Embed Size (px)
Citation preview

Effect sizes of somatic mutations in cancer
Vincent L. Cannataro 1, Stephen G. Gaffney1, Jeffrey P. Townsend 1,2,3†
1Department of Biostatistics, Yale University, New Haven, CT, 2Program in Computational Biology and Bioinformatics and 3Department of Ecology and Evolutionary Biology, Yale University, New Haven, CT †To whom correspondence should be addressed.
1
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Cancer growth is fueled by genomic alterations that confer selective advantage to somatic cells 1. A major goal of cancer biology is determining the relative importance of these alterations. Genomic tumor sequence surveys have frequently ranked the importance of genetic substitutions to cancer growth by P value or a false-discovery conversion thereof 2,3. However, P values are thresholds for belief 4, not metrics of effect 5,6. Their frequent misuse as metrics of effect has often and ineffectively been vociferously decried 5,7–9, even in cases when the only attributable mistake was omission of effect sizes 10,11. Here, we draw upon an understanding of the development of cancer as an evolutionary process 12,13 to estimate the effect size of somatic variants. We estimate the effect size of all recurrent single nucleotide variants in 23 cancer types, ranking their relative importance within and between driver genes. Many of the variants with the highest effect size per tumor, such as EGFR L858R in lung adenocarcinoma and BRAF V600E in colon adenocarcinoma, are within genes deemed significantly mutated by existing whole-gene metrics. Quantifying the effect sizes of somatic mutations underlying cancer has immediate significance to the prioritization of clinical decision-making by tumor boards, selection and design of clinical trials, pharmacological targeting, and basic research prioritization.
2
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Since the advent of whole-exome and whole-genome sequencing of tumor tissues, studies of
somatic mutations have revealed the underlying genetic architecture of cancer 14, producing ordered lists
of significantly mutated genes whose ordering implies their relative importance to tumorigenesis and
cancer development. Typically, differentiation of selected mutations from neutral mutations is performed
by quantifying the over-representation of mutations within specific genes in tumor tissue relative to
normal tissue, and disproportionate prevalence of somatic mutations in a gene has been taken as prima
facie evidence of a causative role for that gene. Two quantifications have implicitly ordered the
importance of discovered cancer “driver” genes: the prevalence of the mutation among tumor tissues
sequenced from that tumor type 15,16, the statistical significance (P value) of the disproportionality of
mutation frequency 2, or both 14. Versions of these metrics have shifted from simple ranks by mutation
prevalence in a tumor population 17–19 to calculation of statistical significance of mutation prevalence over
genome-wide context-specific background mutation rates 20–22, to ratios of the prevalence of
nonsynonymous and synonymous mutations 1,23, to P values based on a gene-specific mutation rate and a
diversity of genomic data 3. Although the approaches used to calculate P values have become more
sophisticated, neither prevalence nor P value is an appropriate metric for quantifying the vital role of
genes or their mutations to tumorigenesis and cancer development.
While prevalence of a somatic mutation in a cancer type has important consequences for biomarker
studies 24 and identification of therapeutic population for a targeted therapy 25, there is only a
correlative—rather than causal—link between prevalence of mutation and its contribution to
tumorigenesis and cancer development. The lack of causal linkage is easily seen by considering the
mutated genes that, in spite of their high prevalence in tumor populations, are universally regarded as
false positives. For example, the gene TTN is a structural protein of striated muscle. Because it is long,
because it replicates relatively late in the synthesis phase of mitosis, and because it is inaccessible to
transcription-mediated repair in non-muscle tissues, it has a high mutation rate and is frequently mutated
3
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

in non-sarcoma cancer tissues. While TTN is an extreme example—sometimes showing up at the top of
lists ordered by prevalence of genic mutation 1,3—it exemplifies the problem with using mutation
prevalence as a proxy for importance. Any consideration of whether mutated genes are contributing to
tumorigenesis and cancer development—or of the degree to which they are contributing—must address
the issue of their underlying mutation rate.
The appearance of such biologically implausible genes in ranked mutation lists prompted the
development of increasingly sophisticated statistical approaches designed to “weed out” false-positives
via calculation of a P value that accounted for gene length and background mutation rate. The classical
evolutionary biology approach is to use the frequency of synonymous site mutations in each gene as a
proxy for mutation rate. As in the divergence of species, synonymous site mutations are presumably
neutral (or nearly so) to the success of cancer lineages during the divergence from normal to resectable
tumor. As the number of mutations observed in a given gene is typically much smaller in the somatic
evolution of cancer than is observed in the divergence between most species, use of synonymous sites
within a single gene leads to many genes with zero synonymous mutations in most cancers, and an
ineffective calculation of P value. Alternative approaches currently in use obtain a reasonably robust
estimate of genic mutation rate using correlates such as gene expression levels, chromatin states, and
replication timing, and are largely successful at excluding known false positives 3.
In genomic tumor surveys, the sample size of tumors varies among studies, posing a problem for
comparison of P values within or between cancer types 5,6. An even more serious issue with using P
values for ranking genes or mutations arises from the same source that obviates use of genic mutation
prevalence: the confounding effect of mutation rate. Because the “sample size” of overall mutations in a
gene is dictated by the genic mutation rate, it is much easier for genes with high mutation rates not only to
reach high genic prevalence, but also to reach statistical significance despite small effect sizes. While
approaches accounting for genic mutation rates will eliminate false positives 3, and the P value will serve
4
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

to exclude genes like TTN that have no role in tumorigenesis and cancer development, the rank order by P
value of genes that do have a role in tumorigenesis and cancer development will remain highly affected
by mutation rate. Genes with higher mutation rate will (correctly) be more likely to achieve statistical
significance, and thus will appear deceptively high on a ranked list whose ordering suggests importance in
tumorigenesis and cancer development.
Because genic mutation prevalence and P value inadequately capture importance to tumorigenesis
and cancer development, another metric must be appropriate. To provide an evaluation of the relative
importance of mutations in diverse cancer types to tumorigenesis and cancer development, we called on
an understanding of the development of cancer as an evolutionary process 12,13, and adapted some
straightforward insights from classical evolutionary theory. The cognate metric in evolutionary theory for
quantifying importance to tumorigenesis and cancer development is the selective effect of the mutation on
the cancer lineage. The appropriateness of this metric is fairly easy to recognize. While mutations are the
ultimate source of variation contributing to tumorigenesis, we do not conduct genomic tumor sequence
surveys to discover neutral mutation rates. We conduct them to determine which mutations spread within
cancer tissues because of the effects of mutations on proliferation and survival. Mutation rate is a
confounding phenomenon: when it is high, it also increases prevalence of mutations. Because silent site
substitutions and other correlates of baseline mutation rate provide a means to independently differentiate
silent mutation rate from the impact of natural selection within the tumor, selection intensities can be
estimated, providing the effect sizes of each mutation.
We calculated cancer effect sizes by comparing the rate of observed substitutions to the rate that
substitutions would be expected to arise in the absence of selection 26. In accordance with population
genetic theory, we specify that the rate neutral mutations arise and the rate that they fix as substitutions
within tumors are equivalent 27, and that non-neutral mutations arise at a consistent rate. Thus any
increase in the flux of substitutions among tumors of a particular context above the baseline silent rate
5
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

would be the appropriate estimate of the intensity of selection on that mutation within the tumor
population (Methods).
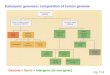
This calculation yielded cancer effect sizes for all fixed substitutions (Methods, Supplemental
Figure 1, Supplemental Table 1) that quantify contribution to the cancer phenotype within 23 tumor types.
Their relative rank corresponds to their relative importance within the respective tumor types. Several
common known oncogenic substitutions, such as BRAF V600E in COAD and EGFR L858R in LUAD,
and substitutions in known tumor suppressor genes, such as APC in READ and TP53 in HPV-negative
HNSCC, are highly selected, and those genes are also determined as significantly mutated by MutSigCV3.
However, genes determined to be significantly altered in cancer by MutSigCV are well-dispersed within a
large range of site-specific cancer effect sizes (Figure 1), illustrating how discrepant Q values are with
cancer effect size. Several substitutions within genes that are not determined to be significantly
over-mutated via MutSigCV are interspersed among more prevalent substitutions within genes that are
estimated to be significantly mutated, for instance Mastermind-like3 (MAML3) G1069A in READ, a
protein that binds to and stabilizes the DNA-binding complex of the Notch intracellular domain 28 and
Nuclear factor (erythroid derived 2)-like 2 (NFE2L2) R34G in UCEC, a protein that is believed to play a
causative role in squamous cell lung cancer 29,30. Indeed, substitutions in this gene comprise two of the top
three most selected substitutions within our analysis of lung squamous cell carcinoma (Figure 1).
6
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Figure 1. Cancer effect sizes of recurrent somatic substitutions in eight of the 23 cancer types analyzed. Effect sizes greater than 1 × 103 are indicated by ticks along the tumor-type axes. The highest 50 effect sizes are labeled within each tumor. Names of genes that have more than one mutation within or between tumors are uniquely colored. Genes deemed significantly burdened with mutation 3 are depicted by a red circle next to mutation labels, and the prevalence of each substitution is represented by the size of this circle. LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma. UCEC: Uterine corpus endometrial carcinoma; LGG: Brain Lower Grade Glioma; HNSC: Head and neck squamous cell carcinoma, broken into HPV positive and HPV negative tumor samples (methods); COAD: Colon adenocarcinoma; READ: Rectum adenocarcinoma. NCSNV refers to a non-coding single nucleotide variant outside an exon (e.g. 5’ or 3’ UTRs). HPV+ and HPV– HNSCC have been demonstrated to have significantly different genetic architectures 31, thus they are presented separately.
7
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Current approaches using conservative P values are particularly underpowered to detect genes that
are of high importance to tumorigenesis and cancer development in some cancer cases because the site or
sites conveying the relevant phenotype are mutated at low rates. For instance, FBXW7 R505G was
estimated to have the highest selection intensity in HPV+ HNSCC, and BRAF V600E was estimated to
have the fifth highest selection intensity in LGG, but both of these genes were classified as not
significantly mutated by MutSigCV within these two cancer types. Mutations within these two
well-known oncogenes were estimated to confer large effect sizes, and these genes were determined to be
significantly mutated in other cancer types, yet their degree of influence within HPV+ HNSCC and LGG
is obscured in cancer-wide significance analyses. The relative effect sizes of cancer mutations can inform
nearly every aspect of basic research related to oncology and should play key roles in clinical
decision-making. They should be integrated into clinical decision-making in tumor boards, where they
would provide crucial insight into the relative upper limits of the efficacy of precision-targeted treatments.
They should guide the selection of clinical trials to target small populations that can benefit from targeted
therapeutics developed for other cancer types. They should guide targeted development of new
therapeutics, indicating the upper limit of effect for a perfect therapeutic ameliorating an oncogenic
mutation. Lastly, they should guide the selection of important areas of basic research that have potential
to lead to therapies and cures for cancer. It does not escape our notice that here we only calculate the
effect size of single nucleotide variants. Importantly, effect sizes of other mutational processes, such as
copy number alterations or epigenetic modifications, could be similarly calculated once estimates of
intrinsic mutation rate in the absence of selection are identifiable for these processes.
8
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

METHODS:
Data acquisition and processing:
All data were obtained either from The Cancer Genome Atlas (TCGA) projects downloaded from
the National Cancer Institute’s Genomic Data Commons 32, or from projects part of the Yale-Gilead
collaboration (Supplemental Table 2). All TCGA data used in this analysis were from GDC version
gdc-1.0.0 and relevant UUID are found in Supplemental Table 3.
All TCGA data were first converted to hg19 coordinates using the liftOver function of the
rtracklayer R package33. To obtain consistency between gene symbols used in MutSigCV and in
mutation data, symbols in the MutSigCV default covariates files were mapped to HUGO symbols.
Non-HUGO symbols were mapped using substitutions from the NCBI Gene database using unambiguous
'synonym' matches; unambiguous 'previous symbol' matches; and manual lookups. CDS coordinates for
each gene were obtained from UCSC's hg19 annotation database. The MutSigCV covariates files, and
gene annotation files used in our analyses can be found at
https://github.com/Townsend-Lab-Yale/SNV_selection_intensity. Nucleotide variants one or two
positions apart in the same tumor sample were removed from the analysis to ensure that we analyzed only
single nucleotide variants. Head and neck squamous cell tumors were designated as HPV positive if they
contained greater than 100 HPV RNA viral transcript reads per hundred million (RPHM) 34 and were
designated positive in clinical data obtained from The Broad GDAC Firehose 35, and tumors were
designated HPV negative if they contained less than 100 HPV RNA viral transcript RPHM and were
designated negative in clinical data obtained from The Broad GDAC Firehose.
Calculating mutation rate and selection intensity:
We define the selection intensity of a single nucleotide variant as the ratio of the flux of fixed
mutations to the expected flux of fixed mutations in the absence of selection. We estimate the expected
rate of fixed mutations in the absence of selection by using MutSigCV to calculate the silent mutation rate
9
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

for each gene. We use the median gene expression from cell lines derived from each analyzed tissue type
(Supplemental Table 3) as the MutSigCV expression covariate for that tissue type. We then normalized
this gene-level rate at every site for every mutation across each gene given the specific trinucleotide
mutation profile of the tissue, such that the average rate of mutations among all trinucleotide
combinations in each gene is equal to the average rate calculated by MutSigCV. The trinucleotide profile
of a tissue is calculated as the average of trinucleotide COSMIC signatures among non-recurrent
mutations calculated by the deconstructSigs R package 36 for all tumors with over 50 single
nucleotide variants.
Formally, we define the rate of non-neutral substitutions, as , where N is the,λ μ u(s)N ×
effective population size of the tumor, is the mutation rate of a substitution, and is the probabilityμ (s)u
the mutation fixes within the population, which is a function of the selection coefficient of the mutation, s .
The probability that a silent mutation fixes is the inverse of the population size within which it fixes, and
thus rate of non-neutral substitutions, divided by the rate of neutral substitutions, is or,λμ = Nμ × 1
N
Nμ × u(s)
, the selection intensity and effect size of the mutation. The term selection intensity is used as a(s)N × u
direct parallel to the classic derivation of “scaled selection coefficient” or “selection intensity” in the
population genetics literature 37–40. Because a tumor sequence is a single snapshot in time, we can only
detect one substitution event per site per tumor. To correct for this issue of detection, we define the rate of
substitution, , as the Poisson rate of occurrence of one or more observed fixation events, i.e. the value ofλ
that maximizes , where is the number of tumors without any substitutionλ e ( −λ) n0 × 1 ( − e −λ) n1 n0
at that site and is the number of tumors with the specified substitution at that site. We definen1
substitutions as single nucleotide variants that are observed in tumors from more than one patient
(recurrent) within our dataset, and we calculate the selection intensity of the recurrently mutated
substitutions to minimize the probability of analyzing passenger mutations.
10
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Scripts used to perform this analysis are available online at
https://github.com/Townsend-Lab-Yale/SNV_selection_intensity
References cited
1. Greenman, C. et al. Patterns of somatic mutation in human cancer genomes. Nature 446, 153–158
(2007).
2. Kan, Z. et al. Diverse somatic mutation patterns and pathway alterations in human cancers. Nature
466, 869–873 (2010).
3. Lawrence, M. S. et al. Mutational heterogeneity in cancer and the search for new cancer-associated
genes. Nature 499, 214–218 (2013).
4. Evans, D. M. & Purcell, S. Power Calculations in Genetic Studies. Cold Spring Harb. Protoc. 2012,
db.top069559 (2012).
5. Nakagawa, S. & Cuthill, I. C. Effect size, confidence interval and statistical significance: a practical
guide for biologists. Biol. Rev. Camb. Philos. Soc. 82, 591–605 (2007).
6. Hojat, M. & Xu, G. A Visitor’s Guide to Effect Sizes – Statistical Significance Versus Practical
(Clinical) Importance of Research Findings. Adv. Health Sci. Educ. Theory Pract. 9, 241–249
(2004).
7. Gardner, M. J. & Altman, D. G. Confidence intervals rather than P values: estimation rather than
hypothesis testing. Br. Med. J. 292, 746–750 (1986).
8. Goodman, S. A dirty dozen: twelve p-value misconceptions. Semin. Hematol. 45, 135–140 (2008).
9. Hayat, M. J. Understanding statistical significance. Nurs. Res. 59, 219–223 (2010).
10. Sullivan, G. M. & Feinn, R. Using Effect Size-or Why the P Value Is Not Enough. J. Grad. Med.
Educ. 4, 279–282 (2012).
11
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

11. Chavalarias, D., Wallach, J. D., Li, A. H. T. & Ioannidis, J. P. A. Evolution of Reporting P Values in
the Biomedical Literature, 1990-2015. JAMA 315, 1141–1148 (2016).
12. Nowell, P. The clonal evolution of tumor cell populations. Science 194, 23–28 (1976).
13. Crespi, B. & Summers, K. Evolutionary biology of cancer. Trends Ecol. Evol. 20, 545–552 (2005).
14. Lawrence, M. S. et al. Discovery and saturation analysis of cancer genes across 21 tumour types.
Nature 505, 495–501 (2014).
15. Miki, Y. et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1.
Science 266, 66–71 (1994).
16. Futreal, P. et al. BRCA1 mutations in primary breast and ovarian carcinomas. Science 266, 120–122
(1994).
17. Zang, Z. J. et al. Exome sequencing of gastric adenocarcinoma identifies recurrent somatic mutations
in cell adhesion and chromatin remodeling genes. Nat. Genet. 44, 570–574 (2012).
18. Kumar, A. et al. Exome sequencing identifies a spectrum of mutation frequencies in advanced and
lethal prostate cancers. Proc. Natl. Acad. Sci. U. S. A. 108, 17087–17092 (2011).
19. Zhao, S. et al. Landscape of somatic single-nucleotide and copy-number mutations in uterine serous
carcinoma. Proc. Natl. Acad. Sci. U. S. A. 110, 2916–2921 (2013).
20. Chapman, M. A. et al. Initial genome sequencing and analysis of multiple myeloma. Nature 471,
467–472 (2011).
21. Wood, L. D. et al. The genomic landscapes of human breast and colorectal cancers. Science 318,
1108–1113 (2007).
22. Sjöblom, T. et al. The consensus coding sequences of human breast and colorectal cancers. Science
314, 268–274 (2006).
23. Krauthammer, M. et al. Exome sequencing identifies recurrent somatic RAC1 mutations in
melanoma. Nat. Genet. 44, 1006–1014 (2012).
12
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

24. Lui, V. W. Y. et al. Frequent mutation of the PI3K pathway in head and neck cancer defines
predictive biomarkers. Cancer Discov. 3, 761–769 (2013).
25. Samuels, Y. et al. High frequency of mutations of the PIK3CA gene in human cancers. Science 304,
554 (2004).
26. Cannataro, V. L. et al. Heterogeneity and mutation in KRAS and associated oncogenes: evaluating
the potential for the evolution of resistance to targeting of KRAS G12C. Oncogene in press, (2017).
27. Gillespie, J. H. Population genetics: a concise guide. 2004.
28. Oyama, T. et al. Mastermind-like 1 (MamL1) and mastermind-like 3 (MamL3) are essential for
Notch signaling in vivo. Development 138, 5235–5246 (2011).
29. Sasaki, H. et al. NFE2L2 gene mutation in male Japanese squamous cell carcinoma of the lung. J.
Thorac. Oncol. 5, 786–789 (2010).
30. Cancer Genome Atlas Research Network. Comprehensive genomic characterization of squamous
cell lung cancers. Nature 489, 519–525 (2012).
31. Seiwert, T. Y. et al. Integrative and comparative genomic analysis of HPV-positive and
HPV-negative head and neck squamous cell carcinomas. Clin. Cancer Res. 21, 632–641 (2015).
32. Grossman, R. L. et al. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 375,
1109–1112 (2016).
33. Lawrence, M., Gentleman, R. & Carey, V. rtracklayer: an R package for interfacing with genome
browsers. Bioinformatics 25, 1841–1842 (2009).
34. Cao, S. et al. Divergent viral presentation among human tumors and adjacent normal tissues. Sci.
Rep. 6, 28294 (2016).
35. Broad Institute. Broad Institute TCGA Genome Data Analysis Center (2016): Firehose
stddata__2016_01_28 run. Broad Institute of MIT and Harvard. doi:10.7908/C11G0KM9. (2016).
36. Rosenthal, R., McGranahan, N., Herrero, J., Taylor, B. S. & Swanton, C. DeconstructSigs:
13
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

delineating mutational processes in single tumors distinguishes DNA repair deficiencies and patterns
of carcinoma evolution. Genome Biol. 17, 31 (2016).
37. Sawyer, S. A. & Hartl, D. L. Population genetics of polymorphism and divergence. Genetics 132,
1161–1176 (1992).
38. Bustamante, C. D. Population Genetics of Molecular Evolution. in Statistical Methods in Molecular
Evolution 63–99 (Springer New York, 2005).
39. Innan, H. & Kim, Y. Pattern of polymorphism after strong artificial selection in a domestication
event. Proc. Natl. Acad. Sci. U. S. A. 101, 10667–10672 (2004).
40. Parsons, T. L. & Quince, C. Fixation in haploid populations exhibiting density dependence I: The
non-neutral case. Theor. Popul. Biol. 72, 121–135 (2007).
Supplementary Information available online. Acknowledgements: This project was supported by Gilead Sciences, Inc. We also thank the Yale High Performance Computing Center for providing computational resources. Author contributions : VLC and JPT designed the analyses. VLC performed the analyses with assistance from SGG. JPT, VLC, and SGG wrote the manuscript. Author Information: The authors declare no competing financial interests. Correspondence and requests for materials should be addressed to [email protected].
14
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

Supplemental figure 1 legend: Selection intensities of recurrent somatic substitutions in 23 cancers. BLCA: Bladder Urothelial Carcinoma; BRCA: Breast invasive carcinoma; Cervical squamous cell carcinoma and endocervical adenocarcinoma; COAD: Colon adenocarcinoma; ESCA: Esophageal carcinoma; GBM: Glioblastoma multiforme; HNSC: Head and neck squamous cell carcinoma, broken into HPV+ and HPV- tumor samples using criteria described within the Methods section; KIRC: Kidney renal clear cell carcinoma; LAML: Acute Myeloid Leukemia; LGG: Brain Lower Grade Glioma; LIHC: Liver hepatocellular carcinoma; LUAD: Lung adenocarcinoma; LUSC: Lung squamous cell carcinoma; OV: Ovarian serous cystadenocarcinoma; PAAD: Pancreatic adenocarcinoma; PRAD: Prostate adenocarcinoma; READ: Rectum adenocarcinoma; SKCM: Skin Curaneous Melanoma, broken into primary skin tumors and metastatic skin tumors; STAD: Stomach adenocarcinoma; THCA: Thyroid carcinoma; UCEC: Uterine corpus endometrial carcinoma.
15
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint

I III I III I III I II III I III III IIII IIII II III I II I II IIII I I I II I II IIIII III I I I I II II II III III II III I I IIII I IIIII II I III II III I IIII II I IIIII I I III I I III III II II I II II IIII II II I II II II I II IIII II I II I II III IIII I II II III III I IIII II II II III II IIII I I II III III III II III IIII I II II IIIII I IIIII IIII I I II I II III II I I IIIII I II III II II I II IIII II I II II II I II I III III II III III III I II II III II II III II IIII I I I I I III III II I IIII II II III II II II I II II III II II I III III II IIII I III II III I II I IIII I II II I III II I III III III I III III II I II II II I I III III I IIIIII II I II I III I II III III II II II I II I III IIII III IIII II I
III II IIII I I IIIII II II I I II II III III II II IIIII IIIII I III II I I I I III I III II I II II I II II I II I II II II IIII II II II I III II I II I II II I I IIII I III II I II II I III III I I I III II II II I II III II I II I III III II II I II II III I III III I I II I II III III I I II I I I II II II II II I I III III II II I I II III II I II I I II III I III II III III II III III II III IIIII I I II III II I II IIII IIIII I III IIII I II II III II II IIII II III III II I III I II II II I IIII III III I I II II II II II II II II III I III II II I IIII II I III III II III II III II II III II II I II II I II I III III I II III I
III II III III I III III III I I III II III I III III I I II II IIII III II IIII I II II IIII II III I III I II II II I II II I II I II II I II II II II III III II II III I II II III IIII I III III I II III IIII I I III IIIIIII III I II II I III I II I II I II II II II III I II II II III I I I IIIIII I III II I II I I III III I III I II I I II III III III II III II IIII II I II I II III II III III I II II I II IIII IIII III II I II IIIIII
I I III I III II II I III I III II I I II II IIIIIII I II II II II IIII II III III I I II I IIIII I I II IIII II I I III I II II I II I III III IIIII II II II II III I II I II III II I III II II I II II I II I IIII IIII II II I II III I IIII IIII III II I II I III III II III I IIII I IIII I III III II I I I I II III II II II I I III I III I I II II III III II II I II II I II I I II II III III I II I II II III III IIIII I II III II I I III III II II II II I IIII II IIII II III II II II I II IIII II I III III II I I II III I III I II I I II IIII I I II IIII III IIII I III II I II I II I II III I II II I I II I III I IIII I III II I II I II I III I II I IIII I I I II III IIIII I II I I II II IIIII II III II IIII I IIII IIII II II II I II I I II II I III I I II II II I III III I II I III III I III I II I III II II I IIII II I IIII I IIII II II I III III IIII I IIII I I II I III III II I III IIII III II III II II II II III II I IIII I II II II III II II II III II III I III III II II II III II III I II III II IIII I III
II I IIII III III I IIII I II II II I II II III I III II III I IIIII I II IIII IIII II II IIIIII III I II I II I III II II II II II II II I II II III IIII II II II II I II III I II I III II II II II I II II II II II I II I I II II II III
I IIIII III I I I III I II I II II I III III I I I I II II III II I III I II I I IIII I I II II II I I II I I II II II I III II II IIII I II II II I II III III II II II I II I IIII I II I IIII II II II II I II IIII I I II II IIII II II IIIII I I III III III II I III II III I I II I II II II II II I III II II I III II II II
IIIIII II II II II
I III I I III II III I IIII II II I II IIII II II II II I IIII I I III II III I I II II I I I I II I II III II I II II I I I II III II II III I IIIIII I IIII IIII IIII I II I II I IIII II II II III I II II IIII II I I II IIII IIII IIII II II I II I II I III II III II IIII I II II I I II I II II III III II I I II II I I IIIII I II II I II I III I II
I IIII I II I I II I II II II I I II III I II II II
I II III III I IIII III I II IIII II I II III III
II III I III III II II III I IIII II I III I I III I II II II I III I II I I I IIII III II II II III I I III II II II I I I II I II II I II II II II II II II II II II II III I II II I III II I I II II III
I I I II II I II III II III I I II I II I II II II III II II I III IIII I III I III I II III II I I II II III I III II I III II III II I II II II I IIII II I II I II III I III II I II I II I I IIII II I I II I II II III I II I III IIII III II I IIII I I IIII II II IIII II II II I I II I III II I III III I I III I III I I I IIII I II III III III I II I IIII I I II II I II II I I IIII II I III II I IIII IIII I I I I II I II II II II II III II I III II II I II II III I III III II II III I I IIII I II II I I III II I IIII II II I II II II III II II I II II I II I II III III II III III II I I I II II III III IIIII II II II II III II II I II I III I IIII II I II II II I II I
IIII I III I IIII III I II II II I III II II II II II IIII II I I I III II I II II II II II I III II II I I I II II I II I I IIII I II III I II II II II III II I I II I II I II I II IIII II III II II II I II I II III IIII III II III IIIII I II I III II III I I IIII I II II IIIIII II I II II II II II IIII III II II I II II II II II II II I IIII I II II I III III I I II III I II II I I IIIII I III I II II II II II IIIIII I II II I III II III II I II I II II II IIIII I I III I II I I IIII I II II III II II I II I I II II III II I II II II II III III I III II I III I II I II II III I III II I I II II III I II II II II IIIIII III I I III III I II IIIII I IIIIII I I II IIII IIII III III II I I III II II I III I III II III I II II III I III I I II I I II II II III I II I III II I III III III II I III II II I
II II III III II II I III III III II I I II II I I II III I II II III I III II I I II I II I II II II I I III IIIII I II I II I II II II IIIII II II II II II I II I I IIIII I III II I II IIIII III II I IIII III II I IIIII I II III I II II II IIIIII III I I III II II III II II IIII II I III I I II II II I III III I I II II III II IIII III II II IIII I IIII I II I II I IIII II III II I I III I I II III I IIIII II II I IIIII III IIII II II II III II II I I IIIII III II II III II I III III I I I II I II II II I IIII IIII II IIIII II I II III IIII II II III I III II II II I II II I I I IIII I II I III III III I III II I III I II I I II II II II III I II III II II III II II I I II III II II I II I II II I II II III III II I I IIII II III I III III II II II II II I I II III I II I III I II II I II II
I I III I II II I II III II I I II II IIII IIII III I II II I IIII II I II III II I II II I I III I I I I II I II III II II II I IIII II II II III III III III III II II II I II II II IIII III II II I II II III II III II I II II II III II III I I II I II III I II II II II I III II I II II II II IIIII II I II II IIII I I II I II I II I II III II II II I IIII III II I II II I I III I I II IIII II II II I II IIII II I II II I III I IIII II I II I I II II II II II II III III I I II
I II I I I II III III II I I II II II II II I II III I I I III II I III IIII I I III I III II I I III IIII
I II III I II II II IIIII II III I II II II II III III II I I III II I II I II I I III III IIII III II II III
III II I II IIIII I I I I IIIII III I I III II III I II IIIIII I III I II III II I I II I II I III II I I III I IIII II I II I II II I III II I II I IIII II II I II II III IIII III I I I I III II I III I I III IIIII III II II I I II II II I II II IIII III I IIII III I II II II III III II I II II II II II I II I III II II III I II II II II I II II IIII I II II III II IIIII II III I III IIII I I III I IIII II II II IIII IIII I II IIIII II II II II II I III I I I III III I II II III I III I III III IIII III II II I II I II II II II II II II II II I III III II I III II II II II II II I I III III I III II II II I II II II II II III I I II I III III I II II II II II II I III II II III II III II I II II I I I II II II II I II II II I III I II I II I I I II I II II II I III I
I III II II IIII III II II III IIII IIII III I I IIII IIIIII I II II II I III II III II II III II I I II I I I IIII III II I II I II II III II I II III III II I II I I III II I III II II II II I II II IIIII II II II II III II II I I II I IIIIII II I III II II II I II I III III II I II II II II I III IIIII II II II III I II II I II II I II II IIII I III II I II II III IIII II I III I II I I IIII II I II II II II IIII I III I II I III II II I II III I I II II II I II IIIII I I IIII II II II I I II I IIIII II II IIII I III II I III III I I III I II II III II II I II II III I I III II III I I II II III I II II II III II II I I IIII III I II II III I II III I III I I II II I I II II I II III II I IIII III III I I II II II I I I I II II III I II I III I II I I I I IIII I II I II II II I IIII I III II I II I I II IIII III IIII III II I II II II I II IIIII III I I IIIII II I II I III II III I II II II III II II II II IIIII I I I III I III III I II II I I I II II II III II II III II I I I I II III I II III II I III II III III III I II II I IIII III II I III II I III I II II I IIII III I II I II II II IIIII IIII II II II III IIII II II II I II II II II I II I II I I III II I II I III III II I IIII I I III III II III II IIIII I II II I I IIIII I I III I IIII II III IIII I II I III I I III III I IIII II III IIIII II II II II II III II IIII II III II II II I II II I II I III II II II I III I I II I I III I II III III II III I II I II I II II II II IIII III I III III I IIII I II II III I II II III II I II II II III I IIII II I III II II I I II III I I II I I III I III I III III II I III I II III III II I II III III II II I II IIII I II II III I III II II II III IIIII I I IIII III I I II I IIIII II II I II II II II II I I II I III II I II I II II I I III II II II I III II I II II I II III III I II I II I IIII I III I I IIII II I II III I II I II II I IIIII III I IIII I I I II II III IIII III II III III II I III I I III II II II II II III I II I III II I II II II II II II II II IIII I I II II I II II I II II IIII II II II I III II IIII III IIII I I III I II III III II I III I III I III II II II I II I II III III II I IIIII II III III I II I III I IIIII II I II I III II IIII II III IIII I III IIII I III I III III I II III I III II II III III III II II I IIIII II I III I I I II I II I II IIII I I II II II I II III III II III II II I II I II II II I I II II IIII II III I I III I II II I I II II I II II II III II II II II I I II III II II II I IIII II I III IIII III I III III II II II IIII II IIII II I I
II I I II I II I II III I I III II I II I III II IIII III I II IIIII I II I I III I I III I I I I II II I I II I II II III III II II III II III II II IIIII I I II I III I II III I II II III II I II III I II III II II III II II I II I II I I I III III II IIII I II I
I II II I II II I III IIII IIIII II II II II I I II IIII II III II III II II IIII II III I IIIII I III III I I IIII I I II I IIII II IIII II II I III IIII III II II II I IIII I I II I I I II II II II I I IIIII III I II III I II I I I II II I II II I II II II IIIIIII I II III I IIII II III III I III III III II II I II II II II I II I II I IIII II I I II II II I IIII II II I II II III II II II I II I III IIII III I I I IIII I III II I I IIII I III II II II II I II I III III II II IIII I IIII I II III III I II II I II II I I I III III III III I II III I II II I III IIIII III II II II II II II II II I III I III II II III I II I III I I II III I II I III I II I IIII I III II I IIII II I II II III II III
I II II I II III II IIII III I II I III II III
I I II III II III III I IIII IIII II I II II I III I IIII II II I I II II III III I IIIII III III II I III I II I II I I II I II II III III II II II II I III III I II II II II III I III II I III IIIII II I III III IIII I I III I II I IIII I I II I I II I III I III III III I II II II II I I II I I II II II III III IIIII II I I II III II I II II I II II I III II IIII III I II II IIII I III I II II II II II II III I I II II II I IIII III II I II I II II I II II II II IIIII II II II I III II I II II I II I I I III III III II II I I III III I III II I III II II I I II I II I I III II III II II III II II I III III III III I I I III III III III I I I II II I II II I III I III II III II III II II IIIII I I IIII II II III III II IIII I II III III III III II I II I III I IIII II I I I IIIII I IIII I II I II II II II II III I III II I I III I I II II I I I III III II I II IIII II I I III I III III II III II II II II I IIII II II III I I IIII III II II III II II II III II II II I II II III III II IIII II III III I II II III I III IIIII I I III II III III II I III III IIII IIII I II I I III I II IIII I IIIII I III I III I III II I III II II II I II I IIII I I II II I II I IIIII II I II III IIIII I IIIII III I IIII I III II II I II I II I II II II II I IIII I III I II II I II I II II II II III II III I II III II IIII I II II I III I II I II I II I III II I III I II I II I II I II I II II III III I I III II III IIIII II I III II III III III I II III II I II II I I III II III II IIII II I I II I II II III I I I I IIII I I III I I III III I III II I II III I IIIII I I I IIII I II I III I III III I I II II I II III IIIII III II IIIII IIIII I II II II II III III I I III II III II I IIII II I IIII III I II II II I II I IIII III II II I I I I II I II I I II I II III II IIII IIII III I I III II I II III II I III II I II II I IIIII II III II II I IIIII II III II II III III II I II III I III I II II II II II IIIII II II I II I II II II II I II II II II IIII I I III I III II I I I II II I IIII I III IIIII III I I IIIII III II III II III III I III II I III III II II II IIIII II I II I I III II I I II II II I II
FB
XW
7 R
425G
ZY
G11
B R
257P
HR
AS
G13
R
EIF
3I R
77P
TB
C1D
12 N
CS
NV
TP
53 D
259V
RA
BE
P2
V35
2L
FG
FR
3 S
249C
BIR
C6
R47
13L
SO
RB
S1
G13
3R
ER
BB
3 V
104L
EX
OC
4 S
21*
EV
I2A
NC
SN
V
NF
E2L
2 R
34G
TB
C1D
12 N
CS
NV
HR
AS
Q61
L
WD
R83
E69
Q
FG
FR
3 Y
373C
PR
DM
16 E
271Q
PIK
3CA
H10
47R
PIK
3CA
H10
47L
NC
OA
6 K
905N
TP
53 L
257Q
HAV
CR
2 N
CS
NV
KR
AS
G12
D
GA
R1
R13
P
TP
53 R
248Q
DS
T N
CS
NV
GT
F2I
RD
1 P
307A
ER
CC
2 N
238S
FAM
82B
R6G
SE
NP
3 N
CS
NV
FG
FR
3 S
371C
GN
A13
R20
0G
MD
GA
2 R
955G
C2o
rf16
E26
8K
HR
AS
Q61
K
PS
D3
NC
SN
V
FG
FR
3 K
650E
FG
FR
3 G
370C
GA
D2
NC
SN
V
HIS
T1H
3B R
132G
WN
K1
R34
1G
TT
N N
CS
NV
SC
AM
P1
NC
SN
V
RB
1CC
1 I4
33V
KC
NK
2 S
6W
ELF
3 E
262Q
KD
M6A
NC
SN
V
KR
AS
G12
V
RA
B11
FIP
5 T
246P
MIN
A N
CS
NV
PIK
3CA
H10
47R
C10
orf7
6 Q
267K
US
P40
NC
SN
V
SF
3B1
K70
0E
EX
OC
4 S
21L
TP
53 Y
220S
FAM
189A
2 I2
41I
TP
53 R
273L
PD
E4D
IP N
CS
NV
KIA
A05
28 R
181Q
PLE
KH
M1
NC
SN
V
LDH
AL6
B N
CS
NV
TP
53 R
342P
MY
LK3
NC
SN
V
TP
53 I1
95T
PIK
3CA
H10
47L
ZM
YM
1 N
CS
NV
TP
53 F
113C
CN
TN
AP
3 N
CS
NV
MLL
3 E
640*
ZN
F69
2 N
CS
NV
MY
L5 R
46P
VP
S13
C T
3732
M
TP
53 L
194R
FR
MD
7 N
CS
NV
US
O1
E61
0Q
AK
AP
9 N
CS
NV
HE
ATR
6 L1
62P
PPA
RG
L24
2V
C1o
rf12
9 N
CS
NV
ER
BB
2 L7
55S
GAT
A3
M29
4K
CD
H1
Q23
*
TP
53 Y
220C
TP
53 H
179R
TP
53 H
193R
PIK
3CA
N34
5K
ZN
F61
1 N
CS
NV
SE
C31
A R
242L
ALD
OA
NC
SN
V
SU
PT
6H D
130Y
PIK
3CA
E54
5A
C12
orf6
9 N
CS
NV
TP
53 N
CS
NV
MY
O15
A K
2535
M
TP
53 H
193L
AK
T1
E17
K
STA
R N
CS
NV
FB
XW
7 R
505G
NO
D2
P17
7R
ST
MN
3 N
CS
NV
ALD
OA
NC
SN
V
ZN
F33
5 N
CS
NV
ME
D12
D23
Y
AN
KR
D20
A4
NC
SN
V
GA
BPA
NC
SN
V
HN
RN
PR
NC
SN
V
TH
AP
1 N
CS
NV
FB
XW
7 R
479G
SIG
LEC
7 Q
309R
TP
63 K
188M
CE
LF4
NC
SN
V
TC
F20
NC
SN
V
PP
P1R
16B
NC
SN
V
CA
CN
G7
V19
7G
C12
orf4
3 E
28Q
TR
MT
2A R
167P
LMN
B2
NC
SN
V
RB
M39
NC
SN
V
ZN
F65
8 N
443K
FAM
155A
NC
SN
V
ST
6GA
L1 N
CS
NV
AK
T1
NC
SN
V
PIK
3CA
C42
0R
CC
ND
1 P
287T
HIS
T2H
2AC
R30
P
GT
F3C
4 F
317F
EP
300
W14
36R
PIK
3CA
H10
47R
HIS
T1H
2AL
R30
P
FAT
2 N
CS
NV
RO
CK
1 N
CS
NV
PR
R22
NC
SN
V
NF
E2L
2 E
82D
KR
AS
G12
D
PIK
3CA
E54
5K
KR
AS
G12
V
ZN
F70
6 I8
N
CA
MK
2D N
CS
NV
BT
G1
NC
SN
V
FAM
19A
5 N
CS
NV
OB
P2B
NC
SN
V
NX
F1
NC
SN
V
IQS
EC
2 P
1185
P
C8o
rf59
NC
SN
V
DE
NN
D4A
R81
4H
MA
PK
1 E
322K
ER
CC
6 E
272K
BR
AF
V60
0E
TP
53 A
159P
AB
CF
3 R
269G
VN
1R5
K35
7M
KR
AS
G12
V
KR
AS
G12
A
KR
AS
G12
D
AP
C K
1370
*
CG
RE
F1
NC
SN
V
ZC
3H11
A M
752T
PIK
3CA
E54
5K
PC
BP
1 L1
00Q
AP
C S
940*
PO
LDIP
3 C
320W
RU
FY
3 N
CS
NV
PIK
3CA
H10
47R
KR
AS
G12
R
ITP
RIP
L2 N
CS
NV
TR
ER
F1
V11
66D
MO
RC
2 N
CS
NV
KR
AS
G12
C
AP
C E
1408
*
GN
G11
NC
SN
V
DS
P I8
38S
TM
ED
4 N
CS
NV
ZN
F18
9 R
392I
GP
R98
S49
92A
PIK
3CA
V34
4G
OR
6C65
L11
2I
NP
NT
NC
SN
V
KR
AS
G13
D
PC
DH
11Y
NC
SN
V
SP
DY
E6
D26
1E
AB
CA
6 T
772A
AP
C E
1397
*
NR
AS
G13
R
ME
P1B
K39
6N
GT
F2B
NC
SN
V
TC
F7L
2 S
46*
TM
EM
132E
A11
G
AP
C Y
935*
AP
C Q
1378
*
PT
PR
K N
915T
AP
C E
941*
PIK
3CA
E54
5A
RB
AK
S99
A
CN
TN
AP
3 N
CS
NV
EYA
2 G
77A
DN
AJC
22 H
30Q
NU
P10
7 T
45A
TP
53 R
175G
AD
AM
TS
3 N
CS
NV
AN
XA
8 N
CS
NV
TP
53 N
CS
NV
PIK
3CA
H10
47L
AR
F6
NC
SN
V
CD
C42
BPA
P67
5T
SU
PT
6H D
130Y
ST
6GA
L1 N
CS
NV
RN
F4
NC
SN
V
TP
53 P
278A
TP
53 C
135F
TP
53 Y
220C
ST
RN
3 N
CS
NV
SLC
38A
11 N
CS
NV
TP
53 R
248Q
RB
BP
8 G
810V
TP
53 C
275F
TP
53 C
176F
RE
RE
NC
SN
V
TP
53 E
198*
TP
53 E
294*
TP
53 R
175H
AM
PH
NC
SN
V
SLI
TR
K6
NC
SN
V
TP
53 R
248W
TT
N T
2290
4P
PIK
3CA
E54
5K
TR
IM13
NC
SN
V
KP
NB
1 L5
52F
NT
RK
3 N
CS
NV
AB
CB
1 K
915T
TP
53 R
213*
DN
AH
10 D
1128
Y
PH
AC
TR
1 N
CS
NV
TP
53 R
282W
GB
E1
F55
1V
HD
AC
4 F
746L
TP
53 R
273C
C10
orf7
6 Q
267K
MB
TP
S1
L77M
ER
BB
2 V
777L
TP
53 R
273H
HE
ATR
7B2
K14
96T
ELA
VL3
NC
SN
V
TP
53 R
196*
TP
53 R
342*
AS
PH
NC
SN
V
AS
PH
NC
SN
V
SN
AI1
NC
SN
V
NE
XN
M38
6R
C10
orf3
5 N
CS
NV
LLG
L1 A
462P
CN
R1
Y27
5S
FH
OD
1 N
CS
NV
DH
X40
E12
1*
BR
AF
V60
0E
FH
L1 K
17Q
SIA
H2
NC
SN
V
AC
TC
1 K
330*
TP
53 Y
220C
NR
AS
Q61
L
GA
BR
A6
V32
F
SID
T2
A63
2P
RG
AG
1 N
CS
NV
TP
53 H
179R
LSS
V48
9G
PIG
O N
CS
NV
PIA
S3
NC
SN
V
EG
FR
G59
8V
PR
DM
11 N
182H
PT
EN
G13
2D
EG
FR
T26
3P
LY6E
NC
SN
V
TP
53 Y
234C
PIK
3R1
G37
6R
TP
53 C
238F
PIK
3R1
NC
SN
V
EG
FR
S64
5C
CT
SA
E42
7D
PT
EN
S17
0N
DA
PK
1 V
1134
A
PD
CD
2L E
344*
EG
FR
A28
9D
TP
53 V
216M
TP
53 C
275Y
PIK
3R1
K37
9N
MU
C2
Y19
77Y
TB
X1
D24
7Y
MLX
NC
SN
V
MU
C6
T10
15M
FAM
120A
H91
8Q
TP
53 R
248Q
EG
FR
A28
9V
DQ
X1
R50
5H
FB
XO
33 S
401R
AN
KR
D17
G17
42G
KC
NT
2 Q
1038
H
TP
53 R
175H
PIK
3CA
M1V
FB
XW
7 R
505G
FG
FR
3 S
249C
MY
H7
E14
9K
IFN
GR
1 R
123*
DH
X15
R39
9H
PIK
3CA
E54
5K
EP
300
D13
99N
PIK
3CA
E54
2K
PIK
3CA
R88
Q
MA
PK
1 E
322K
MU
C16
E14
09Q
ALP
K3
Q86
0Q
PC
DH
A6
S60
8S
CO
L6A
6 L1
125L
TP
53 R
283P
TP
53 E
298*
CC
DC
50 N
CS
NV
TP
53 R
110L
TP
53 V
157F
TP
53 R
196P
TP
53 H
193L
TP
53 H
193P
FB
XW
7 R
505G
HR
AS
G13
R
TP
53 L
201*
TP
53 Y
163C
PIK
3CA
H10
47R
TP
53 R
158L
TP
53 R
337L
TP
53 Y
220C
TP
53 Y
236C
TP
53 R
280G
TM
TC
2 T
409R
TP
53 H
179R
HR
AS
G13
V
EP
HA
3 T
802R
TP
53 R
175H
TP
53 N
CS
NV
TP
53 Y
234C
HR
AS
G12
C
TP
53 R
249S
TP
53 R
282W
CD
KN
2A D
108Y
MA
P2
E86
5*
TP
53 E
294*
TP
53 R
213*
EP
HA
2 S
419*
H3F
3B K
37M
TP
53 C
242F
TP
53 C
238F
TP
53 P
151S
TP
53 V
272M
RA
C1
A17
8V
ITG
B1
D15
8N
CD
KN
2A R
80*
TP
53 N
CS
NV
TP
53 R
196*
TP
53 R
248W
TP
53 R
248Q
TP
53 S
106R
TP
53 P
278S
TP
53 R
273C
TP
53 R
306*
AR
HG
AP
9 N
CS
NV
VH
L S
68*
VH
L S
65*
SE
TD
2 W
1782
*
SIP
A1L
3 G
777A
VH
L L1
69P
VH
L N
78D
VH
L I1
51T
ZN
F38
4 N
CS
NV
SP
SB
3 G
241R
VH
L H
115N
HN
F1B
N30
2K
VH
L L1
28H
VH
L L1
84P
VH
L N
CS
NV
VH
L V
130F
VH
L N
CS
NV
VH
L E
173*
VH
L L1
88P
VH
L L1
58V
BA
K1
R76
W
VH
L N
131K
VH
L N
CS
NV
VH
L R
161*
MTO
R T
1977
R
VH
L W
88*
ALL
C T
83M
PB
RM
1 R
710*
GP
M6A
A50
V
BA
P1
M1I
GA
LNT
6 P
311P
ED
IL3
T34
3M
PB
RM
1 R
850*
NX
PH
4 K
272K
KIT
D81
6V
C1o
rf85
NC
SN
V
TE
T1
N11
34I
KR
AS
G12
V
RIM
S4
V86
L
FLT
3 D
835Y
GA
RT
E94
8G
IDH
1 R
132C
NS
MA
F L
710F
HIS
T1H
2AE
K12
8M
CH
ST
15 N
CS
NV
AR
HG
AP
30 H
1042
H
IDH
2 R
140Q
CA
CN
G7
V19
7G
FLT
3 D
835E
ALO
X12
B N
670T
NR
AS
G13
D
GA
LNT
8 N
CS
NV
ELA
C2
NC
SN
V
IDH
1 R
132H
DN
MT
3A R
882C
MTA
2 N
CS
NV
ZD
HH
C13
S24
2G
PR
DM
2 M
1397
L
DN
MT
3A R
882H
NR
AS
G12
D
C3A
R1
I116
T
CD
4 N
CS
NV
RA
PH
1 K
296M
RU
NX
1 R
201*
IDH
2 R
172K
RU
NX
1 R
162G
AS
B17
W38
R
KR
TAP
12−4
A59
T
RB
BP
4 E
330K
IDH
1 R
132G
IDH
1 R
132S
IDH
1 R
132H
EG
FR
R25
2P
BR
AF
V60
0E
TP
53 R
273L
IDH
1 R
132C
GR
IA4
K29
5E
TP
53 R
158S
CIC
R15
12L
TP
53 Q
136E
TP
53 H
179D
EG
FR
R32
4L
TP
53 Y
220C
TP
53 H
179R
TP
53 R
273C
IDH
2 R
172K
NO
TC
H1
R44
8L
PIK
3CA
H10
47R
IDH
2 R
172W
EG
FR
L62
R
PIK
3CA
M10
43V
PIK
3CA
K11
1E
TP
53 F
270S
PIK
3CA
G11
8D
TP
53 T
155N
TP
53 Y
236C
PIK
3CA
E45
3K
EG
FR
G59
8V
TP
53 C
275F
TP
53 H
193R
TP
53 M
237V
TP
53 M
246V
IDH
2 R
172M
PIK
3R1
N56
4D
C10
orf7
6 R
419*
IDH
2 R
172G
TP
53 V
173L
TP
53 Y
163C
TP
53 R
280K
KR
T13
R20
1C
SD
HA
M1V
TP
53 P
151S
NR
AS
Q61
R
PIK
3CA
E54
2K
PT
PN
11 A
72D
TP
53 R
248Q
ZB
TB
20 N
650S
PLC
G1
E11
63K
CN
OT
4 N
463S
BA
Z2A
R16
42L
ATP
9A N
56I
MY
O7A
S63
W
CD
163L
1 L1
296P
DN
M2
E37
8D
CT
NN
B1
I35S
ZN
F26
4 E
368*
FAM
13C
P20
1Q
RA
LGA
PB
R61
1P
NE
B D
4097
H
C14
orf3
9 V
463L
TE
RT
NC
SN
V
CT
NN
B1
H36
P
NX
F1
R34
2G
FO
XO
1 S
22W
NLR
P6
D44
1H
HC
N4
NC
SN
V
CT
NN
B1
D32
V
CT
NN
B1
S45
P
ES
YT
2 R
569H
CT
NN
B1
S37
A
IL17
RE
L C
23W
ATP
1A1
N40
5T
LRR
C16
B N
CS
NV
CT
NN
B1
D32
G
CT
NN
B1
K33
5I
FAM
193A
L85
2F
OD
F2
E67
6A
PPA
RA
I171
L
TP
53 C
277*
FG
D6
A51
3G
DU
SP
8 T
316M
DB
N1
D41
V
FAM
124B
R47
P
BR
AP
L41
6V
CT
NN
B1
S33
C
TR
PC
4AP
L48
1V
GJC
1 A
37P
TP
53 R
249S
TP
53 V
157F
SP
HK
1 P
417A
TP
53 R
248L
KIA
A03
55 Q
121P
PLA
2G3
K30
5*
UG
T2B
10 D
120H
FE
M1C
L26
6V
ALD
H1L
2 F
107S
ZD
HH
C1
A29
1P
US
P49
I345
L
CT
NN
B1
N38
7K
EG
FR
L85
8R
CT
NN
B1
S37
F
FAM
71A
R30
*
WD
R60
E10
6K
WA
PAL
D42
3N
CT
NN
B1
S45
F
CD
1A N
CS
NV
ZM
PS
TE
24 S
261F
CT
NN
B1
S37
C
BR
AF
V60
0E
GN
AL
P72
P
OR
2W5
H21
4Y
KR
AS
G12
A
CT
NN
B1
G34
V
AK
R1B
15 Q
212H
TD
O2
Q19
7H
HE
ATR
1 F
1877
V
RP
L6 K
221Q
SH
AN
K3
G62
1G
MB
OAT
2 L3
23F
KR
AS
G12
D
AG
AP
7 I5
81I
BR
AF
N58
1S
KR
AS
G12
V
KR
AS
G12
C
EG
FR
G71
9A
PF
KF
B1
R18
6L
EG
FR
L62
R
PT
PR
U P
559S
SM
AR
CC
2 E
855K
CP
SF
1 S
285L
BC
OR
N14
59S
BR
AF
G46
9A
BC
L9L
G98
0C
PIK
3CA
N34
5K
PIK
3CA
H10
47R
FLR
T3
G19
7C
C4B
NC
SN
V
SO
S1
N23
3Y
MU
C2
P24
11H
TP
53 A
159P
TP
53 V
157L
FLR
T3
E49
8K
RA
B8A
NC
SN
V
RE
V1
NC
SN
V
PIK
3CA
E54
5K
TT
C17
R41
3L
RA
D50
D69
Y
KE
AP
1 D
479G
AR
HG
EF
9 R
338L
TP
53 Y
234S
NF
E2L
2 R
34P
NF
E2L
2 R
34G
STO
N1
NC
SN
V
TP
53 V
157F
FR
MD
4B R
900L
TP
53 R
175G
TP
53 E
298*
TP
53 R
158L
TP
53 G
245C
SLC
24A
1 V
455G
ME
GF
8 R
2514
L
CD
KN
2A D
108Y
TT
N R
1073
0G
TP
53 R
273L
TP
53 Y
163C
MD
GA
2 T
517P
MID
N E
122Q
TP
53 Y
234C
TP
53 Y
220C
HO
ME
Z L
306P
TP
53 A
159P
TAF
9 N
CS
NV
NP
R2
E21
1*
LOX
HD
1 T
1789
N
PR
KA
G1
NC
SN
V
DS
T N
CS
NV
CH
D8
E59
9K
CD
KN
2A D
84G
TP
53 N
CS
NV
IQS
EC
2 G
613C
CO
L5A
2 G
936W
DD
R2
D17
1Y
FB
XW
7 R
505G
TP
53 T
125T
TP
53 N
131I
CD
KN
2A D
84Y
NF
E2L
2 G
31A
RO
RC
H49
0P
TP
53 R
248L
C20
orf9
4 T
180R
CLN
3 G
206S
TP
53 Y
236C
TP
53 K
305*
TP
53 Q
144P
C11
orf6
3 K
685M
SLC
45A
2 S
97C
EP
300
D13
99N
TP
53 R
282W
SE
RP
INA
11 R
150P
MO
N2
NC
SN
V
MG
RN
1 G
64A
CO
L14A
1 N
CS
NV
BD
NF
NC
SN
V
PIK
3CD
V23
3G
TOP
BP
1 N
CS
NV
CA
SD
1 N
CS
NV
TP
53 V
157F
TP
53 I2
51S
CC
DC
66 Q
379E
TP
53 I1
95T
RS
F1
NC
SN
V
TP
53 R
273L
RP
S16
NC
SN
V
PD
ILT
Y47
6*
GA
BPA
NC
SN
V
FR
MD
7 N
CS
NV
MA
P2
NC
SN
V
TOP
2A T
215P
PIK
3R4
G60
9D
MA
ML3
G10
69A
CY
P4F
2 A
182G
TP
53 Y
163C
TP
53 Y
220C
SY
TL4
NC
SN
V
TP
53 R
175H
LDH
AL6
B N
CS
NV
UX
S1
V10
5L
TP
53 R
248Q
TP
53 L
194R
NR
IP1
NC
SN
V
TP
53 R
248W
FAM
83H
P70
A
PR
KG
2 N
CS
NV
TF
M45
I
FB
XO
10 S
950T
UT
P20
L38
4H
PN
N S
443A
LRR
C16
B T
804P
UB
E3C
NC
SN
V
SO
RB
S1
NC
SN
V
NR
AS
Q61
R
TP
53 Y
234C
TP
53 S
241F
TP
53 M
237K
RP
F1
F26
7S
TP
53 Q
192*
C1o
rf12
9 N
CS
NV
TP
53 N
CS
NV
TP
53 P
177R
KR
AS
G12
R
KR
AS
G12
V
KR
AS
G12
D
KR
AS
Q61
H
TP
53 V
143E
KR
AS
Q61
R
EE
F2
Y45
7*
TP
53 Y
220C
TP
53 C
275F
TP
53 H
179R
TM
EM
218
NC
SN
V
TP
53 E
294*
IGF
N1
S19
76S
ATP
1B1
E15
4E
TP
53 H
179Y
CD
KN
2A H
83Y
TP
53 R
248W
TP
53 R
213*
TP
53 R
282W
SM
AD
4 S
144*
SM
AD
4 E
520*
TP
53 R
248Q
AB
CA
13 A
2368
V
CD
KN
2A R
80*
SM
AD
4 Q
289*
TP
53 R
175H
GN
AS
R84
4C
TP
53 R
196*
TP
53 G
245S
SM
AD
4 R
361C
TP
53 R
273C
PLE
KH
H1
V86
6M
GN
AS
R84
4H
PX
DN
L R
235Q
TP
53 R
306*
TP
53 R
337C
TR
A2A
R94
*
CD
KN
2A R
58*
CO
L5A
1 P
657L
PLE
KH
A4
R22
5W
TP
53 R
273H
AB
CG
4 T
494M
SLC
12A
6 R
319H
TG
FB
R1
R48
7Q
EH
BP
1L1
A97
7A
OD
Z3
R22
28H
TT
N N
2692
7N
CD
KL4
V10
6I
ER
BB
2 V
842I
RS
PO
1 A
146A
SP
OP
Y87
S
SP
OP
W13
1G
SP
OP
F13
3C
SP
OP
F13
3V
SP
OP
F10
2V
CU
L3 M
305R
SP
OP
F10
2C
SP
OP
Y87
N
TP
53 Y
163H
SP
OP
F10
2S
TP
53 C
141G
SP
OP
F13
3I
RH
OA
L22
R
SP
OP
F13
3L
PC
DH
19 R
395*
SP
OP
Y87
C
SP
OP
W13
1L
TP
53 N
239D
BR
AF
G46
9A
SP
OP
W13
1R
ME
D12
V12
23L
HR
AS
Q61
R
CT
NN
B1
T41
A
TP
53 G
279E
ME
D12
L12
24F
MLL
3 Y
4440
C
AC
P1
NC
SN
V
PIK
3CA
H10
47R
TP
53 R
248Q
PIK
3CA
E54
2K
PO
U2F
1 R
401H
IDH
1 R
132C
C11
orf2
4 A
395V
AK
T1
E17
K
TP
53 G
245S
BD
KR
B1
G11
5R
CAT
SP
ER
G D
966D
TP
53 R
282W
DN
AH
10 R
3649
W
PY
GL
S80
9L
GG
T5
G49
0R
ME
CO
M R
915Q
TP
53 R
175H
CN
OT
3 E
20K
CD
H23
S12
50S
TLR
7 P
147L
ITG
AL
R83
0C
HT
T A
805V
HE
Y2
C24
9C
PAR
VG
A96
T
AP
C E
1209
*
AP
C E
1322
*
AP
C C
1270
*
AP
C K
1165
*
MA
ML3
G10
69A
AP
C E
1353
*
AP
C Q
1367
*
AP
C Q
1378
*
AP
C Q
1406
*
AP
C E
1397
*
AP
C Q
1429
*
AP
C E
1309
*
AP
C S
1327
*
AP
C S
1356
*
AP
C R
1114
*
AP
C R
876*
AP
C E
1306
*
AP
C Q
1338
*
AP
C R
1450
*
AP
C R
302*
AP
C R
213*
NP
HS
1 D
1208
Y
PC
BP
1 L1
00Q
KR
AS
G12
V
AP
C R
216*
C8A
S42
1Y
SLC
5A8
E55
9*
HE
ATR
1 L1
256I
TOP
2A E
1035
*
CC
DC
18 N
CS
NV
AP
1M1
N15
5H
KR
AS
G12
A
AP
C R
283*
AP
C R
348*
AP
C R
564*
AR
ID1A
R19
89*
TP
53 I1
95T
GY
LTL1
B V
553G
PC
YT
1A F
263C
PC
BP
1 L1
02Q
KR
AS
G12
D
NA
A25
S60
6Y
ATP
10B
L76
1H
PD
CD
6IP
R32
7Q
RF
X6
R86
6Q
EYA
2 G
77A
KR
AS
G12
C
TR
PM
7 R
1862
C
AB
CB
1 R
286I
GO
LGB
1 V
1309
L
NR
AS
Q61
R
NR
AS
Q61
K
NR
AS
Q61
L
AP
RT
R87
G
NR
AS
G13
R
BR
AF
V60
0E
CS
DE
1 F
700V
IPM
K L
254V
NR
AS
G12
R
NR
AS
Q61
H
SN
X7
E36
4A
AN
KR
D32
L69
1R
NR
AS
G12
V
SD
CB
P T
268A
EP
RS
I820
L
DH
X32
R51
S
AN
O9
D47
3A
SY
TL2
NC
SN
V
AR
HG
EF
2 R
609G
ZN
F83
0 R
239S
FAM
120A
I302
K
TM
2D3
Y13
6S
KIA
A01
00 R
952L
PPA
T E
11D
SC
UB
E3
L427
V
CD
K4
R24
S
UC
KL1
D47
3H
CP
SF
1 Y
1164
H
TR
IM59
I353
M
FAM
151A
V31
D
US
PL1
A43
5G
PLE
KH
G4B
D33
0A
GP
RA
SP
1 D
145V
SLC
6A2
R21
6G
GC
G N
CS
NV
EIF
4G3
E18
9*
CU
BN
R22
4G
BR
AF
N58
1T
MB
NL3
NC
SN
V
TP
MT
V22
0A
STA
T5A
L35
5P
SLC
25A
46 L
221Q
NR
4A3
NC
SN
V
TLL
2 N
CS
NV
DD
X47
G12
8R
AB
CA
6 N
CS
NV
PIP
4K2A
NC
SN
V
TR
PV
2 L1
27V
CT
RB
2 N
CS
NV
MT
MR
6 F
75C
BR
AF
V60
0E
KIT
K64
2E
NR
AS
Q61
R
NR
AS
Q61
L
RP
S27
NC
SN
V
NR
AS
Q61
K
BR
AF
K60
1E
GN
AQ
Q20
9P
CD
YL2
D31
2E
P2R
Y11
NC
SN
V
BA
GE
2 N
CS
NV
TT
N N
CS
NV
C5
R12
06C
AB
CF
1 R
844*
SN
AP
91 D
723N
SIP
A1L
2 S
1472
L
TP
53 R
342*
CO
L5A
3 N
CS
NV
ZN
F99
NC
SN
V
DD
X27
NC
SN
V
TT
N N
CS
NV
TT
N N
CS
NV
PP
IAL4
A N
CS
NV
SLC
30A
8 L2
90P
TT
N H
1509
3H
KIA
A11
99 G
722E
AQ
P12
B T
16I
CY
P4F
12 P
367S
ZN
F76
8 A
488A
A2M
R14
7C
SA
RS
2 N
CS
NV
AR
NT
S32
1F
MD
N1
A23
8V
CO
L12A
1 R
1414
*
AM
D1
R21
0C
FO
LH1
NC
SN
V
AM
Y1C
K29
3K
RA
C1
P29
S
NS
FL1
C N
CS
NV
MA
GE
A11
S20
8L
DH
TK
D1
P84
2L
SE
RP
INA
1 N
CS
NV
ALP
PL2
G44
8R
GD
F2
F33
6F
RE
G1B
NC
SN
V
RA
SA
L2 P
146L
SY
DE
1 F
703F
NP
HP
1 R
347*
BU
B1
R67
7C
SLC
4A10
R41
0C
AR
ID5B
A10
09G
LHC
GR
NC
SN
V
MA
CC
1 N
CS
NV
RH
OA
Y42
S
VM
O1
NC
SN
V
GA
BR
E N
CS
NV
TT
N N
CS
NV
RN
F17
5 S
215G
PIK
3CA
H10
47R
TT
N N
CS
NV
PIK
3CA
N34
5K
ZC
3H4
E77
9Q
CD
H1
D25
4Y
ZB
TB
7A K
424T
DE
K S
288T
OLI
G2
S88
A
IQS
EC
2 G
283A
TP
53 C
176F
DR
G2
R38
L
KC
NH
2 G
149A
C8B
L13
8R
GN
G7
N45
K
ZD
HH
C8
P45
7A
KR
AS
G12
D
DO
CK
10 K
935M
TP
53 Y
220C
ITG
B1
L378
I
INS
IG2
I63F
CAV
1 N
CS
NV
CT
NN
B1
D32
H
AR
HG
AP
1 N
154I
ZM
YM
5 V
60D
TM
CC
1 N
CS
NV
ER
BB
2 D
769Y
C1o
rf9
NC
SN
V
PIK
3CA
E54
5K
ER
BB
2 N
CS
NV
C8o
rf42
W89
G
DE
FB
104A
E25
A
TP
53 P
151T
MLH
1 R
659*
ER
BB
2 S
310F
RP
L5 M
208T
ZH
X3
N24
9K
WT
1 N
CS
NV
NA
LCN
NC
SN
V
DK
K2
NC
SN
V
GA
RT
Q24
1R
STA
G1
A11
6T
TP
53 V
272M
BR
AF
V60
0E
NR
AS
Q61
R
ST
XB
P3
NC
SN
V
MR
PS
16 N
CS
NV
HR
AS
Q61
R
NR
AS
Q61
K
INT
S2
Y57
8N
SPA
CA
3 N
CS
NV
EIF
1AX
NC
SN
V
MA
RC
H4
NC
SN
V
GN
B2L
1 N
CS
NV
BR
AF
K60
1E
EIF
1AX
G9R
KR
AS
G12
V
KIA
A11
09 V
825F
PLC
L1 V
442I
ATP
6V0A
1 N
CS
NV
PM
PC
B N
CS
NV
EIF
1AX
G9D
HR
AS
Q61
K
MC
PH
1 K
645N
DN
AH
10 I1
660M
TB
C1D
12 N
CS
NV
OG
T P
120L
CC
2D2B
NC
SN
V
PP
P2R
1A A
273V
NU
P93
Q15
*
AK
T1
E17
K
DG
CR
8 E
518K
TB
C1D
12 N
CS
NV
TP
53 Q
192*
OR
52E
2 A
251A
PT
EN
R13
0G
SP
I1 N
CS
NV
NF
E2L
2 R
34G
FG
FR
2 S
252W
VN
1R5
K35
7M
AR
ID1A
R22
36P
PP
P2R
1A P
179R
XPA
NC
SN
V
C2C
D4A
NC
SN
V
CN
O N
CS
NV
MB
D2
NC
SN
V
ST
X5
NC
SN
V
AB
HD
14A
F19
1C
GC
SH
K16
9Q
SS
X2I
P N
CS
NV
RA
B6A
NC
SN
V
ME
TT
L14
R29
8P
P2R
Y13
NC
SN
V
TS
PAN
10 P
115A
SO
X17
S40
3I
ZB
TB
6 T
357I
CB
R3
NC
SN
V
RS
RC
1 S
239Y
FB
XW
7 R
505G
CN
O N
CS
NV
FO
LH1B
NC
SN
V
SO
S1
N23
3Y
DD
X1
NC
SN
V
CN
O N
CS
NV
B2M
NC
SN
V
MO
RC
3 R
896P
C16
orf9
1 N
CS
NV
PC
MT
D1
R16
2Q
PO
LE P
286R
AR
L6IP
6 N
CS
NV
PIK
3CA
H10
47L
LTV
1 R
377*
MU
C2
E26
12K
C2C
D4A
NC
SN
V
INS
M1
NC
SN
V
INS
M1
NC
SN
V
JUN
D N
CS
NV
TM
EM
37 E
165K
DLE
U7
NC
SN
V
ZN
F75
A R
134I
ZN
F75
A R
181I
SO
CS
2 R
96Q
AD
O N
CS
NV
GB
X2
NC
SN
V
AU
P1
NC
SN
V
FB
XW
7
HR
AS
TB
C1D
12
TP
53
FG
FR
3
SO
RB
S1
EX
OC
4
NF
E2L
2
TB
C1D
12
HR
AS
FG
FR
3
PIK
3CA
PIK
3CA
TP
53
KR
AS
TP
53
DS
T
FG
FR
3
MD
GA
2
HR
AS
FG
FR
3
FG
FR
3
TT
N
KR
AS
PIK
3CA
C10
orf7
6
EX
OC
4
TP
53
TP
53
LDH
AL6
B
TP
53
TP
53
PIK
3CA
TP
53
CN
TN
AP
3
MLL
3
TP
53
FR
MD
7
C1o
rf12
9
ER
BB
2
CD
H1
TP
53
TP
53
TP
53
PIK
3CA
ALD
OA
SU
PT
6H
PIK
3CA
TP
53
TP
53
AK
T1
FB
XW
7
ALD
OA
ME
D12
GA
BPA
FB
XW
7
CA
CN
G7
ST
6GA
L1
AK
T1
PIK
3CA
EP
300
PIK
3CA
NF
E2L
2
KR
AS
PIK
3CA
KR
AS
NX
F1
IQS
EC
2
MA
PK
1
BR
AF
TP
53
VN
1R5
KR
AS
KR
AS
KR
AS
AP
C
PIK
3CA
PC
BP
1
AP
C
PIK
3CA
KR
AS
KR
AS
AP
C
PIK
3CA
KR
AS
AB
CA
6
AP
C
NR
AS
AP
C
AP
C
AP
C
PIK
3CA
CN
TN
AP
3
EYA
2
TP
53
TP
53
PIK
3CA
SU
PT
6H
ST
6GA
L1
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TT
N
PIK
3CA
AB
CB
1
TP
53
DN
AH
10
TP
53
TP
53
C10
orf7
6
ER
BB
2
TP
53
TP
53
TP
53
AS
PH
AS
PH
BR
AF
TP
53
NR
AS
TP
53
EG
FR
PT
EN
EG
FR
TP
53
PIK
3R1
TP
53
PIK
3R1
EG
FR
PT
EN
EG
FR
TP
53
TP
53
PIK
3R1
MU
C2
FAM
120A
TP
53
EG
FR
TP
53
PIK
3CA
FB
XW
7
FG
FR
3
PIK
3CA
EP
300
PIK
3CA
PIK
3CA
MA
PK
1
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
FB
XW
7
HR
AS
TP
53
TP
53
PIK
3CA
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
HR
AS
TP
53
TP
53
TP
53
HR
AS
TP
53
TP
53
CD
KN
2A
MA
P2
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
RA
C1
ITG
B1
CD
KN
2A
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
TP
53
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
VH
L
PB
RM
1
PB
RM
1
KIT
KR
AS
FLT
3
GA
RT
IDH
1
IDH
2
CA
CN
G7
FLT
3
NR
AS
IDH
1
DN
MT
3A
DN
MT
3A
NR
AS
RU
NX
1
IDH
2
RU
NX
1
IDH
1
IDH
1 IDH
1
EG
FR
BR
AF
TP
53
IDH
1
TP
53
TP
53
TP
53
EG
FR
TP
53
TP
53
TP
53
IDH
2
PIK
3CA
IDH
2
EG
FR
PIK
3CA
PIK
3CA
TP
53
PIK
3CA
TP
53
TP
53
PIK
3CA
EG
FR
TP
53
TP
53
TP
53
TP
53
IDH
2
PIK
3R1
C10
orf7
6
IDH
2
TP
53
TP
53
TP
53
TP
53
NR
AS
PIK
3CA
TP
53
CT
NN
B1
CT
NN
B1
NX
F1
CT
NN
B1
CT
NN
B1
CT
NN
B1
LRR
C16
B
CT
NN
B1
CT
NN
B1
TP
53
CT
NN
B1
TP
53
TP
53
TP
53
CT
NN
B1
EG
FR
CT
NN
B1
CT
NN
B1
CT
NN
B1
BR
AF
KR
AS
CT
NN
B1
HE
ATR
1
KR
AS
BR
AF
KR
AS
KR
AS
EG
FR
EG
FR
CP
SF
1
BR
AF
PIK
3CA
PIK
3CA
FLR
T3
SO
S1
MU
C2
TP
53
TP
53
FLR
T3
PIK
3CA
TP
53
NF
E2L
2
NF
E2L
2
TP
53
TP
53
TP
53
TP
53
TP
53
CD
KN
2A
TT
N
TP
53
TP
53
MD
GA
2
TP
53
TP
53
TP
53
DS
T
CD
KN
2A
TP
53
IQS
EC
2
FB
XW
7
TP
53
TP
53
CD
KN
2A
NF
E2L
2
TP
53
TP
53
TP
53
TP
53
EP
300
TP
53
TP
53
TP
53
TP
53
TP
53
GA
BPA
FR
MD
7
MA
P2
TOP
2A
MA
ML3
TP
53
TP
53
TP
53
LDH
AL6
B
TP
53
TP
53
TP
53
LRR
C16
B
SO
RB
S1
NR
AS
TP
53
TP
53
TP
53
TP
53
C1o
rf12
9
TP
53
TP
53
KR
AS
KR
AS
KR
AS
KR
AS
TP
53
KR
AS
TP
53
TP
53
TP
53
TP
53
TP
53
CD
KN
2A
TP
53
TP
53
TP
53
SM
AD
4
SM
AD
4
TP
53
CD
KN
2A
SM
AD
4
TP
53
GN
AS
TP
53
TP
53
SM
AD
4
TP
53
GN
AS
TP
53
TP
53
CD
KN
2A
TP
53
TT
N
ER
BB
2
SP
OP
SP
OP
SP
OP
SP
OP
SP
OP
SP
OP
SP
OP
TP
53
SP
OP
TP
53
SP
OP
RH
OA
SP
OP
SP
OP
SP
OP
TP
53
BR
AF
SP
OP
ME
D12
HR
AS
CT
NN
B1
TP
53
ME
D12
MLL
3
PIK
3CA
TP
53
PIK
3CA
IDH
1
AK
T1
TP
53
TP
53
DN
AH
10
TP
53
AP
C
AP
C
AP
C
AP
C
MA
ML3
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
AP
C
PC
BP
1
KR
AS
AP
C
HE
ATR
1
TOP
2A
KR
AS
AP
C
AP
C
AP
C
AR
ID1A
TP
53
PC
BP
1
KR
AS
EYA
2
KR
AS
AB
CB
1
NR
AS
NR
AS
NR
AS
NR
AS
BR
AF
NR
AS
NR
AS
NR
AS
FAM
120A
CP
SF
1
BR
AF
AB
CA
6
BR
AF
KIT
NR
AS
NR
AS
NR
AS
BR
AF
TT
N
TP
53
TT
N
TT
N
TT
N
RA
C1
RH
OA
TT
N
PIK
3CA
TT
N
PIK
3CA
CD
H1
IQS
EC
2
TP
53
KR
AS
TP
53
ITG
B1
CT
NN
B1
ER
BB
2
PIK
3CA
ER
BB
2
TP
53
ER
BB
2
GA
RT
TP
53
BR
AF
NR
AS
HR
AS
NR
AS
EIF
1AX
BR
AF
EIF
1AX
KR
AS
EIF
1AX
HR
AS
DN
AH
10
TB
C1D
12
PP
P2R
1A
AK
T1
TB
C1D
12
TP
53
PT
EN
NF
E2L
2
VN
1R5
AR
ID1A
PP
P2R
1A
C2C
D4A
CN
O
FB
XW
7
CN
O
SO
S1
CN
O
PIK
3CA
MU
C2
C2C
D4A
INS
M1
INS
M1
ZN
F75
A
ZN
F75
A
nTCGA = 137
nTCGA = 143
nTCGA = 184
nTCGA = 178 nYG = 50
nTCGA = 336
nTCGA = 289 nYG = 56
nTCGA = 393
nTCGA = 399
nTCGA = 364 nYG = 41
nTCGA = 412
nTCGA = 65
nTCGA = 412
nTCGA = 436
nTCGA = 437
nTCGA = 495
nTCGA = 508
nTCGA = 492 nYG = 22
nTCGA = 363 nYG = 156
nTCGA = 104 nYG = 47
nTCGA = 530 nYG = 23
nTCGA = 492 nYG = 108
nTCGA = 567 nYG = 108
nTCGA = 986
READ
LAML
ESCA
PAAD
KIRC
CESC
GBM
COAD
LIHC
BLCA
HPV+ HNSC
HPV− HNSC
OV
STAD
PRAD
LGG
THCA
SKCMM
SKCMP
UCEC
LUSC
LUAD
BRCA
103104105106107
Selection intensity
Tum
or ty
pe
Q value
Q ≤ 0.1
Q > 0.1
Prevalence
0.60
0.40
0.20
0.10
0.05
0.01
Supplemental Figure 1
.CC-BY-NC 4.0 International licenseis made available under aThe copyright holder for this preprint (which was not peer-reviewed) is the author/funder. It. https://doi.org/10.1101/229724doi: bioRxiv preprint



![Comparison of 61 Sequenced Escherichia coli Genomes · Comparison of 61 Sequenced Escherichia coli Genomes ... O103:H2 [37] 15578 E. coli E110019 ... Comparison of 61 Sequenced Escherichia](https://img.pdfslide.us/doc/110x75/5af461b97f8b9a92718d78d2/comparison-of-61-sequenced-escherichia-coli-of-61-sequenced-escherichia-coli-genomes.jpg)