Embed Size (px)
Citation preview

ORIGINAL RESEARCH ARTICLE
Effect of Estradiol and SelectedAntiestrogens on Pro- and AntioxidantPathways in Mammalian UterusSudhir Jain,* Deeksha Saxena,* Pradeep G. Kumar,* Samuel S. Koide,† and Malini Laloraya*
In this study, we examined the effect of 17b-estradiol andselected antiestrogens on uterine NADPH-oxidase activity,superoxide dismutase (SOD) activity, hydride (H z2), di-enyl radical and O2
z2-radical generation, and membranefluidity. NADPH oxidase activity was positively modu-lated in estradiol-treated animals and negatively regulatedin animals that received injections of AF-45, RU-39411,tamoxifen, or ICI-182780. The SOD activity was markedlyreduced in estradiol-treated animals when compared withthe control animals. A positive modulation of SOD activ-ity was observed upon treatment with AF45, RU39411,tamoxifen, and ICI 182780, though the potency variedamong the individual test compounds. We observed de-tectable H z2-radical generation as evidenced from MNP-H z adduct formation in the uterine cell preparations fromuntreated control animals. Estradiol produced a tremen-dous augmentation in the superoxide radical profiles inuterine cell preparations compared to the control levels.All the other compounds that were tested significantlylowered the superoxide levels in the test set-up. AF-45,RU-39411, tamoxifen, and ICI-182780 induced varyingorders of suppression of H z2-radical generation in the testsubjects. There was a significant enhancement in mem-brane fluidity, hydride radical levels, and dienyl radicalgeneration in the estradiol-treated group. All the antiestro-gens did not exhibit a similar action on these parameters.RU-39411 exhibited antiestrogen-like activity in modulat-ing hydride levels and membrane fluidity, whereas itstimulated dienyl radical generation. Thus our testsshowed that the selected antiestrogens failed to showestrogen-like activity in these assays. It appears that estra-diol exerts feedback control over pro- and antioxidantpathways and that markers of oxidative status could beused as a measure to evaluate the antiestrogenic activity ofestradiol agonists/antagonists. CONTRACEPTION 1999;60:111–118 © 1999 Elsevier Science Inc. All rights reserved.
KEY WORDS: estradiol, antiestrogens, free radicals, NADPHoxidase, SOD, hydride radical
Introduction
Estrogens regulate growth and cell division intarget tissues such as the uterus. The process ofregulation is thought to involve the initial bind-
ing to high-affinity nuclear receptors, which, in turn,bind to specific DNA sequences that regulate genetranscription.1 The actual series of events that medi-ate the mitogenic effect of estrogen in the uterusremains largely unknown.2 One theory suggests thatestrogens act directly as a mitogen on target cells.Another hypothesis is that estrogens promote uterinegrowth indirectly via a local (paracrine, autocrine) orsystemic (endocrine) regulation of levels of certaingrowth factors, their receptors, or both.2 The rodentuterus has been widely used as a model system tostudy interactions between estrogens and their intra-cellular receptors and the influence of estrogens ontarget organ growth.3,4 In some estrogen target organs,17b-estradiol induces the de novo synthesis of spe-cific proteins that can be used as specific molecularmarkers of hormone action.5–7 Estrogens have beenobserved to increase the overall rate of protein syn-thesis in the uterus,8 and many laboratories havesearched for molecular markers of estrogen action inthe uterus. Several uterine proteins have been sug-gested as markers of estrogen action: peroxidase,9
hydrolase,10 plasminogen activator,11 glucose-6-phos-phate dehydrogenase,12 estrogen receptor,13 proges-terone receptor,14 and induced protein (IP).15–17
There are important practical consequences to fur-ther investigations of estrogen action related to therole of these hormones, because blockade of estrogenaction appears to be a key to the control of certainmalignancies and to the control of fertility. Directlyrelated to this is the controversy surrounding the useof antiestrogens like tamoxifen.18,19 Tamoxifen isrepresentative of the most studied class of antiestro-gens; typically, such agents are nonsteroidal in struc-ture and manifest complex pharmacology encompas-
*School of Life Sciences, Devi Ahilya University, Indore, India; and †ThePopulation Council, Center for Biomedical Research, New York, New York
Name and address for correspondence: Dr. Malini Laloraya, School of LifeSciences, Devi Ahilya University, Vigyan Bhawan, Khandwa Road, Indore 452001, M.P, India; e-mail: [email protected]
Submitted for publication June 23, 1999Revised July 20, 1999Accepted for publication September 8, 1999
© 1999 Elsevier Science Inc. All rights reserved. ISSN 0010-7824/99/$20.00655 Avenue of the Americas, New York, NY 10010 PII S0010-7824(99)00067-0

sing variable tissue-, cell-, and gene-specific effectsfrom estrogen-like actions to complete blockade ofestrogen action characteristic of pure antagonism.20,21
The advent of molecules that bind ER with a highaffinity without activating receptor signaling pro-vided for the first time the opportunity to study in anunequivocal manner the consequences of a full block-ade of estrogen action.22 ICI 182780 is a pure anties-trogen that blocks estrogen action in vitro and in vivoin a concentration (dose)-dependent and completemanner, whereas in the absence of endogenous estro-gens this agent alone produces no estrogen-like ef-fects.23,24 Similarly, the hormonal properties of anor-drin, an antiestrogen that has been found to have apostcoital antifertility effect in women and somelaboratory animals,25–27 or its 1000-times more po-tent metabolite anordiol (AF 45) in terms of anties-trogenic activity are not yet fully understood.RU39411, an antiestrogen that had long been thoughtto possess only antagonistic properties, was recentlyshown to possess some weak, estrogen agonist effectson the oviduct.28
In some of our recent reports, we demonstratedearly pregnancy-associated elevation in free radicalgeneration and the loss of superoxide dismutase(SOD) activity in mouse uteri,29 the resulting lipid-kinking,30 and the increase in membrane fluidity31 asimportant events in mediating blastocyst implanta-tion. Because preimplantation embryo developmentand the embryo implantation in the uterus are largelycontrolled by ovarian steroids, an examination of theeffects of selected estrogenic/antiestrogenic drugs onthe above biophysical and biochemical aspects ofmammalian uterus during early pregnancy couldyield more molecular insight into the mechanism ofaction of estrogens and could define the estrogenic orantiestrogenic nature of the drugs in question usingnovel parameters. This article reports our observa-tions from some of the experiments designed in thisline.
Materials and MethodsReagentsTrizma HCl, Trizma base, diethyl dithiocarbamicacid (DDC), diethylene triamine penta acetic acid(DTPA), and NADPH and N-t-butyl-a-phenyl nitrone(PBN) were purchased from Sigma Chemical (St.Louis, MO). 16-Doxyl stearate (16-NS) and 2-methyl-2-nitrosopropane (MNP) were obtained from AldrichChemical (Milwaukee, WI). The following drugs wereused: anordiol (AF-45, 2a,17a-diethynyl-A-nor-5a-an-drostane-2b,17b-diol) (Shanghai Institute of MateriaMedica, China); tamoxifen ([Z]-2-[4-{1,2-diphenyl-1-butenyl}-phenoxyl]-N,N-dimethylethaneamine, Astra-
Zeneca Pharmaceuticals, Chesire, UK); ICI 182780{7a-(9-4,4,5,5,5-pentafluoropenyl sulphenyl)nonyl-estra-1,3,4(10-triene-3,17b-diol)} from Astra-Zeneca Phar-maceuticals, Chesire, England; RU39411 (3,17c-dihydroxy-11b-(4-dimethylaminoethoxyphenyl)estra-1,3,5-triene) Roussel-Uclaf, France, and 17b-estradiol(1,3,5[10]-estratriene-3,17b-diol, Sigma Chemical, St.Louis, MO). All other reagents were of analyticalgrade, procured from local suppliers.
AnimalsImmature female mice (Mus musculus, Swiss strain),aged 4–6 weeks, bred in our institute animal facilitywere used for all experiments. The experimentalanimals were housed at controlled temperature (27°C6 1°C) and light (14 h light, 10 h dark) regimens atthe facility.
Animals were divided into the following treatmentgroups: group I (vehicle alone, control), group II (17b-estradiol ), group III (tamoxifen), group IV (RU-39411),group V (AF-45), and group VI (ICI-182780). GroupsII–V were divided into three subgroups each for test-ing the dose response of the drugs under investiga-tion. Thus, group II has three subgroups classified aslow dose (0.24 mg/50 mL/day), medium dose (0.60mg/50 mL/day), and high dose (1.50 mg/50 mL/day).AF-45, RU-39411, and ICI-182780 were all tested atthree doses: low dose (8 mg/50 mL/day), medium dose(20 mg/50 mL/day), and high dose (50 mg/50 mL/day).The drugs were delivered through subcutaneous in-jections at 9:00 AM for 3 consecutive days and theanimals were killed at 9:00 AM on day 4 by CO2-asphyxiation. The uteri were excised and cleared ofadhering fat tissues and blood vessels, and were thenwashed in five changes of physiological saline.
Assay of superoxide dismutase SOD activitySOD was extracted as described in one of our earlierreports.29 SOD activity was assayed according to themethod of Marklund and Marklund,32 which uses theinhibition in the rate of autoxidation of pyrogallol asa measure of the activity of SOD. Pyrogallol autoxi-dizes in solution with simultaneous production ofO2
z-anion radical, which, in turn oxidizes other pyro-gallol molecules. The oxidized form of pyrogallolgives an absorption band at 420 nm. Addition of SODinto the system greatly slows down the observed rateof autoxidation of pyrogallol, which is used in theassay of SOD by this method. The enzyme kineticswere monitored on an SLM-AMINCO DW-2000 spec-trophotometer. All calculations were made based onper milligram fresh weight of the tissue.
112 Jain et al. Contraception1999;60:111–118

Spin trapping of oxyradicalsThe superoxide radical was detected by incorporatingDDC as an inhibitor of SOD into the isolation me-dium and then trapping the free radical with PBN.29
The hydride radical (H z2) was trapped as H z2-MNP(methyl nitroso propane) adduct, by incubating thetissue homogenate with 20 mmol/L MNP (final con-centration) for 30 min. After incubation, 50-mL ali-quots were transferred into glass capillaries and oneend flame-sealed. EPR spectra of PBN-O2
z2 adduct andH-MNP adduct were recorded on a Varian E-104 EPRwith TM110 cavity. Instrumental settings employedwere: scan range 100 G, time constant 1 sec, modu-lation amplitude 4G, receiver gain 2.5 3 104 3 10,microwave power 5 mW, field set 3237G, scan time 8min, modulation frequency 100 Hz, temperature27°C, and microwave frequency 9.01 GHz. The EPRabsorption line intensities of the low field, mid-field,and high field lines were calculated employing theequation I 5 kw2h (where k 5 6.51 3 10210, w 5 linewidth, h 5 line height,29 which served as a measure tocompare the quantity of the radical trapped.
Measuring NADPH oxidase activityThe NADPH oxidase activity was measured using adirect method by estimating the NADPH-inducedenhancement in the superoxide production. Each tis-sue homogenate was divided into two aliquots. Onealiquot was incubated with a spin trap PBN forquantifying the basal superoxide anion radical pro-duction. The second aliquot was incubated with PBNin the presence of an excess supplement (20 mmol/L,final concentration) of NADPH. In both cases, incu-bation was carried out for 1 h, after which the sampleswere loaded into glass capillary tubes and the super-oxide quantification was performed using standardspin resonance enhancement protocols. The differ-ence in the spectral response between the NADPH-deficient and NADPH-supplemented incubations wasindicative of the NADPH-oxidase activity.
Spin labeling protocolsThe uterine horns (50 mg of tissue) were mildlysheared in 1 mL of HBSS. A 200 mL aliquot of each ofthe preparations was incubated with 2.6 3 1024
mol/L (final concentration) of the spin label 5-doxylstearate. After 10 min, NiCl2 at a final concentrationof 50 mmol/L was added into the incubation toremove the unincorporated spin label signals and theincubation was continued for another 20 min. Afterthe incubation, the preparations were diluted to 2 mLwith HBSS and were centrifuged at 300 g for 10 min toharvest the cells. The pellet was resuspended in 200mL HBSS. From this suspension, approximately 50 mL
was transferred into glass capillary tubes and wasflame-sealed. The EPR spectra were recorded on aVarian E-104 EPR with TM110 cavity. Instrumentalsettings employed were: scan range 100 G, timeconstant 1.0 sec, modulation amplitude 2 G, receivergain 2.5 3 104 3 10, microwave power 5 mW, field set3237 G, scan time 4 min, modulation frequency 100Hz., temperature 27°C, and microwave frequency9.01 GHz. The rotational correlation time tc wascalculated using the relation tc 5 kwo[(ho/h11)1/221],where w0 is the width, ho and h11 are the mid fieldand low field line heights, and k is a constant with anumerical value of 6.51 3 10210.
Statistical analysisAll the experiments were repeated five times. Obser-vations from the estradiol-treated group were com-pared with their respective controls. Values obtainedin the antiestrogen and RU-39411 groups were com-pared with their respective dosage counterparts fromthe estradiol-treated group. One-way analysis of vari-ance (ANOVA) was run using Introductory StatisticalSoftware Package, version 1.0.33
ResultsThe total SOD activity in the uteri of immature micewas lowered due to administration of estradiol. Al-though the lowest dose (0.24 mg/day/animal) of estra-diol did not bring about a statistically significantdecline in SOD activity, the higher doses broughtabout significant reduction in the activity of thisenzyme. Tamoxifen, ICI-182780, and AF-45 reflecteda dose-dependent increase in SOD activity. Interest-ingly, RU-39411 also caused an increase in the SODactivity in the uterus of immature mice in a dose-dependent fashion (Figure 1).
On the contrary, estradiol resulted in an increase inthe NADPH oxidase activity when compared withthe respective control values. The antiestrogens wereeffective in suppressing the NADPH oxidase activityin a dose-related fashion. RU-39411 was ineffective inaffecting the NADPH oxidase activity at the lowestconcentration tested on comparison with estradioltreated groups. However, at higher doses, RU-39411also introduced inhibition in NADPH oxidase activ-ity (Figure 2).
The quantitative line intensity responses of PBN-O2z2
adduct are plotted graphically in Figure 3. As could beseen in this exhibit, estradiol produced significantaugmentation in the superoxide radical profiles inuterine cell preparations when compared with thecontrol levels. All the other compounds tested signif-icantly lowered (p ,0.01) the superoxide levels in thetest set-up and their effects were dose-dependent.
113Contraception Effect of Estradiol and Antiestrogens on Mammalian Uterus1999;60:111–118
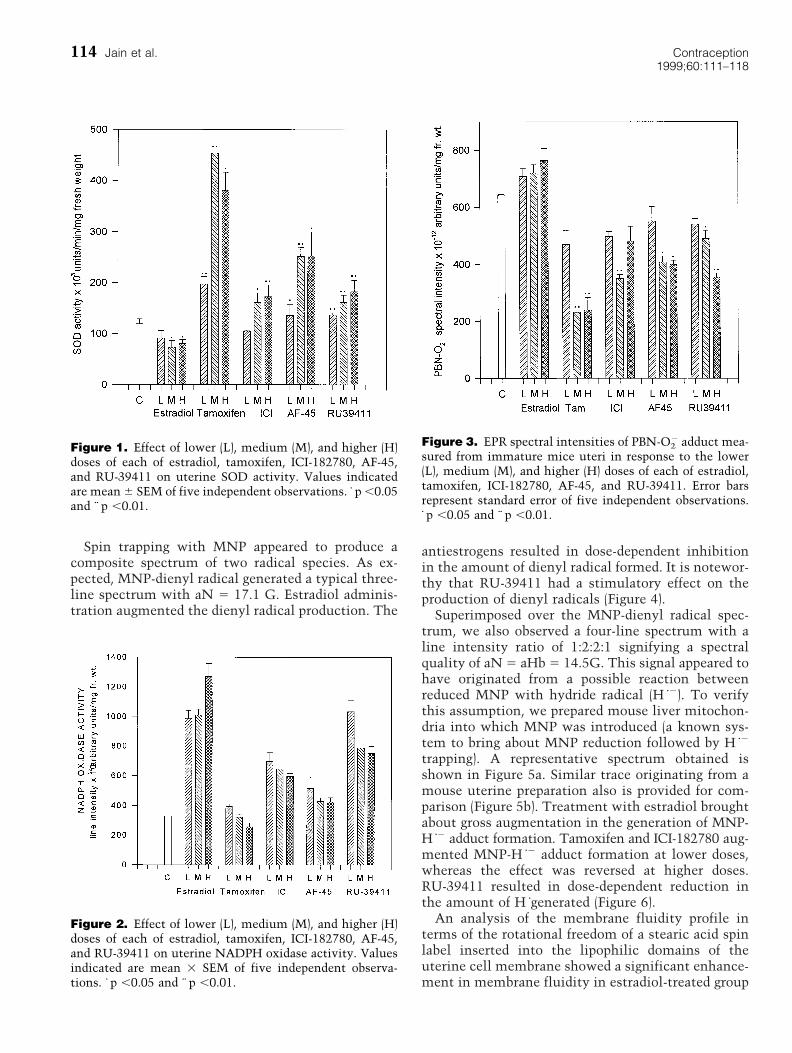
Spin trapping with MNP appeared to produce acomposite spectrum of two radical species. As ex-pected, MNP-dienyl radical generated a typical three-line spectrum with aN 5 17.1 G. Estradiol adminis-tration augmented the dienyl radical production. The
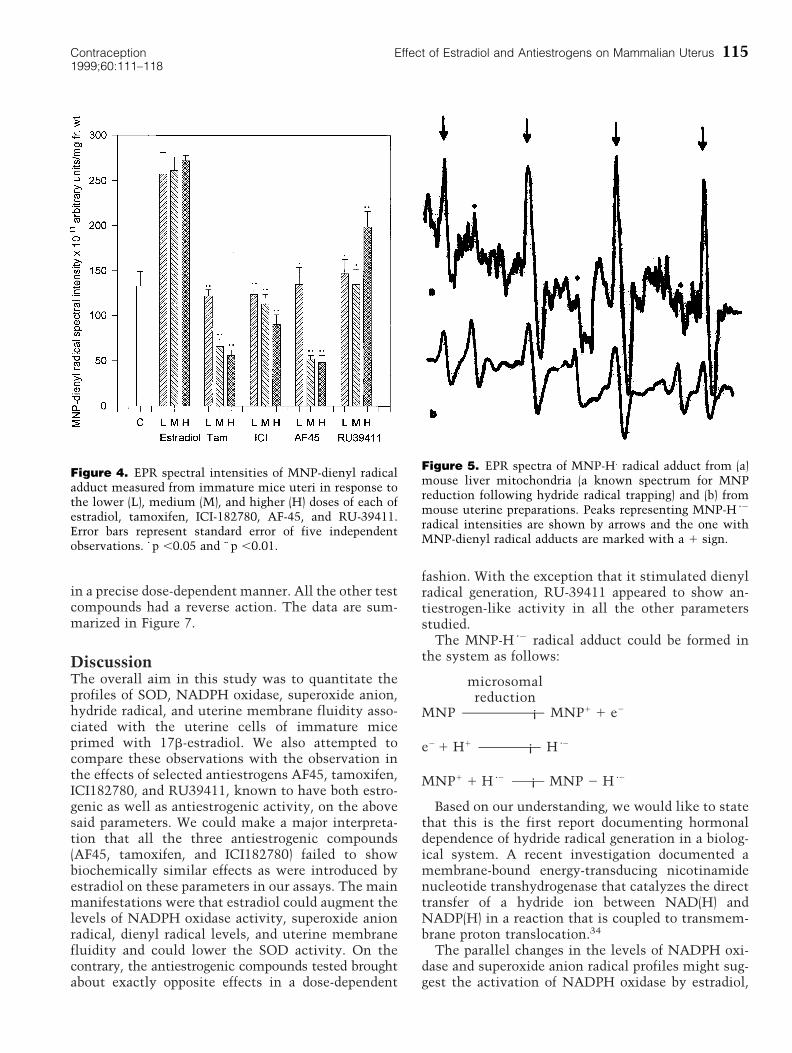
antiestrogens resulted in dose-dependent inhibitionin the amount of dienyl radical formed. It is notewor-thy that RU-39411 had a stimulatory effect on theproduction of dienyl radicals (Figure 4).
Superimposed over the MNP-dienyl radical spec-trum, we also observed a four-line spectrum with aline intensity ratio of 1:2:2:1 signifying a spectralquality of aN 5 aHb 5 14.5G. This signal appeared tohave originated from a possible reaction betweenreduced MNP with hydride radical (H z2). To verifythis assumption, we prepared mouse liver mitochon-dria into which MNP was introduced (a known sys-tem to bring about MNP reduction followed by H z2
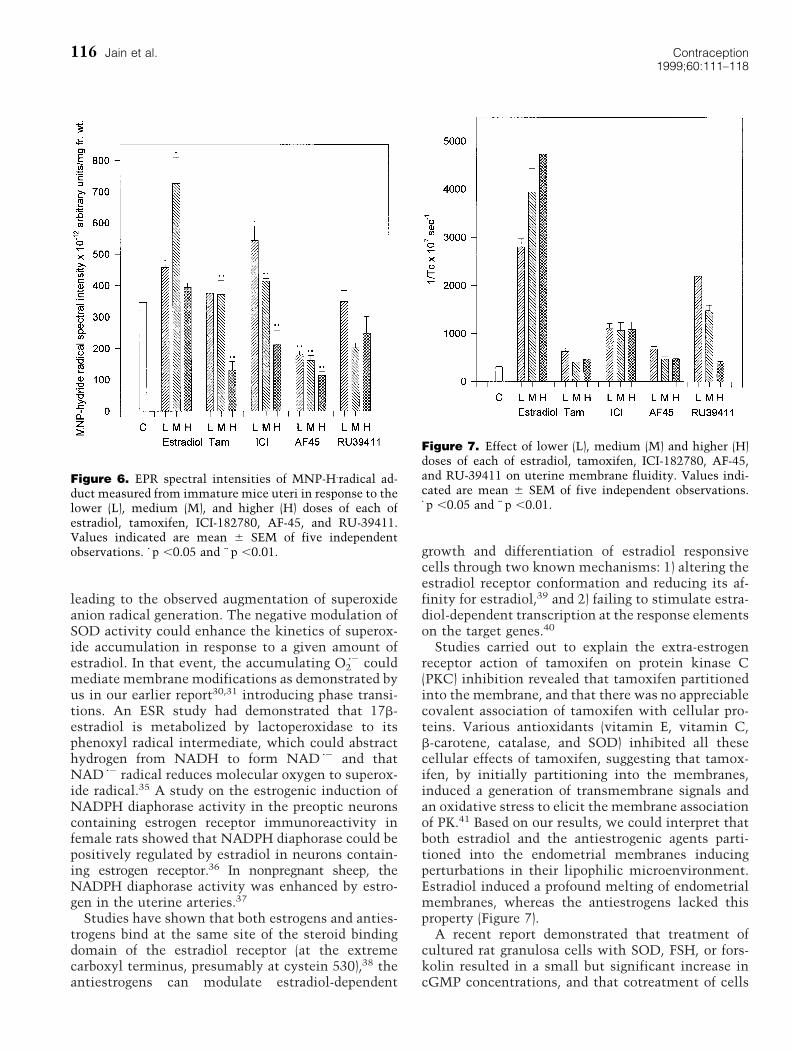
trapping). A representative spectrum obtained isshown in Figure 5a. Similar trace originating from amouse uterine preparation also is provided for com-parison (Figure 5b). Treatment with estradiol broughtabout gross augmentation in the generation of MNP-H z2 adduct formation. Tamoxifen and ICI-182780 aug-mented MNP-H z2 adduct formation at lower doses,whereas the effect was reversed at higher doses.RU-39411 resulted in dose-dependent reduction inthe amount of H zgenerated (Figure 6).
An analysis of the membrane fluidity profile interms of the rotational freedom of a stearic acid spinlabel inserted into the lipophilic domains of theuterine cell membrane showed a significant enhance-ment in membrane fluidity in estradiol-treated group
Figure 1. Effect of lower (L), medium (M), and higher (H)doses of each of estradiol, tamoxifen, ICI-182780, AF-45,and RU-39411 on uterine SOD activity. Values indicatedare mean 6 SEM of five independent observations. z p ,0.05and zz p ,0.01.
Figure 2. Effect of lower (L), medium (M), and higher (H)doses of each of estradiol, tamoxifen, ICI-182780, AF-45,and RU-39411 on uterine NADPH oxidase activity. Valuesindicated are mean 3 SEM of five independent observa-tions. z p ,0.05 and zz p ,0.01.
Figure 3. EPR spectral intensities of PBN-O22 adduct mea-
sured from immature mice uteri in response to the lower(L), medium (M), and higher (H) doses of each of estradiol,tamoxifen, ICI-182780, AF-45, and RU-39411. Error barsrepresent standard error of five independent observations.z p ,0.05 and zz p ,0.01.
114 Jain et al. Contraception1999;60:111–118
in a precise dose-dependent manner. All the other testcompounds had a reverse action. The data are sum-marized in Figure 7.
DiscussionThe overall aim in this study was to quantitate theprofiles of SOD, NADPH oxidase, superoxide anion,hydride radical, and uterine membrane fluidity asso-ciated with the uterine cells of immature miceprimed with 17b-estradiol. We also attempted tocompare these observations with the observation inthe effects of selected antiestrogens AF45, tamoxifen,ICI182780, and RU39411, known to have both estro-genic as well as antiestrogenic activity, on the abovesaid parameters. We could make a major interpreta-tion that all the three antiestrogenic compounds(AF45, tamoxifen, and ICI182780) failed to showbiochemically similar effects as were introduced byestradiol on these parameters in our assays. The mainmanifestations were that estradiol could augment thelevels of NADPH oxidase activity, superoxide anionradical, dienyl radical levels, and uterine membranefluidity and could lower the SOD activity. On thecontrary, the antiestrogenic compounds tested broughtabout exactly opposite effects in a dose-dependent
fashion. With the exception that it stimulated dienylradical generation, RU-39411 appeared to show an-tiestrogen-like activity in all the other parametersstudied.
The MNP-H z2 radical adduct could be formed inthe system as follows:
MNPO¡
microsomalreduction
MNP1 1 e2
e2 1 H1O¡ H z2
MNP1 1 H z2 O¡ MNP 2 H z2
Based on our understanding, we would like to statethat this is the first report documenting hormonaldependence of hydride radical generation in a biolog-ical system. A recent investigation documented amembrane-bound energy-transducing nicotinamidenucleotide transhydrogenase that catalyzes the directtransfer of a hydride ion between NAD(H) andNADP(H) in a reaction that is coupled to transmem-brane proton translocation.34
The parallel changes in the levels of NADPH oxi-dase and superoxide anion radical profiles might sug-gest the activation of NADPH oxidase by estradiol,
Figure 4. EPR spectral intensities of MNP-dienyl radicaladduct measured from immature mice uteri in response tothe lower (L), medium (M), and higher (H) doses of each ofestradiol, tamoxifen, ICI-182780, AF-45, and RU-39411.Error bars represent standard error of five independentobservations. z p ,0.05 and zz p ,0.01.
Figure 5. EPR spectra of MNP-H. radical adduct from (a)mouse liver mitochondria (a known spectrum for MNPreduction following hydride radical trapping) and (b) frommouse uterine preparations. Peaks representing MNP-H z2
radical intensities are shown by arrows and the one withMNP-dienyl radical adducts are marked with a 1 sign.
115Contraception Effect of Estradiol and Antiestrogens on Mammalian Uterus1999;60:111–118
leading to the observed augmentation of superoxideanion radical generation. The negative modulation ofSOD activity could enhance the kinetics of superox-ide accumulation in response to a given amount ofestradiol. In that event, the accumulating O2
z2 couldmediate membrane modifications as demonstrated byus in our earlier report30,31 introducing phase transi-tions. An ESR study had demonstrated that 17b-estradiol is metabolized by lactoperoxidase to itsphenoxyl radical intermediate, which could abstracthydrogen from NADH to form NAD z2 and thatNAD z2 radical reduces molecular oxygen to superox-ide radical.35 A study on the estrogenic induction ofNADPH diaphorase activity in the preoptic neuronscontaining estrogen receptor immunoreactivity infemale rats showed that NADPH diaphorase could bepositively regulated by estradiol in neurons contain-ing estrogen receptor.36 In nonpregnant sheep, theNADPH diaphorase activity was enhanced by estro-gen in the uterine arteries.37
Studies have shown that both estrogens and anties-trogens bind at the same site of the steroid bindingdomain of the estradiol receptor (at the extremecarboxyl terminus, presumably at cystein 530),38 theantiestrogens can modulate estradiol-dependent
growth and differentiation of estradiol responsivecells through two known mechanisms: 1) altering theestradiol receptor conformation and reducing its af-finity for estradiol,39 and 2) failing to stimulate estra-diol-dependent transcription at the response elementson the target genes.40
Studies carried out to explain the extra-estrogenreceptor action of tamoxifen on protein kinase C(PKC) inhibition revealed that tamoxifen partitionedinto the membrane, and that there was no appreciablecovalent association of tamoxifen with cellular pro-teins. Various antioxidants (vitamin E, vitamin C,b-carotene, catalase, and SOD) inhibited all thesecellular effects of tamoxifen, suggesting that tamox-ifen, by initially partitioning into the membranes,induced a generation of transmembrane signals andan oxidative stress to elicit the membrane associationof PK.41 Based on our results, we could interpret thatboth estradiol and the antiestrogenic agents parti-tioned into the endometrial membranes inducingperturbations in their lipophilic microenvironment.Estradiol induced a profound melting of endometrialmembranes, whereas the antiestrogens lacked thisproperty (Figure 7).
A recent report demonstrated that treatment ofcultured rat granulosa cells with SOD, FSH, or fors-kolin resulted in a small but significant increase incGMP concentrations, and that cotreatment of cells
Figure 6. EPR spectral intensities of MNP-H.radical ad-duct measured from immature mice uteri in response to thelower (L), medium (M), and higher (H) doses of each ofestradiol, tamoxifen, ICI-182780, AF-45, and RU-39411.Values indicated are mean 6 SEM of five independentobservations. z p ,0.05 and zz p ,0.01.
Figure 7. Effect of lower (L), medium (M) and higher (H)doses of each of estradiol, tamoxifen, ICI-182780, AF-45,and RU-39411 on uterine membrane fluidity. Values indi-cated are mean 6 SEM of five independent observations.z p ,0.05 and zz p ,0.01.
116 Jain et al. Contraception1999;60:111–118

with FSH plus SOD as well as forskolin plus SOD hada synergistic effect on cGMP content, increasingcGMP levels .100-fold The findings that SOD andactivators of the cAMP-dependent signaling pathwaysynergistically increase the levels of the second mes-senger cGMP and that dibutryl cGMP attenuatedFSH-, forskolin-, and cAMP-induced aromatase activ-ity suggested a potential mechanism of SOD actionand demonstrated the antagonistic action of cGMP oncAMP-mediated estrogen production.42 In our stud-ies, we observed a significant decrease in SOD activ-ity in the uterine tissue after the administration ofestradiol and a significant elevation in SOD activityas a result of antiestrogen treatment (Figure 1). Thus,it appears likely that the SOD and estradiol levels areconnected via a negative feedback regulation. But, atthis point of time, it is difficult to explain the ob-served down-regulation of SOD by estradiol. But it isquite likely that the estrogen receptor bound to an-tiestrogen could have subtle conformational changeswhen compared to estrogen receptor bound to estro-gen. The antiestrogen induced conformation mightbind specifically to some response elements to mod-ulate transcription.43
Considering our observations collectively, estradiolaction on mammalian uterus elevates NADPH oxi-dase activity, augments superoxide anion, dienyl, andhydride radical production, lowers SOD activity, andenhances the membrane fluidity. Tamoxifen, AF-45,and ICI-182780 did not exhibit estrogen-like actionin terms of the parameters of our assays. We believethat the biological activity of these antiestrogensshould be examined using these novel estradiolassay parameters.
AcknowledgmentsThis work was supported by a DST grant SP/SO/B-93/90 to ML and a CSIR-Senior Research Fellowshipto SJ.
References1. Yamamoto KR. Steroid receptor regulation of transcrip-
tion of specific genes and gene networks. Ann RevGenet 1985;19:209–52.
2. Newbold RR, Hanson RB, Jefferson WN. Immaturemouse uterine tissue in organ culture: estrogen-inducedgrowth, morphology and biochemical parameters. InVitro Cell Dev Biol 1994;30A:519–28.
3. Katzenellerbogen BS, Gorski J. Estrogen actions onsynthesis of macromolecules in target cells. In: LitwackG, ed. Biochemical actions of hormones. New York:Academic Press; 1975:187.
4. Markaverich BM, Upchurch S, Clark JH. Two bindingsites for estradiol in rat uterine growth. In: Bresciani F,ed. Perspective in steroid receptor research. New York:Raven Press; 1980.
5. Harris SE, Rosen JM, Means AR, O’Malley BW. Use of aspecific probe for ovalbumin messenger RNA to quan-titate estrogen-induced gene transcripts. Biochemsitry1975;14:20272–81.
6. Deeley RG, Goldberg RF. Regulation of expression ofthe vitellogenin gene in avian liver. In: Hamilton TH,Clark JH, Sadle WA, eds. Ontogery receptors and repro-ductive hormone action. New York: Raven Press; 1979.
7. Westley B, Rochefort H. A secreted glycoprotein in-duced by estrogen in human breast cancer lines. Cell1980;20:353–62.
8. Noteboom WD, Gorski J. An early effect of estrogen onprotein synthesis. Proc Natl Acad Sci USA. 1963;50:250.
9. Lyttle CR, DeSombre ER. Uterine peroxidase as amarker for estrogen action. Proc Natl Acad Sci USA.1977;74:3162–6.
10. Finlay TH, Katz J, Levitz M. Purification and propertiesof an estrogen-stimulated hydrolase from mouseuterus. J Biol Chem 1982;257:10914–9.
11. Kneifel MA, Leytus SP, Fletcher E, Weber T, MangelWF, Katzenellenbogen BS. Uterine plasminogen activa-tor activity modulation by steroid hormones. Endocri-nology 1982;111:493–9.
12. Moulton BC, Barker KL. Synthesis and degradation ofglucose-6-phosphate dehydrogenase in the rat uterus.Endocrinology 1971;89:1131–6.
13. Sarff M, Gorski J. Control of estrogen-binding proteinconcentration under basal conditions and after estrogenadministration. J Biochemistry 1971;10:2557–63.
14. Leavitt WW, Toft DO, Strott CA, O’Malley BW. Aspecific progesterone-receptor in the hamster uterus:physiologic properties and regulation during the estrouscycle. Endocrinology 1974;94:1041–53.
15. Manak R, Wertz N, Slabaugh M, Denari H, Wang J-T,Gorski J. Purification and characterization of the estro-gen-induced protein (i.p.) of the uterus. Mol. Cell Endo-crinol. 1980;17:119–32.
16. Skipper JK, Eakle SD, Hamilton TH. Modulation byestrogen of synthesis of specific uterine proteins. Cell1980;22:69–78.
17. Reiss NA, Kaye AM. Identification of the major com-ponents of the estrogen-induced protein of rat uterus asthe BB isozyme creatine kinase. J Biol Chem 1981;256:5741–9.
18. Fisher B, Constantino JP, Redmond CK, Fisher ER,Wickerham DL, Cronin WMJ. Endometrial cancer intamoxifen-treated breast cancer patients: findings fromthe national surgical adjuvant breast and bowel project(NSABP) B-14. Natl Cancer Inst 1994;86:527–37.
19. Kedar RP, Bourne TH, Powles TJ, Collins WP, AshleySE, Cosgrovo DO, Campbell S. Effects of tamoxifen onuterus and ovaries of postmenopausal women in arandomised breast cancer prevention trial. Lancet 1994;343:1318–21.
20. Wakeling AE. Pharmacology of antiestrogens. Furr BJA,Wakeling AE, eds. Pharmacology and clinical uses ofinhibitors of hormone secretion and action. London:Bailliere Tindall, 1987:1–19.
21. Jordan VC. Biochemical pharmacology of antiestrogenaction. Pharmacol Rev 1984;36:245–76.
22. Wakeling AE. Use of pure antiestrogens to elucidate themode of action of estrogens. Biochem Pharmacol 1995;49:1545–9.
117Contraception Effect of Estradiol and Antiestrogens on Mammalian Uterus1999;60:111–118

23. Wakeling AE, Bowler J. Novel antiestrogens withoutpartial agonist activity. J Steroid Biochem 1991;31:645–53.
24. Wakeling AE, Dukes M, Bowler J. A potent specificpure antiestrogen with clinical potential. Cancer Res1991;51:3867–73.
25. Chin-ping K, Ming-kang C, Hsui-chuan C, Shihi-hsingC, Ta-wei P, Kang T. Pharmacological studies of acontraceptive drug anordrin. Chinese Med J 1976;2:177–4.
26. Crabbe P, Fillion H, Letourneaux Y, Diczfalusy E, AedoAR, Goldzieher JW, Shaikh AA, Castracane VD. Chem-ical synthesis and bioassay of anordrin and dinordrin Iand II. Steroids 1979;33:85–96.
27. Mehta RR, Jenco JM, Chatterton RTJ. Antiestrogenicand antifertility actions of anordrin (2-alpha, 17-alphadiethynyl-A-nor-5 alpha-androstane-2beta-diol 2,17-di-propionate). Steroids 1981;38:679.
28. Kaplankraicer R, Kostovetsky I, Bardin CW, Shalgi R.Tamoxifen and RU39411 synergise with mifepristoneto produce preimplantation pregnancy loss by increas-ing embryo transport in rat. Contraception 1996;53:367–73.
29. Laloraya M, Kumar GP, Laloraya MM. Changes in thelevels of superoxide anion radical and superoxide dis-mutase during the estrus cycle of Rattus norvegicusand the induction of superoxide dismutase in rat ovaryby lutropin. Biochem Biophys Res Commun 1988;157:146–53.
30. Jain S, Thomas M, Kumar GP, Laloraya M. Programmedlipid peroxidation of biomembranes generating kinkedphospholipids permitting local molecular mobility: aperoxidative theory of fluidity management. BiochemBiophys Res Commun 1993;195:574–80.
31. Laloraya M. Fluidity of Phopholipid bilayer of theendometrium at the time of implantation of the blas-tocyst—a spin-label study. Biochem Biophys Res Com-mun 1990;167:561–7.
32. Marklund S, Marklund G. Involvement of superoxideanion radical in the autoxidation of pyrogallol and aconvenient assay for superoxide dismutase. Eur J Bio-chem 1974;47:469–74.
33. Frankenbergre W, Blackmore T. Introductory StatisticsSoftware Package, Verson 1.0, Reading, MA: Addison-Wesley Publishing, 1985.
34. Hatefi Y. Nicotinamide nucleotide transhydrogenase—amodel for utilization of substrate binding energy forprotein translocation. FASEB J. 1996;10:444–52.
35. Sipe HJJ, Jordan SJ, Hanna PM, Mason RP. The metab-olism of 17-beta-estradiol by lactoperoxidase: a possiblesource of oxidative stress in breast cancer. Carcinogen-esis. 1994;15:2637–43.
36. Okamura H, Yokosuka M, Hayashi S. Estrogenic induc-tion of NADPH-diaphorase activity in the preopticneurons containing estrogen receptor immunoreactiv-ity in the female rat. J Neuroendocrinol 1994;6:597–601.
37. Veille JC, Li P, Eisenach JC, Massmann AG, FigueroaJP. Effects of estrogen on nitric oxide biosynthesis andvasorelaxant activity in sheep uterine and renal arteriesin vitro. Am J Obstet Gynecol 1996;174:1043–9.
38. Jordan VC, Murphy CS. Endocrine pharmocology of theantiestrogens as anti tumor agents. Endocr Rev 1990;11:578–612.
39. Sasson S, Notides AC. Mechanism of the estrogenreceptor interaction with 4-hydroxytamoxifen. Mol En-docrinol 1988;2:307–12.
40. Fowell SE, White R, Howe S, Syndeham M, Page M,Parker MG. Inhibition of the estrogen receptor-DNAbinding by the “pure” antiestrogen ICI-164, 384 appearsto be mediated by impaired receptor dimerization. ProcNatl Acad Sci USA 1990;87:6883–92.
41. Gundimeda U, Chen ZH, Gopalakrishna R. Tamoxifenmodulates protein kinase C via oxidative stress inestrogen receptor-negative breast cancer cells. J BiolChem 1996;271:13504–14.
42. Lapolt PS, Hong LS. Inhibitory effects of superoxidedismutase and cyclic guanosine 3959-monophosphateon estrogen production in cultured rat granulosa cells.Endocrinol. 1995;136:5533–9.
43. Macgregor JI, Jordan VC. Basic guide to the mechanismof antiestrogen action. Pharmacol Rev 1998;50:152–96.
118 Jain et al. Contraception1999;60:111–118