Embed Size (px)
Citation preview

[CANCER RESEARCH 46, 5576-5581, November 1986]
Effect of 5-Diazouracil on the Catabolism of Circulating Pyrimidines in Rat Liverand Kidneys1
Axel Holstege,2 JürgenPausch, and Wolfgang Gerok
Medizinische Klinik der UniversitätFreiburg, Hugstetter Strasse 55, D-7800 Freiburg, Federal Republic of Germany
ABSTRACT
The inhibitory effect of 5-diazouracil on the catabolism of circulatingunidl and 5-fIuorouracil was examined in the rat in vivo. Measurements
of the activity of the entire enzymatic pathway of uracil catabolism in thecytosolic supernatant of different rat organs as well as the determinationof the total amount of 5-fluorouracil catabolites, accumulated in these
tissues, served to clarify their role in the complete systemic breakdownof uracil or 5-fluorouracil. The activity of the enzymatic pathway involvedin uracil catabolism was estimated from the I4<O, produced from
|2-'4C]uracil in the cytosolic supernatants. Complete degradation of uracil
was detected only in the liver and, at a much lower rate, in the kidneys.Fifteen min after the i.p. injection of a tracer dose of 5-fluorof6-'4C|-
uracil, more than 90% of the total radioactivity in blood plasma wasassociated with 5-fluorouracil catabolites. The relative amount of themajor catabolite a-fluoro-#-alanine and of dihydrofluorouracil in bloodplasma was considerably suppressed after a pretreatment with 5-di-azouracil inversely correlated with a 27-fold increase in the absoluteamount of unchanged 5-fluorouracil.
Control animals accumulated by far the highest amount of total acid-soluble radioactivity from 5-fluoro|6-MC|uracil in liver and kidneys. Total
radioactivity in all other organs was much lower and was comparable tothe amount of label in blood plasma. In liver and kidneys, the sum oftotal acid-soluble catabolites including dihydrofluorouracil, a-fluoro-/?-ureidopropionic acid, and a-fluoro-/3-alanine made up more than 98% of
the label correlating with minimal salvage utilization of the base analoguein both organs.
Injection of 5-diazouracil 2 h before a tracer dose of 5-fluoro|6-l4C|-
uracil strongly inhibited the accumulation of labeled catabolites in liverand kidneys causing a fall in total acid-soluble radioactivity in both
tissues by 75 and 66%, respectively. In blood plasma and all other organs,however, pretreatment with 5-diazouracil was followed by a 2-fold en
hancement of the radioactivity contents, mostly due to the appearanceof unchanged 5-fluorouracil. Under these conditions, there was a 2.6- to4-fold increase in the relative proportion of cis-diol group-containinganabolites of 5-fluorouracil in liver and in kidneys.
Within 2 h, 12.7% of the administered radioactivity from 5-fluorouracilwas excreted into bile. 5-Diazouracil lowered the biliary excretion of
radioactivity to 2% of the injected dose.Our studies indicate a strong inhibition of the catabolism of 5-fluo
rouracil and uracil by 5-diazouracil in the rat in vivo. Complete degra
dation of circulating pyrimidine bases was restricted to rat liver andkidneys, with the major part occurring in liver. Inhibition of the catabolicpathway by 5-diazouracil considerably increased the bioavailability of 5-
fluorouracil
INTRODUCTION
In mammalian cells the requirements for pyrimidine nucleo-tides are satisfied by synthesis de novo as well as by nucleotideformation from preformed pyrimidines on the salvage pathway(1). Micromolar concentrations of pyrimidines in blood plasmarepresent the extracellular source for salvage nucleotide synthesis in many tissues (1-4). For a certain species, salvage activitiesare different for circulating uridine, cytidine, thymidine, uracil,
Received 6/11/85; revised 3/24/86, 7/14/86; accepted 7/16/86.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by Deutsche Forschungsgemeinschaft, SFB 154, "Klinische undexperimentelle Hepatologie."
2To whom requests for reprints should be addressed.
or pyrimidine deoxyribonucleosides and markedly vary fromone type of tissue to the other (2, 4). The chemotherapeuticefficiency of the antimetabolite FUra1 depends on its conversion
to active FUra nucleotides via the salvage pathway (5). However, the salvage utilization of uridine (2), uracil (4, 6, 7), andFUra (8, 9) often is low under in vivo conditions. Mammaliancells do not possess a specific uracil phosphoribosyltransferaseactivity allowing conversion of uracil into UMP in one enzymatic step (10). Moreover, at physiological pH, uracil in contrast to FUra is only a poor substrate for orotate phosphoribosyltransferase (EC 2.4.2.10) (10, 11). In vivo, uridine phos-phorylase (EC 2.4.2.3) appears to be primarily involved inuridine catabolism rather than in nucleoside formation fromuracil or FUra (11). Accordingly, most of the uracil originatingfrom uridine phosphorolysis or of the exogenously suppliedFUra is catabolized or excreted in urine (6-9).
The activity of the first step in enzymatic uracil or FUradegradation, dihydrouracil dehydrogenase (EC 1.3.1.2), hasbeen demonstrated in several tissues and tumors of humans(12) and rats (13-16). Dihydropyrimidinase (EC 3.5.2.2) is thesubsequent enzyme in pyrimidine base degradation and hasbeen located in liver and kidneys of rats (17). It is absent fromor showed only little activity in normal extrahepatic humantissues but was highly active in several solid tumors (12). Thelast enzyme of the catabolic pathway is /3-ureidopropionase (EC3.5.1.6) which demonstrates a similar distribution pattern likedihydropyrimidinase in both species (12, 14). In normal tissuesof the rat, the complete enzymatic equipment for uracil degradation thus was detected only in liver (12-14, 18-25) and inkidneys (12).
Many attempts have been made to improve the chemotherapeutic effectiveness of FUra by its combination with otherantitumor agents (26) or with naturally occurring pyrimidines(27-30). Protection of FUra from catabolism has been shownto be a major effect of its combination with high doses ofthymidine (31) leading in some tumors to potentiation of itsantineoplastic activity. 5-Diazouracil is an irreversible inhibitorof dihydrouracil dehydrogenase (32, 33) and has been used toincrease the incorporation of FUra into RNA of rat leukemiacells and of normal tissues (21). The aim of our study was tofurther clarify the role of rat liver and kidneys in the catabolismof FUra or uracil and to investigate the effect of a prior dose of5-diazouracil on the metabolic profile of FUra in blood plasma,on the total FUra catabolite contents in different tissues, andon the biliary excretion of radioactivity after administration of[6-14C]FUra.
MATERIALS AND METHODS
ChemicalsandIsotopes
[2-14C]Uracil (52 Ci/mol) and 5-fluoro[6-14C]uracil (56 Ci/mol) were
from Amersham Buchler (Braunschweig, Federal Republic of Germany). Radiochemical purity of the isotopes was controlled by reversedphase HPLC and by counting of the eluate fractions collected every 20
3The abbreviations used are: FUra, 5-fluorouracil; HPLC, high performanceliquid chromatography.
5576
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CATABOLISM OF CIRCULATING PYRIMIDINES
s. Purity was at least 95% for each of the isotopes. FLJra and 5-diazouracil were obtained from Sigma Chemical Co. (St. Louis, MO)and from ICN Pharmaceuticals (Plainview, NY), respectively. Basesand nucleosides were purchased from Boehringer Mannheim (Mannheim, Federal Republic of Germany). a-Fluoro-/3-alanine and dihydro-fluorouracil were kindly supplied by Hoffmann-La Roche (Basel, Switzerland). All other chemicals were of the highest purity available fromE. Merck AG (Darmstadt, Federal Republic of Germany).
Preparation of Tissue Extracts
Female Wistar rats (Ivanovas, Kisslegg, Federal Republic of Germany), weighing 150 to 180 g, had free access to water and a carbohydrate-rich 20% protein diet (Altromin, Lage, Federal Republic of Germany). 5-Diazouracil (5 mg/kg body weight) (21) usually was administered i.p. 2 h before the injection of radioactive pyrimidines.
The different organs used for analysis of the complete pathway ofuracil catabolism were quickly removed under light pentobarbital anesthesia (45 mg/kg body weight) and homogenized at 4°Cwith a motor
ized homogenizer in 2 volumes of a medium containing 0.25 M sucrose,20 mM Tris-HCl (pH 7.4, at 37°C),10 mM 0-mercaptoethanol, and 0.1
HIMEDTA (14). The homogenate was first centrifuged at 30,000 x gfor 30 min at 4°Cand the supernatant was recentrifuged at 105,000 xg for 1 h at 4°C.The 105,000 x g supernatant fluid (cytosol) was used
to estimate the complete catabolic pathway (14). The assay was run induplicate under conditions where the activity was linear with time andenzyme concentration. The complete degradation of uracil was measured by the production of I4CO2 from [2-'4C]uracil (0.5 MM;0.5 /iCi/
ml) (14) using a procedure for quantitating labeled < <>,which has beendescribed recently (24). The reaction was stopped by the addition of0.1 ml 2.5 M perchloric acid. Incubation at 37°Cwas continued for 45
min before the radioactivity trapped in 1 ml l MKOH was determined.Determination of protein concentration was carried out by the methodof Bradford (34) using 7-globulin as a standard.
For metabolic studies, the different rat organs were freeze-clamped,extracted with cold perchloric acid, and neutralized with KHCOs asdescribed (35). Aliquots of the homogenate or the acid-soluble supernatant were counted in Instant Scint Gel (Packard Instrument Company, Downers Grove, IL) at an efficiency of 79%. The neutralizedacid-soluble supernatants were used for metabolite analyses. Blood waswithdrawn from the portal vein with heparinized syringes, immediatelyfollowed by centrifugation and ultrafiltration through Amicon MPSIfilters (Amicon, Witten, Federal Republic of Germany).
Phenylboronate Affinity Chromatography. Phenylboronate affinitychromatography allowed the separation of pyrimidine bases and theircatabolites from anabolites present in the neutralized acid-soluble supernatants of different tissues after i.p. administration of radioactiveuracil or FUra. Anabolites like ribonucleosides and ribonucleotidescontain ci's-diol groups allowing their selective binding to Affi-Gel 601
(Bio-Rad Laboratories, Richmond, CA) (36). Uracil, FUra, and theircatabolites as well as the recently discovered 5-fluorouracil glucuronide(37, 38) lack such a group and do not bind to the gel. Phenylboronateaffinity chromatography of a mixture of 500 to 900 n\ of the acid-soluble supernatant of the different organs and 2.5 M ammoniumacetate (pH 8.8) (10%, v/v) was performed as described (3, 36). Theeluate containing bases, deoxynucleosides, catabolites, and possibly the5-fluorouracil conjugate (37, 38) was collected from the column including a subsequent wash with 10 ml 0.25 Mammonium acetate (pH 8.8).Elution of c/s-diol group-containing anabolites was performed with 6ml of 0. l M formic acid. The eluates were lyophilized to dryness, andthe residue was dissolved in 200 ;/l (formic acid fraction) or 300 n\(ammonium acetate fraction) 0.01 M NH4H2PO4 adjusted to pH 5.Aliquots of both fractions were counted. Dihydrofluorouracil was completely converted into «-fluoro-/î-alanineunder the conditions used forpreparation of the tissue extracts as was tested by the inclusion offurther amounts of blood plasma containing labeled FUra catabolitesto the organ samples before the homogenization procedure. In thedifferent organs, a-fiuoro-/3-alanine thus represented the sum of totalacid-soluble FUra catabolites including dihydrofluorouracil, a-fluoro-0-ureidopropionic acid, and a-fluoro-if-alanine.
Reversed Phase High Performance Liquid Chromatography. Subse
quent to phenylboronate affinity chromatography, 50 n\ of the lyophilized and reconstituted ammonium acetate fraction were analyzed byreversed phase HPLC using a procedure described for the analysis ofribonucleosides (3, 36). The sample was injected onto two Cig-^Bon-dapak columns (600 x 3.9 mm) connected in series with a Guard-PAKprecolumn module containing a C,8-MBondapak insert (Waters, Kö
nigstein, Federal Republic of Germany). The eluent was composed of0.01 MNH4H2PO4 adjusted to pH 5 and 6% methanol (v/v). The flowrate was 1 ml/min at 35°C.The isocratic HPLC system was from Du
Pont De Nemours (Bad Nauheim, Federal Republic of Germany).Fractions of the eluate were collected every 10 or 20 s and counted inInstant Scint Gel. Recovery of FUra after affinity chromatography andHPLC was 85%.
Blood plasma was deproteinized by ultrafiltration and directly injected onto the HPLC columns allowing the separation of labeled 5-fluorouracil, dihydrofluorouracil, a-fluoro-/3-alanine, and possibly a-fluoro-/3-ureidopropionate associated with a fourth labeled minor peak.Except for the latter peak, standards were available which demonstratedidentical retention times as compared to the labeled catabolites, whenUV absorbance was recorded at 200 nm. Under the conditions definedabove, retention times of the unlabeled markers, a-fluoro-/3-alanine,dihydrofluorouracil, and FUra were 5.51 ±0.06, 7.48 ±0.07, and 8.25±0.08 min, respectively (mean of 5 separations ±SD).
Collection of Bile
The peritoneal cavity of female Wistar rats (240 ±32 g) was openedafter tracheotomy under pentobarbital anesthesia. A Teflon cannula(outer diameter, 0.9 mm; inner diameter, 0.5 mm) was introduced intothe bile duct. Bile was collected over 10-min periods for up to 2 h after[6-14C]FUra (8 ^Ci/250 g rat) had been injected into the superior
mesenteric artery. The samples were weighed and counted. Bile flowremained constant in either the presence (131 ±54 mg/10 min) or theabsence (148 ±35 mg/10 min) of a prior dose of 5-diazouracil (5 mg/
kg).
RESULTS
Localization of the Entire Pathway of Uracil Catabolism. Theactivity of the entire pathway of uracil catabolism includingdihydrouracil dehydrogenase, dihydropyrimidinase, and ß-ureidopropionase was measured in the cytosol of spleen, lungs,thymus, brain, intestine, kidney, and liver via the formation ofradioactive CO2 from [2-14C]uracil (14). The highest activity of
the pathway was found in the liver of rats [52.8 ±3.8 nmolCO2 x (mg protein)'1 x h~']. The kidneys possessed 10% of
the catabolic activity detected in the liver (5.3 ±0.9 nmol CO2x (mg protein)"1 x h~'). All other tissues tested did not possess
the complete degradative pathway of uracil leading to therelease of CO2.
Effect of 5-Diazouracil on the Metabolite Pattern of LabeledFUra in Blood Plasma. Total radioactivity in blood plasma 15min after a tracer dose of [6-MC]FUra was increased 1.7-foldafter pretreatment of the rats with 5-diazouracil as comparedto control rats (Table 1). Two h after FUra (100 ^mol/kg; 0.26Ci/mol) there was still a 2-fold rise in total radioactivity whenthe animals had received a prior dose of 5-diazouracil. Catabolism of [2-'4C]uracil does not lead to a labeling of 0-alanine.
Reversed phase HPLC of deproteinized blood plasma revealedthat the 5-fold increase in total radioactivity after administration of [2-14C]uracil (15 ¿/mol/kg)and inhibition of uracil deg
radation (Table 1) was exclusively due to the presence of unchanged uracil (data not shown).
Fifteen min after injection of FUra 75.9 ±19%(n = 4) of thetotal radioactivity in blood plasma was associated with <>fluoro.;•¡ilaniiifversus 7.6 ±0.7% after 5-diazouracil pretreatment(Fig. 1). This inhibition of FUra catabolism was reflected bythe increase in the relative amount of unchanged FUra in
5577
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CATABOLISM OF CIRCULATING PYRIMIDINES
Table 1 Effect of 5-diazouracil on the elimination ofpyrimidine-derivedradioactif ity from rat blood plasma
|6-MC]FUra (35 ^Ci/kg; 56 Ci/mol, t = 15 min; or 26 (¿Ci/kg,0.26 Ci/mol, /= 2 h) or [2-14C]uracil (15 »imol/kg;3 Ci/mol) was injected i.p. Some of the ratswere pretreated with 5-diazouracil 2 h before administration of the labeledp>rimulmc. Total radioactivity was determined in heparinized and deproteinizedblood plasma.
Radioactivity (^Ci/liter)
Pyrimidine[6-14C]FUra(t = 15 min)"
[6-'4C]FUra (1 = 2 h)"[2-14C]Uracil (t = 2 hfNo
pretreatment19.0±2.8*
6.4 ±0.92.8 ±0.45+
5-Diazouracil32.8
±3.812.7 ±1.114.6 ±1.3
" Time interval between pyrimidine injection and blood withdrawal.* Mean ±SD from 3-6 rats.
l3StzsgÌ2C'5£££
1^yfi1it0BLOOD
PLASMAFUro.
(.5-DiOïoUrà )FBALM^{fïo™1-•L.
JL_^^^"*,9™°°^o•iRAT
LIVÇRFBAL(CONTROL)(^5-OiazoUrò)
.FBAL-ieo§¿3£K|"ai4051320a<Xs0
0.KDNEYFBAL(CONTROL)US-DözoUra)FuraFBAL
ïliUjjMsINTESTMFUra/"
\US-OoioUraJFB,—U..,CONTROL)FUTO
FBALf?11
iP-LJÕ-LV4T8 4 8 4 8 "t 8 4 8 " 4 8 4 8 " 48RETENTION
TIME (mn) RETENTION TIME (min)
Fig. 1. Effect of 5-diazouracil (5-DiazoVra) on the metabolite pattern of [6-'4C]FUra in rat plasma, liver, kidney, and intestine. Portal vein blood wasdeproteinized by ultrafiltration and used for reversed phase HPLC (36). FUraand its catabolites were purified from the acid-soluble supernatant of the organsby boronate affinity gel chromatography and separated by reversed phase HPLC.The eluate from the columns was collected every 10 or 20 s and counted. Similarchromatograms were obtained from 4 rats. The value given on the ordinate mustbe multiplied by IO3 (left) or IO2 (right). FBAL, a-fluoro-0-alanine; FH2Ura,
dihydrofluorouracil. Arrow, retention time of a minor radioactive peak presumablyrepresenting a-fluoro-/3-ureidopropionic acid.
control blood plasma from 5.1 ±1.4% (n = 4) to 88.4 ±15.6%(n = 4) after 5-diazouracil. The absolute amount of the basewas increased by a factor of 27 in pretreated rats (Table 1).Dihydrofluorouracil amounted to 15.1 ± 5.0% in controlplasma and 2.9 ±0.7% in pretreated rats (Fig. 1). Anotherminor radioactive peak which eluted between the two otherFUra catabolites from the HPLC columns possibly representeda-fluoro-/3-ureidopropionic acid and contained 3.9 ±2.2 and1.1 ±0.2% in control and pretreated animals, respectively. Twoh after FUra (100 /¿mol/kg)the unchanged base was no longerdetectable in control blood plasma whereas 83.4 ±5.4% of theradioactivity in blood plasma of pretreated animals was associated with intact FUra.
Effect of 5-Diazouracil on the Sum of Total FUra Catabolitesin Different Organs. Total acid-soluble radioactivity was measured in 6 different rat organs 15 min after i.p. administrationof [6-'4C]FUra (Table 2). Liver and kidneys accumulated by far
most of the radioactivity injected. Spleen, intestine, lungs, andthymus contained similar amounts of label as were found inblood plasma (Tables 1 and 2). 5-Diazouracil pretreatmentconsiderably suppressed the accumulation of acid-soluble radioactivity derived from [6-l4C]FUra to 25 and 33% of control
values in liver and kidneys, respectively. In all other tissues,however, inhibition of pyrimidine catabolism was followed bya 2-fold increase in the label of the acid-soluble fraction.
As compared to the amount of FUra catabolized, conversion
of FUra into nucleosides or nucleotides via the salvage pathwaywas almost negligible under the conditions of our experiments.Measurable amounts of cw-diol group-containing acid-solubleanabolites were detected only in liver and kidneys. Pretreatmentof the animals with 5-diazouracil increased the relative proportion of this anabolite fraction in the acid-soluble pool from 0.56±0.22 to 2.23 ±0.37% in liver and from 1.51 ±0.09 to 3.9 ±0.54% in the kidneys. In the other organs tested, the radioactivity in FUra anabolites binding to the phenylboronate gel wasbelow or close to the detection limit of our methods. In thedifferent tissues, a-fluoro-ß-alanine represented the sum of totalacid-soluble catabolites, including dihydrofluorouracil, a-flu-oro-0-ureidopropionic acid, and a-fluoro-/3-alanine, due to theinstability of the former two compounds under our experimental conditions. Accordingly, it was the only catabolite whichcould be measured in the organs (Fig. 1). The 5-diazouracil-induced increase in total acid-soluble radioactivity of spleen,intestine, lungs, and thymus was caused by a strong rise in therelative and absolute amount of unchanged FUra as indicatedin Table 2. In the absence of the inhibitor, intact FUra couldnot be detected in any organ except for spleen and intestine.The latter two organs could have been contaminated by ¡.p.FUra. The fall in total acid-soluble radioactivity of liver andkidneys after 5-diazouracil resulted from a severe depletion of5-fluorouracil catabolites paralleled by the appearance of theunchanged base in both organs (Fig. 1; Table 2).
In addition to the experiments with FUra, we investigatedthe changes in [2-14C]uracil metabolism in 5 different organs 2
h after administration of the labeled pyrimidine (Table 3). 5-Diazouracil prevented the loss of label via exhalation of radioactive CO2 and caused an increase in total radioactivity of liverand kidneys including the label in unchanged uracil, in acid-soluble nucleotides, and in nucleic acids.
Excretion of Radioactivity from [6-14C]FUra into Bile. Theadministration of [6-'4C]FUra to rats was followed by the
appearance of increasing amounts of radioactivity in bile duringthe first 20 min after injection of the label (Fig. 2). At latertime points the rate of biliary excretion of labeled metabolitesslowly declined. Within 2 h 12.7% of the injected radioactivitywas collected in bile. Prior inhibition of pyrimidine catabolismby 5-diazouracil considerably suppressed the label in bile to 2%of the administered dose.
DISCUSSION
FUra has been shown to be rapidly catabolized in the wholeanimal in vivo (8) and in isolated hepatocytes (23). Up to 50%of a hepatic arterial infusion (39) or 67% of an i.p. dose (40) ofFUra is extracted in a single pass by the human liver. Fromthese and other studies (6-8, 12-15, 18-25) it was concludedthat the liver plays a key role in the degradation of uracilpyrimidines. The results from our studies in rats in vivo afteradministration of a tracer dose of [6-14C]FUra support the
assumption that the liver is the primary site in FUra catabolism.As early as 15 min after injection of the label, total acid-solubleradioactivity in the liver accumulated to 440 nCi/kg organ wetweight (Table 2) with more than 99% of the label associatedwith FUra catabolites (Table 2; Fig. 2). This amount representsabout one-half of the total dose administered if one assumes aliver wet weight of 6-7 g in female Wistar rats weighing 160 g.Our experiments in rats in vivo further demonstrate that thekidneys also accumulate FUra catabolites amounting to 275/¿Ci/kg,wet weight. This is in line with the presence of thecomplete enzymatic pathway for uracil catabolism in rat kid
5578
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
CATABOLISM OF CIRCULATING PYRIMIDINES
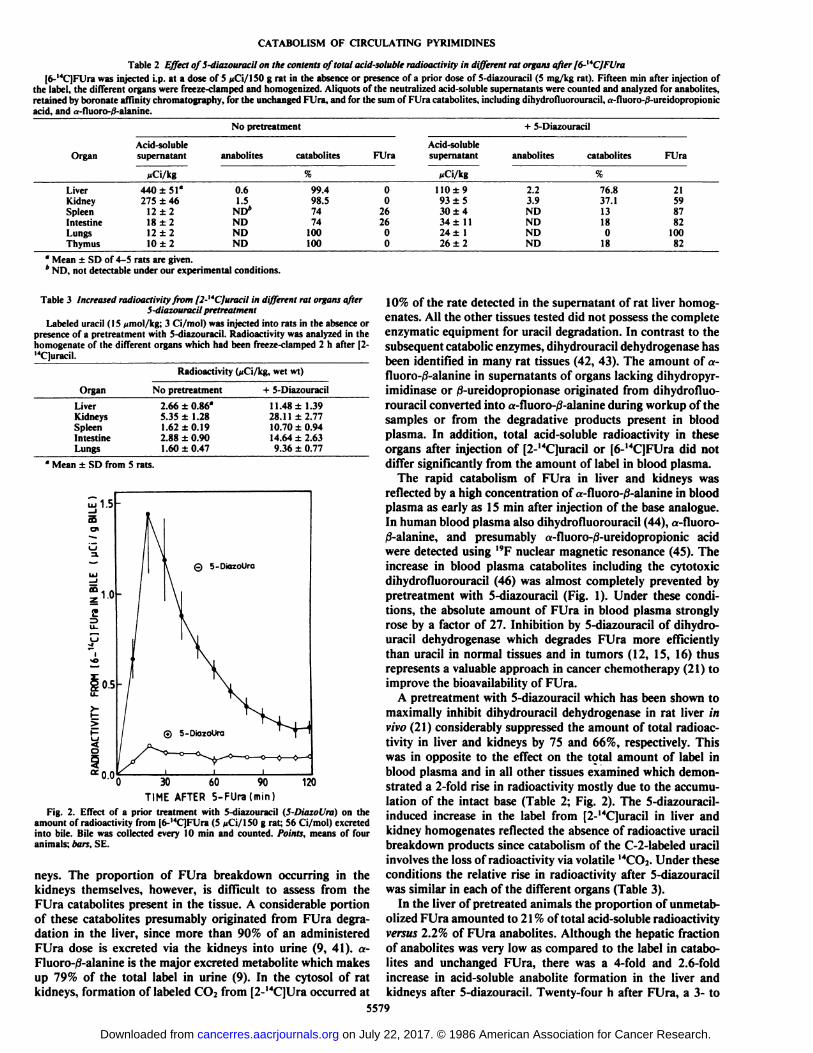
Table 2 Effect of 5-diazouracil on the contents of total acid-soluble radioactivity in different rat organs after f6-"C]FUra[6-'4C]FUra was injected i.p. at a dose of 5 jiCi/150 g rat in the absence or presence of a prior dose of 5-diazouracil (5 mg/kg rat). Fifteen min after injection of
the label, the different organs were freeze-clamped and homogenized. Aliquots of the neutralized acid-soluble supernatants were counted and analyzed for anabolites,retained by boronate affinity chromatography, for the unchanged FUra, and for the sum of FUra catabolites, including dihydrofiuorouracil, a-fluoro-/3-ureidopropionicacid, and a-fluoro-0-alanine.
NopretreatmentOrganLiver
KidneySpleenIntestineLungsThymusAcid-soluble
supernatantMCi/kg440
±51*
275 ±4612±218 ±212 ±210 ±2anabolites0.6
1.5ND*
NDNDNDcatabolites%99.4
98.57474
100100FUra0
02626
00Acid-soluble
supernatant^Ci/kg110±9
93 ±530 ±434 ±1124 ±126 ±2+
5-Diazouracilanabolites2.2
3.9NDNDNDNDcatabolites%76.8
37.113180
18FUra21
598782
10082
°Mean ±SD of 4-5 rats are given." ND, not detectable under our experimental conditions.
Table 3 Increased radioactivity from [2-"CJuracil in different rat organs after5-diazouracil pretreatment
Labeled uracil (15 ^mol/kg; 3 Ci/mol) was injected into rats in the absence orpresence of a pretreatment with 5-diazouracil. Radioactivity was analyzed in thehomogenate of the different organs which had been freeze-clamped 2 h after [2-"Cluracil.
Radioactivity (jiCi/kg, wetwt)OrganLiver
KidneysSpleenIntestineLungsNo
pretreatment2.66±0.86°
5.35 ±1.281.62 ±0.192.88 ±0.901.60 ±0.47+
5-Diazouracil11.48±
1.3928.11±2.77
10.70 ±0.9414.64 ±2.639.36 ±0.77
4 Mean ±SD from 5 rats.
30 60 90TIME AFTER 5-FUra(min)
120
Fig. 2. Effect of a prior treatment with 5-diazouracil (5-DiazoUra) on theamount of radioactivity from [6-14C]FUra (5 (<Ci/150 g rat; 56 Ci/mol) excreted
into bile. Bile was collected every 10 min and counted. Points, means of fouranimals; bars, SE.
neys. The proportion of FUra breakdown occurring in thekidneys themselves, however, is difficult to assess from theFUra catabolites present in the tissue. A considerable portionof these catabolites presumably originated from FUra degradation in the liver, since more than 90% of an administeredFUra dose is excreted via the kidneys into urine (9, 41). a-FIuoro-/3-alanine is the major excreted metabolite which makesup 79% of the total label in urine (9). In the cytosol of ratkidneys, formation of labeled CO2 from [2-14C]Ura occurred at
10% of the rate detected in the supernatant of rat liver homog-enates. All the other tissues tested did not possess the completeenzymatic equipment for uracil degradation. In contrast to thesubsequent catabolic enzymes, dihydrouracil dehydrogenase hasbeen identified in many rat tissues (42, 43). The amount of a-fluoro-ß-alanine in supernatants of organs lacking dihydropyr-imidinase or 0-ureidopropionase originated from dihydrofiuorouracil converted into a-fluoro-/3-alanine during workup of thesamples or from the degradative products present in bloodplasma. In addition, total acid-soluble radioactivity in theseorgans after injection of [2-MC]uracil or [6-MC]FUra did not
differ significantly from the amount of label in blood plasma.The rapid catabolism of FUra in liver and kidneys was
reflected by a high concentration of a-fluoro-/3-aIanine in bloodplasma as early as IS min after injection of the base analogue.In human blood plasma also dihydrofiuorouracil (44), a-fluoro-j8-alanine, and presumably a-fluoro-0-ureidopropionic acidwere detected using I9F nuclear magnetic resonance (45). The
increase in blood plasma catabolites including the cytotoxicdihydrofiuorouracil (46) was almost completely prevented bypretreatment with S-diazouracil (Fig. 1). Under these conditions, the absolute amount of FUra in blood plasma stronglyrose by a factor of 27. Inhibition by 5-diazouracil of dihydrouracil dehydrogenase which degrades FUra more efficientlythan uracil in normal tissues and in tumors (12, 15, 16) thusrepresents a valuable approach in cancer chemotherapy (21) toimprove the bioavailability of FUra.
A pretreatment with 5-diazouracil which has been shown tomaximally inhibit dihydrouracil dehydrogenase in rat liver invivo (21 ) considerably suppressed the amount of total radioactivity in liver and kidneys by 75 and 66%, respectively. Thiswas in opposite to the effect on the total amount of label inblood plasma and in all other tissues examined which demonstrated a 2-fold rise in radioactivity mostly due to the accumulation of the intact base (Table 2; Fig. 2). The 5-diazouracil-induced increase in the label from [2-'4C]uracil in liver and
kidney homogenates reflected the absence of radioactive uracilbreakdown products since catabolism of the C-2-labeled uracilinvolves the loss of radioactivity via volatile I4CC>2.Under theseconditions the relative rise in radioactivity after 5-diazouracilwas similar in each of the different organs (Table 3).
In the liver of pretreated animals the proportion of unmetab-olized FUra amounted to 21 % of total acid-soluble radioactivityversus 2.2% of FUra anabolites. Although the hepatic fractionof anabolites was very low as compared to the label in catabolites and unchanged FUra, there was a 4-fold and 2.6-foldincrease in acid-soluble anabolite formation in the liver andkidneys after 5-diazouracil. Twenty-four h after FUra, a 3- to
5579
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CATABOLISM OF CIRCULATING PYRIMIDINES
4-fold enhanced utilization of the drug has also been observedin leukemia cells, liver, spleen, and small intestine of rats afterthe additional treatment with 5-diazouracil (21).
The methodology used did not allow to discriminate betweenthe individual catabolites of FUra in the organ extracts. Inisolated rat hepatocytes the main intracellular catabolite isdihydrofluorouracil after exposure of the cells to 30 ^M FUra(23). As in the liver in vivo (Fig. 1) unmetabolized FUra hasnot been detected within the cells (23). Thymine (2 HIM)causesa 99% inhibition of the degradative pathway in hepatocytes andis associated both with insignificant synthesis of FUra anabol-ites and the formation of a glucuronide of the FUra base (38).The main reason for the failure to identify the glucuronidederivative of FUra in the liver in vivo in the presence of 5-diazouracil presumably was the use of a low tracer dose ofFUra. The formation of the FUra conjugate has been shown todepend largely on the intracellular concentration of the baseanalogue (37).
In addition to the elimination of FUra and its catabolites inurine (8, 9, 41), we found that a considerable fraction of FUra-derived radioactivity was excreted in bile (Fig. 2). 5-Diazouracilhad no influence on bile flow but depressed the proportion oflabel in bile from 12.7% to 2%. It remains to be establishedwhether the recently discovered glucuronide of FUra (37, 38)is one of the radioactive metabolites in bile and whether 5-diazouracil is able to interfere with its formation.
The results of our experiments indicate that the catabolismof uracil and FUra is strongly inhibited in the whole animal ///vivo by 5-diazouracil through interference with pyrimidine catabolism occurring in liver and kidneys of rats. In addition, theexcretion of label from FUra into bile is considerably suppressed by 5-diazouracil. The bioavailability of FUra and itsconsumption for ribonucleoside or ribonucleotide synthesis ismarkedly increased in the acid-soluble supernatant of liver and
kidneys.
ACKNOWLEDGMENTS
The expert technical assistance by Dagmar Manglitz and ArminDietz is gratefully acknowledged.
REFERENCES
1. Keppler, I)., and Holstege, A. Pyrimidine nucleotide metabolism and itscompartmentation. In: H. Sies (ed.). Metabolic Compartmentation, pp. 147-203. New York: Academic Press, 1982.
2. Moyer, J. I ).. Oliver, J. T., and Handschumacher, R. E. Salvage of circulatingpyrimidine nucleosides in the rat. Cancer Res., 41: 3010-3017, 1981.
3. Holstege, A., Manglitz, I>..and Gerok, W. Depletion of blood plasma cytidinedue to increased hepatocellular salvage in D-galactosamine-treated rats. Eur.J. Biochem., 141: 339-344, 1984.
4. Moyer, J. I>.,Malinowski, N., and Ayers, O. Salvage of circulating pyrimidinenucleosides by tissues of the mouse. J. Biol. Chem., 260: 2812-2818, 1985.
5. Heidelberger, C., Danenberg, P. V., and Moran, R. G. Fluorinated pyrimi-dines and their nucleosides. Adv. Enzymol., 54:57-119, 1983.
6. Rutman, R. J., Cantarow, A., and Paschkis, K. E. Studies in 2-acetylamino-fluorene carcinogenesis. III. The utilization of uracil-2-C'4 by preneoplasticrat liver and hepatoma. Cancer Res., 14: 119-123, 1954.
7. Heidelberger, C., Leibman, K. C., Harbers, E., and Bhargava, P. M. Thecomparative utilization of uracil-2-C'4 by liver, intestinal mucosa, and Flex-ner-Jobling carcinoma in the rat. Cancer Res., 17:399-404, 1957.
8. Chaudhuri, N. K., Montag, B. J., and Heidelberger, C. Studies on fluorinatedpyrimidines. III. The metabolism of 5-fluorouracil-2-C14 and 5-fluoro-orotic-2-C'4 acid in vivo. Cancer Res., IS: 318-328, 1958.
9. Bernadou. J., Armand, J. P., Lopez, A., Malet-Martino, M. C., and Martino,R. Complete urinary excretion profile of 5-fluorouracil during a six-daychemotherapeutic schedule as resolved by "F nuclear magnetic resonance.Clin. Chem., 31: 846-848, 1985.
10. Reyes, P., and Guganig, M. E. Studies on a pyrimidine phosphoribosyltrans-ferase from murine leukemia P1534J. Partial purification, substrate specificity, and evidence for its existence as a bifunctional complex with orotidine5'-phosphate decarboxylase. J. Biol. Chem., 250: 5097-5108, 1975.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
Mclvor, R. S., Wohlhueter, R. M., and Plagemann, P. G. W. Uridinephosphorylase from Novikoff rat hepatoma cells: purification, kinetic properties, and its role in uracil anabolism. J. Cell. Physiol., 722: 397-404, 1985.Naguib, F. N. M., el Kouni, M. H., and Cha, S. Enzymes of uracil catabolismin normal and neoplastic human tissues. Cancer Res., 45: 5405-5412, 1985.Ferdinandus, J. A., Morris, H. P., and Weber, G. Behavior of opposingpathways of thymidine utilization in differentiating, regenerating, and neoplastic liver. Cancer Res., 31: 550-556, 1971.Traut, T. W., and Loechel, S. Pyrimidine catabolism: individual characterization of the three sequential enzymes with a new assay. Biochemistry, 23:2533-2539,1984.Tuchman, M., Ramnaraine, M. L. R., and O'Dea, R. F. Effects of uridineand thymidine on the degradation of 5-fluorouracil, uracil, and thymidine byrat liver dihydropyrimidine dehydrogenase. Cancer Res., 45: 5553-5556,1985.Shiotani, T., and Weber, G. Purification and properties of dihydrothyminedehydrogenase from rat liver. J. Biol. Chem., 256: 219-224, 1981.Dudley, K. H., Butler, T. C., and Bius, D. L. The role of dihydropyrimidinasein the metabolism of some hydantoin and succinimide drugs. Drug. Metab.Dispos., 2: 103-112, 1974.Gasser, T., Moyer, J. D., and Handschumacher, R. E. Novel single-passexchange of circulating uridine in rat liver. Science (Wash. DC), 213: 777-778, 1981.Dahnke, H.-G., and Mosebach, K.-O. In v/vo-Untersuchungen zur Metabol-isierung der Pyrimidinnucleoside. Hoppe-Seyler's Z. Physiol. Chem., 356:1565-1574,1975.Yngner, T., Engelbrecht, C., Lewan, L., and Annerfeldt, J.-E. Anabolismversus catabolism of [5-3H]uridine and its relationship to ribonucleic acidlabelling in mouse liver after partial hepatectomy. Biochem. J., 178: 1-8,1979.Cooper, G. M., Dunning, W. F., and Gréer,S. Role of catabolism inpyrimidine utilization for nucleic acid synthesis in vivo. Cancer Res., 32:390-397, 1972.Fritson, P. The catabolism of C'4-labeled uracil, dihydrouracil, and .¡-iin-idopropionic acid in rat liver slices. J. Biol. Chem., 226: 223-228, 1957.Sommadossi, J.-P., Gewirtz, D. A., Diasio, R. B., Aubert, C., Cano, J.-P.,and Goldman, I. D. Rapid catabolism of 5-fluorouracil in freshly isolated rathepatocytes as analyzed by high performance liquid chromatography. J. Biol.Chem., 257:8171 -8176, 1982.Holstege, A., Leser, H.-G., Pausch, J., and Gerok, W. Uridine catabolism inKupffer cells, endothelial cells, and hepatocytes. Eur. J. Biochem., 149:169-173, 1985.Monks, A., and Cysyk, R. L. Uridine regulation by the isolated rat liver:perfusion with an artificial oxygen carrier. Am. J. Physiol., 242: R465-R470,1982.Ardalan, B., Cooney, D., and MacDonald, J. S. Physiological and pharmacological determinants of sensitivity and resistance to 5-fluorouracil in loweranimals and man. Adv. Pharmacol. Chemother., 17: 289-321, 1980.Spiegelman, S., Nayak, R., Sawyer, R., Stolti, R., and Martin, D. S. Poten-tiation of the anti-tumor activity of 5FU by thymidine and its correlationwith the formation of (5FU) RNA. Cancer (Phila.), 45: 1129-1134, 1980.Au, J. L.-S., Rustum, Y. M., Ledesma, E. J., Mittelman, A., and Creaven,P. J. Clinical pharmacological studies of concurrent infusion of 5-fluorouraciland thymidine in treatment of colorectal carcinomas. Cancer Res., 42:2930-2937, 1982.Santelli, G., and Valeriote, F. In vivo enhancement of 5-fluorouracil cytotox-icity to AKR leukemia cells by thymidine in mice. J. Nati. Cancer lust.. 61:843-847, 1978.Martin, D. S., Nayak, R., Sawyer, R. C., Stolfi, R. L., Young, C. W.,Woodcock, T. M., and Spiegelman, S. Enhancement of 5-fluorouracil chemotherapy with emphasis on the use of excess thymidine. Cancer Bull., 30:219-224, 1978.Young, C. W., Woodcock, T. M., and Martin, D. S. Modulation of 5-FUaction by thymidine in murine and human tumors. Cancer Treat. Rep., 65:83-87, 1981.Agarwal, D. P., Eickhoff, K., and Goedde, H. W. A note on in vitro inhibitionof thymine degradation by some pyrimidine analogues and related products.Z. Naturforsch., 24b: 469-470, 1969.Cooper, G. M., and Greer, S. Irreversible inhibition of dehalogenation of 5-iodouracil by 5-diazouracil and reversible inhibition by 5-cyanouracil. CancerRes., 30: 2937-2941, 1970.Bradford, M. M. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding.Anal. Biochem., 27: 248-254, 1976.Keppler, D., Pausch, J., and Decker, K. Selective uridine triphosphate deficiency induced by D-galactosamine in liver and reversed by pyrimidinenucleotide precursors. Effect on ribonucleic acid synthesis. J. Biol. Chem.,249:211-216, 1974.Gehrke, C. W., and Kuo, K. C. Major and modified nucleosides, RNA, andDNA. In: L. J. Marlon and P. M. Kabra (eds.), Liquid Chromatography inClinical Analysis, pp. 409-443. Clifton, NJ: The Humana Press, 1981.Sommadossi, J.-P., Cross, D. S., Gewirtz, D. A., Goldman, I. D., Cano, J.-I'.. and Diasio, R. B. Evidence from rat hepatocytes of an unrecognizedpathway of 5-fluorouracil metabolism with the formation of a glucuronidederivative. Cancer Res., 45: 2450-2455, 1985.Sommadossi, J.-P., Gewirtz, D. A., Cross, D. S., Goldman, I. D., Cano, J.-P., and Diasio, R. B. Modulation of 5-fluorouracil catabolism in isolated rat
5580
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

CATABOLISM OF CIRCULATING PYRIMIDINES
hepatocytes with enhancement of S-fluorouracil glucuronide formation. Can- activity in normal, differentiating, and regenerating liver and in hepatomas.cer Res., 45: 116-121, 1985. Cancer Res., 31:1004-1009, 1971.
39. Ensminger, W. D., Rosowsky, A., Raso, V., Levin, D. C., Glode, M., Come, 43. Ikenada, K., Shirazaka, T., Kitano, S., and Fujii, S. Effect of uracil onS., Steele, G., and Frei, E., III. A clinical-pharmacological evaluation of metabolism of S-fluorouracil in vitro. Gann. 70:353-359, 1979.hepatic arterial infusions of 5-fluoro-2'-deoxyuridine and 5-fluorouracil. 44. Sommadossi, .1.1'.. Aubert, C., Cano, .1.1'., Gouveia, .1. Ribaud, P., andCancer Res., 38: 3784-3792, 1978. Mathé,G. Kinetics and metabolism of a new fluoropyrimidine 5'-deoxy-5-
40. Speyer, J. L., Sugarbaker, P. H., Collins, J. M., Dedrick, R. L., Klecker, R. fluorouridine in humans. Cancer Res., 43: 930-933, 1983.W., Jr., and Myers, C. E. Portal levels and hepatic clearance of 5-fluorouracil 45. Malet-Martino, M.-C, Martino, R., Lopez, A., Béteille,J.-P., Bon, M.,after intraperitoneal administration in humans. Cancer Res., 41:1916-1922, Bernadou, J., and Armand, J.-P. New approach to metabolism of 5'-deoxy-1981. 5-fluorouridine in humans with fluorine-19 NMR. Cancer Chemother. Phar-
41. Mukherjee, K. L., and Heidelberger, C. Studies on fluorinated pyrimidines. macol., 13:31-35, 1984.IX. The degradation of 5-fluorouracil-6-C14. J. Biol. Chem., 235: 433-437, 46. Diasio, R. B., Schuetz, J. D., Wallace, H. J., and Sommadossi, J.-P. Dihy-
1960. drofluorouracil, a fluorouracil catabolite with antitumor activity in murine42. Queener, S. F., Morris, H. P., and Weber, G. Dihydrouracil dehydrogenase and human cells. Cancer Res., 45: 4900-4903, 1985.
5581
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1986;46:5576-5581. Cancer Res Axel Holstege, Jürgen Pausch and Wolfgang Gerok Pyrimidines in Rat Liver and KidneysEffect of 5-Diazouracil on the Catabolism of Circulating
Updated version
http://cancerres.aacrjournals.org/content/46/11/5576
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on July 22, 2017. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























