Embed Size (px)
Citation preview

Plant Physiol. (1969) 44, 1684-1689
Effect of Sublethal and Lethal Temperatures on Plant Cells'J. W. Daniell2, W. E. Chappell, and H. B. Couch
Department of Plant Pathology and Physiology, Virginia Polytechnic Institute,Blacksburg, Virginia 24061
Received May 28, 1969.
A bstract. Soybean, Glycine max L., and elodea, Elodea canadensis Michx, leaves wereexposed to sublethal and lethal temperatures and examined by light microscopy. Loss ofohlorophyll and swollen chloroplasts were observed in cells of elodea leaves exposed to sub-lethal temperatures. At the thermal death point of leaf cells of elodea and soybean, therewas a disorganization of the tonoplast membrane, plasmalemma, and chloroplast membranes.Approximately 40% of the cells in elodea and 50 % of the cells in soybean leaves exhibitedoriteria of cell death when exposed to a temperature which induced necrotic leaf tissue.Plasmolysis of leaf cells of elodea and soybean occurred at lethal temperatures, but did notappear to be the primary oause of cellular death. The primary effect of lethal temperatureson the leaf cells used in these experiments is disintegration of -the cellular membranes.
Following exposure of attached elodea leaves to lethal temperatures, changes in leaf cellswere periodioally observed with a light microsoope. In low temperature treatments, (43through 520), the percentages of cells exhibiting criteria of death 12 days after treatment didnot change from the percentages dertermined immediately after treatment. All treatmentsabove 520 resulted in 400% or more of the cells exhibiting criteria of cell death immediatelyafter treatment. In these treatments, this resulted in all cells exhibiting criteria of death onthe fourth day after treatment.
Sun-scald of fruits (6), whitespot lesions onstemis (5), and other thermal incited injuries havebeen, reported when plants or fruits have been sub-jected to superoptimal temperatures. Accumulativeheat injury eventually results in total inhibition ofthe vital functions of plants or 'individual cells. Ininitial stages of heat injury,, the effect is reversiblebut finally leads to "thermal death". Nielsen andTodd (13) found that the permeability of cell mem-branes of potato tubers was increased by heating tosulblethal temperatures of 43 to 45°. To determinethe injury to cells from exposure to high tempera-tures, Alexandrov (1) used such criteria as suppres-sion of plasmnolysis and deplasmolysis, exit of pig-ments from the vacuole, loss of vital staining,luminescence of the chloroplasts, and the cessationof protoplasmic streaming. Vital stains have beenused by several workers (11, 15) to determine thedeath of cells.
The cause of death by high temperature usuallyhas been explained as the result of denaturation of
1 Contribution No. 127, Department of Plant Path-ology and Physiology, Virginia Polytechnic Institute,Blacksburg, Virginia. This work was supported bygrants from the Liquefied Petroleum Industry of Vir-ginia. Part of a Ph.D. thesis submitted by J. W.Daniell.
2 Present address: Department of Horticulture, Geor-gia Station, Experiment, Georgia 30212.
proteins (9). Other theories have been reviewedby Levitt (10). Heat denatured proteins may ag-gregate or coagulate, with the coagulation being asecondary phenomenon (14). In 1938, Maximov(12) stated that the action of superoptimal tempera-ture, as explained 'by the coagulation of proteintheory, often is niot sufficient since death may beginat 400, a level which is far below the coagulationpoint of proteins. There appear to be proteins orenzymes which are relatively stable at high tempera-tures. Koffler (8) suggests that the relative heatstability of proteins or enzymes in thermophilicorganisms depends on more effective hydrogen bond-ing. This effectiveness of the bonding may dependon the number, strength, or location of the hydrogenbonds as well as other types of bonds.
The purpose of this investigation was to deter-mine some cellular responses to sublethal and lethaltemperatures and to obtain information on the effectsof superoptimal temperatures at the thermal deathpoint.
Materials and Methods
Soybean plants [Glycine nax (L.) Merr. var.Ogdan] were grown from seed in pots in a green-house until they were 42 days old. Elodea plants(Elodea canadensis Michx) were obtained from afresh water stream when needed.
Soybean. Temperature treatments were appliedto soybean leaves by exposing the plant tissue to a10420 flame for 130 msec. In preliminary tests, it
1684 www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from
Copyright © 1969 American Society of Plant Biologists. All rights reserved.

DANIELL, ET AL.-HIGH HEAT EFFECTS ON PLANT CELLS.
had been determined that leaf tissue exposed to thisflame for 130 msee was killed quickly and verypronounced changes occurred within the cells. Anassembly consisting of 2 Afco gas burners (onemounted on each of the 2 long sides of the assembly)18 inches apart was employed. The burners, sup-plied with liquefied petroleum at a constant pressureof 22 psi, developed a 10420 flame which was 7 inchesacross and 4 inches wide at a distance of 9 inchesfrom the tip. The belt on the assembly was adjustedto travel at a speed of 4.4 feet per sec. Thus, anobject placed on the belt- would move past the burnersand be exposed to the flame for 130 msec. Thesoybean plants were placed on the assembly and anattached leaf was held with a utility clamp on thepetiole so that the upper epidermis of the leaf wasperpendicular to the burner.
Sublethal and lethal temperature treatments wereapplied to soylbean, leaves by placing detached matureleaves in a water bath which was maintained at aconstant temperature within +0.1°. The tempera-ture treatments were from 47 to 570 in 10 increments.Control leaves were immersed in water at 240. De-tached leaves were placed iin cheesecloth and im-mersed in the hot water and stirred constantly. Atthe end of 1 min, the leaves were removed and cooledimmediately by placing them in water at approxi-mately 250. Immediately after treating. 2-cm-squaresections of each leaf were cut and fixed in formalin-acetic acid-alcohol solution. Prepared slides weremade of these sections as outlined by Jensen (7)using the safranin-fast green staining schedule.Cross and longitudinal sections were cut at 9 micronsthickness.
Elodea. Elodea was selected for this study be-cause the cytoplasm and cellular organelles in livingleaf cells could be observed under a light microscopewithout damage to the cells. Temperature treat-ments were applied to elodea leaves by using hotwater treatmen'ts similar to those used for soybeanleaves. Some leaves were mounted in water onslides for immediate study, and others were stainedfor 30 min before mounting. The dyes used wereneutral red, methyl green, toluidine blue 0, methyleneblue, basic fuchsia, and rhodamine B. All of thedyes were used at a concentration of 0.0005 % (w/v)except neultral red, which was used at 0.002 % (w/v).Two of the dyes, neutral red and methylene blue,have been classified as "vital stains". In a livingcell these stains are accumulated in vacuoles, butafter death the cytoplasm and the nucleus stain in-tensely and diffusely. The response of cells to these2 dyes was used as the criterion of cell death. Inthis study, the other dyes stained the cells apparentlyonly after the differential permeability of the cellmembranes had been altered by the high temperatureand were, therefore, used as indicators of changesin cell permeability. The cells in the elodea leafwere observed microscopically to determine theeffect of high temperature on suppression of proto-
plasmic streaming; changes in the cytoplasm, nucleiand chloroplasts; exudation of pigments from thechloroplasts; plasmolysis and deplasmolysis; andsuppressgion of Browniian movement. Photomicro-graphs were taken with a Leica camera and a Zeissmicroscope. The percentage of damaged cells wascalculated from counts of 10 replications of thenumber of damaged and undamaged cells in treatedtissue within the field of a microscope with magnifi-cation of 15OX.
An additional test was conducte-d on attachedelodea leaves using the materials and methods as inthe previous test, except that mounts were made of theintact treated leaves at specific days after treatments.After treatment, the attached leaves were placed inbeakers containing 500 ml of water and kept in agrowth chamber with a 12 hr dark period tempera-ture of 120 and a light period temperature of 170.\Vhole mounts were made using 6 leaves from eachof the treatments on the first, second, fourth, seventh,and twelfth day after the hot water treatments, andchanges in the cells were noted.
Results
Soybean. Leaf studies in this experiment weredirected toward determining the visible effects ofexcessive heat on cells within leaves. The visibleeffects of the flame on the palisade parenchyma cellsof a soybean leaf are shown in Fig. lb. The chloro-plasts, nucleus, and tonoplast appeared to rupture,and leakage of the contents of the cytoplasm into thevacuole occurred.
A second test was designed to determine theminimum temperature at which the above responsescould be observed when the leaves were treated inhot water. At 560 and 570, 90 % or more of thecells exhibited disintegration of the chloroplasts anddisruption of the vacuole. However, all cells didnot exhibit the same degree of injury at a particulartemperature. At 550, approximately 50 % of thepalisade cells exhibited the damage described above,and at 540 approximately 40 % of the cells weredamaged. Little or no damage was observed withtreatments at lower temperatures. Separate tests inwhich one-half of attached soybean leaves receivedthe hot-water-bath treatment, showed that treatmentat 550 or above for 1 min resulted in necrosis of alltreated leaf tissue. A sharp line of demarcation wasapparent between the necrotic tissue and undamaged,non-treated tissue. Also, treatment at 540 resultedin chlorosis of treated tissue whtile treatment at 530and below, resulted in no visible injury.
Elodea. In preliminary tests with attached elodealeaves, a 540 treatment for 1 min produced necrosisof all treated tissue, a 530 treatment produced chloro-sis and necrosis of the summit imbricated leaves, andtreatments below 520 produced no visiible injury.In the present study, it was found that all cells
1685
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
"'.f"-;..i 2I..
i..il
CX W-
^ -.s.N z..i ..PM i.......w. ji;e ,.X2, X't st4 ^ ^ gs
_S . ;_
W e w jt t -f̂.. ..
,wi ..
'S
.. Y. aa_
.~~~~~~~~~~~~~~~~~~~~~~~~~~~iM,.,;~~~~~~~~~~~~~~~~~~~J s F
lbl
1686
....S
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

DANIELL, ET AL.-HIGH HEAT EFFECTS ON PLANT CELLS.
within an elodea leaf did not exhibit the same degreeof injury at a particular temperature as was truewith soybean leaves. Even in the 540 treatment,cytoplasmic streaming was observed in cells adjacentto cells showing a high degree of plasmolysis.
In the treatments from 48 to 510, there appearedto be a reversible cessation of cytoplasmic streamingin all of the cells. This may have been a shockeffect of the treatments, since the cytoplasm resumedstreaming within 6 to 8 hr in the majority of cellsobserved. Vital stains accumulated in the vacuoleof the cells and not the cytoplasm, indicating that thecells were still living. In the check treatment, ap-proximately 3 % of the cells exhibited injury. Thisinjury was classed as mechanical injury and thispercentage was sulbtracted from the total cell injuryfor each treatment.
In the 520 treatment, approximately 60 % of thecells showed no apparent evidence of injury. How-ever, 15 % of these undamaged cells stained witheach of the non-vital stains, indicating that a changein permealbility of membranes had occurred. Thecytoplasm of approximately 15 % of the cells stainedwhen the vital stains were used, indicating they werenon-living. Toluidine blue, which has been used todetermin-e disorganized chloroplasts, stained thechloroplasts in these cells a deep blue or brownishblue color. Approximately one-half of the cells thatstained with the non-vital stains also exhibited plas-molysis and chloroplast leakage. The remaining25 % of the cells exhibited enlarged chloroplasts andnuclei. Cells with enlarged chloroplasts are shownin Fig. lc.
In the 530 treatment, approximately 35 % of thecells showed no evidence of injury. Approximately40 % of the cells stained with the non-vital stainsand approximately the same number of cells wereclassed as "dead" when the vital stains were used.The majority of cells which stained with the non-vital stains also exhibited plasmolysis and bleachedchloroplasts. The remaining 25 % of the cells con-tained seemingly enlarged chloroplasts and nuclei.At 540 and higher temperature treatments, approxi-mately 90 % of the cells showed irreversible plas-
molysis, bleached chloroplasts, and other criteria ofcell death.
Microscopic observations were made with elodealeaves which were mounted on the first, second,fourth, seventh, and twelfth day after the hot watertreatments. In the lower temperature treatments(43 through 520), the percentages of cells exhibitingcriteria of death 12 d-ays after treatment did notchange from the percentages determined immediatelyafter treatment. Also, in the lower temperaturetreatments which produced enlargement of chloro-plasts and cessation of cyclosis, there appeared to beno enlargement of any chloroplasts after the secondday. Approximately 90 % of the cells observed inthe lower temperature treatments exhibited cyclosisat each observation. All of the leaf cells treated at530 and above exhibited characteristics of cell deathon the fourth day after treatment.
Some of the characteristics of cell death andchanges in the cells at or after thermal death whichwere observed in elodea leaves are shown in Fig. ld,le, lf, and lg. In plasmolyzed cells of elodea leavesobserved immediately after treatment (Fig. ld),there appeared to be chlorophyll in the cytoplasm(indicated by the green color in the cytoplasm) andthe chloroplasts appeared shrunken. The chloro-plasts stained dark blue with toluidine blue indicatinga disorganization of the chloroplasts (Fig. le).Cells which had been treated in a water bath at 5302 days previously are shown in Fig. lf and lg. Thechloroplasts appeared intact in some of the cells.The cytoplasm appeared to be coagulated or gelled(also indicated by lack of Brownian movement) andpulled away from the cell wall. Rod-like fibrils wereapparent in the cytoplasm. The plasma memibraneappeared to be breaking down in some of the plas-molyzed cells. Leaf cells which had been treated4 days previously at 540 are shown in Fig. lh. Thechloroplasts were bleached and only remnants of thechloroplasts were apparent in some cells. The proto-plasm of the cells showed characteristics of coagula-tion and the plasma membrane appeared to be dis-rupted. The nucleus appeared to be breaking downin some of the cells at this time.
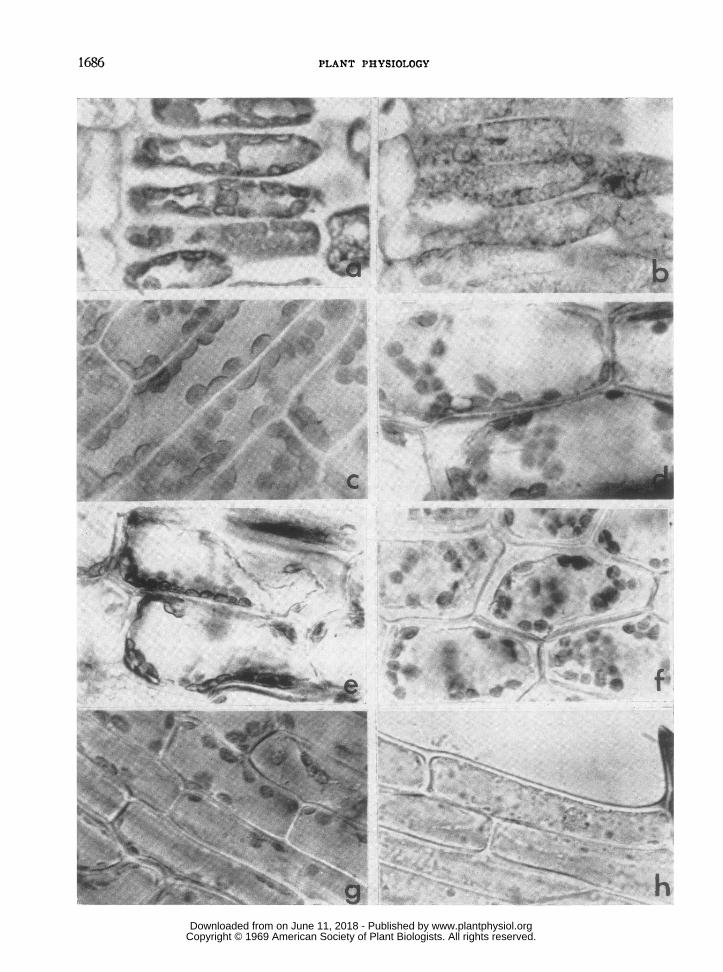
FIG. la. Cross section of untreated control soybean leaf. Chloroplasts appear normal, abutted to each other,and closely appressed to the cell wall. The vacuoles and nuclei are distinct, X 900.
FIG. lb. Cross section of soybean leaf which had been exposed to a flame at 10420 for 130 msec. Note thedisruption of the chloroplasts and nuclei and the leakage of the contents of the cytoplasm into the vacuoles, X 900.
FIG. IC. Cells of Elodea leaf treated with sublethal temperature of 520. The chloroplasts are enlarged (compareIc with Id) and show lack of orientation. Cytoplasmic streaming occurred while the microphotograph was taken,X 900.
FIGS. ld and le. Leaf cells of Elodea showing plasmolysis and damaged chloroplasts immediately after a lethaltreatment. The leaves had been immersed in a hot water bath at 54°. The cells in Fig. le were stained with tolui-dine blue. There appears to be leakage of chlorophyll into the cytoplasm, X 900.
FIGS. If, Ig, and lh. Leaf cells of Elodea which had been treated with lethal temperatures by placing in hotwater baths for 1 min, X 900. Cells in Fig. lf had been treated 2 days previously at 540. Note the plasmolysisand damaged chloroplasts. Cells in Fig. Ig had been treated 2 days previously at 530. Note that 2 of the cells haveintact nuclei while 1 cell exhibits extreme plasmolysis. Cells in Fig. lh had been treated 4 days previously at 540.Only remnants of the chloroplasts are apparent. One peripheral cell shows a nucleus in the process of disintegration.
1687
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
Discussion
In this study, the behavior of "vital stains" wasused to identify dead cells. Most of the cells whichwere classed as "dead" by this method also exhibitedother criteria of cell death or injury, such as sup-pression of protoplasmic streaming, bleached chloro-plasts, plasmolysis, and changes in membrane perme-ability. The other stains were used to show changesin membrane permeability. Some cells showed in-jury, such as enlargement of chloroplasts and cessa-tion of cytoplasmiic streaming, but were not classifiedas "dead" when the vital stains were used. Thisinjury was considered reversible in this study. Theinterpretation that some injury was reversible issupported by the test where mounts of elodea leaveswere observed at specific dates after treatment. Insublethal temperature treatments resulting in cessa-tion of cytoplasm,ic streaming and enlargement ofchloroplasts, the cytoplasm resumed streaming 6 to8 hr after the heat treatments. There appeared tobe no enlargement of chloroplasts beyond the secondday after treatment.
The apparent deleterious swelling and loss ofchlorophyll by the chloroplasts observed in this workmay not be a criterion of cell death. Hartley (5)observed that excessive heat causes whitespot lesionson stems of plants but not necrosis. Observationsmade in a thermal death point experiment (Daniell,unpublished data) suggested that chlorosis in vivoof the leaves of soybeans apparently occurs at atemperature of 1° less than that inducing necrosisof the tissue.
An excellent discussion on the effects of hightemperature on membranes and membrane constitu-ents has been presented by Chapman (4). Someresearch supporting the "lipoid liberation" theory ispresented. With this theory, high temperature in-jury, whether reversible or irreversible, is attributedto the melting of lipid constituents in the cell or cellmemibrane. This theory is linked with the observa-tion that lipid formed by living organisms at hightemperatures is more solid than lipid formed at lowertemperatures. Changes in lipids within the cellmembranes could account for the membrane changesat high temperatures which were observed in thepresent study.
The plasmolysis observed in most of the cells atlethal temperature treatments did not appear to 'be aprimary event at the thermal death of a cell. Plas-molysis was not apparent in all cells which exhibitedother criteria of cell death. It has been suggestedthat plasmolysis is reversible in most cells (2) andmay be a function of living cells (16).
Approximately 40 % of the cells in elodea leavesshow cellular disorganization at a temperature treat-ment which results in death of the whole leaf. Thisagrees with Berkley and Berkley (3) finding thatplasmolysis and deformation of the nuclei were not
uniformly distributed in cells of cotton plants whichhad been subjected to high temperatures.
All enzymes are not inactivated at temperatureswhich produces necrosis of all treated tissue. Pre-vious tests (Daniell, unpublished data) showed thatthe respiration rate of tissue treated at lethal tem-peratures was reduced about 50 % but continued ata steady rate for over 2 hr after the lethal treatment.Isolated chloroplasts subjected to lethal temperaturesstill reduced potassium ferricyanide several hr aftertreatment, but the Hill reaction was significantlyreduced. This could suggest that the action of lethaltemperature is not complete denaturation of proteinsbut is breakage of weak bonds such as those in themembranes of cells.
The results obtained in these cytological testsshow that the primary event at the thermal deathpoint of a cell is the structural change of the cellularmembranes. It also shows that structural changeswithin the cells were similar in the 2 types of treat-ments but were much more pronounced when thetemperature of the cells changes rapidly (as in theflame treatments) than when the temperature changesare more gradual (as in the hot-water-bath treat-ments). Evidence that there are structural changesin the membrane is apparent in the prepared slidesof soybean leaves which had been subjected to sub-lethal or lethal temperatures. In some cells, thetonoplast, the chloroplast membranes, and the nuclearmembranes were broken or extensively altered atlethal temperatures.
Literature Cited
1. ALEXANDROV, V. Y. 1964. Cytophysiological andcytoecological investigations of heat resistance ofplant cells toward the action of high and low temp-erature. Quart. Rev. Biol. 39: 35-77.
2. BENNET-CLARK, T. A. 1959. Water relations ofcells. In: Plant Physiology. F. C. Steward, ed.Academic Press, New York. p 105-91.
3. BERKLEY, D. M. AND E. E. BERKLEY. 1933. Superoptimal and thermal death temperatures of thecotton plant as affected by variations in relativehumidity. Ann. Missouri Botan. Garden 20: 583-604.
4. CHAPMAN, D. 1967. The effect of heat on mem-branes and membrane constituents. In: Thermo-biology. H. Rose, ed. Academic Press, NewYork. p 123-46.
5. HARTLEY, C. 1918. Stem lesions caused by exces-sive heat. J. Agr. Res. 14: 595-610.
6. HARVEY, R. B. 1924. Sunscald of tomatoes. Minn.Studies Plant Sci. Biol. Sci. 5: 229-33.
7. JENSEN, W. A. 1962. Botanical Histochemistry.W. H. Freeman and Company, San Francisco,California. 408 p.
8. KOFFLER, H., G. E. MALLETT, AND J. ADYE. 1957.Molecular basis of biological stability to high tem-peratures. Proc. Natl. Acad. Sci. U. S. 43: 464-77.
1688
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.

DANIELL, ET AL.-HIGH HEAT EFFECTS ON PLANT CELLS.
9. LEPESCHKIN, W .W. 1935. Zur Kenntnis des Hitz-etodes des Protoplasmas. Protoplasma 23: 349-66.
10. LEVITT, J. 1956. The Hardiness of Plants. Aca-demic Press, New York.
11. LEVITT, J. AND G. W. SCARTH. 1936. Frost hard-ening studies with living cells. I. Osmotic andbound water changes in relation to frost resistanceand the seasonal cycle. Can. J. Research (C)14: 267-84.
12. MAxIMov, N. A. 1938. Plant Physiology. Mc-Graw-Hill Book Company Incorporated, NewYork. 473 p.
13. NIELSEN, L. W. AND F. A. TODD. 1946. Bacterialsoft rot of irish potatoes as influenced by suble-thal temperatures. Am. Potato J. 23: 73-87.
14. PUTNAM, F. W. 1953. Protein denaturation. In:The Proteins. H. Neurath and K. Bailey, eds.Academic Press, New York. p 807-92.
15. USHAKOV, V. B. 1964. Problem of the cause ofthermal death of skeletal muscles of cold-bloodedanimals. Acad. Sci. USSR (Botan. Sci. Sec.)155: 178-80.
16. VIRGIN, H. I. 1953. Physical properties of proto-plasm. Ann. Rev. Plant Physiol. 4: 363-82.
1689
www.plantphysiol.orgon June 11, 2018 - Published by Downloaded from Copyright © 1969 American Society of Plant Biologists. All rights reserved.