Embed Size (px)
Citation preview

JURNAL VETERINAR MALAYSIA
Vol. 25 No. 1 & 2 Dec 2013
Table of Contents
Journal Article
Potency and efficacy of a low pathogenic H5N2 inactivated vaccine against
challenge with a Malaysian H5N1 highly pathogenic avian influenza virus S.H. Sharifah, M.N. Surian, M. Maizan, G.H. Ong, D. Azizah, K. Suzana, A.R. Omar
and I. Aini………………………………………………………………………..........1
Effects of Escherichia Coli vaccination in gilts on piglet performance in a farm in
Perak
W.C. Michelle-Fong, P.Y. Choo, B.L. Ong, C.Y. Tee, J.W. Lee, and P.T. Ooi.............7
Occurrence of Salmonella and other enteric microbes in faeces of house lizards
(Hemidactylus frenatus)
Gwen Lo and Saleha A.A……………………………………………………………..........11
Review Article
Feline Hypertrophic Cardiomyopathy – Prevalence, risk factors and
pathological aspect
K.H. Khor…………………………………………………....……………………....15
The role of omega-3 polyunsaturated fatty acids on brain cognitive function -
Review of studies on laboratory animals
A. Hafandi ……………………………………………………………………......…21
Short Communication
Moderately-high humoral antibody responses to a H5N2 inactivated vaccine did
not suppress shedding of highly pathogenic H5N1 Avian Influenza virus during
challenge
S.H. Sharifah, K. Hasuzana, M.N. Suriani, M. Maizan, G.H.. Ong, D. Azizah, A.R.
Omar and I.Aini........................................................................................ ..................27
Jurnal Veterinar Malaysia is the official journal of the Veterinary Association
Malaysia (VAM). It was published formally as Kajian Veterinar and the Malaysian
Veterinary Journal.
Researcher papers on various aspects of veterinary medicine, animal science and
research are invited for publication either as full articles or short communication.
Review papers and abstracts of articles of local interest are also published from time
to time. The publisher (VAM) does not hold itself responsible for statements made in
the journal by contributors. Unless so stated, materials in the Journal do not reflect an
endorsement or an official attitude of VAM or the Editorial Board of JVM.
THE VETERINARY ASSOCIATION MALAYSIA ISSN 9128-2506
EDITORIAL BOARD 2013 Editor-in-Chief: Dr Khor Kuan Hua Advisor: Prof. Dato’ Dr. Abdul Rani Bahaman Board Members: Prof. Dr. Saleha Abdul Aziz Prof. Dr. Rasedee Abdullah Prof. Dr. Mohd. Hair Bejo Prof. Dr. Mohd. Azmi Mohd. Lila Prof. Datin Dr. Kalthum Hashim Assoc. Prof. Dr. Abdul Rahim Mutalib Assoc. Prof. Dr. Latiffah Hassan Assoc. Prof. Dr. Rosnina Yusoff Assoc. Prof. Dr. Sharifah Syed Mohd Hassan Assoc. Prof Dr. Goh Yong Meng Dr. Chen Hui Cheng Dr. Rozanaliza Radzi
EDITORIAL ADVISORY BOARD 2013
Datuk Dr. Abdul Aziz Jamaludin Department of Veterinary Services Malaysia, Putrajaya Prof. Dr. Abdul Aziz Saharee Universiti Putra Malaysia Prof. Dr. Abdul Razak Alimon Universiti Putra Malaysia Prof. Datin Paduka Dr. Aini Ideris Universiti Putra Malaysia Prof. Dr. Eric Gruys Utretch University Utretch, the Netherlands Prof. Dr. Husni Omar Mohamed Cornell University USA Prof. Dr. Mohd Zamri Saad Universiti Putra Malaysia Prof. Dr. Ramli Abdullah Universiti Malaya
Editorial and Business Address:
c/o Faculty of Veterinary Medicine
University Putra Malaysia
43400 UPM Serdang,
Selangor Darul Ehsan
Malaysia
Tel. : (03) 8609 3926
Fax : (03) 8947 1971
E-mail : [email protected]

J. Vet. Malaysia (2013) 25 (1&2):1-6
1
Journal Article
POTENCY AND EFFICACY OF A LOW PATHOGENIC H5N2 INACTIVATED VACCINE AGAINST
CHALLENGE WITH A MALAYSIAN H5N1 HIGHLY PATHOGENIC AVIAN INFLUENZA VIRUS
S.H. Sharifah1*, M.N.Suriani2, M. Maizan2, G.H.Ong2, D. Azizah2, K.Suzana2, A.R. Omar3and I. Aini3
1Monash UniversityMalaysia, Jalan Lagoon Selatan, 46150 Bandar Sunway, Malaysia
2Veterinary Research Institute, Jalan Sultan Azlan Shah, 31400 Ipoh, Perak, 3Faculty of Veterinary Medicine, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
SUMMARY
The potency and efficacy of an avian influenza (AI) H5N2 inactivated vaccine that was developed at Veterinary Research Institute,
Ipoh was tested. The percentage sequence identity of the HA gene of the H5N2 vaccine virus to the challenge virus
[A/chicken/Malaysia/5858/04 (H5N1)] was 88.2% by nucleotide and 90% by amino acid sequences similarities, respectively. As for the
HAI segment, the nucleotide sequence similarities were 88.3 % and by amino acid sequence 87.7%.For potency testing, the heterologous
killed H5N2 AI vaccine, formulated as an oil emulsion was administered only once subcutaneously in twenty five two-week old
commercial broiler chickens. The HI antibodies were not detectable at week 1 post vaccination. The HI GMT attained was 30, 63, 200,
54 and 32 by week 2, 3, 4, 5, and 6 post vaccinations. Efficacy study was conducted on ten SPF chickens at week 3 post vaccination.
60% of the birds (6/10) with HI titres ≥ 64 - 128 survived the challenged. H5N1 challenge virus was reisolated from all the birds with HI
titre ≤ 32 that died, and each of the birds that survived with HI titres of 64 and 128, from the oropharynx and cloaca at day 3 post
challenge. This vaccine protected 60% of chickens against mortality and did not prevent shedding after challenged with a HPAI H5N1
virus.
Keywords: Avian Infuenza, Virus, Vaccine
INTRODUCTION
Since the outbreaks of highly pathogenic avian
influenza (HPAI) H5N1 in poultry in 2000 to 2004, various
countries have adopted several strategies to control or
eradicate the disease. Some have chosen stringent measures
such as killing and destruction of infected poultry.
However, as these methods proved to be expensive and
biosecurity measures and culling cannot be implemented to
successfully control or eradicate the disease for some
countries, an alternative method, is therefore, vaccination.
Vaccination is also one of the tools recommended by
international health organisations in controlling AI (OIE).
For this reasons only two types of vaccines have been
currently approved, (i) heterologous low pathogenic
inactivated vaccines and (ii) recombinant vaccines (Swayne
et al., 2000). Since the emergence of H5N1 in Asia, several
heterologous inactivated vaccines have been developed and
tested against H5 and H7 influenza viruses in poultry and
the use of heterologous inactivated H5N2 vaccines had been
reported in chickens in Hong Kong (2002 - 2006), Pakistan
(2006), India (2006), Russia (2005), Egypt (2006), in ducks,
geese and chickens in China (2004) and Vietnam (2005) to
name a few (Swayne et al., 2001; Swayne et al., 2006;
Swayne 2009). Although these vaccines can protect poultry
from clinical disease, sterile immunity is not achieved under
field conditions, allowing for undetected virus spread and
evolution under immune cover (Fuchs et al., 2009).
However, controlling highly pathogenic H5N1 using
*Corresponding author: Assoc. Prof. Dr. Sharifah Syed Mohd Hassan (S.H. Sharifah) Email: [email protected]
inactivated highly pathogenic H5N1 vaccines are not
permissible for fear that residual viruses that are not fully
inactivated can cause outbreaks. Despite this, in 2003,
Indonesia, however, started using an autologous inactivated
H5N1 vaccine to control the rapid spread of H5N1 in its
poultry population (Swayne, 2009). However, they showed
that the inactivated homologous H5N1 vaccine being
completely protective than the H5N2 virus vaccines against
H5N1 challenged. In using inactivated heterologous
vaccines, where the virus strain used to make the vaccine is
of the same H subtype as the challenging field virus the
clinical protection and the reduction or viral shedding are
ensured by the homologous H group (Capua and Marangon,
2003). Similar HA subtype or high percentage homology
(90 - 96%) between the vaccine strain and the circulating
strain are critical factors for the efficacy of the vaccine.
However, other factors such as antigen quantity and content
and the adjuvant used for the efficacy of the inactivated
vaccines are also important (Swayne et al., 1999; Wood et
al., 1985). The ability of the heterologous vaccine to
provide protection against mortality and morbidity, reduce
cloacal and oropharyngeal shedding and ability to prevent
viral spread to other vaccinated or susceptible birds have
been considered as important factors for protective efficacy
of the vaccine. The aim of the study is to determine the
potency and efficacy of the inactivated H5N2 vaccine
developed, and the ability of the vaccine to invoke sterile
immunity as depicted by shedding of challenge virus, after
challenged with a highly pathogenic Malaysian strain of
H5N1 virus.

J. Vet. Malaysia (2013) 25 (1&2):1-6
2
MATERIALS AND METHODS
Viruses
The vaccine virus A/duck/Malaysia/8443/04 H5N2 was
isolated from the cloacal swab of a duck in a routine
surveillance study in the country. During isolation of the
virus in 9 - 11 day-old SPF embryonated eggs, the HA
activity was detected as early as the first passage. However,
it took 4 passages before the virus kill the SPF embryonated
eggs. The virus was non-pathogenic as determined by the
intravenous pathogenicity index (IVPI) by the standard
procedure (Council Directive 92/40/EEC (1992) Off. J. Eur.
Communities L167, 1 - 16). The sequence of the HA
cleavage site is TIGECPKYVKSDRLVLAKGLRNVPQ----
RETRGLF.
The challenge virus strain used was
A/chicken/Malaysia/5858/04 H5N1. This virus was isolated
from chickens during an outbreak in Malaysia in 2004. The
virus had an intravenous pathogenicity index (IVPI) of 3.0,
where 4 weeks old chickens inoculated with this virus died
within 24 hr (determined by AAHL, Geelong, Australia, the
OIE Reference Centre for Avian Influenza). The presence
of multiple basic amino-acids at the HA cleavage site
sequence of
TIGECPKYVKSNRLVLATGLRNSPQRERRRKKRGLF
indicated the high pathogenicity of the virus. The lethal
dose of the virus was determined to be 105EID50/0.1ml
where it causes 100% mortality of SPF chickens within 48
hr post-infection. All laboratory and animal experiments
using the highly pathogenic H5N1 virus was performed in a
BSL-3 facility of the Veterinary Research Institute, Ipoh,
Malaysia.
Sequencing of the Haemagglutinin gene
PCR was carried out to amplify the full length HA gene
of the H5N1 challenge virus A/chicken/Malaysia/5858/04
H5N1 and the A/duck/Malaysia/8443/04 H5N2, using HA
specific primers as previously described (Hoffmann et al.,
2001). The products were cloned into TOPO PCR vector
and sequenced. Sequences were assembled and edited using
Staden Package, Pairwise sequence alignments and the
nucleotide and amino acid sequence were compared using
the Bio-Edit 7 and Genetyx-Mac programmes.
Preparation of the H5N2 Vaccine
The master seed and working viruses of the duck
isolate were prepared in SPF eggs. A preliminary batch of
vaccine virus was produced by inoculating a batch of 500
SPF eggs with 103.4
EID50/0.1 ml (This dose was found to
give the highest virus titre at day 3 post inoculation).
Vaccine virus infected eggs were incubated for 3 days. The
undiluted allantoic fluid containing virus was inactivated for
18 hr with B-propiolactone at 0.01 %v/v and adjuvanted
with 10%Montanide™ gel. The pre-activation infectivity
titre and the HA titre of the vaccine virus were 107.3
EID50/0.1 ml and 128 HAU respectively.
For determining the potency of the H5N2 inactivated
vaccine, twenty five three-day old commercial broilers
which were not vaccinated with any poultry vaccines were
reared until they reached the age of two weeks-old. The
birds were wing-banded and reared in a non-infectious
animal housing unit.
Vaccination response-experiment
At two weeks old, the birds were immunized with the
H5N2 vaccine. A dosage of 200µl was injected
subcutaneously (SQ) per bird, and the serology of all the
immunized birds were evaluated every week for a period of
6 weeks.
HI assay
The detection of antibodies after vaccination was
studied by the HI assay performed according to the WHO
manual on Animal Influenza diagnosis and Surveillance
(WHO/CDS/CSR/NCS/2002.5). Serum samples were
diluted 2 fold, with the initial serum dilution at 1:2. Titres >
3 log2(8) are considered positive. The serological response
was evaluated for all birds before and after vaccination. The
HI test was performed in V-bottom 96 well microtiter plates
with 8 HAU/50µl of homologous inactivatedH5N1 antigen
per well.
Challenging vaccinated birds with H5N1 virus
In another experiment, ten two-week old SPF chickens
(raised at SPF chicken facility of Veterinary Research
Institute, Ipoh, Malaysia) were vaccinated with 200µl of the
H5N2 vaccine via the SQ route. At 3 weeks post
vaccination (based on 100% seroconversion from earlier
potency study), the birds were challenged with 200µl
containing 105.3
EID50/bird of the H5N1 virus via the
intranasal route. Challenging of the chickens with HPAI
H5N1 virus, was conducted in a negative pressure isolator
cabinet ventilated with HEPA-filtered air in a NATA-
certified biosafety level-3 facility of Veterinary Research
Institute, Ipoh. Water and feed were provided at libitum.
Five SPF birds that had not been vaccinated with the H5N2
vaccine were also challenged with the same dose of virus.
Clinical signs were monitored daily for one week post-
challenged. Cloacal and oropharyngeal swabs of each of the
chickens were sampled at 3 days post challenge for H5N1
virus re-isolation. Virus isolation was performed in 9 - 11
days old SPF embryonated eggs using standard procedures
(OIE, 2012). The presence of H5N1 challenge virus was
detected using the HA test and confirmed using specific
H5N1 haemagglutination-inhibition (HI) serological test.
Three passages were undertaken and HA test performed at
each passage before the samples were considered negative.
J. Vet. Malaysia (2013) 25 (1&2):1-6
3
RESULTS
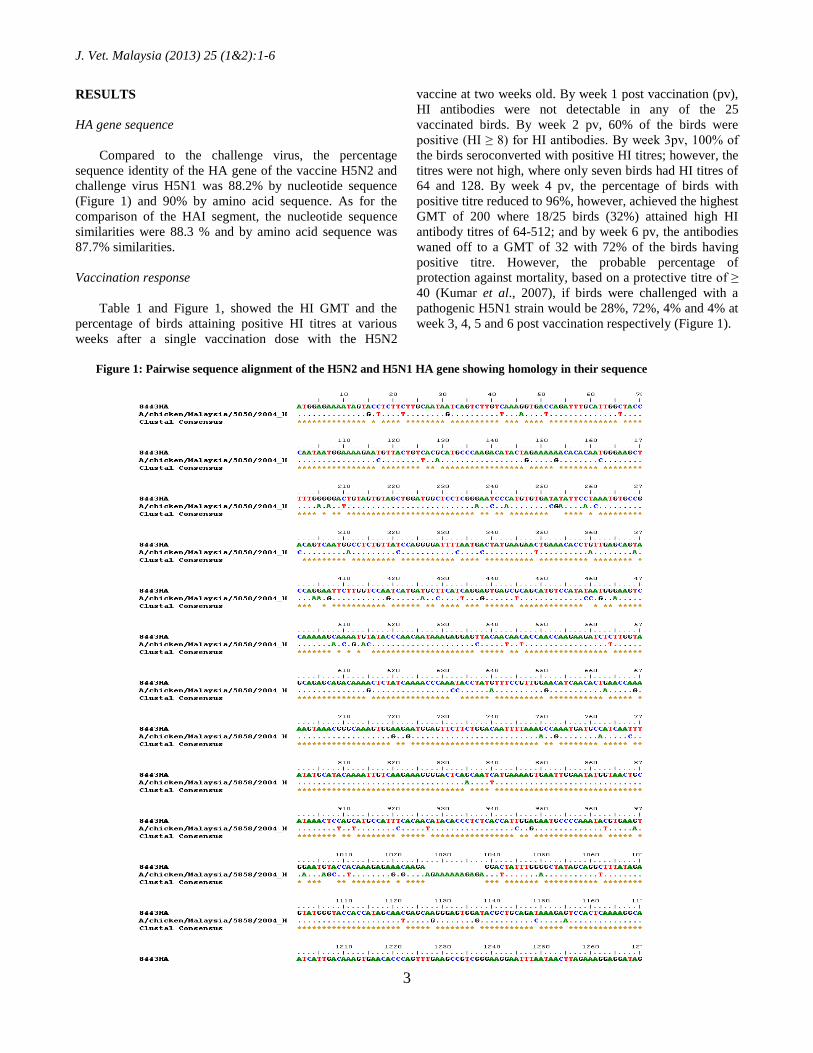
HA gene sequence
Compared to the challenge virus, the percentage
sequence identity of the HA gene of the vaccine H5N2 and
challenge virus H5N1 was 88.2% by nucleotide sequence
(Figure 1) and 90% by amino acid sequence. As for the
comparison of the HAI segment, the nucleotide sequence
similarities were 88.3 % and by amino acid sequence was
87.7% similarities.
Vaccination response
Table 1 and Figure 1, showed the HI GMT and the
percentage of birds attaining positive HI titres at various
weeks after a single vaccination dose with the H5N2
vaccine at two weeks old. By week 1 post vaccination (pv),
HI antibodies were not detectable in any of the 25
vaccinated birds. By week 2 pv, 60% of the birds were
positive (HI ≥ 8) for HI antibodies. By week 3pv, 100% of
the birds seroconverted with positive HI titres; however, the
titres were not high, where only seven birds had HI titres of
64 and 128. By week 4 pv, the percentage of birds with
positive titre reduced to 96%, however, achieved the highest
GMT of 200 where 18/25 birds (32%) attained high HI
antibody titres of 64-512; and by week 6 pv, the antibodies
waned off to a GMT of 32 with 72% of the birds having
positive titre. However, the probable percentage of
protection against mortality, based on a protective titre of ≥
40 (Kumar et al., 2007), if birds were challenged with a
pathogenic H5N1 strain would be 28%, 72%, 4% and 4% at
week 3, 4, 5 and 6 post vaccination respectively (Figure 1).
Figure 1: Pairwise sequence alignment of the H5N2 and H5N1 HA gene showing homology in their sequence

J. Vet. Malaysia (2013) 25 (1&2):1-6
4
Table 1: Relationship of the potency, HI Geometric Mean Titre (GMT) and probable percentage protection afforded by
the H5N2 vaccine on 25 commercial birds. Chicks were vaccinated at two weeks old and each bird was inoculated SQ
with 200µl vaccine (pre-activation titre: 107.3EID50/0.1ml)
Challenged response and shedding
Only ten birds were used for the challenge and
shedding studies as there was limited space in the BSL-
3 cabinet for ease of handling the chickens. As was
observed in the potency study, the rise of humoral HI
antibodies were slow, i.e. it took three weeks post
vaccination for all birds to seroconvert. Challenge was
therefore done at week 3 pv, to ensure that all birds have
antibody titres by then. The birds had pre challenged HI
titres ranging from 8 – 128 i.e two birds with HI titre of
8, two birds with HI titre of 32; four birds with HI titre
of 64 and two birds with HI titre of 128 (Figure 2). All
four birds with HI titre ≤ 32 died during challenged. The
birds died within 3 - 4 days post challenged. The six
birds with HI titre ≥ 64 survived challenged with no
clinical signs observed. Shedding was evaluated at only
one time i.e. at 3 days post-challenged. Challenge H5N1
virus was excreted in the oropharynx and cloaca when
examined at 3 days post challenged in 7/10 birds (70%),
i.e from four birds that died at 3 - 4 days post
challenged, in one bird with HI titre of 64 and one bird
with HI titre of 128. Birds showed signs of depression,
ruffled feathers and loss of appetite before death.
DISCUSSION
The Government of Malaysia does not adopt
the policy of routine vaccination of poultry against
avian influenza. However, in a worst case scenario, the
government recognizes the potential of vaccination as a
complementary measure in the control and eradication
of HPAI, or at least for the vaccination of expensive or
rare exotic birds. In view of this, a pilot batch of vaccine
was prepared using a low pathogenic
A/Duck/Malaysia/8443/04 (H5N2) virus. In our study,
even at a high pre-activation titre of H5N2 virus of
107.3
EID50/0.1 ml, and adjuvanted with 10% montanide
gel (a potent adjuvant), the HI titres invoked with a
single vaccination of this vaccine is moderately low
Week post
vaccination
No of
birds
HI
titre
GMT a/b (Percent) positive
HI titre :HI ≥ 8
Probable percentage of protection based on a
protective titer HI value ≥ 40 (Kumar et al. 2007)
0 (before vaccination) 25 <2 0 0/25 (0%) 0%
1 25 <2 0 0/25 (0%) 0%
2 5
5
5
7
3
<2
4
8
16
32
30 15/25 (60%) 0%
3 4
14
5
2
16
32
64
128
63 25/25 (100%) 7/25 (28%)
4 1
2
2
2
10
6
1
1
4
8
16
32
64
128
256
512
200 24/25 (96%) 18/25 (72%)
5 2
1
2
13
6
1
<2
2
8
16
32
128
54 22/25 (88%) 1/25 (4%)
6 4
1
2
12
5
1
<2
2
4
8
16
64
32 18/25 (72%) 1/25 (4%)

J. Vet. Malaysia (2013) 25 (1&2):1-6
5
with the highest HI titre achieved was 512 in only one
bird. It was only after three weeks post vaccination that
100% seroconversion was observed.
The HI antibody response could not be
detected at 1 week post vaccination, however the GMT
achieved its peak of 200 by week 4 pv but the antibodies
waned off quickly by week 6 pv. This low-moderately
low potency of the vaccine had also been shown by
Kumaret al. (2007), in chickens vaccinated with a
reverse genetic H5N3 isolate where the HA gene was
derived from A/chicken/Vietnam H5N1. The chickens
achieved suboptimal antibody response of HI < 40. He
also showed that chickens with serologic responses of >
40 were protected against challenge with the H5N1
virus. He also showed that, at this protective titre, the
virus could still be reisolated from one out of the 62
birds tested. In our potency study, using Kumar’s value
of HI > 40 as the protective titre, at week 3 and 4 post
challenged, the probable protection afforded would only
be 28% and 72% respectively. However, in our
challenged study, using ten SPF chickens, 60%
protection was afforded when chickens were challenged
at week 3 post vaccination. We were also able to
reisolate the challenge H5N1 virus in 7/10 birds. In
conclusion, the H5N2 inactivated vaccine invoked only
sub-optimal humoral HI antibody titres, not enough to
protect at least 80% of the birds against challenge,
although the HA protein share 90% amino acid
homology with the challenge H5N1 virus. According
to Swayne et al., 1999, the degree of protection of
inactivated vaccines is not strictly correlated to the
degree of homology between the HA gene or protein of
the vaccine and challenge strains, therefore the vaccine
can still be improved to achieve a higher degree of
clinical protection and a better reduction of shedding i.e.
by increasing the antigen mass of the vaccine. Due to
space constrains of the BSL-3 facility, this is only a
preliminary and small study, and therefore there were
insufficient numbers of birds at all the various HI titres
to make statistical inferences of protection associated
with titres.
REFERENCES
Capua, I. and Marangon, S. (2003). The use of vaccination as an
option for the control of avian influenza. Avian
Pathology,32(4), 335-343. Fuchs,W.,Römer-Oberdörfer, A., Veits, J. and Mettenleiter T.C.
(2009). Novel avian influenza virus vaccines-Review. RevSci
Tech. 2009 Apr; 28(1):319-32. Kumar, M., Chu, H.J., Rodenberg, J. and Krauss, S. (2007).
Association of serologic and protective responses of avian
influenza vaccines in chickens. Avian Diseases, 51: 481-483. OIE Terrestrial Manual 2012, Chapter 2.3.4 on avian influenza,
Version adopted by the World Assembly of Delegates of the
OIE in May 2012. 436 http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/
2.03.04_AI.pdf
Figure1: Potency of the H5N2 vaccine. HI-GMT value and
probable protection (HI > 40 based on Kumar et al., 2007)
Figure 2: Challenged Study. 60% (6/10) of the birds was
protected after challenged with the H5N1 virus. The
protected birds had HI titres of 64 and 128
Swayne, D.E (2009). Avian influenza vaccines and therapies for
poultry. Comparative Immunology, Microbiology and
Infectious Diseases, 32: 351-363.
Swayne, D.E., Beck, J.R., Garcia, M. and Stone H.D. (1999).
Influence of virus strain and antigenic mass on efficacy of H5
avian influenza inactivated vaccines. Avian pathology, 28, 245-255.
Swayne, D.E., Beck, J.R., Perdue, M.L. and Beard, C. W.
(2001).Efficacyof vaccines in chickens against highly pathogenic Hong Kong H5N1 avian influenza. Avian
Dis.45:355–365.
Swayne, D.E., Garcia, M., Beck, J.R., Kinney, N. and Suarez, D.L. (2000). Protection against diverse HP H5AI nviruses in
chickens immunized with a recombinant fowl pox vaccine
containing an H5 AI haemagglutinin gene insert. Vaccine, 18: 1088-1095.

J. Vet. Malaysia (2013) 25 (1&2):1-6
6
Swayne, D.E., Lee,C.W. and Spackman,E. (2006). Inactivated North
American and European H5N2 avian influenza virus vaccines protect chickens from Asian H5N1 high pathogenicity avian
influenza virus. Avian Pathol.35:141–146. 2006.
Wood, J.M., Kawaoka, Y., Newberry, L.A., Bordwell, E, and Webster R.G. (1985). Standardization of inactivated H5N2 influenza
vaccine and efficacy against lethal
A/Chicken/Pennsylvania/1370/83 infection. Avian Dis. 29: 867–872. 1985.

J. Vet. Malaysia (2013) 25 (1&2):7-10
7
Journal Article
EFFECTS OF ESCHERICHIA COLI VACCINATION IN GILTS ON PIGLET PERFORMANCE IN
A FARM IN PERAK
Michelle-Fong, W.C.1, Choo, P.Y.1,Ong, B.L.1, Tee, C.Y.2, Lee, J.W.2 and Ooi, P.T.1*
1Faculty of Veterinary Medicine, Universiti Putra Malaysia (UPM), 43400 UPM, Serdang, Selangor, Malaysia.
2Rhone Ma Malaysia (M) Sdn. Bhd., 46100 Petaling Jaya, Selangor, Malaysia
SUMMARY
This study aimed to observe the effects of Escherichia coli (Neocoliporvaccine – Merial) vaccination on diarrhoea percentages,
growth parameters (average weight per piglet and average daily gain) and mortality rate in new-born piglets. A field trial was conducted
in 35 litters of piglets from gilts selected from a farm in Perak. They were randomly allocated into Treatment (16 litters from E. coli
vaccinated gilts) and Control (19 litters) groups respectively. Body weights of the piglets were measured at days 1, 7, 14 and 21 of age
and the episodes of diarrhoea and piglet mortality were monitored daily for each pen. The Treatment group had significantly lower Day 1
neonatal diarrhoea percentage (p < 0.05) and significantly lower mortality rate from Day 1 to Day 7. The total mortality rate for overall
period of 1 - 21 days in the treatment group was at 3.90% when compared to the control group at 8.96%. However, there were no
significant differences (p > 0.05)in the overall diarrhoea percentages (1 - 14 days) and weekly growth parameters between both groups.
Environmental stress and inevitable routine treatment of diarrhoea with antimicrobials within the farm may have affected the significance
of the diarrhoea percentages and growth parameters in this study. In conclusion, E. coli vaccination in gilts was shown to significantly
reduce piglet mortality from Day 1 to Day 7 and neonatal diarrhoeal percentageson1-day-old piglets under typical farm conditions in this
pilot study in Malaysia.
Keywords: Escherichia coli vaccination, neonatal piglet diarrhoea percentage, neonatal piglet mortality rate, average weight per piglet, average daily gain
INTRODUCTION
Escherichia coli, also known as E. coli, is an ubiquitous
organism and one of the leading causes of diarrhoea in
suckling piglets, especially in piglets reared under intensive
management system (Fürer et al., 1982). Neonatal diarrhoea
associated with E. coli is most commonly observed in
piglets aged from 0 – 4 days (Loh et al., 2006; Schwartz,
2009). The severity of neonatal diarrhoea associated with E.
coli is also age-related, and the highest incidence of life-
threatening diarrhoea occurs during the first 2 to 5 days of
life, with less serious diarrhoea occurring later (Loh et al.,
2006; Schwartz, 2009). Neonatal diarrhoea and deaths
caused by enterotoxigenic Escherichia coli (ETEC) were
observed in many herds, especially in piglets farrowed by
gilts, whereas piglets from older sows showed lower
vulnerability (Too, 1997; Riising et al., 2005).
The high pre-weaning mortality rate in Malaysia of
about 12% of total piglets born alive has not changed over a
period of 15 years (1981–1996) and this post a major
problem in the swine industry (Loh et al., 1999). Heavy
losses have been reported in piglets during the first week of
life in Malaysia and many were thought to be as a result of
E. coli infection, or commonly known as colibacillosis,
where more than 95% of E. coli isolated from diarrhoeic
piglets had developed multiple antibiotic resistances, and
more than 50% showed resistance to 10 types of
antimicrobials tested (Loh et al. 2006) can impose a high
economic impact to the producers. * Corresponding Author: Dr.Ooi Peck Toung (Ooi, P.T.); Ph: +603-8946 3916; Email: [email protected]
Therefore, it is highly recommended to prevent this disease
than to continuously fight it (Holden et al., 2006).
Antibodies in colostrum provide passive protection to
suckling piglets from sows that have built up immunity to
specific E. coli strains (Carr, 2006). Active immunization of
sows effectively provided protection to the newborn piglets
through the transfer of antibodies via colostrum (Riising et
al., 2005). The protection of newborn pigs against E. coli
infection by sow vaccination has been a well-established
practice in Denmark for more than 2 decades (Riising et al.,
2005). This practice is not commonly done in farm
conditions in Malaysia.
This study has its aim to observe the effects on the
neonatal (first week) and pre-weaning (second and third
week) performance of piglets sowed by gilts vaccinated
with E. coli vaccine in a farm in Perak, Malaysia for the
following parameters: (1) Diarrhoea percentages, (2)
Growth Performance–average body weight and average
daily gain and (3) Mortality rate.
MATERIALS AND METHODS
Study animals
The field trial was carried out as a prospective study
involving the progeny of 35 in-house bred Landrace-
Yorkshire gilts artificially inseminated with semen from in-
house bred Duroc boars, randomly assigned to Treatment
Group – 16 gilts (vaccinated with E. coli bacterin –
Treatment Group) and Control Group – 19 gilts, from a
farm in Perak with a sow population of 4,000. Gilts were

J. Vet. Malaysia (2013) 25 (1&2):7-10
8
placed into standard size farrowing pens in an intensive
open house system and each litter of respective gilts
consisting of siblings were monitored for 21 days. Cross
fostering of piglets to other sows was strictly prohibited in
this study. Good farm husbandry management and facilities
(heating lamps) were equally provided to both groups.
E. coli vaccine
The vaccine tested is an inactivated vaccine
(Neocolipor–Merial) containing recombinant porcine E. coli
strains with F4 (F4ab, F4ac, F4ad), F5 adhesins and
inactivated field porcine strain with F6, F41 adhesins. This
covers the main adhesins that dominate in neonatal
diarrhoea (Runnels et al., 1987).
E. coli vaccination
Vaccination of the treatment group was done prior to
the study period. The first dose was given by
intramuscular injection in the neck region behind the ear,
5 – 7 weeks before farrowing, with the booster dose given
2 weeks before farrowing. The newborn piglets were
allowed to suckle colostrum.
Evaluated Parameters
The numbers of diarrhoeic piglets observed in each
litter were recorded daily for 14 days. This was done by the
visual observation of every piglet’s anus for stains of
diarrhoea. Litter size and the cumulative weight of siblings
were recorded on Days 1, 7, 14 and 21 using an analogue
weighing scale, with an accuracy of 0.5kg. The average
body weight and the average daily gain were computed.
Piglets that died during the study were recorded as mortality
throughout the 21 days of study. Although, the cause of
death was not thoroughly investigated, piglets crushed by
sow or died due to external trauma were excluded from the
study. All data collected was analysed using Mann-
Whitney-U Test at 95% confidence level.
RESULTS AND DISCUSSION
Diarrhoea percentages
The results for the average percentage of diarrhoea
episodes per litter for Week 1 and 2 are shown in Figure 1.
There were no statistical differences between groups (p >
0.05), despite Treatment group exhibiting lower diarrhoea
episodes in Week 1 and a non-significant slightly higher
diarrhoea score in Week 2 compared to the Control group.
These findings can be attributed partially to the higher
mortality of piglets in the Control group in Week 1 (Figure
1). Piglets with E. coli infection will most commonly suffer
from severe neonatal secretory diarrhoea, leading to the
death. Piglets that had died from the study were omitted, as
no further data could be obtained from them after such
point. These piglets may likely be the ones greatly affecting
the diarrhoea scores, and subsequently the body weight
parameters in the study.
The average diarrhoea percentage per litter was further
analysed on a daily basis for the first week (Figure 2). At
day 1, control group piglets exhibited significantly more
diarrhoea (p < 0.05) and had a higher peak percentage when
compared to the piglets in the treatment group (i.e. 19.0%
versus 13.8%, respectively). On day 2, peak diarrhoea
episodes in both the treatment and control group piglets
were observed.
Figure 1: Average percentage of diarrhoea episodes per litter
in Treatment and Control groups
Figure 2: Average daily percentage of diarrhoea episode per
litter during week 1 in Treatment and Control groups

J. Vet. Malaysia (2013) 25 (1&2):7-10
9
The average percentages of diarrhoea per litter for the
subsequent days were not significantly different (p > 0.05)
and gradually declined in frequency in both groups. The
peak in diarrhoea score at day-2 with subsequent declining
pattern coincides with the manifestation of E. coli infection,
when peak diarrhoea manifestation is usually between 2 to 5
days of age (Schwartz, 2009). Other diseases causing
neonatal diarrhoea were less likely, judging from the clinical
signs and diarrhoeic patterns (Schwartz, 2009).
Growth Performance
The average body weight of the piglets from the Treatment
and Control groups are shown in Figure 3. There were no
significant differences in the average body weights of
piglets (p > 0.05) in both Treatment and Control groups at
all the ages monitored, i.e. day-1, day-7, day-14 and day-21,
despite the higher body weights of 5.7% in the Treatment
group over the Control group, i.e. 5.01kg and 4.74kg
respectively at Day 21.
Figure 3: Average body weight per piglet in Treatment and
Control groups
Figure 4: Average Daily Gain of the piglets in Treatment and
Control groups
The average daily gain (ADG) of the piglets from gilts
vaccinated with E. coli bacterin vaccine and the control
group are shown in Figure 4. Similar to the average body
weight, the ADG of piglets in the Treatment group were
higher than in the Control group, however the differences
were not significant (p > 0.05). Piglets in the Treatment
group had an overall better daily gain of 5.9% over piglets
in the Control group from day 1 – 21.
Mortality Rate
The mortality rate observed for Treatment and Control
groups at Weeks 1, 2 and 3 are shown in Table 1. Piglets in
the Treatment group had a significantly lower mortality (p <
0.05) during the first week as compared with the Control
group (2.6% and 5.9%, respectively). It may be inferred that
the piglets who were highly contributing to the diarrhoea
score in Week 1 in the Control group did not survive to the
second week, whereas, in the Treatment group, piglets
having diarrhoea in Week 1 surviving into the second week
were recorded for the incidence of diarrhoea. The surviving
piglets from a diarrhoeic episode may perform slightly
Table 1: The effects of E. coli vaccination on mortality rate in
Treatment and Control groups
Week Treatment (%) Control (%)
1 2.60a 5.88b
2 1.30 1.60
3 0 1.07
Total Mortality Rate 3.90c 8.56d a, band c, d Values with different superscripts are significantly different (P <0.05)
slower than normal healthy piglets (Edfors-Lilja et al.,
2000), subsequently affecting both the diarrhoea score and
the growth parameters. There was no difference in piglet
mortality in both groups during the second and third week
of age. However, the mortality difference was significantly
lower (p < 0.05) in the Treatment group as compared with
the Control group for the overall period of 1 to 21 days (i.e.
3.9% and 8.6%, respectively). Previous field trials carried
out in Austria and Norway for the same E. coli vaccine
showed piglet mortalities of 4% for the Treatment groups
and 10% in the Control group, similar to the results in this
study. This may suggest the reproducibility of the results
under farm environments.
Piglets in the Control group had consistently higher
neonatal mortality rates when compared to the treatment
group with a peak at days 3 and 4 (1.6%) at first week of
age. The treatment group had peak mortalities at day 4 with
a neonatal mortality rate of 1.3% (Table 2).

J. Vet. Malaysia (2013) 25 (1&2):7-10
10
Table 2: Daily mortality rate of Treatment and Control group
from Day 1 to Day 7
Day Treatment (%) Control (%)
1
2
3
4
5
6
7
0
0.65
0.65
1.30
0
0
0
0.53
1.07
1.61
1.61
0.53
0.53
0
This study did not control for factors that include: (a)
the unavailable data from diarrhoeic piglets that died in the
first week of life, (b) other pathogens (other than E. coli)
involved in neonatal diarrhoea and the diagnosis of the
causes of diarrhoea and mortalities, (c) possible effects of
the environment (cold weather during the study),
management policies and other factors involved in a
actively producing farm (blinded and equal treatment of
diarrhoeic animals with sulphonamide-trimethoprim and
gentamycin sulphate on both groups) to minimize
production losses, and (d) dam effects (variable milk quality
and quantity).
CONCLUSION
In conclusion, this pilot study showed significant
reduction in the first week mortality of neonatal piglets and
Day-one diarrhoeal percentages (p < 0.05) in piglets from
gilts vaccinated with E. coli (Neocolipor – Merial) vaccine.
The result is reproducible as it is in agreement with other
field trials (Anon, 2003) which indicate that E. coli
vaccination in dams could be an alternative way of
moderating mortality due to E. coli in a farm environment.
No significant differences (p > 0.05) were observed in the
overall diarrhoeal percentages and growth parameters at the
other ages monitored. These could be attributed to various
factors, i.e.: (1) unavailable data after the mortalities of
piglets, in particular from the Control group, (2) this study
had no real control over external factors (ambient
temperature, environmental stressors and routine farm
practices such as giving treatment to the diarrhoeic piglets).
More field trials and experimental studies are warranted to
further investigate the effects of E. coli vaccination in gilts
as well as on sows under natural and controlled
environmental conditions to show its benefits on the control
of piglet diarrhoea and subsequent performance of pigs in
farm environments in Malaysia.
CONFLICTS OF INTEREST
None of the authors of this paper has a financial or
personal relationship with other people or organization that
could inappropriately influence or bias the content of the
paper.
ACKNOWLEDGEMENTS
We are grateful to our collaborator, Universiti Putra
Malaysia for availing the results to be presented here.
Partial funding and technical cooperation was provided by
the farm involved in this study. We are grateful to
acknowledge the technical assistance of Chow Guo Hao,
Vania Kiu Tse Ling, Dr Ch’ng Chee Keong and also a few
respectable personnel working in the farm during the course
of this study.
REFERANCES
Anon. (2003). Scientific Discussion on Neocolipor Vaccine.
CVMP/057/98-Rev.3.EMEA/V/C/035. 1-10. Carr, J. (2006). Colibacillosis (infection with Escherichia coli). In:
Whittemore's Science and Practice of Pig Production. Kyriazakis, I.
and Whittemore, C.T. (Eds) 3rd ed., Blackwell Publishing. 296-298. Edfors-Lilja, I. and Wallgren, P. (2000). Escherichia coli and Salmonella
Diarrhoea in Pigs. In: Breeding for Disease Resistance in Farm
Animals.Axford, R.F.E., Bishop, S.C., Nicholas, F.W. and Owen, J.B.
(Eds), CAB international 2000. 253-267.
Fürer, E.C., Cryz, S.J., Dorner, F., Nicolet, J., Wanner, M. and Germanier,
R. (1982). Protection Against Colibacillosis in Neonatal Piglets by Immunization of Dams with Procholeragenoid. Infection and
Immunity, 35:887-894.
Holden, P.J. and Ensminger, M.E. (2006). Swine Health. In: Swine Science.Holden, P.J. and Ensminger, M.E. (Eds), Pearson Education,
Inc. pp. 308-309.
Loh, T.C., Dodds, P.F. and Lean, I.J. (1999). Relationship between Very Low Density Lipoprotein Subtraction 2 and Survival Rate of Pre-
weaning Piglets.J. Vet. Malaysia, 11: 37-39. Loh, T.C., Lim, H.C., Bahaman, A.R. and Foo, H.L. (2006). Prevalence of
Antimicrobial-resistant Escherichia coli Infection in Diarrhoeic
Piglets.J. Vet. Malaysia, 18: 17-20. Riising, H.J., Murmans, M. and Witvliet, M. (2005). Protection Against
Neonatal Escherichia coli Diarrhoea in Pigs by Vaccination of Sows
with a New Vaccine that Contains Purifed EnterotoxicE. coli Virulence Factors F4ac, F4ab, F5, F6 Fimbial Antigens and Het-labile
E. coli Enterotoxin (LT) Toxoid. J. Vet. Med. B, 52: 296-300.
Runnels, P.L. Moseley, S.L. and Moon, H.W. (1987).F41 Pili as Protective Antigens of Enterotoxiggenic Escherichia coli That Produce F41, K99
or Both Pilus Antigens. Infection and Immunity,55: 555-558.
Schwartz, K.J. (2009). Clostridial Diseases in Pigs. In: Proceedings of the 17th Annual Swine Disease Conference for Swine Practitioners, Iowa
State University, Ames. pp. 46-68.
Too, H.L. (1997). Piglet Diarrhoeas. In: A Guide to Pig Disease in Malaysia.Too, H.L. (Ed) Faculty of Veterinary Medicine and Animal
Science, Universiti Putra Malaysia. pp. 31-33.

J. Vet. Malaysia (2013) 25 (1&2):11-14
11
Journal Article
OCCURRENCE OF SALMONELLA AND OTHER ENTERIC MICROBES IN FAECES OF
HOUSE LIZARDS (Hemidactylus frenatus)
Gwen Lo and Saleha A.A*
Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor Malaysia
SUMMARY
Reptiles have been shown to be natural reservoirs of Salmonella and other enteric bacteria and the reptile species close to homes
and eateries are house lizards. This study aimed to determine the occurrence of Salmonella in house lizards at residential and eateries
premises. Fresh faecal samples were collected from live 25 house lizards and 20 pooled dried droppings were collected around premises.
None of the samples were positive for Salmonella. It was probable that a number of lizards may be carrying Salmonella as shown by other
previous studies and in this case they were not shedding the bacteria in the faeces at the time and in the dried faecal droppings,
Salmonellae was probably absent or did not survive the dry and hot condition. Enteric bacteria that were frequently isolated from fresh
droppings were Klebsiella pneumoniae and Citrobacter freundii which were found resistant to amoxicillin + clavulanic and tetracycline.
Keywords: Salmonella, enteric bacteria, house lizards / geckos, antibiotic resistance
INTRODUCTION
Salmonella lives in the intestines of vertebrates and
has been frequently reported in herpatofauna, particularly
reptiles. The presence of Salmonella in reptiles was first
reported in Gila monster and the regal horned lizard
(Chambers et al, 2006). The increase in popularity of exotic
reptiles, such as turtles, snakes and iguanas, as pet animals
has led to increase in reptile-associated salmonellosis, such
as in United States, which is estimated at 93,000 cases
annually (Schroter et al, 2004; Pasmans et al, 2005). One
reptile species that is of interest is the house lizards or
geckos because they are a common sight in residential,
commercial and eateries premises. These house lizards are
native to Southeast Asia but are now invasively distributed
worldwide in other tropical regions and also subtropical
regions such as northern parts of Australia, Africa and
America. They can be seen at night on walls and ceilings of
houses, buildings, eateries, porches and balconies to hunt
for insects that are attracted to lights.
Salmonella presences in reptiles are often
asymptomatic and they shed the organisms, continuously or
intermittently, in the faeces. Humans and pet animals may
be infected if they are in contact with Salmonella-laden
faecal materials. Conditions associated with salmonellosis
in humans include gastroenteritis, bacteraemia, meningitis,
osteomyelitis, peritonitis and pleuritis (Chambers et al,
2006).
Salmonella are ubiquitous in nature and able to
survive for weeks in water and for years in soil and dust in
environmental conditions (such as temperature, humidity
and pH) that are favourable (Todar, 2006). Thus, the
bacteria can survive in dried faeces in the environment for a
period of time. However, the occurrence of Salmonella in
*Corresponding author: Prof. Dr. Saleha Abdul Aziz (Saleha A.A.)
Phone: +603 8609 3459; Email: [email protected].
the faeces was low compared to geckos that were actually
carrying the bacteria (Chan et al, 1982).
Nonetheless, these faeces or droppings are still
considered as route of transmission of the infection. Little is
known on the faecal carrier status of Salmonella in the
house lizards. Apart from Salmonella, not much is known
regarding the occurrence of other pathogenic
microorganisms in their faeces. This is because the
droppings may contaminate food or water supply or the
premises environments and that humans may be in contact
with these faeces. Thus, the aim of this study was to
determine the occurrence of Salmonella and other enteric
microbes in the faeces of house lizards and the antibiotic
resistance of the bacteria isolated.
MATERIALS AND METHODS
Samples collection
The premises identified to trap the live house lizards
were two blocks of a residential college and two cafeterias
in UPM, an apartment block and a water tank on the highest
floor of an apartment building.
A total of 25 live lizards were caught. These lizards
which crept on ceilings and walls were swept down and
then captured using gloved hands. The lizards were then
individually placed in a bottle (5L) containing a small box
in which the lizard can hide inside. The lizards were kept
until they defecated. Fresh faeces were collected within 12
hr after defecation using a sterile glove, placed in a plastic
bag and brought to the Veterinary Public Health
Laboratory, UPM. Most of the lizard defecated within a day
after they were captured but a small number took two days.
Those that did not defecate within two days were fed with a
meal worm until they defecated. Once the faeces were
collected, the lizard was released.

J. Vet. Malaysia (2013) 25 (1&2):11-14
12
Twenty (20) pooled lizard droppings were collected
from different residential and eateries premises. The lizard
droppings could not be differentiated physically in terms of
age but were estimated to be more than 5 - 7 days old. Ten
(10) of the droppings were collected from several eateries
in Seri Kembangan and the other 10 were collected from
various houses in Serdang. These droppings could be found
on walls, furniture tops, window panes and corners and
edges of rooms. The droppings on the floor were not
collected.
Bacteria culture, isolation and identification
Unlike fresh faeces, isolation of enteric microbes was
not done on the dried droppings, that is, isolation of
Salmonella was only conducted on the dried faeces. Each
faecal dropping was added into 10ml of buffered peptone
water in a bottle for pre-enrichment. The mixture was
incubated at 37°C for 24 hr. In the enrichment stage, 1ml of
the preenriched sample was transferred into Rappaport
Vassiliadis Enrichment Broth (Oxoid). The mixture was
then incubated at 37°C for 24 hr. After incubation, a loopful
of the enriched culture was inoculated onto Brilliant Green
Agar (BGA) and Xylose Lysine Tergital 4 (XLT4) agar
(Oxoid) and the plates were incubated at 37°C for 24 hr.
Then, the plates were examined for bacterial growths and
colonial morphology and the cellular morphology was
examined by Gram staining. Different colonies were
subcultured onto the same agar for purification. On BGA,
typical Salmonella colonies appeared red whereas on
XLT4, the bacteria colonies appeared black or black-
centred with a yellow periphery. The suspected Salmonella
colonies from pure cultures were subjected to biochemical
tests for identification which consisted of inoculation into
Triple Sugar Iron agar, Lysine Iron agar, Sulphide Indole
Motility agar and subjected to citrase and urease tests.
Then, suspected Salmonella colonies were subjected to a
serological test, that is, a slide agglutination test using
polyvalent O Salmonella antisera. To isolate other enteric
microbes, the faecal samples were cultured on Blood agar
(Oxoid) and MacConkey agar (Oxoid) to isolate Gram
positive and Gram negative bacteria respectively. All the
different types of colonies obtained were subjected to
appropriate biochemical tests to identify the Gram positive
and Gram negative bacteria as described in Jang et al
(2008).
Antibiotic susceptibility test
Bacteria that were found predominant in the lizards
caught in each premises were subjected to antibiotic
susceptibility test using the disc diffusion method of Kirby
Bauer as described by NCCLS (2002), now known as
Clinical and Laboratory Standards Institute or CLSI. Six (6)
antibiotics were used, namely streptomycin (S, 10µg),
chloramphenicol (C, 30µg), amoxycillin + clavulanic acid
(AMC, 30µg), tetracycline (TE, 30µg), trimethoprim-
sulfamethoxazole (SXT, 25µg), and enrofloxacin (ENR,
5µg). Briefly, each isolate was suspended into 2mL nutrient
broth and standardized to 0.5 Mac Farland standard. A
sterile swab was dipped into the suspension and swabbed
over the entire surface of Mueller Hinton (MH) agar
(Oxoid) plates in three overlapping directions -
horizontally, vertically and obliquely. The plates were
allowed to dry for approximately 5 min. The six antibiotic
discs were dispensed onto the inoculated MH agar plate by
using an antibiotic disc dispenser. After incubation, the
plates were examined against a dark background to clearly
visualize the zone of inhibition around each antibiotic disc.
The zone of inhibition was measured using a calliper. The
measured zones were compared to the zone diameter
interpretive standards breakpoints (NCCLS, 2002) or
following the manufacturer instructions to determine the
susceptibility of the isolates to the antibiotics.
RESULTS AND DISCUSSION
From the study, Salmonella was not isolated from all
the fresh and dried samples. Nine (9) out of 20 dried
droppings that were collected from various residential and
eateries premises showed no bacteria growth. It is probable
that the lizards may be carrying Salmonella but not
shedding the bacteria in the faeces at the time, hence it
would not be present in the sampled faecal droppings. This
is because the study by Fazhana et al (2007) on 32 house
geckos found 31.0% positive, with 43.0% and 28.0% from
houses and eateries, respectively. In that study, Salmonella
was isolated from the gastrointestinal tracts of the house
lizards. A study by Calaway et al (2011) in Townsville,
North Australia found 7% of the house geckos carried
salmonellae in the large intestines. Otokune for et al (2003)
reported Salmonella carriage rate of 32% in the
gastrointestinal tracts of wall gecko (Geckonidae) and 35%
in pooled lizard droppings from various sources. A number
of works had reported that Salmonella is able to survive in
adverse environment for a considerable period of time;
however in the dried lizard droppings sampled, Salmonellae
was probably absent or did not survive the dry and hot
condition. However, according to Otokune for et al (2003),
Salmonella can survive longer in dry as compared to wet
environments – 4 weeks in tap water and up to 6 and 8
weeks in droppings left exposed directly to air and mixed
with dry sand respectively. The frequency of isolation of
different species of enteric bacteria from the faeces of the
house lizards sampled is shown in Table 1.
The predominant enteric bacteria from house lizards at
the residential and eateries premises were Klebsiella
pneumoniae, followed by Citrobacter freundii. Other
bacteria that were isolated from the residential premises
were Klebsiella oxytoca and Proteus penneri whereas from
the eateries were Enterobacter cloacae, Staphylococcus sp
and Bacillus sp. Apart from Salmonella, the enteric
organisms which Gugnani et al (1986) isolated from wall
geckos (Hemidactylus brookei) in Nigeria, included Proteus

J. Vet. Malaysia (2013) 25 (1&2):11-14
13
mirabilis, Pseudomonas aeruginosa, Escherichia coli, K.
pneumoniae and E. cloacae. A number of these enteric
bacteria are opportunistic pathogens that can cause
infections to those who are immunocompromised such as
the young, old, people with debilitating disease, cancer or
Acquired Immunodeficiency Syndrome.
Figure 1 depicts the antimicrobial resistance among all
the K. pneumoniae and C. freundii isolates. The two
bacteria were found resistant to amoxicillin + clavulanic
acid while 80% of K. pneumoniae isolates were resistant to
tetracycline. The bacteria were susceptible to enrofloxacin,
chloramphenicol, trimethoprim + sulfamethoxazole and
streptomycin. The isolated bacteria showed low antibiotic
resistance with no multiple drug resistance (resistant to four
or more antibiotics) detected; it is most likely due to lack of
exposures of the house lizards to environment that were
contaminated with resistant bacteria (Pasmans et al, 2005).
The house lizards are a common sight in residential,
commercial and eateries premises and they indiscriminately
litter the premises with their droppings, also reported by
Otokune et al (2003). Although in this study Salmonella
was not isolated, other studies had shown otherwise. The
presence of house lizards in close proximity to man and the
enteropathogens they may carry could cause infections in
man should man come in contact with enteropathogen-
laden faeces or by contaminating food, water and the floors.
A more detailed epidemiological study with larger sample
size, more premises and wider locations and to include the
zoonotic viruses and enteric parasites would paint a better
picture into the possible role of the house lizards in
sporadic zoonotic infections such as salmonellosis in
households and eateries.
Table 1: Enteric microbes isolated from faeces of house lizards
Bacteria No. of sample with isolates Percentage (%)
Klebsiella pneumoniae 15 44.1
Klebseilla oxytoca 3 8.9
Citrobacter freundii 10 29.4
Proteus penneri 1 2.9
Enterobacter clocae 2 5.9
Staphylococcus sp 2 5.9
Bacillus sp 1 2.9
TOTAL 34 100.0
Figure 1: Antimicrobial resistance among Klebsiella pneumoniae and Citrobacter freundii isolated from house geckos

J. Vet. Malaysia (2013) 25 (1&2):11-14
14
REFERENCES Calaway, Z., A. Thomas, W. Melrose, P. Buttner and R. Speare
(2011). Salmonella Virchow and Salmonella Weltevreden in a
random survey of the Asian house gecko, Hemidactylus frenatus, in houses in Northern Australia. Vector-Borne and
Zoonotic Diseases, 11: 621-625.
Chambers, D.L. and A. C. Hulse (2006). Salmonella serovars in the herpatofauna of Indiana County, Pennsylvania. Applied and
Environmental Microbiology, 72 : 3771 – 3773.
Chan, J.G., C., Shero, L., Young and B. Bereng, B. (1982). Salmonella in two gecko species on the island of Hawaii,
Retrieved 29 December 2012, from http://scholarspace.manoa.
hawaii.edu /bitstream/handle/10123/18 428/fourth-41-47.pdf. Fazhana, I, N. Salim and A.A. Saleha (2007). A study on the
presence of Salmonella spp. in house geckos (Hemidactylus
frenatus). 2nd Proceedings of the Seminar on Veterinary Sciences, Faculty of Veterinary Medicine, UPM, 15 – 19
January 2007, pp. 24-26.
Gugnani, H.C., J.U. Oguike and R. Sakazaki (1986). Salmonellae and other enteropathogenic bacteria in the intestines of wall geckos
in Nigeria. Antonie van Leeuwenhoek, 52: 117 – 120.
Jang, S.S., E.L. Biberstein, and D.C. Hirsch. (2008) A Diagnostic
Manual of Veterinary Clinical Bacteriology and Mycology. Davis : University of California.
NCCLS, National Committee for Clinical Laboratory Standards
(2002). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests For bacteria Isolated from
Animals; Approved Standard – 2nd Edition. Pennsylvania, USA
: NCCLS Otokunefor, T.V., B.I. Kindzeka, I.O Ibiteye, G.U. Osuji, F.O. Obi
and A.W.K. Jack (2003). Salmonella in gut and droppings of
three pest lizards in Nigeria. World Journal of Microbiology & Biotechnology, 19: 545 – 548.
Pasmans, F., A, Martel, F. Boyen, D. Vandekerchove, I. Wybo., F.
Van Immerseel, M. Heyndrickx, J.M. Collard, R. Ducatelle and F. Haesebrouck (2005). Characterization of Salmonella isolates
from captive lizards. Veterinary Microbiology, 110:285-291.
Schroter, M., P. Roggentin, J. Hofmann, A. Speicher, R. Laufs and D. Mack (2004). Pet snakes as a reservoir for Salmonella enteric
subsp. Diarizonae (Serogroup IIIb): a prospectice study.
Applied and Environmental Microbiology, 70: 613 – 615. Todar, K. 2006. Todar's Online Textbook of Bacteriology. University
of Wisconsin, Madison Department of Bacteriology. Available
at: http://www.textbookofbacteriology.net/.

J. Vet. Malaysia (2013) 25 (1&2):15-20
15
Review Article
FELINE HYPERTROPHIC CARDIOMYOPATHY –
PREVALANCE, RISK FACTOR AND PATHOLOGICAL ASPECT
K.H. Khor*
Faculty of Veterinary Medicine, University Putra Malaysia,43400 UPM Serdang, Malaysia
Hypertrophic Cardiomyopathy (HCM)
Prevalence of HCM
HCM is inherited as an autosomal dominant disease in
both humans (Liu et al., 1993; Maron et al., 2003) and cats
(Meurs et al., 2007; Meurs et al., 2005). It is the most
prevalent cardiomyopathy disease in both species; however,
several factors preclude accurate determination of the
disease frequency. It has been estimated to affect 0.1 - 0.2%
of the human population and is the leading cause of sudden
cardiac death in adolescents (including competitive athletes)
(Maron et al., 1995). In cats the prevalence is high and it is
estimated closed to 16% of the overtly healthy cat
population are affected (Cote et al., 2004; Paige et al.,
2009; Riesen et al., 2007a). A retrospective
echocardiographic study reported that 57.5% of cats
initially diagnosed as idiopathic cardiomyopathy (n = 106)
were found to have HCM (Ferasin et al., 2003).
HCM is a disease of young-to-middle age cats between
8 months to 16 year (mean of onset around 6.5 year) (Kraus
et al., 1999; Nakagawa et al., 2002). The youngest
documented was 2 months old (Fujii et al., 2001) and was
commonly observed in males (Fujii et al., 2001; Granstrom
et al., 2011; Liu et al., 1981; Riesen et al., 2007a; Tilley et
al., 1977), a pattern which is not seen in humans. Male cats
were also observed predisposed to HCM earlier in life and
with a more aggressive development of the disease (Atkins
et al., 1992; Rush et al., 2002), compared to female cats.
Generally, cats are a sedentary animal by nature but
stressful situations (e.g. cat fights or chased by dog –
increased sympathetic activity of the heart), may induce
sudden death in cats with asymptomatic HCM (Kittleson
and Kienle, 1998). A cross-sectional echocardiographic
study identified a high frequency of 8 - 10% HCM amongst
overtly healthy cats (Riesen et al., 2007a), which suggested
that substantial changes to the heart were apparent well
before any clinical signs were observed. This asymptomatic
incidence of HCM partially explains episodes of sudden
death or aortic thromboembolism that occurs in apparently
young healthy cats without any obvious clinical signs (Baty
et al., 2001; Cote et al., 2004; Liu and Tilley, 1980; Riesen
et al., 2007b). Two large retrospective studies reported that
the median survival time for cats presented with aortic
thromboembolism, congestive heart failure and
*Corresponding author: Dr Khor Kuan Hua (K.H. Khor);
Phone:+603 8609 3926; Email:[email protected].
asymptomatic cats were between 2 to 6 months, 6 to 18
months and 3 to 5 years, respectively (Atkins et al., 1992;
Rush et al., 2002).
Genetic factors of HCM
Abnormalities in the encoding sarcomeric proteins
identified in HCM patients have led to a theory that it is a
disease of contractile sarcomeric proteins (Marian and
Roberts, 2001). HCM in humans is caused by a genetic
mutation in one of the genes that encode for the sarcomeric
proteins including β-myosin heavy chain (MyHC), cardiac
troponin T and myosin binding protein C gene (MyBP-C)
(Marian and Roberts, 2001; Maron, 2002). Other genes that
accounted for a minority of human HCM cases were cardiac
troponin I, regulatory and essential myosin light chain, titin,
α-tropomysin, α-actin and α-myosin heavy chain (Ommen
and Nishimura, 2004). In human, HCM is inherited, usually
as a heterogeneous autosomal dominant trait, in at least 2/3
of all HCM cases (Marian and Roberts, 2001; Solomon et
al., 1990). To date, more than 1000 different mutations have
been identified within 13 myofilament-related genes in
human (Alcalai et al., 2008; Davies and Krikler, 1994;
Solomon et al., 1990).
In cats, recent studies in Maine Coon and Ragdoll
breeds have identified defects in the same sarcomeric
protein genes, MyBP-C, but in different locations (Kittleson
et al., 1998; Meurs et al., 2008; Meurs et al., 2007; Meurs
et al., 2005), heritable in an autosomal dominant pattern
(Baty and Walkins, 1998; Fananapazir and Epstein, 1994;
Kittleson et al., 1999; Meurs et al., 2005). In addition to
these breeds, there is anecdotal evidence of the familial
heritability of HCM in other breeds, including Persian,
British Shorthair, Norwegian Forest Cat, Turkish Van,
Scottish Fold, Siberian, Sphynx and others (Granstrom et
al., 2011; Meurs et al., 2009; Tilley et al., 2008), although
the exact genetic cause in these breeds have not been well
studied. The familial nature of HCM has also been reported
in domestic (mixed-breed) cats (Kraus et al., 1999;
Nakagawa et al., 2002). A long term observation on the
progression of HCM in a family of domestic shorthair cat
has been documented (Baty et al., 2001).
HCM in Maine Coon cats closely resembles human
familial HCM in terms of mode of inheritance, phenotypic
expression and disease course (Fananapazir and Epstein,
1994; Kittleson et al., 1999; Meurs et al., 2005). Recent
study have found that the MyBP-C gene mutation in Maine
Coon cats is breed specific and may not appears to be

J. Vet. Malaysia (2013) 25 (1&2):15-20
16
associated with the familial HCM in other breed of cats.
The mutation is inherited with incomplete penetrance
(heterozygous) with variable expressivity where not all cats
with the mutation will show the disease or the severity of
the disease may varies among cats (Figure 1) (Carlos
Sampedrano et al., 2009; Lyons, 2010; Meurs et al., 2005).
Carlos Sampedrano et al. (2009) identified 18% of Maine
Coon cats (n = 8/44) with MyBP-C has HCM, hence not all
cats with the identified gene developed HCM. The actual
prevalence may have been underestimated and probably a
long-term follow-up would accurately establish the onset of
disease and its effects on all ages of cats. It was also
suggested that there are likely more than one mutation
responsible for HCM in this breed (Carlos Sampedrano et
al., 2009). In Ragdoll cats, the substitution mutation
identified in the MyBP-C, differs from the Maine Coon cats
because the mutation is located in a different region.
Therefore, Maine Coon and Ragdoll cats’ mutations were
unlikely inherited from a common ancestor. The mode of
inheritance in Ragdoll cats is yet to be identified and this
breed-specific mutation has not been identified in other
breeds of cats. In Ragdoll cats, it was shown that the
homozygous cats appeared to be very severely affected,
often before 2 years of age and the heterozygous cats
appeared to have a milder form of the disease (Meurs et al.,
2007).
In summary, the identification of the first sarcomeric
gene mutation, MyBP-C in Maine Coon cats is highly
significant and supports a role for cats as an alternative to
transgenic mice as a animal model of human familial HCM
(Figure 1) (Baty, 2004; Baty and Walkins, 1998; Hasenfuss,
1998).
/
Unaffected male/female / Affected male/female -------- Mated
The typical features of this mode of inheritance are that it is present in every generation
Figure 1: Example of a colony of cats with familial HCM [an autosomal dominant inheritance with incomplete penetrance
(heterozygous)]
Pathogenesis and progression of HCM
It is a primary disorder of the myocardium
characterised by concentric left ventricular hypertrophy, can
be further described as mild-to-severe (thicken wall of a
normal-to-small chamber size) (Kittleson and Kienle,
1998). To distinguish either: mild hypertrophy from normal,
mild-to-moderate hypertrophy, severe hypertrophy or
secondary due to other abnormalities are often not easy
(Boon, 1998; Fox et al., 1999; Moise et al., 1986). HCM is
phenotypically heterogeneous and no single distribution of
left ventricular hypertrophy is typical of the disease
(Kittleson and Kienle, 1998). Many have compared and
correlated echocardiography findings with other diagnostic
methods such as electrocardiography, necropsy,
histopathology and computed magnetic resonant imaging to
correlated the site or severity of hypertrophy and clinical
findings (Fox, 2003; Sato et al., 1998).
HCM consist of two forms; obstructive and non-
obstructive. Hypertrophy obstructive cardiomyopathy
(HOCM) occurs when the hypertrophied basilar
interventricular septum impinges and results in narrowing
of the left ventricular outflow tract. The mitral valve is
pushed into the outflow tract causing systolic anterior
motion (SAM) of the mitral valve causing further
obstruction of the outflow tract. At the onset of SAM of the
mitral valve in HCM patients, the pushing force of flow is
caused by the dominant hydrodynamic force (Sherrid et al.,
2000), although the Venturi force are present in the outflow
tract but does not contribute to the mechanism of SAM.
Hence, the obstruction may be either dynamic, fixed or both
(Boon, 1998; French, 2008; Takemura et al., 2003). SAM
of the mitral valve is infrequently reported in cats with
HCM but one isolated report by Fox et al. (1995) found that
SAM was a common abnormality identified in 67% of the
HCM cats (n=46). However, this condition may not present
in all HCM cats and some cats may develop this condition
before any evidence of wall thickening or some cats may
have SAM of the mitral valve as a primary disease (Fox et
al., 1995; Kittleson and Kienle, 1998; Klues et al., 1993)
and similar has been widely studied and reported in HCM
patients (Klues et al., 1993; Spirito and Maron, 1984).

J. Vet. Malaysia (2013) 25 (1&2):15-20
17
Therefore, it is important to distinguish between
obstructive and non-obstructive forms of HCM, as clinical
decisions depend on the presence or absence of outflow
obstruction (Kittleson and Kienle, 1998). SAM of the mitral
valve produce two abnormalities where: (i) it obstructs
blood flows out of the left ventricle in systole (dynamic
sub-aortic stenosis) causing an increase blood flow velocity
through the sub-aortic region producing a turbulence and;
(ii) the septal leaflet drawn out from its normal position
creates a mitral regurgitation due to SAM (Kittleson and
Kienle, 1998). Dynamic outflow tract obstruction if severe
will be a stimulus for concentric hypertrophy and
potentially worsen left ventricular diastolic dysfunction.
Hence, coupled with other structural pathological change
(i.e. myocardial fibre disarray, myocardial fibrosis), it may
exacerbate or potentially accelerate progression to heart
failure.
Gross pathology
Cats with severe HCM have severe thickening of the
left ventricular myocardium [the interventricular septum
(IVS) and left ventricular free wall (LVFW)], papillary
muscle hypertrophy and left atrium enlargement with
possible thrombus present. The left ventricular chamber is
smaller than normal due to the inwards myocardial
thickening into the left ventricular cavity (Cesta et al., 2005;
Fox, 2003; Liu et al., 1981; Liu et al., 1993; Tilley et al.,
1977). The distributions of the myocardium thickness in
most cases may not be the same. The IVS and the LVFW
are equally thickened (symmetric hypertrophy) in most cats
with severe HCM. Other cats may have significantly thicker
IVS compared to the LVFW or vice versa. Cats with mild-
to-moderate HCM have lesser myocardial wall thickness,
hypertrophy papillary muscle and probably a normal size of
left ventricular and atrium chamber (Fox et al., 1999;
Kittleson and Kienle, 1998; Tilley et al., 2008). Heart
weight was reported to be a useful indicator for disease
severity or to identify hearts with hypertrophy grossly. The
heart weight in relation to the body weight has been
reported to be 4.8 ± 1.8 g/kg in healthy cats versus 6.0 ±
1.4g/kg in cats with HCM (Fox et al., 1995).
Histopathology
HCM has a wide range of histopathological
abnormalities but myocardial fibre disarray is a distinctive
hallmark of HCM. The myocytes are arranged in chaotic,
disorganised patterns at oblique and/or perpendicular angle
which appeared in a bizarre disorganised cellular
architecture (Baty et al., 2001; Fox, 2003; Kittleson and
Kienle, 1998; Liu et al., 1993; Nakagawa et al., 2002;
Tilley et al., 1977). Other histopathological findings are
hypertrophied myocytes, increased collagen deposition
resulting interstitial fibrosis and abnormalities of the
intramyocardial small vessels (Cesta et al., 2005; Fox,
2003; Liu et al., 1993; Nakagawa et al., 2002; Varnava et
al., 2000). It is likely that these abnormal structural changes
disrupt the transmission of electrophysiology impulses
predisposing the diseased heart to diastolic dysfunction and
ventricular tachyarrhythmia (Fox, 2003; Kittleson and
Kienle, 1998; Liu et al., 1981; Liu et al., 1993; Tilley et al.,
1977). The frequency of each abnormality observed varied
between HCM cats (Liu et al., 1993). Liu et al. (1981)
found 25% of the HCM cats (n=51) observed with
asymmetric left ventricular hypertrophy had myocardial
fibre disarray in the IVS. In other HCM hearts, only
myocyte hypertrophy was evident or some had moderate-to-
severe interstitial. Replacement fibrosis was present in
about 20 - 40% of HCM cases (Liu et al., 1993). The
histopathological findings differ between HCM hearts
probably due to cardiac remodelling which occurs at
different stages of the disease, maybe depending on the
extent of the damage. Hence, whether the asymptomatic
cats with HCM have similar histopathological findings at
the early stage despite detectable functional changes is
unknown and warrant investigation.
Pathophysiology
The pathogenesis of HCM in both humans and cats, to our
knowledge is still not fully understood. It is a known
genetic cardiovascular disease due to the mutations in genes
encoding proteins of the cardiac sarcomere, but the
molecular pathogenesis that leads to the development of
hypertrophy and the variability in the common pathological
phenotypes expression still remain unknown (Abbott, 2010;
Braundwald et al., 2012; Lind et al., 2006; Marian, 2000;
Maron et al., 2009). A mouse model has recently been used
to gain greater understanding of this complex genetic disorder (Berul et al., 2001; Geisterfer-Lowrance et al.,
1996; James et al., 1998; Prabhakar et al., 2003; Shephard
and Semsarian, 2009; Welikson et al., 1999) as studies of
molecular and pathophysiological mechanisms in cats or
human HCM patients is difficult, particularly since the
disease is often well established before diagnosis, especially
Figure 2: Representative photomicrographs of left ventricular
myocardium from a cat with terminal HCM demonstrating
myocardial fibre disarray (H&E stain; x40 magnification)

J. Vet. Malaysia (2013) 25 (1&2):15-20
18
in cats. Pathophysiological changes and/or dysfunction
presence at the early stage of HCM in cats well before the
clinical sign was observed but actual mechanism is still
widely debated. The genetic mutation or the
pathophysiological mechanism is a trigger of the HCM is
still unknown, to our knowledge.
What is understood now are that in any particular HCM
cat, one or more of the pathophysiological changes and
dysfunctions may occur including: (i) left ventricular
diastolic dysfunction; (ii) ventricular and superventricular
tachyarrhythmias; and (iii) myocardial ischemia and fibrosis
(Kittleson and Kienle, 1998; Maron et al., 2009; Tilley et
al., 2008). Coupled with those important derangements,
there are other under-recognised emerging
pathophysiological concerns such as the possible presences
of myocarditis (Bayes-Genis, 2007) and autonomic nervous
dysfunction (Morner et al., 2005) to our knowledge has not
been given emphasis in cats with HCM.
Left ventricular diastolic dysfunction
The main functional implication in cats with HCM is
diastolic dysfunction (Fox et al., 1999; Kittleson and
Kienle, 1998; Tilley et al., 2008). The pathological changes
of the concentric left ventricular hypertrophy from cardiac
remodelling increased myocardial stiffness, increased end
diastolic filling pressure and impaired early diastolic
relaxation (Abbott, 2000; Fox, 2007; Liu et al., 1993).
In HCM, diastolic function is compromised by several
mechanisms. Ventricular compliance is reduced as a result
of left ventricular hypertrophy and small arterial changes
that impair left ventricular perfusion, causing myocardial
ischemia, necrosis and replacement fibrosis (Kitamura et
al., 2001; Kittleson and Kienle, 1998). Compliance is
further compromised by myocardial fibre disarray which
has been shown as the most important factor related to
diastolic dysfunction in humans HCM (Osato et al., 1989)
but assumed to contribute similar consequences in cats with
HCM (Kittleson et al., 1999). Besides that, impaired
sarcolemmal calcium channel regulation and impaired
calcium uptake by the sarcoplasmic reticulum leads to
increased intracellular calcium concentration and impaired
active relaxation (Gwathmey et al., 1991; Opie, 2004).
Ventricular and supraventricular tachyarrhythmias
Correlation between left ventricular hypertrophy and
arrhythmogenic sudden death is well established not only in
HCM, but in other conditions that contribute to left
ventricular hypertrophy (i.e. hypertensive and aortic
stenosis) (Douglas and Tallant, 1991; Piorecka-Makula and
Werner, 2009; Wolk, 2000). The most consistently
observed abnormality that predisposed to arrhythmia is
early or delayed afterdepolarisation and triggered activity
leading to prolong action potential durations and
refractoriness. In addition, non-uniform prolongation of the
action potential may lead to increased dispersion of
repolarisation or refractoriness and favouring re-entry in the
heart (Levick, 2003; Strickland, 1998; Wolk, 2000). In
humans with HCM, the disorganised cellular architecture,
myocardial fibrosis and scarring due to cardiac remodelling
has been suspected as an arrhythmogenic substrate
predisposing to the life-threatening electrical instability
(Marian and Roberts, 2001; Ommen and Nishimura, 2004;
Spirito et al., 1987).
Spirito et al. (1987) documented a strong association of
severe-to-moderate left ventricular hypertrophy with
significantly increased occurrences of ventricular
tachycardia in HCM patients. Cats with a history of
episodic collapse or dyspnoea diagnosed with HCM were
identified with a high frequency of severe ventricular
arrhythmias (i.e. ventricular premature complexes,
ventricular bigeminy, ventricular tachycardia,
supraventricular tachycardia, supraventricular premature
complex) (Bright and Cali, 2000; Ferasin et al., 2003;
Goodwin et al., 1992).
Myocardial ischaemic and fibrosis
Application of positron emission topography and
cardiovascular magnetic resonant has allowed evaluation of
active myocardial ischemia as a determinant of progressive
heart failure in human HCM patients (Harris et al., 2006;
O'Gara et al., 1987; Olivotto et al., 2004). The combination
of increased left ventricular wall thickness (increased
myocardial oxygen demand) and decreased capillary
network (decreased myocardial oxygen supply) will
increase heart rate and afterload, while decreased perfusion
may predispose to myocardial ischemia(Ommen and
Nishimura, 2004).
Presence of myocardial ischemia due to microvascular
dysfunction in HCM has been suggested as an important
pathophysiologic component of the disease progression
(Maron et al., 2009). Cats with HCM have coronary
remodelling (arteriosclerosis or “small vessel disease”)
similar to that described in humans with HCM (Baty et al.,
2001; Nakagawa et al., 2002; Takemura et al., 2003). Liu et
al. (1993) identified intramural coronary arteriosclerosis in
75% (38/51) of cats diagnosed with HCM. Both, Varnava et
al. (2002) and Liu et al. (1993) described a common finding
and relate that the intramural coronary arteriosclerosis are
particularly prominent in tissue sections with moderate-to-
severe fibrosis. There may therefore be a relationship
between arteriosclerosis and myocardial fibrosis. With
compromised blood flow in small vessels triggering
myocardial ischemia, with subsequent cell death (necrosis)
and scarring (fibrosis) affecting the clinical course of the
disease.
Varnava et al. (2000) proposed that myocardial fibre
disarray was a direct response to the functional and
structural abnormalities of the mutated sarcomeric protein,
although the authors also considered that fibrosis and small
vessel disease were secondary response unrelated to
disarray. Myocardial ischemia has been linked to

J. Vet. Malaysia (2013) 25 (1&2):15-20
19
ventricular tachycardia and sudden death (Wolk, 2000) and
its presence in the early stages of HCM may actually be a
pathogenic factor.
REFERANCES Abbott, J.A., 2000. Small animal cardiology secret. Hanley & Belfus,
Philadelphia, Pennsylvania, USA, 371.
Abbott, J.A., 2010. Feline hypertrophic cardiomyopathy: An update. Vet. Clin. North Am. Small Anim. Pract., 40(4): 685-700.
Alcalai, R., Seidman, J.G. and Seidman, C.E., 2008. Genetic basis of
hypertrophic cardiomyopathy: from bench to the clinics. J. Cardiovasc. Electrophysiol., 19(1): 104-110.
Atkins, C.E., Gallo, A.M., Kurzman, I.D. and Cowen, P., 1992. Risk
factors, clinical signs, and survival in cats with a clinical diagnosis of idiopathic hypertrophic cardiomyopathy: 74 cases (1985–1989).
J. Am. Vet. Med. Assoc., 201(4): 613–618. Baty, C.J., 2004. Feline hypertrophic cardiomyopathy: An update. Vet.
Clin. North Am. Small Anim. Pract., 34(5): 1227-1234.
Baty, C.J., Malarkey, D.E., Atkins, C.E., DeFrancesco, T.C., Sidley, J. and Keene, B.W., 2001. Natural history of hypertrophic cardiomyopathy
and aortic thromboembolism in a family of Domestic shorthair cats.
J. Vet. Intern. Med., 15(6): 595-599. Baty, C.J. and Walkins, H., 1998. Familial hypertrophic cardiomyopathy:
Man, mouse and cat. Q. J. Med., 91: 791-793.
Bayes-Genis, A., 2007. Hypertrophy and inflammation: Too much for one heart. Eur. Heart J., 28(6): 661-663.
Berul, C.I., McConnell, B.K., Wakimoto, H., Moskowitz, I.P., Maguire,
C.T., Semsarian, C., Vargas, M.M., Gehrmann, J., Seidman, C.E. and Seidman, J.G., 2001. Ventricular arrhythmia vulnerability in
cardiomyopathic mice with homozygous mutant Myosin-binding
protein C gene. Circulation, 104(22): 2734-2739.
Boon, J.A., 1998. Manual of Veterinary Echocardiography. Lippincott
Williams & Wilkins, Ames, Iowa, USA, 478 pp.
Braundwald, E., Bonow, R.O., Mann, D.L., Zipes, D.P. and Libby, P., 2012. Braunwald's Heart Disease: A textbook of cardiovascular
medicine. Elsevier Saunders, Philadelphia,PA.
Bright, J.M. and Cali, J.V., 2000. Clinical usefulness of cardiac event recording in dogs and cats examined because of syncope, episodic
collapse, or intermittent weakness: 60 cases (1997-1999). J. Am.
Vet. Med. Assoc., 216(7): 1110-1114. Carlos Sampedrano, C., Chetboul, V., Mary, J., Tissier, R., Abitbol, M.,
Serres, F., Gouni, V., Thomas, A. and Pouchelon, J.L., 2009.
Prospective echocardiographic and tissue Doppler imaging screening of a population of Maine Coon cats tested for the A31P
mutation in the myosin-binding protein C gene: A specific analysis
of the heterozygous status. J. Vet. Intern. Med., 23(1): 91-99. Cesta, M.F., Baty, C.J., Keene, B.W., Smoak, I.W. and Malarkey, D.E.,
2005. Pathology of end-stage remodeling in a family of cats with
hypertrophic cardiomyopathy. Vet. Pathol., 42(4): 458-467. Cote, E., Manning, A.M., Emerson, D., Laste, N.J., Malakoff, R.L. and
Harpster, N.K., 2004. Assessment of the prevalence of heart
murmurs in overtly healthy cats. J. Am. Vet. Med. Assoc., 225(3): 384-388.
Davies, M.J. and Krikler, D.M., 1994. Genetic investigation and
counselling of families with hypertrophic cardiomyopathy. Br. Heart J., 72(2): 99-101.
Douglas, P.S. and Tallant, B., 1991. Hypertrophy, fibrosis and diastolic
dysfunction in early canine experimental hypertension. J. Am. Coll. Cardiol., 17(2): 530-536.
Fananapazir, L. and Epstein, N.D., 1994. Genotype-phenotype correlations
in hypertrophic cardiomyopathy. Insights provided by comparisons of kindreds with distinct and identical beta-myosin heavy chain gene
mutations Circulation, 89(1): 22-32.
Ferasin, L., Sturgess, C.P., Cannon, M.J., Caney, S.M.A., Gruffydd-Jones, T.J. and Wotton, P.R., 2003. Feline idiopathic cardiomyopathy: a
retrospective study of 106 cats (1994-2001). J Fel Med Surg, 5(3): 151-159.
Fox, P.R., 2003. Hypertrophic Cardiomyopathy. Clinical and pathologic
correlates. Journal of Veterinary Cardiology, 5(2): 39-45.
Fox, P.R., 2007. Managing feline heart disease - An evidence based
approach, Proceedings of the World Small Animal Veterinary
Association. IVIS, Sydney, Australia, IVIS. Fox, P.R., Liu, S.K. and Maron, B.J., 1995. Echocardiographic assessment
of spontaneously occurring feline hypertrophic cardiomyopathy: An
animal model of human disease Circulation, 92(9): 2645-2651. Fox, P.R., Sisson, D.D. and Moise, N.S., 1999. Textbook of canine and
feline cardiology: Principles and clinical practice., 2nd ed.W.B.
Saunders,Philadelphia, Pennsylvania, USA, 955 pp. French, A., 2008. Feline Cardiomyopathies - An update, Proceedings of the
33rd World Small Animal Veterinary Congress. IVIS, Dublin,
Ireland, pp. 104-106. Fujii, Y., Masuda, Y., Takashima, K., Ogasawara, J., Machida, N.,
Yamane, Y., Chimura, S., Awazu, T., Yamane, T. and Wakao, Y.,
2001. Hypertrophic cardiomyopathy in two kittens. J. Vet. Med. Sci., 63(5): 583-585.
Geisterfer-Lowrance, A.A., Christe, M., Conner, D.A., Ingwall, J.S.,
Schoen, F.J., Seidman, C.E. and Seidman, J.G., 1996. A mouse model of familial hypertrophic cardiomyopathy. Science,
272(5262): 731-734.
Goodwin, J.K., Lombard, C.W. and Ginez, D.D., 1992. Results of continous ambulatory electrocardiography in a cat with hypertrophic
cardiomyopathy. J. Am. Vet. Med. Assoc., 200(9): 1352-1354.
Granstrom, S., Nyberg Godiksen, M.T., Christiansen, M., Pipper, C.B., Willesen, J.T. and Koch, J., 2011. Prevalence of Hypertrophic
Cardiomyopathy in a cohort of British Shorthair Cats in Denmark. J.
Vet. Intern. Med. Gwathmey, J.K., Warren, S.E., Briggs, G.M., Copelas, L., Feldman, M.D.,
Phillips, P.J., Callahan, M., Schoen, F.J., Grossman, W. and
Morgan, J.P., 1991. Diastolic dysfunction in hypertrophic cardiomyopathy. Effect on active force generation during systole. J.
Clin. Invest., 87(3): 1023-1031.
Harris, K.M., Spirito, P., Maron, M.S., Zenovich, A.G., Formisano, F.,
Lesser, J.R., Mackey-Bojack, S., Manning, W.J., Udelson, J.E. and
Maron, B.J., 2006. Prevalence, clinical profile, and significance of
left ventricular remodeling in the end-stage phase of hypertrophic cardiomyopathy. Circulation, 114(3): 216-225.
Hasenfuss, G., 1998. Animal models of human cardiovascular disease,
heart failure and hypertrophy. Cardiovasc. Res., 39(1): 60. James, J.F., Hewett, T.E. and Robbins, J., 1998. Cardiac physiology in
transgenic mice. Circ. Res., 82(4): 407-415.
Kitamura, M., Shimizu, M., Ino, H., Okeie, K., Yamaguchi, M., Funjno, N., Mabuchi, H. and Nakanishi, I., 2001. Collagen remodeling and
cardiac dysfunction in patients with hypertrophic cardiomyopathy: The significance of type III and VI collagens. Clin. Cardiol., 24(4):
325-329.
Kittleson, M.D. and Kienle, R.D., 1998. Small Animal Cardiovascular Medicine. Mosby, St. Louis, USA, 603 pp.
Kittleson, M.D., Meurs, K.M., Kittleson, J.A., Munro, M., Liu, S.K. and
Towbin, J.A., 1998. Heritable characteristics, phenotypic expression, and natural history of hyperttophic cardiomyopathy in
Maine Coon cats (Abstracts). J. Vet. Intern. Med., 12: 198.
Kittleson, M.D., Meurs, K.M., Munro, M.J., Kittleson, J.A., Liu, S.K., Pion, P.D. and Towbin, J.A., 1999. Familial hypertrophic
cardiomyopathy in Maine coon cats - An animal model of human
disease. Circulation, 99: 3172-3180. Klues, H.G., Roberts, W.C. and Maron, B.J., 1993. Morphological
determinants of echocardiographic patterns of mitral valve systolic
anterior motion in obstructive hypertrophic cardiomyopathy. Circulation, 87(5): 1570-1579.
Kraus, M.S., Calvert, C.A. and Jacobs, G.J., 1999. Hypertrophic
cardiomyopathy in a litter of five mixed-breed cats. J. Am. Anim. Hosp. Assoc., 35(4): 293-296.
Levick, J.R., 2003. An introduction to cardiovascular physiology. 3rd Ed.
Arnold, London, UK.
Lind, J.M., Chiu, C. and Semsarian, C., 2006. Genetic basis of
hypertrophic cardiomyopathy. Expert Review Cardiovascular
Therapy, 4(6): 927-934. Liu, S.K., Maron, B.J. and Tilley, L.P., 1981. Feline hypertrophic
cardiomyopathy: Gross anatomic and quantitative histologic
features. Am. J. Pathol., 102(3): 388-395.

J. Vet. Malaysia (2013) 25 (1&2):15-20
20
Liu, S.K., Roberts, W.C. and Maron, B.J., 1993. Comparison of
morphologic findings in spontaneously occuring hypertrophic
cardiomyopathy in humans, cats and dogs. Am. J. Cardiol., 72(12): 944-951.
Liu, S.K. and Tilley, L.P., 1980. Animal models of primary myocardial
diseases. Yale J. Biol. Med., 53(3): 191-211. Lyons, L.A., 2010. Feline genetics: Clinical applications and genetic
testing. Top Companion Animal Medicine, 25(4): 203-212.
Marian, A.J., 2000. Pathogenesis of diverse clinical and pathological phenotypes in hypertrophic cardiomyopathy. Lancet, 355(9197): 58-
60.
Marian, A.J. and Roberts, R., 2001. The molecular genetic basis for hypertrophic cardiomyopathy. J. Mol. Cell. Cardiol., 33(4): 655-
670.
Maron, B.J., 2002. Hypertrophic cardiomyopathy - A systematic review. Journal of the American Medical Association, 287(10): 1308-1320.
Maron, B.J., Gardin, J.M., Flack, J.M., Gidding, S.S., Kurosaki, T.T. and
Bild, D.E., 1995. Prevalence of hypertrophic cardiomyopathy in a general population of young adults: Echocardiographic analysis of
4111 subjects in the CARDIA study. Circulation, 92(4): 785-789.
Maron, B.J., McKenna, W.J., Danielson, G.K., Kappenberger, L.J., Kuhn, H.J., Seidman, C.E., Shah, P.M., Spencer, W.H., III, Spirito, P., Ten
Cate, F.J. and Wigle, E.D., 2003. ACC/ESC clinical expert
consensus document on hypertrophic cardiomyopathy: A report of the American College of Cardiology Task Force on Clinical Expert
consensus Documents and the European Society of Cardiology
committee for Practice Guidelines (Committee to Develop an Expert Consensus Document on Hypertrophic Cardiomyopathy). J. Am.
Coll. Cardiol., 42(9): 1687-1713.
Maron, M.S., Olivotto, I., Maron, B.J., Prasad, S.K., Cecchi, F., Udelson, J.E. and Camici, P.G., 2009. The case for myocardial ischemia in
hypertrophic cardiomyopathy. J. Am. Coll. Cardiol., 54(9): 866-875.
Meurs, K.M., Norgard, M., Haggstrom, J. and Kittleson, M., 2008.
Mutational analysis of four sarcomeric genes in three breeds of cats
with hypertrophic cardiomyopathy. J. Vet. Intern. Med., 22(3): 758-
758. Meurs, K.M., Norgard, M.M., Ederer, M.M., Hendrix, K.P. and Kittleson,
M.D., 2007. A substitution mutation in the myosin binding protein C
gene in ragdoll hypertrophic cardiomyopathy. Genomics, 90(2): 261-264.
Meurs, K.M., Norgard, M.M., Kuan, M., Haggstrom, J. and Kittleson, M.,
2009. Analysis of 8 sarcomeric candidate genes for feline hypertrophic cardiomyopathy mutations in cats with hypertrophic
cardiomyopathy. J. Vet. Intern. Med., 23(4): 840-843. Meurs, K.M., Sanchez, X., David, R.M., Bowles, N.E., Towbin, J.A.,
Reiser, P.J., Kittleson, J.A., Munro, M.J., Dryburgh, K.,
MacDonald, K.A. and Kittleson, M.D., 2005. A cardiac myosin binding protein C mutation in the Maine Coon cat with familial
hypertrophic cardiomyopathy. Hum. Mol. Genet., 14(23): 3587-
3593. Moise, N.S., Dietze, A.E., Mezza, L.E., Strickland, D., Erb, H.N. and
Edwards, N.J., 1986. Echocardiography, electrocardiography, and
radiography of cats with dilatation cardiomyopathy, hypertrophic cardiomyopathy, and hyperthyroidism. Am. J. Vet. Res., 47(7):
1476-1486.
Morner, S., Wiklund, U., Rask, P., Olofsson, B.O., Kazzam, E. and Waldenstrom, A., 2005. Parasympathetic dysfunction in
hypertrophic cardiomyopathy assessed by heart rate variability:
Comparison between short-term and 24-h measurements. Clincal Physiology & Functional Imaging, 25(2): 90-99.
Nakagawa, K., Takemura, N., Machida, N., Kawamura, M., Amasaki, H.
and Hirose, H., 2002. Hypertrophic cardiomyopathy in a mixed breed cat family. J. Vet. Med. Sci., 64(7): 619-621.
O'Gara, P.T., Bonow, R.O., Maron, B.J., Damske, B.A., Van Lingen, A.,
Bacharach, S.L., Larson, S.M. and Epstein, S.E., 1987. Myocardial
perfusion abnormalities in patients with hypertrophic
cardiomyopathy: Assessment with thallium-201 emission computed
tomography. Circulation, 76(6): 1214-1223. Olivotto, I., Cecchi, F. and Camici, P.G., 2004. Coronary microvascular
dysfunction and ischemia in hypertrophic cardiomyopathy.
Mechanisms and clinical consequences. Ital. Heart J., 5(8): 572-580.
Ommen, S.R. and Nishimura, R.A., 2004. Hypertrophic cardiomyopathy.
Curr. Probl. Cardiol., 29(5): 239-291.
Opie, L.H., 2004. Heart Physiology: From Cell to Circulation, 4th edition. Lippincott Williams & Wilkins, Philadelphia, PA, USA, 648 pp.
Osato, K., Shimizu, M., Sugihara, N., Konishi, K. and Takeda, R., 1989.
Histopathological factors related to diastolic function in myocardial hypertrophy. Jpn. Circ. J., 53(6): 595-596.
Paige, C.F., Abbott, J.A., Elvinger, F. and Pyle, R.L., 2009. Prevalence of
cardiomyopathy in apparently healthy cats. J. Am. Vet. Med. Assoc., 234(11): 1398-1403.
Piorecka-Makula, A. and Werner, B., 2009. Prolonged QT dispersion in
children with congenital valvular aortic stenosis. Medical Science Monitor, 15(10): CR534-538.
Prabhakar, R., Petrashevskaya, N., Schwartz, A., Aronow, B., Boivin, G.P.,
Molkentin, J.D. and Wieczorek, D.F., 2003. A mouse model of familial hypertrophic cardiomyopathy caused by a alpha-
tropomyosin mutation. Mol. Cell. Biochem., 251(1-2): 33-42.
Riesen, S.C., Kovacevic, A., Lombard, C.W. and Amberger, C., 2007a. Echocardiographic screening of purebred cats: An overview from
2002 to 2005. Schweiz. Arch. Tierheilkd., 149(2): 73-76.
Riesen, S.C., Kovacevicz, A., Lombard, C.W. and Amberger, C., 2007b. Prevalence of heart disease in symptomatic cats: An overview from
1998 to 2005. Schweiz. Arch. Tierheilkd., 149(2): 65-71.
Rush, J.E., Freeman, L.M., Fenollosa, N.K. and Brown, D.J., 2002. Population and survival characteristics of cats with hypertrophic
cardiomyopathy: 260 cases (1990-1999). J. Am. Vet. Med. Assoc.,
220(2): 202-207. Sato, T., Nakamura, K., Yamanari, H., Yoshinouchi, T. and Ohe, T., 1998.
Relationship between electrocardiographic features and distribution
of hypertrophy in patients with hypertrophic cardiomyopathy. Japanese Circulation Journal-English Edition, 62(7): 483-488.
Shephard, R. and Semsarian, C., 2009. Role of animal models in HCM
research. Journal of Cardiovascular Translation Research, 2(4): 471-
482.
Sherrid, M.V., Gunsburg, D.Z., Moldenhauer, S. and Pearle, G., 2000.
Systolic anterior motion begins at low left ventricular outflow tract velocity in obstructive hypertrophic cardiomyopathy. J. Am. Coll.
Cardiol., 36(4): 1344-1354.
Solomon, S.D., Jarcho, J.A., McKenna, W., Geisterfer-Lowrance, A., Germain, R., Salerni, R., Seidman, J.G. and Seidman, C.E., 1990.
Familial hypertrophic cardiomyopathy is a genetically
heterogeneous disease. J. Clin. Invest., 86(3): 993-999. Spirito, P. and Maron, B.J., 1984. Patterns of systolic anterior motion of
the mitral valve in hypertrophic cardiomyopathy: Assessment by two-dimensional echocardiography . Am. J. Cardiol., 54(8): 1039-
1046.
Spirito, P., Watson, R.M. and Maron, B.J., 1987. Relation between extent of left ventricular hypertrophy and occurrence of ventricular
tachycardia in hypertrophic cardiomyopathy Am. J. Cardiol., 60(14):
1137-1142. Strickland, K.N., 1998. Advances in antiarrhythmic therapy. Veterinary
Clinics of North America Small Animal Practice, 28(6): 1515-1546.
Takemura, N., Nakagawa, K., Machida, N., Washizu, M., Amasaki, H. and Hirose, H., 2003. Acquired mitral stenosis in a cat with hypertrophic
cardiomyopathy. J. Vet. Med. Sci., 65(11): 1265-1267.
Tilley, L.P., Liu, S.K., Gilbertson, S.R., Wagner, B.M. and Lord, P.F., 1977. Primary myocardial disease in the cat. A model for human
cardiomyopathy. Am. J. Pathol., 86(3): 493-522.
Tilley, L.P., Smith, F.W., Oyama, M.A. and Sleeper, M.M., 2008. Manual of Canine and Feline Cardiology. 4th ed. Saunders, Elsevier,
St.Loius, Missouri, USA, 443 pp.
Varnava, A.M., Elliott, P.M., Sharma, S., McKenna, W.J. and Davies, M.J., 2000. Hypertrophic cardiomyopathy: The interrelation of
disarray, fibrosis, and small vessel disease. Heart, 84(5): 476-482.
Welikson, R.E., Buck, S.H., Patel, J.R., Moss, R.L., Vikstrom, K.L.,
Factor, S.M., Miyata, S., Weinberger, H.D. and Leinwand, L.A.,
1999. Cardiac myosin heavy chains lacking the light chain binding
domain cause hypertrophic cardiomyopathy in mice. Am. J. Physiol., 276(6 Pt 2): H2148-2158.
Wolk, R., 2000. Arrhythmogenic mechanisms in left ventricular
hypertrophy. Europace, 2(3): 216-223.

J. Vet. Malaysia (2013) 25 (1&2):21-26
21
Review Article
THE ROLE OF OMEGA-3 POLYUNSATURATED FATTY ACIDS ON BRAIN COGNITIVE
FUNCTION - REVIEW OF STUDIES ON LABORATORY ANIMALS
A. Hafandi*
Faculty of Veterinary Medicine, University Putra Malaysia, Malaysia
OVERVIEW
Fatty acids are essential components of the diet and
sources of food energy. A study carried out by Burr and
Burr, (1929) first acknowledged that specific components of
fatty acids may be necessary for the proper growth and
development of animals and humans. They proposed that
clinical signs of essential fatty acid (EFA) deficiency was
correlated with cardiovascular disease and compromised
immune system. In mammals, the fatty acids have been
recognized as important nutrition factors for brain cognitive
performance. Nutrient supplementation such as omega-3 (ω-
3) fatty acids are widely accepted to improve diets, and
contribute to the maintenance of good health, especially on
the cardiovascular system and other aspects of health, such
as cognition throughout life. Fish oil from tuna and salmon
are the richest source of docosahexaenoic acid (DHA) that
affects the cognitive function of the brain. This could
indicate why fish has been called "brain food" and why
DHA deficiencies can have a life-long impact on
intelligence. Thus, knowledge of health benefits fatty acid as
a dietary trend to increase good health status. In fact, world
health organizations such as the UK Food Standards Agency
and the Japan Society for Lipid Nutrition, advise that more
people should aim to increase their intake of ω-3 fatty acid
supplementation. As these fatty acids have an essential role
during brain development, the first section of this review
examines the importance of ω-3 fatty acid in relation to the
metabolism. This will be followed by a review of the role of
fatty acids on spatial recognition memory such as neural
membrane function, cholinergic neurotransmitter and animal
behavior studies. Experimental works in mice and rats have
shown that ω-3 fatty acid supplementation can increase
levels of synaptic vesicles, neuron dendrites and cholinergic
neurotransmitters. The mechanism physiological of brain
gene expression and cognitive function by ω-3 fatty acid
also will be examines in this section. Thus, this review is
focused on dietary ω-3 fatty acid supplementation on brain
cognitive function and the mechanism interaction between
ω-3 fatty acid and genetic functions on laboratory animals.
Fatty acids
Fat is a substance containing one or more fatty acids
bound to a glycerol backbone (Rudin and Felix, 1996; for
review see Sprecher, 2000). Fats play major roles in the
*Corresponding Author: Dr Hafandi Ahmad (A. Hafandi)
Ph: +603 8609 3416; Email: [email protected].
metabolic, storage and protective functions of the
mammalian body (Gur and Harwood, 1991). Practically,
fatty acids can be from four to twenty-eight carbons in the
chain, with can be classified as short-, medium-, or long-
chained. In addition, fatty acids can also be classified as
saturated fatty acid (SFA) and unsaturated fatty acid (UFA)
(Rudin and Felix, 1996). In SFA, the carbons in the chain
are completely saturated with hydrogen atoms. The results
are a dense and solid fat such as the white fat in beef and
lamb produced butter that does not melt at room temperature
(Rudin and Felix, 1996). However with UFAs, especially
polyunsaturated fatty acids, the carbons carry less hydrogen.
Polyunsaturated fatty acids (PUFAs) are essential for
normal growth and development. The PUFAs are classified
according to the position of the first double bond from the
methyl terminal end. The first double bond in ω-3 is found at
the third carbon atom from the methyl terminal, whereas in
ω-6 the first double bond is located after the sixth carbon
atom from the methyl terminal. The precursors of two
families of PUFAs namely, linoleic acid (LA, ω-6) and -
linolenic acid (ALA, ω-3) are termed essential because they
are cannot be produced by the animal or human body and
must be supplied from the diet (Birberg-Thornberg et al.,
2006). Sources of ω-6 PUFAs and LA are found mainly in
vegetable products such as soybean, corn, nut, and
sunflower oils (Bouziane et al., 1992; Madani et al., 1998).
However, sources of ω-3 fatty acids are based on fish
(menhaden, mackerel, herring, and salmon) and vegetable
(rapeseed, soybean and nut) oils (Aid et al., 2005). However,
the ALA is also found in the chloroplast of green leafy
vegetables, such as spinach, seeds of flax and linseed
(Kitessa et al., 2003).
A significant proportion of the fatty acids are present as
PUFA derivatives of the two parent essential fatty acids,
ALA and LA. The precursor essential fatty acids ALA and
LA are metabolized by a process of desaturation and chain
elongation. These precursors undergo sequential
desaturation through the addition of double bonds and
elongation by addition of carbon atoms. The DHA is
synthesized from ALA by the addition of a double bond by a
Δ6-desaturase to form stearidonic acid (SDA, C18:4n-3).
The elongation of SDA forms eicosatetraenoic acid (ETA,
C20:4n-3) and the addition of another double bond by a Δ5-
desaturase produces eicosapentaenoic (EPA, C20:5n-3). The
elongation of EPA forms docosapentaenoic (DPA, C22:5n-
3), and the final addition of a double bond produces DHA
(for review see Sprecher et al., 1995). The long chain ω-6
fatty acid synthesis such as the elongation of LA to DPA

J. Vet. Malaysia (2013) 25 (1&2):21-26
22
occurs via the same alternating desaturation and elongation
steps.
Role of ω-3 fatty acids on spatial recognition memory
ω-3 fatty acids and neural membrane function
ω-3 fatty acids are the major structural components of
membrane phospholipids (Bertrand et al., 2006). They
influence membrane fluidity and ion transport across cell
membranes (Ehringer et al., 1990). Among the long chain
fatty acids, DHA is important structural components of the
highly lipid biomembrane of neuron cell (for review, see
Lauritzen et al., 2001). Previous studies report that DHA
deficiency caused a reduction in the size of neurons of the
brain region in the hippocampus (Ahmad et al., 2002). These
authors showed that neuron size in the hippocampus,
hypothalamus and parietal cortex are decreased in rats which
are fed a DHA deficient diet containing 1.9gm/100gm
safflower oil compared to those fed on a DHA adequate diet
(Flaxseed oil, 0.48gm/100gm). The authors also reported
that the brains of these rats exhibited a nearly 90% decrease
of DHA. Thus, these results indicate that membrane neuron
cell depends on DHA content for giving an optimal function
in neurons such as signal transduction and synaptic activity.
Further evidence that ω-3 fatty acids promotes neurite
growth in hippocampal neurons comes from the previous
works reporting that DHA supplementation of about 2.6% in
culture increased the population of neurons with longer
neurite length per neuron and with a higher number of
branches (Calderon and Kim, 2004). The author suggested
that rats containing a lower level of DHA (about 0.1%)
showed decreased neurite length, branches and neuron
population, and consequently impaired their cognitive
performance. This was supported by previous studies
reporting that the DHA promotes neurite growth and
survival in hippocampal neurons (Cao et al., 2005). These
findings suggest that an ω-3 fatty acid family, especially
DHA, selectively promotes the development of hippocampal
neurons, which in turn affects the number and quality of
synaptic connections during brain function.
Another approach valuable for the evaluation of the
role of ω-3 fatty acids comes from the previous works
reporting that an ω-3 fatty acid sufficient diet can promote
structural changes in hippocampal neurons (Yoshida et al.,
1997). The studies explained that rats with an ω-3 fatty acid
deficient diet containing safflower oil show a 30% decreased
density of synaptic vesicles in the terminal of the brain
region compared with those in the DHA group (perilla oil).
These results suggest that an ω-3 fatty acid deficient diet
decrease lead to synaptic vesicle functions such as synthesis
movement to releasable sites and fusion with synaptic
membrane in the hippocampal region. Additionally, ω-3
fatty acid supplementation increased the number of synaptic
vesicles (Weisinger et al., 1995) and improved fluidity of
the synaptic membrane which improved interneuron
communication and signal transduction (Ahmad et al., 2002;
Calderon and Kim, 2004; Yoshida et al., 1997).
Functionally, the fluidity of neuronal membranes affects the
signal processing properties of neurons and can improve
neural performance (Ehringer et al., 1990). Overall, it can be
suggested that ω-3 fatty acid is essential in normal
neurogenesis and synaptogenesis, and is also linked to
improvement in learning and memory function.
ω-3 fatty acids and cholinergic neurotransmitter
Acetylcholine (ACh) is a neurotransmitter
which is found in both the peripheral nervous
system and central nervous system (CNS) in many
organisms (Mathew et al., 2007). This
neurotransmitter is released from the brain neurons
to extracellular fluids and plays important roles in
various biological processes such as cognitive
functioning, memory and emotion (Zhang et al .,
2002). In addition, central cholinergic activity
facilitates human and animal cognitive function
(Harmon and Wellman, 2003). A disturbance in the
central cholinergic systems such as decreased ACh
levels are partly responsible for the decline in
cognitive functions in Alzheimer’s patients (Bennett
et al., 2007) and aging humans (Leung et al., 2003).
Previous studies report that cholinergic activity
facilitates long-term potentiation in various areas of
the brain such as cerebral regions and cerebrospinal
fluid levels of choline and acetylcholine (Dash et
al., 2007). Thus, this could indicate that high level
of cholinergic neurotransmitter such ACh is
important for the improvement cognitive and
memory functions.
It is known that ω-3 fatty acids can modulate the ACh
in the brain by diet with ω-3 supplementation in rats (for
review, see Young and Conquer, 2005). Previous studies
report that an increase ACh cerebral levels following
administration of dietary 5% DHA are correlated with an
improvement performance in passive avoidance tasks in a
rats (Minami et al., 1997). This theory is supported by the
finding that ω-3 fatty acid sufficiency from tuna oil acts to
enhance the stimulated synaptic release process of ACh in
the hippocampus, which consequently contributes to the
improvement of learning and memory performance in rats
(Aid et al., 2005; Aid et al., 2003). This could indicate that
ω-3 fatty acid plays a key role in ACh function via changes
in the brain phospholipids composition and may
consequently improve learning and memory function.
A previous study reported that the behavioral and
cognitive changes in rats induced by ω-3 fatty acid
deficiency could be due to changes in cholinergic
neurotransmitters (Delion et al., 1994). Similarly,
cholinergic neurotransmission in the hippocampus is
specifically affected by a diet-induced lack of neuronal ω-3
fatty acid (Aid et al., 2003). Furthermore, there are
functional interactions between cholinergic and
monoaminergic systems that are altered by an ω-3 fatty acid

J. Vet. Malaysia (2013) 25 (1&2):21-26
23
deficient diet (Delion et al., 1994). Both systems contribute
to the impairment of hippocampal function and induce
behavior performance disturbances in rats. Thus, the
hippocampal cholinergic system plays a major role in the
regulation of cognitive functions and its modification might
contribute to the cognitive and behavioral disturbances that
occur in ω-3 fatty acid deficient diet.
ω-3 fatty acids and cognitive functions
A diet with ω-3 fatty acid especially DHA maintains a
fluid synaptic membrane and consequently improves the
ability of learning and memory function in mammals
(Suzuki et al., 1998). Previous studies have shown that ω-3
fatty acid deficient mice demonstrated impaired learning in
the memory version of the Barnes circular maze as they
spent more time and made more errors in search of an
escape tunnel (Fedorova et al., 2007). In the brain’s fatty
acid profiles, this study found a 51% loss of total brain DHA
in mice with an ω-3 fatty acid deficient diet compared to ω-3
fatty acid sufficient mice. Similarly, previous studies report
that the level of brain DHA decreased about 50% in mice on
an ω-3 fatty acid deficient diet compared to control group on
an ω-3 fatty acid sufficient diet (Carrie et al., 1999). These
studies suggest that the ω-3 fatty acid deficient diet
significantly decreased learning performance and retinal
DHA level in adult mice. In a follow up study, the authors
confirmed that an ω-3 fatty acid deficient diet altered fatty
acid composition in brain regions and significantly reduced
spatial learning as well as mice behavior (Carrie et al.,
2000).
Further evidence that ω-3 fatty acid is required for
cognitive function comes from the previous work reporting
that ω-3 fatty acid deficient diet significantly decreased
learning performance in adult mice (Umezawa et al., 1999).
It was observed that mice which were fed on ω-3 fatty acid
deficient diet consisting of 250.3g/100g safflower oil had
significantly reduced learning performance compared with
mice with ω-3 fatty acid sufficient diet (Perilla oil;
0.24g/100g). Deficiency of ω-3 fatty acid in the diet also
leads to reduced brain DHA levels in rats (Moriguchi and
Salem, 2003). The reduced levels of DHA lead to a loss in
brain function as reflected in poorer spatial task
performance. However, the spatial task performance of
DHA deficient rats can be normalized after dietary ω-3 fatty
acid supplementation for 13 weeks to restore brain DHA.
Thus, this could indicate that learning and memory
performance are correlated with the brain level of ω-3 fatty
acid especially DHA.
Many studies have shown that ω-3 fatty acids from fish
oil are essential for normal neurological development
associated in learning and memory function (Carrie et al.,
2000; Chung et al., 2008; Joshi et al., 2004). One example
of such evidence comes from previous work reporting that a
fish oil diet from sardine oil (100g/kg) induced a significant
increase in exploratory activity and learning ability in young
mice (Carrie et al., 2000). A similar effect suggested that
adults mice which were fed on the sardine oil diet for a long
period maintain higher levels of DHA brain phospholipids
and improved learning ability (Suzuki et al., 1998). This was
in line with the reports that fish oil (70g/kg; cod liver oil)
supplementation during pregnancy improved cognitive
performance in dams and their offspring (Chung et al., 2008;
Joshi et al., 2004). In addition, fish oil supplements might
reduce the risk of memory loss or Alzheimer disease in
human populations (Cole et al., 2005). Overall, it is clear
that dietary fish oil is important in neurological development
associated with an improvement in brain cognitive function.
ω-3 fatty acids and animal behavior task
Previous studies have shown that there are several
maze tasks that assess cognitive function including the
Morris water maze (MWM), Y-maze and elevated plus maze
(Abumrad et al., 2005; Astur et al., 2004). Dietary ω-3 fatty
acid deficiency over three generations disrupted learning and
memory performance in the MWM task by adult rats
(Moriguchi and Salem, 2003). In the case of the MWM task,
the ω-3 fatty acid deficient group showed longer escape
latency and delayed acquisition of this task compared with
the ω-3 fatty acid sufficient group. Previous studies report
that rats with DHA (1%) and LNA (3.1%) dietary
supplementation have a shorter escape latency in the MWM
task compared to the rats in DPA (1%) and LA (1%) dietary
supplementation (Lim et al., 2005; Lim et al., 2005).
However, dietary fish oil induced a significant increase in
exploratory and locomotor activity in the MWM test in
young mice (Carrie et al., 2000).
Another approach valuable for evaluation of the role of
ω-3 fatty acid in neuronal and cognitive function comes
from the works reported previously (Bluthe, 2005). This
study demonstrated that mice were fed ω-3 fatty acid
deficient diet containing peanut oil and the control-lipid diet
containing a mixture of peanut and rapeseed oil (1200 mg
LA and 300mg ALA per 100g diet). The spatial task
performance of these animals was then compared using a
two-trial recognition task in the Y-maze. The outcome of
that study indicated that during the acquisition phase, the
activity (number of visits/2-open arms) was the same
whatever the dietary treatment administered. However,
during the restitution phase, 45 min after the end of the
acquisition phase or when the three arms were all open for
exploration, animals in ω-3 fatty acid deficient group were
less able to recognize the new arm compared to the mice fed
the control-lipid diet. Based on these findings, it is quite
evident that ω-3 fatty acid family (especially ALA) provided
in the diet plays a key role in cognitive functions and is
required for optimal brain function.
In the elevated plus maze, a test of anxiety, the time
spent on open arms of the maze was significantly lower in
ω-3 fatty acid deficient mice compared to the sufficient mice
which received rapeseed oil 30g/kg and peanut oil 30g/kg
(Carrie et al., 2000). Similarly, the time spent in open arms
as well as the frequency of entry into the open arms tended

J. Vet. Malaysia (2013) 25 (1&2):21-26
24
to be higher in the ω-3 fatty acid sufficient mice compared to
deficient mice in the elevated plus maze of anxiety protocol
(Nakashima et al., 1993). The findings of increased anxiety
in ω-3 fatty acid deficient animals are supported by previous
studies which also showed that ω-3 fatty acid deficient rats
spent less time in the open arms compared to the sufficient
group, signaling an anxiogenic response. However, after one
week of supplementation with ω-3 fatty acid, the rats
demonstrated a significant improvement in terms of the
number of entries into the open arms. This condition may
explain the improved behavior performance of anxiety
protocol in the elevated plus maze as a result of the ω-3 fatty
acid sufficient diet.
Effect of ω-3 fatty acids on brain gene expression
Fatty acid regulation of gene expression occurs in
unicellular and complex organisms. Fatty acid also plays a
role in controlling gene expression in a variety of tissues
such as nerves and brain tissues (Barcelo-Coblijn et al.,
2003). It has become evident that ω-3 fatty acid can also act
as signaling molecules involved in regulating gene
expression, eicosanoid synthesis and membrane structure
(Duplus et al., 2000). Previous studies have report that
several genes are activated by dietary ω-3 fatty acid and
some gene products have beneficial effect on brain functions
such as learning and memory (Kitajka et al., 2002; Kitajka et
al., 2004; Puskas et al., 2003). Further to this, DNA
microarray technology is a good approach for identifying
changes in transcription of multiple genes in certain brain
regions such as hippocampus (Puskas et al., 2003) and
dentate gyrus (Burger et al., 2007).
As has been reported previously, 55 genes were
detected as overexpressed and 47 were suppressed in rats fed
ω-3 fatty acid supplementation (Kitajka et al., 2004). These
finding shows that the expression of genes can be altered by
DHA supplementation, using cDNA microarray analysis.
Several genes such as transthyretin participating in signal
transduction processes were overexpressed in rat brains
receiving a DHA-enriched diet for one month (Barcelo-
Coblijn et al., 2003). Similarly, transthyretin gene was
expressed by fish oil supplementation in a rat’s brain
hippocampus over one month (Puskas et al., 2003). This
gene binds thyroid hormones and plays an important role in
cognitive function. In such studies, thyroid hormone
deficiency during brain development impairs performance of
cognitive function (Wilcoxon et al., 2007). This finding was
similar in previous studies reporting that a low level of
calmodulin-dependent protein kinase-II activation in a
transgenic mouse model, resulted in enhanced performance
in cognitive function which was associated with an increased
transthyretin transcription (Butler et al., 1995). Thus, these
could indicate that transthyretin gene has a marked
influenced in synaptic plasticity, learning and memory.
Further evidence that ω-3 fatty acid can modulate the
suppression and enhancement of expression genes comes
from the previous studies reporting that ω-3 fatty acid
induces various genes involved in diverse functions in
different brain regions (Kitajka et al., 2004). These findings
proposed an experimental feeding protocol containing perilla
oil, which is rich in ALA (39% ALA/29% LA) and fish oil
rich in DHA (27% DHA/23% LA/3% ALA/12% EPA). This
resulted in gene encoding synuclein α and γ which were
over-expressed. These genes participated in signal
transduction processes, synaptosomes and ion channel
formation. In addition, synuclein possibly related to
cognitive functions in young rats receiving ω-3 fatty acid
from conception until adulthood (Barcelo-Coblijn et al.,
2003). The synuclein is shown to accumulate in the brain of
song birds during the period of song learning (Recchia et al.,
2004). In has been shown that over-expression of synucleins
appear to be associated with the development and maturation
of neurons and neurotransmission (Eslamboli et al., 2007).
Furthermore, genes participating in signal transduction
processes such as calmodulins also were up-regulated by the
dietary of high LNA (perilla oil; 8%) or high EPA + DHA
(fish oil; 8%) (Kitajka et al., 2002). Interestingly,
calmodulins may enhance communication between neurons
during signal transduction process and have a special role in
the stimulant-induced plasticity of the CNS (Jordan et al.,
2007). Overall, dietary ω-3 fatty acid influences the
transcription of key genes involved in cognitive function as
well as being important for normal brain function and
exerting protection against the incidence of
neurodegenerative diseases such as Alzheimer disease.
CONCLUSION
Nutritional status is one of the factors that can
influence learning and memory function in mammals. Food
restriction is also increased the animal to lead learning and
memory deficits. Increasing number of evidence shows that
dietary ω-3 fatty acid in brain cell membranes is important
to improve learning and memory function. In biochemistry
study, ω-3 fatty acids also have important roles to regulate
and modulate brain gene expression associated learning and
memory function. Therefore, future study needs to address
this issue especially dietary ω-3 fatty acid deficiency
interacts with neural membrane function and cholinergic
neurotransmitter associated with cognitive function. The
mechanism by which ω-3 fatty acids modulate the gene
expression associated with cognitive function is yet to be
explored. It is important to understand the possible effect
since this area of study is limited. Thus, a number of
experiments need to be conducted to demonstrate the
mechanism effects of ω-3 fatty acid in brain gene expression
associated with cognitive functions.
REFERANCES
Abumrad, N. A., Ajmal, M., Pothakos, K. and Robinson, J. K. (2005).
CD36 expression and brain function: Does CD36 deficiency impact learning ability? Prostaglandins & other Lipid Mediators, 77, 77-83.

J. Vet. Malaysia (2013) 25 (1&2):21-26
25
Ahmad, A., Moriguchi, T. and Salem, N. (2002). Decrease in neuron size in
docosahexaenoic acid-deficient brain. Pediatrics Neurology, 26, 210-
218. Ahmad, A., Murthy, M., Greiner, R. S., Moriguchi, T. and Salem, N. Jr.
(2002). A decrease in cell size accompanies a loss of
docosahexaenoate in the rat hippocampus. Nutritional Neuroscience, 5(2), 103-113.
Aid, S., Vancassel, S., Linard, A., Lavialle, M. and Guesnet, P. (2005).
Dietary docosahexaenoic acid [22: 6(n-3)] as a phospholipid or a triglyceride enhances the potassium chloride–evoked release of
acetylcholine in rat hippocampus. The Journal of Nutrition, 135,
1008-1013. Aid, S., Vancassel, S., Poumes-Ballihaut, C., Chalon, S., Guesnet, P. and
Lavialle, M. (2003). Effect of a diet-induced n-3 PUFA depletion on
cholinergic parameters in the rat hippocampus. Journal of Lipid Research, 44, 1545-1551.
Astur, R. S., Tropp, J., Sava, S., Constable, R. T. and Markus, E J. (2004).
Sex differences and correlations in a virtual Morris water task, a virtual radial arm maze, and mental rotation. Behavior Brain
Research, 151, 103-115.
Barcelo-Coblijn, G., Hogyes, E., Kitajka, K., Puskas, L. G., Zvara, A., Hackler, Jr., L., Nyakas, C., Penke, Z. and Farkas, T. (2003).
Modification by docosahexaenoic acid of age-induced alterations in
gene expression and molecular composition of rat brain phospholipids. Proceedings of the National Academy of Sciences,
100(20), 11321-11326.
Bennett, B. M., Reynolds, J. N., Prusky, G. T., Douglas, R. M., Sutherland, R. J. and Thatcher, G. R. J. (2007). Cognitive deficits in rats after
forebrain cholinergic depletion are reversed by a novel NO mimetic
nitrate ester. Neuropsychopharmacology, 32, 505-513. Bertrand, P. C., O’Kusky, J. R. and Innis, S. M. (2006). Maternal dietary (n-
3) fatty acid deficiency alters neurogenesis in the embryonic rat
brain. The Journal of Nutrition, 136, 1570-1575.
Birberg-Thornberg, U., Karlsson, T., Gustafsson, P. A. and Duchen, K.
(2006). Nutrition and theory of mind-The role of polyunsaturated
fatty acids (PUFA) in the development of theory of mind. Prostaglandins, Leukotrienes and Essential Fatty Acids, 75, 33-41.
Bluthe, R.-M., Mingam, R., Moranis, A., DeSmedt, V., Lavialle, M.,
Dantzer, R. and LayE, S. (2005). Effects of n-3 fatty acid dietary deficiency on cognitive function and LPS-induced sickness behavior
in mice. Brain, Behavior, and Immunity, 19, e1-e42.
Bouziane, M., Prost, J. and Belleville, J. (1992). Changes in serum and lipoprotein fatty acids of growing rats fed protein-deficient diets with
low or adequate linolenic acid concentrations. The Journal of Nutrition, 122, 2037-2046.
Burr, G. O., & Burr, M. (1929). On the nature and role of the fatty acids
essential in nutrition. J. Biol. Chem, 82, 345-367. Burger, C., López, M. C., Feller, J. A., Baker, H. V., Muzyczka, N. and
Mandel, R. J. (2007). Changes in transcription within the CA1 Weld
of the hippocampus are associated with age-related spatial learning impairments. Neurobiology of Learning and Memory, 87, 21-41.
Butler, L. S., Silva, A. J., Abeliovich, A., Watanabe, Y., Tonegawa, S. and
McNamara, J. O. (1995). Limbic epilepsy in transgenic mice carrying a Ca2+/calmodulin dependent kinase II a-subunit mutation.
Proceedings of the National Academy of Sciences, 92, 6852-6855.
Calderon, F., & Kim, H.-Y. (2004). Docosahexaenoic acid promotes neurite growth in hippocampal neurons. Journal of Neurochemistry, 90, 979-
988.
Cao, D., Xue, R., Xu, J. and Liu, Z. (2005). Effects of docosahexaenoic acid on the survival and neurite outgrowth of rat cortical neurons in
primary cultures. Journal Nutrition Biochemistry, 16, 538-546.
Carrie, I., Clement, M., de Javel, D., Frances, H. and Bourre, J-M. (2000). Specific phospholipid fatty acid composition of brain regions in
mice: Effects of n–3 polyunsaturated fatty acid deficiency and
phospholipid supplementation. The Journal of Lipid Research, 41,
465-472.
Carrie, I., Clement, M., De Javel, D., Frances, H. and Bourre, J. M. (1999).
Learning deficits in first generation OF1 mice deficient in (n-3) polyunsaturated fatty acids do not result from visual alteration.
Neuroscience Letters, 266, 69±72.
Carrie, I., Guesnet, P., Bourre, J-M. and Frances, H. (2000). Diets containing long-chain n-3 polyunsaturated fatty acids affect
behaviour differently during development than ageing in mice.
British Journal of Nutrition, 83, 439-447.
Chung, W.-L., Chen, J-J. and Su, H-M. (2008). Fish oil supplementation of control and (n-3) fatty acid-deficient male rats enhances reference
and working memory performance and increases brain regional
docosahexaenoic acid levels. Journal of Nutrition, 138, 1165-1171. Cole, G. M., Lim, G. P., Yang, F., Teter, B., Begum, A., Ma, Q., Harris-
White, M. E. and Frautschy, S. A. (2005). Prevention of Alzheimer’s
disease: Omega-3 fatty acid and phenolic anti-oxidant interventions. Neurobiology of Aging, 26S, S133-S136.
Dash, P. K., Moore, A. N., Kobori, N. and Runyan, J. D. (2007). Molecular
activity underlying working memory. Learning & Memory, 14, 554-563.
Delion, S., Chalan, S., Harault, J., Galloteau, D., Besnard, J-C. and Durand,
G. (1994). Chronic dietary alpha-linolenic acid deficiency alters dopaminergic and serotoninergic neurotransmission in rats. Journal
of Nutrition, 124, 2466-2476.
Duplus, E., Glorian, M. and Foresti, C. (2000). Fatty acid regulation of gene transcription. The Journal of Biological Chemistry, 275(40).
Ehringer, W., Belcher, D., Wassal, S. R. and Stillwell, W. (1990). A
comparison of the effects of linolenic (18:3[Omega]3) and docosahexaenoic (22:6[Omega]3) acids on phospholipid bilayers.
Chemistry and Physics of Lipids, 54(2), 79-88.
Eslamboli, A., Romero-Ramos, M., Burger, C., Bjorklund, T., Muzyczka, N., Mandel, R. J., Baker, H., Ridley, R. M. and Kirik, D. (2007).
Long-term consequences of human alpha-synuclein overexpression
in the primate ventral midbrain. Brain, 10, 1-17. Fedorova, I., Hussein, N., Martino, C. D., Moriguchi, T., Hoshiba, J.,
Majchrzak, S. and Salem Jr. N. (2007). An n-3 fatty acid deficient
diet affects mouse spatial learning in the barnes circular maze. Prostaglandins, Leukotrienes and Essential Fatty Acids, 77(5-6), 269-
277.
Gurr, M. I., & Harwood, J. L. (1991). Lipid biochemistry. An introduction.
Fourth edition. Chapman and Hall. London, New York, Tokyo,
Melbourne and Madras Press, 69-83.
Harmon, K. M., & Wellman, C. L. (2003). Differential effects of cholinergic lesions on dendritic spines in frontal cortex of young
adult and aging rats. Brain Research, 992, 60-68.
Jordan, S., Regardie, K., Johnson, J.L., Chen, R., Kambayashi, J., McQuade, R., Kitagawa, H., Tadori, Y. and Kikuchi, T. (2007). In
vitro functional characteristics of dopamine D2 receptor partial
agonists in second and third messenger-based assays of cloned human dopamine D2 long receptor signalling. Journal of
Psychopharmacology, 21, 620-627. Joshi, S., Rao, S., Girigosavi, S., Daware, M., Kale, A. and Hegde, M.
(2004). Differential effects of fish oil and folic acid supplementation
during pregnancy in rats on cognitive performance and serum glucose in their offspring. Nutrition, 20, 465- 472.
Kitajka, K., Puskas, L. G., Zvara, A., Hackler, Jr., L., Barcelo-Coblijn, G.,
Yeo, Y. K. and Farkas, T. (2002). The role of n-3 polyunsaturated fatty acids in brain: Modulation of rat brain gene expression by
dietary n-3 fatty acids. Proceedings of the National Academy of
Sciences, 99(5), 2619-2624. Kitajka, K., Sinclair, A. J., Weisinger, R. S., Weisinger, H. S., Mathai, M.,
Jayasooriya, A. P., Halver, J. E. and Puskas, L. G. (2004). Effects of
dietary omega-3 polyunsaturated fatty acids on brain gene expression. Proceedings of the National Academy of Sciences,
101(30), 10931-10936.
Kitessa, S. M., Peake, D., Bencini, R. and Williams, A.J. (2003). Fish oil metabolism in ruminants III. Transfer of n-3 polyunsaturated fatty
acids (PUFA) from tuna oil into sheep’s milk. Animal Feed Science
Technology, 108, 1-14. Lauritzen, L., Hansen, H. S., Jùrgensen, M. H. and Michaelsen, K. F.
(2001). The essentiality of long chain n-3 fatty acids in relation to
development and function of the brain and retina. Progress in Lipid
Research, 40, 1±94.
Leung, L. S., Shen, B., Rajakumar, N. and Ma, J. (2003). Cholinergic
activity enhances hippocampal long-term potentiation in CA1 during walking in rats. The Journal of Neuroscience, 23(28), 9297-9304.
Lim, G. P., Calon, F., Morihara, T., Yang, F., Teter, B., Ubeda, O., Salem,
N. Jr., Frautschy, S. A. and Cole, G. M. (2005). A diet enriched with the omega-3 fatty acid docosahexaenoic acid reduces amyloid burden

J. Vet. Malaysia (2013) 25 (1&2):21-26
26
in an aged Alzheimer mouse model. The Journal of Neuroscience,
25(12), 3032-3040.
Lim, S.-Y., Doherty, J. D. and Salem Jr., N. (2005). Lead exposure and (n-3) fatty acid deficiency during rat neonatal development alter liver,
plasma and brain polyunsaturated fatty acid composition. Journal of
Nutrition, 135, 1027-1033. Madani, S., Lopez, S., Blond, J. P., Prost, J. and Belleville, J. (1998).
Highly purified soybean protein is not hypocholesterolemic in rats
but stimulates cholesterol synthesis and excretion and reduces polyunsaturated fatty acid biosynthesis. The Journal of Nutrition,
128, 1084-1091.
Mathew, S. V., Law, A. J., Lipska, B. K., Davila-Garcıa, M. I., Zamora, E. D., Mitkus, S. N., Vakkalanka, R., Straub, R. E., Weinberger, D. R.,
Kleinman, J. E. and Hyde, T. M. (2007). Alfa-7 nicotinic
acetylcholine receptor mRNA expression and binding in postmortem human brain are associated with genetic variation in neuregulin 1.
Human Molecular Genetics, 16(23), 2921-2932.
Minami, M., Kimura, S., Endo, T., hamaue, N., Hirafuji, M., Togashi, H., Matsumoto, M., Yoshioka, M., Saito, H., Watanabe, S., Kobayashi,
T. and Okuyama, H. (1997). Dietary docosahexaenoic acid increases
cerebral acetylcholine levels and improves passive avoidance performance in stroke-prone spontaneously hypertensive rats.
Pharmacology Biochemistry and Behavior, 58(4), 1123-1129.
Moriguchi, T., & Salem, J. N. (2003). Recovery of brain docosahexaenoate leads to recovery of spatial task performance. Journal of
Neurochemistry, 87, 297-309.
Nakashima, Y., Yuasa, S., Hukamizu, Y., Okuyama, H., Ohhara, T., Kameyama, T. and Nabeshimat, T. (1993). Effect of a high linoleate
and a high dinolenate diet on general behavior and drug sensitivity in
mice. Journal of Lipid Research, 34, 239-247. Puskas, G. S., Kitajka, K., Nyakas, C., Barcelo-Coblijn, G. and Farkas, T.
(2003). Short-term administration of omega 3 fatty acids from fish oil
results in increased transthyretin transcription in old rat
hippocampus. Proceedings of the National Academy of Sciences,
100(4), 1580-1585.
Recchia, A., Debetto, P., Negro, A., Guidolin, D., Skaper, S. D. and Giusti, P. (2004). Alpha-synuclein and Parkinson’s disease. Journal
of the Federation of American Societies for Experimental Biology,
18, 617-626.
Rudin, D., & Felix, C. (1996). Omega-3 oils. New York Press, 215. Sprecher, H. (2000). Metabolism of highly unsaturated n-3 and n-6 fatty
acid. Biochemistry Biophysics Acta, 1486, 219-231.
Sprecher, H., Luthria, D.L., Mohammed, B.S. and Baykousheva, S.P. (1995). Reevaluation of the pathway for the biosynthesis of
polyunsaturated fatty acids. Journal Lipid Research, 36, 2471-2477.
Suzuki, H., Park, S. J., Tamura, M. and Ando, S. (1998). Effect of the long-term feeding of dietary lipids on the learning ability, fatty acid
composition of brain stem phospholipids and synaptic membrane
fluidity in adult mice: A comparison of sardine oil diet with palm oil diet. Mechanisms of Ageing and Development, 101, 119-128.
Umezawa, M., Kogishi, K., Tojo, H., Yoshimura, S., Seriu, N., Ohta, A.,
Takeda, T. and Hosokawa, M. (1999). High-linoleate and high-alpha-linolenate diets affect learning ability and natural behavior in
SAMR1 mice. Journal of Nutrition, 129, 431-437.
Weisinger, H. S., Vingrys, A. J. and Sinclair, A. J. (1995). Dietary manipulation of long-chain polyunsaturated fatty acids in the retina
and brain of guinea pigs. Lipids, 30(5), 471±473.
Wilcoxon, J. S., Nadolski, G. J., Samarut, J., Chassande, O. and Redei, E. E. (2007). Behavioral inhibition and impaired spatial learning and
memory in hypothyroid mice lacking thyroid hormone receptor α.
Behavioural Brain Research, 177(1), 109-116. Yoshida, S., Miyazaki, M., Takeshita, M., Yuasa, S., Kobayashi,T.,
Watanabe, S. and Okuyama, H. (1997). Functional changes of rat
brain microsomal membrane surface after learning task depending on dietary fatty acids. Journal of Neurochemistry, 68, 1269-1277.
Young, G., & Conquer, J. (2005). Omega-3 fatty acids and neuropsychiatric
disorders. Reproduction Nutrition Development, 45, 1-28. Zhang, W., Cao, X., Wan, F., Zhang, S. and Jin, L. (2002). In vivo
determination of the monoamine neurotransmitters in rat brain by
liquid chromatography with a thioctic acid/iridium oxide–palladium
modified electrode. Analytica Chimica Acta, 472, 27-35.

J. Vet. Malaysia (2013) 25 (1&2):27-30
27
Short Communication
MODERATELY-HIGH HUMORAL ANTIBODY RESPONSES TO A H5N2 INACTIVATED VACCINE DID NOT
SUPPRESS SHEDDING OF HIGHLY PATHOGENIC H5N1 AVIAN INFLUENZA VIRUS DURING
CHALLENGE.
S.H. Sharifah1*, K. Hasuzana2,M.N. Suriani2, M. Maizan2, G.H.. Ong2, D. Azizah2, A.R. Omar3and I.Aini3
1Monash University Malaysia,Jalan Lagoon Selatan, 46150 Bandar Sunway, Malaysia 2Veterinary Research Institute, Jalan Sultan Azlan Shah, 31400 Ipoh, Perak,
3Faculty of Veterinary Medicine, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
SUMMARY
Twelve SPF chickens with moderately high-titred HI antibodies of 7 log2 and 8 log2achieved at week 4 post-vaccination after a
single vaccination with an inactivated whole-H5N2 virus vaccine developed at Veterinary Research Institute, Ipoh were selected for
efficacy and shedding studies. The H5N2 vaccine virus shares approximately 88.2% homology to the HA gene of the H5N1 challenge
virus. The chickens with moderately high titred-HI humoral antibodies provided 100% protection against mortality and morbidity after
challenged with a lethal highly pathogenic H5N1 Malaysian strain. The challenge H5N1 virus was reisolated from pooled cloacal swabs
of chickens with HI titres of 7 log2 and 8 log2 at 3 days post challenged, however, was not reisolated from the pooled oropharyngeal
swabs. The virus titre at reisolation was 101EID50/0.1 ml in pooled cloacal samples from both the 7 log2 and 8 log2 HI titred-chickens.
There was no challenge H5N1 virus reisolated from chickens with HI titre of 9 log2. This study demonstrated that chickens with
moderately high HI humoral antibodies protect chickens against clinical disease and mortality did not fully prevent infection, however,
was able to reduce virus shed via the cloaca and oropharynx.
Keywords: Avian Infuenza, Virus, Vaccine
INTRODUCTION
The H5N1 epizootic outbreak has resulted in the
increase in the vaccination against H5N1 in poultry
population of countries such as China, Italy, Mexico,
Pakistan and Indonesia. For these countries, vaccination
seemed to be one of the principal means of combating
highly pathogenic (HP) avian influenza, as vaccination can
control infection and reduce the incidence of clinical
disease, thus reduce viral load in the environment (Swayne
and Suarez, 2007; Capua et al., 2007). As inactivated HP
H5N1 vaccines are not feasible to be made into vaccines,
due to reasons, such as incomplete inactivation which may
result in disease and spread, and difficulty in differentiating
from wild field and vaccine strains, inactivated vaccines
based on reverse genetics and heterologous HA and NA
antigens are therefore the best options. Inactivated vaccines,
however, seemed to give variable results and booseters need
to be given to achieve high immune responses. A study by
Terrigino et al. (2006) showed that vaccination with an
inactivated H5N9 subtype vaccine at 3 weeks and
boostering at 7 weeks of age induced a very high immune
response, GMT 10.3 log2. This immune response was
protective against challenge with 106 EID50/0.1 ml of the
highly pathogenic A/chicken/Yamaguchi/7/2004 H5N1
subtype and suppress shedding after challenge. Studies in
ducks and geese also showed variable results. A study on an
inactivated reverse genetic (RG) vaccine H5N1/PR8 for
*Corresponding author : Assoc. Prof. Dr. Sharifah Syed Mohd Hassan Email: [email protected]
ducks and geese, where the HA gene implicated in virulence
is replaced with a non-pathogenic H5 gene, HI antibodies
were only detectable at week 1 post vaccination (pv), and
by 6 weeks pv, the antibody rose to a high HI titre of 1024
and waned off to a titre of 16 by 43 weeks (Tian et al.,
2005). A bivalent inactivated vaccine of H5N9 +H7N1and a
monovalent H5N3 which was given to 1 day old and 3
weeks old ducks , induced only low titres of 2 log2 - 3 log2
and 3 log2 – 6 log2 respectively, which was achieved at
week 2 post vaccination. Despite the failure to stimulate
significant HI titres, the bivalent vaccine did offer
protection however, did not stop virus replication as seen in
the seroconversion or the rise in antibody titres following
challenge. The monovalent H5N3 vaccine, however,
provided solid protection with no evidence of shedding of
the challenge virus and no serological response to the H5N1
challenge virus (Middleton et al., 2007). Lee et al. (2007),
however, showed that doses or quantity of antigen of the
same HA subtype and boostering are important for
protective efficacy of the vaccine against H5N1 challenge.
In their study, they showed that one dose of 128 HAU and
64 HAU homologous H5 vaccine induced 100% and 50%
protection respectively. Virus shedding was prevented with
the 128 HAU but not with the 64 HAU antigen quantities.
They also showed that two doses at a 3-week interval with
64 HAU as well as an extra one dose of 1024 HAU of
heterologous H5N3 vaccine provided 100% protection and
prevent viral shedding completely. This is in agreement
with studies by Swayne et al. (1999) who showed that there
was a correlation between the antigen quantity or antigenic

J. Vet. Malaysia (2013) 25 (1&2):27-30
28
content and protective efficacy of the vaccine. In contrast,
others reported that high virus titres can still be present in
vaccinated chickens that are protected against clinical
disease (Maas et al., 2009) where in one study with H9N2
vaccine, after vaccination with as much as 128 or 1024
HAU low pathogenic (LP) H9N2 virus and despite the
induction of high antibody titres, the LP H9N2 challenge
virus could still be isolated from the vaccinated chickens
(Choi et al., 2008).
The molecular and antigenic similarities of 96.8 - 100%
between the individual H5 avian influenza strains were
sufficient to elicit solid cross protection against emerging
HP H5N1 viruses, however, there was no positive
correlation between sequence identity and the ability to
reduce the quantity of challenge virus shed. For example, a
study by Kumar et al. (2007) showed that chickens with
high HI titres (> 640) elicited by a reverse genetics H5N3
avian influenza isolate and challenged with the original
H5N3 virus i.e 100% similarity in the HA genes, did not
result in sterile immunity as virus can be reisolated from at
least 1/16 chickens with high antibody titres (HI:160 - 640).
The purpose of the study was, therefore, to determine
the protection and effect on challenge virus shedding by
chickens with moderately high titred-antibody as induced
by a heterologous whole-H5N2 virus vaccine strain, with an
HA gene homology of 88.2% with the challenge H5N1
virus. The practical implication of this trial is that, if
protection is afforded and is effective in decreasing virus
excretion at such moderately high titres, this vaccine need to
be further improved eg. in its antigenic content during
delivery, formulation with effective oil adjuvants and
boostering effect, so as this vaccine can induced high titres
in at least 80% of chickens in a single or multiple
vaccination.
MATERIALS AND METHODS
Viruses
The Malaysian vaccine virus strain A/duck/Malaysia
/8443/04 H5N2 isolated from ducks was developed as an
inactivated whole-virus oil emulsion vaccine at Veterinary
Research Institute, Ipoh. The pre-activation titre was
107.3
EID50/0.1ml. Cleavage site sequencing and the IVPI
showed that the virus is of low pathogenicity. Table 1
showed the comparison between the H5N2 vaccine and
H5N1 challenged strain. The challenge virus strain used
was A/chicken/Malaysia/5858/04 H5N1.
Compared to the challenge virus, the percentage
sequence identity of the HA gene of the vaccine H5N2 and
challenge virus H5N1 was 88.2% by nucleotide sequence
and 90% by amino acid sequence. As for the comparison of
the HAI segment, the nucleotide sequence similarities were
88.3 % and by amino acid sequence was 87.7% (Sharifah et
al., 2012).
Table 1: Characterization of the H5N2 vaccine virus strain
compared to the challenged H5N1 virus
Comparison Percentage
HA gene
Sequence identity
HAI segment
Nucleotide
sequence
88.2 88.3
Amino-acid
sequence
90 87.7
Vaccination of SPF birds and selection of chickens with
high HI titre ≥ 128
Thirty two-week old SPF chickens (raised in Veterinary
Research Institute, Ipoh, Malaysia) were vaccinated with
200µl of the H5N2 vaccine via the SQ route. In our studies
(Sharifah et al., 2012) we found that 100% of birds had
seroconverted by week 4 post vaccination and demonstrated
the highest titres at this week of vaccination. At week 4 post
vaccination, twelve chickens with high HI antibody titres of
7 log2 - 9 log2 (128 - 512) were selected, tagged and
challenged with 200µl H5N1 virus by the intranasal route
with 106.0
EID50/0.1ml of the virus. This dose was earlier
shown to induce full mortality of unvaccinated controls with
a MDT of 36 - 48 hr.
Challenged of the chickens with HPAI H5N1 virus was
conducted in a negative pressure isolator cabinet ventilated
with HEPA-filtered air in a NATA-certified biosafety level-
3 facility of VRI, Ipoh. Water and feed were provided at
libitum. Three SPF birds of the same batch and age that had
not been vaccinated with the H5N2 vaccine were used as
the challenge control. Mean Death Time was determined for
all birds that died.
HI assay
The detection of antibodies after vaccination was
studied by the HI assay performed according to the WHO
manual on Animal Influenza diagnosis and Surveillance
(WHO/CDS/CSR/NCS/2002.5). Serum samples were
diluted 2 fold, with the initial serum dilution at 1:2. Titres >
3 log2 were considered positive. The serological response
was evaluated for all birds before and after vaccination. The
HI test was performed in V-bottom 96 well microtiter plates
with 8 HAU/50µl of homologous inactivatedH5N1 antigen
per well.
Oropharyngeal and cloacal shedding of virus
Clinical signs were monitored daily for one week post-
challenged. Cloacal and oropharyngeal swabs of each of the
chickens were sampled at 3 days post challenge for virus
reisolation. The cloacal and oropharyngeal swabs of
chickens with similar HI titres (i.e birds with HI titres of 7
log2 (128), 8 log2 (256) and 9 log2 (512) were pooled and
virus isolation performed in 9-11 day SPF embryonated
eggs using standard procedures (Krauss et al., 2004).
Personnel of the Virology Lab of VRI who conducted the

J. Vet. Malaysia (2013) 25 (1&2):27-30
29
re-isolation of the virus were blinded by the treatment
group. The presence of H5N1 challenge virus in the
allantoic fluid was detected using the HA test and
confirmed using specific H5-haemagglutination-inhibition
test. Three passages were undertaken and HA test
performed at each passage before the samples were
considered negative. Positive viral titres were expressed as
EID50/0.1ml.
RESULTS
Shedding of challenged virus
Twelve chickens with high HI antibody titres of 7log2-9
log2 (128-512) did not show any clinical signs and survived
the challenged. Table 2 showed the HI titre at which the
chickens were challenged, the morbidity, mortality and
virus re-isolation data. H5N1 virus was however, isolated
from the pooled cloacal swabs of the group with HI titre of
128 and 256, but not from the pooled oropharyngeal swabs.
No virus was isolated from the pooled samples of the other
two groups i.e. the group with HI titre: 256 and HI titre:
512. The H5N1 virus was only detectable at the third
passage in SPF eggs and the titre was calculated to be 1 log
10 EID50/0.1ml. This shows that chicken/s in the HI titre:
128 and 256 group of chickens shed detectable amount of
challenge virus by the cloacal route. This is a small study
limited by space of the BSL-3 facility, to make statistical
inferences of protection associated with these moderately
high titres not possible.
DISCUSSION
From our previous study (Sharifah et al., 2012), we
showed that with a single vaccination of commercial birds
with the inactivated whole H5N2 vaccine that was
developed at VRI, Ipoh, the HI antibody titers did not rise to
high levels. The highest HI titre achieved was 9 log2 (512)
and only in 4% (1/25) of vaccinated chickens. Titres of 7-9
log2 (128-512) was only achieved at week 4 post
vaccination (Sharifah et al., 2012). In this study, out of the
30 SPF chickens vaccinated only 40% (12/30) achieved a
titre 7-9 log2. From the 30 vaccinated birds, only 5, 4 and 3
chickens attained an HI titre of 128, 256 and 512
respectively, at week 4 post vaccination. In this study, we
inoculated 30 SPF birds with the aim of selecting
moderately high HI titred chickens for efficacy and
shedding evaluation. From the 30 vaccinated birds, only 5,
4 and 3 chickens attained an HI titre of 128, 256 and 512
respectively, at week 4 post vaccination. In this experiment
we wanted to determine whether chickens with HI titres of
7-9 log2 (maximum titres achieved using this vaccine in a
single vaccination) can still protect birds against morbidity,
mortality and shedding. There were no clinical signs
observed in chickens after challenged. However, chickens
seemed to excrete virus at 3 days post challenged via the
cloacal route. The loads of virus isolated from the cloaca
however, was very much reduced (101 EID50/0.1 ml)
compared to the load of virus reisolated from the cloaca and
also the oropharynx of the challenged control unvaccinated
groups. This study confirms that heterologous vaccine and
moderately high titred responses protected against clinical
signs and mortality, and significantly decreased shedding
after intranasal challenge, but they did not fully prevent
infection or provide sterile immunity. This inactivated
H5N2 vaccine sharing an 88.2 % nucleotide and 90% amino
acid similarities with the HA gene and protein respectively
of the challenge H5N1 virus, was able to protect chickens
with moderately high HI titres of 7 - 9 log2. Although this
vaccine protected chickens with moderately high titred-HI
antibody against challenged, chickens with lower HI titres
were not protected (Sharifah et al., 2012).
Table 2: Response of 12 SPF chickens (at 6 weeks old) with moderately high HI titres after a single vaccination with the
H5N2 inactivated vaccine and challenged with pathogenic H5N1 virus
Challenged time No of
birds
HI titre No. of
morbidity
No of
mortality
Isolation of H5N1 from pooled samples
(titre of virus)
Week 4 post-
vaccination
(6 weeks old)
5
128
0/12 0/12 H5N1 virus was reisolated at the third
passage in SPF embryonated eggs (EE)
only from the pooled cloacal swabs of
chickens with HI titre of 128 and 256.
Virus load in the cloacal swabs from
chickens of both HI titres was 101
EID50/0.1 ml
4 256
3 512
Controls
6-wks old SPF
chickens
3 < 2 3/3 3/3
H5N1 virus was reisolated from the pooled
samples from cloacal swabs and
orpharyngeal swabs of dead chickens.
H5N1 virus reisolated at 1st passage in
SPF EE. Virus load was 103.6EID50/0.1ml

J. Vet. Malaysia (2013) 25 (1&2):27-30
30
Boostering might result in chickens achieving much
higher HI titres, however, this was not done for this
vaccine.
All the studies showed that no matter how high the
titre achieved by vaccination with a heterologous NA
vaccine, high titres still cannot afford sterile immunity
to all the birds. Some of the reasons for this could be
due to such factors as host genetic and immune system,
immunosuppression by other diseases the level of
virulency of HPAI strains and partial cross protection,
where for example, any one virus that escape the
immunity, has the ability to multiply and replicate in
the susceptible cells of the oropharynx and the
intestines, i.e. areas not reachable by humoral HI
antibodies.
In this study we were not able to isolate virus from
the oropharynx, however, many workers including
Swayne et al. (1999) and Kumar et al. (2007) were
able to isolate the challenge virus from the oropharynx.
The load of virus in the oropharynx was shown to be
higher than in the cloaca. According to Swayne, a
100% homology between the haemagglutinin of
vaccine and the challenge virus can protect birds
against clinical disease but did not result in the
prevention of infection by the challenge virus and
shedding from the oropharynx. The differences in total
amino acid sequence of the HA1 protein of vaccines,
however, was also shown not to correlate with
reductions in challenged virus titres shed from the
oropharynx or cloaca.
Many workers have reported that vaccination
cannot prevent infection whether the vaccine is
heterologous or homologous to the virus challenged
strain, therefore it is essential that as long as
vaccination decreases virus excretion to levels that are
insufficient for virus transmission within poultry
flocks, than the vaccine should be acceptable. The
variable results achieved by the various workers
reflects the diverse variables used in each of the studies
which include the use of different vaccines, and
challenge viruses, different routes of administration of
challenge viruses and the different doses and antigenic
content of the vaccine at delivery.
Controlling of widespread transmission of the
H5N1 virus is a major issue in countries where H5N1
is present and the use of inactivated vaccines is an
effective control strategy (Swayne and Suarez, 2000).
However, for eradication purposes, a mechanism for
the differentiation between infected and vaccinated
chickens needs to be introduced (Capua, 2007). It is
clear that, because of the human health implications of
AI infections, control plans must aim at the elimination
of the infection, based on any strategy that is chosen.
REFERENCES
Capua, I. (2007). Vaccination for notifiable avian influenza in
poultry. Rev Sci Tech. Apr; 26(1): 217-227. Review.
Choi, J.G., Lee, Y.J., Kim, Y.J., Lee, E.K., Jeong, O.M., Sung, H.W.,
Kim, J.H. and Kwon, J.H. (2008). An inactivated vaccine to control the current H9N2 low pathogenic avian influenza in
Korea.J Vet Sci. Mar; 9(1): 67-74.
Krauss S, Walker D, Pryor SP, Niles L, Chenghong L, Hinshaw VS, Webster RG. Influenza A viruses of migrating wild aquatic
birds in North America Vector Borne Zoonotic Dis. Fall;
4(3):177-189. Kumar, M., Chu, H.J., Rodenberg, J. and Krauss, S. (2007).
Association of serologic and protective responses of avian
influenza vaccines in chickens. Avian Diseases, 51: 481-483. Lee, Y.J., Sung, H.W., Choi, J.G., Lee, E.K., Jeong, O.M., Kwon,
Y.K., Kwon, J, H., Song, C.S. and Kimd, J.H. (2007). Effects
of homologous and heterologous neuraminidase vaccines in chickens against H5N1 highly pathogenic avian
influenza.Avian Diseases, Mar; 51 (1 Suppl): 476-478. Maas, R., Tacken, M., van Zoelen, D. and Oei, H. (2009). Dose
response effects of avian influenza (H7N7) vaccination of
chickens: serology, clinical protection and reduction of virus excretion.Vaccine, Jun 2; 27(27):3592-3597.
Middleton, D., Bingham, J., Selleck, P., Lowther, S., Gleeson, L.,
Lehrbach, P., Robinson, S., Rodenberg, J., Kumar, M. and Andrew, M. (2007). Efficacy of inactivated vaccines against
H5N1 avian influenza infection in ducks.Virology, Mar
1;359(1): 66-71. Epub 2006 Oct 9. Sharifah, S.H., Suriani, M.N., Maizan, M., Ong, G.H., Azizah, D.,
Hasuzana, K., Omar, A.R. and I. Aini. (2012). Potency and
efficacy of a low pathogenic H5N2 inactivated vaccine against challenge with a Malaysian H5N1 highly pathogenic avian
influenza virus. JurnalVeterinar Malaysia (submitted)
Swayne, D.E. and Suarez, D.L. (2007). Current developments in avian influenza vaccines, including safety of vaccinated birds
as food.DevBiol (Basel), 7;130:123-133.
Swayne, D.E., Beck, J.R., Garcia, M. and Stone H.D. (1999). Influence of virus strain and antigenic mass on efficacy of H5
avian influenza inactivated vaccines. Avian pathology, 28, 245-
255. Tian, G., Zhang, S., Li,Y., Bu, Z., Liu, P., Zhou, J., Li, C., Shi, J.,
Yu, K. and Chen, H. (2005). Protective efficacy in chickens,
geese and ducks of an H5N1-inactivated vaccine developed by reverse genetics. Virology, Oct 10; 341(1):153-62.
Terregino, C., Toffan, A., Beato, M.S., De Nardi, R., Drago, A. and
Capua, I.(2007).Conventional H5N9vaccine suppresses shedding in specific-pathogen-free birds challenged with HPAI
H5N1 A/chicken/Yamaguchi/7/2004. Avian Diseases, Mar;
51(1 Suppl):495-7.

Guidelines for authors
31
INSTRUCTIONS TO AUTHORS
Jurnal Veterinar Malaysia is a peer-reviewed journal that publishes original research work, reviews, case reports, short communications and
letters to the editor from any veterinary related fields. The copyright of
papers accepted for publication is that of the Veterinary Association Malaysia and the journal is published twice a year. Jurnal Veterinar
Malaysia is published in English.
Submission of manuscripts Submission of manuscripts to the Jurnal Veterinar Malaysia is via email
to [email protected]. An acknowledgment of receiving submission of manuscripts will be provided. Authors must submit articles in WORD
format (saves as .doc or .docx) and not as PDF files. PDF proofs will be
automatically generated from uploaded files and these are used for subsequent reviewing. Queries concerning the submission process or
journal procedures should be sent by e-mail to: [email protected].
The Corresponding Author, who is normally the Author submitting the
paper, will be asked to confirm that the article is original and is not being
considered for peer-reviewed publication elsewhere. Submission also implies that all of the Authors have approved the paper for release and are
in agreement with its content. The Corresponding Author will also be
required to confirm that all Authors have made substantial contributions to (1) the conception and design of the study or acquisition of data or analysis
and interpretation of data, (2) drafting the article or revising it critically for
important intellectual content, and (3) final approval of the version to be submitted. Contributors who do not meet these criteria for authorship
should be listed in an Acknowledgements section. Upon acceptance of the
article by Jurnal Veterinar Malaysia, the Author(s) will be asked to transfer the copyright of the article to the Publisher. This transfer will
ensure the widest possible dissemination of information.
Plagiarism
Plagiarism is globally recognised as a serious academic offence. The
Corresponding Author will be asked to tick a box to confirm acceptance of these guidelines before approval of the PDF file and completion of the
submission process.
Animal welfare
Where animals have been used in a study, the institutional ethical or
animal welfare Authority under which the work was conducted must be stated, along with the specific authorisation reference number and the date
of approval (if applicable). The Jurnal Veterinar Malaysia will reject any paper where there is reason to believe that animals have been subjected to
unnecessary or avoidable pain or distress.
Conflict of interest
At the end of the text, under a subheading "Conflict of interest” statement,
all Authors must disclose any financial and personal relationships with other people or organisations that could influence (bias) their work.
Examples of potential conflicts of interest include employment,
consultancies, stock ownership, honoraria, paid expert testimony, patent applications or registrations. Conflicts of interest may also exist where a
commercial company donates (or provides funding for a study that uses)
drugs, equipment, test kits, vaccines, reagents or other products manufactured or marketed by that company. If no conflicts of interest
exist, this should be stated as "None of the authors of this paper has any
financial or personal relationship with other people or organisations that could inappropriately influence or bias the content of the paper".
Role of the funding source All sources of funding should be declared in an Acknowledgements
section at the end of the text. Authors should declare the role of study
sponsors, if any, in the study design, in the collection, analysis and
interpretation of data, in the writing of the manuscript and in the decision
to submit the manuscript for publication.
PREPARING THE MANUSCRIPT
Title page
A title page must be included at the start of the article. This should give the paper's title, names of Author(s), the name(s) and address(es) of the
institution(s) where the work was done and other Authors' addresses where
these differ. If the article is a Review or Short Communication, this should
be clearly indicated at the left top on the title page. Insert a page break at the bottom of the title page before the Abstract.
Except where all Authors come from the same department, each Author should be identified using a number superscript (1,2,3 etc.), and the
Corresponding Author should be designated by an asterisk (*) as follows,
example:
HEMATOLOGICAL PROFILES OF THE MALAYSIAN BEAR
(Helarctos Malayanus) KEPT IN CAPTIVITY
C.A. Azlan1*, A. Siti Suri1, H. Latiffah1, A.R. Bahaman1, R. Mat Naim2 and L.Kevin3
1Faculty of Veterinary Medicine, Universiti Putra Malaysia (UPM), 43400
UPM, Serdang, Selangor, Malaysia. 2Zoo Negara, Hulu Kelang, 68000 Ampang, Selangor, Malaysia. 3Zoo Taiping, Jalan Pekeliling, Taman Tasik Taiping, 34000 Taiping, Perak.
The contact details of the corresponding Author [telephone number, fax number (if available), and E-mail address] should then be given using the
following format:
* Corresponding author: Dr Azlan Che Mat (C.A. Azlan); Phone No : +603 123 4567; Fax No: +603 234 5678; Email:
If it is desirable to indicate that more than one author contributed equally
to the work, the numeric superscript (a) should be placed after the names. Then below the corresponding author details you should state: a These authors contributed equally to the work. It is not usually acceptable
for all authors to be acknowledged as equal contributors to a study.
Format and layout
Authors submitting papers that are suitable for consideration but do not comply fully with this guide will be asked to amend the text and re-submit.
Articles must be written in the English language. UK English is preferred.
Please use the spelling as printed by The Oxford English Dictionary. Jurnal Veterinar Malaysia will not edit submitted manuscripts for style or
language; therefore Authors are advised to write clearly and simply, and to
have their article checked by English proficient writers before submission.
Use double-spacing except for Title page and References, which
should be in single spacing. Time New Roman, font size 12 point, is preferred for all text (including Tables, Figures, References and all
symbols). A smaller font size maybe used in Tables if space is limiting, and line spacing may also be adjusted in Tables where
necessary. Formatting must not be customized since this impedes
editing.
Continuous line numbers are required throughout the text.
Bacteria, viruses, genes, mutations, genotypes, and alleles should be indicated in italics as appropriate.
1. FULL PAPER (Original Articles)
Manuscripts of a full paper should be arranged as follows: (1) Title page;
(2) Summary and up to five Keywords in alphabetical order to be supplied below the Summary; (3) the main text must be sub-divided into
Introduction, Materials and methods, Results, Discussion, and
Conclusions; (4) Conflict of interest statement; (5) Acknowledgements; (6) References; (7) Tables; (8) Figure legends; (9) Figures (uploaded as
separate files). The sections should not be numbered. The text should not
exceed 5,000 words in length or about 6 pages of the Journal including
tables, illustrations and references.
SUMMARY. The summary should not exceed 250 words (with no sub-headings), which should emphasise objectives/brief background, the
experimental procedure, results and conclusions.

Guidelines for authors
32
KEYWORDS. Please include 3 to 5 keywords representing the main
content of the article.
INTRODUCTION. This should describe briefly the background and any
previously published literatures related to the work. It should indicate the
justification and objective(s) of the work being presented.
MATERIALS AND METHODS. An account of the animals or samples
as well as the experimental design, methods and techniques used should be described. Detailed descriptions that have been cited in appropriate
references should be avoided.
RESULTS. These should be concise and the data should be logically
analysed and presented. Tables and figures illustrating the same data
should be avoided.
DISCUSSION. This section evaluates and interprets the findings and
should not repeat the results. It also compares the result of present study with those of earlier studies and should include the author’s conclusion.
ACKNOWLEDGEMENTS. Only persons who have made substantial contributions to the study should be acknowledged.
REFERENCES. Only pertinent references should be cited. Authors are strongly advised to use reference management software such as EndNote.
However, references should be checked carefully for accuracy and
corrected manually to ensure the format matches exactly the Jurnal Veterinar Malaysia style described below. In the text, the name(s) of
author(s) should be as follows: Lim and Yusof (1985) or (Lim and Yusof,
1985); Saroja (1985a); Saroja (1985b); papers with more than two authors should be referred to by the first author followed by et al. The list of
references should be single spaced arranged in alphabetical order and
further sorted chronologically. All authors’ name should be included.
Titles of journal should be abbreviated according to the ISI Journal Title
Abbreviations. References to books and monograph should include the
name(s) of author(s) or editor(s) followed by the date of publication in brackets, the title of the chapter or article, the full title of the book, the
edition (if any), page referred to, the publisher and place of publication.
The following are examples for guidance.
Book
Turner, H.N. and Yong, S.S.Y. (1990): Quantitative Genetics in Sheep Breeding. Cornell University Press, Ithaca.
Serial Article
Moore, L.J. and Rutter, J.M. (1989): Attachment of Moraxella bovis to
calf corneal cells and inhibition by antiserum. Aust. Vet. J. 66: 39-42.
Paper in Edited Book
Roberts, S.J. (1986): Abortion and other gestational diseases in mares. In: Current Therapy in Theriogenology. Morrow, D. A. (Ed.) 2nd. ed., W.B.
Saunders Co., Philadelphia. pp. 705-710.
Conference Proceedings
Seetha, D.Y., Dahlia, W. and Sivagurunathan, S. (1991): Acetylcysteine
treatment in a cat with acetaminophen toxicosis. In Proceedings of the 7 th Veterinary Association Malaysia Scientific Congress, Seremban,
Veterinary Association Malaysia. pp. 83-84.
Unpublished Materials (e.g. thesis, reports, etc.)
Ali, A.H. (1980): Studies on the prevention of neonatal calf diarrhoea,
MVM Thesis, Glasgow.
Please note:
Insert a page break only after the Title page, after the Abstract with
Keywords, after the References section, between each Table, and
before the Legends to figures.
The Results and Discussion sections must be distinct and not
combined.
Avoid sub-headings in the Discussion section.
References must not be included within the Conclusions section.
2. CLINICAL PAPERS AND CASE REPORTS
These are either short or full papers that follow the overall arrangements described above. However, they may include the following:
i. Introduction: explaining why the case is being reported, with
reference where necessary to previous reports. ii. An account of the history and clinical examination of the case
and laboratory workup sufficiently complete to enable a
similar case to be recognised. iii. The diagnosis and, where applicable, a differential diagnosis.
iv. An account of the treatment and the subsequent course of the
condition. v. An account of the findings at post mortem examination where
appropriate.
vi. A discussion.
3. SHORT COMMUNICATIONS
Short communications are short papers and should follow the requirements
for full manuscripts, but the text must not exceed 2,000 words or 3 printed
pages of the Journal (including tables, figures and plates). The paper should not be divided into conventional sections. They describe either the
results of complete experiments but are less comprehensive than full
length articles or short clinical reports. Headings for the Summary, Keywords, Acknowledgements, Conflict of interest statement and
References should be included, but there should be no other headings or
subheadings in the main text. There should be no more than 10 references in a Short Communication. A Summary of not more than 125 words is
required and up to five Keywords should be supplied below it.
4. REVIEW PAPERS
Review papers are accepted only if they conform to the following criteria:
i. The author or co-author(s) must have publication (in refereed
journals) in the reviewed area.
ii. Provide new knowledge or a high-calibre synthesis or important knowledge.
iii. Critical review of a specific area of scientific research.
iv. Preferably pertinent to the Malaysian context.
Review papers may be commissioned or proposed. Authors wishing to
submit a review article are advised to contact the Editor at [email protected]. Reviews are about 4000 words in length and may
cover any relevant aspect of veterinary science or comparative medicine. Reviews should follow the layout for Original Articles, but with the main
text subdivided as appropriate to the subject matter, starting with a
Summary and Introduction and incorporating Conclusions and a Conflict of interest statement. Sections should not be numbered.
5. LETTER TO EDITOR
The purpose of “Letters to Editor” are:
i. to report preliminary accounts of research findings for publication, and
ii. to discuss or expand on scientific points made in articles
recently published in the Journal.
Letters are limited to ONE (1) printed page. Acceptability of letters will
be decided by the Editorial Board.
Abbreviations and symbols Only standard abbreviations and symbols should be used. Abbreviations in the title and at the beginning of a sentence should be avoided. The full
term for which an abbreviation stands should precede its first use in the
text, e.g. agar gel precipitation tests (AGPT). Standard units of
measurement are exempted. Whenever possible, measurements should be
in the International System of Units.
Manufacturers
Manufacturers and suppliers should be indicated within the text after the
name of the product. For example: 'diazepam (Valium, Roche)' or 'using an infusion pump (Medfusion 2010, Medex)'. Addresses/locations of

Guidelines for authors
33
manufacturers is not necessary and the use of ® or ™ should be avoided.
Note: proprietary names must not appear in the title or summary. Dosage
of drugs should also be given, e.g. 6 x 106 i.u. procaine penicillin (Bipen; Gist-Brocades).
Tables
Tables should be few, simple and typed with double spacing on separate
sheets. Each table should have a title (above the table) that summarizes the
whole table, numbered and cited in sequence using Arabic numerals (i.e. Table 1, 2, 3 etc.), and place after the reference list. Complex tables are not
acceptable and should be summarized rather than tabulated. Footnotes to
tables should be indicated by a, b etc. and typed at the bottom of the relevant table. Information in tables should not be duplicated in figures
and vice versa. The tables should be placed at the end of the main text after
the References but before the Figure Legends, with one table per page.
Figure and illustrations
This should be provided after tables (if present). The Editors will reject Figures of an unacceptable standard or ask the Authors to replace them.
Figure should be numbered in sequence (using Arabic numerals - i.e.
Figure 1, 2, 3 etc), have short title of figure and figure legend. Do not write the legends on the figures. Scale bars must be provided on all
photomicrographs and electron micrographs.
In preparing Figures, Authors should note the following:
Use uniform lettering and sizing of the original artwork.
Use only the following fonts in your figures: Times New Roman,
Arial, Courier, Helvetica, Symbol.
Number the figures according to their sequence in the text.
Provide all figures as a separate files (Figure 1, Figure 2 etc).
Do not included in the main manuscript.
Artwork formats
Regardless of the application used, when your electronic artwork is
finalised, please 'save as' or convert the images to one of the following formats (Note the resolution requirements for line drawings, half-tones,
and line/half-tone combinations given below):
EPS: Vector drawings. Embed the font or save the text as 'graphics'.
TIFF: Colour or greyscale photographs (half-tones): Always use a
minimum of 300 dpi. TIFF: Bitmapped line drawings: Use a minimum of 1,000 dpi.
TIFF: Combinations of bitmapped line/half-tone (colour or greyscale): A
minimum of 500 dpi is required.
DOC, XLS or PPT: If your electronic artwork is created in any of these
Microsoft Office applications, please supply 'as is'.
All Figures will be published in colour on-line, but colour will only be
used in the hard copy of the Jurnal Veterinar Malaysia where it is considered to be essential to the presentation of the paper (colour
reproduction charges may apply).
The Editorial Board reserves the right to reject any manuscript
Proofs The corresponding Author will be advised by the Editor when the paper
has been accepted for publication. One set of page proofs in PDF format
will be sent by e-mail to the corresponding Author.
The Jurnal Veterinar Malaysia will do everything possible to publish an
accepted article quickly and accurately. Therefore, it is important to ensure that all author corrections are sent back to in one communication; Authors
should check proofs very carefully before replying, since inclusion of any
subsequent corrections cannot be guaranteed. Proof reading is the
responsibility of the authors, but final Editor’s correction may also be
incorporated at proof stage. Once the final corrections have been made,
Jurnal Veterinar Malaysia will aim to publish the paper electronically within 6-8 weeks. The hard copy version containing the paper may follow
later, normally within 12 months of acceptance.
Offprints
The Corresponding author will, at no cost, be provided with a PDF file of
the article via e-mail. The PDF file is a watermarked version of the published article and includes a cover sheet with the journal cover image
and a disclaimer outlining the terms and conditions of use.





























