Embed Size (px)
Citation preview

Peptides. Vol. 6, pp. 555-561, 1985. ~ Ankho International Inc. Printed in the U.S.A. 0196-9781/85 $3,00 + .00
Effect of Oral Contraceptives on the Rat Brain and Pituitary Opioid Peptides
G O P I A. T E J W A N I , 1 K U L D E E P K. V A S W A N I , 2 A N D J O S E P H I N E C. B A R B A C C I
Department o f Pharmacology, College o f Medicine, The Ohio State University 5086 Graves Hall, 333 W. lOth Avenue, Columbus, OH 43210
R e c e i v e d 3 J a n u a r y 1985
TEJWANI, G. A,, K. K. VASWAN1 AND J. C. BARBACCI. Effect of oral contraceptives on the rat brain and pituitary opioid peptides. PEPTIDES 6(3) 555-561, 1985.--This study was designed to explore the hormonal regulation of CNS opioid peptide levels in female Sprague Dawley rats. Forty-eight animals were divided into 2 equal groups for acute and chronic studies. Each group was further divided into 4 subgroups, each containing 6 animals. Each rat in the control group received an inert pill (in 0.25 ml corn oil daily by gavage); the second group, 15 #.g norethindrone (NE, a potent progestin present in the oral contraceptive Micronor~); the third group, 15 p.g NE and 1/zg ethinyl estradiol, EE2 (present in the oral contraceptive Modicon ®) and the fourth group, 10 times the dose of the third group. Rats were treated either acutely for 5 days or chronically for 7 weeks. Opioid peptides were estimated by radioimmunoassay. Acute administration of 150/xg NE + 10/xg EE.2 decreased the levels of methionine-enkephalin (ME), leucine-enkephalin (LE), dynorphin (DYN) and fl-endorphin like immunoreactivity (B-EI) by about 50% in the pituitary. The same dose on chronic administration also decreased DYN, but increased the levels of ME and LE in the pituitary by 331 and 69%, respectively. In the hypothalamus, chronic administration of NE + EE2 increased the level of ME (155%) and LE (87%) as well as of DYN (97%). In the striatum, the levels of LE (33%) and DYN (115%) were elevated during chronic administration. It is concluded that the acute administra- tion of NE + EE2, in general, reduces the levels of ME, LE, DYN and/3-EI. The extent of this decrease is about the same in the pituitary, hypothalamus and striatum. Chronic administration of these hormones, however, results in a reversal of this decrease (except for fl-El) and actually can increase the levels of ME, LE and DYN in all three tissues.
B-Endorphin Dynorphin Methionine-enkephalin Leucine-enkephalin Oral contraceptives Gonadotropins Opioids
THE pituitary and hypothalamus contain a variety of opioid peptides such as fl-EI, ME, LE and DYN. The chemical nature and biosynthetic pathways for these peptides have been identified, but very little is known about their regula- tory mechanisms and physiological phenomenon [37]. Lo- calization of these peptidergic neurons, in close proximity to the LHRH neurons in the hypothalamus, has aroused considerable interest in their physiological role in the regula- tion of pituitary LH release [13].
Endogenous opioids and morphine have been shown to decrease gonadotropin secretion in various species, includ- ing humans, apparently by suppressing the release of LHRH from the hypothalamus [4, 6, 27, 34, 36]. The opiate antagonist naloxone causes several fold increase in the se- cretion of gonadotropin [2, 4, 7, 8, 9, 23, 27, 28], suggesting a regulatory interaction between endogenous opioids, gonadotropin and gonadal steroids. Naloxone-induced re- lease of LH is facilitated by estradiol in humans [3, 13, 40], suggesting an antagonistic effect of estradiol on the endoge- nous opioids. Opioids may play a significant role in neuroen- docrine mechanisms of gonadotropin release in the human menstrual cycle. A preovulatory increase of/3-EI in serum occurs 2 days prior to the LH surge and a postovulatory decrease in/3-EI occurs 5 days later. Decrease in/3-EI levels
is preceded by an increase in estradiol and is associated with an increase in progesterone levels in serum, suggesting that these gonadal steroids may be responsible for decreasing fl-EI levels [43]. Male rats have twice as much pituitary ME as compared to females, indicating sex-related differences [20]. The ME content of the anterior pituitary is highest in the morning of proestrous during the estrous cycle of female rats [25], indicating possible regulation of these peptides by gonadal steroids.
RIA B-El ME LE DYN LH LHRH NE
EEe
ABBREVIATIONS
Radioimmunoassay B-Endorphin like immunoreactivity Methionine-enkephalin Leucine-enkephalin Dynorphin Luteinizing hormone Luteinizing hormone releasing hormone Norethindrone ( 17a-Ethinyl- 19-nor-testosterone,
a potent progestin present in the oral contraceptive Micronor ® and Modicon ®)
Ethinyl estradiol (present in Modicon ®)
1Requests for reprints should be addressed to Gopi A. Tejwani, Ph.D., Department of Pharmacology, College of Medicine, The Ohio State University, 5086 Graves Hall, 333 W. 10th Avenue, Columbus, OH 43210.
ePresent address: Department of Neurology F-140, Albert Einstein College of Medicine, Bronx, NY 10461.
555
556 T E J W A N I , V A S W A N I A N D B A R B A C C I
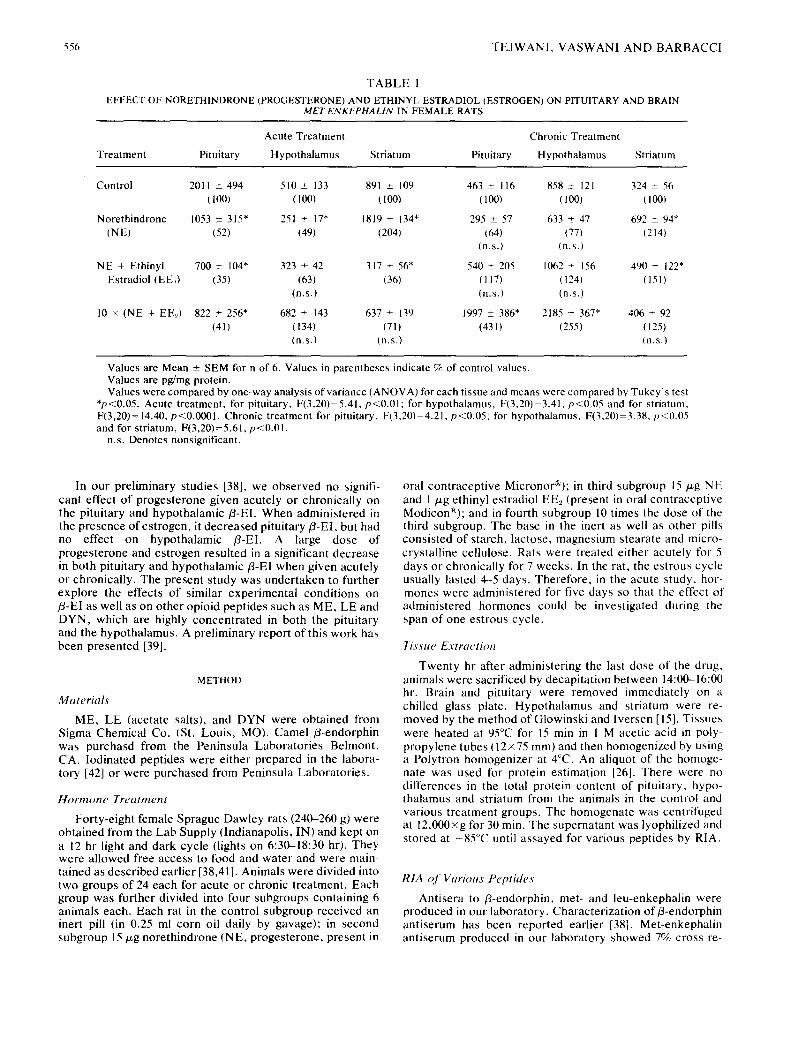
T A B L E I
EFFECT OF NORETHINDRONE (PROGESTERONE) AND ETHINYL ESTRADIOL (ESTROGEN) ON PITUITARY AND BRAIN MET-ENKEPHALIN IN FEMALE RATS
Acute Treatment Chronic Treatment
Treatment Pituitary Hypothalamus Striatum Pituitary Hypothalamus Striatum
Control 2011 ± 494 510 + 133 891 _+ 109 463 _+ 116 858 _+ 121 324 + 56 (100) (100) (100) (100) (100) (100)
Norethindrone 1053 ± 315" 251 _+ 17" 1819 + 134" 295 + 57 633 ± 47 692 _+ 94* (NE) (52) (49) (204) (64) (77) (214)
(n.s.) (n.s.)
NE + Ethinyl 700 ± 104" 323 + 42 317 + 56* 540 ± 205 1062 + 156 490 _+ 122" Estradiol (EE2) (35) (63) (36) (117) (124) (151)
(n.s.) (n.s.) (n.s.)
10 × (NE + EE_,) 822 ± 256* 682 _+ 143 637 ± 139 1997 +_ 386* 2185 _+ 367* 406 + 92 (41) (134) (71) (43 I) (255) (125)
(n.s.) (n.s.) (n.s.)
Values are Mean _+ SEM for n of 6. Values in parentheses indicate % of control values. Values are pg/mg protein. Values were compared by one-way analysis of variance (ANOVA) for each tissue and means were compared by Tukey's test
*p<0.05. Acute treatment, for pituitary, F(3,20)=5.41, p<0.01; for hypothalamus, F(3,20)=3.41, p<0,05 and for striatum, F(3,20)= 14.40, p<0.0001. Chronic treatment for pituitary, F(3,20)=4.21, p<0.05; for hypothalamus, F(3,20)=3.38, p<0.05 and for striatum, F(3,20)=5.61, p<0.01.
n.s. Denotes nonsignificant.
In our prel iminary studies [38], we observed no signifi- cant effect o f proges terone given acutely or chronical ly on the pituitary and hypothalamic fl-EI. When adminis tered in the presence of estrogen, it decreased pituitary f l -EI , but had no effect on hypothalamic fl-EI. A large dose of proges terone and es t rogen resulted in a significant decrease m both pituitary and hypothalamic 13-EI when given acutely or chronical ly. The present study was under taken to further explore the effects o f similar exper imental condi t ions on /3-EI as well as on o ther opioid peptides such as ME, LE and D Y N , which are highly concent ra ted in both the pituitary and the hypothalamus. A prel iminary report of this work has been presented [39].
METHOD
Materials
ME, LE (acetate salts), and DYN were obtained from Sigma Chemical Co. (St. Louis , MO). Camel f l -endorphin was purchasd from the Peninsula Labora tor ies Belmont , CA. Iodinated peptides were ei ther prepared in the labora- tory [42] or were purchased from Peninsula Labora tor ies .
Hormone Treatment
Forty-eight female Sprague Dawley rats (240-260 g) were obtained f rom the Lab Supply (Indianapolis, IN) and kept on a 12 hr light and dark cycle (lights on 6:30-18:30 hr). They were al lowed free access to food and water and were main- tained as descr ibed earl ier [38,41]. Animals were divided into two groups o f 24 each for acute or chronic t reatment . Each group was further divided into four subgroups containing 6 animals each. Each rat in the control subgroup rece ived an inert pill (in 0.25 ml corn oil daily by gavage); in second subgroup 15 keg norethindrone (NE, proges terone , present in
oral cont racept ive Micronor®); in third subgroup 15 keg NE and 1 keg ethinyl estradiol EE2 (present in oral cont racept ive Modicon®); and in fourth subgroup 10 t imes the dose of the third subgroup. The base in the inert as well as other pills consis ted of starch, lactose, magnesium stearate and micro- crystal l ine cellulose. Rats were treated ei ther acutely for 5 days or chronical ly for 7 weeks. In the rat, the estrous cycle usually lasted 4-5 days. Therefore , in the acute study, hor- mones were administered for five days so that the effect of administered hormones could be investigated during the span of one estrous cycle.
7issue Extraction
Twenty hr after administer ing the last dose of the drug, animals were sacrificed by decapitat ion be tween 14:00--16:00 hr. Brain and pituitary were removed immediately on a chilled glass plate. Hypotha lamus and striatum were re- moved by the method of Glowinski and Iversen [15]. Tissues were heated at 95°C for 15 min in 1 M acetic acid in poly- propylene tubes (12 × 75 mm) and then homogenized by using a Polytron homogenizer at 4°C. An aliquot of the homoge- hate was used for protein est imation [26]. There were no differences in the total protein content of pituitary, hypo- thalamus and striatum from the animals in the control and various t reatment groups. The homogenate was centrifuged at 12,000×g for 30 rain. The supernatant was lyophilized and stored at - 85°C until assayed for various peptides by RIA.
RIA o f Various Peptides
Antisera to /3-endorphin, met- and leu-enkephalin were produced in our laboratory. Character izat ion of f l-endorphin ant iserum has been reported earl ier [38]. Met-enkephal in ant iserum produced in our laboratory showed 7%, cross re-
OPIOID PEPTIDES AND F E M A L E SEX HORMONES 557
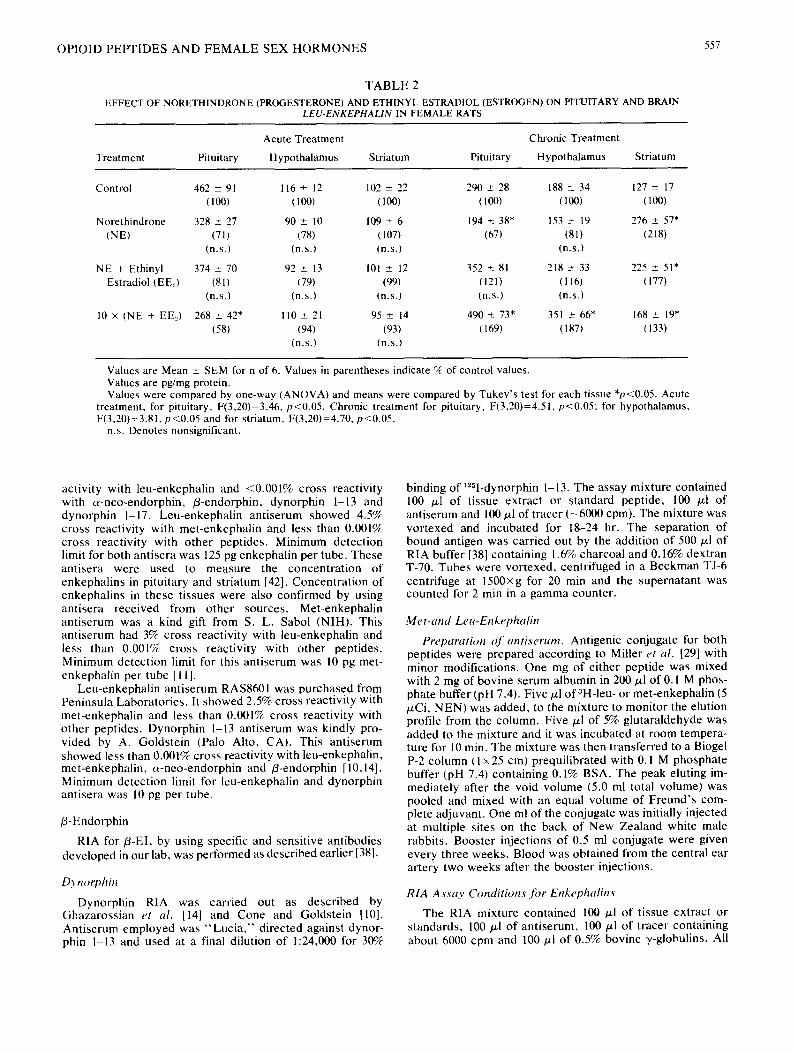
TABLE 2
EFFECT OF NORETHINDRONE (PROGESTERONE) AND ETHINYL ESTRADIOL (ESTROGEN) ON PITUITARY AND BRAIN LEU-ENKEPHALIN IN FEMALE RATS
Acute Treatment Chronic Treatment
Treatment Pituitary Hypothalamus Striatum Pituitary Hypothalamus Striatum
Control 462 _+ 91 116 + 12 102 ± 22 290 _+ 28 188 ± 34 127 ± 17 (100) (100) (100) (100) (100) (100)
Norethindrone 328 ± 27 90 ± 10 109 + 6 194 ± 38* 153 ± 19 276 ± 57* (NE) (71) (78) (107) (67) (81) (218)
(n.s.) (n.s.) (n.s.) (n.s.)
NE + Ethinyl 374 + 70 92 + 13 101 ± 12 352 ± 81 218 ± 33 225 ± 51" Estradiol (EEl) (81) (79) (99) (121) (116) (177)
(n.s.) (n.s.) (n.s.) (n.s.) (n.s.)
10 × (NE + EE.,) 268 ± 42* 110 + 21 95 _+ 14 490 ± 73* 351 ± 66* 168 ± 19" (58) (94) (93) (169) (187) (133)
(n.s.) (n.s.)
Values are Mean ± SEM for n of 6. Values in parentheses indicate % of control values. Values are pg/mg protein. Values were compared by one-way (ANOVA) and means were compared by Tukey's test for each tissue *p<0.05. Acute
treatment, for pituitary, F(3,20)=3.46, p<0.05, Chronic treatment for pituitary, F(3,20)=4.51, p<0.05; for hypothalamus, F(3,20)=3.81, p<0.05 and for striatum, F(3,20)=4.70, p<0.05.
n.s. Denotes nonsignificant.
activity with leu-enkephalin and <0.001% cross reactivity with c~-neo-endorphin, /3-endorphin, dynorphin 1-13 and dynorphin 1-17. Leu-enkephalin antiserum showed 4.5% cross reactivity with met-enkephalin and less than 0.001% cross reactivity with other peptides. Minimum detection limit for both antisera was 125 pg enkephalin per tube. These antisera were used to measure the concentration of enkephalins in pituitary and striatum [42]. Concentration of enkephalins in these tissues were also confirmed by using antisera received from other sources. Met-enkephalin antiserum was a kind gift from S. L. Sabol (NIH). This antiserum had 3% cross reactivity with leu-enkephalin and less than 0.001% cross reactivity with other peptides. Minimum detection limit for this antiserum was 10 pg met- enkephalin per tube [111.
Leu-enkephalin antiserum RAS8601 was purchased from Peninsula Laboratories. It showed 2.5% cross reactivity with met-enkephalin and less than 0.001% cross reactivity with other peptides. Dynorphin 1-13 antiserum was kindly pro- vided by A. Goldstein (Palo Alto, CA). This antiserum showed less than 0.001% cross reactivity with leu-enkephalin, met-enkephalin, c~-neo-endorphin and /3-endorphin [10,14]. Minimum detection limit for leu-enkephalin and dynorphin antisera was 10 pg per tube.
/3-Endorphin
RIA for/3-EI, by using specific and sensitive antibodies developed in our lab, was performed as described earlier [38].
Dynorphin
Dynorphin RIA was carried out as described by Ghazarossian et al. [14] and Cone and Goldstein [10]. Antiserum employed was " L u c i a , " directed against dynor- phin 1-13 and used at a final dilution of 1:24,000 for 30%
binding of lzSI-dynorphin 1-13. The assay mixture contained 100 /xl of tissue extract or standard peptide, 100 /xl of antiserum and 100 p.l of tracer (-6000 cpm). The mixture was vortexed and incubated for 18--24 hr. The separation of bound antigen was carried out by the addition of 500 ~1 of RIA buffer [38] containing 1.6% charcoal and 0.16% dextran T-70. Tubes were vortexed, centrifuged in a Beckman TJ-6 centrifuge at 1500×g for 20 min and the supernatant was counted for 2 min in a gamma counter.
Met-and Leu-Enkephalin
Preparation o f antiserum. Antigenic conjugate for both peptides were prepared according to Miller et al. [29] with minor modifications. One mg of either peptide was mixed with 2 mg of bovine serum albumin in 200/~1 of 0. I M phos- phate buffer (pH 7.4). Five/~1 of3H-leu - or met-enkephalin (5 p.Ci, NEN) was added, to the mixture to monitor the elution profile from the column. Five ~1 of 5% glutaraldehyde was added to the mixture and it was incubated at room tempera- ture for 10 min. The mixture was then transferred to a Biogel P-2 column (1 × 25 cm) prequilibrated with 0. I M phosphate buffer (pH 7.4) containing 0.1% BSA. The peak eluting im- mediately after the void volume (5.0 ml total volume) was pooled and mixed with an equal volume of Freund's com- plete adjuvant. One ml of the conjugate was initially injected at multiple sites on the back of New Zealand white male rabbits. Booster injections of 0.5 ml conjugate were given every three weeks. Blood was obtained from the central ear artery two weeks after the booster injections.
RIA Assay Conditions fi~r Enkephalins
The RIA mixture contained 10g p.l of tissue extract or standards, 100/~1 of antiserum, 100 ~tl of tracer containing about 6000 cpm and 100 ~1 of 0.5% bovine y-globulins. All

558 T E J W A N | , VASWANI AND BARBACCI
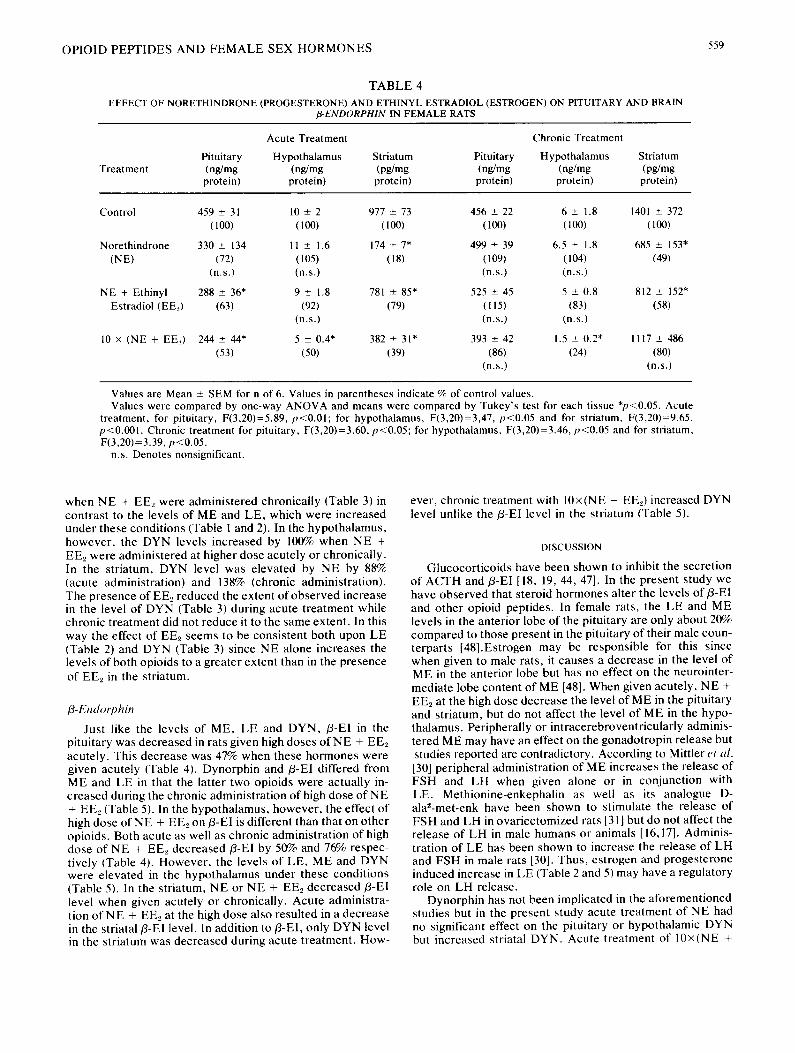
TABLE 3
EFFECT OF NORETHINDRONE (PROGESTERONE) AND ETHINYL ESTRADIOL (ESTROGEN) ON PITUITARY AND BRAIN D YNORPHIN IN FEMALE RATS
Acute Treatment Chronic Treatment
Treatment Pituitary Hypothalamus Striatum Pituitary Hypothalamus Striatum
Control 400 ± 82 134 ± 22 83 ± 8 693 ± 121 368 _+ 47 58 ± 5 (100) (100) (I00) (100) (100) (100)
Norethindrone 481 ± 38 111 ± 19 155 ± 21" 434 ± 83* 282 ± 15" 138 _+ 21" (NE) (120) (83) (188) (63) (77) (238)
(n.s.) (n.s.)
NE + Ethinyl 488 + 71 165 ± 27 33 ± 5* 361 ± 77* 317 ± 39 89 ± 5* Estradiol (E E._,) (122) (123) (40) (52) (86) (152)
(n.s.) (n.s.) (n.s.)
10 × (NE + EEl) 236 ± 38* 275 ± 48* 33 ± 3* 476 ± 63* 725 ± 118" 125 ± 34* (59) (205) (39) (69) (197) (125)
Values are Mean _+ SEM for n of 6. Values in parentheses indicate % of control values. Values are pg/mg protein. Values were compared by one-way ANOVA and means were compared by Tukey's test for each tissue *p<0.05. Acute
treatment, for pituitary, F(3,20)=3.68, p<0.05; for hypothalamus, F(3,20)=4.03, p<0.05 and for striatum, F(3,20)=7.61, p<0.01. Chronic treatment for pituitary, F(3,20)=4.92, p<0.05; for hypothalamus, F(3,20)=5.21, p<0.05 and for striatum, F(3,20)=6.97, p<0.01.
n.s. Denotes nonsignificant.
dilutions were carried out in the RIA assay buffer as de- scribed for/~-EI [38]. Tubes were vortexed and incubated for 18-24 hr at 4°C. Bound antigen was separated by adding 400 /~1 of 30% polyethylene glycol (8000). After mixing, the tubes were further incubated at 4°C for 20 min. After centrifuging at 1500xg for 20 min, the precipitate was counted for 2 min in a gamma counter.
Statistical Analysis
The results obtained were compared using Analysis of Variance (ANOVA) and means were compared by Tukey's test. In this paper, the concentrations of opioid peptides are expressed as pg/mg protein. Differences in the results be- tween the control and various treatment groups were the same whether the concentrations of opioid peptides were expressed per mg of protein or per g of tissue.
RESULTS
Met-Enkephalin (ME)
Norethindrone (NE) when given acutely (for five days) caused about a 50% decrease in the level of ME, in both the pituitary and the hypothalamus. This inhibition was slightly increased in the presence of ethinyl estradiol (EE2, Table 1). However, chronic administration of EEz in the presence of NE (10xdose) increased ME by 155% in the hypothalamus and 331% in the pituitary. Striatal levels of ME were in- creased by 104% by NE when given acutely and 114%, when given chronically. Acute administration of EE2 in the pres- ence of NE not only abolished the increase in the level of striatal ME, but decreased it further by 64%. Thus, it appears that NE + EE2, when given acutely, decrease levels of ME, but when given chronically increase the level of this
enkephalin in the hypothalamus and pituitary. However, in the case of striatum, this increase in the level of ME was observed in the presence of NE alone. We observed that ME levels in the control animals receiving " iner t " pill chroni- cally, decreased by 64 and 77% in striatum and pituitary, respectively (Table 1). This decrease in ME levels may be due to the presence of a number of ingredients in the "inert pill" (see the Method section) or due to the stress experi- enced by animals during daily garage for seven weeks.
Leu-Enkephalin (LE)
Low doses of NE or NE + EE., unlike their effect on ME levels, did not significantly affect the levels of LE in the hypothalamus or pituitary, whether given acutely or chroni- cally. Acute administration of high doses of NE + EE2 de- creased LE levels in the pituitary by 42% (Table 2). How- ever, like the levels of ME, LE levels were also substantially increased in the hypothalamus as well as in the pituitary upon chronic administration of 10x dose of NE + EEl. In the striatum, NE alone, caused a greater increase in the level of LE compared to when NE was given together with EE.,, chronically. Chronic administration of these hormones especially at higher dose, results in a substantial increase in the levels of ME and LE. In the striatum also, chronic ad- ministration of these hormones results in an increase in the levels of both ME and LE, but NE alone seems to be more effective in causing this increase than NE and EE~ together.
Dynorphin 1-13
Like ME and LE, the level of DYN in the pituitary de- creased upon acute administration of high dose of NE + EE~ (Table 3). Dynorphin levels in the pituitary remained decreased
OPIOID PEI:q'IDES AND F E M A L E SEX HORMONES 559
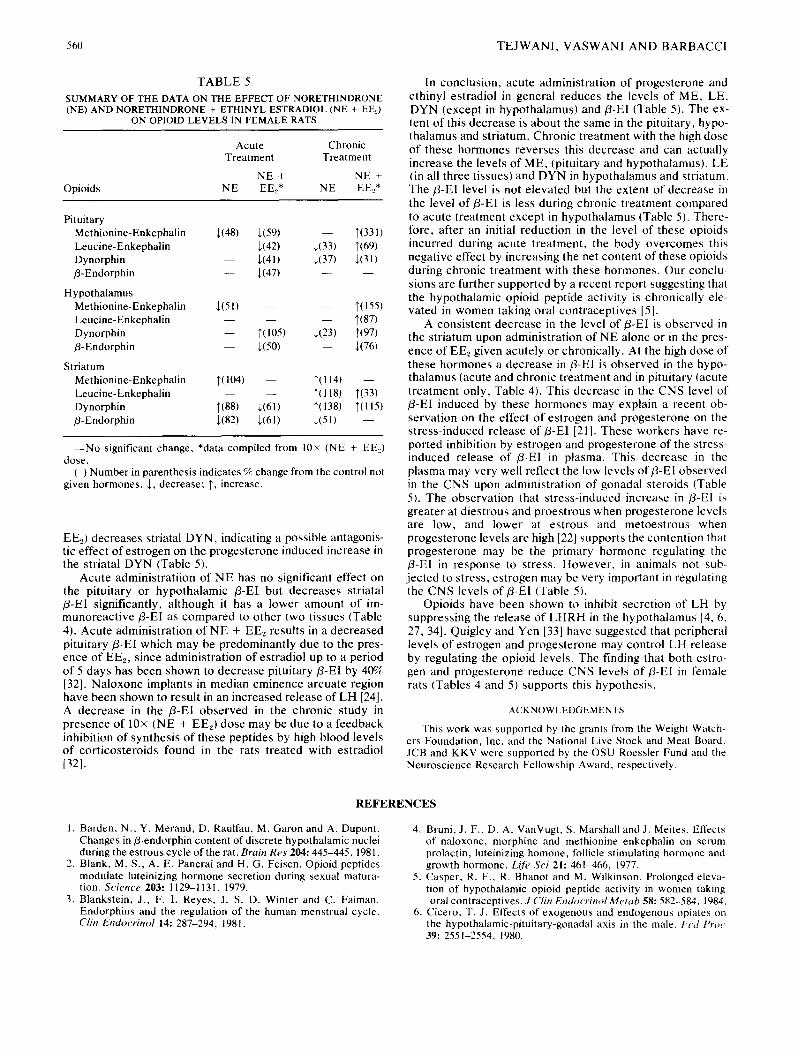
TABLE 4
EFFECT OF NORETHINDRONE (PROGESTERONE) AND ETHINYL ESTRADIOL (ESTROGEN) ON PITUITARY AND BRAIN fl-ENDORPHIN IN FEMALE RATS
Treatment
Acute Treatment Chronic Treatment
Pituitary Hypothalamus Striatum Pituitary Hypothalamus Striatum (ng/mg (ng/mg (pg/mg (ng/mg (ng/mg (pg/mg protein) protein) protein) protein) protein) protein)
Control 459 ± 31 10 +_ 2 977 ± 73 456 ± 22 6 ± 1.8 1401 ± 372 (100) (lO0) (100) (100) (100) (100)
Norethindrone 330 _+ 134 11 ± 1.6 174 + 7* 499 ± 39 6.5 + 1.8 685 ± 153" (N E) (72) (105) (18) (109) (104) (49}
(n.s.) (n.s.) (n.s.) (n.s.)
NE + Ethinyl 288 ± 36* 9 ± 1.8 781 ± 85* 525 ± 45 5 ± 0.8 812 + 152" Estradiol (EEe) (63) (92) (79) (115) (83) (58)
(n.s.) (n.s.) (n.s.)
10 × (NE + EE._,) 244 ± 44* 5 ± 0.4* 382 ± 31" 393 ± 42 1.5 + 0.2* 1117 ± 486 (53) (50) (39) (86) (24) (80)
(n.s.) (n.s.)
Values are Mean ± SEM for n of 6. Values in parentheses indicate % of control values. Values were compared by one-way ANOVA and means were compared by Tukey's test for each tissue *p<0.05. Acute
treatment, for pituitary, F(3,20)=5.89, p<0.01; for hypothalamus, F(3,20)=3,47, p<0.05 and for striatum, F(3,20)=9.65, p<0.001. Chronic treatment for pituitary, F(3,20)=3.60, p<0.05; for hypothalamus, F(3,20)=3.46, p<0.05 and for striatum, F(3,20)=3.39, p<0.05.
n.s. Denotes nonsignificant.
when NE + EE2 were administered chronically (Table 3) in contrast to the levels of ME and LE, which were increased under these conditions (Table 1 and 2). In the hypothalamus, however, the DYN levels increased by 100% when NE + EEz were administered at higher dose acutely or chronically. In the striatum, DYN level was elevated by NE by 88% (acute administration) and 138% (chronic administration). The presence of EE~ reduced the extent of observed increase in the level of DYN (Table 3) during acute treatment while chronic treatment did not reduce it to the same extent. In this way the effect of EE2 seems to be consistent both upon LE (Table 2) and DYN (Table 3) since NE alone increases the levels of both opioids to a greater extent than in the presence of EEz in the striatum.
fl-Endorphin
Just like the levels of ME, LE and DYN, fl-El in the pituitary was decreased in rats given high doses o f N E + EEe acutely. This decrease was 47% when these hormones were given acutely (Table 4). Dynorphin and fl-EI differed from ME and LE in that the latter two opioids were actually in- creased during the chronic administration of high dose of NE + EE~ (Table 5). In the hypothalamus, however, the effect of high dose o f N E + EE2 on fl-EI is different than that on other opioids. Both acute as well as chronic administration of high dose of NE + EE2 decreased fl-EI by 50% and 76% respec- tively (Table 4). However, the levels of LE, ME and DYN were elevated in the hypothalamus under these conditions (Table 5). In the striatum, NE or NE + EE2 decreased fl-E1 level when given acutely or chronically. Acute administra- tion of NE + EE2 at the high dose also resulted in a decrease in the striatal fl-E1 level. In addition to fl-E1, only DYN level in the striatum was decreased during acute treatment. How-
ever, chronic treatment with 10×(NE + EE2) increased DYN level unlike the fl-EI level in the striatum (Table 5).
DISCUSSION
Glucocorticoids have been shown to inhibit the secretion of ACTH and fl-EI [18, 19, 44, 47]. In the present study we have observed that steroid hormones alter the levels of fl-El and other opioid peptides. In female rats, the LE and ME levels in the anterior lobe of the pituitary are only about 20% compared to those present in the pituitary of their male coun- terparts [48].Estrogen may be responsible for this since when given to male rats, it causes a decrease in the level of ME in the anterior lobe but has no effect on the neurointer- mediate lobe content of ME [48]. When given acutely, N E + EE2 at the high dose decrease the level of ME in the pituitary and striatum, but do not affect the level of ME in the hypo- thalamus. Peripherally or intracerebroventricularly adminis- tered ME may have an effect on the gonadotropin release but studies reported are contradictory. According to Mittler et al. [30] peripheral administration of ME increases the release of FSH and LH when given alone or in conjunction with LE. Methionine-enkephalin as well as its analogue D- ala2-met-enk have been shown to stimulate the release of FSH and LH in ovariectomized rats [31] but do not affect the release of LH in male humans or animals [16,17]. Adminis- tration of LE has been shown to increase the release of LH and FSH in male rats [30]. Thus, estrogen and progesterone induced increase in LE (Table 2 and 5) may have a regulatory role on LH release.
Dynorphin has not been implicated in the aforementioned studies but in the present study acute treatment of NE had no significant effect on the pituitary or hypothalamic DYN but increased striatal DYN. Acute treatment of 10×(NE +
560 T E J W A N I , V A S W A N I A N D B A R B A C C I
T A B L E 5
SUMMARY OF THE DATA ON THE EFFECT OF NORETH1NDRONE (NE) AND NORETHINDRONE + ETHINYL ESTRADIOL (NE + EE0
ON OPIOID LEVELS IN FEMALE RATS
Opioids
Acute Chronic Treatment Treatment
NE + NE + NE EE_,* NE EEe*
Pituitary Methionine-Enkephalin ~(48) ~(59) - - 1"(331) Leucine-Enkephalin - - ~(42) ~(33) 1'(69) Dynorphin - - ~,(41) ~(37) ~(31) /3-Endorphin - - ~(47) - - - -
Hypothalamus Methionine-Enkephalin ,~(51 ) - - - - T(155) Leucine-Enkephalin - - - - - - I'(87) Dynorphin - - T(105) +(23) 1"(97) /3-Endorphin - - ¢(50) - - ~(76)
Striatum Methionine-Enkephalin 1"(104) - - 1'(114) - - Leucine-Enkephalin - - - - ~'(118) T(33) Dynorphin 1'(88) +(61) ]'(138) l'(115) /3-Endorphin ~,(82) ~(61) ~,(51) - -
- - N o significant change, *data compiled from 10x (NE + EE=,) dose.
( ) Number in parenthesis indicates % change from the control not given hormones. ], decrease; T, increase.
EEO decreases striatal DYN, indicating a possible antagonis- tic effect of es t rogen on the proges terone induced increase in the striatal DYN (Table 5).
Acute administrat i ion of N E has no significant effect on the pituitary or hypothalamic /3-EI but decreases striatal /3-El significantly, al though it has a lower amount of im- munoreac t ive /3 -EI as compared to o ther two tissues (Table 4). Acute administrat ion of N E + EE~ results in a decreased pituitary f l-EI which may be predominant ly due to the pres- ence of EE2, since administrat ion o f estradiol up to a period of 5 days has been shown to decrease pi tui tary/3-EI by 40% [32]. Na loxone implants in median eminence arcuate region have been shown to result in an increased release of L H [24]. A decrease in the /3-EI observed in the chronic study in presence of 10x (NE + E E 0 dose may be due to a feedback inhibition of synthesis of these peptides by high blood levels o f cor t icosteroids found in the rats treated with estradiol [32].
In conclusion, acute administrat ion of progesterone and ethinyl estradiol in general reduces the levels of ME, LE, DYN (except in hypothalamus) and fl-EI (Table 5). The ex- tent o f this decrease is about the same in the pituitary, hypo- thalamus and striatum. Chronic t reatment with the high dose of these hormones reverses this decrease and can actually increase the levels of ME, (pituitary and hypothalamus) , LE (in all three tissues) and D Y N in hypothalamus and striatum. T h e / 3 - E l level is not e levated but the extent of decrease in the level o f /3 -E l is less during chronic t reatment compared to acute t reatment except in hypothalamus (Table 5). There- fore, after an initial reduct ion in the level of these opioids incurred during acute t reatment , the body ove rcomes this negat ive effect by increasing the net content of these opioids during chronic t rea tment with these hormones . Our conclu- sions are further supported by a recent report suggesting that the hypothalamic opioid peptide act ivi ty is chronical ly ele- vated in women taking oral cont racept ives [5].
A consis tent decrease in the level of fl-EI is observed in the striatum upon administrat ion of N E alone or in the pres- ence of EE., given acutely or chronically. At the high dose of these hormones a decrease in fl-EI is observed in the hypo- thalamus (acute and chronic t rea tment and in pituitary (acute t rea tment only, Table 4). This decrease in the CNS level of f l-EI induced by these hormones may explain a recent ob- servat ion on the effect o f es t rogen and progesterone on the s tress- induced release o f /3 -E l [21]. These workers have re- ported inhibition by es t rogen and proges terone of the stress- induced release of /3-El in plasma. This decrease in the plasma may very well reflect the low levels o f f l -E l observed in the CNS upon administrat ion of gonadal steroids tTable 5). The observat ion that s tress-induced increase in f l-Ei is greater at diestrous and proestrous when progesterone levels are low, and lower at estrous and metoes t rous when proges terone levels are high [22] supports the content ion that proges terone may be the primary hormone regulating the fl-EI in response to stress. Howeve r , in animals not sub- j ec ted to stress, es t rogen may be very important in regulating the C N S levels of /3-El (Table 5).
Opioids have been shown to inhibit secret ion of LH by suppressing the release of L H R H in the hypothalamus [4, 6, 27, 34]. Quigley and Yen [33] have suggested that peripheral levels of estrogen and proges terone may control LH release by regulating the opioid levels. The finding that both estro- gen and progesterone reduce CNS levels of /3-EI in female rats (Tables 4 and 5) supports this hypothesis .
ACKNOWLEDGEMENTS
This work was supported by the grants from the Weight Watch- ers Foundation, Inc. and the National Live Stock and Meat Board. JCB and KKV were supported by the OSU Roessler Fund and the Neuroscience Research Fellowship Award, respectively.
R E F E R E N C E S
I. Barden, N., Y. Merand, D. Raulfau, M. Garon and A. Dupont. Changes in/3-endorphin content of discrete hypothalamic nuclei during the estrous cycle of the rat. Brain Res 204: 445-445. 1981.
2. Blank, M. S., A. E. Panerai and H. G. Feisen. Opioid peptides modulate luteinizing hormone secretion during sexual matura- tion. Science 203:1129-1131, 1979.
3. Blankstein, J., F. 1. Reyes, J. S. D. Winter and C. Faiman. Endorphins and the regulation of the human menstrual cycle. CHn Endocrinol 14: 287-294, 1981.
4. Bruni, J. F., D. A. VanVugt, S. Marshall and J. Meites. Effects of naloxone, morphine and methionine enkephalin on serum prolactin, luteinizing homone, follicle stimulating hormone and growth hormone. Life Sci 21: 461-466, 1977.
5. Casper, R. F., R. Bhanot and M. Wilkinson. Prolonged eleva- tion of hypothalamic opioid peptide activity in women taking oral contraceptives. J Clin Endocrinol Metab 58: 582-584, 1984.
6. Cicero, T. J. Effects of exogenous and endogenous opiates on the hypothalamic-pituitary-gonadal axis in the male. Fed Proc 39: 2551-2554, 1980.

OPIOID P E P T I D E S A N D F E M A L E S E X H O R M O N E S 561
7. Cicero, T. J., B. A. Schainker and E. R. Meyer. Endogenous opiates participate in the regulation of the hypothalamic- pituitary-lutenizing hormone axis and testosterone's negative feedback control of lutenizing hormone. Endocrinology 104: 1286-1291, 1979.
8. Cicero, T. J., D. P. Owens, P. F. Schmoeker and E. R. Meyer. Morphine induced supersensitivity to the effects of naloxone on lutenizing-hormone secretion in the male rat. J Pharmacol E~p Ther 225: 35-41, 1983.
9. Cicero, T. J.. D. P. Owens, K. S. Newman, P. F. Schmoeker and E. R. Meyer. Exogenous and endogenous opioid-induced enhancements of naloxone's effect on serum luteinizing hor- mone levels in the male rat. Eur J Pharmaeol 105: 273-284, 1984.
10. Cone, R. 1. and A. Goldstein. A specific radioimmunoassay for the opioid peptide dynorphin-B in neural tissues. Neuropeptides 3: 97-106, 1982.
11. Dandekar, S. and S. L. Sabol. Cell free translation and partial characterization of mRNA coding for enkephalin precursor protein. Proc Natl Aead Sci USA 79: 1017-1021, 1982.
12. Forman, L. J., W. E. Sonntag, V. W. Hylka and J. Meites. lmmunoreactive /3-endorphin in the plasma, pituitary and hypothalamus of young female rats on the day of estrous in intact and chronically castrated old constant estrous female rats. Lt:Jb Sei 33: 993-999, 1983.
13. Gabriel, S. M.. J. W. Simpkins and S. P. Kalra. Modulation of endogenous opioid influence on leuteinizing hormone secretion by progesterone and estrogen. Endocrinology 113:1806-1811, 1983.
14. Ghazarossian, V. E., C. Chavkin and A. Goldstein. A specific radioimmunoassay for the novel opioid peptide dynorphin. Li/b Sci 27: 75-86, 1980.
15. Glowinski, J and L. L. Iversen. Regional studies of catechola- mines in the rat brain-l. The disposition of [3H]-norepinephrine, [3H]-dopamine and [3H]-DOPA in various regions of the brain. J Neuroehem 13: 655-669, 1966.
16. Goldberg, R., N. Conforti and 1. M. Spitz. The effect of methionine enkephalin on prolactin and luteinizing hormone levels in intact and castrated rats. Horm Metab Res 14: 89-92, 1982.
17. Grossman, A., P. J. A. Moult, R. C. Gaillard, G. Delitala, W. D. Toff, L. H. Rees and G. M. Besser. The opioid control of LH and FSH release: Effects of a met-enkephalin analogue and naloxone. Clin Endoerinol 14: 41-47, 1981.
18. Guillemin, R., T. Vargo, J. Rossier, S. Minick, N. Ling, C. Rivier, W. Vale and F. E. Bloom. /3-endorphin and adrenocor- ticotropin are secreted concimitantly by the pituitary gland. Sci- em'e 197: 1367-1369, 1977.
19. Hollt, V., O. A. Muller and R. Fahlbusch./3-endorphin in human plasma: basal and pathologically elevated levels. Life Sci 25: 37-44, 1979.
20. Hong, J. S., K. Yoshikawa and C. A. Lamartiniere. Sex-related difference in the rat pituitary (met5)-enkephalin level-altered by gonadectomy. Brain Res 251: 380-383, 1982.
21. Hcilse, G. K. and G. J. Coleman. Effect of stress on tissue and plasma levels of immunoreactive/3-endorphin in ovariectomized rats primed with estrogen and progesterone. J Endoerinol 100: 277-280, 1984.
22. H~ilse, G. K., G. J. Coleman, D. L. Copolov and J. A. Cle- ments. Relationship between endogenous opioids and estrous cycle in the rat. J Endocrinol 100: 271-275, 1984.
23. leiri, T., H. T. Chen, G. A. Campbell and J. Meites. Effects of naloxone and morphine on the proestrous surge of prolactin and gonadotropins in the rat. Endocrinology 106: 1568-1570, 1980.
24. Kalra, S. P. Neural loci involved in naloxone induced lutenizing hormone release: Effects of a norepinephrine synthesis in- hibitor. Endocrinology 109: 19805-1810, 1981.
25. Kumar, M. S. A., L. Chen and T. F. Muther. Changes in the pituitary and hypothalamic content of met-enkephalin during the estrous cycle of rats. Ltfe Sei 25: 1687-1696, 1979.
26. Lowry, O. H., N. J. Rosenbrough, A. L. Farr and R. J. Randall. Protein measurement with Folin-phenol reagent. J Biol Chem 193: 265-275, 1951.
27. Meites, J., J. F. Bruni, D. A. VanVugt and A. F. Smith. Rela- tionship of endogenous opioid peptides and morphine to neuroendocrine function. L~f~, S('i 24: 1325-1336, 1979.
28. Meites, J. and K. Briski. Relation of endogenous opiates to secretion of gonadotropins. In. Hormonal Cantrol ~f" the hypothalamo-Pituitary-Gonadal Axis, edited by K. W. McKerns and Z. Naor. New York: Plenum Press, 1984, pp. 50%519.
29. Miller, R. J., K. J. Chang, B. Cooper and P. Cuatrecasas. Radioimmunoassay and characterization of enkephalins in rat tissue. J Bh~l Chem 253: 531-538, 1978.
30. Mittler, J. C., P. B. May and N. H. Ertel. Leucine-enkephalin modification of pituitary response to LH-releasing hormone. Neuroendoerino/Lett 5: 193-200, 1983.
31. Motta, M. and L. Martini. Effect of opioid peptides on gonado- tropin secretion. Aeta Endocrinol 99: 321-325, 1982.
32. Mueller, G. P. Attenuated pituitary/3-endorphin release in es- trogen treated rats. Proe Sot" Exp Biol Med 165: 75-81, 1980.
33. Quigley, M. E. and S. S. C. Yen. The role of endogenous opiates on LH-secretion during the menstrual cycle. J Clin Endocrinol Metub 51: 17%181. 1980.
34. Reid, R. L., J. D. Hoff, S. S. C. Yen and C. H. Li. Effects of exogenous/3-h-endorphin on pituitary hormone secretion and its disappearance rate in normal human subjects. J Clin Endoerinol Metab 52: 117%1184, 1981.
35. Rossier, J. Opioid peptides have found their roots. Nature 298: 221-222, 1982.
36. Rotsztejn, W. H.. S. V. Drouva, E. Patton and C. Kordon. Met-enkephalin inhibits in vitro dopamine-inducedoLHRH re- lease from mediobasal hypothalamus of male rats. Nature 274: 281-282, 1978.
37. Tejwani. G. A. Pharmacologic and other effects of opiate agonists and antagonists. Recent Adv C/in Ther 1: 193-216, 1981.
38. Tejwani, G. A., K. K. Vaswani, J. C. Barbacci, C. W. Richard and J. R. Bianchine. Effect of oral contraceptives on the rat brain and pituitary/3-endorphin. Life Sci 33: Suppl 1, 51%522, 1983.
39. Tejwani, G. A., K. K. Vaswani and J. C. Barbacci. Differential effects of oral contraceptives on CNS levels of enkephalins, dynorphin and /3-endorphin in female rats. Fed Prae 43: Abstract 4009, 1984.
40. VanVugt, D. A., G. Bakst, I. Dyrenfurth and M. Ferin. Naloxone stimulation of luteinizing hormone secretion in the female monkey: Influence of endocrine and experimental con- ditions. Endocrinology 113: 1858-1864, 1983.
41. Vaswani, K. K., G. A. Tejwani and S. Mousa. Stress induced differential intake of various diets and water by rat: The role of opiate system. Lift ~ Sei 32: 1983-1996. 1983.
42. Vaswani, K. K. Role of endogenous opioid peptides in stress induced eating. Ph.D Thesis, The Ohio State University, 1984.
43. Vrbricky, K. W., J. S. Baumstark, I. C. Wells, T. W. Hilger, W. T. Kable and C. J. Elias. Evidence for the involvement of /3-endorphin in the human menstrual cycle. Fertil Steril 38: 701-704, 1982.
44. Wardlaw, S. L. and A. G. Frantz. Measurement of/3-endorphin in human plasma. J Clin Endoerinol Metab 48: 176-180, 1979.
45. Wardlow, S. L., L. Thoron and A. G. Frantz. Effects of sex steroids on brain/3-endorphin. Brain Res 245: 327-331, 1982.
46. Wehrenberg, W. B., S. L. Wardlaw, A. G. Frantz and M. Ferin. /3-endorphin in hypophyseal portal blood; Variations throughout the menstrual cycle. Endo¢'rinology 111: 87%881, 1982.
47. Weidenmann, E., T. Saito, J. A. Linfoot and C. H. Li. Specific radioimmunoassay of human j3-endorphin in unextracted plasma. J Clin Endocrinol Metab 49: 478-480, 1979.
48. Yoshikawa, K. and J. S. Hong. The e~kephalin system in the rat anterior pituitary: Regulation by gonadal steroid hormones and psychotrophic drugs. Endocrinology 113: 1218-1227, 1983.





























