Embed Size (px)
Citation preview

686
Effect of Vitamin E on Pentane Exhaled by Rats Treated with Methyl Ethyl Ketone Peroxide
L.A. HERSCHBERGER and AL L. "I'APPEL*, Department of Food Science and Technology, University of California, Davis, CA 95616
ABSTRACT
One useful method to monitor in vivo lipid peroxidation is the measurement of volatile hydro- carbons, mainly pentane and ethane, that derive from unsaturated fatty acid hydroperoxides. Vitamin E, the biological antioxidant, inhibits lipid peroxidation and the production of pentane and ethane. The rates of pentane production by male Sprague-Dawley rats fed a diet that contained 10% vitamin E-stripped corn oil and 0, 1, 3, 5 or 10 IU dl-a-tocopherol acetate/kg were monitored over a 12-wk period. During the eleventh and twelfth weeks, the rats were injected intraperitoneally with 3.3 and 13 mg of methyl ethyl ketone peroxide (MEKP)/kg body wt, respectively. Pentane production was then measured at intervals over a 50-rain period, and the total amount of pentane produced over this time interval was estimated. An asymptotic function was found to describe the relationship between exhaled pentane and the low levels of dietary vitamin E that were fed to the rats. As measured by pentane production, rats had a higher minimal vitamin E requirement after they were treated with the potent peroxidation initiator MEKP than they did prior to treatment. The level of pentane exhaled by rats injected with 13 mg MEKP/kg body wt was significantly correlated with kidney and spleen tocopherol levels. Lipids 17:686-691, 1982.
Lipid peroxidation has been implicated as a cause of cellular damage (1) and has been asso- ciated with a variety of diseases (2). Vitamin E, an in vivo biological antioxidant, protects mem- branes against lipid peroxidation (3). Vitamin E-deficient experimental animals exhibit diverse symptoms, some of which are species-specific (4). Deficiencies of most vitamins result in characteristic pathological symptoms, but vita- min E deficiency symptoms are more difficult to recognize and to quantify (5). Vitamin E deficiency symptoms have been used to deter- mine minimal vitamin E requirements (6-10) and relative potencies of tocopherol isomers and esters (11-14).
One of the newest methods to detect lipid peroxidation in vivo directly is the measure- ment of exhaled volatile hydrocarbons, particu- larly pentane and ethane. Pentane and ethane are minor products formed during decomposi- tion of w6- and 603-unsaturated fatty acid hydroperoxides, respectively (15). A variety of oxidants initiate lipid peroxidation in vivo. Riely et at. (16) first demonstrated that the level of ethane increased in breath from mice injected with carbon tetrachloride. Ethane and/or pentane were elevated in expired breath of animals treated with halogenated hydro- carbons (17), ethanol (18), ozone (19), iron (20) or methyl ethyl ketone peroxide (MEKP)
*Author to whom correspondence should be addressed.
(2 I), and vitamin E inhibited the ensuing lipid peroxidation.
MEKP is used industrially to initiate poly- merization reactions in the manufacture of plastics and fabrics. Unlike most other organic peroxides, MEKP decomposes to free radicals in the presence of metal ions at room tempera- ture. Litov et at. (21) reported that injected
MEKP caused severe lipid peroxidation as deter- mined by measurement of the immediate high- level production of pentane by vitamin E- deficient rats. The measurement of expired volatile hydrocarbons to index in vivo lipid peroxidation has gained acceptance in recent years.
The possibility that the technique could be used in bioassay of antioxidant status was sug- gested (22). Dietary vitamin E has its major effect on basal pentane production by rats in the range of 0-10 IU dl-a-tocopherol acetate/kg diet (19,23). However, at low levels of dietary vitamin E, the relationships between expired pentane and dietary vitamin E levels and between expired pentane and tissue vitamin E levels have not been adequately investigated.
The quantitative relationships between low levels of dietary vitamin E and both basal pen- tane production and MEKP-induced pentane production are examined in this study. Kidney and spleen vitamin E levels were measured to correlate pentane production with tissue vitamin E status. The possible use of the tech- nique in bioassay of antioxidant status is discussed.
LIPIDS, VOL. 17, NO. 10 (1982)

VITAMIN E PROTECTION AGAINST A PEROXIDE 687
M ETH ODS
Animals and Diets
Nineteen weanling male Sprague-Dawley rats were divided into five groups and were fed a basal 10% tocopherol-stripped corn oil diet (24) with mineral mix 4179 (Teklad Test Diets) for 12 wk and 0, 1, 3, 5 or 10 IU dl-a-tocopherol acetate/kg for 12 wk. All of the groups con- tained four rats except there were only three rats in the group fed 1 IU vitamin E/kg diet. The rats were housed singly or in pairs in hang- ing wire cages and were kept on a 14-hr light and 10-hr dark cycle at 22-24 C. The diets and water were provided ad libitum.
Pentane Measurements and Oxidant Treatments
Sample collection and measurement of pentane were done as described by Dillard et al. (22). The rats were fasted 18-24 hr before their rates of pentane production were determined. Measurements of exhaled pentane were made during weeks 3, 7, 10, 11 and 12. After a 10- min exposure to hydrocarbon-free air, a 500-ml air-breath sample was collected and analyzed by flame-ionization gas chromatography on a 10-ft alumina column at 160 C. The nitrogen carrier gas flow rate was 25 ml/min. The injec- tor and detector temperatures were 210 and 300 C, respectively. The amount of pentane was calculated from the peak height x width of the peak at half-height. The instrument was calibrated daily with a 0.89-ppm pentane standard. The basal rates of pentane produc- tion are expressed as pmol pentane/100 g body wt/min.
Basal pentane production by the rats was measured prior to each treatment with MEKP. Each rat was injected intraperitoneally (ip) with 3.3 mg MEKP/kg body wt during week 11 and 13 mg MEKP/kg body wt during week 12. The solution injected was 6.6 mg MEKP/ml tri- butyrin. Tributyrin was used to maximize the absorption of MEKP. The lower level of MEKP was chosen to ensure that the rats would sur- five. The higher MEKP dose was used to obtain highly significant differences among the rates of pentane production by the rats in the different dietary groups. Pentane was measured in samples of breath 10, 20, 35 and 50 min fol- lowing injections. The basai measurement of pentane was used as a zero time point. After MEKP was injected into a rat, the total number of pmol of pentane exhaled above the basal level of pentane was estimated by integrating the rate of pentane production over 50 rain and subtracting from this the number of pmol that would have been produced during the 50 min under basal conditions. The latter was esti-
mated by multiplying the basal rate of pentane production (pmol/100 g body wt/min) by 50 min. The integrated amount of pentane exhaled following the oxidant treatment is expressed as pmol pentane/100 g body wt/50 min.
Tissue Tocopherol Analysis
The rats were anesthetized with penta- barbital ca. 75 rain after the second injection of MEKP was given. The blood was removed by heart puncture to remove as much blood from the tissues as possible. The kidney and spleen were removed, rinsed in 0.9% KC1 and frozen at -10 C under nitrogen. The kidney and spleen were analyzed for total tocopherol by fluores- cence assay (25).
Statistical Analysis
The curves obtained by plotting pentane production vs dietary vitamin E were modeled to the general equation Y = a + br x (26) using the Biomedical Computer Program BMD06R. This equation describes a decrease in the pen- tane response, y, that reaches an asymptote at high levels of x, the dietary vitamin E. In this
equa t ion , a is the minimal pentane level obtain- able, b is the difference in the rate of pentane production between rats fed no vitamin E and those fed higher amounts of vitamin E and r is a dimensionless number that describes how quickly the asymptote is approached. To deter- mine if the data fit the asymptotic model, a comparison was made between the residual sum of squares, calculated using the asymptotic model (SSEMoDEL), and the minimal residual sum of squares possible over all models (SSEMEANS). The SSEMEAN S is the sum of the squares of the deviation of each observation from its group mean. The ratio (SSEMoDE L - SSEMEANS)/SSEMODE L was calculated for each fitted line. A Burroughs 7800 computer was used for the above computations. All p values < 0.05 were considered to be significant.
RESULTS
Basal Pentane Measurements
The amounts of pentane produced by rats fed varying vitamin E levels for 3, 7, 11 and 12 wk are shown in Figure 1. The data obtained during the tenth week were similar to the eleventh week data, but they were excluded from the figure for clarity. Over the concentra- tion range of dl-a-tocopherol acetate used in the diets, pentane production was inversely related to vitamin E intake. During the third week, the levels of exhaled pentane were higher than at other time points. The 7-wk curve and
LIPIDS, VOL. 17, NO. 10 (1982)
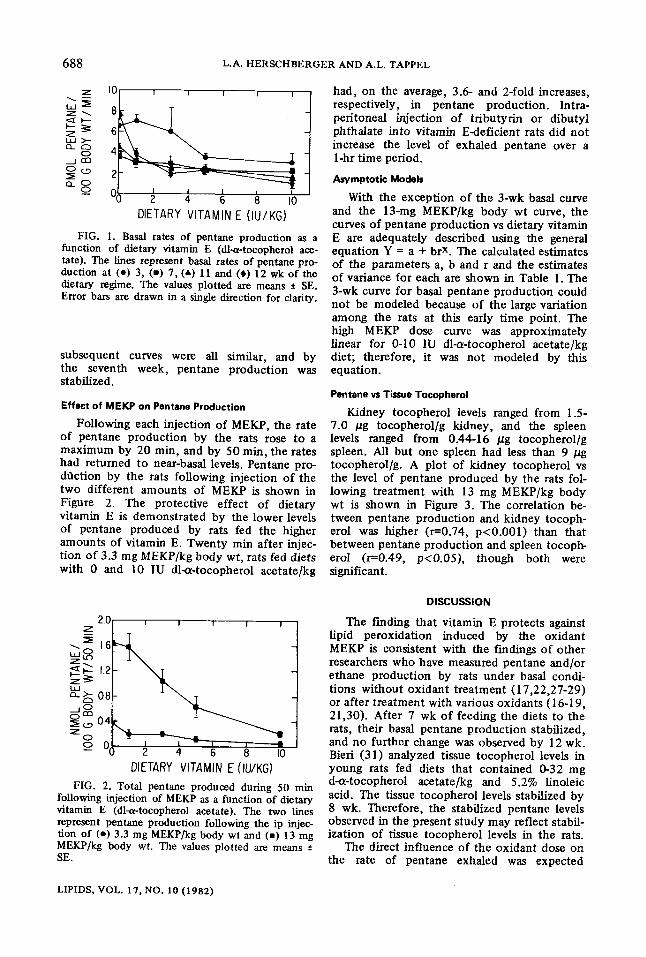
688 L.A. HERSCHBERGER AND A.L. TAPPEL
z I0 I I I I I
,z ~ 6
~ 2
2 4 8 ,o DIETARY VITAMIN E (IU/KG)
FIG. 1. Basal rates of pentane production as a function of dietary vitamin E (dl-a-tocopherol ace- late). The lines represent basal rates of pentane pro- duction at (e) 3, (-) 7, (A) 11 and (#) 12 wk of the dietary regime. The values plotted are means • SE. Error bars are drawn in a single direction for clarity.
subsequent curves were all similar, and by the seventh week, pentane production was stabilized.
Effect of MEKP on Pentene Production
Fol lowing each injection o f MEKI ), the rate of pentane product ion by the rats rose to a maximum by 20 min, and by 50 min, the rates had returned to near-basal levels. Pentane pro- dt~ction by the rats following injection of the two different amounts of MEK.P is shown in Figure 2. The protective effect of dietary vitamin E is demonstrated by the lower levels of pentane produced by rats fed the higher amounts of vitamin E. Twenty min after injec- tion of 3.3 mg MEKP/kg body wt, rats fed diets with 0 and 10 IU dlax-tocopherol acetate/kg
had, on the average, 3.6- and 2-fold increases, respectively, in pentane production. Intra- peritoneal injection of tributyrin or dibutyl phthalate into vitamin E-deficient rats did not increase the level of exhaled pentane over a 1-hr time period.
Asymptotic Models
With the exception of the 3-wk basal curve and the 13-mg MEKP/kg body wt curve, the curves of pentane production vs dietary vitamin E are adequately described using the general equation Y = a + br x. The calculated estimates of the parameters a, b and r and the estimates of variance for each are shown in Table 1. The 3-wk curve for basal pentane production could not be modeled because of the large variation among the rats at this early time point. The high MEKP dose curve was approximately linear for 0-10 IU dl-~-tocopherol acetate/kg diet; therefore, it was not modeled by this equation.
Pentane vs Tissue Tocopherol
Kidney tocopherol levels ranged from 1.5- 7.0 /~g tocopherol/g kidney, and the spleen levels ranged from 0.44-16 /zg tocopherol/g spleen. All but one spleen had less than 9 /ag tocopherol/g. A plot of kidney tocopherol vs the level of pentane produced by the rats fol- lowing treatment with 13 mg MEKP/kg body wt is shown in Figure 3. The correlation be- tween pentane production and kidney tocoph- erol was higher (r=0.74, p<0.001) than that between pentane production and spleen tocoph- erol (1"=0.49, p<0.05), though both were significant.
20 I I ! I I g
"~ 1.6
~ - ~ 08 ~ o
2 4 - V-- o DIETARY VITAMIN E (IU/KG)
FIG. 2. Total pentane produced during 50 min following injection of MEKP as a function of dietary vitamin E (dl-~-tocopherol acetate). The two lines represent pentane production following the ip injec- tion of (e) 3.3 mg MEKP/kg body wt and (-) 13 mg MEKP/kg body wt. The values plotted are means • SE.
DISCUSSION
The finding that vitamin E protects against lipid peroxidation induced by the oxidant MEKP is consistent with the f'mdings of other researchers who have measured pentane and/or ethane production by rats under basal condi- tions without oxidant treatment (17,22,27-29) or after treatment with various oxidants (16-19, 21,30). After 7 wk of feeding the diets to the rats, their basal pentane production stabilized, and no further change was observed by 12 wk. Bieri (31) analyzed tissue tocopherol levels in young rats fed diets that contained 0-32 mg d-a-tocopherol acetate/kg and 5.2% linoleic acid. The tissue tocopherol levels stabilized by 8 wk. Therefore, the stabilized pentane levels observed in the present study may reflect stabil- ization of tissue tocopherol levels in the rats.
The direct influence of the oxidant dose on the rate of pentane exhaled was expected
LIPIDS, VOL. 17, NO. 10 (1982)
VITAMIN E PROTECTION AGAINST A PEROXIDE
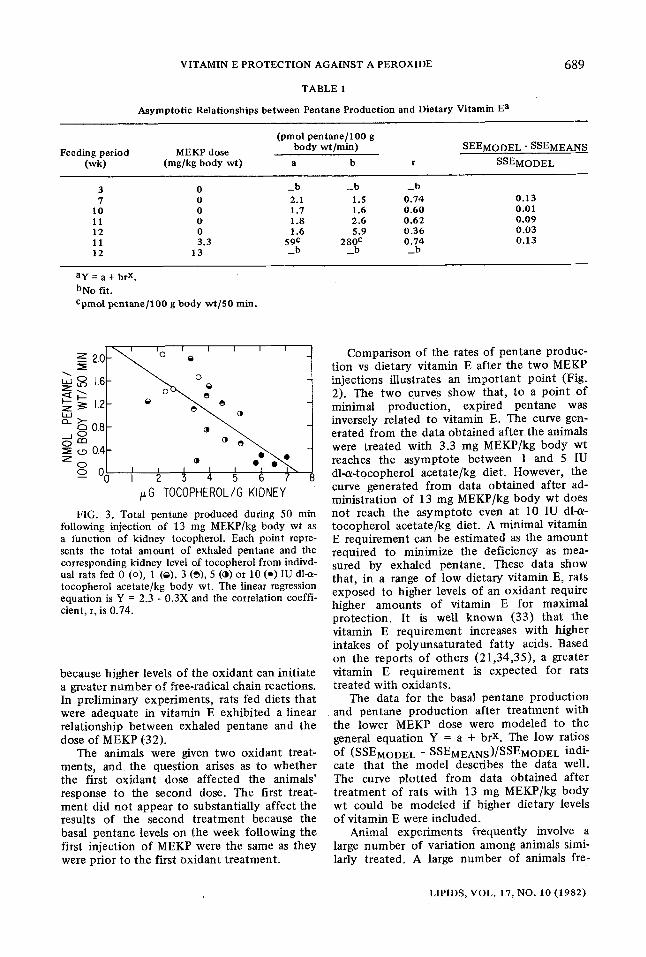
TABLE 1
Asymptotic Relat ionships be tween Pentane Production and Dietary Vitamin E a
689
Feeding period (wk)
(pmol p e n t a n e / 1 0 0 g MEKP dose body wt/min) SEEMODEL - SSEMEANS
(mg/kg body wt) a b r SSEMoDEL
3 7
I0 l l 12 11 12
0 _ b _ b _ b 0 2.1 1.5 0 .74 0 .13 0 1.7 1.6 0 .60 0 .01 0 1.8 2 .6 0 .62 0 .09 0 1.6 5.9 0 .36 0 .03 3.3 59 c 280 c 0 .74 0 .13
13 _ b _ b _ b
ay = a + br x. bNo fit. epmol pentane]100 g body wt]50 min.
o 1.6 oo-,,. ~
z ~ : 1.2 I..iJ a-~O. 8 o _ o ~ , ~ ~' , " ~ , " ~ 0 . 4
Z o O 0 I I I I I I I% - - ~) I 2 3 4 5 6 7 8
/zG TOCOPHEROL/G KIDNEY
FIG. 3. Total pentane produced during 50 min following injection of 13 mg MEKP/kg body wt as a function of kidney tocopherol. Each point repre- sents the total amount of exhaled pentane and the corresponding kidney level of tocopherol from indivd- ual rats fed 0 (o), 1 (~), 3 (~), 5 (O) or 10 (-) IU dl-~- tocopherol acetate/kg body wt. The linear regression equation is Y = 2.3 - 0.3X and the correlation coeffi- cient, r, is 0.74.
because higher levels of the oxidant can initiate a greater number of free-radical chain reactions. In preliminary experiments, rats fed diets that were adequate in vitamin E exhibited a linear relationship between exhaled pentane and the dose of MEKP (32).
The animals were given two oxidant treat- ments, and, the question arises as to whether the first oxidant dose affected the animals' response to the second dose. The first treat- ment did not appear to substantially affect the results of the second treatment because the basal pentane levels on the week following the first injection of MEK_P were the same as they were prior to the first oxidant treatment.
Comparison of the rates of pentane produc- tion vs dietary vitamin E after the two MEKP injections illustrates an important point (Fig. 2). The two curves show that, to a point of minimal production, expired pentane was inversely related to vitamin E. The curve gen- erated from the data obtained after the animals were treated with 3.3 mg MEKP/kg body wt reaches the asymptote between 1 and 5 IU dl-ct-t0copherol acetate/kg diet. However, the curve generated from data obtained after ad- ministration of 13 mg MEKP/kg body wt does not reach the asymptote even at 10 IU dl-~- tocopherol acetate/kg diet. A minimal vitamin E requirement can be estimated as the amount required to minimize the deficiency as mea- sured by exhaled pentane. These data show that, in a range of low dietary vitamin E, rats exposed to higher levels of an oxidant require higher amounts of vitamin E for maximal protection. It is well known (33) that the vitamin E requirement increases with higher intakes of polyunsaturated fatty acids. Based on the reports of others (21,34,35), a greater vitamin E requirement is expected for rats treated with oxidants.
The data for the basal pentane production and pentane production after treatment with the lower MEKP dose were modeled to the general equation Y = a + br x. The low ratios of (SSEMoDE L - SSEMEANS)/SSEMODEL indi- cate that the model describes the data well. The curve plotted from data obtained after treatment of rats with 13 mg MEKP/kg body wt could be modeled if higher dietary levels of vitamin E were included.
Animal experiments frequently involve a large number of variation among animals simi- larly treated. A large number of animals fre-
LIPIDS, VOL. 17, NO. 10 (1982)

690 L.A. HERSCHBERGER AND A.L. TAPPEL
quently must be used to detect significant differences among groups treated differently. The experimental design of the present study circumvents the problem in two ways. First, the rates of pentane production were mon- itored in the same animals throughout the feed- ing program and also following their treatment with the oxidant MEKP. Second, a potent initiator of lipid peroxidation was administered to accentuate the differences in vitamin E status among the groups fed various levels of vitamin E. High levels of a toxicant are fre- quently used to minimize the number of ani- mals required to detect significant differences.
The bioassay described in this study uses the response of in vivo pentane production to determin0 the antioxidant effectiveness in the whole animal. When 13 mg MEKP/kg body wt was injected into the rats, there were highly significant differences in pentane production among rats fed low levels of vitamin E. The incorporation of an oxidant in antioxidant bioassays is often used in tests of erythrocyte hemolysis. There are some distinct advantages to using exhaled pentane to assess vitamin E status in animals. One advantage is that t h e exhaled pentane derives from hydroperoxides, which are the initial products of peroxidation. A second advantage is that the amount of pen- tane exhaled by an animal is a reflection of its in vivo antioxidant status. The level of pentane produced is a function of the absorption, dis- tribution and excretion of the vitamin E and te specific isomers ingested. The pentane level thus reflects the in situ antioxidant concentra- tions and effectiveness. A third advantage of the pentane methodology is that it is theoret- ically applicable to most animal species (16,22, 36-38). A fourth advantage of this in vivo bio- assay is that incorporation of MEKP could be used to test the effectiveness of many different antioxidants.
The pentane method also has some disad- vantages that may limit its use in a bioassay of vitamin E that does not include an oxidant. It was observed in some studies (21) that differ- ences in pentane production between rats fed high and low vitamin E occurred only after an oxidant was used to initiate peroxidation. Another possible disadvantage is that a lower minimal vitamin E requirement was estimated by measurement of exhaled pentane than was estimated in studies using erythrocyte hemol- ysis as an index of vitamin E deficiency. Under basal conditions, this requirement was esti- mated to be between 1 and 5 IU dl-~-tocoph- erol acetate/kg diet. The National Research Council vitamin E requirement for rats (39) was based on 'a study by Jager and Houtsmiiller
(10). In their study, erythrocyte hemolysis was used to index vitamin E deficiency in rats fed diets containing 3.5% linoleic acid. The minimal vitamin E requirement was estimated to be 13 mg d-0t-tocopherol acetate/kg diet (39). This is equivalent to 17.6 mg dl-a-tocopherol acetate/ kg diet (39). When the higher dose of the MEKP was included, the minimal vitamin E requirement was greater than 10 IU dl-a- tocopherol acetate, which is closer to that estimated by Jager and Houtsmiiller (10). Other incongruities among bioassay methods used to determine vitamin E requirements can be found in the literature (11 ).
Litov et al. (21) found a high correlation between pentane exhaled by rats injected with MEKP and the ratio of mg tocopherol/g lipid found in plasma. In the present study, analysis of tissue for tocopherol was done to determine whether tissue vitamin E levels were more highly correlated with pentane levels than were the dietary vitamin E levels. If this were true, some of the variation in the pentane levels could be explained by differences in vitamin E absorption, distribution or excretion. I~e cor- relations of pentane exhaled by rats injected with 13 mg MEKP/kg body wt with tocopherol levels in the kidney and spleen were low, but both were significant (p<O.05). The correlation coefficient between the exhaled pentane and log(dietary vitamin E + 1) was much higher. Liver may be the most important organ related to lipid peroxidation. Over 95% of conjugated dienes in organs of rats treated with CC14 was found in the liver (40). Another possible ex- planation for the low correlation of exhaled pentane with the tissue tocopherol levels may be that the MEKP oxidized the low levels of tocopherol to tocopherol quinone, which is n o t d e t e c t e d by the fluorescence assay.
The measurement of in vivo pentane produc- tion as an index of lipid peroxidation was shown in this study to be useful for studying the effects of low levels of vitamin E in rats. Under conditions of high oxidant stress, the minimal vitamin E requirement was greater than that estimated for rats under basal conditions.
ACKNOWLEDGMENT
This research was supported by funds from Best F o o d s - a unit o f CPC North America, and by research grant AM 09933 from the National Institute of Arthritis, Diabetes, Digestive and Kidney Diseases.
REFERENCES
1. Bus, J.S., and Gibson, J.E. (1979) in Reviews in Biochemical Toxicology (Hodgson, E., Bend, J.R., and Philpot, R.M., eds.), vol. I, pp. 125-149,
LIPIDS, VOL. 17, NO. 10 (1982)

VITAMIN E PROTECTION AGAINST A PEROXIDE 691
Elsevier North Holland, Inc., New York, NY. 2. DelMaestro, R.F. (1980) Acta Physiol. Seand.
Suppl. 492,153-168. 3. Molennaar, I., Hulstaert, C.E., and Hardonk,
M.J. (1980) in Vitamin E: A Comprehensive Treatise (Machlin, L.J., ed.), pp. 372-389, Marcel Dekker, Inc., New York, NY.
4. Desai, I.D. (1980) in Vitamin E: A Comprehen- sive Treatise (Machlin, L.J., ed.), pp. 67-98, Marcel Dekker, Inc., New York, NY.
5. Horwitt, M.K. (1980) in Vitamin E: A Compre- hensive Treatise (Maehlin, L.J., ed.), pp. 621-636, Marcel Dekker, Inc., New York, NY.
6. Ames, S.R. (1974) Am. J. Clin. Nutr. 27, 1017- 1025.
7. Gabriel, E., Machlin, L.J., Filipski, R., and Nelson, ]. (1980) J. Nutr. 110, 1372-1379.
8. Bieri, J.G., and Poukka, R.K.H. (1970) J. Nutr. 100, 557-564.
9. Jager, F.C. (1972) Nutr. Metabol. 14, 1-7. 10. Jager, F.C., and Houtsmiiller, V.M.T. (1970)
Nutr. Metabol. 12, 3-12. 11. Bieri, J.G., and Evarts, R.P. (1974) J. Nutr. 104,
850-857. 12. Ames, S.R. (1971) Lipids 6,281-290. 13. Ames, S.R. (1979) J. Nutr. 109, 2198-2204. 14. Scott, M.L., and Desai, I.D. (1964) J. Nutr. 83,
39-43. 15. Dumelin, E.E., and Tappel, A.L. (1977) Lipids
11,894-900. 16. Riely, C.A., Cohen, G., and Lieberman, M.
(1974) Science 183,208-210. 17. Hafeman, D.G., and Hoekstra, W.G. (1977) J.
Nutr. 107,656-665. 18. Litov, R.E., Irving, D.H., Downey, J.E., and
Tappel, A.L. (1978) Lipids 13, 305-307. 19. Dumelin, E.E., Dillard, C.J., and Tappel, A.L.
(1978) Arch. Environ. Health 33, 129-134. 20. Dougherty, J.J., Croft, W.A., and Hoekstra,
W.G. (1981) J: Nutr. 111, 1784-1786. 21. Litov, R.E., Matthews, L.C., and Tappel, A.L.
(1981) Toxicol. Appl. Pharmacol. 59, 96-106.
22. Dillard, C.J., Dumelin, E.E., and Tappel, A.L. (1977) Lipids 12, 109-114.
23. Litov, R.E., Gee, D.L., Downey, J.E., and Tappel, A.L. (1981) Lipids 16, 52-57.
24. Draper, H.H., Bergan, J.G., Chiu, M., and Csal- lany, A.S. (1964) J. Nutr. 84, 395-400,
25. Taylor, S.L., Lamden, M.P., and Tappel, A.L. (1976) Lipids 11,530-538.
26. Stevens, W.L. (1951) Biometrics 7, 247-267. 27. Dillard, C.J., Litov, R.E., and Tappel, A.L.
(1978) Lipids 13,396-402. 28. Hafeman, D.G., and Hoekstra, W.G. (1977) J.
Nutr. 107, 666-672. 29. Downey, J.E., Irving, D.H., and Tappel, A.L.
(1978) Lipids 13,403-407. 30. Burk, R.F., and Lane, J.M. (1979) Toxicol.
Appl. Pharmacol. 50,467-478. 31. Bieri, J.G. (1972) Ann. N.Y. Acad. Sei. 203,
181-191. 32. Hersehberger, L.A. (1982) M.S. thesis, University
of California, Davis. 33. Harris, P.L., and Embree, N.D. (1963) Am. J.
Clin. Nutr. 13, 385-392. 34. Chen, L.H. (1981) Am. J. Clin. Nutr. 34, 1036-
1041. 35. Chow, C.K., and Tappel, A.L. (1972) Lipids 7,
518-524. 36. Dillard, C.J., Litov, R.E., Savin, W.M., Dumalin,
E.E., and Tappel, A.L. (1978) J. Appl. Physiol. Respirat. Environ. Exercise Physiol. 45,927-934.
37. Young, R.G., and Tappel, A.L. (1978) Exp. Gerontol. 13, 457-459.
38. Dumelin, E.E., Diilard, C.J., and Tappel, A.L. (1978) Environ. Res. 15, 38-43.
39. National Research Couneil/NAS, Nutrient Requirements of Laboratory Animals, No. 10, National Academy of Sciences, Washington, DC, 1978, pp. 21-22.
40. Sagai, M., and Tappel, A.L. (1979) Toxicol. Appl. Pharmacol. 49, 283-291.
[Received March 8, 1982]
LIPIDS, VOL. 17, NO. 10 (1982)





























