Embed Size (px)
Citation preview

Bioresource Technology 102 (2011) 1649–1655
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Effect of nutrients on growth and lipid accumulation in the green algaeDunaliella tertiolecta
Meng Chen a, Haiying Tang a, Hongzhi Ma a, Thomas C. Holland b, K.Y. Simon Ng a, Steven O. Salley a,⇑a Department of Chemical Engineering and Materials Science, Wayne State University, 5050 Anthony Wayne Drive, Detroit, MI 48202, USAb Department of Immunology and Microbiology, School of Medicine, Wayne State University, 540 East Canfield Avenue, Detroit, MI 48201, USA
a r t i c l e i n f o
Article history:Received 29 May 2010Received in revised form 9 September 2010Accepted 15 September 2010Available online 13 October 2010
Keywords:Dunaliella tertiolectaNutrientsNitrateNitrogen starvationFatty acid composition
0960-8524/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.biortech.2010.09.062
⇑ Corresponding author. Tel.: +1 313 577 5216; faxE-mail address: [email protected] (S.O. Salley).
a b s t r a c t
Production of biofuel from algae is dependent on the microalgal biomass production rate and lipid con-tent. Both biomass production and lipid accumulation are limited by several factors, of which nutrientsplay a key role. In this research, the marine microalgae Dunaliella tertiolecta was used as a model organ-ism and a profile of its nutritional requirements was determined. Inorganic phosphate PO3�
4 and trace ele-ments: cobalt (Co2+), iron (Fe3+), molybdenum (Mo2+) and manganese (Mn2+) were identified as requiredfor algae optimum growth. Inorganic nitrogen in the form of nitrate NO�3 instead of ammonium (NH4
+)was required for maximal biomass production. Lipids accumulated under nitrogen starvation growthcondition and this was time-dependent. Results of this research can be applied to maximize productionof microalgal lipids in optimally designed photobioreactors.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction (2) application of stress conditions to large cultures to induce
Development of new means of biofuel production is expected tobecome increasingly important in coming years. Large-scaledomestic production of biofuels appears to be feasible, reducingdependence on imported petroleum and petroleum-derived fuelsand improving energy security. In addition, biofuels, being derivedby reduction of atmospheric CO2, have the potential to greatly re-duce net CO2 emissions. Biofuels may be derived from a variety ofsources, but microalgae have attracted particular interest as one ofthe most promising sources of biomass for biofuel production. Asaquatic organisms, microalgae can be produced on non-arableland, minimizing the impact of algal biomass production on agri-culture. Additionally, it has been estimated that biomass produc-tion per hectare may be 10 or more times higher for microalgaethan for conventional crops (Chisti, 2007a,b).
For many microalgae, excess solar energy captured by photo-synthesis is primarily stored as lipids, often with high levels of tria-cylglycerols (TAG). Biodiesel is produced by the transesterificationof TAG with methanol (or other alcohol) in the presence of anappropriate catalyst, with glycerol as a byproduct. However, mostmicroalgal species produce large amounts of TAG only under stressconditions, e.g., under nitrogen starvation. Since stress conditionsgenerally lead to low growth rates, economical production of algalbiodiesel will require optimization of two phases of microalgalgrowth: (1) rapid production of large amounts of microalgae and
ll rights reserved.
: +1 313 577 3810.
TAG synthesis. The utility of this strategy has been noted by others(Benamotz, 1995; Schenk et al., 2008).
A genus of unicellular green marine microalgae, Dunaliellatertiolecta (D. tertiolecta, UTEX# LB999), has been reported to havean oil yield of 36–42% (Tsukahara and Sawayama, 2005). In addi-tion, D. tertiolecta is simple to cultivate, does not clump on the sur-face and is highly salt tolerant, which might be useful in large scaleoutdoor cultivation (Elenkov et al., 1996). In this study, the com-plete profile of nutrients assimilated by D. teriolecta was quantifiedand the nutritional elements that limit its growth were determinedin order to ascertain conditions for maximal biomass production.The lipid content of cells cultivated under several different starva-tion conditions was also determined. Finally, the fatty acid compo-sition of cells cultivated under normal and nitrogen starvationcondition was measured.
The conclusions from this research are important for obtainingmaximal microalgal biomass production, for optimal photobiore-actor design, for the development of catalysts for conversion oflipids to biodiesel and for the determination of the pollutant for-mation chemistry of algae-derived biofuels.
2. Methods
2.1. Algal source and culture medium
Marine microalgae D. tertiolecta (UTEX#: LB999) was obtainedfrom the culture collection of the University of Texas, Austin, Texas,

Table 1Components and their concentrations in Erdschreiber’s medium.
Component Stock solution concentration Final concentration
Seawater 0.3 mMNa2EDTA�2H2O 2 mM 2.4 � 10�2 mMFeCl3�6H2O 3.6 � 10�1 mM 4.32 � 10�3 mMMnCl2�4H2O 2.1 � 10�1 mM 2.52 � 10�3 mMZnCl2 3.7 � 10�2 mM 4.44 � 10�4 mMCoCl2�6H2O 8.4 � 10�3 mM 1.008 � 10�4 mMNa2MoO4�2H2O 1.7 � 10�2 mM 2.04 � 10�4 mMVitamin B12 12 g/l 1.2 � 10�2 g/lNaNO3 0.7 M 2.3 mMNa2HPO4�7H2O 0.2 M 0.67 mMSoilwater 24.40% 1.20%
1650 M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655
USA. The cells were cultured in Erdschreiber’s medium (Table 1),consisting of 3 L of pasteurized artificial seawater (Kent Marine,Franklin, Wisconsin), 36 mL of P-IV metal solution (0.75 g/L ofNa2EDTA�2H2O (Sigma ED255), 0.097 g/L of FeCl3�6H2O (Sigma1513), 0.041 g/L of MnCl2�4H2O (Baker 2540), 0.005 g/L of ZnCl2
(Sigma Z-0152), 0.002 g/L of CoCl2�6H2O (Sigma C-3169), and0.004 g/L of Na2MoO4�2H2O (J.T. Baker 3764)), 10 mL of NaNO3
(Fisher BP360–500), 10 mL of Na2HPO4�7H2O (Sigma S-9390),150 mL of soil water (1 teaspoon of garden soil (Schultz enrichedgarden soil, St. Louis, Missouri) was added to 200 mL of deionized(DI) water). The soil water was pasteurized by a gradual increase intemperature to approximately 95 �C in 15 min and 98 �C for 3 h on2 consecutive days. Soil water was filtered through a 0.22 lm filter(Corning 431097, Corning, NY) before using.) and 3 mL of VitaminB12 (0.1 mM of Vitamin B12 (cyanocobalamin, (Sigma V-6629) in DIwater was added in 50 mM HEPES buffer (Sigma H-3375) and ad-justed the pH to 7.8)(Schreiber, 1927).
2.2. Growth conditions
2.2.1. Culture flasksAlgae cultures were set up in 650 mL culture flasks (4.45 �
10.16 � 14.38 cm, Greiner Bio-One Gmbh, Germany) illuminatedwith one 12 inch, 8 watt fluorescent lamp (Phillips), and suppliedwith a feed of air/CO2 (4%) mixture delivered at a total gas flow rateof 60 mL/min bubbled through an aquafizz 1 inch air stone (Petso-lutions, Beavercreek, OH). The temperature of the culture wasmaintained at 23 �C and pH was not controlled. During the cultiva-tion, cell density, temperature, and pH were recorded daily.
2.2.2. 96-well microplate culturesMicroalgal cultures were set up in 96-well microtiter plates
(Greiner bio-one, Monroe, NC) with 240 lL culture volume. Theplates were incubated at 32 �C in an incubator (NuAire, Plymouth,MN) with double 12 inch 8 watt fluorescent light with 5% CO2. The32 �C incubation temperature was chosen to maximize the cellgrowth rates.
2.3. Growth measurements
2.3.1. Optical density (OD)Optical density was used to monitor the algae growth. OD of
bulk cultures was measured at 680 nm with an Evolution 60 spec-trophotometer, (Thermo Scientific, Waltham, MA). Each samplewas measured twice and the average value was used. The OD ofalgae cultivated in 96-well plates was measured at 405 nm in a96-well microplate reader INFINITE 200 (Tecan, Seestrasse 103CH-8708 Männedorf, Switzerland). Microalgal cultures were mixedby shaking for 30 s before reading. The value of optical density wasquantified using Magellan software. For each sample, the averagevalue for eight wells was computed.
2.3.2. Biomass determinationAlgae were cultivated in flasks for biomass determination. Ten
milliliter samples were collected daily from the cultures. Opticaldensities were determined as described above. After centrifuga-tion, the supernatant was carefully removed by pipette.Wet algae biomass was determined using an analytical scale(AB304-S, Mettler Toledo, Switzerland).
2.4. Inorganic nitrogen measurement
Inorganic nitrogen was quantified in the two forms, nitrate andammonium, using a portable spectrophotometer (DR 2800, Hach,Loveland, Colorado). Briefly, 5 mL algae culture was collected everyday. The culture was centrifuged and the supernatant was trans-ferred to another clean tube for analysis. Nitrate was measuredusing reaction kit Nitrate TNTplus, HR (5–35 mg/L NO3-N) (TNT836, Hach, Loveland, Colorado). Ammonium was measured usingreaction kit Ammonia TNTplus, Low Range (1–12 mg/L NH3-N)(TNT 831, Hach, Loveland, Colorado).
2.5. Inductively coupled plasma (ICP)
The concentrations of inorganic phosphorus and trace metalelements were measured by inductively coupled plasma (ICP) Op-tima 2000 DV ICP-OES (N0800443, PerkinElmer, Waltham, Massa-chusetts). Briefly, 5 mL samples were collected daily from algaecultures cultivated in flasks. After centrifugation, the supernatantwas transferred to clean tubes and diluted with 10 volumes of5% nitric acid prior to IPC analysis.
2.6. Lipid analysis
2.6.1. 96-well microplate Nile Red fluorescence assayA stock solution of Nile Red (NR) (Sigma, 72485) was prepared
by adding 5 mg of NR to100 mL of acetone. The solution was keptin an amber colored bottle and stored in the dark at 4 �C. Algal cellcultures that were cultivated in 96-well plates under variousgrowth conditions were transferred to black 96-well plates withtransparent bottoms (Greiner bio-one, Monroe, NC). 5 lL of NileRed solution was added to each well and mixed, followed by20 min incubation in the dark. Finally, cellular neutral lipids werequantified using a 96-well microplate reader (Tecan infinite 200,Seestrasse 103 CH-8708 Männedorf, Switzerland) with an excita-tion wavelength of 485 nm and an emission wavelength of595 nm. The lipid standard triolein (Sigma T9275) was used to pro-duce a standard curve.
2.6.2. Lipid extractionAlgal cultures were harvested by centrifugation (Eppendorf cen-
trifuge 5804R, Germany) at 5000 rpm for 5 min. Cell pellets werefrozen at �20 �C for at least 6 h, and then 10 g of wet algae was dis-rupted with glass beads (Sigma, St. Louis) in a vortex mixer (Scien-tific Industries Inc., Bohemia, NY) for 10 min. The lipid wasextracted with 10 mL of chloroform: methanol (2:1), and the ex-tract was centrifuged at 5000 rpm (Eppendorf centrifuge 5804R,Germany) for 10 min for phase separation. The upper layer (meth-anol–water layer) was removed and the chloroform layer includinglipid was collected. Chloroform was evaporated from extracts toyield the resultant algae oil, which was analyzed by GC-FID andGC–MS.
2.6.3. Fatty acid determinationThe fatty acid profile of the oil sample was determined by con-
verting the fatty acids in the oil to fatty acid methyl esters (FAMEs).The FAME composition was determined using a PerkinElmer Clarus500 GC–mass spectrometer (GC–MS) with a split automatic
M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655 1651
injector and an Rtx-WAX (Restek, Bellefonte, PA) column (length:60 m; ID: 0.25 mm, coating: 0.25 lm). Details of the procedure
OD 680
Bio
mas
s (m
g/m
l)
0
1
2
3
4
5
6
7
8
9
0 0.5 1 1.5 2
C
A
B
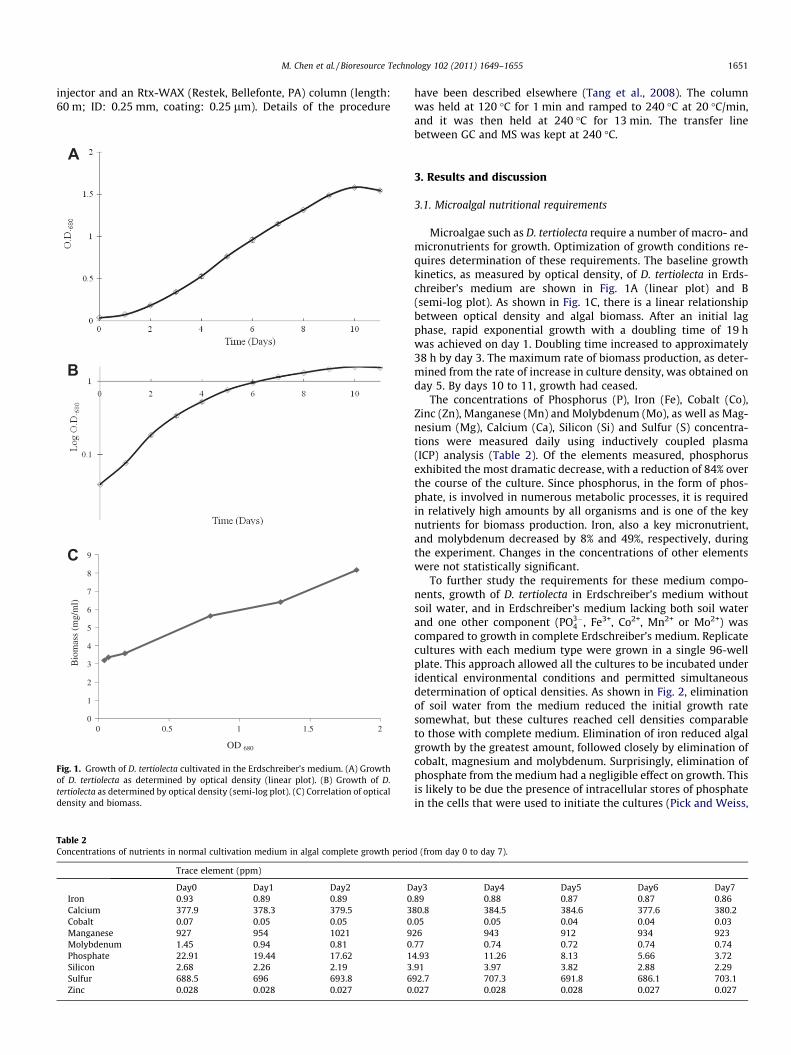
Fig. 1. Growth of D. tertiolecta cultivated in the Erdschreiber’s medium. (A) Growthof D. tertiolecta as determined by optical density (linear plot). (B) Growth of D.tertiolecta as determined by optical density (semi-log plot). (C) Correlation of opticaldensity and biomass.
Table 2Concentrations of nutrients in normal cultivation medium in algal complete growth perio
Trace element (ppm)
Day0 Day1 Day2 DIron 0.93 0.89 0.89 0Calcium 377.9 378.3 379.5 3Cobalt 0.07 0.05 0.05 0Manganese 927 954 1021 9Molybdenum 1.45 0.94 0.81 0Phosphate 22.91 19.44 17.62 1Silicon 2.68 2.26 2.19 3Sulfur 688.5 696 693.8 6Zinc 0.028 0.028 0.027 0
have been described elsewhere (Tang et al., 2008). The columnwas held at 120 �C for 1 min and ramped to 240 �C at 20 �C/min,and it was then held at 240 �C for 13 min. The transfer linebetween GC and MS was kept at 240 �C.
3. Results and discussion
3.1. Microalgal nutritional requirements
Microalgae such as D. tertiolecta require a number of macro- andmicronutrients for growth. Optimization of growth conditions re-quires determination of these requirements. The baseline growthkinetics, as measured by optical density, of D. tertiolecta in Erds-chreiber’s medium are shown in Fig. 1A (linear plot) and B(semi-log plot). As shown in Fig. 1C, there is a linear relationshipbetween optical density and algal biomass. After an initial lagphase, rapid exponential growth with a doubling time of 19 hwas achieved on day 1. Doubling time increased to approximately38 h by day 3. The maximum rate of biomass production, as deter-mined from the rate of increase in culture density, was obtained onday 5. By days 10 to 11, growth had ceased.
The concentrations of Phosphorus (P), Iron (Fe), Cobalt (Co),Zinc (Zn), Manganese (Mn) and Molybdenum (Mo), as well as Mag-nesium (Mg), Calcium (Ca), Silicon (Si) and Sulfur (S) concentra-tions were measured daily using inductively coupled plasma(ICP) analysis (Table 2). Of the elements measured, phosphorusexhibited the most dramatic decrease, with a reduction of 84% overthe course of the culture. Since phosphorus, in the form of phos-phate, is involved in numerous metabolic processes, it is requiredin relatively high amounts by all organisms and is one of the keynutrients for biomass production. Iron, also a key micronutrient,and molybdenum decreased by 8% and 49%, respectively, duringthe experiment. Changes in the concentrations of other elementswere not statistically significant.
To further study the requirements for these medium compo-nents, growth of D. tertiolecta in Erdschreiber’s medium withoutsoil water, and in Erdschreiber’s medium lacking both soil waterand one other component (PO3�
4 , Fe3+, Co2+, Mn2+ or Mo2+) wascompared to growth in complete Erdschreiber’s medium. Replicatecultures with each medium type were grown in a single 96-wellplate. This approach allowed all the cultures to be incubated underidentical environmental conditions and permitted simultaneousdetermination of optical densities. As shown in Fig. 2, eliminationof soil water from the medium reduced the initial growth ratesomewhat, but these cultures reached cell densities comparableto those with complete medium. Elimination of iron reduced algalgrowth by the greatest amount, followed closely by elimination ofcobalt, magnesium and molybdenum. Surprisingly, elimination ofphosphate from the medium had a negligible effect on growth. Thisis likely to be due the presence of intracellular stores of phosphatein the cells that were used to initiate the cultures (Pick and Weiss,
d (from day 0 to day 7).
ay3 Day4 Day5 Day6 Day7.89 0.88 0.87 0.87 0.8680.8 384.5 384.6 377.6 380.2.05 0.05 0.04 0.04 0.0326 943 912 934 923.77 0.74 0.72 0.74 0.744.93 11.26 8.13 5.66 3.72.91 3.97 3.82 2.88 2.2992.7 707.3 691.8 686.1 703.1.027 0.028 0.028 0.027 0.027
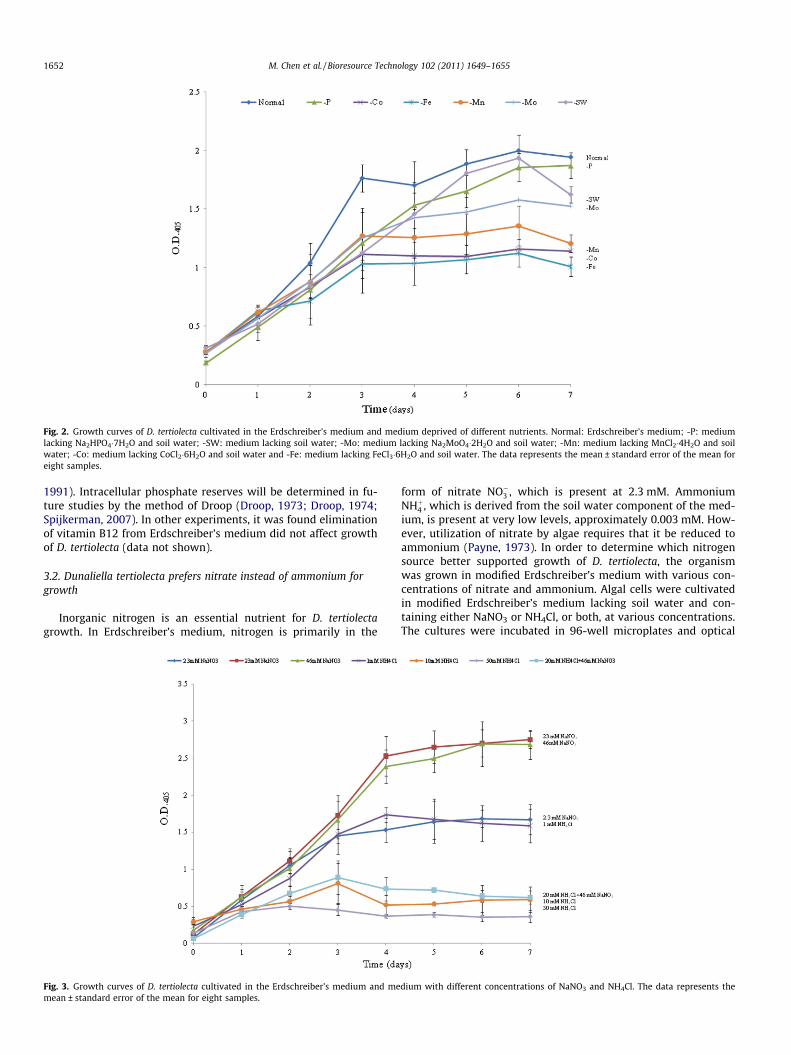
Fig. 2. Growth curves of D. tertiolecta cultivated in the Erdschreiber’s medium and medium deprived of different nutrients. Normal: Erdschreiber’s medium; -P: mediumlacking Na2HPO4�7H2O and soil water; -SW: medium lacking soil water; -Mo: medium lacking Na2MoO4�2H2O and soil water; -Mn: medium lacking MnCl2�4H2O and soilwater; -Co: medium lacking CoCl2�6H2O and soil water and -Fe: medium lacking FeCl3�6H2O and soil water. The data represents the mean ± standard error of the mean foreight samples.
1652 M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655
1991). Intracellular phosphate reserves will be determined in fu-ture studies by the method of Droop (Droop, 1973; Droop, 1974;Spijkerman, 2007). In other experiments, it was found eliminationof vitamin B12 from Erdschreiber’s medium did not affect growthof D. tertiolecta (data not shown).
3.2. Dunaliella tertiolecta prefers nitrate instead of ammonium forgrowth
Inorganic nitrogen is an essential nutrient for D. tertiolectagrowth. In Erdschreiber’s medium, nitrogen is primarily in the
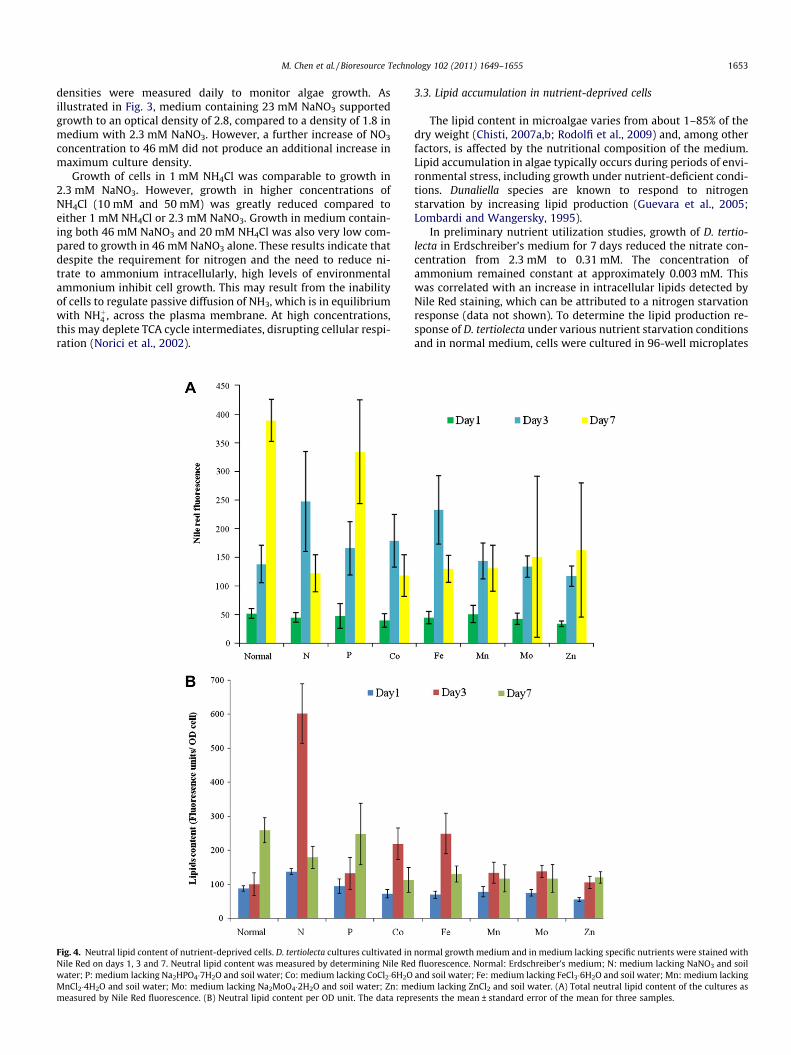
Fig. 3. Growth curves of D. tertiolecta cultivated in the Erdschreiber’s medium and memean ± standard error of the mean for eight samples.
form of nitrate NO�3 , which is present at 2.3 mM. AmmoniumNHþ4 , which is derived from the soil water component of the med-ium, is present at very low levels, approximately 0.003 mM. How-ever, utilization of nitrate by algae requires that it be reduced toammonium (Payne, 1973). In order to determine which nitrogensource better supported growth of D. tertiolecta, the organismwas grown in modified Erdschreiber’s medium with various con-centrations of nitrate and ammonium. Algal cells were cultivatedin modified Erdschreiber’s medium lacking soil water and con-taining either NaNO3 or NH4Cl, or both, at various concentrations.The cultures were incubated in 96-well microplates and optical
dium with different concentrations of NaNO3 and NH4Cl. The data represents the
M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655 1653
densities were measured daily to monitor algae growth. Asillustrated in Fig. 3, medium containing 23 mM NaNO3 supportedgrowth to an optical density of 2.8, compared to a density of 1.8 inmedium with 2.3 mM NaNO3. However, a further increase of NO3
concentration to 46 mM did not produce an additional increase inmaximum culture density.
Growth of cells in 1 mM NH4Cl was comparable to growth in2.3 mM NaNO3. However, growth in higher concentrations ofNH4Cl (10 mM and 50 mM) was greatly reduced compared toeither 1 mM NH4Cl or 2.3 mM NaNO3. Growth in medium contain-ing both 46 mM NaNO3 and 20 mM NH4Cl was also very low com-pared to growth in 46 mM NaNO3 alone. These results indicate thatdespite the requirement for nitrogen and the need to reduce ni-trate to ammonium intracellularly, high levels of environmentalammonium inhibit cell growth. This may result from the inabilityof cells to regulate passive diffusion of NH3, which is in equilibriumwith NHþ4 , across the plasma membrane. At high concentrations,this may deplete TCA cycle intermediates, disrupting cellular respi-ration (Norici et al., 2002).
Fig. 4. Neutral lipid content of nutrient-deprived cells. D. tertiolecta cultures cultivated inNile Red on days 1, 3 and 7. Neutral lipid content was measured by determining Nile Redwater; P: medium lacking Na2HPO4�7H2O and soil water; Co: medium lacking CoCl2�6H2OMnCl2�4H2O and soil water; Mo: medium lacking Na2MoO4�2H2O and soil water; Zn: memeasured by Nile Red fluorescence. (B) Neutral lipid content per OD unit. The data repr
3.3. Lipid accumulation in nutrient-deprived cells
The lipid content in microalgae varies from about 1–85% of thedry weight (Chisti, 2007a,b; Rodolfi et al., 2009) and, among otherfactors, is affected by the nutritional composition of the medium.Lipid accumulation in algae typically occurs during periods of envi-ronmental stress, including growth under nutrient-deficient condi-tions. Dunaliella species are known to respond to nitrogenstarvation by increasing lipid production (Guevara et al., 2005;Lombardi and Wangersky, 1995).
In preliminary nutrient utilization studies, growth of D. tertio-lecta in Erdschreiber’s medium for 7 days reduced the nitrate con-centration from 2.3 mM to 0.31 mM. The concentration ofammonium remained constant at approximately 0.003 mM. Thiswas correlated with an increase in intracellular lipids detected byNile Red staining, which can be attributed to a nitrogen starvationresponse (data not shown). To determine the lipid production re-sponse of D. tertiolecta under various nutrient starvation conditionsand in normal medium, cells were cultured in 96-well microplates
normal growth medium and in medium lacking specific nutrients were stained withfluorescence. Normal: Erdschreiber’s medium; N: medium lacking NaNO3 and soil
and soil water; Fe: medium lacking FeCl3�6H2O and soil water; Mn: medium lackingdium lacking ZnCl2 and soil water. (A) Total neutral lipid content of the cultures asesents the mean ± standard error of the mean for three samples.
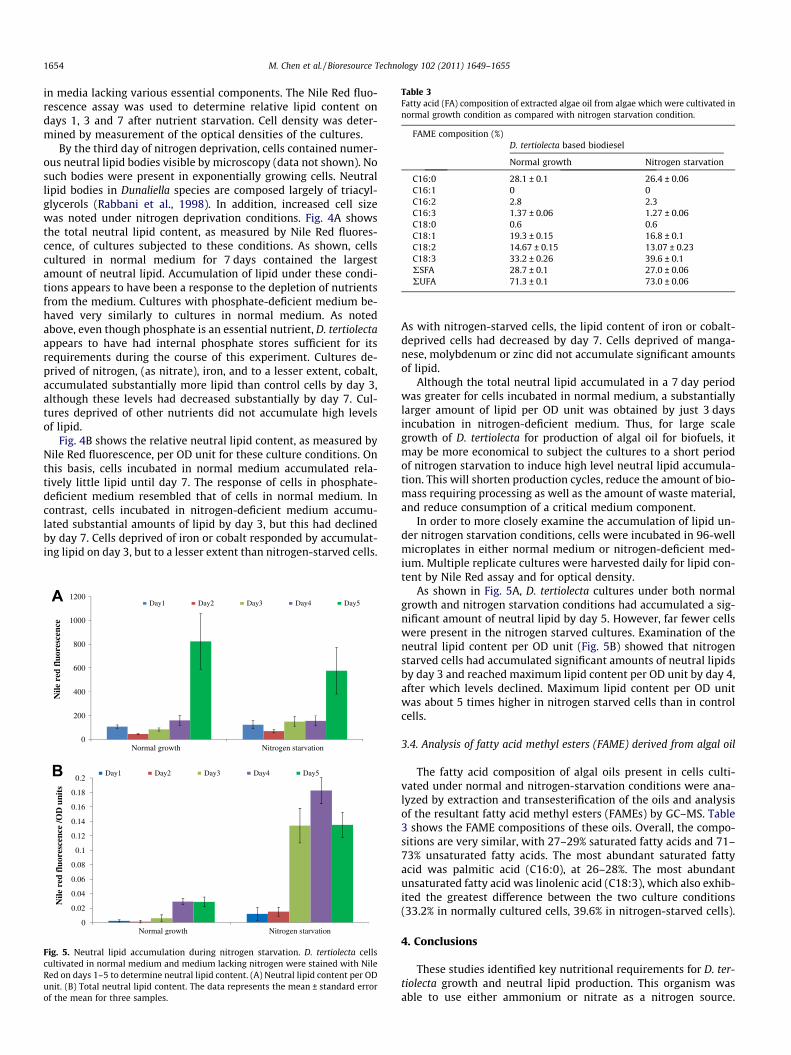
Table 3Fatty acid (FA) composition of extracted algae oil from algae which were cultivated innormal growth condition as compared with nitrogen starvation condition.
FAME composition (%)D. tertiolecta based biodiesel
Normal growth Nitrogen starvation
C16:0 28.1 ± 0.1 26.4 ± 0.06C16:1 0 0C16:2 2.8 2.3C16:3 1.37 ± 0.06 1.27 ± 0.06C18:0 0.6 0.6C18:1 19.3 ± 0.15 16.8 ± 0.1C18:2 14.67 ± 0.15 13.07 ± 0.23C18:3 33.2 ± 0.26 39.6 ± 0.1RSFA 28.7 ± 0.1 27.0 ± 0.06RUFA 71.3 ± 0.1 73.0 ± 0.06
1654 M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655
in media lacking various essential components. The Nile Red fluo-rescence assay was used to determine relative lipid content ondays 1, 3 and 7 after nutrient starvation. Cell density was deter-mined by measurement of the optical densities of the cultures.
By the third day of nitrogen deprivation, cells contained numer-ous neutral lipid bodies visible by microscopy (data not shown). Nosuch bodies were present in exponentially growing cells. Neutrallipid bodies in Dunaliella species are composed largely of triacyl-glycerols (Rabbani et al., 1998). In addition, increased cell sizewas noted under nitrogen deprivation conditions. Fig. 4A showsthe total neutral lipid content, as measured by Nile Red fluores-cence, of cultures subjected to these conditions. As shown, cellscultured in normal medium for 7 days contained the largestamount of neutral lipid. Accumulation of lipid under these condi-tions appears to have been a response to the depletion of nutrientsfrom the medium. Cultures with phosphate-deficient medium be-haved very similarly to cultures in normal medium. As notedabove, even though phosphate is an essential nutrient, D. tertiolectaappears to have had internal phosphate stores sufficient for itsrequirements during the course of this experiment. Cultures de-prived of nitrogen, (as nitrate), iron, and to a lesser extent, cobalt,accumulated substantially more lipid than control cells by day 3,although these levels had decreased substantially by day 7. Cul-tures deprived of other nutrients did not accumulate high levelsof lipid.
Fig. 4B shows the relative neutral lipid content, as measured byNile Red fluorescence, per OD unit for these culture conditions. Onthis basis, cells incubated in normal medium accumulated rela-tively little lipid until day 7. The response of cells in phosphate-deficient medium resembled that of cells in normal medium. Incontrast, cells incubated in nitrogen-deficient medium accumu-lated substantial amounts of lipid by day 3, but this had declinedby day 7. Cells deprived of iron or cobalt responded by accumulat-ing lipid on day 3, but to a lesser extent than nitrogen-starved cells.
0
200
400
600
800
1000
1200
Normal growth Nitrogen starvation
Normal growth Nitrogen starvation
Day1 Day2 Day3 Day4 Day5
Nile
red
flu
ores
cenc
e
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2Day1 Day2 Day3 Day4 Day5
Nile
red
flu
ores
cenc
e /O
D u
nits
A
B
Fig. 5. Neutral lipid accumulation during nitrogen starvation. D. tertiolecta cellscultivated in normal medium and medium lacking nitrogen were stained with NileRed on days 1–5 to determine neutral lipid content. (A) Neutral lipid content per ODunit. (B) Total neutral lipid content. The data represents the mean ± standard errorof the mean for three samples.
As with nitrogen-starved cells, the lipid content of iron or cobalt-deprived cells had decreased by day 7. Cells deprived of manga-nese, molybdenum or zinc did not accumulate significant amountsof lipid.
Although the total neutral lipid accumulated in a 7 day periodwas greater for cells incubated in normal medium, a substantiallylarger amount of lipid per OD unit was obtained by just 3 daysincubation in nitrogen-deficient medium. Thus, for large scalegrowth of D. tertiolecta for production of algal oil for biofuels, itmay be more economical to subject the cultures to a short periodof nitrogen starvation to induce high level neutral lipid accumula-tion. This will shorten production cycles, reduce the amount of bio-mass requiring processing as well as the amount of waste material,and reduce consumption of a critical medium component.
In order to more closely examine the accumulation of lipid un-der nitrogen starvation conditions, cells were incubated in 96-wellmicroplates in either normal medium or nitrogen-deficient med-ium. Multiple replicate cultures were harvested daily for lipid con-tent by Nile Red assay and for optical density.
As shown in Fig. 5A, D. tertiolecta cultures under both normalgrowth and nitrogen starvation conditions had accumulated a sig-nificant amount of neutral lipid by day 5. However, far fewer cellswere present in the nitrogen starved cultures. Examination of theneutral lipid content per OD unit (Fig. 5B) showed that nitrogenstarved cells had accumulated significant amounts of neutral lipidsby day 3 and reached maximum lipid content per OD unit by day 4,after which levels declined. Maximum lipid content per OD unitwas about 5 times higher in nitrogen starved cells than in controlcells.
3.4. Analysis of fatty acid methyl esters (FAME) derived from algal oil
The fatty acid composition of algal oils present in cells culti-vated under normal and nitrogen-starvation conditions were ana-lyzed by extraction and transesterification of the oils and analysisof the resultant fatty acid methyl esters (FAMEs) by GC–MS. Table3 shows the FAME compositions of these oils. Overall, the compo-sitions are very similar, with 27–29% saturated fatty acids and 71–73% unsaturated fatty acids. The most abundant saturated fattyacid was palmitic acid (C16:0), at 26–28%. The most abundantunsaturated fatty acid was linolenic acid (C18:3), which also exhib-ited the greatest difference between the two culture conditions(33.2% in normally cultured cells, 39.6% in nitrogen-starved cells).
4. Conclusions
These studies identified key nutritional requirements for D. ter-tiolecta growth and neutral lipid production. This organism wasable to use either ammonium or nitrate as a nitrogen source.

M. Chen et al. / Bioresource Technology 102 (2011) 1649–1655 1655
However, only moderate concentrations of ammonium were toler-ated and high levels inhibited growth. In contrast, high concentra-tions of nitrate increased maximum cell densities. Deprivation ofcells of nitrogen, or to a lesser extent, iron or cobalt, caused rapidaccumulation of neutral lipids. Although phosphate is essentialfor growth, phosphate deprivation had little effect in these studies,apparently due to intracellular phosphate stores. Further studieson the phosphate requirements of D. tertiolecta are in progress.
Acknowledgement
Financial support from the Department of Energy (Grant DE-FG36-05GO85005) for this research is gratefully acknowledged.
References
Benamotz, A., 1995. New mode of Dunaliella biotechnology-2-phase growth forbeta-carotene production. Journal of Applied Phycology 7, 65–68.
Chisti, Y., 2007a. Biodiesel from microalgae. Biotechnology Advances 25, 294–306.Chisti, Y., 2007b. Biodiesel from microalgae beats bioethanol. Trends in
Biotechnology 26, 6.Droop, M.R., 1973. Some thoughts on nutrient limitation in algae. Journal of
Phycology 9, 9.Droop, M.R., 1974. The nutrient status of algal cells in continuous culture. Journal of
the Marine Biology Association 55, 31.Elenkov, I., Stefanov, K., DimitrovaKonaklieva, S., Popov, S., 1996. Effect of salinity on
lipid composition of Cladophora vagabunda. Phytochemistry 42, 39–44.Guevara, M., Lodeiros, C., Gómez, O., Lemus, N., Núñez, P., Romero, L., Vásquez, A.,
Rosales, N., 2005. Carotenogénesis de cinco cepas del alga Dunaliella sp.
(Chlorophyceae) aisladas de lagunas hipersalinas de Venezuela. Revista DeBiologia Tropical 55, 7.
Lombardi, A.T., Wangersky, P.J., 1995. Particulate lipid class composition of 3marine phytoplankters chaetoceros-gracilis, isochrysis-galbana(Tahiti) andDunaliella tertiolecta grown in batch culture. Hydrobiologia 306, 1–6.
Norici, A., Dalsass, A., Giordano, M., 2002. Role of phosphoenolpyruvate carboxylasein anaplerosis in the green microalga Dunaliella salina cultured under differentnitrogen regimes. Physiologia Plantarum 116, 186–191.
Payne, W., 1973. Reduction of nitrogenous oxides by microorganisms. Bacteriol Rev4.
Pick, U., Weiss, M., 1991. Polyphosphate hydrolysis within acidic vacuoles inresponse to amine-induced alkaline stress in the halotolerant alga Dunaliellasaline. Plant Physiology 97, 1234–1240.
Rabbani, S., Beyer, P., Von Lintig, J., Hugueney, P., Kleinig, H., 1998. Induced beta-carotene synthesis driven by triacylglycerol deposition in the unicellular algaDunaliella bardawil. Plant Physiology 116, 1239–1248.
Rodolfi, L., Zittelli, G.C., Bassi, N., Padovani, G., Biondi, N., Bonini, G., Tredici, M.R.,2009. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis andOutdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnology andbioengineering 102, 100–112.
Schenk, P.M., Thomas-Hall, S.R., Stephens, E., Marx, U.C., Mussgnug, J.H., Posten, C.,Kruse, O., Hankamer, B., 2008. Second Generation Biofuels: High-EfficiencyMicroalgae for Biodiesel Production. Bioenergy research 1, 24.
Schreiber, E., 1927. Die Reinkultur von marinen Phytoplankton und derenBedeutung für die Erforschung der Produktionsfähigkeit des Meerwassers.Wissenschaften Meeresuntersuch N.F. 10, 1–34.
Spijkerman, E., 2007. Phosphorus acquisition by Chlamydomonas acidophila underautotrophic and osmo-mixotrophic growth conditions. Journal of ExperimentalBotany 58, 4195–4202.
Tang, H.Y., Salley, S.O., Ng, K.Y.S., 2008. Fuel properties and precipitate formation atlow temperature in soy-, cottonseed-, and poultry fat-based biodiesel blends.Fuel 87, 3006–3017.
Tsukahara, K., Sawayama, S., 2005. Liquid fuel production using microalgae. Journalof the Japan Petroleum Institute 48, 251–259.