Embed Size (px)
Citation preview

Cell Biology International 2000, Vol. 24, No. 6, 365–373doi:10.1006/cbir.1999.0519, available online at http://www.idealibrary.com on
EFFECT OF ABSCISIC ACID AND COLD ACCLIMATION ON THE CYTOSKELETALAND PHOSPHORYLATED PROTEINS IN DIFFERENT CULTIVARS OF
TRITICUM AESTIVUM L.
OLGA V. OLINEVICH1*, LUDMILA P. KHOKHLOVA1 and MARJATTA RAUDASKOSKI2
1Department of Plant Physiology, Kazan State University, Kremlyovskaya str. 18, 420008, Kazan, Russia2Division of Plant Physiology, Department of Biosciences, P.O. Box 56, FIN-00014 University of Helsinki, Finland
Received 30 April 1999; accepted 9 August 1999
In winter wheat, the tubulin and 60 kDa-phosphorylated proteins/actin ratio is considerablyhigher in the roots than in the leaves. Differences in the content of the main cytoskeletal proteinswere also found in the leaves of the different cultivars. It is suggested that the lower amount ofthe tubulin and 60 kDa-phosphorylated proteins and higher content of actin determine thegreater tubulin cytoskeletal stability in the leaves and their higher frost resistance, as comparedwith the roots. Also, it is possible that the higher content of the tubulin and 60 kDa-phosphorylated proteins defines the lower microtubule (MT) stability in the leaves of the lowfrost resistant cultivar than in the leaves of the more frost resistant ones. In the roots and leavesof the low frost resistant cultivar, the low stability of the numerous tubulin structures isapparently one reason for the abscisic acid (ABA)-induced reduction of the cytoskeletal and60 kDa-phosphorylated proteins in the cells. The cold acclimation compensated the ABA effectin the roots of the very frost resistant cultivar in the most extent. This suggests the existence ofthe different pathways in the increased plant cell frost resistance through the action of ABA andlow temperature. � 2000 Academic Press
K: Triticum aestivum L.; leaves; roots; abscisic acid; cold acclimation; tubulin; actin;phosphorylated proteins; wheat cultivars.A: ABA, abscisic acid; MTs, microtubules; MFs, microfilaments.
*To whom correspondence should be addressed: Olga V. Olinevich,Department of Plant Physiology, Kazan State University,Kremlyovskaya str. 18, 420008, Kazan, Russia; e-mail:[email protected]
INTRODUCTION
Exogenous abscisic acid (ABA) as a cold acclim-ation agent increases the frost resistance of manyplants (Tignor, 1992; Lang et al., 1989), includingwinter wheat (Veisz et al., 1996). During the coldacclimation of winter cereals, increase in endogen-ous levels of ABA in cells has been observed by anumber of authors (Lalk and Dorffling, 1985; Veiszet al., 1996). Cold acclimation of plants correlateswith the de novo protein synthesis (Guy et al., 1990;Welbaum et al., 1997), some of these proteins beingproduced in tissues after ABA treatment (Gustaet al., 1996). This indicates that the same genes are
1065–6995/00/060365+09 $35.00/0
induced by both ABA and low temperature(Hughes and Dunn, 1996). Several reports havedescribed genes that are induced by cold stress,but these were shown not to be responsive toexogenous ABA treatments (Nordin et al. 1993;Yamaguchi-Shinozaki and Shinozaki, 1994). Fromthese findings, the existence of both ABA-independent and ABA-dependent pathways of theplant frost resistance increase is suggested (Dallaireet al., 1994; Gusta et al., 1996; Welbaum et al.,1997).
The cytoskeleton may be the sensitive systemwhich perceives signals both from ABA and lowtemperature, and the dynamic reorganization ofthe cytoskeleton may influence the frost resistanceof plants. Previously we have shown that the treat-ment of wheat seedlings with the tubulin proteinstabilizers and inhibitors changed the water status
� 2000 Academic Press

366 Cell Biology International, Vol. 24, No. 6, 2000
of plants differently and the cold acclimationcompensated the action of these compounds(Khokhlova et al., 1997). A change in the tubulincytoskeleton stability is one of the responses ofwinter cereal cells to the cold treatment (Kerrand Carter, 1990a; Pihakaski-Maunsbach andPuhakainen, 1995). Rikin et al. (1983) suggestedthat the depolymerization of microtubules (MTs) isone of the causes of cold injury, and showed thatABA pretreatment of cotton plants preventedthis process and increased their cold tolerance.Sakiyama and Shibaoka (1990) also noted theeffect of ABA on stabilization and orientation ofMTs. However, Chu et al. (1992) discovered thatthe hormone did not change MT stability of cornplants. Moreover, Jiang et al. (1996) showed thatABA-induced destruction of tubulin cytoskeletonin guard cells of Vicia. Thus the literature contain-ing contradictory data about the hormone effect onorientation and cold stability of MTs and infor-mation on ABA-induced changes in the totalamount of cytoskeletal proteins in cells during thecold acclimation of plants is lacking.
The aims of our work were to compare thecontent of the cytoskeletal and phosphorylatedproteins in the root and leaf tissues of the seedlingsof different age, and to investigate the ABA andlow temperature actions on the levels of theseproteins in different winter wheat cultivars in re-lation to frost resistance. For these purposes pro-teins were extracted from the control and treatedseedlings, separated by one dimensional electro-phoresis and immunoblotted with antibodiesagainst tubulin, actin and phosphothreonine. Apolyclonal antibody against phosphorylatedthreonine was used since it had been shownrecently to interact with phosphorylated thylakoid(Rintamaki et al., 1997) and cytoplasmic proteins(Raudaskoski et al., 1997). Recent experimentshave also suggested that the phosphorylation ofcytoskeletal or cytoskeleton associated proteinsmay play an important role in freeze tolerance ofplants (Danyluk et al., 1996).
MATERIALS AND METHODS
The cultivars and culture methods
Three winter wheat cultivars which had beenshown to differ in the frost resistance were chosenfor the investigation: Bezostaya1 (low frost resist-ant), Mironovskaya 808 (moderately frost resist-ant), and Albidum 114 (highly frost resistant). Theplants were grown in hydroculture at an illumi-
nation of 100 W/m2 and with a photoperiod of12 h. Non-acclimated plants were grown at 23�Cfor 5 or 9 days. Abscisic acid (Serva) 30 � wasadded to the 7-day-old plant roots and the plantswere grown then in ABA solution for three days.For cold acclimation, 8-day-old plants were trans-ferred to 3�C for 7 days. In the treatments, in whichthe effect of ABA together with cold acclimationwas studied, the addition of ABA was done oneday before the beginning of acclimation.
One-dimensional electrophoresis andimmunoblotting
Roots or leaves from control and treated seedlings,300 mg from each organ, were quickly transferredto liquid nitrogen and homogenized to a finepowder. After homogenization, the powder wascollected in 200 �l SDS-extraction buffer whichcontained 30 m Tris HCl, pH 8.5, 20% glycerol,2% 2-mercaptoethanol, and for the inhibition ofprotease activity 25 �g/ml leupeptin (BoehringerMannheim Biochemicals), 25 �g/ml pepstatinand 1 m PMSF (Sigma Chemical Co). Sampleswere boiled for 5 min and insoluble material wasremoved by the centrifugation in the EppendorfMicrofuge (Sigma 201 ) for 5 min at 14,000�g.For protein determination, 10 �l of sample extractwas precipitated with four volumes of cold acetoneand the solution was incubated at �20�C over-night. The precipitate was washed three times with100% and once with 80% acetone at 3�C in a coldroom, dried and dissolved in 10�l of 1 NaOHand then diluted to 800 �l with water. Proteindetermination was made with Bio-Rad’s micro-assay procedure. Samples containing 15 and 30 �gof proteins were analyzed on 10% SDS-polyacrylamide mini-gels prepared and run inBio-Rad’s Mini Protein� II dual slab cell system.After electrophoresis the polypeptides were trans-ferred to a PVDF membrane from Bio-Rad (for usewith monoclonal actin and tubulin antibodies) orto a PVDF membrane from Millipore (for usewith polyclonal phosphothreonine antibody) ina Trans-Blot apparatus (Bio-Rad Laboratories,Richmond, U.S.A.). For the immunostaining, thefilters were first incubated for 5–6 h in 1% fat freeBSA in TTBS, pH 7.5 (0.02 Tris-HCl, 0.15 NaCl, 0.1% Tween 20), after which the membraneswere exposed overnight to the monoclonal �- and�-tubulin (Amersham N.356 and N.357) or actin(Amersham N.350) antibodies diluted 1:1000 inTBS, pH 7.5, (0.02 Tris-HCl, 0.15 NaCl) con-taining 0.1% fat free BSA. For the identificationof proteins with phosphorylated threonines, the

Cell Biology International, Vol. 24, No. 6, 2000 367
membranes were exposed overnight to the poly-clonal phosphothreonine antibody (Zymed Lab-oratories, N. 61-8200) diluted 1:3000 in TBScontaining 0.1% fat free BSA. After washing (fivetimes for 5 min in TTBS), the membranes treatedwith the monoclonal �-tubulin, �-tubulin or actinantibodies were incubated for 2 h with anti-mouse IgG conjugated to alkaline phosphatase(Promega). The membranes treated with the poly-clonal phosphothreonine antibody were incubatedfor 2 h with anti-rabbit IgG conjugated to alkalinephosphatase (Caltag Laboratories). Both second-ary antibodies were diluted 1:3000 in TBS, pH 7.5containing 0.1% fat free BSA. After 5 rinses inTTBS, pH 7.5 and 3 rinses in TBS, pH 7.5, 5 mineach, membranes were treated with Chemilumines-cent Substrate Kit solution (Bio-Rad) for 5 min.Vizualisation of the proteins was obtained byexposing the membranes to X-ray films (FujifilmRX ID N. 03E310).
�
�
� � �
� � �
� � � � � �
� � �
�
� � �
Fig. 1. Effects of ABA (30 �) and cold acclimation (3�C, 7days) on the levels of the �-tubulin proteins in the roots (5–8)and leaves (1–4) of the 9-day-old seedlings of Bezostaya 1(A),Mironovskaya 808 (B), Albidum 114 (C) winter wheat. a, b:15 �g proteins in the leaf and root samples, respectively; c:30 �g proteins in the leaf samples; 1, 5: non-cold-acclimated(�ABA); 2, 6: non-cold-acclimated (+ABA); 3, 7: cold-acclimated (3�C, 7 days, �ABA); 4, 8: cold-acclimatedseedlings (3�C, 7 days, +ABA).
RESULTS
When the extracts from 9-day-old control leavesand roots containing 15�g of the proteins wereimmunoblotted with �- or �-tubulin antibodies, aclear signal was obtained from the roots (Fig. 1b;Fig. 2b) but not from the leaves (Fig. 1a; Fig. 2a).The increase in protein from 15 to 30 �g made itpossible to obtain a weak signal only from �- and�-tubulins from leaves of the low frost resistantcultivar (Fig. 1Ac; Fig. 2Ac) compared with themore frost resistant ones (Fig. 1Bc, Cc). Thismeans that, in the leaf cells of Bezostaya 1 wheat,the level of tubulin proteins is higher than inMironovskaya 808 and Albidum 114. In theMironovskaya 808 roots, the content of �- and�-tubulin was considerably higher (Fig. 1Bb;Fig. 2Bb) compared with the roots of Albidum 114and Bezostaya 1 (Fig. 1Ab and Cb; Fig. 2Cb).
During the immunoblotting of the leaf and theroot extracts of the young 5-day-old seedlingscontaining 15 �g of the protein the signal from thetubulin proteins was obtained both from roots andfrom leaves (Fig. 3Aab). The differences in theintensity of the �-tubulin signal from tissues ofboth organs were not found, but the signal wasstronger in the leaves and roots of Mironovskaya808 (Fig. 3Aab lanes 3 and 6), and weaker in theleaves and roots of Albidum 114 (Fig. 3Aab, lanes2 and 5).
During the investigation of the effects of bothABA and low temperature action on the content of�- and �-tubulin proteins in the cells of 9-day-old
seedlings, we found that ABA decreased theamount of a-tubulins in roots of three cultivars(Fig. 1Ab, Bb, Cb lanes 6 and 5) and in leaves oflow frost resistant cultivar (Fig. 1Ac lanes 2 and 1).Also the levels of �-tubulins were lower in ABAtreated roots than in control roots (Fig. 2Bb, Cblanes 6 and 5). In the cold-acclimated roots of threecultivars and in leaves of Bezostaya 1, the levels of�- and �-tubulins were similar to those in controltissues (Fig. 1Aa, Bb, Cb lanes 7 and 5; Fig. 1Aclanes 3 and 1; Fig. 2Bb, Cb lanes 7 and 5).
The idea that cold acclimation increases the levelof tubulin is supported by the higher amount of �-and �-tubulins in the ABA-treated and cold-acclimated samples than in the samples treatedwith ABA alone (Fig. 1Ab, Bb, Cb lanes 8 and 6;Fig. 2Bb, Cb lanes 8 and 6). This means that coldacclimation removes the ABA effect in roots of thevery frost resistant cultivar to the greatest extent(Fig. 1Cb lanes 8 and 6).
In difference from tubulin proteins the signalsfrom actin ones were more intensive in the leaves of9- and 5-day old seedlings (Fig. 3Ba; Fig. 4Bc, Cc)

368 Cell Biology International, Vol. 24, No. 6, 2000
�
�
� � �
� �
� � � �
� �
�
� �
� � � � � �
Fig. 2. Effects of ABA (30 �) and cold acclimation (3�C, 7days) on the levels of the �-tubulin proteins in the roots (5–8)and leaves (1–4) of the 9-day-old seedlings of Bezostaya 1(A),Mironovskaya 808 (B), Albidum 114 (C) winter wheat. a, b:15 �g proteins in the leaf and root samples, respectively; c:30 �g proteins in the leaf samples; 1, 5: non-cold-acclimated(�ABA); 2, 6: non-cold-acclimated (+ABA); 3, 7: cold-acclimated (3�C, 7 days, �ABA); 4, 8: cold-acclimatedseedlings (3�C, 7 days, +ABA).
�
� �
� � � �
� �
Fig. 3. Content of �-tubulin (A) and actin (B) proteins in roots(4–6) and leaves (1–3) of the 5-day-old seedlings of Bezostaya1 (1, 4), Mironovskaya 808 (3, 6), Albidum 114 (2, 5) winterwheat. a, b: 15 �g proteins in the leaf and root samples,respectively.
compared with the roots (Fig. 3Bb; Fig. 4Bb, Cb).The roots of Mironovskaya 808 wheat containedmore actin (Fig. 3Bb lane 6; Fig. 4Bb) than twoother cultivars (Fig. 3Bb lanes 4 and 5; Fig. 4Ab,Cb). The same result was also obtained for theleaves of young plants (Fig. 3Ba lanes 3 and 1, 2).ABA induced a decrease in actin levels in the low-and moderately-frost resistant cultivar roots only(Fig. 4Ab, Bb lanes 6 and 5), but not in the rootsof Albidum 114 (Fig. 4Cb lanes 6 and 5). In thecold-acclimated and ABA-treated and cold-acclimated roots of the cultivars Bezostaya 1 andMironovskaya 808, changes in the levels of actinwere comparable to those of tubulin (Fig. 4Ab, Bblanes 8 and 7). The cold acclimation removed theABA effect in Mironovskaya 808 to a greaterextent (Fig. 4Bb lanes 8 and 6) than in Bezostaya 1
(Fig. 4Ab lanes 8 and 6). No variation in theactin level in response to a specific treatment wasobserved in samples from leaves of 3 cultivars androots of the very frost-resistant cultivar (Fig. 4Aa,Bc, Cc, Cb).
Two to five clear bands were detected in leaves(Fig. 5Aa, Ba, Cc) and three to seven in roots in theimmunoblots with antibody against phosphothreo-nine (Fig. 5Ab, Bb, Cb). In root extracts from allcultivars the most significant band revealed by theantibodies was located below but close to the67 kDa molecular weight marker, and was there-fore called the 60 kDa-phosphoprotein (Fig. 5Ab,Bb, Cb lane 5). The signal from the 60 kDa-proteinwas more intensive in roots of the Bezostaya 1wheat (Fig. 5Ab lane 5). This polypeptide wasnot detected in leaf extract containing 15 �g pro-tein (Fig. 5Cc lane 1). After increasing theprotein concentration 30 �g in the leaf extracts,the 60 kDa-phosphoprotein was revealed by thephosphothreonine antibody in the leaves of the lowfrost resistant cultivar to the greatest extent (Fig.5Aa lane 1). By the analogy with a change of thetubulin signal, ABA treatment decreased the signalfrom the 60 kDa-phosphorylated proteins in theroot cells of three cultivars and in the leaves of lowfrost resistant cultivar only (Fig. 5Ab, Bb, Cb lanes6 and 5; Fig. 5Aa lanes 2 and 1), but cold acclim-ation removed this ABA effect (Fig. 5Ab, Bb, Cb
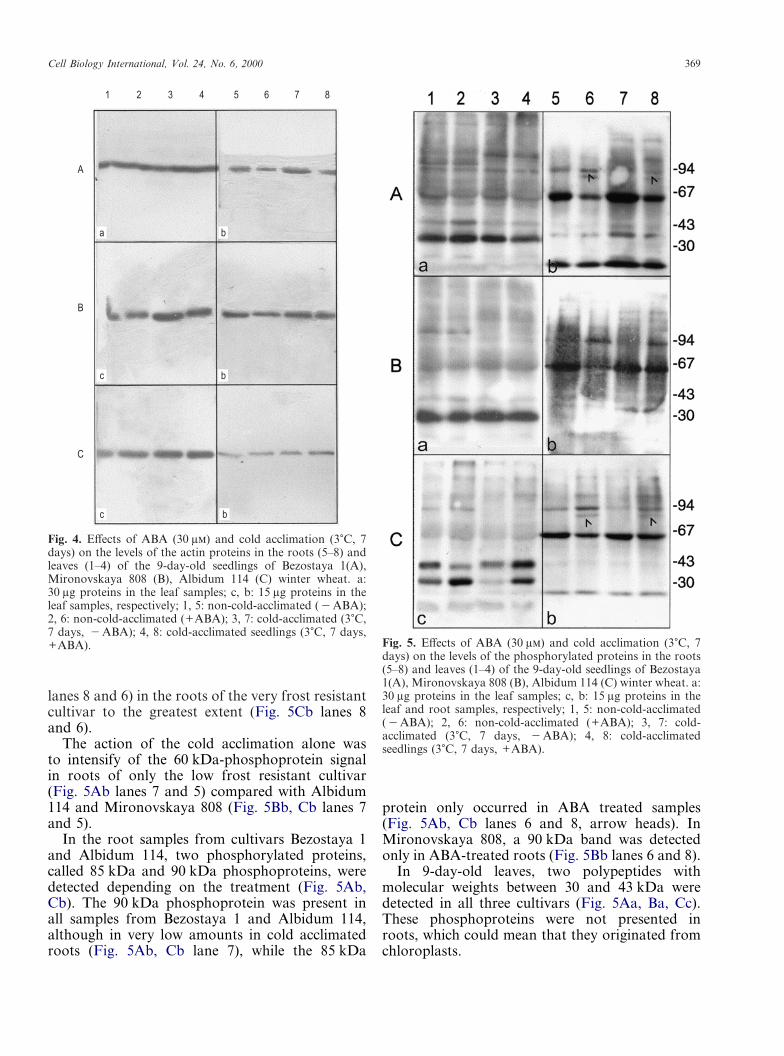
Cell Biology International, Vol. 24, No. 6, 2000 369
�
�
� � �
� �
� �
�
� �
� �
Fig. 4. Effects of ABA (30 �) and cold acclimation (3�C, 7days) on the levels of the actin proteins in the roots (5–8) andleaves (1–4) of the 9-day-old seedlings of Bezostaya 1(A),Mironovskaya 808 (B), Albidum 114 (C) winter wheat. a:30 �g proteins in the leaf samples; c, b: 15 �g proteins in theleaf samples, respectively; 1, 5: non-cold-acclimated (�ABA);2, 6: non-cold-acclimated (+ABA); 3, 7: cold-acclimated (3�C,7 days, �ABA); 4, 8: cold-acclimated seedlings (3�C, 7 days,+ABA).
Fig. 5. Effects of ABA (30 �) and cold acclimation (3�C, 7days) on the levels of the phosphorylated proteins in the roots(5–8) and leaves (1–4) of the 9-day-old seedlings of Bezostaya1(A), Mironovskaya 808 (B), Albidum 114 (C) winter wheat. a:30 �g proteins in the leaf samples; c, b: 15 �g proteins in theleaf and root samples, respectively; 1, 5: non-cold-acclimated(�ABA); 2, 6: non-cold-acclimated (+ABA); 3, 7: cold-acclimated (3�C, 7 days, �ABA); 4, 8: cold-acclimatedseedlings (3�C, 7 days, +ABA).
lanes 8 and 6) in the roots of the very frost resistantcultivar to the greatest extent (Fig. 5Cb lanes 8and 6).
The action of the cold acclimation alone wasto intensify of the 60 kDa-phosphoprotein signalin roots of only the low frost resistant cultivar(Fig. 5Ab lanes 7 and 5) compared with Albidum114 and Mironovskaya 808 (Fig. 5Bb, Cb lanes 7and 5).
In the root samples from cultivars Bezostaya 1and Albidum 114, two phosphorylated proteins,called 85 kDa and 90 kDa phosphoproteins, weredetected depending on the treatment (Fig. 5Ab,Cb). The 90 kDa phosphoprotein was present inall samples from Bezostaya 1 and Albidum 114,although in very low amounts in cold acclimatedroots (Fig. 5Ab, Cb lane 7), while the 85 kDa
protein only occurred in ABA treated samples(Fig. 5Ab, Cb lanes 6 and 8, arrow heads). InMironovskaya 808, a 90 kDa band was detectedonly in ABA-treated roots (Fig. 5Bb lanes 6 and 8).
In 9-day-old leaves, two polypeptides withmolecular weights between 30 and 43 kDa weredetected in all three cultivars (Fig. 5Aa, Ba, Cc).These phosphoproteins were not presented inroots, which could mean that they originated fromchloroplasts.

370 Cell Biology International, Vol. 24, No. 6, 2000
DISCUSSION
The absence of signals from �- and �-tubulinproteins in the leaf extracts and their presence inthe root samples provide evidence that the contentof tubulin proteins in roots of the 9-day-old seed-lings is considerably higher than in the leaves. Byincreasing the protein concentration from 15 to30 �g in the samples, the appearance of a weaksignal from tubulins of the Bezostaya 1 leavesconfirms this. Further, the latest result also indi-cates a higher amount of tubulin proteins in theleaves of the low frost resistant cultivar comparedwith more frost resistant ones.
In contrast to these data, the content of actinproteins is higher in leaves than in roots. Thecultivar peculiarities were revealed in the highercontent of both the actin proteins and the �-,�-tubulin ones in the roots of the 9-day-old seed-lings of the Mironovskaya 808 wheat. Similar datawere obtained for roots and leaves of the 5-day-oldplants. From this the conclusion is reached that theroots and the leaves are characterized by the differ-ent ratio of the tubulin and actin proteins. Alsothe leaves of the different cultivars do not have thesame ratio of the main cytoskeletal proteins in thecells. We believe that the tubulin/actin ratio deter-mines the low temperature stability of MTs intissues. In the roots the higher ratio of theseproteins (more tubulins and less actin) correspondsto the low stability of their tubulin cytoskeleton. Incontrast, in the leaves the lower tubulin/actin ratio(less tubulins and more actin) correlates with ahigher cytoskeletal stability. Apparently, in the leafcells the higher actin level than tubulin is necessaryfor the maintenance of a greater stability of thesmall tubulin cytoskeleton. Our assumption aboutthe lower microtubule stability in roots as com-pared with leaves is confirmed by the results ofPihakaski-Maunsbach and Puhakainen (1995).The authors came to the same conclusion usingthe indirect immunofluorecence microscopy for theinvestigation of the MT stability to cryostress in thewinter rye roots and leaves. In explaining ourresults, we have based much on data concerning theclose interaction between MTs and microfilaments(MFs), especially in the cortical layer of the cyto-plasm which, together with plasma membrane,forms the single structural-functional complex(Kobayashi et al., 1988). Under this, the corticalMFs may control the orientation of MTs andstabilize them (Ding et al., 1991). After detection ofthe interactions between MFs and the cold-stableMTs along the length of the latter, Mizuno (1992)concluded about the participation of MFs in the
low temperature stability of MTs. In the opinion ofthis author, the loss of MT cold stability is evokedby the separation of the actin proteins from the MTsurface. Therefore, the increase of the MT coldstability could be bound with the increase of theactin filament content and their ability to associatewith the MT surface. Using the younger 5-day-oldplants, we did not show any differences in the levelsof the �-tubulin proteins in roots and leaves. Thetubulin/actin ratio was considerably higher inleaves of the 5-day-old seedlings compared with9-day-old ones. At the same time, the ratio of thecytoskeletal proteins was similar or lower slightlyin the roots of the 5-day-old plants than in the9-day-old roots. We assume that the content ofthe tubulin proteins decreased with the increaseof the leaf age, but the MT stability increased,whereas in the roots the amount of MTs and theirquality changed little. Such explanation is con-firmed by the results of the Joyce et al. (1992) inmaize, who showed that the abundance of the�-tubulin transcripts is lower in mature leaf tissueand higher in root tips, and that the transcripts ofall �-tubulin isotypes were reduced when the leafmatured.
The result of the lowest tubulin/actin ratio inroots and leaves of the young plants of Albidum114 compared with Bezostaya 1 and Mironovskaya808 supports our assumption that a low amount oftubulins and a high amount of actin determines thehigh frost resistance of cells.
The ABA treatment of seedlings decreased thecontent of the �- and �-tubulin proteins in roots ofthree cultivars and leaves of low frost resistantcultivar. This could be a consequence of thedecrease in the cytoskeletal protein synthesis in thecells. Such suggestion agrees with the reports aboutthe reduction of seedling length in wheat plants orfresh weight and protein synthesis in wild-typeArabidopsis plants treated with ABA (Finkelsteinand Somerville, 1990; Dallaire et al., 1994). Thedestructive action of ABA on the cytoskeleton isnot excepted unsupported, but is confirmed byJiang et al. (1996), who observed destruction ofMTs in Vicia guard cells treated by the hormone.Similar to the effects of ABA on the tubulins, thehormonal action on actin levels was shown in rootsof low- and moderately-frost resistant cultivarsonly. Apparently, in leaves of three cultivars and inroots of Albidum 114, the actin proteins were notsensitive to the treatments used, from which weconclude that the higher actin stability comparedwith tubulins accords with data in the literature(Chu et al., 1993). Furthermore, it is possible thatthe MF stability in root cells of the very frost

Cell Biology International, Vol. 24, No. 6, 2000 371
resistant cultivar is higher than in cells of theseorgans of the less frost-resistant cultivars. It couldbe the consequence of the cytoskeleton-membranecomplex existence seen best in Albidum 114 cells.
In several plants, cold acclimation is corre-lated with the synthesis of new proteins which areassumed to play a role in the increase of thefreezing tolerance (Welbaum et al., 1997). In thepresent study, the levels of tubulins and actin incold acclimated wheat roots were comparable tothose in control plants. In the plants treated withABA and cold acclimated the levels of tubulins andactin in the roots were higher than in the rootstreated with ABA alone. Therefore, we concludethat the effect of the cold acclimation on thecytoskeletal proteins is revealed during the changeof the hormonal status of cells only. Possibly theprotein synthesis induced by cold acclimationcompensated for the decrease induced by ABA.The increase in the cytoskeletal protein contentobserved in cold acclimation indicates that, inroots, tubulins and actin belong to the proteins, thesynthesis of which is induced by cold acclimationperhaps through expression of new isotypes oftubulin or actin in winter rye roots (Kerr andCarter, 1990b). As shown with the �-tubulin pro-tein immunoblots, low temperature treatment com-pensated most for the ABA effect in roots of thevery frost resistant cultivar. Possibly Albidum 114root cells have the best mechanisms for low-temperature regulation of tubulin synthesis andtheir qualitative composition.
We began the investigation of the content of thephosphorylated proteins in cells because it wasshown that phosphorylation and dephosphoryl-ation of molecules which interacted with the micro-tubules influenced on the microtubule dynamicturnover rate. In plants, characterization of micro-tubule associated proteins has only just started(Chang-Jie and Sonobe, 1993; Rutten et al., 1997),and their phosphorylation stage is unknown. How-ever, inhibitors of protein kinases and phosphataseshave already been shown to have an effect on theMT orientation and stabilization in plant cells(Baskin and Wilson, 1997). This suggested that inplant cells, as in animal cells, phosphorylation andthe dephosphorylation of microtubule associatedproteins affects the microtubule turnover rates.Recently it was shown that in plant cell extracts,protein phosphatase activity was strongly decreasedwhile individual protein kinases were differentiallyreduced and some of the kinases even retainedsubstantial activity at low temperature (Monroyet al., 1997). As a consequence the phosphorylationlevel of a 58-kDa protein increased sharply when
the temperature of the reaction mixture fell below12�C in an in vitro phosphorylation experiment withplant cell extract (Monroy et al., 1997).
It is interesting, therefore, that in the frost-sensitive winter wheat cultivar Bezostaya 1, thesignal for phosphorylation—especially of the60 kDa protein—is more intense than in the otherless frost sensitive cultivars. Also in the cold-sensitive rice variety, the phosphorylation of a60-kDa protein in leaves was stimulated by coldstress (Komatsu and Kato, 1997). Differentsensitivity of phosphatases and kinases to lowtemperature could cause hyperphosphorylation ofmicrotubule associated proteins. Phosphorylationof microtubule associated proteins is known toprevent their interaction with MTs (McNally,1996) and hyperphosphorylation may lead to un-regulated turnover of MT cytoskeleton. This inturn could cause permanent defects in subcellularorganization in frost-sensitive plant cultivars, suchas Bezostaya 1.
The result of the similar tubulin changes causedby the action of ABA and cold acclimation may bethat the 60 kDa-phosphorylated protein belongs toa group of proteins associating with MTs. ABAdecreased the level of 60 kDa proteins in roots ofthree cultivars and leaves of low frost resistantcultivar, but the cold acclimation compensated forthis ABA effect in roots of the very frost resistantcultivar in the greatest extent. Only during theincrease of the protein concentration from 15 to30 �g similar to the tubulin proteins, could weinvestigate the action of the hormone and lowtemperature on the 60 kDa-phosphorylated proteincontent in leaves. Evidently, the result of the ABA-induced reduction of the content of the maincytoskeletal and 60 kDa-phosphorylated proteins isdetermined by the creation of too high a con-centration of the endogenous hormone in roots.According to the Welbaum et al. (1997), ABAcontent is higher in roots than in leaves. It ispossible that the additional treatment of roots withABA breaks the hormonal balance in cells andconsequently decreases the level or the synthesisof the cytoskeletal and 60 kDa-phosphorylatedproteins.
During investigation of the differences in thetissue responses of three cultivars on the ABAtreatment, we supposed that in roots of all cultivarsand leaves of low frost resistant cultivar the syn-thesis of the phosphorylated proteins was moresensitive to the exogenous ABA, compared withleaves of moderately and very frost resistantcultivars—where we did not observe ABA-inducedreduction in protein synthesis. This indicates the

372 Cell Biology International, Vol. 24, No. 6, 2000
weak stability of three cultivar roots and Bezostaya1 leaves to the low temperature.
In spite of general reduction of the cytoskeletaland 60 kDa-phosphorylated protein levels in rootsof three cultivars and Bezostaya 1 leaves exposed toABA, an 85 kDa phosphoprotein was induced byABA treatment in the root cells of Albidum 114and Bezostaya 1 and a 90 kDa protein in the rootcells of Mironovskaya 808. In Picea abies, ABAtreatment induces proteins to individual embryoniccell lines (Sabala et al., 1996). One of these proteinswas shown to secrete in growth medium and tohave amino acid structure of the antifreeze pro-teins. In wheat seedlings, the absence of the 85 and90 kDa phosphoproteins from cold-acclimatedroots and their presence in ABA-treated plus cold-acclimated roots supported the idea that the ex-pression of these proteins or their phosphorylationwas a response to ABA treatment. The data ofHong et al. (1997) also favour this conclusion. Theauthors identified a rapidly induced receptor-likeprotein kinase gene by the exogenous ABA treat-ment in Arabidopsis thaliana. The absence of theABA-induced creation of the phosphorylatedproteins in leaves in contrast to roots may indicatethe higher sensitivity of the latter to changes inhormone balance.
The high level of the phosphorylated changesinduced by ABA in cells indicates that the hormoneeffect on cytoskeletal protein content and possiblyon MT-associated proteins had occurred viachanges in intracellular Ca2+. Cramer and Jones(1996) showed that a concentration-dependentABA effect could increase or decrease the amountof this ion in the Arabidopsis roots. Ca2+ is asecond messenger in signal transduction in a var-iety of organisms, including plants (Gilroy et al.,1991). This ion is involved in the regulationof protein phosphorylation (Veluthambi andPoovaiah, 1984), and in changes in cytoskeletalstructure stability (Cyr, 1991). Calcium ions playan important role in regulating cold-induced geneexpression and the development of freezing toler-ance (Monroy et al., 1993). However, high Ca2+
concentrations can disrupt MT structures (Cyr,1991). Apparently, cold acclimation regulates hor-mone and ion balance and restores the levels of thecytoskeletal and 60 kDa-phosphorylated proteinsin roots of the very frost resistant cultivar in thegreatest extent.
We have shown organ-specific differences inthe composition and the content of the cyto-skeletal and phosphorylated proteins, and in theirresponses to ABA and low temperature. We sug-gest that in the root tissue of three cultivars and
in leaves of low frost resistant cultivar the hightubulin and 60 kDa-phosphorylated protein/actinratio determines the low stability of their tubulincytoskeleton. The last might be the reason for theABA-induced reduction of the tubulin and 60 kDa-phosphorylated protein amount or the decrease oftheir synthesis in cells. In contrast, the lower ratioof the tubulin and 60 kDa-phosphorylated protein/actin determines the higher stability of the MTstructures and their lower sensitivity to the ex-ogenous hormone in leaf cells compared with roots.Apparently, during the increase of the plant agefrom 5 to 9 days the high ratio of these proteinsdecreases, indicating the considerable lability of theyoung seedling tubulin cytoskeleton compared withmore old plants. In comparison with ABA action,cold acclimation increases the synthesis of the maincytoskeletal and 60 kDa-phosphorylated proteins,and possibly low temperature assists in microtubulestability in root cells treated with ABA. Thissuggests the existence of different pathways ofincreasing plant cell frost resistance through theaction of ABA and low temperature.
REFERENCES
B T-I, W J, 1992. Inhibitors of protein kinasesand phosphatases alter root morphology and disorganizecortical microtubules. Plant Physiol 113: 493–502.
C-J J, S S, 1993. Identification and prelimi-nary characterization of a 65 kDa higher-plant microtubule-associated protein. J Cell Sci 105: 891–901.
C B, X Z, L P, C J, 1992. Depolymerization ofcortical microtubules is not a primary cause of chillinginjury in corn (Zea mays L. cv. Black Mexican Sweet)suspension culture cells. Plant Cell Environ 15: 307–312.
C B, S P, C J, 1993. Alteration of �-tubulingene expression during low-temperature exposure in leavesof Arabidopsis thaliana. Plant Physiol 103: 371–377.
C GR, J R, 1996. Osmotic stress and abscisic acidreduce cytosolic calcium activities in roots of Arabidopsisthaliana. Plant Cell and Envir 19: 1291–1298.
C RJ, 1991. Calcium-calmodulin affects microtubulestability in lysed protoplasts. J Cell Sci 100: 311–317.
D S, H M, G Y, S H, B S,C N, S F, 1994. ABA and low temperatureinduce freezing tolerance via distinct regulatory pathways inwheat. Plant Cell Physiol 35: 1–9.
D J, C E, S F, 1996. Identificationand characterization of a low temperature regulated geneencoding an actin-binding protein from wheat. FEBS Lett389: 324–327.
D A, T R, P M, 1991. Microfila-ment organization and distribution in freeze substitutedtobacco plant tissues. Protoplasma 165: 96–105.
F R-R, S C, 1990. Three classes ofabcsisic acid (ABA)-insensitive mutations of Arabidopsisdefine genes that control overlapping subsets of ABAresponses. Plant Physiol 94: 1172–1179.

Cell Biology International, Vol. 24, No. 6, 2000 373
G S, F MD, R ND, T AJ, 1991. Roleof calcium in signal transduction in Commelina guard cells.Plant Cell 3: 333–344.
G LV, W R, F P, 1996. Low-temperature stresstolerance: The role of abscisic acid, sugars, and heat-stableproteins. Hort Science 31: 39–46.
G CL, 1990. Cold acclimation and freezing stress tolerance:role of protein metabolism. Ann Rev Plant Physiol Plant MolBiol 41: 187–223.
H SW, L JH, K JM, N HG, 1997. Identificationof a receptor-like protein kinase gene rapidly induced byabscisic acid, dehydration, high salt, and cold treatments inArabidopsis thaliana. Plant Physiol 113: 1203–1212.
H MA, D M, 1996. The molecular biology of plantacclimation to low temperature. J Exper Bot 47: 291–305.
J CJ, N N, K N, 1996. Disruption ofmicrotubules by abscisic acid in guard cells of Vicia faba L.Plant Cell Physiol 37: 697–701.
J CM, V R, S D, S C, 1992.Tubulin gene expression in maize (Zea mays L.). Change inisotype expression along the developmental axis of seedlingroot. J Mol Biol 227: 97–107.
K GP, C J, 1990a. Relationship between freezingtolerance of root-tip cells and cold stability of microtubulesin rye (Secale cereale L. cv. Puma). Plant Physiol 93: 77–82.
K GP, C J, 1990b. Tubulin isotypes in rye roots arealtered during cold acclimation. Plant Physiol 93: 83–88.
K L, O O, P E, T N,A E, V I, 1997. Effect of tubulin proteinmodifiers on the water exchange of non-hardened andcold-hardened plants of winter wheat. Acta Agro Hungarica45: 377–389.
K H, F H, S H, 1988. Interrelationbetween the spatial disposition of actin filaments and micro-tubules during the differentiation of tracheary elements incultured Zinnia cells. Protoplasma 143: 29–37.
K S, K A, 1997. Varietal differences in proteinphosphorylation during cold treatment of rice leaves.Phytochemistry 45: 1329–1335.
L I, D K, 1985. Hardening, abscisic acid, prolineand freezing resistance in two winter wheat varieties. PhysiolPlant 63: 287–292.
L V, H P, P E, 1989. Low temperature accli-mation and treatment with exogenous abscisic acid inducecommon polypeptides in Arabidopsis thaliana (L.). Theorand Applied Genetics 77: 729–734.
MN F, 1996. Modulation of microtubule dynamicsduring the cell cycle. Curr Op Cell Biol 8: 23–29.
M K, 1993. Induction of cold stability of microtubules incultured tobacco cells. Plant Physiol 100: 740–748.
M A-F, S F, D R, 1993. Cold-inducedchanges in freezing tolerance, protein phosphorylation, andgene expression. Plant Physiol 102: 1227–1235.
M AF, L E, D R, 1997. Low temperatureperception in plants: effect of cold on protein phos-
phorylation in cell-free extracts. FEBS Letters 410:206–209.
N K, V T, P E, 1993. Differential expres-sion of two related, low-temperature-induced genes inArabidopsis thaliana kin 1 gene. Plant Physiol 98:1532–1534.
P-M K, P T, 1995. Effectof cold exposure on cortical microtubules of rye (Secalecereale) as observed by immunocytochemistry. PhysiolPlantarum 93: 563–571.
R M, T M, A H, M H,L E, 1997. The regulation of pollen tube cyto-skeleton by protein phosphorylation and dephosphoryl-ation. 12th Meeting of European Cytoskeletal Forum.Euroconference n. ERBFMMACT950040. p. 131.
R A, A D, G C, 1983. Quantitation of chill-induced release of a tubulin-like factor and its prevention byabscisic acid in Gossypium hisutum L. Plant Physiol 71:747–748.
R E, S M, S U, C I,A B, A E, 1997. Phosphorylation of light-harvesting complex II and photosystem II core proteinsshows different irradiance-dependent regulation in vivo.J Biol Chem 272: 30,476–30,482.
R T, C J, L C, 1997. A 60-kDa plantmictotubule-associated protein promotes the growth andstabilization of neurotubules in vitro. Proc Natl Acad SciUSA 94: 4469–4474.
S I, E U, F H, A S, 1996.Abscisic acid-induced secretion of an antifreeze-like proteinin embryogenic cell lines of Picea abies. J Plant Physiol 149:163–170.
S M, S H, 1990. Effect of abscisic acid on theorientation and cold stability of cortical microtubulesin epicotyl cells of the dwarf pea. Protoplasma 157: 165–171.
T ME, 1992. The effect of exogenous abscisic acid anda,a�-dipyridyl on cold acclimation and physical characteris-tics of Pisum sativum ‘Alaska’ seedlings. MS Thesis Depart-ment of Horticulture, Virginia Tech, Blacksburg, VA,U.S.A.
V O, G G, S J, 1996. Effect of abscisic acid onthe cold hardiness of wheat seedlings. J Plant Physiol 149:439–443.
V K, P B, 1984. Calcium-promoted proteinphosphorylation in plants. Science 223: 167–169.
W G-E, B D, H D, G R, GM, 1997. Freezing tolerance, protein composition, andabscsic acid localization and content of pea epicotyl, shoot,and root tissue in response to temperature and water stress.J Exper Bot 48: 643–654.
Y-S K, S K, 1994. A novelcis-acting element in an Arabidopsis gene is involved inresponsiveness to drought, low-temperature, or high-saltstress. Plant Cell 6: 251–264.




























