Embed Size (px)
Citation preview

1179
PALM OIL CAROTENOIDS
hydrocarbon carotenoids.In this paper we have separated and identified 11
hydrocarbon carotenoids, including seven not previ-ously isolated, in palm oil processed fractions. Theincreasing interest in the medical applications of thisclass of compounds makes t h e i r isolation and character-ization crucial. Palm oil, one of the world's most widelyconsumed vegetable oils and a proven non-toxic food (16),c o u l d become an important commerc i a l source ofcarotenoids. F u t u r e work will focus on high pressureliquid chromatographic analysis of t h e s e compoundsand t h e i r potential anti-carcinogenic and/or antioxida-tive effects.
ACKNOWLEDGMENTSThis work was supported in part by research grants from the PalmOil Research Institute of Malaysia (PORIM) and a BiomedicalResearch Support Grant from the University of Massachusetts toB. Tan. A.S.H. Ong of PORIM and C.T. Chuah of Yee LeeOils andFoodstuffs Co. provided advice.
REFERENCES1. Gawienowski, A.M., M. Stacewicz-Sapuncakis and R.
Longley, J. L i p i d Res. 15:375-379 {1974).2. Haeger-Aronsen, B., G. Krook and M. Abdulla, Internat. J.
Dermat. 18:73-82 (1979).3. Bianchi, A., A. Santagati, L. Andreoni, P. 13ermond and L.
Santamaria, Medicine Biologie Environment 10:25-31 (1982).4. Hunter, R.F., and A.D. Scott, Biochem. J. 35:31-37 (1941}.5. Hunter, R.F., A.D. Scott and J.R. Edisbury, Ibid. 36:697-702
(1942).6. Hunter, R.F., A.D. Scott and N.E. Williams, I b i d .
38:209-211 (1944).7. Hunter, R.F., and A.D. Scott, Ibid. 38:211-213 (1944).8. Hunter, R.F., and R.M. Krakenberger, I b i d . 40:492-
493 (1946).9. DeRitter, E., and A.E. Purcell, in Carotenoids as Colorants and
Vitamin A Precursors, edited by J.C. Baurenfeind, AcademicPress, New York, 1981, pp. 815-836.
10. Davies, B.H., in C h e m i s t r y and B i o c h e m i s t r y o f P l a n tPigments, edited by T.W. Goodwin, Academic Press, London,1976, Vol. 2 , pp. 54-165.
11. DeRitter, E., in Carotenoids as Colorants and Vitamin APrecursors, edited by J.C. Baurenfeind, Academic Press, NewYork, 1981, pp. 883-923.
12. Taylor, R.F., and B.H. Davies, Biochem. J. 139:751-760(1974).
13. Davies, B.H., and A.F. Rees, Phytochemistry 12:2745-2750(1973}.
14. Liaaen-Jensen, S., Kgl. N o r s k e Videnskab. Selskabs S k r i f t e rNo. 8 (1962).
15. Zechmeister, L., in Cis-Trans Isomeric Carotenoids, VitaminsA , and Arylpolyenes, Academic Press, New York, 1962, pp.21-24.
16. Cohen, D., S. Yannai and S. Mokady, Arch. Toxicol., Suppl.6:182-185 (1983).
[Received A u g u s t 19, 1985]
Effect of Culture Conditions on Fatty Acid Composition in LipidsProduced by the Yeast Cryptococcus albidus var. albidusLena Hansson* and Mllan DostalekDepartment of Applied Microbiology, Chemical Center, University of Lund, P.O. Box 124, S-221 O O Lund, Sweden
T h e influence of culture condit ions on the f a t t y acidcomposit ion in lipids produced by Cryptococcus albidusvar. albidus CBS 4517 was studied.
T h e major f a t t y acids in C. albidus var. albidus wereoleic {18:1}, linoleic (18:2) and palmitic {16:0) acid. T h erelative amounts of f a t t y acids produced variedconsiderably during growth and lipid accumulationphases in nitrogen-limited as well as excess-nitrogencultures. T h e degree of unsaturation correlated to thelipid content in the biomass and decreased withincreasing amounts of cellular lipid. A f t e r glucoseexhaustion, no further changes in the f a t t y acidcomposit ion nor in the lipid content of the cells wereobserved. A number of carbon and nitrogen sourcescould b e utilized for lipid synthesis , but they influencedthe f a t t y acid composit ion only to a minor extent.
Lipid production from microorganisms provides a m p l eopportunity for research activities, and the subjectrecently has been reviewed by Ratledge IlL A grea t dealof the work has been performed on lipid-accumulating
*Towhom correspondence should be addressed.
yeast strains because a n u m b e r of them are capable ofa c c u m u l a t i n g large a m o u n t s of intracellular lipids{50-70%, w/w) {2,3}. The lipid fraction usually has a grea tsimilarity t o p l a n t oils (1,4), and with the aim ofproducing substitutes for some of the more expensivetypes , it is important t o gain increased understandingand control over fat ty acid biosynthesis in microorgan-isms.
The y e a s t C r y p t o c o c c u s t e r r i c o l u s Pedersen (nowre-named C. a l b i d u s v a r . a l b i d u s (5) has been reportedt o have a high lipid-accumulating capacity, w h i c h is notdependent on a high C/N r a t i o in the med ium {2,6}. Thisis a u n i q u e characteristic because lipid accumulation inoleaginous yeasts is r ega rded as a two-stage processwhere lipid starts t o be produced from e x c e s s c a r b o na f t e r exhaustion of nitrogen (or other nutrient e x c e p tcarbon} from the medium. Despite its interestingproperties as a lipid producer, very little work has beenreported on this s t r a i n of C. a l b i d u s in recent years.B o u l t o n and Rat ledge (61 c o m p a r e t h e i r r e s u l t sconcerning the constitutive lipid-producing capacitywith those of Pedersen (2,7), whereas Krylova et al. (8,9)deal with lipid synthesis in C. a l b i d u s var. a e r i u s IBFMy-229 u s i n g ethanol as the c a r b o n source. Results from
JAOCS, Vol, 63, no. 9 (September 1986)

1180L. HANSSONAND M. DOST,h,LEK
our laboratory (10) confirm t h a t C. albidus var. albiduscan p roduce s u b s t a n t i a l amounts o f l ipids in t h epresence of excess nitrogen, but the lipid productivitywas f o u n d to be markedly increased u n d e r nitrogen-limited conditions, suggesting an intermediate lipidaccumulation pat tern between the t radi t ional oleagin-ous y e a s t s and t h e originally described C. terricolus.
No investigation has, so far, been repor ted concerningt h e effect o f culture conditions on the fa t ty acidcomposition in C. albidus var. albidus. Such knowledgeis of interest, as control of certain growth parametersmay provide a means to influence the final qual i ty andva lue of the product.
In this work we report on the effects of ca rbon source,n i t rogen source and temperature on fa t ty acid composi-t ion in Cryptococcus albidus var. albidus. Alterations int h e var ious fa t ty ac ids dur ing cul t iva t ion u n d e rnitrogen-l imited and excess-nitrogen conditions aredescribed, as are a correlation between the degree o funsaturation in the lipid fract ion and t h e lipid content ofthe cells.
EXPERIMENTAL PROCEDURES
Microorganism. Cryptococcus albidus var. albidus CBS4517 was used. C. albidus var. albidus was kept onYM-agar slants a t 8 C and was transferred every secondmonth .
Cultivation medium. The nitrogen-limited mediumcontained, per liter: NH4C1, 1.0 g; y e a s t extract , 1.0 g;tryptone, 1.0 g; KH2PO4, 3.0 g; MgSO4.7H~O, 1.0 g;FeC13.6H20, 15 mg; ZnSO~'7H20, 7.5 mg, and CuSO~.5H20, 0.5 rag. The excess-nitrogen medium was thesame as above, but the NH4C1 concentration was 5 g/1.The glucose concentration was 30 g/1 in fermentorcul tures and 20 g/1 in s h a k e r f lask experiments . No p Hadjus tments were performed in s h a k e r f lask cultures; ino rde r to shorten the cultivation time a t decreased pH,the init ial glucose concentration was lowered in theseexperiments . In the nitrogen-limited fed-batch cul tu re ,additional amounts of glucose in concentrated solutionswere added a t 40 and 65 h r of cultivation t o give aconcentration of 30 g/1.
When growing the inoculum, NH~C1 concentration was2.5 g/1 and glucose concentration was 15 g/1. The mediumwas otherwise the same as above. Nei ther ammoniumions n o r glucose were exhaus ted a t t h e time forinoculation. A b o u t 70 ml inoculum (cell suspension)from the logarithmic growth p h a s e was used p e r l i terfermentor volume, resu l t ing in a n init ial optical densityof ~0.6.
Investigations concerning effects o f different carbonor nitrogen sources were performed in nitrogen-limiteds h a k e r f lask cultures. All f lasks contained 0.67 molecarbon/1 (corresponding t o 20 g/1 glucose), a n d then i t rogen source t o be t e s t e d was a d d e d to give aconcentration of 0.019 mole nitrogen/1 (corresponding t o1 g/1 NH,C1).
The p H was adjusted t o 5.5 with NaOH. The ca rbonsource and t h e magnes ium solution were autoclavedseparately. Urea and amino acids were sterilized b ymembrane fil trat ion (0.2 ~m).
Cultivation equipment and conditions. Fermentorcultures were performed in a fermentor (ChemofermAB,
Hfigersten, Sweden) with a working volume o f 3.0 1. Thecultivation temperature was 20 C, and p H was keptconstant a t 5.5 b y t i t ra t ion with 1.0 M NaOH. Air wassparged t h r o u g h t h e b ro th with a flow rate o f 0.5 VVM(volume/volume, min). The dissolved oxygen tension wasmeasured continuously with a galvanic oxygen electrode(11). No oxygen limitation occurred dur ing t h e cultiva-tions. The st i r rer speed was 350 rpm. Foaming wascont ro l led b y addi t ion o f Adekanol LG-109 (AsahiElectro Chemical Co., Japan) .
Shaker f lask experiments were performed in one-Ibaff led flasks, containing 150 ml of cell suspension, on awate r b a t h rotary shaker .
In the inves t iga t ion o f temperature effects t h einoculum was grown a t 22.5 C and then used fo rcul turesa t 20, 25 and 30 C. In all o t h e r experiments , t h eincubation temperature was 20 C for inoculum as well asfor subsequent cultures.
Growth measurements. Cell g rowth was followed bymeasur ing opt ica l density (OD) a t 620 nm in a Turne rpho tome te r (G.K. Turne r Associates, Palo Al to , Califor-nia). The dry weight ofb iomass was determined by centri-fuging 15 ml of cell suspension, washing the pel let with0.9% NaCI and dry ing a t 105 C for 17 hr.
Lipid determination. 100 ml (OD~< 10) or 50 ml (OD > 10) ofcell suspension was removed from the culture. Each samplewas centrifuged at 20,000 xg for 10 min at 10 C and washedonce in 0.9% NaC1. The pellet was kept in a sealedcentrifuge tube and s t o r e d a t - 2 0 C.
Before extract ion of l ipids was performed, t h e cellswere disintegrated. Two ml of wate r was added t o thethawed pellet and the cell suspension was sonicated for 6min on ice in a n MSE Soniprep 150 UltrasonicDis in tegra tor(MSE, Crawly, England). Thereafter, 5 g ofglass beads (~ = 0.5-0.75 mm) were a d d e d and the cellswere d i s rup ted in a Lab-Line Mul t iwr is t Shaker(Lab-Line Instr . , Inc. , Melrose Park, Illinois) a t m a x i m u mintensity for 1.5 hr.
Lipid extract ion was performed with n-butanol as t h esolvent. 10 ml of b u t a n o l was a d d e d t o t h e cellsuspension, and the tube was shaken, as describedabove, for 20 min. The butanol phase was separated fromcells and g l a s s beads b y centrifugation. The extract ionprocedure was repeated twice, and the b u t a n o l extractswere pooled. The extracts were washed according t o themethod of Folch e t al. (12) by shak ing with 5 ml of wate rfor 5 min. Af t e r centrifugation, the b u t a n o l p h a s e wast ransfer red to a weighed vessel and evapora t ed u n d e rvacuum. The lipid was dried over anhydrous P205 in anevacuated desiccator, and the amount of lipid wasdetermined gravimetrically.
Fat t y acid composition. The fa t ty acid compositionwas determined by gas chromatography of t h e fa t ty acidmethyl esters. The lipids to be transesterified weredissolved in hexane , and methyl es te r s were preparedaccording to the procedure o f Glass e t al. (13).
The ins t rument u s e d was a V a r i a n 3700 GasChromatograph. Samples were separated on a stainlesssteel column (1.8 m × 2 mm) containing 10% DEGSliquid p h a s e coated on Chromosorb W HP (80-100mesh). Ni t rogen with a flow rate of 50 ml/min was usedas the carr ier gas. The oven temperature was pro-grammed from 130 t o 180 C a t the ra te of 2 C/min. Peakswere identified by means of fa t ty acid methyl e s t e r
JAOCS, Vol, 63, no. 9 (September 1986)

1181
FATTYACID COMPOSITION IN C. ALBIDUS
s t andard FO6 (Larodan Fine Chemicals, Analy t ica lS t anda rds AB, Kungsbacka, Sweden}.
The t o t a l amount of fa t ty acids in t h e lipid fract ionconstitutes 100%. Besides those fa t ty acids t h a t wereidentified by u s i n g the FO6 s tandard (14:0, 16:0, 16:1,18:0, 18:1, 18:2, 18:3}, a small pa r t , less t h a n 4% of t h efraction, consisted o f o t h e r fa t ty acids, viz. 15:0, 17:0,17:1, 20:0, 20:1, 20:2, 20:3, 22:0, 22:1, 24:0, and 24:1.T h o s e were identified in a few samples by capillarycolumn chromatography by A B Karlshamns Oljefabriker.Unsa tu ra t ed fa t ty acids from this part o ft h e lipid fract ionwere not included in the calculation of the degree o f un-saturation.
The degree o f unsa tu ra t i on (A/mole} in the lipidfract ion was calculated according t o Kates and Baxter{14}. The formula used is:
A/mole =[1.0 I% monoene} + 2.0 (% diene) + 3.0 (%triene)]/100
Analyses o f glucose and NH4Cl. All assays wereperformed on samples where cells had been removedfrom the cul t iva t ion b ro th b y cent r i fugat ion a n dmembrane fi l t rat ion. Glucose was measured b y u s i n g aBoehringer t e s t kit (Boehringer, Mannheim, Federa lRepublic o f Germany}, and NH4C1 was assayed asammonium ions according to Chaney and M arbach (15).
RESULTS A N D DISCUSSION
Effect o f carbon source, nitrogen source and g r o w t htemperature on fa t ty acid composition. The influence ofdifferent ca rbon sources on growth and fa t ty acidcomposition was investigated in nitrogen-limited (1 g/1NH4C1} shaker-flask cultures. The cells were harvestedin the stationary phase, and dry weigh t , lipid contentand fa t ty acid composition were determined {Table 1}. C.albidus var. albidus was able t o utilize all the carbonsources t e s t e d except ethanol, b u t very poor growthtook place on glycerol. It is c lear from Table 1 t h a t only
m i n o r changes were o b s e r v e d in t h e f a t t y acidcompos i t ion , a n d consequen t ly in t h e degree o funsa tu ra t i on (/l/mole}.
The stabi l i ty in fa t ty acid composition regardless o fthe ca rbon source utilized is in agreement with resul t srepor tedfo rCandida 107 (16,17) and R h o d o torula gracilisLundin R-gl 172 {18}. The m o s t pronounced alterationsin fa t ty acid composition as a function o f carbon sourcehave been reported for n-alkanes {1,19}. The abil i tyamong y e a s t s to utilize n-alkanes is, however, res t r ic tedt o a limited number o f strains.
The effect o f different nitrogen sources on the fa t tyacid composition is shown in Table 2. The ca rbon sourcewas glucose, 20 g/1. The growth curves (not shown} werea lmos t identical regardless of t h e n i t rogen source used.A drop in pH, most marked in NH4C1, (NH4hSO4 andL-ornithine cultures, was observed. C. albidus var.albidus has, however, previously been f o u n d t o have ab road pH-opt imum for growth (20, and Hansson andDost~lek, unpublished results} and because growthrates were n o t affected, the changes in p H were assumednot t o affect markedly the resul t s presented below.
The relat ive amount of stearic acid (18:0) was a b o u t5.6% when t h e cells were grown on inorganic and~10.5% on organic nitrogen sources. A s l igh t decreasein palmitic acid (16:0} was observed when L-arginine orL-ornithine was used, and an increase in linoleic acid(18:2) was n o t e d when cells were grown on any of theamino acids. The degree o funsa tu ra t i on was in the sameorde r o f magni tude fo r all the n i t rogen sources tested.
The n i t rogen source used might affect the fa t ty acidcomposition b y differences in t h e uptake rates fo rorganic a n d inorganic n i t rogen o r in t h e n i t rogencatabolism. The intracel lular NH~ concen t ra t ionrecent ly was reported t o affect lipid production ra te andt h e regulation o f lipid biosynthesis in Rhodosporidiumtoruloides Banno CBS 14 {21,22}. R h . toruloides con-t a ined , in c o n t r a s t to C. albidus var. albidus, h ighe ramounts o f lipid when grown on organic n i t rogensources than on inorganic. No da t a concerning fa t ty acid
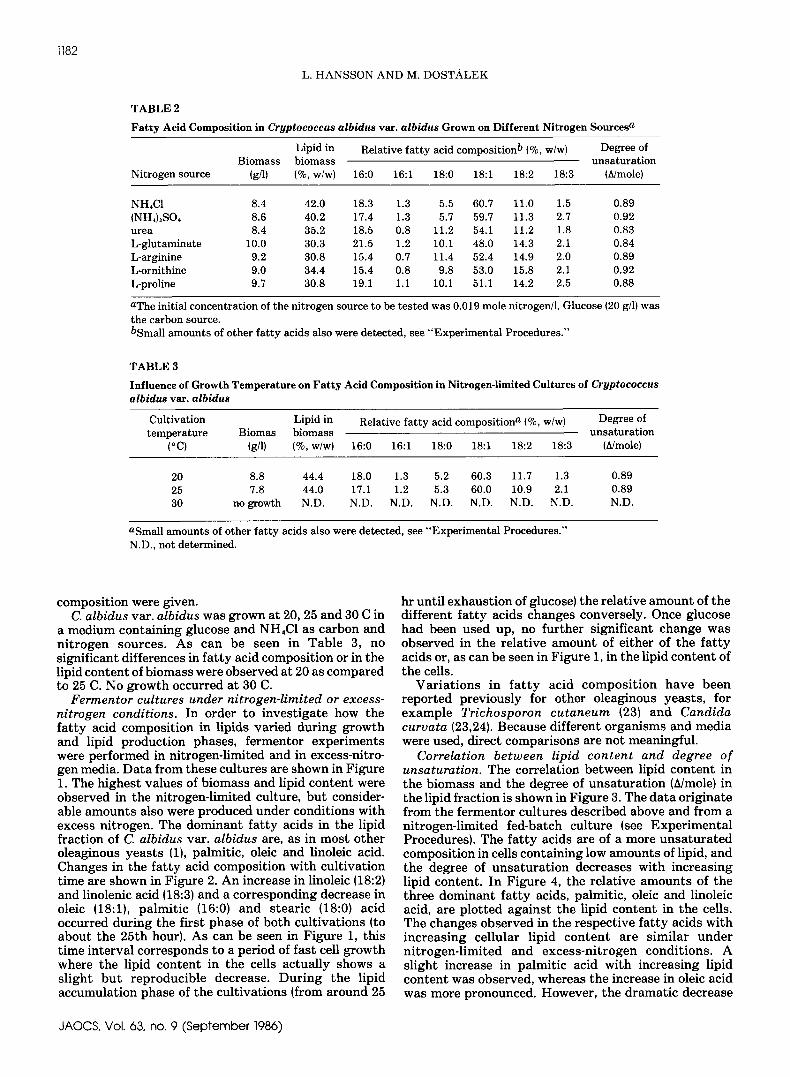
T A B L E 1
F a t t y A c i d C o m p o s i t i o n i n Cryptococcus a l b i d u s va t . a l b i d u s Grown o n Different C a r b o n Sources i nNitrogen-Limited Cultures I1 g / l NH4CI)a
Lipid in Relative fatty acid composition b (%, w/w) DegreeofBiomass biomass unsaturation
Carbon source (g/l} (%,w/w} 16:0 16:1 18:0 18:1 18:2 18:3 (A/mole)
xylose 7.1 33.0 21.9 1.7 4.3 54.5 13.8 1.1 0.87glucose 8.3 40.1 18.6 1.2 5.4 60.9 11.2 1.1 0.88maltose 8.2 37.7 22.2 1.2 6.0 53.9 13.2 1.2 0.85lactose 6.5 26.3 25.8 1.2 5.1 47.4 16.6 1.4 0.86sol. starchc 4.7 33.1 22.0 0.9 5.1 51.9 16.1 1.5 0.90mannitol 7.8 43.5 22.3 0.9 8.2 55.4 10.4 1.3 0.81glycerol c 1.5 43.8 20.5 1.1 5.9 52.4 14.8 1.3 0.87ethanol d nogrowth N.D. N.D. N.D. N.D. N.D. N.D. N.D. N.D.
aInitial carbon source concentration was 0.67 mole carbon/1 unless otherwise indicated.bSmall amounts ofother fatty acids also were detected, see "Experimental Procedures."CInitial carbon source concentration, 15 g/1.dInitial carbon source concentration, 1Og/1.N.D., not determined.
JAOCS, Vol. 63, no. 9 (September 1986)
1182
L. HANSSON AND M. DOSTfi.LEK
TABLE 2
F a t t y A c i d C o m p o s i t i o n i n Cryptococcus albidus va t . albidus Grown o n Different Ni trogen Sources a
Lipid in Relative fatty acid compositionb (%, w/w) Degree ofBiomass biomass unsaturation
Nitrogen source (g/l) (%, w/w) 16:0 16:1 18:0 18:1 18:2 18:3 (A/mole)
NH4C1 8.4 42.0 18.3 1.3 5.5 60.7 11.0 1.5 0.89(NH4)2SO4 8.6 40.2 17.4 1.3 5.7 59.7 11.3 2.7 0.92urea 8.4 35.2 18.5 0.8 11.2 54.1 11.2 1.8 0.83L-glutaminate 10.0 30.3 21.5 1.2 10.1 48.0 14.3 2.1 0.84L-arginine 9.2 30.8 15.4 0.7 11.4 52.4 14.9 2.0 0.89L-ornithine 9.0 34.4 15.4 0.8 9.8 53.0 15.8 2.1 0.92L-proline 9.7 30.8 19.1 1.1 10.1 51.1 14.2 2.5 0.88
aThe initial concentration of the nitrogen source to be tested was 0.019 mole nitrogen/1. Glucose(20 g/l) wasthe carbon source.bSmall amounts of other fatty acids also were detected, see "Experimental Procedures."
T A B L E 3
Influence of Growth Temperature o n F a t t y A c i d C o m p o s i t i o n i n Ni trogen- l imi ted Cultures of Cryptococcusalbidus var . albidus
Cultivation Lipid in Relative fatty acid compositiona (%, w/w) Degree oftemperature Biomas biomass unsaturation
(°C) (g/l) (%, w/w) 16:0 16:1 18:0 18:1 18:2 18:3 (A/mole)
20 8.8 44.4 18.0 1.3 5.2 60.3 11.7 1.3 0.8925 7.8 44.0 17.1 1.2 5.3 60.0 10.9 2.1 0.8930 no growth N.D. N.D. N.D. N.D. N.D. N.D. N.D. N.D.
aSmall amounts of other fatty acids also were detected, see "Experimental Procedures."N.D., not determined.
composition were given.C. albidus var. albidus was g r o w n at 20, 25 and 30 C in
a med ium containing glucose and NH4C1 as carbon andn i t r o g e n sources. As can be seen in T a b l e 3, nosignificant differences in fat ty acid composition or in thelipid content of biomass were observed at 20 as comparedt o 25 C. No g r o w t h occurred at 30 C.
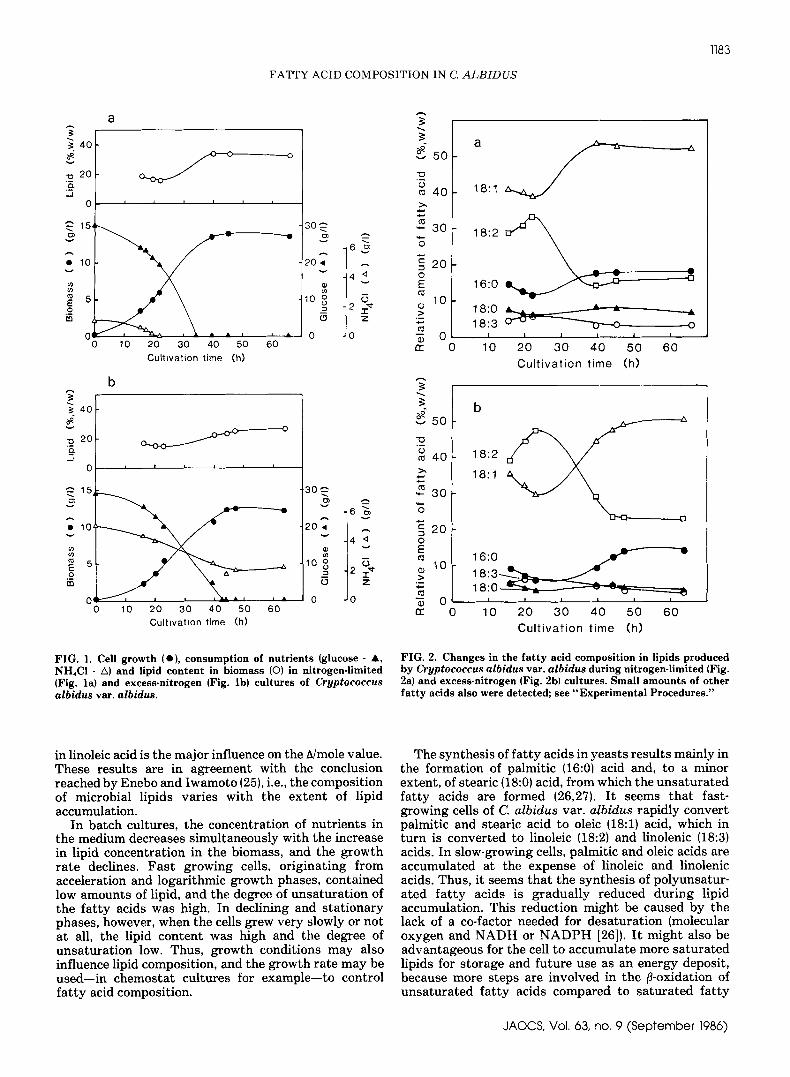
Fermentor cultures under nitrogen-limited or excess-nitrogen conditions. In o r d e r t o investigate how thefat ty acid composition in lipids v a r i e d d u r i n g g r o w t hand lipid production phases, fermentor experimentswere performed in nitrogen-limited and in excess-nitro-gen media. Data from these cultures are shown in F i g u r e1. The highest values of biomass and lipid content wereobserved in the nitrogen-limited culture, but consider-able a m o u n t s also were produced u n d e r conditions withexcess nitrogen. The dominant fat ty acids in the lipidfraction of C. albidus vat. albidus are, as in most otheroleaginous yeasts (1), palmitic, oleic and linoleic acid.Changes in the fat ty acid composition with cultivationtime are shown in F i g u r e 2. An increase in linoleic (18:2}and linolenic acid (18:3) and a corresponding decrease inoleic (18:1), palmitic (16:0) and s t e a r i c (18:0) acidoccurred d u r i n g the first p h a s e of both cultivations (toa b o u t the 25th hour}. As can be seen in F i g u r e 1, thistime interval corresponds t o a period of fast cell g r o w t hwhere the lipid content in the cells actually shows as l i gh t but reproducible decrease. D u r i n g the lipidaccumulation p h a s e of the cultivations (from a r o u n d 25
hr u n t i l exhaustion of glucose} the relative a m o u n t of thedifferent f a t t y acids changes conversely. Once glucosehad been used up , no further significant change wasobserved in the relative a m o u n t of either of the f a t t yacids or, as can be seen in F i g u r e 1, in the lipid content ofthe cells.
V a r i a t i o n s in f a t t y acid composition have beenreported previously for other oleaginous yeasts, forexample Trichosporon c u t a n e u m (23} and Candidacurvata {23,24}. B e c a u s e different organisms and med iawere used, direct comparisons are not meaningful.
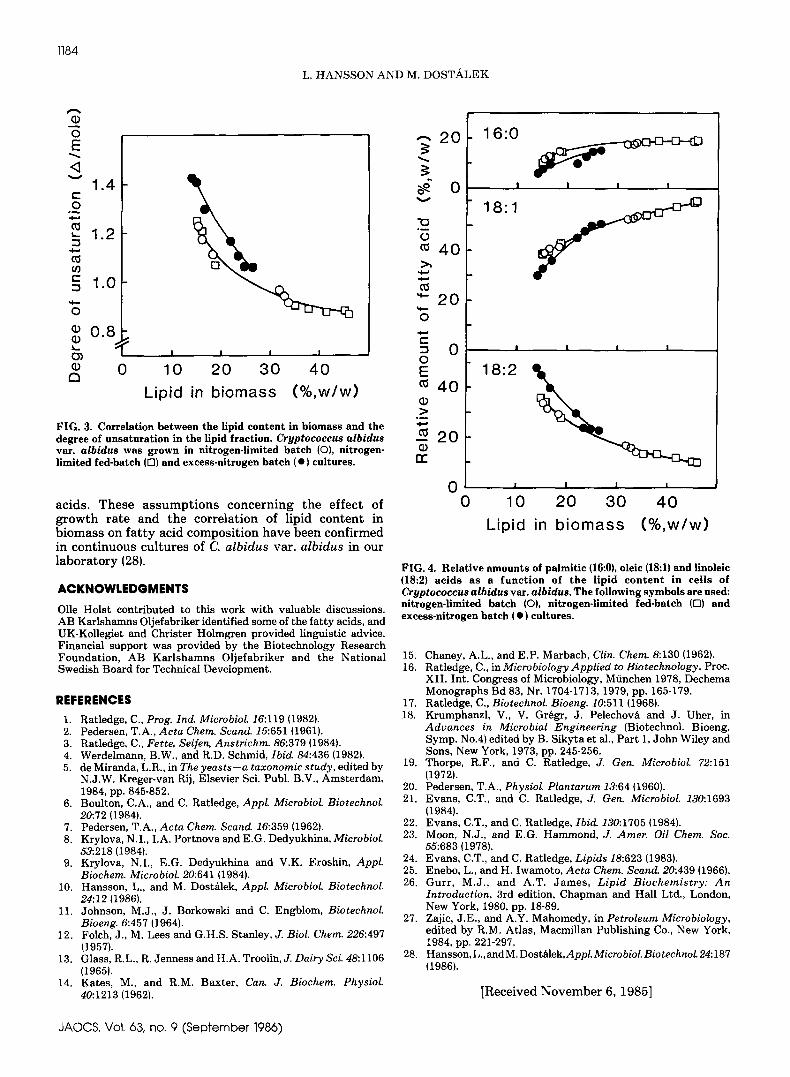
Correlation b e t w e e n l i p i d c o n t e n t and degree o funsaturation. The correlation between lipid content inthe biomass and the degree of unsaturation (A/mole) inthe lipid fraction is shown in F i g u r e 3. The data originatefrom the fermentor cultures described a b o v e and from anitrogen-limited fed-batch culture {see ExperimentalProcedures}. The fat ty acids are of a more unsaturatedcomposition in cells containing low amounts of lipid, andthe degree of unsaturation decreases with increasinglipid content. In F i g u r e 4, the relative a m o u n t s of thet h r e e dominant fat ty acids, palmitic, oleic and linoleicacid, are plotted a g a i n s t the lipid content in the cells.The changes observed in the respective fat ty acids withi n c r e a s i n g cellular lipid content are s i m i l a r u n d e rnitrogen-limited and excess-nitrogen conditions. Aslight increase in palmitic acid with increasing lipidcontent was observed, whereas the increase in oleic acidwas more pronounced. However, the d r a m a t i c decrease
JAOCS, Vol. 63, no. 9 (September 1986)
FATTY ACID COMPOSITION IN C. A L B I D U S
1183
A
40
"o 20"E_ J
0
15~
• 10
0
40
•"u 20
J
o
--'~ 15,C3~
v
• 10,
E 50
¢ ,n
i i i I
_ A
10 20 30 40 50 60Cultivation time (h)
0
00 10 20 30 40 50 60
Culhvatmn time (h)
3 0 ~
20 ~ <
lO =o
e =7
o o
~o~
(D
lot ~ 2
o o
FIG. 1. Cell growth (e), consumpt ion of nutrients ( g l u c o s e - A ,NH4CI - z~) and l i p i d content in b i o m a s s (©) i n ni trogen- l imited( F i g . l a ) and excess-ni trogen ( F i g . lb) cultures of Cryptococcusalbidus var . a lb idus .
A
{ 5o- O
"~ 4 0 i> ,
~ a o
2 o0E
10
-~ 0~: 0
18:1
18:2 ~ e
16:0 --m
18:0 ~ , t ~ _ . ~ -"18:3 o---x.r~ ~ - - ~ ~o
I I I I I I
10 20 30 40 50 6 0Cultivation time (h)
5 0" O
0 40> ,
~ 3O
e ooE
10, , . . ,
"~ 0rr
b
18:2
18:1
rl
16:0 ~ 1
1 8 : 0 ~ -
i I I I I I
10 20 30 40 50 60Cultivation time (h)
FIG. 2. Changes i n the fat ty a c i d c o m p o s i t i o n i n l ip ids producedby Cryptococcus albidus va t . albidus during ni trogen- l imited ( F i g .2a} and excess-ni trogen ( F i g . 2b) cul tures . S m a l l amounts of otherfat ty acids also were detected; see "Experimental Procedures."
in linoleic acid is t h e major influence on the h/mole value.These resul t s are in agreement with the conclusionreached by Enebo a n d Iwamoto (25}, i.e., t h e compositiono f microbial l ipids varies with the extent o f lipidaccumulation.
In b a t c h cultures, t h e concentration of nut r ien ts inthe medium decreases simultaneously with t h e increasein lipid concentration in the biomass, and the growthra te declines. F a s t growing cel l s , or ig inat ing fromacceleration and logarithmic growth phases, containedlow amounts of l ipid, a n d t h e degree of unsaturation ofthe fa t ty acids was high. In declining and s ta t ionaryphases, however, when the cells grew very slowly or nota t all, the lipid content was high a n d the degree ofunsaturation low. T h u s , g rowth conditions may alsoinfluence lipid composition, and t h e growth ra te may beused-- in cbemos ta t cultures for example--to controlfa t ty acid composition.
The synthesis of fa t ty ac ids in yeasts resul t s mainly inthe formation of palmitic (16:0) acid and, to a minorextent , o f stearic (18:0) acid, from which t h e u n s a t u r a t e dfa t ty acids are formed (26,27}. It seems t h a t f a s t -growing cells of C. a l b i d u s var. a l b i d u s rapidly conver tpalmitic and stearic acid to oleic (18:1) acid, which inturn is converted to linoleic (18:2) a n d linolenic (18:3)acids. In slow-growing cells, palmitic and oleic acids areaccumulated a t the expense o f linoleic and linolenicacids. T h u s , it seems t h a t the synthesis of polyunsatur-ated fa t ty acids is gradually reduced dur ing lipidaccumulation. This reduction might be caused b y thelack of a co-factor needed for desaturation {molecularoxygen and NADH or N A D P H [26]}. It might also b eadvantageous for the cell t o accumulate more sa tu r a t edlipids fo r storage and future use as an ene rgy deposit,because more s t e p s are involved in t h e ~-oxidation ofu n s a t u r a t e d fa t ty acids compared to sa tu r a t ed fa t ty
JAOCS, Vol. 63, no. 9 (September 1986)
1184
L. H A N S S O N AND M . D O S T A L E K
0E
<11.4c-
O
~- 1.2
ffl
~- 1.0
0
~ •
~ 0121
I I I I
10 20 3 0 40Lipid in b i o m a s s ( % , w / w )
FIG. 3 . C o r r e l a t i o n between the l i p i d content i n b i o m a s s and thedegree of unsaturat ion i n the l i p i d fract ion. Cryptococcus albidusva t . albidus w a s grown i n ni trogen- l imited batch (O), ni trogen-l imi ted fed-batch ([:]} and excess-ni trogen batch ( • ) cul tures .
a c i d s . T h e s e a s s u m p t i o n s c o n c e r n i n g t h e e f f e c t ofg r o w t h r a t e a n d t h e c o r r e l a t i o n o f l i p i d c o n t e n t inb i o m a s s o n f a t t y acid c o m p o s i t i o n h a v e b e e n c o n f i r m e din c o n t i n u o u s c u l t u r e s of C. a l b i d u s v a r . a l b i d u s in o u rl a b o r a t o r y (28).
A C K N O W L E D G M E N T S
Ol le H o i s t contr ibuted to th i s work with valuable discuss ions .AB Karlshamns Oljefabriker ident if ied some of the f a t t y acids , andUK-Kollegiet and Chris ter Holmgren provided l ingu i s t i c advice.Financial support was provided by the Biotechnology ResearchFoundat ion , AB Karlshamns Oljefabriker and the NationalSwedish B o a r d for Technical Development .
R E F E R E N C E S
i . Ratledge, C., Prog. Ind. Microbiol. 16:119 (1982}.2. Pedersen, T.A., A c t a Chem. Scand. 15:651 {1961}.3. Ratledge, C., Fette, Seifen, Anstrichm. 86:379 {1984}.4. Werdelmann, B.W., and R.D. Schmid , Ibid. 84:436 (1982).5 . de Miranda, L.R., in The y e a s t s - - a taxonomic s t u d y , edited by
N . J . W . Kreger-van Rij, Elsevier Sci. Pub l . B.V., Amsterdam,1984, pp . 845-852.
6. Bou l ton , C.A., and C. Ratledge, Appl. Microbiol. Biotechnol.20:72 (1984).
7 . Pedersen, T.A., A c t a Chem. Scand. 16:359 (1962).8 . Krylova, N.I., I.A. Portnova and E.G. Dedyukhina, Microbiol.
53.'218 (1984).9 . Krylova, N.I., E.G. Dedyukhina and V.K. Eroshin , Appl.
Biochem. Microbiol. 20:641 (1984).10. Hansson, L., and M . Dost&lek, Appl. Microbiol. Biotechnol.
24:12 (1986).11. Johnson , M . J . , J . Borkowski and C. Engblom, Biotechnol.
Bioeng. 6:457 (1964).12 . Folch , J., M . Lees and G.H.S. Stanley , J. Biol. Chem. 226:497
(1957).13 . Glass , R.L., R . Jenness and H.A. Trool in , J. D a i r y Sci. 48:1106
(1965).14 . Kates, M., and R.M. Baxter, Can. J. Biochem. Physiol.
40:1213 (1962).
-~ 20
~ 0
"0. m
0m 4o
~ 200¢-= 00Em 4O>
o m
--~ 2Or r
16:0
I I I I
18:1 S
I I I I
18"2
0 i I I i0 10 2 0 3 0 4 0
L i p i d in b i o m a s s (%,w/w)
FIG. 4. Re la t ive amounts of p a l m i t i c (16:0), o l e i c 118:1) and l i n o l e i c(18:2} acids a s a funct ion of the l i p i d c o n t e n t i n ce i l s o fCryptococcus albidus var . a lb idus . The fo l lowing s y m b o l s are used:ni trogen- l imited b a t c h (O}, n i trogen- l imited fed-batch (D) andexcess-ni trogen batch { • } cul tures .
15. Chaney, A.L., and E.P . Marbach, Clin. Chem. 8:130 (1962).16. Ratledge, C., in Microbiology A p p l i e d to Biotechnology, Proc.
XII. In t . Congress of Microbiology, M(inchen 1978, DechemaMonographs Bd 83, Nr. 1704-1713,1979, pp . 165-179.
17. Ratledge, C., Biotechnol. Bioeng. 10:511 {1968}.18. Krumphanzl , V., V. Gr~gr, J . Pelechov~ and J . U h e r , in
A d v a n c e s in Microbial Engineering (Biotechnol . Bioeng.Symp . No.4) edited by B . S i k y t a et al., Part 1, John Wiley andSons , New York, 1973 , pp . 245-256.
19. Thorpe, R.F., and C. Ratledge, J. Gen. Microbiol. 72:151(1972).
20. Pedersen, T.A., Physiol. Plantarum 13:64 {1960}.21. E v a n s , C.T., and C. Ratledge, J. Gen. Microbiol. 130:1693
(1984).22. E v a n s , C.T., and C. Ratledge, Ibid. 130:1705 {1984}.23. Moon, N . J . , and E.G. Hammond, J. A m e r . Oil Chem. Soc.
55:683 (1978).24. E v a n s , C.T., and C . Ratledge, L i p i d s 18:623 (1983}.25. Enebo, L., and H. Iwamoto, A c t a Chem. Scand. 20:439 (1966).26. G u r r , M . J . , and A.T. J a m e s , L i p i d B i o c h e m i s t r y : A n
Introduction, 3rd edi t ion , Chapman and Hall Ltd., London,New York, 1980, pp . 18-89.
27 . Zajic, J . E . , and A.Y. Mahomedy, in Petroleum Microbiology,edited by R.M. At las , Macmil lan Publ i sh ing Co., New York,1984 , pp . 221-297.
28 . Hansson, L.,andM.Dostalek,AppLMicrobioLBiotechnol.24:187(1986).
[ R e c e i v e d N o v e m b e r 6 , 1 9 8 5 ]
JAOCS, Vol. 63, no. 9 (September 1986)