effects in the ileum are less evidentthan in the duodenum by effective buffering of the digesta by intestinalsecretions (Braude et al, 1976). The pH level in the ileum measured in ourstudy was relatively high, not only with respect to weanling pigs (Gabertand Sauer, 1995), but also with respect to growing pigs fed high fibre diets(Bach Knudsen etal, 1991; van derMeulen etal, 1991).
The presence of soluble saccharides did not affect the osmolality in theileum as suggested by Wiggins (1984). Also, Veldman et al. (1993) didnot find any affect on osmolality in piglets fed diets with and withoutß-galactosidase. Although the concentration of soluble saccharides in theileum was higher for (some of) the enzyme-treated diets, such an increase isvery limited with respect to all solutions in the ileum. Moreover, any effect
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

114 J.VANDERMEULENefai.
of soluble saccharides on osmolality may have been compensated by fluidretention.
From this study it can be concluded that using cell wall degradingenzyme-preparations may increase the amount of soluble saccharides instomach and small intestine. This effect depends upon the nature of theenzyme preparation used and the way of enzyme application. However, pHand osmolality in stomach and small intestine are not affected, but using cellwall degrading enzyme preparations may increase VFA fermentation in thesmall intestine.
Acknowledgements
The authors would like to thank Mr. R. A. Dekker, Mr. J. Veldhuis and thestaff of the metabolism unit and the laboratory for assisting with the samplecollection and analysis. This investigation was partly financed by the DutchFund for Manure and Ammonia Research, Wageningen, the Netherlands andFinnfeeds International Ltd., Marlborough, United Kingdom.
References
Annison, G. (1992) Commercial enzyme supplementation of wheat-based diets raises ilealglycanase activities and improves apparent metabolisable energy, starch and pentosandigestibilities in broiler chickens, ,4mm. Feed Sci. Technol., 38, 105-121.
Annison, G., Choct, M. and Cheetham, N. W. (1992) Analysis of wheat arabinoxylans from alarge-scale isolation. Carbohyd. Polym., 19, 151-159.
Argenzio, R. A. and Southworth, M. (1975) Sites of organic acid production and absorption ingastrointestinal tract of the pig. Am. J. Physiol., 228, 454-460.
Bach Knudsen, K. E. and Hansen, I. (1991) Gastrointestinal implications of wheat and oatfractions. 1. Digestibility and bulking properties of polysaccharides and other major con-stituents. Brit. J. Nutr., 65, 217-232.
Bach Knudsen, K. E., Jensen, B. B., Andersen, J. O. and Hansen, I. (1991) Gastrointestinalimplications of wheat and oat fractions. 2. Microbial activity in the gastrointestinal tract.Brit. J. Nutr., 65, 233-248.
Braude, R., Fulford, R. J. and Low, A. G. (1976) Studies on digestion and absorption in theintestines of growing pigs. Measurements of the flow of digesta and pH. Brit. J. Nutr., 36,497-510.
Chesson, A. (1993) Feed enzymes. Anim. Feed Sci. Technol., 45, 65-79.Clemens, E. T., Stevens, C. E. and Southworth, M. (1975) Sites of organic acid production and
pattern of digesta movement in the gastrointestinal tract of swine. J. Nutr., 105, 759-768.Dierick, N. A. and Decuypere, J. A. (1994) Enzymes and growth in pigs. In: Cole, D. J. A.,
Wiseman, J. and Varley, M. A. (Eds.), Principles of Pig Science, Nottingham UniversityPress, Nottingham, pp. 169-195.
Dierick, N. A. and Decuypere, J. A. (1995) Advances in the use of enzymes in pig nutrition. In:van Hartingsveldt, W., Hessing, M., van der Lugt, J. P. and Somers, W. A. C. (Eds.), Second
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4

ENZYMES AND SACCHARIDES IN THE GUT OF PIGS 115
European Symposium of Feed Enzymes, TNO Nutrition and Food Research Institute, Zeist,pp. 23-29.
Gabert, V. M. and Sauer, W. C. (1995) The effect of fumaric acid and sodium fumarate sup-plementation to diets for weanling pigs on amino acid digestibility and volatile fatty acidconcentrations in ileal digesta. Anim. Feed Sci. Technol., 53, 243-254.
Graham, H., Hesselman, K. and Aman, P. (1986) The influence of wheat bran and sugar-beetpulp on the digestibility of dietary components in a cereal-based pig diet. J. Nutr., 116,242-251.
Inborr, J., Puhakka, J., Bakker, J. G. M. and van der Meulen, J. (1999) ß-glucanase and xylanaseactivities in stomach and ileum of growing pigs fed wheat bran based diets with and withoutenzyme treatment. Arch. Anim. Nutr., 52, 263-274.
Lawrence, T. L. J. (1972) The effect of certain dietary factors on in vivo pH changes and pepsinactivity in the stomach of the growing pig. Brit. Vet. J., 128, 402-411.
Low, A. G. (1985) The role of dietary fibre in digestion, absorption and metabolism. In:Just, A., Fernandez, J. and J0rgensen, H. (Eds.), Proceedings 3rd International Symposiumon Digestive Physiology in the Pig, National Institute of Animal Science, Copenhagen,pp. 157-179.
Nahm, K. H. and Carlson, C. W. (1985): Effects of cellulase from Trichoderma Vtride onnutrient utilisation by broilers. Poult. Sci., 64, 1536-1540.
Neudoerffer, T. S. and Smith, R. E. (1969) Enzymic degradation of wheat bran to improve itsnutritional value for monogastrics. Can. J. Anim. Sci., 49, 205-214.
Payne, R. W., Lane, P. W., Ainsley, A. E., Bicknell, K. E., Digby, P. G. N., Harding, S. A., Leech,P. K, Simpson, H. R., Todd, A. D., Verrier, P. J., White, R. P., Gower, J. C , TunnicliffeWilson, G. and Paterson, L. J. (1987) Genstat 5 Reference Manual, Oxford UniversityPress, Oxford.
Robinson, P. H., Tamminga, S. and van Vuuren, A. M. (1986) Influence of declining level offeed intake and varying the proportion of starch in the concentrate on rumen fermentationin dairy cows. Livest. Prod. Sci., 15, 173-189.
Schutte, J. B., van Kempen, G. J. M. and Hamer, R. J. (1990) Possibilities to improve the utili-sation of feed ingredients rich in non-starch polysaccharides for poultry. In: Proceedings8th European Poultry Conference, Barcelona, Spain, pp. 128-135.
Schutte, J. B., de Jong, J., Polziehn, R. and Verstegen, M. W. A. (1991) Nutritional implicationsof D-xylose in pigs. Brit. J. Nutr., 66, 83-93.
Schutte, J. B., de Jong, J., van Weerden, E. J. and Tamminga, S. (1992) Nutritional implicationsof L-arabinose in pigs. Brit. J. Nutr., 68, 195-207.
van der Meulen, J. and Bakker, J. G. M. (1991) Effect of various sources of dietary fibre onchemico-physical characteristics of digesta in the small intestine of the pig. In: Verstegen,M. W. A., Huisman, J. and den Hartog, L. A. (Eds.), Proceedings 5th InternationalSymposium on Digestive Physiology in the Pig, Pudoc, Wageningen, pp. 440-445.
van Lonkhuijsen, H., van Gelder, A., Jetten, J., Lichtendonk, W. and Cone, J. (1997) 3-StepNSP analysis. Feed Mix, 5, 14-16.
Veldman, A., Veen, W. A. G., Barug, D. and van Paridon, P. A. (1993) Effect of ß-galactosidesin feed on ileal piglet digestive physiology. J. Anim. Physiol. a Anim. Nutr., 69, 57-65.
Wiggins, H. S. (1984) Nutritional value of sugars and related compounds undigested in thesmall gut. Proc. Nutr. Soc., 43, 69-75
Yin, Y. L., McEvoy, J., Schulze, H. and McCracken, K. J. (1997) Effect of levels of dietarynon-starch polysaccharides (NSP) and feed enzymes on digestibility and ileal concen-tration of volatile fatty acids (VFA) in growing pigs. In: Laplace, J. P., Février, C. andBarbeau, A. (Eds.), Proceedings 7th International Symposium on Digestive Physiology inPigs, EAAP Publication No. 88, pp. 502-505.
Dow
nloa
ded
by [
The
Uni
vers
ity o
f M
anch
este
r L
ibra
ry]
at 1
1:07
10
Oct
ober
201
4