Embed Size (px)
Citation preview

1FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
EDITORIAL
networks (see back page of the issue)and the Coordination Centre. This struc-ture is regarded as too extensive. Duringthe Ninth Meeting of the ESCORENA Co-ordinators held at the Institut NationalAgronomique Paris-Grignon (INA, P-G),23-25 November 2000, the simplificationof the structure of the Network to make itmore compact was discussed. This sug-gestion has been made several times asthe Network is expensive to run. Origina-lly in 1989, the Network was structured onfive crop-based subnetworks: almond,hazelnut, pecan, pistachio and walnut.The subnetworks on Chestnut and Stone-pine were established in 1991 and 1995,respectively. Two disciplinary, rather thanspecies oriented, subnetworks wereadded in 1994: Economics and GeneticResources. Five Coordination Boardmeetings have already been held (Spain,1991; Turkey, 1994; Portugal, 1995; Mo-rocco, 1996 and Italy, 1997). During theFifth Coordination Board Meeting held in1997 in Rome, it was decided that the fo-llowing meeting (the Sixth Coordination
Activities 2000During the year 2000 some important ac-tivities were carried out within the FAO-CIHEAM Interregional Cooperative Re-search Network on Nuts following its pro-gramme (1997-2002). The First Interna-tional Symposium on Stonepine was heldin late February in Valladolid, Spain. ThisSymposium was organized jointly by Jun-ta de Castile and Leon, Instituto Nacionalde Investigación y Tecnología Agraria yAlimentaria (INIA) and the Confederaciónde Organizaciones de Selvicultores deEspaña (COSE) together with the colla-boration of another ten organizations, theFAO-CIHEAM Nut Network included.
Genetic resources inventoriesRegarding the Inventories on Germ-plasm, Research and References, the se-cond Inventory on Hazelnut, following thefirst on Almond, was published after be-ing edited by A.I. Köksal and various co-llaborators. This inventory published as aREU Technical Series, is an importantcompilation of the currently available ha-zelnut genetic resources and informationon on-going research projects and biblio-graphy. In addition, three more invento-ries are being compiled and are at diffe-rent stages of completion. The Inventory onChestnut, edited by G. Bounous, is almostready, the Inventory on Walnut being com-piled by E. Germain is close to completionand the Inventory on Pistachio is being co-llated by N. Kaska and B.E. Ak. All thesecatalogues are being funded by FAO’s Re-gional Office for Europe and the Seed andPlant Genetic Resources Service (AGPS)together with CIHEAM-IAMZ.
Network restructurizationThe current FAO-CIHEAM Research NutNetwork structure is based on nine sub-
Board Meeting) should be held in 1999where steps on this issue would be taken.However, this meeting has been postpo-ned until 26-28 April 2001, to be held inZaragoza, Spain and the renewal processstill stands. There is an ever-increasinglack of funding to organize the Coordina-tion Board meetings for discussion andplanning of future activities. In addition,due to the forthcoming retirement of someliaison officers, it is necessary to under-take a process of change and renewal.Largely the process would be a reductionin the number of subnetworks and jointmanagement.
Response to the NUCIS 8questionnaireA questionnaire requesting informationabout the FAO-CIHEAM Research NutNetwork and this Newsletter was atta-ched to the NUCIS 8 issue. In addition, inorder to update our database, there wasa section to be filled with the completeaddress, telephone, fax and electronicmail address and field of interest (al-
Almond tree blooming in Tarragona, Spain
N U C I SN E W S L E T T E R
Number 9 December 2000Information Bulletin of the Research Network on Nuts (FAO-CIHEAM)
IRTA - Mas Bové Coordination Centre of the Research Network on Nuts
FAO CIHEAMNut Network
CIHEAM

2 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
mond, chestnut, economics, genetic re-sources, pecan, pistachio, stone pine,quality and/or walnut). Also, there was ashort questionnaire about NUCIS to as-sess the level of interest raised by eachindividual section including a place formaking comments. Readers from 27 di-fferent countries sent back the completedforms. However the overall response (7percent) is worrying as it was clearly sta-ted to readers that to carry on receivingthe bulletin is was necessary to fill out theform and return it to us. A short summaryof the answers to the questionnaire ispresented on page 59 of this issue. Themajor species and/or fields of main inter-est were: almond, walnut, pistachio andgenetic resources. The most valuableNUCIS sections considered were: Articlesand Reports and Bibliography. A few in-teresting comments were made on diffe-rent aspects of the Newsletter and mostare already included in this issue. I thankall of the readers who responded to thequestionnaire.
Nut Network on the webFrom July 2000 the FAO European Sys-tem of Cooperative Research Networks inAgriculture (ESCORENA) has opened awebsite Nuts Network at (http://www.fao.org/regional/europe/escorena/nut-crops.html) on which up-to-date infor-mation regarding the Network and sub-networks can be found. This is a usefulstep forward to becoming more global.
NUCIS on the webA short version of the Newsletter (edito-rial, contents and back page) from issuenumber 6 and onwards, is available onthe Internet web pages of both FAO(http://www.fao.org/regional/europe/pu-blic-e/nucis.htm) and CIHEAM (http://www.iamz.ciheam.org/ingles/nucis6.htm).The contents of this Newsletter can bebrowsed through and also copied and pr-inted. This will further help to disburseyour information. In addition, general in-formation on the Nut Network’s activitiescan be found in English at (http://www.iamz.ciheam.org/ingles/nuts.htm).
Contributions to NUCISAs in past NUCIS editorials, we againstress that this Newsletter must be aneffective vehicle of communication for allthe Network members. The pages of thisbulletin are open to all readers who wouldlike to suggest ideas or to express theiropinion about the work developed by theNetwork (activities carried out and plan-ned) or to publish short articles and re-ports on relevant horticultural subjects ofgeneral interest. We receive a sufficientnumber of contributions from the Medite-rranean Basin and overseas for the arti-cles and reports section. However, thesections on news and notes and also on
congresses and meetings are usually di-fficult to cover due to the scarce informa-tion received and thus, contributions aremost welcomed. Otherwise, the Editorhas to report on the issues he is aware of,but certainly there must be many more is-sues on-going throughout the year whichmerit reporting. Also, the place for ‘grey’bibliography (references and documentswhich are difficult to search like Mastersor Ph Theses) is scarcely filled.
The exchange of information betweenNetwork members through the pages ofthis Newsletter is the basis for developingcollaboration. The editing task in the nineNUCIS issues already published hasbeen huge (NUCIS 1, 9 pages; 2, 20 pa-ges; 3, 24 pages; 4, 28 pages; 5, 36 pa-ges; 6, 52 pages, 7, 44 pages, 8, 46 pa-ges and 9, 68 pages). This time-consu-ming major editing can no longer be pro-vided by the Coordination Centre of ourNut Network as time and resources are li-mited. Therefore we are asking contribu-tors who send articles, news, notes, bi-bliographic references, etc., to the diffe-rent sections to provide them well organi-zed and elaborated. Information shouldbe sent in satisfactory English. Contribu-tions could be send through Internetusing the Editor’s email. The alternativeis to provide them on diskette and also inprinted format. This bulletin is reproducedin black and white only, including slidesand photographs. We thank all who havecontributed to this issue. Please sendyour contributions for the next issue,number 10 (December 2001) by the endof October 2001. Finally, we wish all NutNetwork members and collaborators arewarding and Happy 2001.
The Editor
The designations employed and the pre-sentation of material in this publication donot imply the expression of any opinionwhatsoever on the part of the Food andAgriculture Organization of the UnitedNations concerning the legal status ofany country, territory, city or area or of itsauthorities, or concerning the delimitationof its frontiers or boundaries.
This publication contains the collectiveviews of an international group of ex-perts and does not necessarily repre-sent the decisions or the stated policyof the Food and Agriculture Organiza-tion of the United Nations nor of the In-ternational Centre for Advanced Medite-rranean Agronomic Studies of the Orga-nization for the Economic Cooperationand Development.
Contributions should be written conci-sely in English. Please send contributio-ns on paper and diskette (MicrosoftâWord or Word Perfectâ). Authors areresponsible for the content of their pa-pers. Reproduction of the articles is au-thorised, provided that the original sour-ce is clearly stated.
EDITORIAL .......................................... 1
ARTICLES AND REPORTS
• Almond growing in Turkey .......................................... 3
• Almond production and characteristics .........................in Algarve, Portugal ..................................................... 6
• Inbreeding consequences related ..................................to self-compatibility in almond .................................... 9
• Increasing yields and profitability ..................................of almond culture in Israel ......................................... 13
• Three new self-compatible almond cultivars .................from Zaragoza ........................................................... 15
• About the possible relationships between Amygdalus ..webbi Spach and Amygdalus communis L. ............... 17
• Dwarf almond (Amygdalus nana L.) in Yugoslavia .... 19
• Integral study of the ecophysiological responses ..........to water stress: characterization of almond varieties 20
• Identification, isolation and characterization .................of genes related to drought stress in almond ............ 22
• Hazelnut production in Portugal ................................ 24
• Nutritional value of hazelnut (Corylus avellana L.) .... 25
• Postharvest Physiology of Turkish Filbertt ....................(Corylus colurna L.) Seeds ........................................ 27
• Walnut situation in Iran ............................................. 32
• Hedgerow walnut planting system: ...............................experiences in Spain .................................................. 33
• Selection from natural population and hybridization .....of walnut at the Faculty of Agriculture in Novi Sad,, ......Yugoslavia ................................................................. 37
• The taxonomic ranking of Pistacia terebinthuss ............and P. Palaestina ....................................................... 39
• The influences of different irrigation programmes ........on weight of pistachio leaves .................................... 40
• Cutting propagation of mastic tree ................................(Pistacia lentiscus var. Chia Durham.) ...................... 42
• The chestnut: the ultimate energy source nutritionall ...value and alimentary benefits .................................... 44
• The stonepine (Pinus pinea L.) breeding ................... 50programme in Castile-Leon (Central Spain) .............. 50
• Pollen tube growth and fertilization ...............................in carob (Ceratonia siliqua L.) ................................... 55
NOTES AND NEWS
• Consumer awareness ................................................ 58
• Opportunities for walnut growing ..................................and research in the UK .............................................. 58
• Response to NUCIS 8 Questionaire ........................... 59
CONGRESSES AND MEETINGS
• I International Symposium on Stonepine ......................held in Valladolid, Spain ............................................ 60
• V Inter. Congress on Hazelnut held in Oregon, USA .. 60
• III International almond workshop held .........................in Tras-os-Montes, Portugal ..................................... 61
• Spanish workshop on almond technology ....................and economics held in Almería, Spain ....................... 61
• Workshop on soil management and erosion .................control in almond orchards held in Córdoba, Spain .. 62
• Workshop on tree physiology and pathogens ...............and pests of chestnut held in Sopron, Hungary ......... 62
• IX Meeting of the ESCORENA Coordinators ..................held at Grignon, France ............................................. 62
TO BE HELD ................................................................. 63BIBLIOGRAPHY ........................................................... 64BACKPAGE ................................................................... 68
CONTENTS Page
3FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
ARTICLES ANDREPORTS
ALMOND GROWINGIN TURKEY
PRODUCTIONAlmond is one of the most important nutspecies in Mediterranean countries andmore than half of the world production isplaced in this region. Major almond pro-ducing countries and their productionsare given in Table 1.
Turkey is an important nut producingcountry in the world. It produces approxi-mately 4% of world production (Köksal etal 1995). In Turkey, almond productionwhich ranks as the 4th among nuts has in-creased by 39% in the last 15 years (Sen-gül and Emeksiz 1999). Almond produc-tion according to years is presented inTable 2. Almond production in Turkeyshowed some fluctuations during 1978-1987 period, whereas an increase occur-red between 1988-1994. Similar fluctua-tions were observed during the last fewyears (Anonymous, 1997).
Generally, average yield per tree and perhectare is low owing to different factorsas late spring frosts, lack of vegetativepropagation and inefficient management.
The number of mono-crop almond or-chards is limited and most almond treesare planted at field borders. This hasmade it impossible to give a definite figu-re for the amount of land devoted to al-mond culture.
Production could not reach the expectedlevel due to both limiting ecological condi-tions and self or cross incompatibility.Therefore, there are no regularly establis-hed orchards except in Datca township ofMugla province. Irrigated conditions willbe provided for almond growing within theSoutheast Anatolia Project. Thanks to thisirrigation project, increases in productionare expected in the near future.
GROWING DISTRICTSAlmond is grown almost in every region ofTurkey, except the North-East and higherelevations in Eastern Anatolia (Dokuzo-guz and Gülcan, 1979). However, it iswidely spread in the Aegean, Mediterra-nean and South-Eastern regions wheredry climate and poor soil conditions deter-mine the fruit species to be grown. Re-cently, the number of trees has been in-creasing especially in the western part ofTurkey. Izmir, Mugla and Denizli are themost important almond growing provinces
in this region. Mugla is the most conspi-cuous among the others with regard toproduction of almond. Almond trees arewidely grown in Datça district of Mugla.They are one of the main sources of agri-cultural income for the growers in the re-gion. In some microclimates such as Datca,there are still some problems in almondgrowing even though the temperature de-creases to below zero very rarely and mostof the orchards have been establishedusing grafted trees. One of the reasons ofunsuccessful almond growing in this area isthe bacterial cancer disease, Pseudomo-nas amygdali (Gündogdu, 1990).
The important production areas of the Ae-gean and Marmara regions are Aydin, Iz-mir, Denizli, Isparta, Burdur and Ca-nakkale. There are almond populationswhich have thick hull in Acipayam andTavas township of Denizli provinces andKeçiborlu, Uluborlu and Senirkent town-ship of Isparta province. Thin hull andgood kernels are obtained from the pro-duction areas in Cesme, Menemen andFoça in Izmir, and Didim in Aydin andUmurlu in Canakkale provinces. Almondtypes which have thin and hard shell arefound in Dösemealti, Side, Alanya andGazipasa towns in Antalya (Dokuzoguz et
al., 1968; Dokuzoguz and Gülcan, 1972).On the other hand, almonds in centralAnatolia have very thick hull and are small.The important production areas ofSoutheastern Anatolia are Mardin, Elazigand Diyarbakir.
There are good prospects for almondgrowing in Harran Plain in Sanliurfa.Drying, dehulling, shelling, storing andtransportation of almonds are much easierdue to suitable ecological conditions (Kas-ka, 1999). In addition, Upper Firat river ba-sin and Çoruh valley are the most importantmicro climate areas in the North EasternAnatolia (Aslantas and Güleryüz 1999).
CLIMATETemperatureTurkey is situated between 36°N and 42°Nlatitude and extends for nearly 2000 kilo-metres in the east-west direction, and is atransition between the temperate and sub-tropical climates. This has made it possibleto grow many fruit species in this country.
The almond requires less winter chillingthan most of the other deciduous fruitspecies. A relatively short and mild winterexists in those regions. Temperature inNovember and December is adequate for
Table 1. Main almond producing countries and their production (t)
1989-91 1997 1998
World 1.288.000 1.598.000 1.268.000USA 414.000 574.000 393.000Spain 278.000 367.000 217.000Italy 106.000 105.000 88.000Iran 67.000 76.000 76.000Syria 27.000 26.000 67.000Morocco 61.000 87.000 66.000Tunisia 42.000 51.000 59.000Pakistan 31.000 49.000 49.000Lebanon 13.000 38.000 39.000Greece 57.000 43.000 35.000Turkey 46.000 33.000 34.000
Table 2. Number of bearing and non-bearing almond trees and production valuesbetween 1987 and 1997
Year Nr. of bearing Nr. of non-bearing Productiontrees (x 1000) trees (x 1000) (t)
1987 4054 761 330001988 4034 735 420001989 4040 752 460001990 4040 775 460001991 4019 755 460001992 3980 747 470001993 3965 730 480001994 3906 723 470001995 3865 700 370001996 3825 677 430001997 3775 640 33000

4 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
the dormancy period in coastal line of Me-diterranean region. Average temperatu-res in January and February cause earlyflowering. For this reason, most almondtrees bloom in January. Thus, this crop isconsumed as green fresh almond. Thefirst green almonds come to the marketfrom this area. The lowest temperatureranges from -1. 6°C (Finike) to -6. 2°C(Adana) in that region every 10 years(Gülcan, 1982). It is seen that flowers andgreen almond are not generally damagedby frost. On the other hand, in the areasexcept those at sea level, precautionmust be taken to prevent frost damage foralmond growing. In these areas such asOsmaniye, Islahiye and Mut, choosinglate flowering types is necessary. Springfrost always occurs in important growingdistricts such as Akseki, Burdur and Is-parta. Thus, it is determined that produc-tion losses occur quite often. Controver-sy, Southeastern Anatolia is too safe interms of late spring frosts.
RainfallRegarding almond production, the Ae-gean and Mediterranean regions are ofgreat importance. In the Aegean region,the amount of annual rainfall varies from600 to 900 mm. , and the larger amountof the rain occurs during the period fromOctober to May. Besides, being dry, thetemperature is very high during July andAugust. Almond trees can tolerate very li-mited summer rainfall.
Although, 65% of the precipitation is inwinter in the western part of the Mediter-ranean, it is 45 % and 35 % in Adana andHatay respectively. Spring rainfall is 15%, 25 % and 29 % in these placesrespectively. The distribution of summer
rainfall in the three districts is 2 %, 5 %and 12 %, respectively (Gülcan 1982).
The amount of annual rainfall is adequatefor water requirement of almond. Waterrequirement starts to increase in Apriland reaches maximum in July. Takingthis into consideration, it may be accep-ted that Adana and Hatay districts aremore suitable than the other provinces.
Almond trees are well adapted to the dryhot climates and especially to the stonyand calcareous soils of the western andsouthern of Turkey.
CULTIVAR SITUATIONAlmonds have been cultivated in Turkeyby seeds since the beginning. The exis-tence of a large number of bearing treesgrown from seeds under various ecologi-cal conditions provides an invaluablesource for varietal selections.
Because of the heterozygosity and theneed for cross fertilization, vegetativepropagation is necessary to obtain a uni-form crop in almond. In Turkey, almondhas been grown from seeds thus hinde-ring the possibilities of producing a stan-dard product.
In Turkey, the first attempt for selectionwas made in 1966 with the aim of impro-ving and standardizing productionthrough selected clones (Dokuzoguz etal., 1968). The existing trees in variouslocalities differ widely from each other insuch characters as vigour, yield, nut qua-lity and flowering time. The aim of thefirst project was to select the best indivi-duals among the almond trees of the Ae-gean and Marmara regions which are all
propagated by seeds and to look for lateflowering types which are very importantin almond production. During 1966 and1967 harvest seasons, 167 individualtrees were marked. Sixteen types of the-se have been selected for their bettercommercial characters.
In order to improve the yield and qualityby vegetative propagation with selectedcultivars, trials were established in Wes-tern Anatolia including Izmir, Manisa,Aydìn, Mugla, Denizli, Canakkale and Te-kirdag. In this project, standard almondcultivars with high agricultural and com-mercial values and late flowering typeswere selected. 38-44 days of differenceshave been observed between the firstand the last flowering selections (Dokuzo-guz and Gülcan, 1972).
In another project on the selected clonespollination requirements, yield and vigourwere searched. Some other importantcharacteristics of the selected cloneswere determined (Dokuzoguz et al.,1979). In addition, selected clones werepropagated by the state’s own nurseriesand distributed to growers in the regionswhere they were selected, the Aegeanand the Mediterranean regions.
Selection programmes still continue indifferent regions. For instance, 28 typeswere selected from wild almonds grown inTokat and studies are continued on thismatter (Gerçekçioglu and Günes, 1999).Similarly, a selection programme wasconducted to select late flowering in theNorth Eastern Anatolia during 1992-1995and 1996-1997. As a result, a collectionwas established at the Horticultural Re-search Institute in Erzincan (Aslantas andGüleryüz, 1999). In addition almond spe-cies are scattered all over Turkey. Forexample, Amygdalus orientalis and Amyg-dalus turcomanica grow naturally inSoutheastern Anatolia (Ak et al, 1999).
There are some investigations in relationto foreign cultivars as in the case of anexperiment which was carried out during1988-1993 in Adana and Pozanti, where16 foreign and 3 local almond types andcultivars were tested. Generally, the treesshowed a better growth and developmentin Adana than in Pozanti. As a result ofthis study, some cultivars such as ‘Fer-ragnes’, ‘Ferraduel’, ‘Cristomorto’,‘Drake’, ‘Texas’ and ‘Nonpareil’ werefound promising. It will be useful if thesecultivars are grown in different locationsof the Aegean and Mediterranean regions(Kaska et al., 1993).
Another experiment was carried out withabout 25 almond cultivars (Turkish, French,Spanish, Italian and American) at threelocations in South-eastern Anatolia and
Almond orchard growing under dry conditions

5FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Eastern Mediterranean region. Anothertrial was started by a private firm (NUR-MET) with foreign cultivars and all plantsare drip irrigated (Kaska, 1999). In ano-ther study, the performance of local typesand foreign cultivars was determined inHatay under the Mediterranean regionconditions (Polat et al., 1999).
CULTURAL PRACTICESOrchard establishmentMost existing almond trees have beengrown either as border trees or scatteredin grain fields or vineyards. Trees aregrown from seeds and are receiving regu-lar care.In recent years, very few plantations wereestablished with nursery stocks buddedwith selected clones. Trees are spaced at7-10 m distance depending upon soil,rainfall and irrigation conditions.
Propagation and rootstocksIn Turkey, up to now almond production isalmost totally based on trees propagatedby seeds. Recently, some of the nativeand foreign cultivars are being reprodu-ced by budding in the western part of Tur-key. In propagating almonds, budding ispractised either at the seed beds or at thenursery. “In situ” budding is the methodthat is used in Datça, the main almondgrowing district in Turkey. In the other re-gions, almond is propagated by buddingseedlings. Due to the tap root formation,bud taking is low in this way of establis-hing the orchard. However, this method isnot widely spread. Even though to a les-ser extent, in propagation by budding, therootstocks generally used are seedlingsof bitter almond or other almond cultivars.Thus, in Turkey there is not a standardseedling rootstock. Almond seedling isthe only rootstock used for the time be-ing. In grafted tree propagation, someclones such as 17-2, 2-32, 6-1 and‘Texas’ cultivar is used for seedling pro-pagation (Dokuzoguz and Gülcan, 1979).The outstanding feature of soil in Turkeyis the high calcium content in the majorityof the regions. Therefore attention hasbeen given to this fact in rootstock stu-dies. In the experiment in which somefeatures of almond cultivars ‘Afyon sivri’,‘Afyon bitter’, ‘Mordogan bitter’, ‘Texas’,01-12, 21-10, 47-10, 48-4 and 104-1 wereinvestigated as rootstock, ‘Texas’ and 48-4 were found to be superior in quality dueto seedling growth, the distance up to thefirst shoot and root branching. However,47-10 which was unacceptable due to itslow ratio of root branching displayed bet-ter results in terms of other parameters.Bitter cultivars together with 01-12 for-med primarily carrot roots. Carrot rootedseedlings present a lower performance inorchard establishment. However, this cri-terion looses its value if the seedlings arefield budded. ‘Afyon sivri’ and ‘Afyon bit-
ter’ cultivars, despite their low performan-ce in terms of most of the assessed cha-racters, could be considered as suitablebecause of the low number of shoots for-med on the seedlings (Unal et al, 1994).In addition, it can be said that the besttime of budding in Erzincan condition was15-25 August (Aslantas and Güleryüz,1999). In Bursa, about 10.000 grafted al-mond seedlings are propagated andnearly all of them are produced by privatesectors (Barut et al, 1999).
Soil managementThe soil management is seldom carriedout in arid and hot regions due to lost oforganic material from the soil. The cleancultivation system has generally beenused for many orchards. Deeply plowingof the soil in late fall is the general practi-se. The soil may be cultivated once ortwice throughout the spring and tillage isgenerally stopped at early summer.
FertilizationManure has been the universal fertilizerfor almond trees in Turkey. Fall or winterapplication is the general practise espe-cially in the Aegean and Mediterraneanregions. It is applied as a ring around thetree trunk. Many almond trees showsymptoms of nitrogen deficiency. The useof fertilizers will raise with the increasingacreage of mono-crop almond orchards.
IrrigationIrrigation is not practised for most almondtrees grown throughout the country.Water is the limiting factor in many regionsof Turkey and the almond has been con-sidered as a drought tolerant nut speciesalthough it produces larger and bettercrop where irrigation is possible. Many re-gions of the country have very little or norain during summer months. The almondis one of the few tree fruit species grownin rain-fed orchard systems in dry regionsof Turkey. Recently, drip irrigation is be-ing applied in some places.
PruningGenerally, young almond trees are trai-ned to form a leader system. It is obser-ved that almond trees are not pruned du-ring the productive stage.
DiseaseIn some microclimates such as Datça,
one of the reasons of unsuccessful al-mond growing is the bacterial cancer di-sease. According to Gündogdu and Kaya(1976), 13 % of the existing trees in thisarea are infected by the bacteria produ-cing damage to the twigs, branches andtrunks of the tree. Taking into account theimportant damages caused in almond or-chards in Turkey, an experiment was car-ried out by Gündogdu and Demir (1990),on the control measures and the suscep-tibility of some economically important al-mond cultivars. According to their results,48-2 and 48-5 almond clones are foundsusceptible, 101-23 clone being resistantto Pseudomonas amygdali. In addition,breeding work was carried out in order tocombine the unique characteristics of 48-2, 48-5 and ‘Nonpareil’ with resistance toP. amygdali and late flowering of 101-23(Misirli et al, 1999).
HARVESTINGAlmond harvest starts from middle of Julyin coastal line of the Aegean and Mediter-ranean regions. For example, harvest pe-riod is between 15-20 July and the firstweek of August in Alanya, Finike, Kas,Fethiye and Datça. Harvest is later in theplaces above the sea level. In Burdur, Is-parta and Denizli harvest starts during thesecond half of August and ends in thefirst week of September. In South-easternAnatolia, almonds are harvested in Au-gust. Almond harvest can take place inSeptember in Central and Eastern Anato-lia.
UTILIZATIONLargest part of almond production in Tur-key is domestic consumed and a limitedamount is exported. The almond is a uni-que nut species that can be consumed atdifferent stages of maturity from thegreen fruit stage to fully ripe. It is a com-mon usage in Turkey to eat the greennuts with salt when the kernel is still cris-py before the endocarp hardening occurs.The second stage for almond con-sumption is the time when embryos arecompletely developed, however, the useof this way is very limited. It is largelyconsumed as an edible nut either raw ordry roasted with salt, the kernel is eatenwithout blanching. Almond is a major in-gredient in confectionary and pastry formaking various kinds of products. In con-fectionary, kernels are blanched and coa-
Table 3. Almond export of Turkey
1995 1996 1997
Quantity Value Quantity Value Quantity Value(1000 US $) (t) (1000 US $) (t) (1000 US $) (t)
Almond (shell) 74 95 202 747 299 1.103Almond (kernel) 140 654 233 1.164 267 1.110

6 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
ted with sugar or chocolate. Another im-portant use of almond in confectionary isthe almond paste. The blanched kernelsare ground and made into a paste withsugar, then it is prepared as candies invarious kinds.
EXPORTTurkey’s almond export, which was negli-gible anyway, has declined from 500 t in1980-1982 period to 300 t in 1992-1994period. In the same period, however, al-mond production has increased conside-rably. The domestic consumption and in-dustrial use of almond has not increasedeven if nuts can be utilized in 500 to 1000different products in nut related industries(Sengül and Emeksiz, 1999). Almond ex-port of Turkey is given in Table 3.
CONCLUSIONAlthough Turkey has a big potential thereis not an industrialized almond produc-tion. Thus the almond growing should becarried out under irrigated conditions andwith standard cultivars. An important partof South–Eastern region will be irrigated.Considering this condition, this region isforeseen as an almond production centrein the future.
REFERENCES
Ak, B. E., I. Acar and E. Sakar, 1999.An investigation on determination of po-mological and morphological traits ofwild almond grown at Sanliurfa provin-ce. XI GREMPA Meeting on Pistachios& Almonds Sept 1-4, 1999. SanliurfaTurkey (In press)
Anonymous, 1997. The Summary ofAgricultural Statistics, 1997.
Aslantas, R. and M. Güleryüz, 1999. Al-mond selection in microclima areas ofNorth East Anatolia. . XI G R E M P AMeeting on Pistachios & Almonds Sept1-4, 1999. Sanliurfa Turkey (In press)
Aslantas, R. and M. Güleryüz, 1999Effects of budding time on success andsapling growth in almond in Erzincan lo-cation. XI. GREMPA Meeting on Pista-chios & Almonds Sept 1-4, 1999. San-liurfa Turkey (In press)
Barut, E., 1999. Almond growing in Bur-sa vicinity. XI GREMPA Meeting on Pis-tachios & Almonds Sept 1-4, 1999. San-liurfa Turkey (In press)
Dokuzoguz, M., R: Gülcan and A. Atilla,1968. Seleksiyon Yoluyla Bedem Islahi,TUBITAK TOAG-37 no’lu Proje SonuçRaporu.
Dokuzoguz, M., R: Gülcan, 1972. EgeBölgesi Bademlerinin Seleksi yon Yolu-yla Islah ve Seçilmis Tiplerin Adaptas-yonu Üzerinde Arastirmalar, TUBITAKTOAG-80 no’lu Proje Sonuç Raporu.
Dokuzoguz, M., R: Gülcan, 1979. Ba-dem Yetistiriciligi ve Sorunlari. TübitakYayinlari No: 432, Seri, No:90. Türkiye.
Dokuzoguz, M., R: Gülcan and M.N. Ka-rakir, 1979. Seçilmis Badem Tiplerinin8. Mukayesesi ve StandardizasyonuÜzerinde Arastirmalar, TUBITAKTOAG-203 no’lu Proje Sonuç Raporu.
Eti, S., S.Peydas, A.B. Küden, N. Kas-ka, S. Kurnaz and M. Ilgin, 1993.Çukurova Kosullarda Yetistirilen BaziBadem Çesitlerinin Döllenme Biyolojisive Embriyo Gelismesi Üzerine Çalisma-lar. TUBITAK TOAG-675 no’lu Proje So-nuç Raporu.
Gerçekçioglu, R. and M. Günes, 1999.A research on improvement of almond(P. amygdalus) by selection of wildplants grown in Tokat central district. XIGREMPA Meeting on Pistachios & Al-monds Sept 1-4, 1999. Sanliurfa Turkey(In press)
Gülcan, R., 1982. Akdeniz Bölge Ba-dem Yetistiriciligi Özellikleri ve Sonuçla-ri (unpublished).
Gündogdu, M. and Kaya, 1976. Prelimi-niary studies on a new bacterial diseaseof almond. J. Turkish Phytopat. V.5Num: 2-3:87-98.
Gündogdu, M. and G. Demir, 1990. In-vestigation on the susceptibility of eco-nomically important almond varietiesagainst “Pseudomonas amygdali” Psal-lidas control measures in Aegan RegionAlmond Orchards- Turkey J. Türk.Phytopath, 19 (1): 7-12.
Kaska, N. , 1999. Southeast Anatoliacan be an important almond growing re-gion of Turkey. XI GREMPA Meeting onPistachios & Almonds Sept 1-4, 1999.Sanliurfa Turkey (In press)
Kaska, N., A. Küden, AB. Küden, 1993.Özellikle geç çiçek açan yabanci ve yer-li badem çesitlerinin Adana vePozanti’da yetistirilmeleri üzerine aras-tirmalar. TUBITAK TOAG-670 no’lu Pro-je Sonuç Raporu.
Köksal, I., Y. Okay and B: Kunter, 1995.Consumption projections and produc-tion targets of nuts. National IV. Techni-que Congress of Turkish Agriculture En-gineer, Ankara.
MisirliΩ, A., A: Küden, G. Demir andR.Gülcan, 1999. Determine of phenoliccompounds in some almond hybrids va-rying in resistance to Psendomonasamygdali. XI GREMPA Meeting on Pis-tachios & Almonds, Sanliurfa, Türkey.
Polat, A. A. , C. Durgaç and O. Kamilo-glu, 1999. Determination of pomologicalcharacteristics of some local and fo-reign almond cultivars in Yayladagi (Ha-tay) ecological conditions. XI GREMPAMeeting on Pistachios & Almonds Sept1-4, 1999. Sanliurfa Turkey (in press)
Sengül, S and F. Emeksiz, 1999. Poten-tial almond production and developmentpossibility of domestic consumption andexport in Turkey. XI GREMPA Meeting
on Pistachios & Almonds Sept 1-4,1999. Sanliurfa Turkey (in press)
Unal, A., R. Gülcan and A. Misirli, 1994.A study on seedling rootstock proper-ties of some almond cultigens. ActaHorticulturae 373: 105-110.
Zeybekoglu, N., R: Gülcan and A. Misir-li, 1995. Bazi Seçilmis Badem TiplerininDöllenme Biyolojisi Üzerinde Arastirma-lar. Türkiye II. Ulusal Bahçe BitkileriKongresi, Adana, Türkiye.
A. Misirli; R. GülcanEge University, Agricultural Faculty, Depart-
ment of Horticulture35100 Bornova, Izmir – Turkey
Phone: 232 388 0110Fax: 232 388 1864
E-mail: [email protected]
ALMOND PRODUCTION ANDCHARACTERISTICS
IN ALGARVE, PORTUGAL
INTRODUCTIONAlmond is an Algarve (southern region ofPortugal) (Fig.1) traditional product, whe-re almond tree (Prunus amygdalus Batsh)is being cultivated for several centuries,on account of edaphologic and climateconditions of this region be very advanta-geous to its development.
VIANA DOCASTELO
PORTO
VILAREAL
BRAGANCA
COIMBRA
CASTELO BLANCO
SERRA
BARROCAL
FARO
BEJA
LISBOA
LITORAL
EVORA
Fig. 1. Portugal and Algarve region
ALGARVE
7FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Algarve climate is classified as mediterra-nean (Cs) of Koppen, with warm summer,annual mean temperature of 17ºC and22ºC of maxim mean temperature. It ismoderately rainy, with an annual meanprecipitation about 500 mm, whose distri-bution is irregular during the year, with pre-dominance between October and April. In-solation is superior to 3000 hours/ year.
Almond tree is at majority spread, gene-rally associated with other crops (fig andcarob tree), occupying mainly an area
called “Barrocal” (geological band bet-ween “Serra” and “Litoral”) (Fig. 1). In thisband, soils are mostly calcareous andstony, a fact that difficults the develop-ment of land.
In spite of this situation, almond tree hascomposed the main fruit-crop of Algarvetill 1960 decade, with a prominence posi-tion, for some decades, as much the por-tuguese market as the international mar-ket. From 1970 a decrease of yield wasregistered, which progressively incremen-
ted, reaching 1990 decade at a positionof a wide abandon.
From 1995 on a team of technicians andresearchers began works and studies forthe recovery of regional almond. Theexisting situation of abandon, despite theregional consumption be superior to pro-duction, contributes to import, more andmore product from USA and Spain.
Therefore, some research and experi-mentation projects, between 1995 and
Fig. 2. Almond and kernel of ‘Boa Casta’
Fig. 5. Almond and kernel of‘Duro Amarelo Grado’
Fig. 8. Almond and kernel of ‘Laja’
Fig. 11. Almond and kernel of ‘Patarata’
Fig. 3. Almond and kernel of‘Bonita de San Bras’
Fig. 6. Almond and kernel of‘Duro da Estrada Grado’
Fig. 9. Almond and kernel of ‘Lourencinha’
Fig. 12. Almond and kernel of‘Quinta de Flandres’
Fig. 4. Almond and kernel of ‘Do Prato’
Fig. 7. Almond and kernel of ‘Galamba’
Fig. 10. Almond and kernel of ‘Matias’
Fig. 13. Almond and kernel of ‘Zé Sales’
8 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
1999, were developed. They were inte-grated at communitarian programmes na-med PRAXIS XXI and PAMAF-IED, withparticipation of Direcção Regional deAgricultura do Algarve, Algarve Universityand some portuguese institutions, in or-der to recover existent traditional varie-ties, through the knowledge of its physi-co-chemical characteristics as well as ofits genetic identification.
AREA AND PRODUCTIONArea occupied by almond tree, at lastyears, is coming to decrease, due to theappearing of more profitable crops, na-mely citrines. Nowadays, almond treereaches, at Algarve, an area of about 15
Table 1. Evolution of almond production (kernel) along the last ten years (estimation)
Year 1990 1994 1995 1996 1998 1999 Average
1000 kg 9773 3000 1369 3642 1792 5300 4146
000 ha. Annual mean production, alongthe last ten years, was approximately4100 t of kernel. Its evolution can be seenon Table 1.
CultivarsDuring the last six years, 45 Algarve al-mond varieties were studied, throughphysical, chemical, and organolepticanalyses. They were settled as a collec-tion for later genetic and agronomical stu-dy.
As result of realised studies, 12 varieties(Figures 2 to 13) were selected for theirquality characteristics. Plants of these va-rieties were implanted at an experimental
field, with 6m x 5m spacing, and drip irri-gation (2 drips/plant). Physical and che-mical characteristics of these varietiesare registered on the following Tables 2and 3.
UsesKeeping in view almond quality characte-ristics, namely its composition on monoin-saturated fat acids, sugars, and naturalantioxidants, it has many applications,being an excellent food when consumednaturally, raw or roasted.
It is used, at Algarve, mainly in the wellappreciated regional sweets, as well as inother products combined with fig, carob
Table 2. Physical characteristics of 12 varieties of regional almond
Variety Mean weight Fruit mean size (with kernel)(with kernel) (mm)
(g) Longitudinal Transversal Thickness Yield Hardnesslength length (%) (N)
‘Boa Casta’ 3,52 29,21 19,71 13,54 31,3 773‘Bonita de S. Brás’ 3,14 28,41 17,85 15,16 30,6 746‘Do Prato/Bico de Papagaio’ 3,82 38,52 22,65 15,84 38,7 315‘Duro Amarelo Grado’ 4,30 26,61 23,00 18,04 20,9 1020‘Duro da Estrada Grado’ 4,04 27,84 22,34 15,98 23,0 1053‘Galamba’ 4,28 29,49 22,12 15,07 23,4 1078‘Laja’ 1,99 28,50 21,78 15,49 50,3 1029‘Lourencinha’ 3,60 27,27 20,94 14,71 31,4 1132‘Matias’ 2,27 26,98 22,35 16,78 51,5 229‘Patarata’ 4,32 28,76 23,66 16,08 21,1 1085‘Quinta de Flandres’ 3,58 26,02 19,43 17,32 27,4 771‘Zé Sales’ 2,78 24,27 13,55 8,25 39,2 952
N= Newton
Table 3. Chemical characteristics of 12 varieties of regional almond
Variety ° Brix Ash Moisture Fat Fatty acids (%)
(%) (%) (%) (%) Palmitic PalmitoleicEsteáric Oleic Linoleic Linolénic Others
‘Boa Casta’ 18,3 2,9 4,3 45,5 6,003 0,375 3,132 67,596 22,125 0,034 0,734‘Bonita de S. Brás’ 29,0 4,0 4,3 30,1 6,544 0,368 3,132 67,597 22,126 0,033 0,732‘Do Prato/Bico de Papagaio’ 21,5 2,9 5,4 40,6 7,312 0,408 2,035 58,961 29,886 0,032 1,366‘Duro Amarelo Grado’ 19,0 3,3 4,3 51,0 5,936 0,370 2,631 67,567 22,971 0,096 0,430‘Duro da Estrada Grado’ 18,0 3,2 4,3 48,6 6,153 0,409 2,811 69,687 20,285 0,121 0,533‘Galamba’ 20,3 2,9 6,6 49,1 6,245 0,404 3,194 68,334 20,971 0,303 0,548‘Laja’ 25,0 3,5 4,5 46,4 6,485 0,414 2,917 68,225 21,410 0,079 0,469‘Lourencinha’ 25,0 4,0 3,5 42,5 6,802 0,458 2,859 70,889 17,518 0,070 1,412‘Matias’ 27,0 3,6 4,1 41,1 6,224 0,443 2,626 66,404 23,774 0,038 0,492‘Patarata’ 27,0 3,8 3,8 45,3 6,095 0,354 2,145 63,531 26,303 0,037 1,535‘Quinta de Flandres’ 24,0 3,3 4,3 47,0 6,409 0,379 2,796 65,412 24,515 0,092 0,397‘Zé Sales’ 27,5 2,1 4,3 31,5 7,258 0,388 2,219 60,194 28,069 0,045 1,828
9FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
and honey, also traditional regional com-modities. According to its varietal charac-teristics, almond should be used in seve-ral ways, in order to be better appreciatedand valorised.
Future prospectsConversion of traditional almond orchardswill be a reality, since attractive Europeansupports in relation to some more profita-ble regional crops subsist, and promotionand diffusion actions of the Algarve al-mond also take place.
Consumers’ demand for healthy food pro-ducts (like Algarve almond, with hardshell varieties without application ofphytosanitary treatments) will induce toincrease the regional almond con-sumption and, in this way, farmers can re-new old orchards, at short or middle term.
A. Neto Martins1; C. Gomes1; L. Ferreira2
Direcção Regional de Agricultura do Algarve1Laboratory of Agricultural Technology;
2 Fruit-growing DepartmentApartado 282, Patacão
8001-904 Faro, PortugalTel: 351 289 870 756; Fax: 351 289 816 048E-mail: [email protected]
INBREEDING CONSEQUENCESRELATED TO
SELF-COMPATIBILITYIN ALMOND
INTRODUCTIONSelf-compatibility (SC) in almond (Prunusdulcis (D. A. webb) Miller) is an importantissue of research in many countries, theMediterranean south shore is also inte-rested in this trait and by the self-compa-tible varieties. It was a subject of manyworks, particularly its transmission. Vari-ous works have shown that the use ofsome sources of SC (inter-specific hybri-dizations and induced mutations) havemany disadvantages. Some undesirabletraits, found in other species, are trans-mitted together with self-compatibility tothe offspring. Many back-crosses are ne-cessary to recover the almond type (Kes-ter and Asay, 1975). If the mutations ap-pear to be interesting to apply, they arenot selective and the results obtained areuncertain. Most breeding programmes forself-compatibility are based on the use ofself-compatible genotypes. Cultivars fromApulia (Italy) constitute a principal sourceof self-compatibility. They have allowedto select some self-compatible cultivarslike ‘Lauranne’, ‘Steliette’, ‘Ayles’, ‘Mon-cayo’ ‘Guara’ and recently ‘Mandaline’(Duval, 1999), ‘Antoñeta’ and ‘Marta’(Egea et al., 1999).
Like other breeding programmes, theexistence of the inbreeding abnormalities,hinders experimentation and slows theprogress made in the selection. The un-lucky effects of inbreeding were attributedto unfavourable genes expression (Seavyand Bawa, 1986; Krebs and Hancock,1988, Matthew et al., 1994). They can beexpressed at different phases of plant life(Melville et al., 1980; Karron, 1989) andthe importance of their effects variesbetween species (Schemske, 1983). The-se abnormalities concern also the almond(Grasselly and Olivier, 1981; 1987, So-cias I Company, 1990; Garcia and Dicen-ta, 1995). They were not subject of anyparticular studies, and their descriptionwere reviews and cannot allow to knowthe expression levels of undesirable ge-
nes and assess their implication on theoffspring performance.
The inbreeding consequences, related tothe self-compatibility of the cultivated al-mond have been studied with the cultivar‘Tuono’ and evaluated histologically andmorphologically. The following contribu-tion concerns three topics related to thepollen tube growth, the process of fertili-zation, the fruitfulness and the pomologi-cal traits of nuts.
MATERIAL AND METHODSThe self compatible cultivar ‘Tuono’growing in INRA Aïn Taoujdate collection,was self- and cross- pollinated by threegenotypes of different genetical origin:‘Marcona’ (Spain), ‘Ferragnès’ (France)
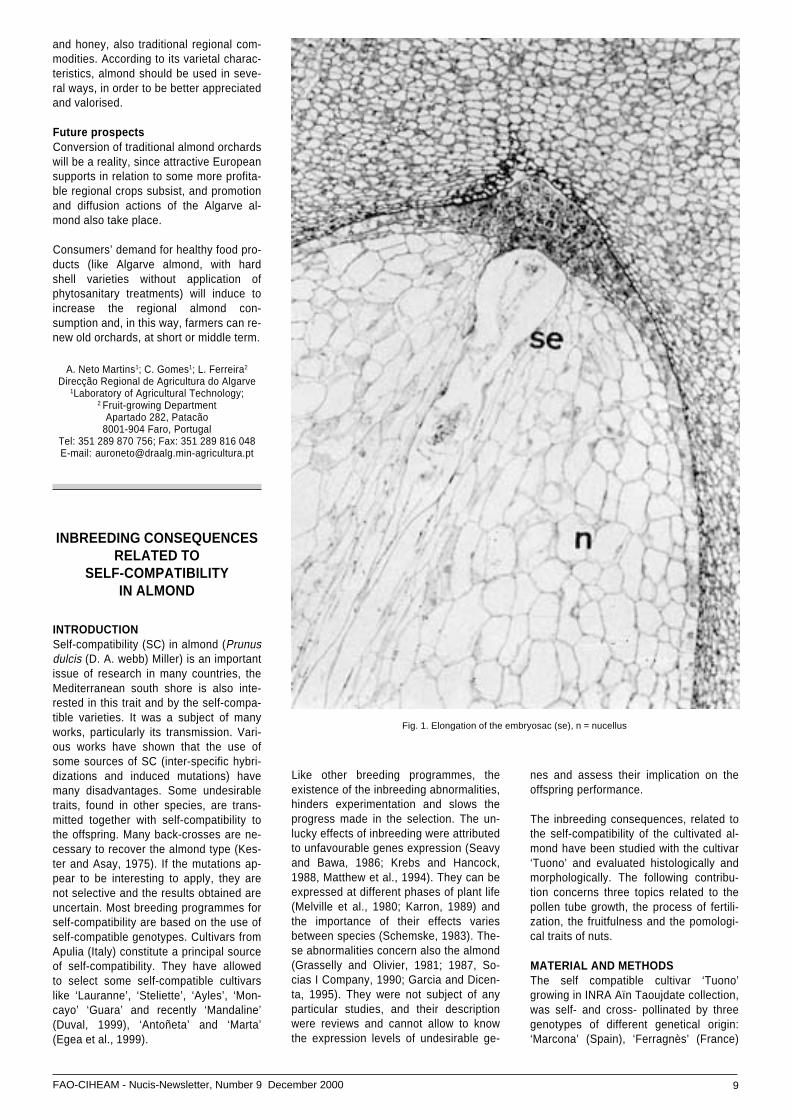
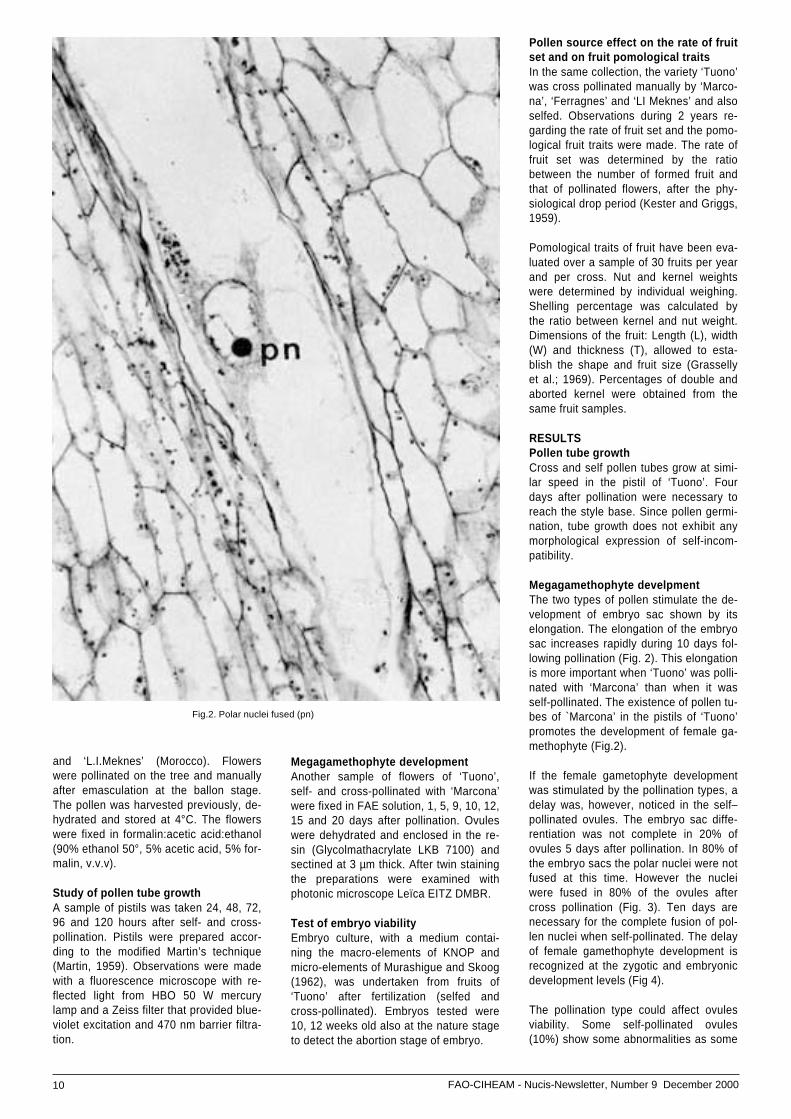
Fig. 1. Elongation of the embryosac (se), n = nucellus
10 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
and ‘L.I.Meknes’ (Morocco). Flowerswere pollinated on the tree and manuallyafter emasculation at the ballon stage.The pollen was harvested previously, de-hydrated and stored at 4°C. The flowerswere fixed in formalin:acetic acid:ethanol(90% ethanol 50°, 5% acetic acid, 5% for-malin, v.v.v).
Study of pollen tube growthA sample of pistils was taken 24, 48, 72,96 and 120 hours after self- and cross-pollination. Pistils were prepared accor-ding to the modified Martin’s technique(Martin, 1959). Observations were madewith a fluorescence microscope with re-flected light from HBO 50 W mercurylamp and a Zeiss filter that provided blue-violet excitation and 470 nm barrier filtra-tion.
Megagamethophyte developmentAnother sample of flowers of ‘Tuono’,self- and cross-pollinated with ‘Marcona’were fixed in FAE solution, 1, 5, 9, 10, 12,15 and 20 days after pollination. Ovuleswere dehydrated and enclosed in the re-sin (Glycolmathacrylate LKB 7100) andsectined at 3 µm thick. After twin stainingthe preparations were examined withphotonic microscope Leïca EITZ DMBR.
Test of embryo viabilityEmbryo culture, with a medium contai-ning the macro-elements of KNOP andmicro-elements of Murashigue and Skoog(1962), was undertaken from fruits of‘Tuono’ after fertilization (selfed andcross-pollinated). Embryos tested were10, 12 weeks old also at the nature stageto detect the abortion stage of embryo.
Pollen source effect on the rate of fruitset and on fruit pomological traitsIn the same collection, the variety ‘Tuono’was cross pollinated manually by ‘Marco-na’, ‘Ferragnes’ and ‘LI Meknes’ and alsoselfed. Observations during 2 years re-garding the rate of fruit set and the pomo-logical fruit traits were made. The rate offruit set was determined by the ratiobetween the number of formed fruit andthat of pollinated flowers, after the phy-siological drop period (Kester and Griggs,1959).
Pomological traits of fruit have been eva-luated over a sample of 30 fruits per yearand per cross. Nut and kernel weightswere determined by individual weighing.Shelling percentage was calculated bythe ratio between kernel and nut weight.Dimensions of the fruit: Length (L), width(W) and thickness (T), allowed to esta-blish the shape and fruit size (Grassellyet al.; 1969). Percentages of double andaborted kernel were obtained from thesame fruit samples.
RESULTSPollen tube growthCross and self pollen tubes grow at simi-lar speed in the pistil of ‘Tuono’. Fourdays after pollination were necessary toreach the style base. Since pollen germi-nation, tube growth does not exhibit anymorphological expression of self-incom-patibility.
Megagamethophyte develpmentThe two types of pollen stimulate the de-velopment of embryo sac shown by itselongation. The elongation of the embryosac increases rapidly during 10 days fol-lowing pollination (Fig. 2). This elongationis more important when ‘Tuono’ was polli-nated with ‘Marcona’ than when it wasself-pollinated. The existence of pollen tu-bes of `Marcona’ in the pistils of ‘Tuono’promotes the development of female ga-methophyte (Fig.2).
If the female gametophyte developmentwas stimulated by the pollination types, adelay was, however, noticed in the self–pollinated ovules. The embryo sac diffe-rentiation was not complete in 20% ofovules 5 days after pollination. In 80% ofthe embryo sacs the polar nuclei were notfused at this time. However the nucleiwere fused in 80% of the ovules aftercross pollination (Fig. 3). Ten days arenecessary for the complete fusion of pol-len nuclei when self-pollinated. The delayof female gamethophyte development isrecognized at the zygotic and embryonicdevelopment levels (Fig 4).
The pollination type could affect ovulesviability. Some self-pollinated ovules(10%) show some abnormalities as some
Fig.2. Polar nuclei fused (pn)

11FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
failure occurred in the polar nucleus unitand lack of embryo sac elongation thatcould induce ovular degeneration. Crosspollination of ‘Tuono’ by ‘Marcona’ leadsto fast ovule development than self polli-nation. The type of pollination seems tohave a potential effect on ovular viabilitywhich suggests that the inbreeding be-gins to take place after fertilization.
Embryo viabilityDespite the type of pollination, the em-bryos of ‘Tuono’ are able to germinate “invitro”. The viability of embryos, tested 11weeks after pollination, had shown a sig-nificant difference between self- andcross-pollination. The rate of viability was89% in the first case and only 60% in thesecond. About 40% of embryos achievedtheir development at this time when ‘Tuo-no’ was selfed. This delay could be a pro-blem in early stage of embryo develop-ment. A proportion of young fruits drop-ped after self-pollination. At the maturestage, the rate of embryo viability was si-milar for the two types of pollination andthe mean was 91%.
Effect of the pollen source on the fruitset rate and on pomological fruit traitsThe rate of fruit set differs according tothe type of pollination. The mean fruit setobtained in 2 years is 25.6% in cross-pollination and was higher to that obtai-ned in self-pollination, reaching 19.2%.Cross-pollination is more efficient thanself-pollination.
Fruit size is reduced when ‘Tuono’ wasself-pollinated in comparison with crosspollination, particularly in weight, lengthand thickness of the nut. The decrease ofthe average fruit weight ranges from 4 to17.5%.
Considerable influence of the pollen pa-rent has been observed on nut size (Ku-mar and Das, 1996). But in our case, thisreduction is linked to inbreeding effects.Self-fertilization would have favoured thecombination of unfavourable recessivealleles controlling pomological traits.
DISCUSSION AND CONCLUSIONSInbreeding effect on reproduction and onvigour are known in many vegetal spe-cies. Inbreeding causes many genetic,morphologic and physiologic abnormali-ties. In self-compatible almond, effectsare very severe after self-pollination andaffect at different stages since seeds for-mation and establishment of seedling.
During the reproductive phase, self polli-nation of ‘Tuono’ does not lead to a lowspeed of pollen tube growth. They grewat similar speed to cross pollen tube andreached the style base after 4 days. No
Fig.3. Embryo (em)
morphological manifestation of incompati-bility was observed in the two types ofpollination. Pre-zygotic inhibition can be agenetical barrier to self-pollination whichwas not shown in the style. At this level,the embryo sac development and its ma-turation were stimulated by the two typesof pollination. The polar nuclei fusion andembryo sac elongation are produced aspollen tubes are at the base of style. Adelay in the female gamethophyte deve-lopment however was noticed when self-pollination. The latter could affect theovular potential viability and embryo de-velopment.
After fertilization, fruit set rates were dif-ferent between types of pollination. Fruitset is better when this cultivar is crossedmanually. This difference is linked to ge-
netical factors which have a depressiveeffect. This later were detectables at thepomological traits of fruit. Kernel length,weight and weakness were reduced when‘Tuono’ is self-pollinated.
Embryonic abortion of seed may not be atthe origin of this problemme. The rates ofembryo germination, tested at maturestage “in vitro” culture, were similar be-tween self- and cross-pollination. Sincethe raising, a proportion of young seed-lings coming from self-pollination havesome difficulties in their growth phaseand their development. The inbred seed-lings have a low vigour of 20% in compa-rison with seedlings from crosses. Someof the 28% of seedlings died and the dea-th concerned the less vigorous (Oukabliet al., 2000).

12 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Many traits are subject to inbreeding de-pression and the effects are clear whenthey are quantitative. Fruit set, kernel po-mological traits, seed germination, deve-lopement and seedling growth, vigour areaffected by self-pollination. An unfavoura-ble gene, present at recessive state in thegenome of ‘Tuono’ seems to be at homo-zygotic state.
Some inbreeding barriers are disclosed atdifferent phases. One of the most impor-tant inbreeding barriers is self-incompati-bility. This study has concluded that thisbarrier forces self-fertilization at an earlystage, and it operates in the style, afterwhich it ceases. It is genetically raised bythe self-compatibility condition. However,tree barriers remain operational and actat different levels:
1st barrier: post-zygotic, it could be functio-nal at the ovular level. The embryothrowing, in self-pollination, could be linkedto its incapacity to continue its develop-ment. This incapacity could be linked to un-desirable genes that conduct to loss of em-bryonic vigour or to endosperm abortion.
2nd barrier: it is situated at the germinationlevel of seed and constitutes an obstaclethat reduces germination rate. This stickinglevel could be the result of accumulation, inthe genotype of any embryo, of genes thatinhibit or affect unrolling of any biochemicalprocess during germination.
3rd barrier: it concerns one part of plantsthat escape from the two first barriers. Itis exhibited during growth and develop-ment of the inbred descendants and redu-ces seedling vigour.
These operational barriers are geneticabnormalities caused by deleterious ge-
nes that are masked by the heterosisstructures. Self-compatibility acquisition,for one allogamous species is complexand striking at the inbreeding effects.This is one of the mechanisms that act toincrease heterozigousity in the genomeby eliminating unfavourable genes.
The importance of inbreeding effects ob-served in this study, reinforces the ideathat ‘Tuono’ could have undergone yet aninbreeding process in its native area. Theinbreeding is difficult to avoid when thegenetic base of the breeding programmeis narrow. The breeder should plan to usea high number of genitors.
LITERATURE CITED
Kester, D.E; and R. Asay, 1975. Al-monds; in advances in fruit breeding.Edited by Jules Janick and James v.Moore: 387-420.
Duval, H., 1999. ‘Mandaline’, a newfrench almond variety. Nucis 8, 36.
Egea J. 1999. ‘Antoñeta’ and ‘Marta’two new self-compatible and late flowe-ring almond varieties. Nucis 8, 36.
Seavey, S. and K. S. Bawa. 1986. Late-acting self-incompatibility in Angios-perms. The Botanical Review 52 (2):195-219.
Krebs, S.L. and J.F. Hancock. 1988.The consequence of inbreeding on ferti-lity in Vaccinium corymbosum, L. J.Amer. Soc. Hort. Sci. 113 (6): 914-918.
Matthew, B. Hamilton and Thomas Mit-chell-Olds, 1994. The mating systemand relative performance of selfed andoutcrossed progeny in Arabis feconda(Brassicacea). American Journal of Bo-tany 81(10): 125-128.
Melville, A. H.; G. T. Galleta and A. D.Draper, 1980. Seed germination andearly seedling vigor in progenies of in-bred strawberry selections. HortScien-ce, 15(6): 749-750.
Karron, J. D. 1989. Breeding systemsand levels of inbreeding depression ingeographically restricted and widspreadspecies of Astragalus (Fabaceae).American Journal of Botany 76: 331-340.
Schemske, D. W., 1983. Breeding sys-tem and habit effects on fitness compo-nents in three neotropical Costus (Zingi-beracea). Evolution 37: 523-539.
Grasselly, Ch.; P. Crossa-Raynaud; G.Olivier; et H. Gall.,1981. Transmissiondu caractère d’auto-compatibilité chezl’amandier. Colloque GREMPA, Optionsméditerranéennes, CIHEAM/IAMZ 81/I:71-75.
Socias I Company, R. 1990. Breedingself-compatible almonds. Plant Bree-ding Review 8: 313-337.
Garcia, J. E. and F. Dicenta, 1994. Eva-luation of losses of plant material duringan almond breeding programme. J. Ge-net. and Breed. 48:155-160.
Martin, F. W. 1959. Staining and obser-ving pollen tubes in the styles by meansof fluoresence. Stain technol. 34: 125-128.
Kester, D.E. and W.H. Griggs. 1959.Fruit setting in the almond: the effect ofcross pollinating various percentages offlowers. Proc. Amer. Soc. Hort. Sci.74:206-213.
Grasselly, Ch.; H. Gall; et P. Leglise.1969. Etude pomologique de quarantevariétés d’amandier. Bull. Techniqued’information, 241: 507-522.
Murachine, J. and F. Skoog. 1962. A re-vised medium for rapid growth and bio-assays with tobacco tissue cultures.Physiol. Plant 15: 473-497.
Kumar, K. and B. Das. 1996. Studies onxenia in almond [Prunus dulcis (Miller)D.A. Webb]. Journal of HorticulturalScience 71(4): 545-549.
Oukabli, A., A. Lansari, D.E. Wallali andA. Abousalim, 2000. Effets endogami-ques sur la germination et la croissancedes semis d’amandier autocompatiblesPrunus dulcis (D. A. Webb) Miller cv‘Tuono’. Fruits (in press).
Oukabli A.1, A. Lansari2, Wallali L.D.E3
and A. Abousalim3
1 INRA, Centre Régional de la RechercheAgronomique
Programme Arboriculture Fruitière, BP 578,Meknès, Morocco
2 Ecole Nationale D’Agriculture, Meknes,Morocco
3 IAV Hassan II, Département d’Horticulture,Rabat, Morocco
Fig. 4. Almonds from ‘Tuono’ selfed, ‘Tuono’ x ‘Marcona’ and ‘Tuono’ x ‘Ferragnès’
13FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
INCREASING YIELDS ANDPROFITABILITY OF ALMOND
CULTURE IN ISRAEL
Almond is a very old crop in Israel and inthe rest of the Mediterranean area. Al-mond, a dominant culture in Israel a cen-tury ago, was replaced by the more profi-table citrus. Currently, almonds are grownon 2,500 ha , producing 2,000 tons of ker-nels per year at a value of $9,000,000.There is a total demand for 4,000 t andUS $10,000,000 worth of import. The pri-ce of the almond is constantly increasingand has risen from $2 per kg three yearsago to $5-6 from 1999 to this year 2000.Although this price might drop somewhat,there is an increasing world and local de-mand evaluated at 15% per annum withnewly opened markets in the Far East. Inaddition to natural and roasted unshellednuts, there is a constant increase in theindustrial use of almonds; processed al-monds receive very high prices.
Almond culture is of great interest in Is-rael because of its new profitability, me-chanical harvesting and pruning and au-tomated fertirrigation. This enables sol-ving of problemmes resulting from thefact that in Israel the cost of labour is highand workers are difficult to find. In addi-tion yields in Israel are low, approximate-ly 750 kg kernel/ha on the multiannualaverage. During the past two years, wehave found orchards yielding 1t/ha usingnew agrotechniques. In California, thereare orchards with excellent cultural condi-tions that reach 3t kernels/ha. In Israel al-mond culture is profitable from 1t/ha inthe new intensified orchards. Theblooming period in our area occurs earlyin January, when the rain and cold wea-ther harm flowers and interfere with polli-nation and fertilization, frequently causinglow yields and alternate bearing.
Our cultivars have a paper shell and arevery susceptible to all pests, especially inspring. The leading cultivar is ‘Um El Fa-hem’ (local selection). In our climate,hard shell and late flowering cultivarswould be expected to result in higher andmore consistent yields with fewer pestici-de applications.
For many years, the rootstock best adap-ted to our region was the bitter almondseedling, which is resistant to droughtand suitable for dry land farming. In arootstock trial, we observed that GF-677and GF-749 (peach x almond), bred inFrance, gave the best results and largeryields when culture intensification isused. The trees were taller, double insize, with 140% more yield and withouttree decline (while in bitter almond 30%of the trees died). The phenomena mani-
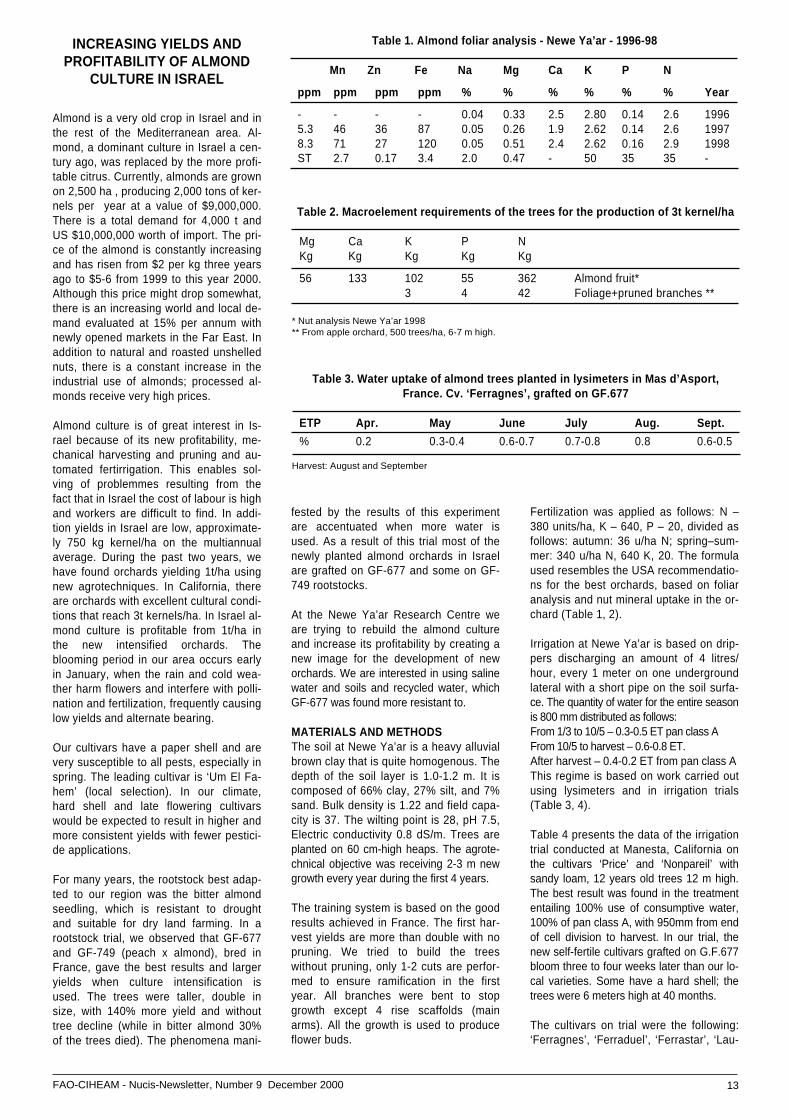
Table 1. Almond foliar analysis - Newe Ya’ar - 1996-98
Mn Zn Fe Na Mg Ca K P N
ppm ppm ppm ppm % % % % % % Year
- - - - 0.04 0.33 2.5 2.80 0.14 2.6 19965.3 46 36 87 0.05 0.26 1.9 2.62 0.14 2.6 19978.3 71 27 120 0.05 0.51 2.4 2.62 0.16 2.9 1998ST 2.7 0.17 3.4 2.0 0.47 - 50 35 35 -
Table 3. Water uptake of almond trees planted in lysimeters in Mas d’Asport,France. Cv. ‘Ferragnes’, grafted on GF.677
ETP Apr. May June July Aug. Sept.
% 0.2 0.3-0.4 0.6-0.7 0.7-0.8 0.8 0.6-0.5
Harvest: August and September
Table 2. Macroelement requirements of the trees for the production of 3t kernel/ha
Mg Ca K P NKg Kg Kg Kg Kg
56 133 102 55 362 Almond fruit*3 4 42 Foliage+pruned branches **
* Nut analysis Newe Ya’ar 1998** From apple orchard, 500 trees/ha, 6-7 m high.
fested by the results of this experimentare accentuated when more water isused. As a result of this trial most of thenewly planted almond orchards in Israelare grafted on GF-677 and some on GF-749 rootstocks.
At the Newe Ya’ar Research Centre weare trying to rebuild the almond cultureand increase its profitability by creating anew image for the development of neworchards. We are interested in using salinewater and soils and recycled water, whichGF-677 was found more resistant to.
MATERIALS AND METHODSThe soil at Newe Ya’ar is a heavy alluvialbrown clay that is quite homogenous. Thedepth of the soil layer is 1.0-1.2 m. It iscomposed of 66% clay, 27% silt, and 7%sand. Bulk density is 1.22 and field capa-city is 37. The wilting point is 28, pH 7.5,Electric conductivity 0.8 dS/m. Trees areplanted on 60 cm-high heaps. The agrote-chnical objective was receiving 2-3 m newgrowth every year during the first 4 years.
The training system is based on the goodresults achieved in France. The first har-vest yields are more than double with nopruning. We tried to build the treeswithout pruning, only 1-2 cuts are perfor-med to ensure ramification in the firstyear. All branches were bent to stopgrowth except 4 rise scaffolds (mainarms). All the growth is used to produceflower buds.
Fertilization was applied as follows: N –380 units/ha, K – 640, P – 20, divided asfollows: autumn: 36 u/ha N; spring–sum-mer: 340 u/ha N, 640 K, 20. The formulaused resembles the USA recommendatio-ns for the best orchards, based on foliaranalysis and nut mineral uptake in the or-chard (Table 1, 2).
Irrigation at Newe Ya’ar is based on drip-pers discharging an amount of 4 litres/hour, every 1 meter on one undergroundlateral with a short pipe on the soil surfa-ce. The quantity of water for the entire seasonis 800 mm distributed as follows:From 1/3 to 10/5 – 0.3-0.5 ET pan class AFrom 10/5 to harvest – 0.6-0.8 ET.After harvest – 0.4-0.2 ET from pan class AThis regime is based on work carried outusing lysimeters and in irrigation trials(Table 3, 4).
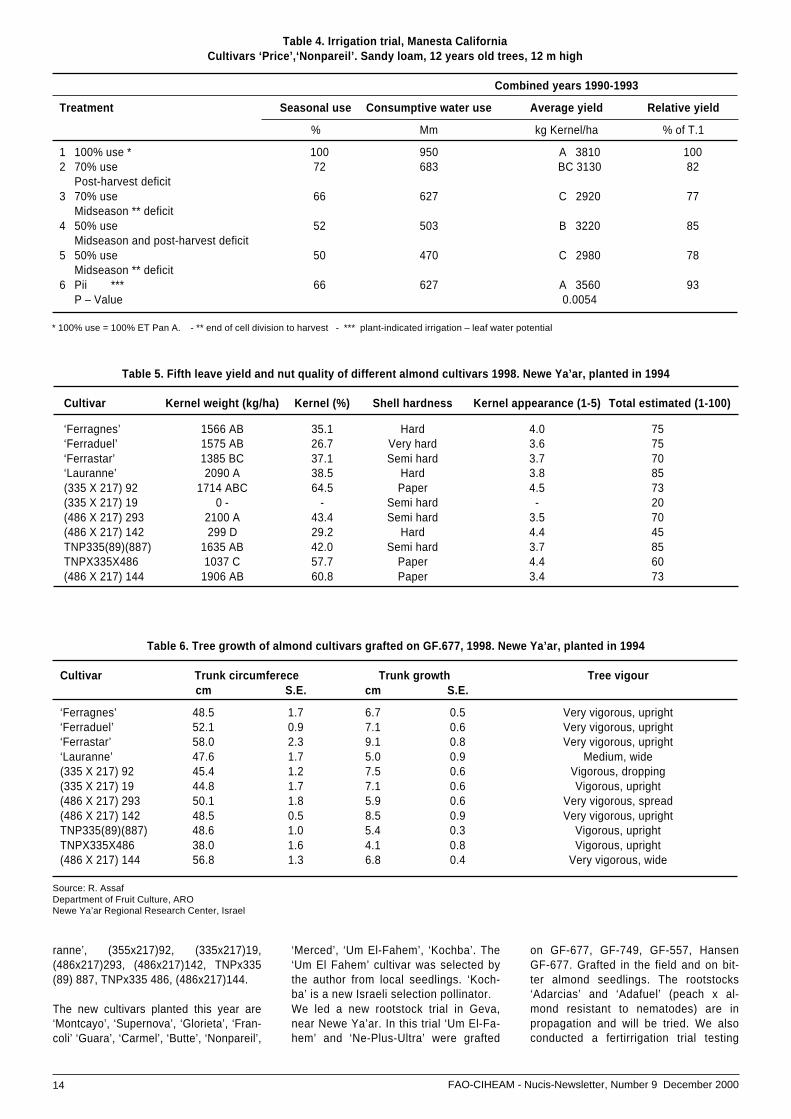
Table 4 presents the data of the irrigationtrial conducted at Manesta, California onthe cultivars ‘Price’ and ‘Nonpareil’ withsandy loam, 12 years old trees 12 m high.The best result was found in the treatmententailing 100% use of consumptive water,100% of pan class A, with 950mm from endof cell division to harvest. In our trial, thenew self-fertile cultivars grafted on G.F.677bloom three to four weeks later than our lo-cal varieties. Some have a hard shell; thetrees were 6 meters high at 40 months.
The cultivars on trial were the following:‘Ferragnes’, ‘Ferraduel’, ‘Ferrastar’, ‘Lau-
14 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Table 5. Fifth leave yield and nut quality of different almond cultivars 1998. Newe Ya’ar, planted in 1994
Cultivar Kernel weight (kg/ha) Kernel (%) Shell hardness Kernel appearance (1-5) Total estimated (1-100)
‘Ferragnes’ 1566 AB 35.1 Hard 4.0 75‘Ferraduel’ 1575 AB 26.7 Very hard 3.6 75‘Ferrastar’ 1385 BC 37.1 Semi hard 3.7 70‘Lauranne’ 2090 A 38.5 Hard 3.8 85(335 X 217) 92 1714 ABC 64.5 Paper 4.5 73(335 X 217) 19 0 - - Semi hard - 20(486 X 217) 293 2100 A 43.4 Semi hard 3.5 70(486 X 217) 142 299 D 29.2 Hard 4.4 45TNP335(89)(887) 1635 AB 42.0 Semi hard 3.7 85TNPX335X486 1037 C 57.7 Paper 4.4 60(486 X 217) 144 1906 AB 60.8 Paper 3.4 73
Table 6. Tree growth of almond cultivars grafted on GF.677, 1998. Newe Ya’ar, planted in 1994
Cultivar Trunk circumferece Trunk growth Tree vigourcm S.E. cm S.E.
‘Ferragnes’ 48.5 1.7 6.7 0.5 Very vigorous, upright‘Ferraduel’ 52.1 0.9 7.1 0.6 Very vigorous, upright‘Ferrastar’ 58.0 2.3 9.1 0.8 Very vigorous, upright‘Lauranne’ 47.6 1.7 5.0 0.9 Medium, wide(335 X 217) 92 45.4 1.2 7.5 0.6 Vigorous, dropping(335 X 217) 19 44.8 1.7 7.1 0.6 Vigorous, upright(486 X 217) 293 50.1 1.8 5.9 0.6 Very vigorous, spread(486 X 217) 142 48.5 0.5 8.5 0.9 Very vigorous, uprightTNP335(89)(887) 48.6 1.0 5.4 0.3 Vigorous, uprightTNPX335X486 38.0 1.6 4.1 0.8 Vigorous, upright(486 X 217) 144 56.8 1.3 6.8 0.4 Very vigorous, wide
Source: R. AssafDepartment of Fruit Culture, ARONewe Ya’ar Regional Research Center, Israel
Table 4. Irrigation trial, Manesta CaliforniaCultivars ‘Price’,‘Nonpareil’. Sandy loam, 12 years old trees, 12 m high
Combined years 1990-1993
Treatment Seasonal use Consumptive water use Average yield Relative yield
% Mm kg Kernel/ha % of T.1
1 100% use * 100 950 A 3810 1002 70% use 72 683 BC 3130 82
Post-harvest deficit3 70% use 66 627 C 2920 77
Midseason ** deficit4 50% use 52 503 B 3220 85
Midseason and post-harvest deficit5 50% use 50 470 C 2980 78
Midseason ** deficit6 Pii *** 66 627 A 3560 93
P – Value 0.0054
* 100% use = 100% ET Pan A. - ** end of cell division to harvest - *** plant-indicated irrigation – leaf water potential
ranne’, (355x217)92, (335x217)19,(486x217)293, (486x217)142, TNPx335(89) 887, TNPx335 486, (486x217)144.
The new cultivars planted this year are‘Montcayo’, ‘Supernova’, ‘Glorieta’, ‘Fran-coli’ ‘Guara’, ‘Carmel’, ‘Butte’, ‘Nonpareil’,
‘Merced’, ‘Um El-Fahem’, ‘Kochba’. The‘Um El Fahem’ cultivar was selected bythe author from local seedlings. ‘Koch-ba’ is a new Israeli selection pollinator.We led a new rootstock trial in Geva,near Newe Ya’ar. In this trial ‘Um El-Fa-hem’ and ‘Ne-Plus-Ultra’ were grafted
on GF-677, GF-749, GF-557, HansenGF-677. Grafted in the field and on bit-ter almond seedlings. The rootstocks‘Adarcias’ and ‘Adafuel’ (peach x al-mond resistant to nematodes) are inpropagation and will be tried. We alsoconducted a fertirrigation trial testing

15FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
five treatments on ‘Um El-Fahem’ and‘Ne Plus Ultra’, grafted on GF-677.
RESULTSTable 5 shows the second yield and nutquality of the almond (5th leaf). In the 4th
leaf we received a yield of 1400 kg/ ker-nels/ ha in the best cultivar and in the 5th
leaf the yield was 2090 kg/ha.
In relation to accumulated yield promisingcultivars are:First group: (486 X 217) 293, ‘Lauranne’Second group: TNP 335 (89) (887),‘Ferraduel’, ‘Ferragnes’, (486 X 217) 144,(335x217) 92.Third group: ‘Ferrastar’, TNPX335X486,(486 X 217) 142.The cultivar (335 X 217) 19 failed.
The nuts of all of the different cultivarswere free of pests, with no double or shri-veled kernel. All of the new cultivars weretested by a panel and found to be of avery good nut quality. The kernel percen-tage was as follows: hard shell almonds –35% kernel; very hard shell – 27% kernel;paper shell – 50 -60% kernel.
Trunk growth is presented in Table 6. Wefound that all of the trees manifested thedesired improved growth. The largesttrees were ‘Ferrastar’ and (486x217)144.The smallest tree was TNPx335x486.The rest of the cultivars were placed inbetween.
The trees were tall and well developed.The distance between the trees within therow are already narrow. However, the or-chard is young, and it is too early to con-sider any conclusions. In summary, thebest cultivars, according to the first re-sults of the field test and nut judgementare ‘Lauranne’, TNP 335 (89) (887), ‘Fe-rragnes’ and ‘Ferraduel’.
The second best cultivars were (335 x217) 92 and (486 x 217) 144. Cultivar(335 x 217) 19 failed. The reproductivityof the best cultivars in the first years washigh and promising, resembling that ofthe orchards in Paramont California andMonduelle Mas d’Asport in France, yiel-ding 3 t/ha.
It is expected to obtain higher and betterquality yields in these new orchards. Thetrees in our lot were not pruned during thefirst years, while building the trees. All ofthe branches were banded, leaving onlyfour main upright branches. This proce-dure enables all of the growth to be usedfor fruiting. The fertirrigation practiceswere similar to those we applied to earlypeaches and nectarines and they will bemodified in accordance with the results ofour trials. Irrigation is given daily, using asolution applied by drippers. This orchard
has been visited by many Israeli and fo-reign almond growers. All of them werevery impressed with the capacity, poten-tial yields, and nut quality of the trees.Last year 300 ha were planted in Israelusing our new concepts. However, welack the vegetative material of GF-677.Solving this problem will enable us toplant larger areas, especially in order toreplace low-profit orchards such as avo-cado and citrus in unfavourable regions.
ACKNOWLEDGEMENTSWe thank our technicians, Y. Bar-Yacovand C. Khatib who have helped us ingrowing the orchard and collecting thedata. We would also like to thank theExtension Service officers, R. Birgir andA. Lin, who participated in all of the al-mond trials.
R. AssafDepartment of Fruit Culture, ARO
Newe Ya’ar Regional Research Center, Israel
THREE NEWSELF-COMPATIBLE ALMOND
CULTIVARS FROM ZARAGOZA
INTRODUCTIONSince the beginning of almond (Prunusamygdalus Batsch) research in 1966 atZaragoza it was evident that the most im-portant problem in the Spanish almondproduction was its very low productivity,as it did not even reach 125 kg/ha, whatwas often not enough to cover the fixedcosts of almond growing (Felipe, 1984).This low productivity was mainly due totwo causes: the incidence of spring frostsand a deficient pollination. The incidenceof frosts at bloom is important in all inlandgrowing regions of Spain, but also inter-mittently in the coastal regions. Conse-quently, late blooming became a trait ofthe most importance for almond cultivarsas this has been the first fruit species tobloom. One of the first steps then under-taken was the establishment of a collec-tion to include the most important culti-vars of almond grown throughout theworld, but mainly the late-blooming culti-vars from different foreign countries(France, Italy and the then Soviet Union)as most of the Spanish cultivars resultedearly or only middle blooming.
Late blooming cultivars offer two evidentadvantages. Firstly they can bloom whenthe risk of late frosts is low or even mini-mal, thus escaping from the loss causedby low temperatures. Secondly, theybloom when temperatures are higher andthus more favourable for the processes ofpollination and fertilization.
At the same time it was shown that a defi-cient pollination was a problem found inmost almond orchards. This was mainlydue to the fact that many of them wereplanted with the two main Spanish culti-vars, ‘Marcona’ and ‘Desmayo Largueta’,
Fig. 1.- Fruits of the three new cultivars.

16 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
which do not coincide in bloom sufficient-ly to allow a good pollination betweenthem. Besides, the presence of pollina-ting insects was very low as it only reliedon the presence of wild bees and beehi-ves were not introduced at bloom into theorchards. The weather conditions duringthe early bloom of almond are often ina-dequate for bee flight, decreasing the effi-ciency of the low population of pollinatinginsects. For all these reasons from thevery beginning self-compatibility was con-sidered as a primary objective of the al-mond breeding programme to avoid therequirement of two simultaneouslyblooming cultivars, the presence of polli-nating insects and the occurrence of fa-vourable weather conditions, becauseself-compatible cultivars can be pollina-ted by the pollen of the same flowerwithout the requirements of any foreignintervention.
The low productivity of many Spanish al-mond orchards was aggravated by thepresence of a very large number of localcultivars, often of low quality and comingfrom the empirical selection made bygrowers along several centuries. Only afew cultivars, as those mentioned, ‘Mar-
cona’ and ‘Desmayo Largueta’, are reallyof good quality, although early blooming.
To solve these problems an almond bree-ding programme was started in 1974 (Fe-lipe and Socias i Company, 1985). At thesame time, the study of the pollination re-quirements of the late blooming cultivarsintroduced in the collection (Herrero etal., 1977) allowed to identify the self-com-patibility of some of them (Herrero andFelipe, 1975). The first step in this selec-tion and breeding work allowed the rea-lease of three new cultivars, ‘Guara’,‘Aylés’ and ‘Moncayo’ (Felipe and Sociasi Company, 1987), but the breeding pro-gramme was followed because none ofthese three cultivars fulfilled all the requi-rements looked for in a cultivar, takingalso into account that these requirementsmay change depending on the growingregion (Socias i Company et al., 1998).
Now three new cultivars from our bree-ding programme are described, characte-rized by their different blooming times(Fig. 1) and kernel type but with two com-mon traits: the three are autogamous,thus not requiring any foreign interventionfor their proper pollination and conse-
quently for the production of a commer-cial crop, and besides their kernels showno doubles.
ORIGIN OF THE NEW CULTIVARSThe three new cultivars come from artifi-cial or natural pollinations, following thetraditional steps in a fruit breeding pro-gramme (Socias i Company and Felipe,1987). The fruits were stratified and theseedlings were planted in a bed previous-ly to their transfer to the field, where theywere studied to proceed to their scree-ning according to the selection process.The first characters to be consideredwere self-compatibility, blooming time,morphological traits of the plant and thefruit, kernel quality... Once the best seed-lings were selected, they were graftedand planted in a study plot in order todeeply characterize the most importanttraits, as the effectiveness of self-pollina-tion (Ben Njima and Socias i Company,1995a and b), the morphological and bio-chemical characteristics of the plants(Bernad and Socias i Company, 1994),the flower and blooming characteristics(Bernad and Socias i Company, 1995)and the branching habit and bloom densi-ty (Bernad and Socias i Company, 1998).
‘Blanquerna’ comes from ‘Genco’ self-pollinated. ‘Genco’ is probably the self-compatible cultivar from the Italian regionof Puglia of higher quality, but it has notbeen widely used in the almond breedingprogrammes because of its bloomingtime, a little earlier than ‘Tuono’, whichotherwise has been repeatedly utilized asa parent. The blooming time of ‘Blanquer-na’ is middle and, consequently, is onlyrecommended for planting in regions withlow frost risk. Its ripening time is very ear-ly, which allows harvest when temperatu-res are high for a quick drying processand thus an early marketing. Its name co-mes from the philosophical novel ‘Blan-querna’, a book written by Ramon Llull(1232/33-1315/16), probably the mostuniversal man born in Majorca, the homeisland of one of the obtentors (RSiC).
‘Cambra’ comes from the cross ‘Tuono’ x‘Ferragnès’, having inherited many goodtraits from ‘Ferragnès’, but also with agood level of autogamy. Its name is ahommage to Mariano Cambra Ruiz deVelasco (1916-1985), a remarkable fruitresearcher, interested in all fruit speciesand a pioneer in the study of almond polli-nation (Cambra, 1954).
‘Felisia’ comes from the cross ‘Titan’ x‘Tuono’. ‘Titan’ is a Californian cultivarwhich has transmitted an allele for lateblooming (Socias i Company et al., 1999),making ‘Felisia’ the latest blooming culti-var so far released. Its name is a homma-ge to one of the obtentors (AJF) and the
Young almond orchard

17FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
institution where the work has been deve-loped, Servicio de Investigación Agroali-mentaria (SIA).
REFERENCES
Ben Njima N. and Socias i Company, R.1995a. Efecto del estilo en la velocidad decrecimiento de los tubos polínicos. Invest.Agrar., Prod. Prot. Veg., 10 (1): 39-45.
Ben Njima N. and Socias i Company, R.1995b. Characterization of some self-compatible almonds. I. Pollen tubegrowth. HortScience, 30 (2): 318-320.
Bernad D. and Socias i Company, R.1994. Caracterización morfológica ybioquímica de algunas selecciones au-tocompatibles de almendro. Inf. Técn.Econ. Agrar., 90V (2): 103-110.
Bernad D. and Socias i Company, R.1995. Characterization of some self-compatible almonds. II. Flower phenolo-gy and morphology. HortScience, 30(2): 321-324.
Bernad D. and Socias i Company, R.1998. Bud density and shoot morpholo-gy in some self-compatible almond se-lections. Acta Hort., 470: 273-279.
Cambra M. 1954. Polinizaciones en al-mendro ‘Desmayo’. An. Estac. Exp.Aula Dei, 3 (2): 229-232.
Felipe, A.J. 1984. Profitability of almond or-chards in Spain. Acta Hort., 155: 287-290.
Felipe, A.J. and Socias i Company, R.1985. L’amélioration génétique del’amandier à Saragosse. Options Médi-terr. CIHEAM/IAMZ, 85/I: 9-14.
Felipe, A.J. and Socias i Company, R.1987. ‘Aylés’, ‘Guara’, and ‘Moncayo’almonds. HortScience, 22: 961-962.
Herrero M., Cambra M. and Felipe, A.1977. Interpolinización de variedadesde almendro. An. Inst. Nac. Invest.Agrar., Ser. Prod. Veg., 7: 99-103.
Herrero M. and Felipe, A.J. 1975. Polli-nisation de l’amandier. Incompatibilitépollen-style. II Coll. GREMPA, Mont-pellier-Nîmes, Septembre 1975.
Socias i Company R. and Felipe, A.J.(1987). La mejora genética del almen-dro. Frutic. Prof., 11: 64-66.
Socias i Company R., Felipe A.J. andGómez Aparisi, J. 1999. Genetics oflate blooming in almond. Acta Hort.,484: 261-265.
Socias i Company, R., Felipe, A.J., Gó-mez Aparisi, J., García J.E. and Dicen-ta, F. 1998. The ideotype concept in al-mond. Acta Hort., 470: 51-56.
R. Socias i Company and A.J. FelipeUnidad de Fruticultura, SIA-DGA
Apartado 727, 50080 Zaragoza, SpainE-mail: [email protected]
ABOUT THE POSSIBLERELATIONSHIPS BETWEEN
Amygdalus webbi Spach andAmygdalus communis L.
On the “Genetic Resources and CropEvolution” journal, No 46; 1999, a paperentitled “On the origin of almond” by G.Ladizinsky from the Hebrew University ofJerusalem, Faculty of Agriculture, waspublished. In his work, G. Ladizinsky exa-mines the domestication of A. communisand explains the difficulties of identifyingthe almond’s wild ancestors. The authortakes into account some wild almond spe-cies and at the end of the paper consi-ders both A. korshinskyi and A. webbii asderived forms and not genuine wild typesand concludes finding out A. fenzliana as“the most plausible ancestor of almond”.
Writing about A. communis and A. webbiiG. Ladizinsky repeatedly calls me intoquestion, often without the expected ac-curacy. That is why I decided to answer inorder to clarify some of the unconvincingstatements of G. Ladizinsky.
G. Ladizinsky reports the content of a vi-sit to Apulia on 1996. Actually, in the mor-ning of May 30, I was pleased to take theIsraeli colleague and show him one of thesites where A. webbii is naturally spread inthe province of Bari. The right name of thesite is Contrada Ferratelle between Masse-ria Nova del Duca and Murgia del Ceraso.The site is the same where I use to take allvisiting scientists dealing with cultivatedand wild almonds, as, for instance, C.Grasselly, D.E. Kester and his colleagues(see Rivista di Frutticoltura, 1, 1996, page73). It is the first site where I found A. web-bii in Apulia and furthermore it is not farfrom Bari and easily reachable by car.
I own that the article of G. Ladizinsky hada positive effect on me because it urgedme to take up again field prospections.The result was the finding in Apulia of se-veral additional sites of natural diffusionof many hundreds individuals of A. webbiiin late summer 1999. Everywhere in theprovince of Bari, including the site visitedby G. Ladizinsky, A. webbii populatessunny places and is associated mainlywith P. amygdaliformis and also with C.monogyna: I never found the dwarf A.webbii and P. amygdaliformis mingledwith the tall, invading, dominant and sha-ding Quercus sspp. In the province of Fo-ggia, on Gargano headland, important A.webbii populations are mingled almostexclusively with bushy wild olive (Oleaeuropaea var. oleaster L.).
Regarding the commercial distribution offruit groves in Apulia region, it is possible
to summarize that in rainfed areasgrowers used to keep best soils as for po-sition, depth and fertility for grapevine,assigning low hilly, medium fertility anddepth soils to olive. From time immemo-rial almond crop is forced to utilize theworst arable soils, i.e. those very poor,arid and shallow. Poor soils mean alsovery low yield and contribute to explainthe progressive decline of almond indus-try in the region during the past thirtyyears. Soils worse than those destined tocultivated almond as for altitude, fertilityand depth are unsuited for agriculturalpurposes. The remote moorland, wherethe human presence is scarse and whereany other fruit crop would be unsuccess-ful is the natural habitat of A. webbii andother wild forms/species. However, it isclear that it would be impossible to G. La-dizinsky “at an altitude of approximately500m, where hard limestone and terrarossa soil are common” to face somewhatdifferent from sporadic and very old com-mercial almond orchards, forming an in-termittent crown around the prominentand rocky centre populated by A. webbii,together with P. amygdaliformis and C.monogyna. In this way, it could be usefulto reappraise the importance of the ob-servation of the “exact planting pattern inall almond plantations in that place” con-sidering that the phenomenon really con-cerns only a few dozens of almond trees.
G. Ladizinsky reports that “in one particu-lar plot...a spinescent tree has emergedfrom the bottom of a dry trunk, indicatingthe kind of the rootstock”. I can add thatthe old time died trunk seems belongingto A. communis while the living rootstockbelongs to A. webbi. Grafting “in situ” cul-tivars is an ancient and common practicein the region, particularly for olive (ontoOlea europaea var. oleaster L.), fig (ontoFicus carica caprificus Risso) and pear(onto Pyrus amygdaliformis L.) but not foralmond, due to the absence of wild al-mond forms in plains, fertile and arablelands and the difficulties of grafting suc-cessfully adult stone fruit trees. The re-sults of the technique are plantations withspacing sintomatically erratic and wide.Numerous examples concerning oliveand fig grafted “in situ” onto the aboverootstock respectively still existeverywhere in the region. Cultivated pear(local early ripening cultivars) is presentin the region only as isolated trees, nearthe towns, mingled with major fruit crops,used for home consumption rather thanfor the market. Considering the pear fruittypology, I wonder why somebody shouldtake care of grafting P. amygdaliformis ina deserted and remote hilly moorland, farfrom any farmhouse? The truth is thatwhat observed by G. Ladizinsky is thesame, perhaps one and only example ofA. communis grafted long time ago onto

18 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
A. webbii which I repeatedly show andwill continue to show to all colleagues vi-siting that area, at least until it will exist.
What G. Ladizinsky affirms concerningthe gradual replacement of almond by vi-neyard in that area is not correct. Is it truethat vineyards are present only a few kilo-meters far as the crow flies, but in a com-pletely different environment regarding al-titude, soil depth and fertility.
Later on G. Ladizinsky writes that I wouldhave told him that “A. webbii from thatarea has a single flower bud per leaf axisand a low percentage of twins per nut, asin the common almond cultivars of the re-gion”. It is impossible that I stated theabove sentence because since 1979 Ihave been repeatedly writing just the op-posite. I wrote that A. webbii possessessome peculiar morphological traits and, inparticular, the tendency of the flowerbuds to bear two flowers instead of asingle one (and not a single flower budper leaf axis) and the tendency to givehigh amount of doubles (twins is a com-pletely different kernel characteristic),and that these traits can be easily foundin cultivated almond population native ofApulia (Godini, 1979).
Moreover G. Ladizinsky affirms that “the-re is a notion that the self-compatibility ofsome almond cultivars of southern Italyhas been contributed by A. webbii, butaccording to Dr. Godini self-compatibilityhas not been found yet in this taxon”. Itcould be of interest to recall that self-compatibility has been found to be natu-rally possessed exclusively by 30 Apulianalmond cultivars so far and the concer-ning list is reported in Table 1. In particularthe four Apulian cultivars found to be self-compatible in 1992 belong to the called bit-ter types because of the high amygdalincontent in the kernel. Maybe an intermedia-te step of the long chain confirming the re-lationships between the two extreme rings,A. webbii and A. communis?
Therefore I do not fear confessing the au-thorship of the above “notion”. In 1979 Iwrote a paper entitled “Ipotesi sulla com-parsa dell’autocompatibilità nel mandorlo”(Hypothesis about the origin of self-com-patibility in almond”), Scienza e TecnicaAgraria, 19 (2-3):3-9, where I put forwardthe hypothesis that the origin of self-com-patibility was due to remote and sponta-neous hybridization between cultivatedalmond (A.communis) and the only self-compatible wild almond known up to now
(A. webbii). Really, no man’s-land rollingstrips normally separate the typical areasof diffusion of true to type A. webbii andtrue to type A. communis and over there itis possible to observe isolated almondseedlings at different hybridization de-gree between the two species concerningtree habit, shoot thorniness, shell size,shape, thickness and hardness, doublesand kernel taste. Nowadays, the aboveintuition has been unanimously acceptedby the whole of the pomologists whichdeal with almonds, such as I. Batlle, F.Dicenta, H. Duval, A.J. Felipe, T.M. Grad-ziel, Ch. Grasselly, D.E. Kester, R. So-cias i Company, F.J. Vargas, in rigorousalphabetical order.
As concerns the presence of self-com-patibility in the taxon A. webbii the sen-tence of G. Ladizinsky must be correc-ted as follows:”… but according to Dr.Godini self-compatibility has not beensearched yet in this taxon”. Really, atthat time I based my intuition on i) thepersonal finding of several self-compati-ble Apulian almonds, ii) the statementsof Ch. Grasselly (1977) and Felipe andSocias i Company (1977) about the self-compatibility of A. webbii and iii) thepresence of A. webbii in Apulia.
I already wrote that Ladizinsky’s paperhad positive effects on me. Besides thenew A. webbii findings, this current year Istarted with some researches aiming tocheck the presence of self-compatibility inA. webbii individuals. Work is in progressand I can advance that the use of fluores-cence microscopy is now allowing me andcoworkers to find several biotypes in whi-ch the pollen tubes grow down to the ownstylar tissues therefore resulting presu-mably able to set fruits with their own ga-metes. This ability has been ascertainedin a high percentage of the biotypes tillnow observed.
I confess not to be an expert on botani-cal taxonomy and yet I trust in scientistswho continue to consider A. webbi atrue species and recognize this wild al-mond among the species belonging tothe genus Amygdalus (Spach, 1843;Kester and Asay, 1975; Crossa Ray-naud and Grasselly, 1980; Browzic andZohary, 1996, etc.). In a work entitled“Random amplified DNA polymorphismsin Amygdalus communis L. and A. web-bii Spach” by P. Resta, G. Corona, G.Fanizza, M. Palasciano and A. Godiniand presented as oral paper at the II In-ternational Symposium on Pistachiosand Almonds held in Davis, USA, Au-gust 24-29, 1997, the authors concludedthat the group including A. webbii plantsgave “the most clearly distinguishedcluster of the dendogram, as expectedfor a related species”.
Table 1. Naturally self-compatible Apulian almonds till now individuated
Cultivar Reference
1. ‘Tuono’ (Syn. ‘Mazzetto’) Jaouani, 1973; Godini, 1975; Grasselly and Olivier, 19762. ‘Falsa Barese’ Godini, 19753. ‘Ferrante’ Godini, 19754. ‘Filippo Ceo’ Godini, 1975; Grasselly and Olivier, 19765. ‘Genco’ Grasselly and Olivier, 1976; Godini, 19776. ‘Patalina’ Godini, 19777. ‘Sannicandro’ Godini, 19778. ‘Scorza Verde’ Godini, 19779. ‘Antonio De Vito’ Reina, Giorgio, Godini, 1986
10. ‘Bianchi’ Reina, Giorgio, Godini, 198611. ‘Calò’ Reina, Giorgio, Godini, 198612. ‘Dicoladonato’ Reina, Giorgio, Godini, 198613. ‘Gargano’ Reina, Giorgio, Godini, 198614. ‘Irene Lanzolla’ Reina, Giorgio, Godini, 198615. ‘Laterza’ Reina, Giorgio, Godini, 198616. ‘Montenegro’ Reina, Giorgio, Godini, 198617. ‘Pepparudda’ Reina, Giorgio, Godini, 198618. ‘Pignatelli’ Reina, Giorgio, Godini, 198619. ‘Piscalze’ Reina, Giorgio, Godini, 198620. ‘Rachelecchia’ Reina, Giorgio, Godini, 198621. ‘Salvella’ Reina, Giorgio, Godini, 198622. ‘Torre’ Reina, Giorgio, Godini, 198623. ‘Ventura’ Reina, Giorgio, Godini, 198624. ‘Viapiano’ n 1 Reina, Giorgio, Godini, 198625. ‘Viapiano’ n 2 Reina, Giorgio, Godini, 198626. ‘Andria’ (bitter) Godini, de Palma, Palasciano, 199227. ‘Cicerchia’ (bitter) Godini, de Palma, Palasciano, 199228. ‘Gaetanuccia’ (bitter) Godini, de Palma, Palasciano, 199229. ‘Padula di Ruvo’ (bitter) Godini, de Palma, Palasciano, 199230. ‘Garibaldina’ Palasciano, Godini, 1999

19FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
I confirm the likelihood that A. webbi con-tributed to the origin of self-compatibilityin cultivated almond apart from its taxo-nomic classification. Thus the wild A.webbii could be considered an ancestorof A. communis at least as donor of self-compatibility.
Concluding, I can not believe that G. La-dizinsky upheld a preconceived idea inorder to affirm that A. fenzliana is themost plausible ancestor of almond. Really,the arguments advanced in favour of histhesis seem to me weak and unconvin-cing, as well as weak and unconvincingappears its refusal of admittance of anyrelationships between A. communis andA. webbii. It is possible that the inexact-nesses reported by G. Ladizinsky whichoriginated this clarification were due i) tothe time elapsed between the visit andthe drafting of his article or ii) to incom-prehension due to my approximateknowledge of English language. I preferto believe that the latter was the true cau-se. It remains to be seen whether andhow much the above misunderstandingscould have influenced the statementsmade by Ladizinsk in his paper.
REFERENCES
Browicz K., Zohary D., 1996. The genusAmygdalus L. (Rosaceae): Species re-lationships, distribution and evolutionunder domestication. Genetic Resour-ces and Crop Evolution, 43: 229-247.
Crossa Raynaud P., Grasselly C., 1980.L’Amandier. Maisonneuve et Larose,Paris.
Felipe A., Socias I Company R.,1977.Prima segnalazione di un mandorlo sel-vatico, probabilmente Amygdalus web-bii, in Spagna. III GREMPA Meeting,Valenzano (Italy), 3-7 October.
Godini A., 1975. Il mandorlo in Puglia:panorama della situazione varietale.Notiziario Ortoflorofrutticolo, 6.
Godini A., 1977b. Contributo alla conos-cenza delle cultivar di mandorlo (P.amygdalus Batsch) della Puglia: 2) unquadriennio di ricerche sull’autocom-patibilità. III GREMPA Meeting, Valen-zano (Italy), 3-7 October.
Godini A., 1979. Ipotesi sulla comparsadell’autocompatibilità nel mandorlo.Scienza e Tecnica Agraria, 2-3.
Godini A., de Palma L., Palasciano M.,1992a. Sur l’autocompatibilité de quel-ques cultivars amers parmi la popula-tion d’amandiers des Pouilles. 8th
GREMPA Meeting, Nimes (France), 26-27 June 1990. Rapport EUR 14081fr.
Grasselly Ch., 1977. Considerazioni su-lle caratteristiche di alcune specie sel-vatiche di mandorlo e sulle loro possibi-lità di utilizzazione in programmi di mi-
glioramento genetico. III GREMPAMeeting, Valenzano (Italy), 3-7 October.
Grasselly Ch., Olivier G., 1976. Mise inévidence de quelques types autocom-patibles parmi les cultivars d’amandier(P. amygdalus Batsch) de la populationdes Pouilles. Ann. Amélior. Plantes, 26.
Jaouani A., 1973. Etude de l’auto-compatibilité de la varieté “Mazzetto”.Activ. Lab. Arbor. Fruit., INRAT (In:Grasselly Ch. e Olivier G., 1976. Mise inévidence de quelques types autocom-patibles parmi les cultivars d’amandier(P. amygdalus Batsch) de la populationdes Pouilles. Ann.Amélior. Plantes,26.).
Kester D.E., Asay R., 1975. Almonds.In: Advances in fruit breeding (J.Janickand J.N. Moore editors). Purdue Univ.Press. pp: 387-419.
G. Ladizinsky, 1999. On the origin of al-mond. Genetic Resources and CropEvolution, 46: 143-147.
Palasciano M., Godini A:, 1999. Gari-baldina, an old and interesting self- fruitfulApilian almond. XI GREMPA Meeting,Sanliurfa (Turkey), 1-4 September.
Reina A., Giorgio V., Godini A., 1986.Cultivar autocompatibili nella popolazio-ne pugliese di mandorlo. Rivista di Frut-ticoltura, 12.
Spach E., 1843. Monographia generisAmygdalus. Ann. Sci. Nat. (Paris), 2,19.
A. GodiniDipartimento di Scienze delle Produzioni
VegetaliUniversità di Bari, Italy
E-mail: [email protected]
DWARF ALMOND (Amygdalusnana L.) IN YUGOSLAVIA
INTRODUCTIONAs a result of unfavourable climatic condi-tions for almond growing in the large partof FR Yugoslavia it is not grown widely.Production is estimated to be less than1% of the world (Korac et al, 1996). Forthe same reason even this small produc-tion varies between years. Almond in Yu-goslavia is mostly damaged by low tem-peratures at flowering time, being an ear-ly-flowering plant. One of the main aimsof almond breeding is, therefore, develop-ment of late flowering, productive culti-vars bearing high quality nuts, resistant tolow temperatures and pathogens. Con-cerning almond breeding, dwarf almond(Amygdalus nana L.) is very important be-cause it starts vegetation late and flowerslate avoiding spring and autumn frostsand it is resistant to low winter temperatu-
res and pathogens. It is also slightly vigo-rous which is an important characteristicrequired for intensive fruit production.Comparing to peach and almond thereare no high-quality vegetative rootstockswhich are not vigorous. Dwarf almond istherefore very important for developmentof such rootstocks.
MATERIALS AND METHODSIn the spring of 1990 we found dwarf al-monds in Deliblatska pe√cara (Cerovic etal, 1996) and transplanted thirty of themin the area of Novi Sad for research anduse in hybridization. As dwarf almond isreproduced by sprouts spontaneously,the material taken was rather uniform asit was taken from one tree.
Almond growth was studied by examininggrowth and diameter of the trees at theheight of 5 cm from the ground, at thestart and end of vegetation, start and endof flowering, as it is an important phenolo-gy, we assessed resistance to pathogensaccording to IBPGR nomenclature (1985)and it was crossed with vineyard peach(selections D-4 and D-6 from Novi Sad)and also with two late-flowering almondselections from Novi Sad.
RESULTSThe dwarf almond started its vegetationand flowered late. During four years theaverage date of flowering was 19th April -only six days before apple cultivar ‘Jona-than’, well known for its late flowering.Flowering phenology and foliation weresimultaneous. Corollas were intensivelypink.
Dwarf almond shows extremely low vi-gour. Five years old trees were, on avera-ge, 82 cm high and 11 mm thick at theheight of 5 cm from the ground. Nuts ofthe examined almond were small (avera-ge nut mass was 1.3 g), and the kernelswere bitter. Leaves were small, prolongedand elliptical and in September, evenwithout protection, completely healthy -they were assessed grade 1 (affected -grade 9).No freezing was recorded during the pe-riod of six years (1990-1996)). Dwarf al-mond is extremely resistant to low tempe-ratures which can be proved by the factthat its natural habitat is the surroundingsof the forest of Deliblatska pe√cara wheretemperature easily changes and reaches-30oC. It is also resistant to high tempera-tures and droughts which can also be in-ferred on the basis of its natural habitat.
Dwarf almond can be crossed with al-mond cultivars and selections and vine-yard peach. According to Rihter (1972)haploid number of chromosomes of com-mon almond (A. communis L.), dwarf al-mond (A. nana L.), and most other kinds

20 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
of almond (except A. pedunculata Maxim)is eight. According to the same author, Mi-churin often used A. nana var. sibirica (syn.A. ledebouculata) for hybridization. He cla-ims that cross genus hybridization can re-sult in self-pollinating almond cultivars aswell as cultivars resistant to Monilia laxa.
Considering characteristics of the exami-ned almond, it can be used for develop-ment of non vigorous vegetative root-stocks for peach and almond grafting,which are rather tolerant to high lime con-tents in the soil and resistant to Capnodistenebrionis L., as well as for developmentof almond cultivars characterized by lessvigour, higher resistance to low tempera-tures, droughts and pathogens. It is re-commended to use A. nana L. with sweetkernel (form dulcis), which is however di-fficult to find as dwarf almond rarelyyields in nature. We found that, under fa-vourable conditions, dwarf almond has a
higher growth, few deformed flowers andit yields well if it was provided a good po-llinator because, like most almond culti-vars, it is a cross-pollinated plant.
CONCLUSIONSOur research showed that dwarf almondfrom Deliblatska pescara is slightly vigo-rous. The average height of the five-years-old trees was 82 cm and averagetree circumference was 11 mm.
The examined almond flowered and leafedat the same time and very late. The start offlowering was between 11th April (1994)and 25th April (1996). In the period of sixyears (1990-1996) there were no damagesas a result of low temperatures.
There were no damages of pathogenseven before the protective treatment. InSeptember the leaves were on the treesand they fell in October affected by lowtemperatures.
It can be crossed with cultivars and selec-tions of almond and vineyard peach. It canbe used for development of non-vigorousvegetative rootstocks for almond andpeach grafting and for development of newcultivars of late-flowering almond with aslight vigour, high resistance to low tempe-ratures, drought and pathogens.
REFERENCES
Cerovic, S., Duric, B., Gasic, Ksenija,Golosin, B., Gvozdenovic, D., Kesero-vic, Z., Korac, M., Krstic, M., Ninic-To-dorovic, J., Ognjanov, V., Vujanic-Var-ga, D., 1996. Conservation of FruitGermplasm in the Balkan Peninsula.Budapest.
Korac,M., Golosin B., Ninic – TodorovicJ., Cerovic,S., 1996. Amygdalus nanaL. and its importance in breeding. Jour-nal of Yugoslav Pomology. No 113-114:151-154, Cacak.
Rihter, A.A., 1972. Mindal. Jalta.
M. Korac, B. Golosin, J. Ninic Todorovic,and S. Cerovic
Faculty of AgricultureNovi Sad, Yugoslavia
Table 1. - Beginning of flowering of dwarf almond and Novi Sad late-floweringalmond selection in Veternik (near Novi Sad)
Year Dwarf almond Novi Sad late-flowering(Amygdalus nana L.) almond selection
1993 24th April 23rd April1994 11th April 11th April1995 16th April 15th April1996 25th April 25th April
Almond blooming
INTEGRAL STUDYOF THE ECOPHYSIOLOGICAL
RESPONSES TO WATERSTRESS: CHARACTERIZATION
OF ALMOND VARIETIES.
Abstract of the PhD Thesis.Barcelona University,
Spain.
Almond (Prunus amygdalus Batsch, sin.dulcis (Miller) D.A. Webb) is a woodycrop, typically Mediterranean, which easi-ly adapts to region with scarce water re-sources (Kester and Asay, 1975; Grassellyand Duval, 1997). Mediterranean climatepromotes water stress in plants leading toreductions in yield and responses in frontof the environment (Rundel, 1998). In thiscontext, a better knowledge about plantmaterial characteristics can enable an op-timisation of its agronomic managementand running of plant breeding program-mes.
With these objectives, eight commercialalmond varieties from different originsand ranging in empirical drought resistan-ces, were ecophysiologically characteri-sed: ‘Desmayo Largueta’, ‘Falsa Barese’,‘Garrigues’, ‘Lauranne’, ‘Marcona’, ‘Mas-bovera’, ‘Nonpareil’ and ‘Ramillete’. Un-der field and semi-controlled conditionstheir water relations and hydraulic, mor-phologic and photosynthetic characteris-tics were studied. As a practical applica-tion for plant breeding in this species, thedescendants F2 from the hybrid crossingalmond cv. ‘Texas’ x peach (Prunus per-sica) cv. ‘Earlygold’ were characterisedand classified regarding their drought re-sistance.
Almond tree shows high response plasti-city on its tolerance mechanisms. ‘Mas-bovera’ and ‘Ramillete’ varieties have os-motic adjustment (Morgan, 1984 and1192; Turner, 1986); ‘Garrigues’, ‘Nonpa-reil’, ‘Marcona’ and ‘Lauranne’ have elas-tic adjustment (Savé et al., 1993); no os-motic neither elastic adjustment werefound in ‘Desmayo Largueta’. Some avoi-dance mechanisms, as cuticular transpi-ration rate and leaf abscission, were alsodescribed, contributing on a higher de-gree to the final response of this speciesin front of drought.
The vulnerability to cavitation of almondtrees shows intermediate behaviour be-tween temperate mesophyllous deciduo-us species and Mediterranean sclerophy-llous species. At water potential at turgorloss point, this species shows around a70% of percentage of loss of xylemhydraulic conductivity. The relationshipbetween maximum specific conductivityand hydraulic conductivity loss at turgorloss point, leads to higher water trans-

21FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
port efficiency, under water deficit con-ditions, on those varieties with highermaximum specific conductivity and in-termediate vulnerability to cavitation(Sperry, 1995; Jones and Sutherland,1991). This technique (de Herralde etal., 1997; Cochard et al., 1992; Sperryet al, 1988) has proved to be an easy,reproducible and reliable method for thehydraulic characterization of almondtrees; it also allows determining diffe-rences between cultivars.
Crown morphology determines the perfor-mance of photosynthesis and water rela-tions, and potentially modifies plant pro-ductivity. Open crowns with large leavesand balancedly distributed branch inclina-tion angles were considered advanta-geous for almond tree.
Almond is a species with high photosyn-thetic capacity, with high light saturationpoints (de Jong, 1983). Under irrigation,the different limitations to CO2 assimila-tion between cultivars are due to metabo-lism. Under water deficit conditions, sto-matal limitation is high in all the studiedcultivars, but no damage was detected onthe photosynthetic apparatus. Water useefficiency was one of the most discrimina-tory parameters between cultivars. Higherwater use efficiency, along the whole ve-getative and reproductive growth season,could be the most determinant factor onalmond production under dry growingconditions. The analysis of carbon isoto-pic composition and derived parametersare an easy and reproducible method,which gives an integrative value of wateruse efficiency (Farquhar and Richards,1984; Farquhar et al., 1989; Febrero etal., 1998). It also has the advantage ofbeing a reasonable cost measure, whichrequires little manipulation.
As species, almond submitted to suddenand severe water stress suffers reduc-tions of leaf water potential, promotingstomatal closure that leads to decrea-sing CO2 assimilation rates. However,when water stress is progressive alongthe whole growing season, water poten-tial is more negative but gas exchangeis only slightly reduced. This indicatesthe high degree of adaptation of thisspecies to dry conditions, which arecommon in the Mediterranean climaticregions.
According to the results obtained, the stu-died cultivars could be classified for theirdrought resistance and potential producti-vity under dry Mediterranean conditions.From high to low: ‘Masbovera’ and ‘FalsaBarese’, ‘Garrigues’, ‘Ramillete’ and ‘Mar-cona’, ‘Lauranne’, ‘Nonpareil’ and ‘Des-mayo Largueta’.
The methods and the information obtai-ned in the work above mentioned, allo-wed discarding 33% individuals from a F2,taking the characters that confer droughtresistance as a selection criteria. The me-thod was validated by the position obtai-ned by almonds, peach and the hybridwithin the classification. This method isproposed as an improvement on the earlyselection methods in fruit and nut treebreeding.
REFERENCES
Cochard, H.; Cruiziat, P.; Tyree, M.T.1992. Use of positive pressures to esta-blish vulnerabilty curves. Further sup-port for the air-seeding hypothesis andimplications for pressure-volume analy-sis. Plant Physiology, 100 (1): 205-209.
de Herralde, F.; Biel, C.; Cohen, M.; Bat-lle, I.; Savé, R. 1997. Vulnerability cur-ves to embolism and drought resistan-ce in two almond cultivars. Nucis, 6:12-14.
de Jong, T.M. 1983. CO2 assimilationcharacteristics of five Prunus tree fruitspecies. Journal of American Societyof Horticultural Science, 108: 303-307.
Farquhar, G.D. y Richards, R.A. 1984.Isotopic composition of plant carboncorrelates with water-use efficiency ofwheat genotypes. Australian Journal ofPlant Physiology, 11: 539-552.
Farquhar, G.D., Elheringer, J.R. y Hu-bick, K.T. 1989. Carbon isotope discri-mination and photosynthesis. AnnualReview of Plant Physiology and PlantMolecular Biology, 40: 503-537.
Febrero, A., Fernández, S., Molina-Cano, J.L. y Araus, J.L. 1998. Yield,carbon isotope discrimination, canopyreflectance and cuticular conductanceof barley isoline of differing glaucous-ness. Journal of Experimental Botany,49(326):1575-1581.
Grasselly, C. y Duval, H. (eds.) 1997.L’amandier. Centre technique interpro-fessionnel des fruits et légumes (Ctifl).167 pp.
Jones, H.G. y Sutherland, R.A. 1991.Stomatal control of xylem embolism.Plant, Cell and Environment, 14 (6):607-612.
Kester, D.E. y Asay, R. 1975. Almonds.pp. 387-419. En: Advances in FruitBreeding. Purdue University Press.
Morgan, J.M. 1984. Osmorregulationand water stress in higher plants. An-nual Review of Plant Physiology, 35:299-319
Morgan, J.M. 1992. Osmotic compo-nents and properties associated withgenotypic differences in osmoregulationin wheat. Australian Journal of PlantPhysiology, 19:67-76.
Rundel, P.W., 1998. Landscape distur-bance in Mediterranean-type ecosys-tems: an overview. Pp. 3-22. En: P.W:Rundel, G. Montenegro y F.M. Jaksic(eds.) Landscape Disturbance and Bio-diversity in Mediterranean-Type Eco-systems. Ecological Studies 136. Sprin-ger-Verlag, Berlin.
Savé, R., Alegre, H., Pery, M. y Terra-das, J. 1993. Ecophysiology of after-fireresprouts of Arbutus unedo L. Orsis 8:107-119.
Sperry, J.S. 1995. Limitations on stemwater transport and their consequen-ces. pp. 105-124. En: B.L. Gartner(ed.). Plant Stems: Physiology andFunctional Morphology. PhysiologicalEcology Series. Academic Press. SanDiego.
Sperry, J.S., Donelly, J.R. y Tyree,M.T. 1988. A method for measuringhydraulic conductivity and embolism inxylem. Plant, Cell and Environment,11: 35-40.
Turner, N.C., 1986. Adaptation to waterdeficits: a changing in perspective. Aus-tralian Journal of Plant Physiology, 13:175-190.
F. de HerraldeIRTA – Centre de Cabrils
Dept. Tecnologia HortícolaCrta. de Cabrils, s/n
08348 – Cabrils, SpainPhone: 34- 93 750 75 11
Fax: 34- 93 753 39 54E-mail: [email protected]
Almond potted trees of the drought trial at IRTACabrils, Spain

22 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
IDENTIFICATION, ISOLATIONAND CHARACTERIZATION OF
GENES RELATED TO DROUGHTSTRESS IN ALMOND
INTRODUCTIONThe Mediterranean region is an importantalmond (Prunus amygdalus [L.] Batsch)producing area, where the characteristicsevere drought periods together with latespring frost are the main limiting factorsreducing productivity. In an attempt to in-troduce drought resistance at IRTA’sscion almond breeding programme, bothIRTA and CSIC started a multidisciplinaryproject that includes different approachessuch as ecophysiological characterizationof different almond cultivars, the study ofagronomical parameters and moleculargenetics.
One of the challenges of plant biotechno-logy is to elucidate the complexity of thisresponse in order to improve the capacityof plant survival and to give high yieldsunder unfavourable conditions. Duringthe last few years, many efforts havebeen focused on the identification of rele-vant genes and on the study of their func-tion during plant dehydration. A variety ofdifferent biological systems had beenused in order to identify some of the ge-nes involved in the molecular responsesof plants against dehydration. 1) Plantseeds have been largely used as a modelto analyse gene expression due to theirability to survive severe water loss duringtheir final maturation stage of develop-ment (Goday et al., 1988). Furthermore,many of the genes and proteins that areinduced or accumulated in the matureembryo are also detected in vegetativetissues during water deficit, and thushave been related to desiccation toleran-ce. 2) Another approach that has givenvery useful information is the study of ve-getative tissues of resurrection plants thatcan survive severe water loss (Bartels etal., 1990). The genes identified on thisspecies are quite similar to those expres-sed in seeds of other species. 3) A widerange of mutants affected in the responseto dehydration have also been very usefulin order to analyze crucial steps in thetransduction signal that follows waterstress perception (Giraudat et al., 1994).4) And finally, many different crop plantshave also been used to analyse gene ex-pression during drought stress. Amongthe most studied species we can includecotton, rice, maize and barley. The infor-mation generated with this kind of studiesis very useful because it can be applied tothe molecular breeding programmes inagronomically important species.
The abiotic stresses such as drought,cold and high salinity represent the most
limiting factors in crop production, and allof them have as a consequence changesin water relation in plant tissues, andthus, lead to similar molecular responsesin the plant. Once plants have a loss ofwater, a cascade of events occur at thebiochemical and genetic level, such aschanges in membrane characteristics,protein turnover, osmolyte biosynthesis,ion uptake, hormone sensitivity and geneinduction. The plant hormone abscisicacid (ABA) plays an important role in theregulation of several aspects during seeddevelopment and in the plant response toenvironmental stresses such as desicca-tion, salt or cold. ABA levels increase du-ring embryo development and in vegetati-ve tissues under water deficit conditionsand it is associated to the expression ofspecific genes involved in the response todehydration (Campalans et al., 1999; Le-ung and Giraudat, 1998). The genes in-duced during water deficit codify for pro-teins involved in a high diversity of cellu-lar mechanisms. Some of these proteinshave known functions in protecting andrepairing cellular damage caused bywater loss, such as proteins involved inthe metabolism of various osmoprotec-tants, transport proteins, proteins invol-ved in protein degradation and repair pro-cesses and detoxifying enzymes. Otherproteins that are also induced during
water deficit, have an unknown functionsuch as cell wall proteins, lipid transferproteins, histones and LEA proteins (Fi-gure 1).
ALMOND: ANALYSIS OF A COMPLEXRESPONSEAlthough the molecular responses to de-hydration have been extensively studiedand a lot of information has been obtai-ned from the different approaches, thecomplexity of the response has not yetbeen completely elucidated. Furthermore,most of the information has been obtai-ned from herbaceous species, and reportsinvolving woody plant are limited (Artlip etal., 1997). In order to identify some of thegenes involved in drought response in al-mond we have mainly used two experi-mental models: the embryogenesis/ger-mination process and “in vitro” culturedalmond plantlets submitted to differenttreatments. The almond cultivar used inthese studies was ‘Masbovera’, obtainedat IRTA Mas Bové, Reus (Figure 2), dueto its good productivity and tolerance todehydration conditions.
Embryo development in almond is a pro-cess that takes about 200 days from theperiod of maximum flowering (DAF), be-ginning the desiccation around 100 DAFand being maximum at the mature embryo
detoxification, degradation and repairpipid metabolismtransportbiosynthesis of osmolytescell wall compositionhistonesLEA proteisn
Water deficit
leaf abscisionstomatal closureembryo developmentgerminaton
ABA
GENE INDUCTION
singnal transduction
protein degradationand repair
mRNA stability
Embryo development
Low temperatures
dehydration
Salinity
accumilation ofcompatible solutes
ion uptake, sequestrationand partitioning
membrane composition
Figure 1. Schematic representation of the complexity of plant responses to environ-mental and developmental conditions. Different situations lead to similar molecular
and biochemical responses in plant tissues. A variety of genes are induced as aresponse to water deficit or ABA and have very different roles in the plant response
to dehydration.

23FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
with water contents below 10%. In orderto identify some of the polypeptides thataccumulate in the mature embryo, two-di-mensional protein patterns were analysedat different stages of embryo develop-ment. Those proteins that highly accumu-late in the last stages of embryogenesisare of special interest in this study becau-se they may be related to desiccation to-lerance.
In order to directly identify some of thegenes involved in drought tolerance in al-mond, we have also used a differentialexpression technique such as cDNA-AFLP (Amplification fragment lenght poly-morphism) (Bachem et al., 1996) to com-pare the mRNA population between ma-ture and developing embryos and in vitrocultured plantlets submitted to waterstress and ABA treatments. By using thisapproach we have been able to identifydifferent genes possibly involved in thealmond response against dehydration.Some of the identified genes have beenpreviously described in other plant spe-cies to be induced under water stressconditions such as low molecular weightheat shock proteins, possibly involved intissues protection against dehydration,cysteine proteases that should be relatedto protein degradation processes thattake place during water loss, or enzymes
involved in the biosynthesis of lipids thatcould be responsible for some of the alte-rations on membrane composition thattake place during water deficit. Other ge-nes identified in this study are of un-known function at the moment and mayrequire further investigation.
Late embryogenesis abundantproteinsA very well studied group of proteins thathighly accumulate during the last stagesof embryo development are LEA proteins(for late embryogenesis abundant), alsonamed RAB for their responsiveness tothe phytohormone abscisic acid (ABA).LEA proteins are a complex family of pro-teins ubiquitous among photosyntheticorganisms that have been identified incyanobacteria and plants, both monocotsand dicots, and they are thought to be in-volved in protecting proteins and mem-branes against dehydration (Close,1996). Using the information on LEA pro-teins available from different plant spe-cies, we have been able to identify twoLEA proteins from almond (Campalans etal., 2000). One of these proteins belongsto the group 2 of LEA proteins that havebeen postulated to protect cells againstdehydration by maintaining protein andmembrane structure, sequestering ions,binding water and acting as molecular
chaperones. The other LEA protein iden-tified belongs to group 5, that although itis the only group of LEA proteins that arenot highly hydrophobic, has also been re-lated to desiccation protection but with apossibly different way of action (Arenas-Mena et al., 1999).
THE USE OF GENETIC MAPSThe resistance to abiotic stresses, and inparticular to drought stress, is very com-plex, and it is not possible to measure itas a discrete response. This means that itis necessary to develop new systems toallow the genetic study of these complextraits by using quantitative genetics. Thedevelopment of molecular markers inplant species has opened the possibilityto construct dense genetic maps in anyplant species. The main advantage ofthese kind of maps, in terms of plantbreeding, is the coverage of the wholegenome. This allows us to screen geno-mes in order to find markers linked to anyquantitative trait (QTL), and to select foror against the trait using the marker inany tissue or developmental stage. Fur-thermore, if the molecular marker con-sists of a known gene with a known func-tion, the information of the map is muchmore useful, and can allow for the corre-lation of a gene with a specific trait. Thegenes identified in this study have been lo-cated in the Prunus genetic map (Joobeuret al., 1998) in order to be used as mole-cular markers in the breeding program-mes for the future generation of almondcultivars.
CONCLUDING REMARKSFrom an agronomically point of view,more than the generation of dehydrationresistant cultivars, it is interesting to ob-tain crops that can produce at optimal le-vels in unfavourable environmental condi-tions. Taking into account the complexresponse of plants against water deficit,with the induction of so many different ge-nes involved in such a variety of cellularmechanisms, we can conclude that it isnot possible to explain a specific respon-se just considering the expression of oneor a few number of genes.
According to that, the identification ofquantitative-trait loci (QTLs) for droughtresistance may be an effective analyticaltool. This type of approach is particularlypromising considering the saturatedDNA-marker maps that are now availablefor many different plant species, includingmodel and crop plants.
The information obtained with the analy-sis of the almond response to dehydrationcan be very useful for the future genera-tion of new cultivars with better produc-tion rates during dry periods. Furthermo-re, this information can also be applied to
Figure 2. ‘Masbovera’ almond trees growing at IRTA-Mas Bové, Reus.
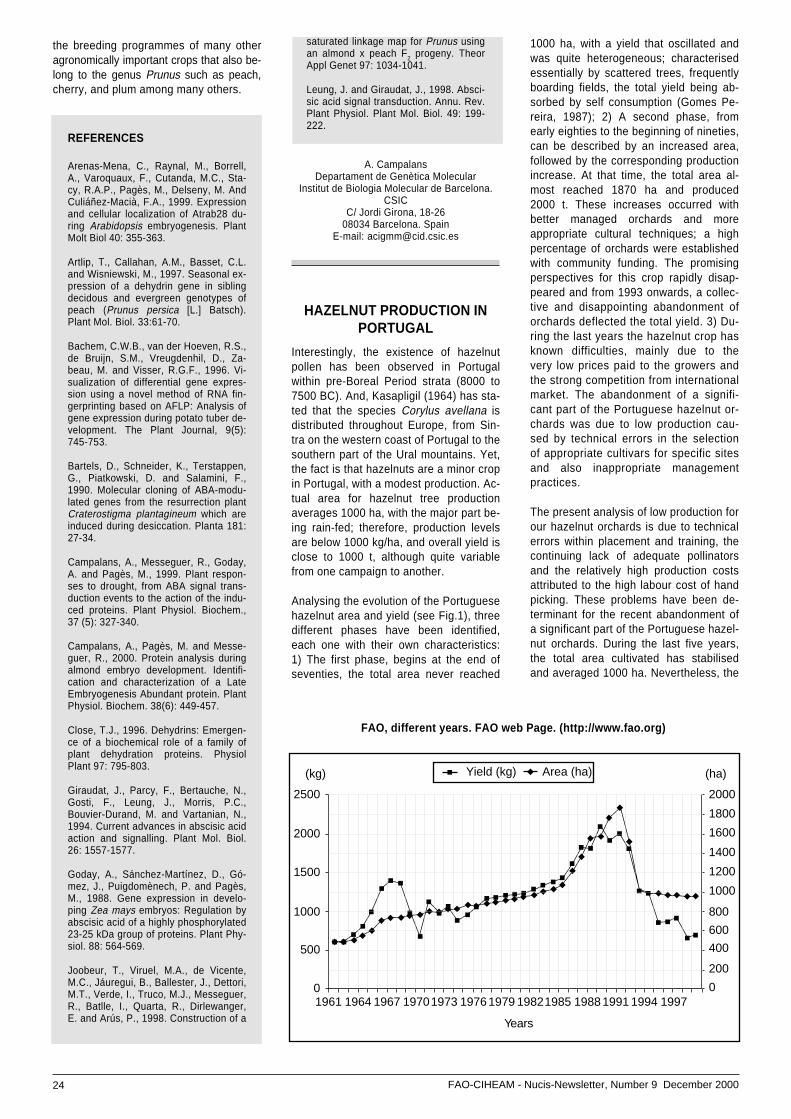
24 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
the breeding programmes of many otheragronomically important crops that also be-long to the genus Prunus such as peach,cherry, and plum among many others.
REFERENCES
Arenas-Mena, C., Raynal, M., Borrell,A., Varoquaux, F., Cutanda, M.C., Sta-cy, R.A.P., Pagès, M., Delseny, M. AndCuliáñez-Macià, F.A., 1999. Expressionand cellular localization of Atrab28 du-ring Arabidopsis embryogenesis. PlantMolt Biol 40: 355-363.
Artlip, T., Callahan, A.M., Basset, C.L.and Wisniewski, M., 1997. Seasonal ex-pression of a dehydrin gene in siblingdecidous and evergreen genotypes ofpeach (Prunus persica [L.] Batsch).Plant Mol. Biol. 33:61-70.
Bachem, C.W.B., van der Hoeven, R.S.,de Bruijn, S.M., Vreugdenhil, D., Za-beau, M. and Visser, R.G.F., 1996. Vi-sualization of differential gene expres-sion using a novel method of RNA fin-gerprinting based on AFLP: Analysis ofgene expression during potato tuber de-velopment. The Plant Journal, 9(5):745-753.
Bartels, D., Schneider, K., Terstappen,G., Piatkowski, D. and Salamini, F.,1990. Molecular cloning of ABA-modu-lated genes from the resurrection plantCraterostigma plantagineum which areinduced during desiccation. Planta 181:27-34.
Campalans, A., Messeguer, R., Goday,A. and Pagès, M., 1999. Plant respon-ses to drought, from ABA signal trans-duction events to the action of the indu-ced proteins. Plant Physiol. Biochem.,37 (5): 327-340.
Campalans, A., Pagès, M. and Messe-guer, R., 2000. Protein analysis duringalmond embryo development. Identifi-cation and characterization of a LateEmbryogenesis Abundant protein. PlantPhysiol. Biochem. 38(6): 449-457.
Close, T.J., 1996. Dehydrins: Emergen-ce of a biochemical role of a family ofplant dehydration proteins. PhysiolPlant 97: 795-803.
Giraudat, J., Parcy, F., Bertauche, N.,Gosti, F., Leung, J., Morris, P.C.,Bouvier-Durand, M. and Vartanian, N.,1994. Current advances in abscisic acidaction and signalling. Plant Mol. Biol.26: 1557-1577.
Goday, A., Sánchez-Martínez, D., Gó-mez, J., Puigdomènech, P. and Pagès,M., 1988. Gene expression in develo-ping Zea mays embryos: Regulation byabscisic acid of a highly phosphorylated23-25 kDa group of proteins. Plant Phy-siol. 88: 564-569.
Joobeur, T., Viruel, M.A., de Vicente,M.C., Jáuregui, B., Ballester, J., Dettori,M.T., Verde, I., Truco, M.J., Messeguer,R., Batlle, I., Quarta, R., Dirlewanger,E. and Arús, P., 1998. Construction of a
saturated linkage map for Prunus usingan almond x peach F2 progeny. TheorAppl Genet 97: 1034-1041.
Leung, J. and Giraudat, J., 1998. Absci-sic acid signal transduction. Annu. Rev.Plant Physiol. Plant Mol. Biol. 49: 199-222.
A. CampalansDepartament de Genètica Molecular
Institut de Biologia Molecular de Barcelona.CSIC
C/ Jordi Girona, 18-2608034 Barcelona. Spain
E-mail: [email protected]
HAZELNUT PRODUCTION INPORTUGAL
Interestingly, the existence of hazelnutpollen has been observed in Portugalwithin pre-Boreal Period strata (8000 to7500 BC). And, Kasapligil (1964) has sta-ted that the species Corylus avellana isdistributed throughout Europe, from Sin-tra on the western coast of Portugal to thesouthern part of the Ural mountains. Yet,the fact is that hazelnuts are a minor cropin Portugal, with a modest production. Ac-tual area for hazelnut tree productionaverages 1000 ha, with the major part be-ing rain-fed; therefore, production levelsare below 1000 kg/ha, and overall yield isclose to 1000 t, although quite variablefrom one campaign to another.
Analysing the evolution of the Portuguesehazelnut area and yield (see Fig.1), threedifferent phases have been identified,each one with their own characteristics:1) The first phase, begins at the end ofseventies, the total area never reached
1000 ha, with a yield that oscillated andwas quite heterogeneous; characterisedessentially by scattered trees, frequentlyboarding fields, the total yield being ab-sorbed by self consumption (Gomes Pe-reira, 1987); 2) A second phase, fromearly eighties to the beginning of nineties,can be described by an increased area,followed by the corresponding productionincrease. At that time, the total area al-most reached 1870 ha and produced2000 t. These increases occurred withbetter managed orchards and moreappropriate cultural techniques; a highpercentage of orchards were establishedwith community funding. The promisingperspectives for this crop rapidly disap-peared and from 1993 onwards, a collec-tive and disappointing abandonment oforchards deflected the total yield. 3) Du-ring the last years the hazelnut crop hasknown difficulties, mainly due to thevery low prices paid to the growers andthe strong competition from internationalmarket. The abandonment of a signifi-cant part of the Portuguese hazelnut or-chards was due to low production cau-sed by technical errors in the selectionof appropriate cultivars for specific sitesand also inappropriate managementpractices.
The present analysis of low production forour hazelnut orchards is due to technicalerrors within placement and training, thecontinuing lack of adequate pollinatorsand the relatively high production costsattributed to the high labour cost of handpicking. These problems have been de-terminant for the recent abandonment ofa significant part of the Portuguese hazel-nut orchards. During the last five years,the total area cultivated has stabilisedand averaged 1000 ha. Nevertheless, the
1961 1964 1967 19701973 19761979 19821985 19881991 19940
200
400
600
800
1000
1200
1400
1600
1800
2000
1997
2500
2000
1500
1000
500
0
(kg)
Years
(ha)Area (ha)Yield (kg)
FAO, different years. FAO web Page. (http://www.fao.org)

25FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
yield has been decreasing, mainly due tounfavourable climatic conditions.
As shown on Fig. 2, hazelnut groves aremainly located in Beira Litoral (Viseu re-gion), Trás-os-Montes (Moimenta, Ser-nancelhe) and Beira Interior (Guarda,Fundão), which respectively account for52%, 37% and 6% of the country’s totaloutput. The remainder is produced in theAlentejo (Portalegre, Marvão) and EntreDouro e Minho (Amarante, Baião). Themap also reflects the statement of Carta-geno Ferreira, (1992) about the appro-priate ecological conditions for hazelnutproduction under most of the agriculturalregions in the North and central part ofPortugal.
Local cultivars are rare, but the produc-tion areas often include the utilization of alarge number of cultivars, sometimes ofundefined characteristics and low produ-cing. The main cultivars in old plantingsare “Fertile de Coutard”, “Grada de Vi-seu”, “Negreta”, “Tonda de Giffoni”,“Grossal”; most commonly used polini-zers are “Gunslebert”, “Merveille de Bo-
llwiller” and “Daviana”. “Butler” and “En-nis” are usually found in the new or-chards. Our current recommendation forthe ecological conditions of northern Por-tugal is to select among “Butler”, “Ennis”,“Tonda de Giffoni”, “Segorbe” and “F. deCoutrad” as producers and “M. de Bol-lwiller”, “Daviana” and “Gunslebert” aspolinizers, according to the results obtai-ned by Santos et al. (1999).
Most of the production is sold to the ma-jor population centres (Lisbon and Opor-to) by regional retailers and only smallquantities go directly from producers toconsumers. During the last two campa-igns, imports of hazelnut kernels wentdown from 39 to 15 t, while imports ofshelled hazelnuts increased by 36 t to atotal of 114 t (Anuário Hortofrutícola,2000).
Most hazelnut kernels are imported fromItaly, and shelled hazelnuts come fromSpain and Turkey. Portugal’s hazelnutexports go to the PALOP (African Coun-tries of Portuguese Official Language)nations, especially Angola, althoughquantities are almost insignificant.
ACKNOWLEDGEMENTSThanks are due to: J. Soeiro de Carvalhoe O. Borges, Direcção Regional de Agri-cultura de Trás-os-Montes e Alto Douro.A. Assunção, Direcção Regional de Agri-cultura do Minho Litoral. A. Dias Lopes,Direcção Regional de Agricultura do Min-ho Litoral
LITERATURE CITED
Anuário Hortofrutícola, 2000. GPPAA,Gabinete de Planeamento e PolíticaAgro-Alimentar, 288 pp.
Cartageno Ferreira, A.J., 1992. El ave-llano. Perspectivas de cultivo. Fruticul-tura Profesional, 48:21-24.
FAO, different years. FAO web Page.(http://www.fao.org).
Gomes Pereira, J., 1987. Frutos secosem Portugal. A história de um projecto.Vida Rural, 24:16-23.
Kasapligil, B., 1964. A contribution tothe histotaxonomy of Corylus (Betula-ceae). Adansonia 4:43-90.
Santos, A. e Silva, A. P., 1999. Comporta-mento agronómico de once cultivares deavellano. Fruticultura Professional, nº 104,Especial Frutos Secos II, 67-70.
A.P. Silva and A. Álvares dos Santos.Crop Science Department
University of Trás-os-Montes e Alto DouroPortugal
Phone: 351 59 32 0450Fax: 351 59 32 0629
E-mail: [email protected]
NUTRITIONAL VALUE OFHAZELNUT (Corylus avellana L.)
INTRODUCTIONIn 1994 Oldways Preservation & Exchan-ge Trust and WHO/FAO proposed thepyramid for the Mediterranean Diet, inclu-ding hazelnuts and other nuts near itsbase and between legumes, edible fruitsand vegetables. Hazelnut belongs to avery important group of foods, from thenutritional point of view, due to its chemi-cal composition.This note resumes actual knowledgeabout hazelnut composition. Data aretaken from both bibliography and resultsfrom IRTA’s research programme on ha-zelnut quality. For more information aboutthis topic check the references and web-sites at the end of this report.
HAZELNUT MEAN COMPOSITIONHazelnut kernel has low water level (5 %)that makes the concentration in any nu-trient very high. Major components of thekernel are (Figure 1): lipids (62 %), pro-teins (13 %), available carbohydrates (11%), dietary fiber (7 %) and ash (2 %). Allof them are mean values which can chan-ge depending on variety, climate and soilconditions of the production zone, theyear characteristics, etc. However, fromthe nutritional point of view, these varia-tions should not be considered since theirfinal influence on the consumers’ totaldiet is not relevant.With 600 kcal/100g, hazelnut shows avery high energy value. The presence ofan important fraction of substanceswithout any energy contribution (fiber andminerals) implies that hazelnut is a foodwith a very high nutritional density andnot only a caloric food.In relation to Mediterranean Diet, the oilcharacteristics (mainly fatty acid composi-tion) and dietary fiber (cellulose, hemice-llulose and lignin) with beneficial effectson health should be stood out. It must beremarked that nuts, legumes and whole-wheat bread are the main sources of die-tary fiber. Also, biophenols contents mustbe considered for their antioxidant activityand sensorial properties.
LipidsIn relation to hazelnut fat, its high contentin monounsaturated fatty acids (79 %),with 13 % of polyunsaturated and lessthan 8 % of saturated fatty acids must beemphasised. These are common to vege-tal fats in the Mediterranean Diet andshow a good effect on blood cholesterolmetabolism, reported in several epide-miological studies.The main monounsaturated fatty acid isoleic acid (C18:1) while linoleic acid(C18:2) is the main polyunsaturated (thisis an essential fatty acid belonging to w-6
Fig.2. Hazelnut production areas

26 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
group, which cannot be synthesised byhumans and should be supplied with thediet). Palmitic (C16:0) and stearic (C18:0)acids are the most usual saturated fattyacids. From the technological point ofview, this highly unsaturated compositionmeans that hazelnut is very sensible torancidity and must be handled with carein order to preserve its quality.
CarbohydratesThis group includes dietary fiber (7 %)and available carbohydrates (11 %) likestarch and dextrine (6 %) and availablesugars (5 %).
The major available sugar is sucrose (4.6%) followed by raffinose (0.3 %), glucose(0.2 %), fructose (0.2 %), myo-inositol(0.1 %) and other alcohol-sugars. From atechnical point of view, the distribution ofsugars is related to the preservation qua-lity of hazelnuts, as glucose and fructosecontents rises with storage time by sucro-se hydrolysis.
ProteinProtein content is high (13 %), mainly (80%) formed by globulin (most usual reser-
vation proteins in seeds) and albumin (withmetabolic functions, like enzymes), glutein(15 %) and very low levels of prolamin.
However, the most interesting is the qua-lity of hazelnut proteins, as they includeall the essential amino acids (aminoacids) in suitable proportions. Table 1shows amino acids composition in hazel-nut protein compared to ideal levels pro-posed by FAO (1973). It can be observedthat hazelnut provides almost all the ami-no acids in the correct levels, with sulphu-red amino acids group (methioni-ne+cystine) as limiting factor although itslevel is 94 % of FAO’s recommendation.
MineralsMajor minerals in the kernel are: K (6000ppm), P (3000 ppm), Ca (1940 ppm) andMg (1320 ppm). In a lower proportion Fe(33 ppm) is one of the main vegetal sour-ces for this element. However, to improveFe assimilation it must be combined withfoods rich in Vitamin C. Sodium (Na) levelis usually low (29 ppm), although high fluc-tuations may be observed, and this factallows to use hazelnuts in low sodium con-tent diets.
VitaminsHazelnut is very rich in vitamins, inclu-ding vitamin E (295 ppm) and C (35 ppm),Niacin (13.5 ppm), Pantothenic acid (11.5ppm), Vit B6 (4.5 ppm), Thiamine (3.9ppm), Riboflavin (2.1 ppm), Folate (0.71ppm) and Carotene (0.29 ppm).Hazelnut is the second natural source ofvitamin E after vegetal oils. It shows an-tioxidant activity scavenging free radicals.
CONCLUDING REMARKSPrevious data show that hazelnut has avery high nutritional value, rich in mo-nounsaturated fat (oleic acid), with goodlevel of dietary fiber, high content of mi-nerals (low in Na), with high quality pro-tein and very rich in vitamin E.In conclusion, hazelnut is a completefood with very good organoleptical cha-racteristics, that can be included easily inany kind of recipes and with various diffe-rent commercial presentations. Its inclu-sion in the pyramid of Mediterranean Dietis completely justified.
REFERENCES
Alphan, E.; Pala, M.; Açkurt, F.; Yilmaz,T., 1996. Nutrition composition of hazel-nuts and its effects on glucose and lipidmetabolism. Acta Horticulturae 445ISHS: 305-310.
Andreoni, N., 1996. Hazelnut phenolicsubstances as natural antioxidants.Acta Horticulturae 445 ISHS: 217-221.
Ayfer, M.; Uzun, A.; Bas, F. 1986. Tur-kish hazelnut cultivars. Black Sea Re-gion Hazelnut Exporters Union. 1-95.
Beuchat, L.R.; Worthington, R.E. 1978.Technical note: fatty acid compositionof tree nut oils. J. Fd. Technol. Vol. 13:355-358.
Botta, R.; Gianotti, C.; Me, G., 1996.Kernel quality in hazelnut cultivars andselections analysed for sugars, lipidsand fatty acid composition. Acta Horti-culturae 445 ISHS: 319-326.
García, R.; Carballido, A.; Díaz, A.1978. Contribución al estudio de losaceites de frutos secos españoles.Composición del insaponificable: II)Fracción metil esterólica. Anales deBromatología. vol.30(2): 175-186.
García, R.; Valls, C.; Díaz, A. 1979.Study of constituents of Spanish nutoils. Characteristics of hazelnut oilsfrom Tarragona (Harvest, 1976). Anal.Bromatol. XXXI-2: 137-158.
Garrone, W.; Vacchetti, M. 1991. Consi-derazioni sulla qualità delle nocciole uti-lizzate nell’industria alimentare. TheCracker. Nº 1: 19.
Hadorn, H.; Keme, T.; Kleinert, J.; Mes-serli, M.; Zürcher, K. 1978. Lagerungs-versuche und Qualitätsprüfungen anHaselnüssen. Gordian. vol.10: 300-310.
Water5% Protein
13%
Availablecarbohydrates
11%
Dietary fiber7%
Lipids61%
Minerals2%
Fig.1. Hazelnut mean composition
Table 1.- Comparison between hazelnut amino acid (a.a.) composition and thatproposed by FAO (1973). Last column shows their ratio as %.
Amino-acid Protein FAO (1973) Hazelnut Protein hazelnut/FAO(mg a.a. / g protein) (mg a.a. / g protein) (%)
Leucine 7.0 8.4 127Isoleucine 4.0 6.3 158Lysine 5.5 6.1 111Methionine+Cystine 3.5 3.3 94Phenylalanine+Tyrosine 6.0 15.8 263Threonine 4.0 4.0 100Valine 5.0 5.6 112Tryptophan 1.0 2.0 200

27FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
POSTHARVEST PHYSIOLOGYOF TURKISH FILBERT
(Corylus colurna L.)SEEDS
ABSTRACTIn this paper the results of the examinedinfluential factors related to the produc-tion of seeds material of Turkish filbert(Corylus colurna L.). Forty-five mothertrees were selected from the green areasof Novi Sad as a potential source of seedmaterial.
Experimental work in seed productionwas carried out in ecological conditions,involving micro-climate conditions in thegreen areas and the nursery, includedphenological observation, determinationof the right time of seeds collection, ex-traction of the nuts from cupules, prepa-ration, determination of the contents ofwater, proteins and oils, storage in labo-ratory conditions, cold stratification andtesting seed material in green housesand nursery conditions.
Results of the long-term research showedsignificant individual differences whichwere caused by genotype and influenceof ecological factors. Thirteen genotypeswith seed material showing extraordinaryphysiological results in seedling produc-tion were separated for special protectionand preservation of germplasm. The sug-gestion is to direct the production of plan-ting material of Turkish filbert to the diffe-rent needs and study biodiversity in foresthabitats of relict phytocenoses as in theIron gate region, oak and beach forests inSerbia and the western areas of Monte-negro.
INTRODUCTIONTurkish filbert (Corylus colurna L.) is atertiary relict. It is interesting for preserva-tion of biological diversity in habitats ofrelict phytocenoses, oak and beech fo-rests in Serbia and western areas of Mon-tenegro, and also as a grown species inpopulations of secondary origin. Pursuantto the Law on protection of the nature it isa law-protected species.
Phytocenoses of Turkish filbert in Yugos-lavia were investigated and their structuredescribed by Milevic (1951), Velickovic(1959), Misic (1966), Jankovic et al.(1984), Stojanovic and Jovanovic (1987).
Experimental research and a detailed stu-dy of the most dominant factors and de-termination of the most suitable technolo-gical methods in production of seed andplanting material of Turkish filbert(Corylus colurna L.) in Yugoslavia startedin 1983 (Ninic-Todorovic, 1990). The pre-selection included groups of trees and
populations of Turkish filbert in greenareas of Vojvodina, Novi Sad and sur-roundings, which adapted to urban condi-tions, considerably different from ecologi-cal conditions within the natural habitatsof this species.
Periods of intense flowering and seedproduction in cultivated trees are morefrequent comparing to the trees in the na-tural population, such as in relict phytoce-noses of the Iron Gate area, which wasincluded in the initial phase of the rese-arch. A small distance of the green areasin Novi Sad enabled a continuous obser-vation of all dominant factors of the seedsconcerning production of planting mate-rial of Turkish filbert. A close selectionwas made on 45 genotypes, 13 having anextremely high yield. These are groups ofphysiologically mature trees in the Futos-ki park, proposed for a special protectionand germplasm preservation.
Lagerstedt (1975) in his book “Advancesin fruit breeding” points out that Turkishfilbert has been used as a rootstock forhazel grafting since 1930. In Yugoslaviathe first results of hazel grafted onto Tur-kish filbert, using the method of graftingon a dormant bud, were first reported byKorac and Slovic (1973/74).
MATERIAL AND METHODSThe selection of Turkish filbert treesfrom the green areas was made accor-ding to the criteria for selection of horti-cultural seed objects (Stilinovic and Tu-covic, 1977). Phenological observationswere adjusted for Turkish filbert and thespecific ecological conditions (Ninic-To-dorovic, 1990). Collection, preparation,and determination of morphological,technological and the most importantnutritive components of the seeds aswell as the treatment and storage untilinvestigation of physiological characte-ristics were done by systematic obser-vation and with personal effort of the au-thor during the research for the doctoraldissertation and according to the metho-dology of Stilinovic (1985). The exami-nation of germination in controlled con-ditions was conducted according to acombination of Yugoslav and ISTA regu-lations for Corylus sp. (1985, 1993 cit.IPGRI Tec. Bul. 1996).
Phenological observations were based oninter-phases of development including thewhole period from the very beginning tothe full opening of male and female flo-wers, phenophase of foliage formation,cupules formation, development of cupu-les and nuts, physiological maturity anddetermination of right time for picking cu-pules, technological maturity and separa-tion of the nuts from the shell and leaf fall.Separation of the nuts from cupules and
Parcerisa, J.; Boatella, J.; Codony, R.;Farràn, A.; García, J.; López, A.; Ràfe-cas, M.; Romero, A. 1993. Influence ofvariety and geographical origin on the li-pid fraction of hazelnuts (Corylus ave-llana L.) from Spain: I. Fatty acid com-position. Food Chemistry. (48): 411-414.
Parcerisa, J.; Rafecas, M.; Castellote,A.I.; Codony, R.; Farràn, A.; García, J.;Gonzalez, C.; López, A.; Romero, A.;Boatella, J. 1995. Influence of varietyand geographical origin on the lipidfraction of hazelnuts (Corylus avellanaL.) from Spain: (III) Oil stability, toco-pherol content and some mineral con-tents (Mn, Fe, Cu). Food Chemistry.(53): 71-74.
Pszczola, D.E. 2000. Health and func-tionality in a Nutshell. Food Technology.54 (2): 54-59.
Quattrucci, E. 1996. Nutritional qualityof treenuts & how they complementother foods. The Cracker. September:14-15.
Richardson, D.G., 1996. The health be-nefits of eating hazelnuts: Implicationsfor blood lipid profiles, coronary heartdisease and cancer risks. Acta Horticul-turae, ISHS - 445: 295-300.
Romero, A.; Tous, J.; Plana, J.; Díaz, I.;Boatella, J.; García, J.; López, A., 1997.Commercial quality characterization ofSpanish ‘Negret’ cultivar. Acta Horticul-turae, ISHS - 445: 157-163
Ruggeri, S.; Cappelloni, M.; Gambelli,L.; Nicoli, S.; Carnovale, E., 1998. Che-mical composition and nutritive value ofnuts grown in Italy. Ital. J. Food Sci.Vol.10(3): 243-252.
Sánchez, G.; Malo, M., 1996. Reco-mendaciones nutricionales en la UniónEuropea. Política Científica, (45): 51-55.
Serra, J.; Ventura, F. 1995. Study of thecarbohydrate fraction of the principalvarieties of Tarragona hazelnuts (Cory-lus avellana L.). Food Chemistry. 46:285-288.
WEBSITES REFERENCES
Agricultural Research Servicehttp://www.ars-grin.gov/cgi-bin/duke/
farmacy2.pl?314FAO International Network of Food
Data Systemshttp://www.fao.org/infoods/
Swiss Food Composition Databasehttp://food.ethz.ch/swifd/
USDA Nutrient Data Laboratoryhttp://www.nal.usda.gov/fnic/foodcomp/
A. Romero and J. TousDivisió Alimentària-Dept. d’Arboricultura
Mediterrània,IRTA - Centre de Mas Bové. Apartat 415,
43280 Reus, SpainPhone: 34 977 343252
Fax. 34 977 344055E-mail: [email protected]

28 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
their preparation provided information onmorphological characteristics of a samplecontaining 50 nuts of a specific genotype(length, width, thickness, diameter of thepericarp, nut weight, kernel weight).Technological characteristics were highcontent of the kernel in the nut and abso-lute weight, used to calculate the numberin a kilogram for the purpose of sowing inthe nursery. The depth of sowing andplanting distance were adjusted to thesize of the nuts. Raw oil, total proteinsand water make the biggest part of co-tyledons resulting in physiological proces-ses in the seed, germination and develop-ment of the bud, until the plant is capableof subsisting on the food from its ownroot. The kernel of Corylus colurna L. is asignificant nutritive component. (Ninic-To-dorovic, 1992).
Prolonged seed dormancy is caused bycharacteristics of the embryo, thicknessof the pericarp and, slightly, by inhibitoryeffect of the acids present in fleshy cupu-les. Change of the color of cupule intoyellowish-green is an indicator for pic-king. While shaking the trees, cupules felleasily due to their weight and physiologi-cally separated stems, while green cupu-les remained in the treetop. The succes-sive maturity resulted in the beginningand duration of the phenophase of flowe-ring. Climate, microclimate and other en-vironmental conditions in January, Fe-bruary and March (depending on the yearof observation of the flowering phase)had impact on pollination, formation ofthe cupule, formation of the ovary and itsorganelles, the manner of pollination, in-tensive nut development in June, develo-pment of the embryo and fruits with layersuntil the phase of physiological and tech-nological maturity.
Prolonged seed dormancy of Corylus co-lurna L. caused by insufficiently develo-ped embryo or its segments and thick-ness of the pericarp in laboratory condi-tions, was performed by stratification(4x50) of the nuts in pure river sand con-taining 27 - 30% moisture. The nuts werefirst saturated in water for 24 to 48 hoursand after that disinfected by 0.3% solu-tion of Orthocide to prevent growth of mil-dew although the phytopathological testdid not prove presence of any pathoge-nes on the pericarp. The moisture contentwas controlled continuously in cycles ofseven days during the cold stratificationperiod at the temperature of +4oC to +6oCand relative moisture of the refrigerator of95%.
The proper time for placing the seed un-der conditions needed for the beginningof germination was indicated by openingof the pericarp, in other words by ruptu-ring of the seed coat at the joint. The nuts
with ruptured pericarp were placed in dis-hes with river sand at the depth of 4 to 5cm. Technical germination, absolute ger-mination, energy of germination and ave-rage duration of germination were investi-gated in laboratory. The experimentswere carried out at the Institute of Pomi-culture, Viticulture and Horticulture andthe greenhouse of the Institute of Biology,Department of Mathematics and Sciencesin Novi Sad. Examination of the field ger-mination was based on seed samples,containing 100 nuts, sowed in three suc-cessive cycles in blocks with a randomorder of treatments and in three terms(September, October and November). Inthe later phase the field germination wasstudied concerning the total number ofnuts of a specific genotype which wassown. The experiments and production ofplanting material of Corylus colurna L.were carried out in the plant nursery ofthe Department of Agriculture in RimskiSancevi.
RESULTS AND DISCUSSIONThe largest number of the mother-trees ofCorylus colurna L. was chosen by indivi-dual selection from the areas of Novi Sadand surroundings. The mother-trees wereplanted for esthetic functions and melio-ration purposes required by an urbanarea (Ninic, J., 1981). In the course oftime they became physiologically mature,with well-developed treetops. The treesgave abundant yield, alternatively, de-pending on the year. The target was tostudy the trees thoroughly, examine thegenerative reproductive material and re-gister them in the Catalogue of SeedSources and Nurseries of Forest and Hor-ticultural Stock Production in Serbiamainly for different purposes in horticultu-re and landscape architecture. The initialcriteria for selection of mother-trees werephenotypic features, health condition, vi-tality, ornamentality, abundance of flowe-ring and yielding depending on the yearof observation.
Hazelnut orchard growing at Constantí, Spain
29FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Subselection included 45 genotypes ofCorylus colurna L. grown in the Futoskipark, Dunavski park, plant nursery of theCity Greenery of Novi Sad, park of theInstitute of Cardiovascular diseases inSremska Kamenica and park “Dvorskabasta” in Sremski Karlovci.
Optimal physiological maturity of the indi-vidual trees, quantity of the yield and re-sults of the several-year research showedthat groups of the trees from the Futoskipark in Novi Sad have priority over theothers. The examined individuals arecharacterized by straight trees of 12 to 18meters in height and by trunks withoutbranches up to the treetop at the height of1.8 m to 3.2 m. The tops are regular co-nes in shape and in older trees wide-coni-cal. The skeletal branches are arrangedalternatively. The oldest trees were plan-ted in 1910 - at the time the park was es-tablished. No significant individual diffe-rences in morphological features and the
leaf form were found within groups oftrees of the same age, besides the habi-tat taken as a whole.
In phenophases of flowering of male andfemale flowers, however, significant indi-vidual differences took place. Extremelyearly and extremely late flowering treescan be observed. The earliest floweringwas recorded on January 05, 1983 at theaverage daily temperature of 3.6o C withinthe period of 15 days and the intensity offlowering being 80% (B5). The most im-portant impact on the beginning, durationand end of flowering phenophase wasthat of daily air temperatures (minimal,maximal and average daily temperatures)and the exposure of the treetop to thesunlight. The phenophase of foliage for-mation began at the end of March and thebeginning of April, and cupule formationwas observed in April. The process of fer-tilization took place in May and the begin-ning of June followed by the period of in-
tensive nut growth which reached itsmaximal development at the end of Juneand the beginning of July.
According to Tucovic (1970), in relation tothe characteristics of cupules, the diver-gence of the Corylus colurna L. variety ismanifested as a specific form (C. ColurnaL. f. Colurna Kit., 1966) in which petiolesand parts of the nut layer are smooth orslightly fuzzy , with no gland fuzzes (treesbelonging to group C: C2, C3, C4, C5) andthe form of glandulifers (Corylus ponticaf. glandulifera Koch., 1849) in which bothpetiole and the nut layer are covered inplentiful gland fuzzes (denoted by marksof other trees presented in Table 1).
During the harvesting period, gatheringand extraction of the nuts, which takesplace in August and at the beginning ofSeptember, it was concluded that it waspossible to find cupules containing a sin-gle nut, but also cupules containing up to
Table 1. Average occurrence of nuts in cupules and morphological features (several years)
Tree Mark Number of Length of nuts Width of nuts Diameterof nuts Pericarp thickness Nut weight Kernel weightnuts in a cupule (mm) (mm) (mm) (mm) (g) (g)
A1 6 15,91 14,19 11,31 1,45 1,30 0,57B1 7 15,70 13,63 11,17 1,29 1,04 0,42B2 6 16,23 13,50 10,76 1,35 1,12 0,46B4 6 17,12 13,74 11,01 1,67 1,13 0,46B5 7 17,04 13,85 11,09 1,61 1,16 0,45B7 7 16,99 13,79 11,64 1,75 1,12 0,45B8 11 16,71 13,71 10,64 1,57 1,05 0,42B9 5 18,18 14,05 10,88 1,88 1,16 0,47B11 5 17,14 15,47 11,88 1,79 1,40 0,54C2 5 16,75 14,11 11,19 1,45 1,28 0,53C3 5 16,99 15,16 11,82 1,57 1,29 0,59C4 5 16,48 15,86 12,29 2,07 1,44 0,55C5 4 16,29 14,89 12,32 1,58 1,41 0,52
Table 2. Average values of technological and biochemical characteristics of the Corylus colurna L. nuts(in the period of several years)
Technological Biochemical characteristics of cotyledonscharacteristics (nuts prepared for storage)
TreeMark Kernel content Absolute weight No. of nuts in kg Water content Protein content Oil Content(%) (g) (%) (%) (%)
A1 83,66 1139 905 4,27 16,51 71,36B1 94,33 1070 1030 3,98 16,57 68,05B2 91,00 1033 988 3,29 14,85 78,69B4 97,33 1054 963 4,25 20,24 63,97B5 95,00 1156 867 4,22 16,49 67,44B7 87,33 1114 912 3,58 19,06 66,36B8 91,00 1085 925 4,36 17,98 73,14B9 84,50 1210 827 5,87 20,70 68,03B11 84,66 1222 825 4,06 19,64 69,00C2 86,33 1058 983 4,22 18,99 65,57C3 88,66 1115 899 4,94 18,87 69,36C4 92,50 1441 698 4,07 19,83 67,65C5 86,66 1404 717 4,50 21,08 65,90
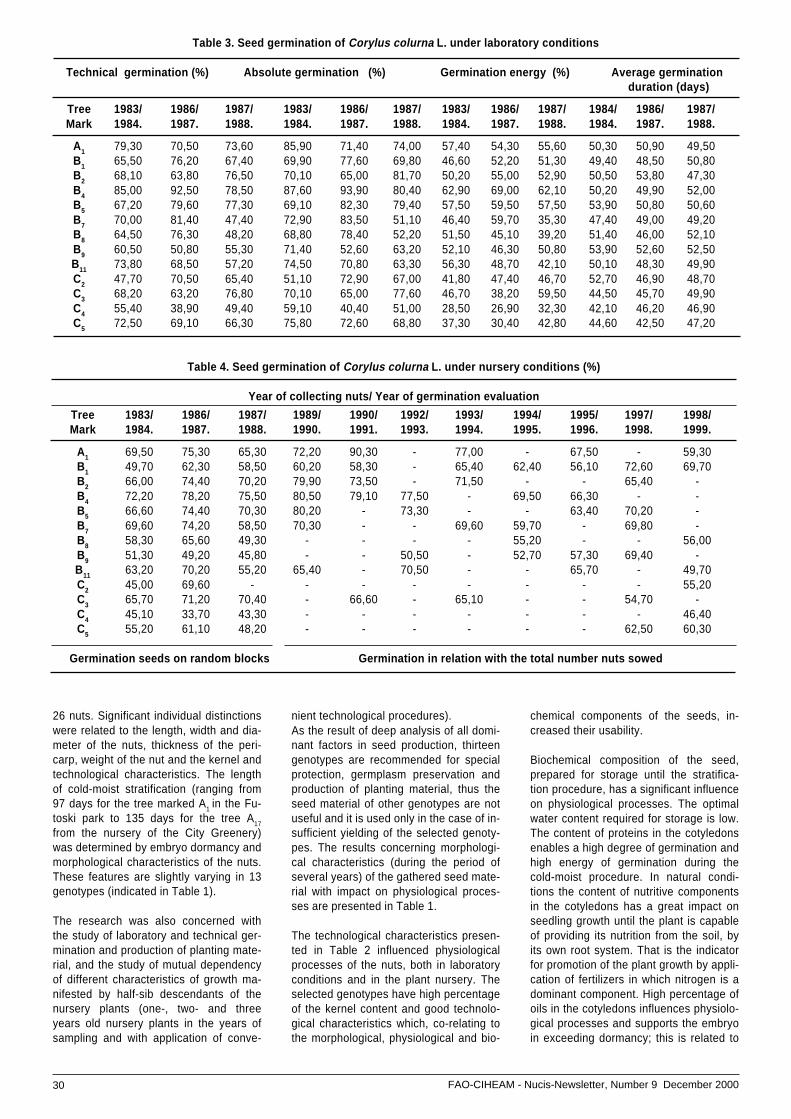
30 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
26 nuts. Significant individual distinctionswere related to the length, width and dia-meter of the nuts, thickness of the peri-carp, weight of the nut and the kernel andtechnological characteristics. The lengthof cold-moist stratification (ranging from97 days for the tree marked A1 in the Fu-toski park to 135 days for the tree A17
from the nursery of the City Greenery)was determined by embryo dormancy andmorphological characteristics of the nuts.These features are slightly varying in 13genotypes (indicated in Table 1).
The research was also concerned withthe study of laboratory and technical ger-mination and production of planting mate-rial, and the study of mutual dependencyof different characteristics of growth ma-nifested by half-sib descendants of thenursery plants (one-, two- and threeyears old nursery plants in the years ofsampling and with application of conve-
nient technological procedures).As the result of deep analysis of all domi-nant factors in seed production, thirteengenotypes are recommended for specialprotection, germplasm preservation andproduction of planting material, thus theseed material of other genotypes are notuseful and it is used only in the case of in-sufficient yielding of the selected genoty-pes. The results concerning morphologi-cal characteristics (during the period ofseveral years) of the gathered seed mate-rial with impact on physiological proces-ses are presented in Table 1.
The technological characteristics presen-ted in Table 2 influenced physiologicalprocesses of the nuts, both in laboratoryconditions and in the plant nursery. Theselected genotypes have high percentageof the kernel content and good technolo-gical characteristics which, co-relating tothe morphological, physiological and bio-
chemical components of the seeds, in-creased their usability.
Biochemical composition of the seed,prepared for storage until the stratifica-tion procedure, has a significant influenceon physiological processes. The optimalwater content required for storage is low.The content of proteins in the cotyledonsenables a high degree of germination andhigh energy of germination during thecold-moist procedure. In natural condi-tions the content of nutritive componentsin the cotyledons has a great impact onseedling growth until the plant is capableof providing its nutrition from the soil, byits own root system. That is the indicatorfor promotion of the plant growth by appli-cation of fertilizers in which nitrogen is adominant component. High percentage ofoils in the cotyledons influences physiolo-gical processes and supports the embryoin exceeding dormancy; this is related to
Table 3. Seed germination of Corylus colurna L. under laboratory conditions
Technical germination (%) Absolute germination (%) Germination energy (%) Average germination duration (days)
Tree 1983/ 1986/ 1987/ 1983/ 1986/ 1987/ 1983/ 1986/ 1987/ 1984/ 1986/ 1987/Mark 1984. 1987. 1988. 1984. 1987. 1988. 1984. 1987. 1988. 1984. 1987. 1988.
A1 79,30 70,50 73,60 85,90 71,40 74,00 57,40 54,30 55,60 50,30 50,90 49,50B1 65,50 76,20 67,40 69,90 77,60 69,80 46,60 52,20 51,30 49,40 48,50 50,80B2 68,10 63,80 76,50 70,10 65,00 81,70 50,20 55,00 52,90 50,50 53,80 47,30B4 85,00 92,50 78,50 87,60 93,90 80,40 62,90 69,00 62,10 50,20 49,90 52,00B5 67,20 79,60 77,30 69,10 82,30 79,40 57,50 59,50 57,50 53,90 50,80 50,60B7 70,00 81,40 47,40 72,90 83,50 51,10 46,40 59,70 35,30 47,40 49,00 49,20B8 64,50 76,30 48,20 68,80 78,40 52,20 51,50 45,10 39,20 51,40 46,00 52,10B9 60,50 50,80 55,30 71,40 52,60 63,20 52,10 46,30 50,80 53,90 52,60 52,50B11 73,80 68,50 57,20 74,50 70,80 63,30 56,30 48,70 42,10 50,10 48,30 49,90C2 47,70 70,50 65,40 51,10 72,90 67,00 41,80 47,40 46,70 52,70 46,90 48,70C3 68,20 63,20 76,80 70,10 65,00 77,60 46,70 38,20 59,50 44,50 45,70 49,90C4 55,40 38,90 49,40 59,10 40,40 51,00 28,50 26,90 32,30 42,10 46,20 46,90C5 72,50 69,10 66,30 75,80 72,60 68,80 37,30 30,40 42,80 44,60 42,50 47,20
Table 4. Seed germination of Corylus colurna L. under nursery conditions (%)
Year of collecting nuts/ Year of germination evaluation
Tree 1983/ 1986/ 1987/ 1989/ 1990/ 1992/ 1993/ 1994/ 1995/ 1997/ 1998/Mark 1984. 1987. 1988. 1990. 1991. 1993. 1994. 1995. 1996. 1998. 1999.
A1 69,50 75,30 65,30 72,20 90,30 - 77,00 - 67,50 - 59,30B1 49,70 62,30 58,50 60,20 58,30 - 65,40 62,40 56,10 72,60 69,70B2 66,00 74,40 70,20 79,90 73,50 - 71,50 - - 65,40 -B4 72,20 78,20 75,50 80,50 79,10 77,50 - 69,50 66,30 - -B5 66,60 74,40 70,30 80,20 - 73,30 - - 63,40 70,20 -B7 69,60 74,20 58,50 70,30 - - 69,60 59,70 - 69,80 -B8 58,30 65,60 49,30 - - - - 55,20 - - 56,00B9 51,30 49,20 45,80 - - 50,50 - 52,70 57,30 69,40 -B11 63,20 70,20 55,20 65,40 - 70,50 - - 65,70 - 49,70C2 45,00 69,60 - - - - - - - - 55,20C3 65,70 71,20 70,40 - 66,60 - 65,10 - - 54,70 -C4 45,10 33,70 43,30 - - - - - - - 46,40C5 55,20 61,10 48,20 - - - - - - 62,50 60,30
Germination seeds on random blocks Germination in relation with the total number nuts sowed

31FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
the cold-moist treatment in laboratoryconditions or sowing the nuts in the plantnursery in the October term.Laboratory germination was examined inthe period lasting 70 days, the first recor-ding being made on the 7th day and thelast on the 70th day. The energy of germi-nation was determined on the 21st day.The Regulations of International SeedTesting Association (ISTA), concerninggermination of Corylus sp., require thefirst counting of the seeds germinated onthe 28th day and the last on the 70th day.The Yugoslav Regulations on determina-tion of germination of Corylus sp. Seedssuggest the first counting on the 7th andthe last on the 28th day from transporta-tion of the seeds in favourable conditionsfor germination taking into considerationthe fact that both parts of pericarp shouldbe removed. During the examination ofgermination, the pericarp of the nuts wasnot removed in this examination. After thegermination period, we determined thenumber of fresh non-germinated seeds(treated by solution of three-phenyl – te-trasolium chloride), empty and putridseeds, as well.
The results of examined laboratory germi-nation of Corylus colurna L. seeds showthat the best physiological and reproduc-tive characteristics were found in seedmaterial of the trees indicated in Table 3.The average value of technical germina-tion of the genotype B4 was 85.00 %, ab-solute germination was 87.60 %, germi-nation energy 62.90 % and the averageduration of germination 52.00 days. Theresults concerning the seed quality of thetrees marked B5, A1, B2, C3, B7, B11 and theothers are presented in the table. Absolu-te seed germination does not vary fromthe results achieved for technical germi-nation in laboratory conditions. Some ofthe dominant factors were content of thekernel and germination speed of a heal-thy seed. Germination energy is extreme-ly high, which is important for simulta-neous appearance of the seedling andapplication of suitable technological pro-cedures. A seed with low germinationspeed has a low value of the average du-ration of germination, which, relating toexamined characteristics of Corylus co-lurna L. nuts, determines the quality andits usability. The percentage of fresh non-germinated seeds with completely deve-loped embryo is the result of extreme dor-mancy of the embryo and characteristicsand permeability of the pericarp.
Optimal results under nursery conditionswere achieved by sowing of the nuts inOctober. Field germination (presented inTable 4) stands in correlation with the re-sults of technical germination achieved inlaboratory conditions. In natural condi-tions the opening of the pericarp takes
place in March, and the seedling appearsin May. The table presents the averagevalues of the field germination recordedin May (after the appearance of seedling),in June (during the developments of theplants) and in July (when the stem beco-mes woody). The impact of the environ-ment and manual and mechanical treat-ments also have a great influence on theresults of the field germination in a plantnursery.
CONCLUSIONThe results of the study of the most im-portant factors concerning production ofCorylus colurna L. seeds, which includedlaboratory examination and a parallelexamination in a nursery, showed a gene-tic divergence in the used seed material.Comparative observation of biologicalfeatures and examination of morphologi-cal, technological, physiological and nu-tritive values and preservation of nut ger-mination, when stored according to diffe-rent methods drew guidelines of techno-logical procedures to achieve high pro-ductivity and effectiveness of productionof seedling material of Turkish filbert. Inthe nursery of Rimski Sancevi, this spe-cies is used as a rootstock for grafting ofconventional hazel cultivars. Graftingtechnology, applying methods of mergingand tongue grafting (English grafting),was improved. The rootstocks are two,three or four years old seedlings of Tur-kish filbert grafted in situ. The purpose isto produce tree-like plants for orchards,where they would allow a full applicationof machinery, and for planting aroundhouses. Five and six years old nurserytrees can be used in horticulture andlandscape architecture, in forestry andmelioration of erosive areas and planta-tions intended for exploitation of this va-luable tree.Turkish filbert is characterized by a widerange of application possibilities, veryhigh resistance to diseases and pests, itis proposed that production of the nurseryplants should be increased and directedto forest ecosystems of Yugoslavia andalso to other areas with suitable ecologi-cal conditions.
BIBLIOGRAPHY
Hong, T. D. and Ellis, R. H., 1996. Aprotocol to determine seed storage be-haviour. IPGRI Technical Bull. No 1.Department of Agriculture, The Univer-sity of Reading, UK.
Jankovic, M. , Pantic, N., Mi√ic, V.,Diklic, N., Gajic, M., 1984. VegetacijaSR Srbije. SANU, Beograd.
Korac, M. Slovic, D., 1973/1974. Uticajnekih faktora na uspeh kalemljenja les-
ke. Letopis naucnih radova, Sveska 17-18. Poljoprivredni fakultet, Novi Sad.
Lagerstedt, H. B., 1971. Filbert treeGrafting. Ann. Report Oregon. St. Hort.So. 62:60-63.
Lagerstedt, H. B., 1975. Filberts PurdueUniversity Press. West Lafayette. Indiana
Milevic, K., 1951. Mecja leska (Coryluscolurna L.). sumarstvo br 6. Beograd.
Mi√ic, V., 1966. Vegetacija Djerdapskogpodrucja. Za√tita prirode br. 33. Zbornikradova Republickog zavoda za za√tituprirode SR Srbije,. Beograd.
Ninic, J., 1981. Funkcionalnost zelenihpovr√ina Novog Sada. Magistarski rad.sumarski fakultet. Beograd
Ninic-Todorovic, J., 1988. Determina-tion of the most suitable methods ofhandling Corylus colurna L. fruit. Actahort. 226, Wageningen
Ninic-Todorovic, J., 1990. A study of do-minant factors and determination of opti-mal technological methods for productionof high quality nursery plants of Turkishfilbert (Corylus colurna L.). Doctoral the-sis. Faculty of Forestry. Belgrade
Ninic-Todorovic, J. Biochemical compo-sition of seed in Turkish filbert (Coryluscolurna L.). Journal of Yugoslav Pomo-logy. Vol. 26 (99-100): 23-30, Cacak.
Ninic-Todorovic, J., 1998. Preservationof biological diversity of Turkish filbert(Corylus colurna L.). Nature protectionNo 50. P. 99-105. Belgrade.
Prljincevic, M., Mancic, A., 1997. Cata-logue of seed sources and nurseries offorests and horticultural stock produc-tion in Serbia. Ministry of Agriculture,Forestry and Water power engineeringof Serbia. Belgrade
Stilinovic, S. Tucovic, A., 1977. Prilograzmatranju kriterijuma za izdvajanjehortikulturnih semenskih objekata.Glasnik sumarskog fakulteta. Serija C.Univerzitet u Beogradu.
Stilinovic, S., 1985. Semenarstvo √ums-kog i ukrasnog drveca i zbunja. Univer-zitet u Beogradu.
Stilinovic, S., 1987. Proizvodnja sadnogmaterijala √umskog i ukrasnog drveca izbunja. Univerzitet u Beogradu.
Stojanovic, Lj., Jovanovic, B., 1987.Ceno-ekolo√ke i uzgojne karakteristikesastojine bukve i jele sa plemenitimli√carima u zapadnoj Srbiji. sumarstvo br.3-4, Beograd.
Tucovic, A., 1970. Familija Corylaceae.Flora SR Srbije, tom II. SANU, Beograd
Velickovic, M., 1959. Nalazi√ta mecjeleske (Corylus colurna L.) na podrucju√umskog gazdinstva Vranje. sumarstvobr. 11/12. Beograd
J. Ninic-TodorovicDepartment of Agriculture
University of Novi SadYugoslavia

32 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
WALNUT SITUATION IN IRAN
INTRODUCTIONWalnut is one of the Iranian traditional nutcrops, found in 29-39° latitude and 45-64°longitudes, from lowlands up to 2500 me-ters height. Dominant walnut species isJuglans regia. Wild and natural sites ofthis species are scattered in north, westand centre of the country. Many scientistsknow Iran as the origin of J. regia and callit Persian walnut. J. regia is native to wet-ter areas in the mountains of Iran. Walnutstands are found in the Kopet DaghMountains, the low land along thesouthern shore of Caspian Sea and insporadic locations throughout the rangesof western and southern Iran. The KopetDagh Mountains along the northern bor-der of Iran are quite dry, especially on thenorthern slope and do not support fo-rests. Native J. regia is found here only ina few of the wetter gorges and deepvalleys.
Walnuts are grown to a great extent onElburz, Zagros, Lalezar and Jabal Barezmountain feet (Fig 1). Most of these treesare seedlings and show a wide variationin their characteristics. Therefore it canbe easily found precociously, lateral bea-ring, big kernel and other valuable cha-racters in these genotypes. In some dis-tricts, plantings expanded along the ri-vers, around the orchards or mixed withother fruit trees. There are many old andhuge trees of this species in Iran. Theage of the oldest is estimated to be about1400 years. This tree is situated in “Ore-gan”, a village of Shahre Kord city. Thereare also many other old walnut trees inKhansar, Isfahan, Qazvin, Taleghan andTooiserkan regions with the age rangingfrom 300 to 700 years (Fig 1). Thesetrees produce about 50-100 thousandnuts each. Unfortunately, many of thesetrees have been destroyed by English,Italian and Russian timber buyers aftersecond world war (1951-1961).
In recent years, with the government en-couragement, identification of superiorgenotypes and establishing grafted or-chards using these genotypes have beenstarted. In addition, walnut price incre-ment in domestic markets has promotedgrowers to establish new walnut or-chards. Thus, planted area of non-bea-ring orchards increased to five folds du-ring last decade (Fig 2). At the presenttime, number of walnut growers is estima-ted around 30,000.
Islamic Republic of Iran occupies an im-portant niche among the world’s leadingproducers. With its 45 thousand hectaresof walnut crops which produce an avera-ge of 145 thousand MT annually, or 10%
of world production, Iran ranks thirdamong leading producers (Tab 1). Strongdemand and good price of walnut in do-mestic markets has decreased tends toexport this crop. Walnut kernel is a valua-ble snack in Iran, sometimes eaten withcheese for breakfast. It is also a suitableingredient for sweets. This is a very va-luable tree for Iranian growers because ofits nuts as well as its timber.
TECHNICAL INFORMATIONDistributionAll over Iran but particularly in northern,western and central district on mountainfeet of Alborz, Zagros, Lalezar and JabalBarez in Kerman, Hamedan, Qazvin,Azarbaijan, Kermanshah, Khorasan andFars provinces (Fig 3).
CultivarsThere are important populations of Per-sian walnut in Iran and most of them areseedlings. These populations haveshown relatively stable characteristics forlong time. The biggest walnut populationsare situated in Tooiserkan, Kerman, Qa-zvin and Shahmirzad district. These po-pulations have not been much studied.Many of these trees are very old, indicati-ve of resistance to much stress duringhundreds of years.
GermplasmIn northern Iran, J. regia is found in theCaspian lowlands and Elburz Mountainregion of Mazandaran, Gorgan and Gillanprovinces at elevations of 60-2400 m.Collections are noted here from the Ma-zandaran Mountains, the Haraz valleyand the mountains around Gorgan. Thesewetter mountain regions of Iran exhibit atypical holarctic deciduous forest. Also,there are scattered J. regia collections inthe mountain ranges extending from eas-tern Turkey to southern Pakistan throughthe Iranian provinces of Azarbaijan-eBakhtari, Kordestan, Kermanshah, Lores-tan, Bakhtiari, Kerman and Khorasan. Inthe south Caspian lowlands, north face of
Elburz Mountains, the mountains of Kho-rasan and Zagros ranges densely fores-ted areas have been severely degradedand the natural vegetation has been redu-ced to mere remnants as a result of agri-cultural activity and cutting.
Orchard featuresWalnut trees traditionally planted alongthe rivers, around the orchards, interplantwith the other fruits or exclusively at dis-tances of 10-15 m. In new specialised or-chards, 9-10 x 10-11 m spaces are used.
PropagationMainly by seed. Recently, grafting usedfor propagation of superior genotypes.Some research projects on micro-propa-gation are underway.
Training and pruning systemsCommon training system is modified cen-tral leader. Pruning is not common in tra-ditional orchards and some growers be-lieve that pruning is not necessary forwalnut trees, but in new orchards pruningis done.
FertilizationScarcely used. In specialised orchards,manure (40-60 t/ha), 400 kg/ha KCl and250 kg/ha P2O5 used at planting and 100g of N unit for each year of the age of thetree is necessary.
IrrigationWalnut trees require about 10,000 to12,000 m3/ha during growing season. Innorth of Iran, near the Caspian Sea, inwhich there is more than 700 mm raining,irrigation is not necessary but in otherparts of Iran orchards are irrigated every10-15 days during summer. The irrigationsystem in traditional orchards is furrow orbasin but in new plantings there is dripirrigation.
Diseases and pestsIn spite of rarely application of pesticidesagainst pests and diseases, most of wal-
Table 1. Walnut planted area and production in Iran
Years Planted area (ha) Production (t)Fruit bearing Non-fruit bearing
1988 11644 5196 378211989 13243 5395 460091990 15025 9027 444821991 17973 12067 644861992 22696 10089 712371993 25976 12460 1108191994 28975 14563 1172181995 32290 14978 1192181996 35783 17889 1131871997 41766 20837 1248721998 45152 27373 1458201999 50161 32698 142906

33FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
nut trees are healthy in appearance.Blight and anthracnose are observed insome regions with the high rainfall duringspring and summer, especially around theCaspian Sea. Black line disease has notbeen reported yet, maybe because allwalnut trees are grafted on J. regia roots-tock. Aphids, codling moth, bark and treeborers, scales and mites are seen insome districts. Birds, especially crow,attack the nuts during harvest. Its dama-ge is more severe in orchards, which har-vesting is delayed.
Physiological disordersWalnut kernel browning is a major pro-blem some years when summer is dryand hot. Some of the growers try to de-crease this disorder by making holes onthe tree trunk using an axe or long nail.They claim that walnut bleeding is effecti-ve on kernel colour.
HarvestFrom 5-10 September to mid October de-pending on variety and climate. Becauseof the variation of walnut seedlings, har-vesting is manually done.
Post-harvest systemsHulling is done traditionally by hand. Re-cently some hullers and dryers have beenimported or were made by local experts.
Import and exportAs far as we know, there is no import ofwalnut. Export of in-shell dried walnut isnot so much due to high demand of do-mestic markets. Top export destinationsfor Iranian walnut kernel are: the UnitedArab Emirates, Germany, Turkey, SaudiArabia, Qatar, Liban, Kuwait, Ukraine,Sweden and Syria.
Research
Main institution AreaMinistry of AgricultureAgricultural ResearchCentrein Karaj, Shahrood,Mashhad, Ormieh,Azar Shahr
Departmentof HorticultureFaculty of AgricultureUniversity of TehranKaraj
Prospective1. Identification and evaluation of superiorgenotypes using IPGRI walnut descriptorsinto the rich walnut populations of Iran.
2. Estimating mathematical models be-tween qualitative and quantitative charac-ters of walnut trees and environmentalconditions of their growing district.
3. Improvement of grafting methods forasexual propagation of superior genoty-pes and releasing new cultivars.
4. Research on other propagation me-thods (ex: micropropagation, stooling,apomixis, etc.).
5. Establishing modern orchards usingsuperior cultivars.
6. Mechanisation of harvest and post-harvest systems.
ACKNOWLEDGEMENTThe author wishes to acknowledge Dr. EricGermain, Director of UREFV, INRA for hiskindness in scientific editing of this paper.
REFERENCES
Agricultural Statistics and InformationDepartment, 1999. Dried fruits, figuresand views. Ministry of Agriculture. Iran.
ASID, 2000. Agricultural statistics year-book. Ministry of Agriculture. Iran.
Atefi J., 1990. Preliminary research onPersian walnut and correlation betweenpair characters. 1st Intl. Symp. on Wal-nut Production, 25-29 Sep. 1989, Buda-pest (Hungary). Acta Hort. 284: 97-104.
Atefi J., 1997. Study on phonologicaland pomological characters on walnutpromising clones in Iran. 3rd Intl. WalnutCongress, 13-16 June 1995, Alcobaça(Portugal). Acta Hort. 442: 101-108.
Dehlavi A., 1994. Walnut growing.Baghdar. No. 1: 40-42.
Farshi AA, 1997. An estimate of waterrequirement of main field crops and or-chards in Iran. Soil and Water ResearchInstitute. Ministry of Agriculture. Iran.
Germain E., 1993. The Persian walnutin Iran. Nucis. No. 1: 5 – 6.
Grassi G., 1998. Walnut. In: Grassi etal. (eds.) Italian horticulture. SocietaOrticola Italiana. Italy.
INC, 2000. World consumption and pro-duction trends. The cracker. No. 1: 33-49.
Leslie Ch. and McGranahan G., 1988.Native populations of Juglans regia –adraft, Intl. Conference on Walnuts.Sep.19-23. Yalova, Turkey.
Pourbabaee H. et al., 1999. Diversity ofwoody species of common walnut (Ju-glans regia L.) sites in Guilan forests.Iranian Journal of Natural Resources.52 (1): 35-45.
Visan consulting engineers, 1995. Stu-dy of Iran horticulture project report.Vol. 7. Ministry of Agriculture. Iran.
K. VahdatiDepartment of Horticulture, Faculty ofAgriculture
University of Tehran - Karaj-IranEmail: [email protected]
Tel: ++98 21 7811963 - Fax: ++98 21 8078611
Genotypeevaluation,germplasm,breeding,irrigation,grafting,apomixis
Tissueculture,stooling,grafting
HEDGEROW WALNUTPLANTING SYSTEM:
EXPERIENCES IN SPAIN
INTRODUCTIONWalnut orchards using grafted trees star-ted to be planted in Spain around 1976when Californian and French cultivarswere introduced. Walnut orchard mana-gement was unknown and technologiesapplied in California and France were as-sessed.
At present the surface of grafted walnutorchards is around 3500 ha, and the Spa-nish walnut production from regular plan-tations is about 4000 t, considering thatmost orchards are still young. Main culti-vars in adult orchards are: ‘Franquette’, inareas having late spring frosts, and ‘Hart-ley’ and ‘Serr’ in warm areas. Since 1990the most favourite cultivar is ‘Chandler’.
So far Spanish technology to producewalnuts has been developed and or-chards are well managed. Pests and di-seases are detected and treated in time.Xanthomonas arboricola pv. juglandis(Blight) and Cydia pomonella are themain phytosanitary affections. In someplaces, spray orchard programme shouldinclude Gnomonia leptostyla control.
The Extensive and semi-intensive planta-tions are still the most common plantingsystems in Spain although some farmersstarted planting cultivars like‘Chico’,‘Vina’ or ‘Chandler’ in hedgerow.Experimental hedgerow orchards are be-ing controlled by IRTA (Figure 1), someuseful results of these trials are reportedin this paper. Mechanical pruning is alsoevaluated.
Description of IRTA's experimentalwalnut hedgerow plotIn 1988, an intensive walnut orchard wasplanted in the Northeast of Spain, at Bí-tem, near Tortosa, Tarragona, as a hed-gerow. Two cultivars were chosen, ‘Chi-co’ and ‘Vina’ both had shown a high andprecocious productivity in Mediterraneanenvironment (Aletà and Ninot, 1993). Twoplanting distances were considered 7 m x5 m and 7 m x 3.5 m, trees were trainedin central leader but their primary bran-ches were selected on the row direction.Two assays were established, the first tocompare the cited planting distancesusing ’Chico’ and the second to comparecultivar behaviour of ‘Chico’ and ‘Vina’ at7 m x 5 m. Two pollinators were also in-cluded ‘Serr’ and ‘Amigo’ in parallel rowsto those of main cultivars. Both trials wereplanned as complete blocks with 3 repli-cations. The number of observations pertreatment varied from 5, at 7 m x 5 m, to8, at 7 m x 3.5 m. Experimental plots are
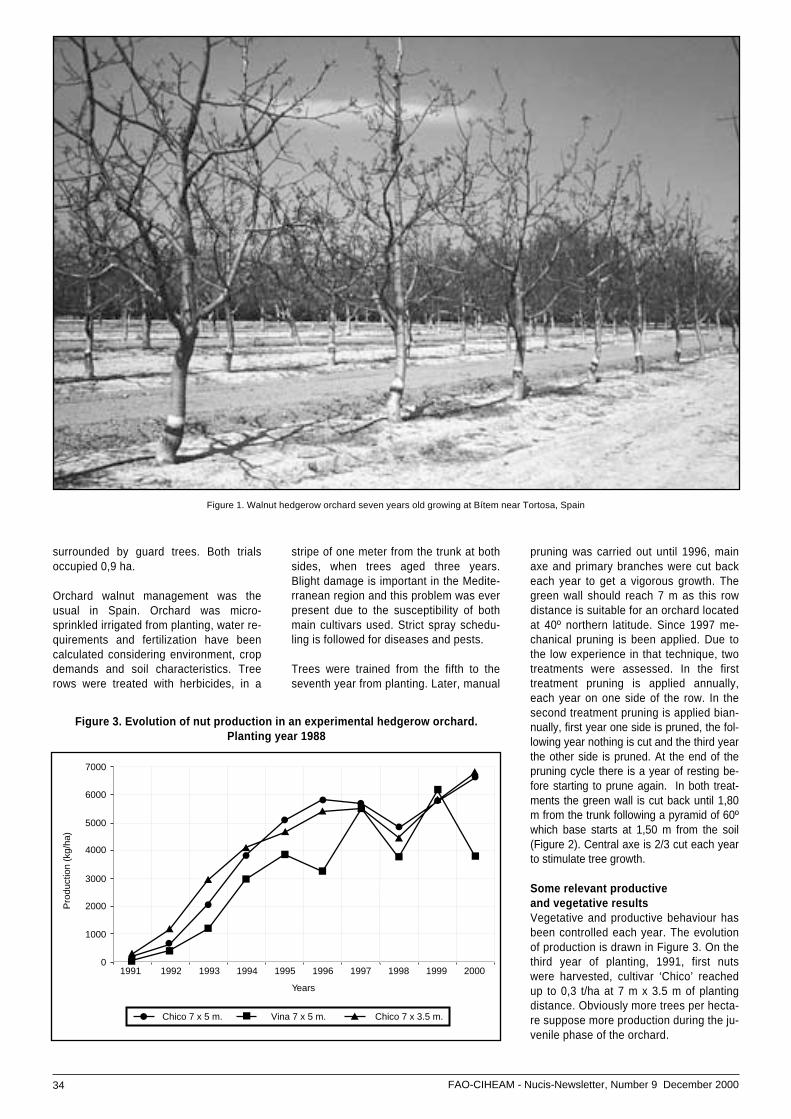
34 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
surrounded by guard trees. Both trialsoccupied 0,9 ha.
Orchard walnut management was theusual in Spain. Orchard was micro-sprinkled irrigated from planting, water re-quirements and fertilization have beencalculated considering environment, cropdemands and soil characteristics. Treerows were treated with herbicides, in a
stripe of one meter from the trunk at bothsides, when trees aged three years.Blight damage is important in the Medite-rranean region and this problem was everpresent due to the susceptibility of bothmain cultivars used. Strict spray schedu-ling is followed for diseases and pests.
Trees were trained from the fifth to theseventh year from planting. Later, manual
pruning was carried out until 1996, mainaxe and primary branches were cut backeach year to get a vigorous growth. Thegreen wall should reach 7 m as this rowdistance is suitable for an orchard locatedat 40º northern latitude. Since 1997 me-chanical pruning is been applied. Due tothe low experience in that technique, twotreatments were assessed. In the firsttreatment pruning is applied annually,each year on one side of the row. In thesecond treatment pruning is applied bian-nually, first year one side is pruned, the fol-lowing year nothing is cut and the third yearthe other side is pruned. At the end of thepruning cycle there is a year of resting be-fore starting to prune again. In both treat-ments the green wall is cut back until 1,80m from the trunk following a pyramid of 60ºwhich base starts at 1,50 m from the soil(Figure 2). Central axe is 2/3 cut each yearto stimulate tree growth.
Some relevant productiveand vegetative resultsVegetative and productive behaviour hasbeen controlled each year. The evolutionof production is drawn in Figure 3. On thethird year of planting, 1991, first nutswere harvested, cultivar ‘Chico’ reachedup to 0,3 t/ha at 7 m x 3.5 m of plantingdistance. Obviously more trees per hecta-re suppose more production during the ju-venile phase of the orchard.
Figure 1. Walnut hedgerow orchard seven years old growing at Bítem near Tortosa, Spain
1991 1992 1993 1994 1995 1996 1997 1998 1999 20000
1000
2000
3000
4000
5000
6000
7000
Pro
duct
ion
(kg/
ha)
Years
Chico 7 x 5 m. Vina 7 x 5 m. Chico 7 x 3.5 m.
Figure 3. Evolution of nut production in an experimental hedgerow orchard.Planting year 1988
35FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
‘Chico’, planted at 7 m x 5 m, was alwayssignificantly more productive than ‘Chico’at 7 m x 3.5 m after 1995. In Figure 4atrunk section evolution could give a res-ponse to those productive results. Vege-tative competition seems to have startedin 1993, trunk section growth was signifi-cantly less at low planting distance. Diffe-rences in growing affected nut productionof the following harvest, as it was fore-seen. The 1994 harvest was similar be-tween both planting spaces but the follo-wing year and later the largest tree spa-cing was always performing as the besttreatment. Trunk section in ‘Vina’ is alsosignificantly less than ‘Chico after 1995. Iftree height is considered, differences be-tween planting distances are present onlyduring first years, later indiscriminate pru-ning makes trees more vigorous (Figure4b). ‘Vina’ responds less to pruning than‘Chico’, ‘Vina’ is more difficult to managein hedgerow system than ‘Chico’.
In juvenile phase nut size, measured asthe mean weight per nut, starts to be hig-her than expected in all cultivars. Whenorchard production increases it tends togo down. This general rule should be par-ticularly considered in intensive orchardswhere nut production increases very fast.In Figures 5 we show that cultivar ‘Chico’spectacularly reduced its nut size in1997. This year corresponds to the se-cond consecutive year in which the pro-duction gets up to 5 t/ha. Mechanical pru-ning applied in February 1998 stabilisedproduction and even increased it whilenut weight showed to be difficult to in-crease. However it seems that equili-brium around 11g nut weight could bereached. Nut size of ‘Vina’ is describedas bigger than ‘Chico’ (Aletà et al., 1993;Hendricks et al., 1997) but its high sus-ceptibility to stress production does not
Years
1993 1994 1995 1996 1997 1998 1999
0
50
100
150
200
250
300
350
Sec
tion
(cm
2 )
Trunk section
Chico 7 x 5 m. Vina 7 x 5 m. Chico 7 x 3.5 m.
Years
1993 1994 1995 1996 1997 1998 1999
0
100
300
400
500
600
700
900
Hei
ght (
cm)
Tree height
Chico 7 x 5 m. Vina 7 x 5 m. Chico 7 x 3.5 m.
800
200
Figure 4a and 4b. Evolution of vegetative growth, trunk section and tree height, in a hedgerow experimental orchard.Planting year 1988.
Figure 2. Walnut orchard eleven years old. Pyramidal hedged trees at Bitem near Tortosa, Spain
Figure 4a Figure 4b
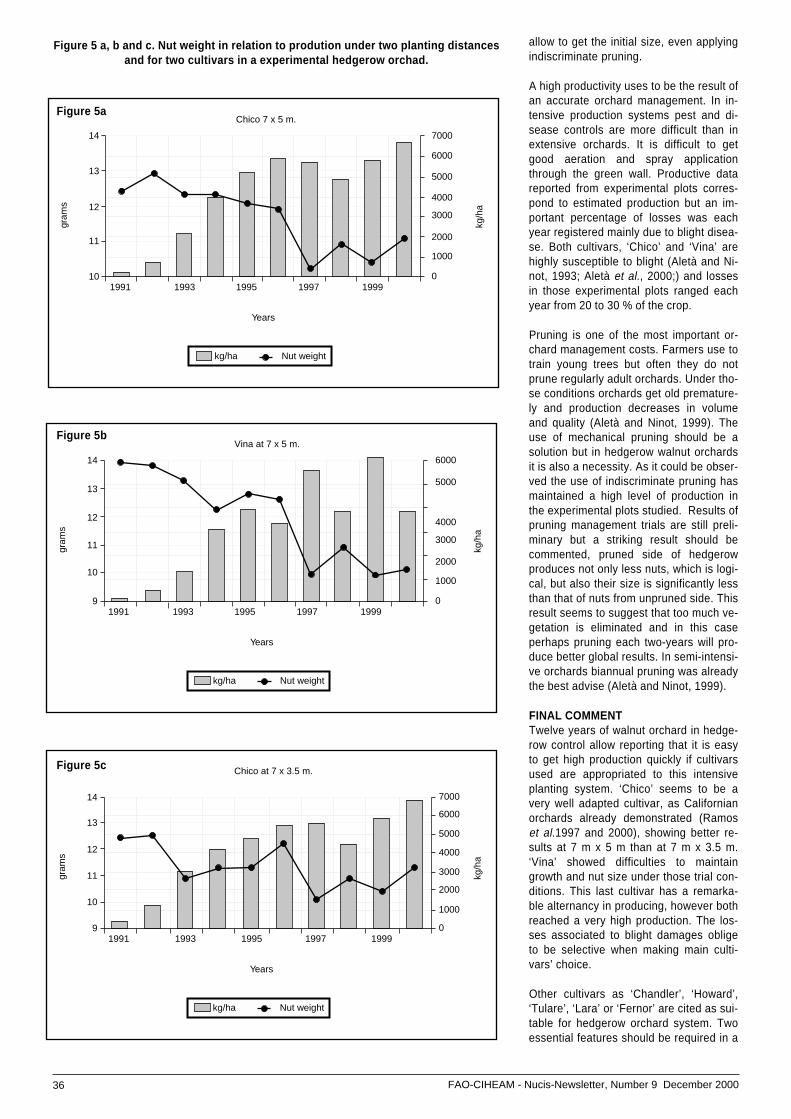
36 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
allow to get the initial size, even applyingindiscriminate pruning.
A high productivity uses to be the result ofan accurate orchard management. In in-tensive production systems pest and di-sease controls are more difficult than inextensive orchards. It is difficult to getgood aeration and spray applicationthrough the green wall. Productive datareported from experimental plots corres-pond to estimated production but an im-portant percentage of losses was eachyear registered mainly due to blight disea-se. Both cultivars, ‘Chico’ and ‘Vina’ arehighly susceptible to blight (Aletà and Ni-not, 1993; Aletà et al., 2000;) and lossesin those experimental plots ranged eachyear from 20 to 30 % of the crop.
Pruning is one of the most important or-chard management costs. Farmers use totrain young trees but often they do notprune regularly adult orchards. Under tho-se conditions orchards get old premature-ly and production decreases in volumeand quality (Aletà and Ninot, 1999). Theuse of mechanical pruning should be asolution but in hedgerow walnut orchardsit is also a necessity. As it could be obser-ved the use of indiscriminate pruning hasmaintained a high level of production inthe experimental plots studied. Results ofpruning management trials are still preli-minary but a striking result should becommented, pruned side of hedgerowproduces not only less nuts, which is logi-cal, but also their size is significantly lessthan that of nuts from unpruned side. Thisresult seems to suggest that too much ve-getation is eliminated and in this caseperhaps pruning each two-years will pro-duce better global results. In semi-intensi-ve orchards biannual pruning was alreadythe best advise (Aletà and Ninot, 1999).
FINAL COMMENTTwelve years of walnut orchard in hedge-row control allow reporting that it is easyto get high production quickly if cultivarsused are appropriated to this intensiveplanting system. ‘Chico’ seems to be avery well adapted cultivar, as Californianorchards already demonstrated (Ramoset al.1997 and 2000), showing better re-sults at 7 m x 5 m than at 7 m x 3.5 m.‘Vina’ showed difficulties to maintaingrowth and nut size under those trial con-ditions. This last cultivar has a remarka-ble alternancy in producing, however bothreached a very high production. The los-ses associated to blight damages obligeto be selective when making main culti-vars’ choice.
Other cultivars as ‘Chandler’, ‘Howard’,‘Tulare’, ‘Lara’ or ‘Fernor’ are cited as sui-table for hedgerow orchard system. Twoessential features should be required in a
Figure 5 a, b and c. Nut weight in relation to prodution under two planting distancesand for two cultivars in a experimental hedgerow orchad.
1997 1999199519931991
Chico at 7 x 3.5 m.
14
13
12
10
9
7000
6000
5000
4000
3000
2000
1000
0
Years
gram
s
kg/ha Nut weight
kg/h
a
11
1997 1999199519931991
Chico 7 x 5 m.
14
13
12
11
10
7000
6000
5000
4000
3000
2000
1000
0
Years
gram
s
kg/ha Nut weight
kg/h
a
1997 1999199519931991
Vina at 7 x 5 m.
14
13
12
10
9
6000
5000
4000
3000
2000
1000
0
Years
gram
s
kg/ha Nut weight
kg/h
a
11
Figure 5a
Figure 5b
Figure 5c

37FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
cultivar to be hedged: high lateral bearingand low to medium vigour. It will be alsoadvisable to use cultivars easy to train(Germain et al., 1999; Ramos et al.,2000).
REFERENCES
Aletà, N.; Ninot, A. 1993. Variedades denogal. Fruticultura Profesional 54: 93-100.
Aletà, N.; Ninot, A. 1999. La poda en elnogal. Fruticultura Profesional 104. Es-pecial Frutos Secos II: 85-91
Aletà, N.; Ninot, A.; Moragrega, C.; Llo-rente, I.; Montesinos, E. 2000. ActaHorticulturae 544: 353-362.
Germain, E.; Prunet, J.P.; Garcin, A.1999. Le noyer. Ed. CTIFL. 279 pp.
Hendricks, L.C; Coates, W.W.; Elkins,R.B.; McGranahan, G.H.; Philips, H.A.;Ramos, D.E.; Reil, W.O.; Snyder, R.G.1997. Selection of varieties. In: WalnutManual Production. Publication 3373 ofthe Univ. of California: 84-89.
Ramos, D.; DeJong, T.M.; Ryugo, K.;Olson, W.H.; Reil, W.O.; Sibbett, G.S.;Krueger, W.H.; Snyder, R.G. 1997. Pru-ning and tree thinning.
Ramos, D.; Kelly, K.; Reil, W.; SibertG.S.; Snyder, R. 2000. Establishmentand management considerations forwalnut hedgerow orchards. Acta Horti-culturae 544: 427-435.
N. Aletà and A. Ninot.Institut de Recerca i Tecnologia
Agroalimentàries (IRTA)Mas Bové. apartat 415.
43280 Reus. spain.E-mail. [email protected]
SELECTION FROM NATURALPOPULATION AND
HYBRIDIZATION OF WALNUTAT THE FACULTY
OF AGRICULTURE IN NOVISAD YUGOSLAVIA
ABSTRACTDespite the fact that in the largest part ofYugoslavia weather and soil conditionsare favourable for walnut growing, it isstill not wide spread (approximately 2.3 %comparing with other fruit trees). If wewant to increase walnut production, re-search on walnut selection should beco-me more important.
As walnut reproduction is almost exclusi-vely generative, its selection from the na-tural population is significant to enablefast and economical development of va-
rieties and selections which are to be re-produced vegetatively or used for plan-ned crossing.
The Faculty of Agriculture in Novi Sadhas been engaged in walnut selection formore than four decades, since it wasfounded. Rudic was the first at the Facul-ty who was involved in the work.
This paper presents results of walnut se-lection at the Faculty of Agriculture inNovi Sad where all the researchers whoselected the only existing five Yugosla-vian walnut cultivars are from. Consideringyield and nut quality, many selections, un-der the specific conditions of the area, ex-ceed the most famous world varieties. Inthe first place we will mention selection‘Rasna’ which is characterised by a veryhigh nut quality and high yield from thenursery. This paper shows tables of cha-racteristics of our varieties and selectionscomparing to well known cultivars ‘Sejno-vo’ and ‘Franquette’.
INTRODUCTIONWalnut trees in Yugoslavia have been re-produced generatively for centuries. Thisway of reproduction resulted in a large po-pulation. Among many genotypes, thosewith less quality nuts prevail, however tho-se of excellent quality are also possible tobe found. Besides, by crossing varietiesand selections with excellent separate cha-racteristics it is possible to produce hybridswhich could exceed the existing.
Researchers involved in walnut selectionin Yugoslavia are Fazlagic (1935), Rudic(1962), Jelenkovic (1966), Djurdjevic(1968), Manusev (1970), Hlisc (1972),Korac, Cerovic, Golosin (1974, 1976,1982, 1989, 1990, 1992, 1995, 1997),Kuzmanovski (1976), Miletic (1990), Bu-garcic (1985) etc.
MATERIAL AND METHODSThe Faculty of Agriculture in Novi Sadhas been engaged in walnut selectionsince 1957. At the beginning Rudic(1962) selected high quality types princi-pally from tree rows and walnut centres inVojvodina at the scene. The resultsachieved were only fair as research timewas short and the number of observedtrees small.
An original method of assembling primarymaterial has been applied since 1973. Byannouncing an open competition “Loo-king for high quality walnuts” each yearwe gathered fruits which were consideredhigh-quality by growers. The quality of thefruits was assessed at laboratory. A scoreof 20 points was given to the nuts with alight colored shell, high kernel-shell ratio(ranging from 54 to 58%) and fruit weightexceeding 14 g. Ten points were given for
the best taste, nice appearance of theshell (smooth and firmly closed), suitabili-ty for storage (iodine number 145 andless) and high commercial value of thenuts. A sample could be given 100 pointsat the most.
More than 1000 samples from all over thecountry were assessed in this way. Abroad selection was made, which inclu-ded types with high-quality nuts. After themain trees were observed over a long pe-riod of time, several types were chosenon the basis of the nut quality, yield, re-sistance to frost and diseases and thenthey were reproduced vegetatively andexamined together with famous foreignwalnut cultivars in the research orchardsin Beska and Veternik. The best five se-lections were named as cultivars: ‘Sam-pion’, ‘Srem’ and ’Tisa’ in 1987 and ‘Bac-ka’ and ‘Mire’ in 1988 and two more se-lections are in the process of selection.
In the first place hybridization was con-ducted to produce hybrids characterizedby a shorter period of vegetation compa-ring to our variety ‘Sampion’. ‘Sampion’and ‘Rasna’ were used as mothers and‘Kasni rodni’ and ‘Geisenheim 251’ as thefathers of the selection.
RESULTSBy assessment of seven most importantcharacteristics of the fruits, our twelvecultivars and selections were given scoreof 91 or more points and, according to thefruit quality, they were classified into theextra class.
‘Sampion’ scored 96, ‘Backa’ 95 and ‘YUprvenac’ 95 (this selection is susceptibleto Gnomonia juglandis, and after a longresearch it was eliminated from the res-tricted choice of the best cultivars and se-lections). The total of 68 selections hadfruits belonging to the first class (whichscored 81 to 90). Tables 1 and 2 presentthe fruit characteristics of our varietiesand selections comparing to varieties‘Sejnovo’ and ‘Franquette’.
Considering the data presented in Table1 it can be concluded that varieties ‘Sam-pion’, ‘Srem’ and ’Tisa’ and selections‘Rasna’ and ‘Macva’ have very large fruitsand the others medium large fruits. Mostcultivars and selections are characterizedby a very thin shell. Table 2 shows that allour cultivars and selections have a highor very high kernel / shell ratio. Oil con-tent is high and in ‘Backa’ very high. Thecolour of the kernel is the most beautifulin ‘Backa’, and in ‘Sampion’, ‘Rasna’,‘Kasni rodni’ and the French variety‘Franquette’.
The results of some phenological obser-vations are presented in Table 3. Most of
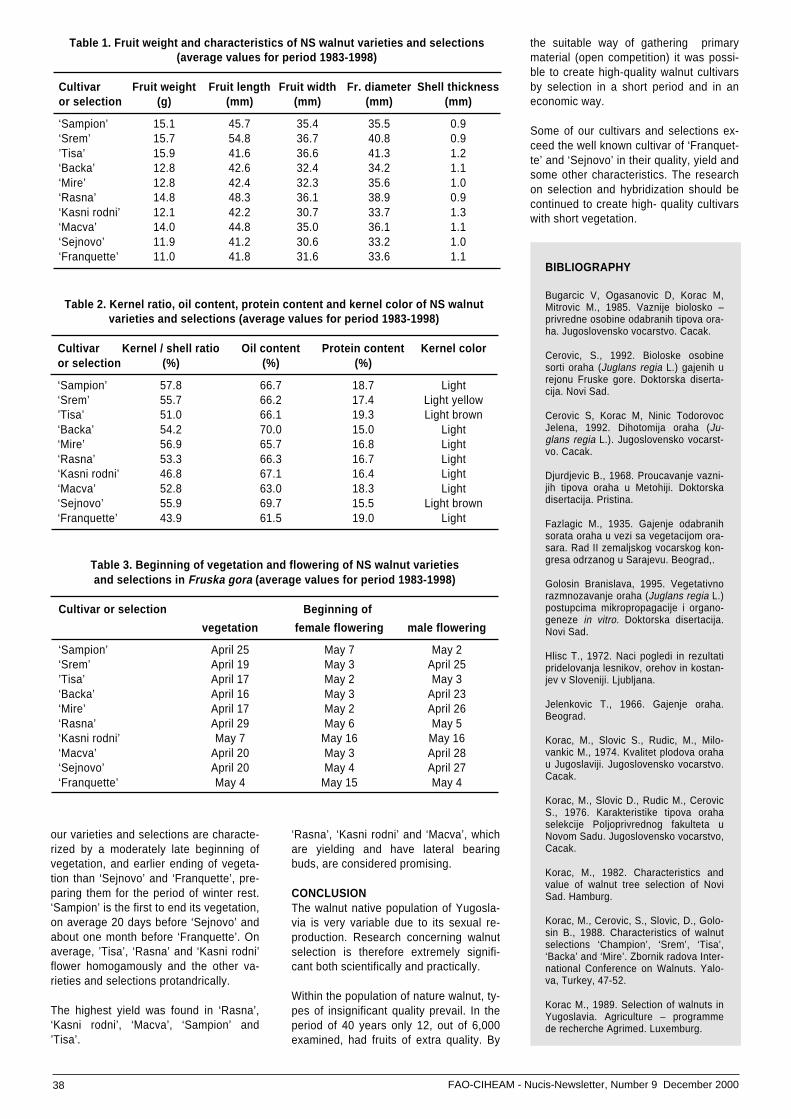
38 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
our varieties and selections are characte-rized by a moderately late beginning ofvegetation, and earlier ending of vegeta-tion than ‘Sejnovo’ and ‘Franquette’, pre-paring them for the period of winter rest.‘Sampion’ is the first to end its vegetation,on average 20 days before ‘Sejnovo’ andabout one month before ‘Franquette’. Onaverage, ’Tisa’, ‘Rasna’ and ‘Kasni rodni’flower homogamously and the other va-rieties and selections protandrically.
The highest yield was found in ‘Rasna’,‘Kasni rodni’, ‘Macva’, ‘Sampion’ and’Tisa’.
‘Rasna’, ‘Kasni rodni’ and ‘Macva’, whichare yielding and have lateral bearingbuds, are considered promising.
CONCLUSIONThe walnut native population of Yugosla-via is very variable due to its sexual re-production. Research concerning walnutselection is therefore extremely signifi-cant both scientifically and practically.
Within the population of nature walnut, ty-pes of insignificant quality prevail. In theperiod of 40 years only 12, out of 6,000examined, had fruits of extra quality. By
Table 1. Fruit weight and characteristics of NS walnut varieties and selections(average values for period 1983-1998)
Cultivar Fruit weight Fruit length Fruit width Fr. diameter Shell thicknessor selection (g) (mm) (mm) (mm) (mm)
‘Sampion’ 15.1 45.7 35.4 35.5 0.9‘Srem’ 15.7 54.8 36.7 40.8 0.9’Tisa’ 15.9 41.6 36.6 41.3 1.2‘Backa’ 12.8 42.6 32.4 34.2 1.1‘Mire’ 12.8 42.4 32.3 35.6 1.0‘Rasna’ 14.8 48.3 36.1 38.9 0.9‘Kasni rodni’ 12.1 42.2 30.7 33.7 1.3‘Macva’ 14.0 44.8 35.0 36.1 1.1‘Sejnovo’ 11.9 41.2 30.6 33.2 1.0‘Franquette’ 11.0 41.8 31.6 33.6 1.1
Table 2. Kernel ratio, oil content, protein content and kernel color of NS walnutvarieties and selections (average values for period 1983-1998)
Cultivar Kernel / shell ratio Oil content Protein content Kernel coloror selection (%) (%) (%)
‘Sampion’ 57.8 66.7 18.7 Light‘Srem’ 55.7 66.2 17.4 Light yellow’Tisa’ 51.0 66.1 19.3 Light brown‘Backa’ 54.2 70.0 15.0 Light‘Mire’ 56.9 65.7 16.8 Light‘Rasna’ 53.3 66.3 16.7 Light‘Kasni rodni’ 46.8 67.1 16.4 Light‘Macva’ 52.8 63.0 18.3 Light‘Sejnovo’ 55.9 69.7 15.5 Light brown‘Franquette’ 43.9 61.5 19.0 Light
Table 3. Beginning of vegetation and flowering of NS walnut varietiesand selections in Fruska gora (average values for period 1983-1998)
Cultivar or selection Beginning of
vegetation female flowering male flowering
‘Sampion’ April 25 May 7 May 2‘Srem’ April 19 May 3 April 25’Tisa’ April 17 May 2 May 3‘Backa’ April 16 May 3 April 23‘Mire’ April 17 May 2 April 26‘Rasna’ April 29 May 6 May 5‘Kasni rodni’ May 7 May 16 May 16‘Macva’ April 20 May 3 April 28‘Sejnovo’ April 20 May 4 April 27‘Franquette’ May 4 May 15 May 4
the suitable way of gathering primarymaterial (open competition) it was possi-ble to create high-quality walnut cultivarsby selection in a short period and in aneconomic way.
Some of our cultivars and selections ex-ceed the well known cultivar of ‘Franquet-te’ and ‘Sejnovo’ in their quality, yield andsome other characteristics. The researchon selection and hybridization should becontinued to create high- quality cultivarswith short vegetation.
BIBLIOGRAPHY
Bugarcic V, Ogasanovic D, Korac M,Mitrovic M., 1985. Vaznije biolosko –privredne osobine odabranih tipova ora-ha. Jugoslovensko vocarstvo. Cacak.
Cerovic, S., 1992. Bioloske osobinesorti oraha (Juglans regia L.) gajenih urejonu Fruske gore. Doktorska diserta-cija. Novi Sad.
Cerovic S, Korac M, Ninic TodorovocJelena, 1992. Dihotomija oraha (Ju-glans regia L.). Jugoslovensko vocarst-vo. Cacak.
Djurdjevic B., 1968. Proucavanje vazni-jih tipova oraha u Metohiji. Doktorskadisertacija. Pristina.
Fazlagic M., 1935. Gajenje odabranihsorata oraha u vezi sa vegetacijom ora-sara. Rad II zemaljskog vocarskog kon-gresa odrzanog u Sarajevu. Beograd,.
Golosin Branislava, 1995. Vegetativnorazmnozavanje oraha (Juglans regia L.)postupcima mikropropagacije i organo-geneze in vitro. Doktorska disertacija.Novi Sad.
Hlisc T., 1972. Naci pogledi in rezultatipridelovanja lesnikov, orehov in kostan-jev v Sloveniji. Ljubljana.
Jelenkovic T., 1966. Gajenje oraha.Beograd.
Korac, M., Slovic S., Rudic, M., Milo-vankic M., 1974. Kvalitet plodova orahau Jugoslaviji. Jugoslovensko vocarstvo.Cacak.
Korac, M., Slovic D., Rudic M., CerovicS., 1976. Karakteristike tipova orahaselekcije Poljoprivrednog fakulteta uNovom Sadu. Jugoslovensko vocarstvo,Cacak.
Korac, M., 1982. Characteristics andvalue of walnut tree selection of NoviSad. Hamburg.
Korac, M., Cerovic, S., Slovic, D., Golo-sin B., 1988. Characteristics of walnutselections ‘Champion’, ‘Srem’, ‘Tisa’,‘Backa’ and ‘Mire’. Zbornik radova Inter-national Conference on Walnuts. Yalo-va, Turkey, 47-52.
Korac M., 1989. Selection of walnuts inYugoslavia. Agriculture – programmede recherche Agrimed. Luxemburg.

39FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Korac M., Cerovic S., Golisin B., MileticR., 1990. Population variability of do-mestic walnut and selection results. Ju-goslovensko vocarstvo, Cacak.
Korac M., Cerovic S., Golosin B., Ogn-janov V., Miletic R., 1993. PerspectiveYugoslavian walnut selections with late-ral fruit buds. Acta Horticulturae, 311:41-46.
Korac M., Cerovic S., Golosin B, MileticR., 1997. Collecting, evaluation and uti-lization of walnut (Juglans regia L.) inYugoslavia. Plant Genetic ResourcesNewsletter.
Kuzmanovski I., Georgiev D., PopovskiH., 1976. Biolosko – morfoloske osobi-ne nekih tipova oraha. Jugoslovenskovocarstvo, Cacak.
Rudic M., 1962. Selekcije oraha uVojvodini. Arhiv za poljoprivrednenauke, Beograd.
Korac, M.; Cerovic, S.; Golosin, B.; Korac, J.University of Novi SadFaculty of Agriculture
D. Obradovica, 821000 Novi Sad, Yugoslavia
THE TAXONOMICRANKING OF
Pistacia terebinthusand P. palaestina
Botanists dealing with the flora and vege-tation of the Mediterranean basin recogni-ze in this phyto-geographical region twoclosely related terebinth-type wild pista-chio taxa. The classic attitude (see Bois-sier 1872; M. Zohary 1952) has been torank them as full-fledged vicarious spe-cies: (i) Pistacia terebinthus L. (in thewestern part of the Mediterranean re-gion); and (ii) P. palaestina Boiss. (in theeastern part of this phyto-geographic re-gion). However, several workers (e.g.Engler 1883; Yaltirik 1967; Meikle 1977;Browicz 1984) considered them conspeci-fic, and treated them only as two subspe-cies within P. terebinthus. Both taxa resem-ble each other rather closely - both intheir habit and in their morphology. Theyare small (up to 6 (8) m high), ramified,deciduous trees, frequently developingseveral trunks. They also show parallelecological specificities: Both are commonconstituents of the Mediterranean ever-green maquis vegetation; and as the caseis in numerous arboreal elements in thisformation, their populations are frequentlyquite variable. The main traits used todistinguish between these taxa (Boissier1872; M. Zohary 1952) are leaf morpholo-gy and geographic distribution. P. tere-binthus [syn. P. terebinthus L. subsp. te-rebinthus] is described as having impari-
pinnate leaves that carry a fully develo-ped terminal leaflet; and its lateral leafletsare obtuse (Fig. 1). In contrast, P. palaes-tina Boiss. [syn. P. terebinthus L. subsp.palaestina (Boiss.) Engl.] has - almostalways - paripinnate leaves in which theterminal leaflet is absent or drastically re-duced, and the lateral leaflets are acumi-nate (Fig. 1). Phyto-geographically, P. te-rebinthus has been classified as a west-Mediterranean element, distributed fromthe Aegean basin westward to the Medi-terranean parts of Morocco and southPortugal; and it extends also to thesouthern coast of the Black Sea. P. pa-laestina has been regarded as an east-Mediterranean element, confined to theLevant countries and to south Anatolia.
The problem how to rank taxonomicallythese two Pistacia taxa is still valid today.More so since the terebinthus-palaestina‘aggregate’ is one of the characteristicfeatures of the Mediterranean vegetation,and because there is a growing interest inthese plants among the pistachiogrowers. The main questions that have tobe answered in order to clarify their sta-tus are as follows:
(1) What is the pattern of morphologi-cal variation in the terebinthus-palaestina‘aggregate’ in the Mediterranean region?Is the variation in it continuous or discon-tinuous? In other words, do we have hereclines and intermediate populations thatbridge the morphological features of thetypical terebinthus and palaestina taxa?Alternatively, are these two taxa separa-ted by distinct discontinuities?
(2) Is the distribution area of the‘aggregate’ more or less continuous? Al-
ternatively, is a morphological distinctionbetween these two taxa accompanied bygeographic separation?
(3) How well do we know today the va-riation patterns in, and the distribution of,the terebinthus-palaestina ‘aggregate’?Have we already sufficient information formaking a decision; or are we still liable toarrive at wrong conclusions because ofmajor gaps in our information?
Until the middle of the 20th Century, thetaxonomic treatment of the terebinthus-palaestina ‘aggregate’ (particularly in theeastern part of the Mediterranean basin)was based - almost entirely – on a very li-mited number of herbarium collections.Surveys of variation in populations, parti-cularly in the critical area of the Aegeanbasin and south and west Turkey, werepractically non-existent. Also the informa-tion about the spread of these wild pista-chios over the eastern Mediterranean ba-sin was fragmentary. On basis of theavailable herbarium sheets the impres-sion of both the author of P. palaestina(Boissier 1872), and the monographer ofthe genus (M. Zohary 1952) was that pa-laestina material obtained from the Le-vant is distinct, and deserves specificrank. The situation changed drastically inthe last forty years. Numerous new her-barium specimens and records wereadded, including careful examination ofthe variation in populations in the field. Amost extensive survey was carried out byBrowicz and his associates (Browicz1984, Boratynski et al. 1992). It includedseveral field trips to Turkey, to the Ae-gean islands and to Greece. Similar exa-minations were made by Meikle (1977) in
Figure 1. Leaf morphology in typical Pistacia terebinthus (left) and P.palaestina (right).Adapted from M. Zohary (1952)

40 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Cyprus. In addition, several other bota-nists examined the terebinthus- palaesti-na ‘aggregate’ in the field, and while theirobservations remained mostly unpublis-hed, they shared them with their collea-gues. All in all, today the information wehave on these taxa is indeed much morereliable than 30-40 years ago. It revealsthat the distribution of the terebinthus-pa-laestina ‘aggregate’ in the Mediterraneanvegetation belt in Greece, Turkey, Cyprusand the Levant is rather continuous (seeBrowicz 1984, Map 3). It also shows thatthe two types are not separated by mor-phological discontinuities. Particularly insouth-west Turkey and in Cyprus one isfaced with massive, intermediate popula-tions, comprising a whole range of inter-mediate and recombinant forms that brid-ge the terebinthus and palaestina “pure”types. In other words, a west to east clineexists in this aggregate, particularly be-tween the Aegean basin and the Levant.
In conclusion, the available informationstrongly suggests that P. terebinthus andP. palaestina are not independent vica-rious species but only two intergradinggeographic races or subspecies. Theirappropriate botanical naming is thereforeP. terebinthus L. subsp. terebinthus andP. terebinthus L. subsp. palaestina(Boiss.) Engl.
REFERENCES
Boissier, E., 1872. Flora orientalis Vol.2, p. 6. H. George, Geneva and Basel.
Boratynski, A. Browicz, K. and Zielinsky, J.,1992. Chorology of trees and shrubs inGreece. Institute of Dendrology, Polish Aca-demy of Sciences. Kornik, Poznan. p.
Browicz, K., 1984. Chorology of treesand shrubs in south-west Asia and adja-cent regions Vol. 3. Polish scientific Pu-blishers, Warszawa-Poznan. 88 pp.
Engler, A., 1883. Pistacia. In: A. and C.De Candole (eds.) Monographiae Pha-nerogamarum, Vol. 4, pp. 284-293
Meikle, R. D., 1977. Flora of Cyprus Vol.1.Royal Botanical Gardens, Kew, UK.
Yaltirik, F., 1967. Pistacia L. In: P. H.Davis (ed.) Flora of Turkey and the eastAegean islands Vol. 2, pp. 544-9. Edin-burgh University Press, Edinburgh, UK
Zohary, M., 1952. A monographic studyof the genus Pistacia. Palestine Jour.Bot. (Jerusalem series) 5, 187-228.
D. ZoharyDepartment of Evolution, Systematics
and EcologyThe Hebrew University, Jerusalem 91904, Israel
E-mail: [email protected]
THE INFLUENCESOF DIFFERENT IRRIGATION
PROGRAMMES ON WEIGHT OFPISTACHIO LEAVES
ABSTRACTThe study was carried out in Gaziantepregion, Turkey, between the middle ofMay and early September in 1999. Pista-chio orchard where this study was carriedout is almost 3 ha and contains 280 pista-chio trees being 25 years old. In the stu-dy, the characteristics of ‘Uzun’ pistachiovariety, which is a P. vera type, was stu-died. Different deficit irrigation program-mes; 7 days (I1), and 14 days (I2) were im-plemented, and different crop coeffi-cients; 0.60 (kcp), and 0.90 (kcp) wereused during the study (I1 0.60, I1 0.90 andI2 0.60, I2 0.90). At the same time, diffe-rent nitrogen doses (NO, N1, N2, N3), andsame amounts of potassium (K) andphosphorus (P) were applied togetherwith different irrigation deficits.
522 leaf samples were collected at ran-dom from all trees. These leaves weredried and weighed at the laboratory.
Pistachio orchard at Ceylanpinar farm in Anatolia, Turkey
Dried leaves were observed at differentcrop coefficients: 0.60 (kcp) and 0.90(kcp). Results of observations highlightedthat dried weight of samples at 0.90 (kcp)coefficient were found high in nitrogen indifferent deficit irrigation. The highest dryweight of leaf sample was found atI2N290. Dry weight of leaf samples at 0.60(kcp) was found high except nitrogen (N1)dose in all treatments (NO, N2, N3) at diffe-rent deficit irrigation ranges. The highestdry weight of leaf sample was observedat I2N060. The lowest dried leaf weight ofsamples was found at I1N260.
Amount of irrigation, and applied fertili-zers, and nitrogen with different irrigationdeficit are shown in Table 1, Table 2, andFigure 1a and 1b.
S. ÖzmenUniversity of Cukurova, Faculty of Agriculture
Department of Agricultural Structures &Irrigation.
Adana, TurkeyPhone: 90 322 338 63 88
Fax: 90 322 458 4909E-mail: [email protected]
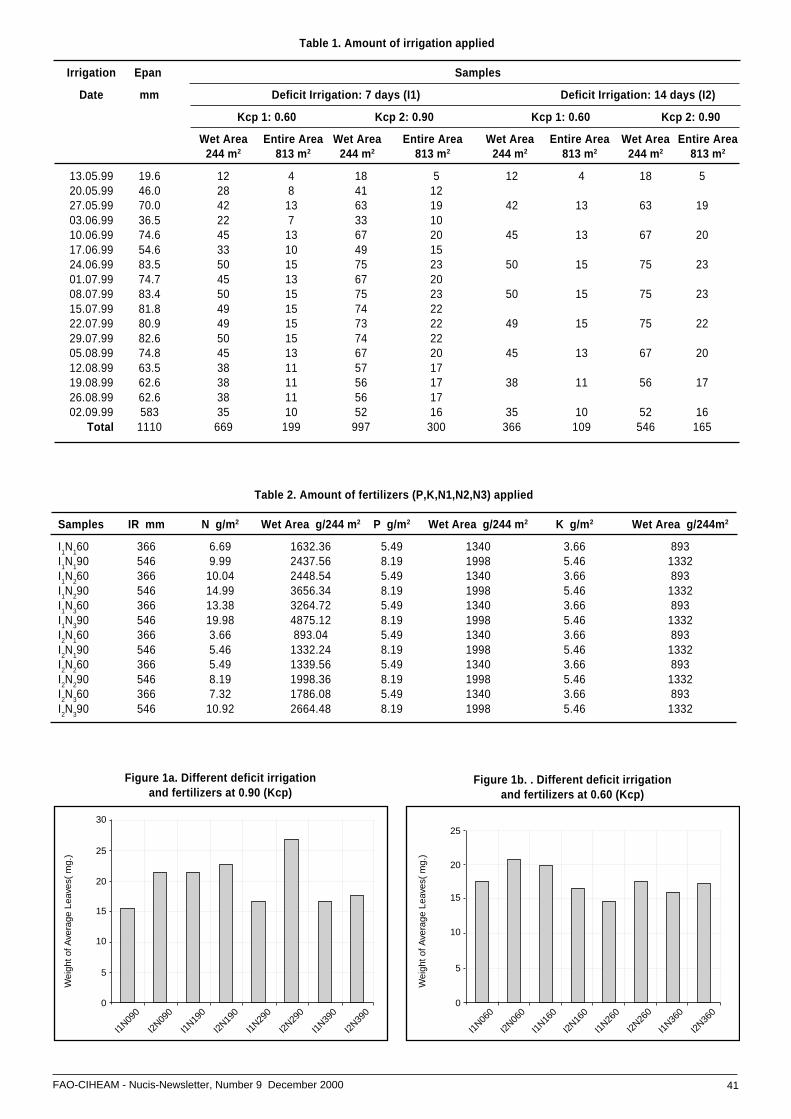
41FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Table 1. Amount of irrigation applied
Irrigation Epan Samples
Date mm Deficit Irrigation: 7 days (I1) Deficit Irrigation: 14 days (I2)
Kcp 1: 0.60 Kcp 2: 0.90 Kcp 1: 0.60 Kcp 2: 0.90
Wet Area Entire Area Wet Area Entire Area Wet Area Entire Area Wet Area Entire Area244 m2 813 m2 244 m2 813 m2 244 m2 813 m2 244 m2 813 m2
13.05.99 19.6 12 4 18 5 12 4 18 520.05.99 46.0 28 8 41 1227.05.99 70.0 42 13 63 19 42 13 63 1903.06.99 36.5 22 7 33 1010.06.99 74.6 45 13 67 20 45 13 67 2017.06.99 54.6 33 10 49 1524.06.99 83.5 50 15 75 23 50 15 75 2301.07.99 74.7 45 13 67 2008.07.99 83.4 50 15 75 23 50 15 75 2315.07.99 81.8 49 15 74 2222.07.99 80.9 49 15 73 22 49 15 75 2229.07.99 82.6 50 15 74 2205.08.99 74.8 45 13 67 20 45 13 67 2012.08.99 63.5 38 11 57 1719.08.99 62.6 38 11 56 17 38 11 56 1726.08.99 62.6 38 11 56 1702.09.99 583 35 10 52 16 35 10 52 16 Total 1110 669 199 997 300 366 109 546 165
25
20
15
10
5
I1N06
0
I2N06
0
I1N16
0
I2N16
0
I1N26
0
I2N26
0
I1N36
0
I2N36
00
Wei
ght o
f Ave
rage
Lea
ves(
mg.
)
30
25
20
15
10
5
I1N09
0
I2N09
0
I1N19
0
I2N19
0
I1N29
0
I2N29
0
I1N39
0
I2N39
00
Wei
ght o
f Ave
rage
Lea
ves(
mg.
)
Figure 1a. Different deficit irrigationand fertilizers at 0.90 (Kcp)
Figure 1b. . Different deficit irrigationand fertilizers at 0.60 (Kcp)
Table 2. Amount of fertilizers (P,K,N1,N2,N3) applied
Samples IR mm N g/m2 Wet Area g/244 m2 P g/m2 Wet Area g/244 m2 K g/m2 Wet Area g/244m2
I1N160 366 6.69 1632.36 5.49 1340 3.66 893I1N190 546 9.99 2437.56 8.19 1998 5.46 1332I1N260 366 10.04 2448.54 5.49 1340 3.66 893I1N290 546 14.99 3656.34 8.19 1998 5.46 1332I1N360 366 13.38 3264.72 5.49 1340 3.66 893I1N390 546 19.98 4875.12 8.19 1998 5.46 1332I2N160 366 3.66 893.04 5.49 1340 3.66 893I2N190 546 5.46 1332.24 8.19 1998 5.46 1332I2N260 366 5.49 1339.56 5.49 1340 3.66 893I2N290 546 8.19 1998.36 8.19 1998 5.46 1332I2N360 366 7.32 1786.08 5.49 1340 3.66 893I2N390 546 10.92 2664.48 8.19 1998 5.46 1332

42 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
CUTTING PROPAGATION OFMASTIC TREE
(Pistacia lentiscus var. Chia Duham.)
INTRODUCTIONPistacia lentiscus L. is one of the mostcommon plant species of coastal Mediter-ranean. It is an evergreen shrub (rarelysmall trees) with leathery leaves andconspicously winged rachis. A circum Me-diterranean species, and a leading com-ponent of thermophylous low-altitude ma-quis vegetation in the Mediterranean ba-sin, extending to Madeira island (Zohary1995).
It is the unique species appreciated forgum mastic obtained by wounding thetrunk and branches of the plant. Gummastic is a natural resin, or more correct-ly an oleoresin obtained from P. lentiscusvar. chia Duham. (syn: P. lentiscus var.latifolia Coss.) differing from the real spe-cies through its leaf morphology and tree-like growth (Baytop, 1968; Browicz,1987). Only male trees are being used formastic production (Perikos, 1993).
Apart from pharmaceutical properties,mastic has been widely used to provideflavour for a range of traditional foods andalcoholic distilled beverages. Mastic treecultivation and gum mastic productionhave been associated for a long time withthe Greek island of Chios. This relic in-dustry has a significant role in island eco-nomy (Perikos, 1993).
Although it is not as extensive and inquantity as on Chios island, mastic hadbeen produced in the adjacent westernAnatolia, on Çesme peninsula where theecological conditions are identical (Bay-top, 1968). However, agricultural activi-ties gradually diminished as a result of ra-pid socio-economic transformation in pe-ninsula. Thus, cultivation was even nega-tively affected by these circumstances,number of trees dramatically decreasedand mastic production has stopped.
Recent field observations indicated thatthere were less than 150 male individualswho showed the characteristics of realvariety in the vicinity of Çesme. But theyare very old and destructed in various de-grees. Remnants of old plantations havebeen legally conserved since 1995.Without regarding the economic profits,mastic tree should be considered as animportant loss from Pistacia genetic re-sources of Turkey.
It was supposed that one of the limitingfactors to extend mastic tree growing isthe difficulty in clonal propagation. Tradi-tional propagation has been carried outby direct sticking of hardwood cuttings to
the soil where the plantation will be esta-blished (Perikos, 1993). In this method,rooting success is generally low and acci-dental. No budding or grafting successhas been reported so far.
Cuttings from mature trees of Pistaciaspecies are considered very difficult-to-root (Joley and Opitz, 1971). Proper cut-ting collection timing is crucial for rootingsuccess in Pistacia species. Cuttingscould be rooted during a very short periodof the year if they were prepared from ac-tively growing shoots. In this case, eventhe effect of applied auxin is more pro-nounced (Avanzato and Damiano, 1990;Dunn et al., 1996a). Attempts to root Pis-tacia cuttings from mature trees havebeen limited with high concentrations ofauxin or auxin combinations (Al Baraziand Schwabe, 1982; Dunn et al., 1996b).Juvenility in Pistacia species generallyseems to be lost after two years (Pair andKhatamian, 1983; Lee et al., 1976; Al Ba-razi and Scwabe, 1984), making cuttingpropagation more difficult. Semi-hardwo-od cuttings of mature mastic tree had re-latively high rooting percentage with15000 ppm IBA when taken in March.Further softwood cuttings did not survive(Isfendiyaroglu and Karakir, 1994).
The objectives of this study were to (1)determine the most proper cutting collec-tion time, and (2) optimum concentrationsof auxins and their combinations to opti-mize rapid cutting propagation of mastictree. Thus, survival of this genetic resour-ce would be ensured and the sufficientquantities of nursery stock could be provi-ded to establish the new plantations. Twoseparate plantations of 30 and 300 treeshave already been established.
MATERIALS AND METHODSSemi-hardwood leafy terminal cuttingswere collected from old but relatively wellprotected male trees. In the first year ofexperiments, cuttings were taken at 1-month or sometimes shorter intervalsfrom 15 November 1995 through 7 April1996 and 5 September 1996 through 20November 1996. They were treated withindole butyric acid (IBA) at concentrationsvarying from 0 to 20 g.l-1 dissolved in 50% isopropanol by 5 sec. quick dip. Takinginto account the first years results, in thesecond year, cuttings were taken on 15January, 1 March and 1 April 1997. Theywere treated with 0, 20, 30, 40 g.l-1 IBA;10, 20 g.l-1 naphtalene acetic acid (NAA);10, 20 g.l-1 indole acetic acid (IAA); 10 g.l-1 IBA + 2.5, 5, 10 g.l-1 NAA solutions. Be-fore auxin treatments, cuttings were recutto 15-20 cm long and two third of the lea-ves and whole male inflorescence wereremoved. Cuttings were placed in rootingbed containing pure perlite under the lowtunnel misting system. Misting intervals
controlled by an electronic leaf and bot-tom heat was adjusted to 25°C. The per-centage of rooted cuttings were recordedapproximately 12 weeks after planting. Inexperiments, a randomized block designwas used with three replications of tencuttings for each treatment combination.Statistical analysis were carried out usingSPSS 5.0. Data were analysed by ANO-VA and Duncan’s multiple-range test wasused to discern differences at the 5 per-cent level of significance.
RESULTS AND DISCUSSIONThere was a significant interaction bet-ween cutting date and IBA concentrationin the first year (Table 1).
Cuttings that were taken on 15 Februaryand treated with 20 g.l-1 IBA had highest(76.6 %) rooting percentage (increasinglevels of IBA raised rooting compared tocuttings that were taken on all other da-tes). Despite the IBA treatments, rootingpercentages were sharply decreased on2 March and the effect of IBA also de-creased after a certain level. Followingdates cuttings did not respond well to in-creasing concentrations of IBA and therooting percentages had lower values.Mature Pistacia chinensis has very shortperiod (4 weeks) of rootability. Rootingwas highest only in softwood cuttingstaken in May and treated with 8.750 mg l-1
IBA, two or four fold increase in concen-tration decreased rooting (Dunn et al.,1996a). Only softwood cuttings collected30 and 60 days after sprouting rootedwell with 4000 ppm IBA, while semi hard-wood cuttings failed to root in Pistacia at-lantica (Avanzato and Damiano, 1990).Despite only semi-hardwood cuttingswere used in all experiments, mastic treealso has greatest potential to root forma-tion in a very limited period as reported inother Pistacia species.
In the second year, significant interactionwas predicted between cutting date anddifferent auxin concentration (Table 2).The highest percentage of rooted cuttingswas obtained when they were taken on15 February and treated with 10 g.l-1 IBA+ 2.5 g.l-1 NAA combination. Increase inNAA proportion in combinations signifi-cantly reduced the rooting percentage.Similarly, it has been reported that soft-wood cuttings of mature P. chinensisgave the highest rooting percentagewhen IBA combined with NAA in equalparts (5 g.l-1) and proportional increasesin NAA reduced rooting (Dunn et al.,1996b).
Rooting percentages significantly increa-sed in parallel with IBA concentrations upto 40 g.l-1 on 15 February (Table 2). Butall IBA treatments were less effectivethan 10 g.l-1 IBA+2.5 g.l-1 NAA combina-
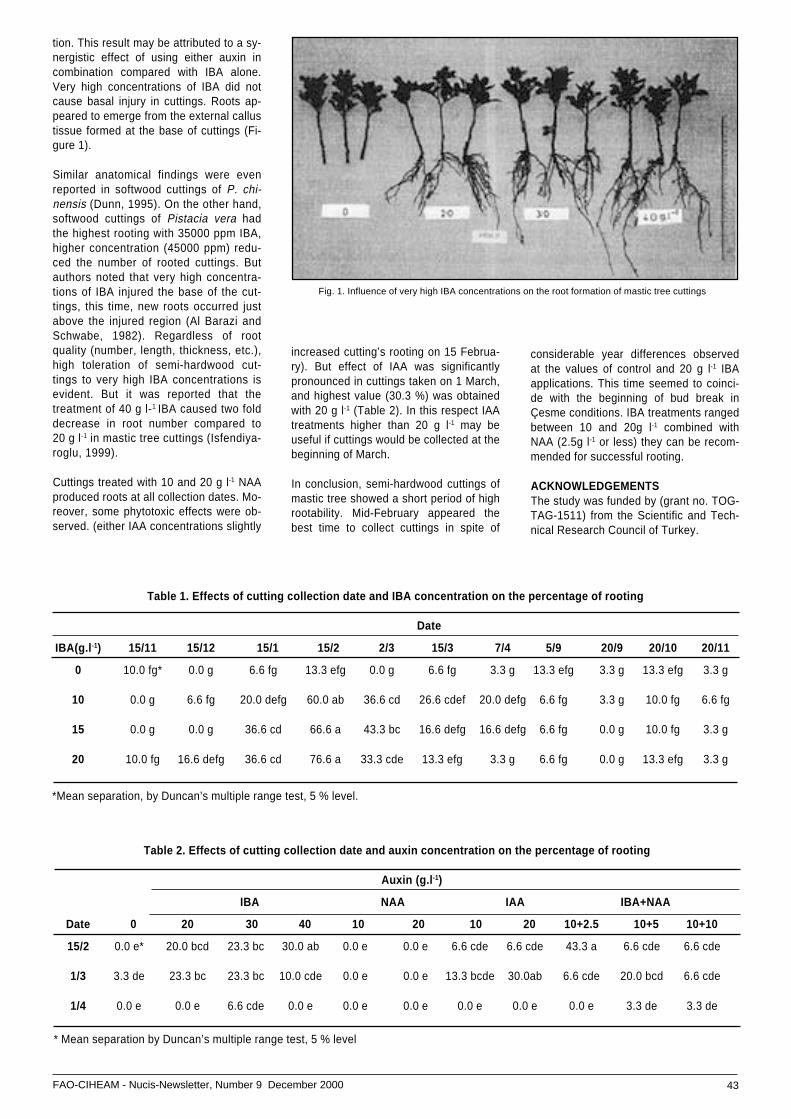
43FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
tion. This result may be attributed to a sy-nergistic effect of using either auxin incombination compared with IBA alone.Very high concentrations of IBA did notcause basal injury in cuttings. Roots ap-peared to emerge from the external callustissue formed at the base of cuttings (Fi-gure 1).
Similar anatomical findings were evenreported in softwood cuttings of P. chi-nensis (Dunn, 1995). On the other hand,softwood cuttings of Pistacia vera hadthe highest rooting with 35000 ppm IBA,higher concentration (45000 ppm) redu-ced the number of rooted cuttings. Butauthors noted that very high concentra-tions of IBA injured the base of the cut-tings, this time, new roots occurred justabove the injured region (Al Barazi andSchwabe, 1982). Regardless of rootquality (number, length, thickness, etc.),high toleration of semi-hardwood cut-tings to very high IBA concentrations isevident. But it was reported that thetreatment of 40 g l-1 IBA caused two folddecrease in root number compared to20 g l-1 in mastic tree cuttings (Isfendiya-roglu, 1999).
Cuttings treated with 10 and 20 g l-1 NAAproduced roots at all collection dates. Mo-reover, some phytotoxic effects were ob-served. (either IAA concentrations slightly
increased cutting’s rooting on 15 Februa-ry). But effect of IAA was significantlypronounced in cuttings taken on 1 March,and highest value (30.3 %) was obtainedwith 20 g l-1 (Table 2). In this respect IAAtreatments higher than 20 g l-1 may beuseful if cuttings would be collected at thebeginning of March.
In conclusion, semi-hardwood cuttings ofmastic tree showed a short period of highrootability. Mid-February appeared thebest time to collect cuttings in spite of
considerable year differences observedat the values of control and 20 g l-1 IBAapplications. This time seemed to coinci-de with the beginning of bud break inÇesme conditions. IBA treatments rangedbetween 10 and 20g l-1 combined withNAA (2.5g l-1 or less) they can be recom-mended for successful rooting.
ACKNOWLEDGEMENTSThe study was funded by (grant no. TOG-TAG-1511) from the Scientific and Tech-nical Research Council of Turkey.
Table 1. Effects of cutting collection date and IBA concentration on the percentage of rooting
Date
IBA(g.l-1) 15/11 15/12 15/1 15/2 2/3 15/3 7/4 5/9 20/9 20/10 20/11
0 10.0 fg* 0.0 g 6.6 fg 13.3 efg 0.0 g 6.6 fg 3.3 g 13.3 efg 3.3 g 13.3 efg 3.3 g
10 0.0 g 6.6 fg 20.0 defg 60.0 ab 36.6 cd 26.6 cdef 20.0 defg 6.6 fg 3.3 g 10.0 fg 6.6 fg
15 0.0 g 0.0 g 36.6 cd 66.6 a 43.3 bc 16.6 defg 16.6 defg 6.6 fg 0.0 g 10.0 fg 3.3 g
20 10.0 fg 16.6 defg 36.6 cd 76.6 a 33.3 cde 13.3 efg 3.3 g 6.6 fg 0.0 g 13.3 efg 3.3 g
*Mean separation, by Duncan’s multiple range test, 5 % level.
Table 2. Effects of cutting collection date and auxin concentration on the percentage of rooting
Auxin (g.l-1)
IBA NAA IAA IBA+NAA
Date 0 20 30 40 10 20 10 20 10+2.5 10+5 10+10
15/2 0.0 e* 20.0 bcd 23.3 bc 30.0 ab 0.0 e 0.0 e 6.6 cde 6.6 cde 43.3 a 6.6 cde 6.6 cde
1/3 3.3 de 23.3 bc 23.3 bc 10.0 cde 0.0 e 0.0 e 13.3 bcde 30.0ab 6.6 cde 20.0 bcd 6.6 cde
1/4 0.0 e 0.0 e 6.6 cde 0.0 e 0.0 e 0.0 e 0.0 e 0.0 e 0.0 e 3.3 de 3.3 de
* Mean separation by Duncan’s multiple range test, 5 % level
Fig. 1. Influence of very high IBA concentrations on the root formation of mastic tree cuttings

44 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
REFERENCES
Al Barazi, Z. and Schwabe, W.W., 1982.Rooting softwood cuttings of adult Pis-tacia vera. Journal of HorticulturalScience, 57 (2): 247-252.
Al Barazi, Z. and Schwabe, W.W., 1984.The possible involvement of polyphe-nol-oxidase and the auxin-oxidase sys-tem in root formation and developmentin cuttings of Pistacia vera. Journal ofHorticultural Science, 59 (3): 453-461.
Avanzato, D. and Damiano, C., 1990.Study on rooting of Pistacia atlanticaDesf. XXIIIrd International HorticulturalCongress, Firenze (Italy), Abstracts ofContributed Papers, No. 4032.
Baytop, T., 1968. Les Possibilités deProduction du Mastic en Turquie. J.Fac. Pharm. of Istanbul Univ., 4 (1): 31-35. (in Turkish)
Browicz, K., 1987. Pistacia lentiscus cv.Chia (Anacardiaceae) on Chios island.Pl. Syst. Evol, 155: 189-195.
Dunn, D.E., 1995. Vegetative Propaga-tion of Chinese Pistache. MSc Thesis,Oklahoma State University, 155 p(unpublished).
Dunn, D.E., Cole, J.C. and Smith, M.W.,1996a. Timing of Pistacia chinensisBunge. rooting using morphologicalmarkers associated with calendar dateand degree days. J. Amer. Soc. Hort.Sci., 121 (2): 269-273.
Dunn, D.E., Cole, J.C. and Smith, M.W.,1996b. Position of cut, bud retentionand auxins influence rooting of Pistaciachinensis. Scientia Horticulturae 67:105-110.
Isfendiyaroglu, M. ve Karakir, M.N.,1994. Studies on the effects of variousfactors on rooting of leafy cuttings ofsome woody ornamental plants. J. ofEge Univ. Grad. Sch. of Nat. and App.Sci. (in Turkish, in press).
Isfendiyaroglu, M. 1999. Investigationson cutting propagation of the mastictree (Pistacia lentiscus var. Chia Du-ham.) and anatomical-physiological stu-dy of root formation. (in Turkish) PhDThesis, Ege University, Izmir, Turkey,123pp.
Joley, L.E. and Opitz, K.W., 1971. Fur-ther experiences with propagation ofPistacia. Proc. Int. Plant. Prop. Soc.,21: 67-76.
Lee, C.I., Paul, J.L. and Hackett, W.P.,1976. Root promotion on stem cuttingsof several ornamental plants by acid orbase pretreatment. Comb. Proc. Intl.Plant Prop. Soc. 26: 95-99.
Pair, J.C. and Khatamian, H., 1983.Propagation and growing of the chinesepistache. Proc. Int. Plant Prop. Soc., 32:497-503.
Perikos, J., 1993. The Chios Gum Mas-tic. Print All Ltd., ISBN 960-85009-3-1,Athens, Greece, 95p.
Zohary, D., 1995. The genus PistaciaL., 1-11, Taxonomy, Distribution, Con-servation and Uses of Pistacia GeneticResources. Padulosi, S., Caruso, T.and Barone, E. (Eds.), Report of aworkshop, 29-30 June 1995, Palermo,Italy, Int. Plant Gen. Res. Ins. Rome,Italy, 69p.
M. IsfendiyarogluEge University
Faculty of AgricultureDepartment of Horticulture
35100 Bornova / Izmir-TurkeyTel: 90 232 388400/2621
Fax: 90 232 3881864E-mail: [email protected].
THE CHESTNUT:THE ULTIMATE ENERGYSOURCE NUTRITIONAL
VALUE AND ALIMENTARYBENEFITS
Chestnut spreading areaChestnut tree species are present in theirnatural state in three major geographicalareas: Europe, where the European spe-cies (Castanea sativa Mill.) is cultivatedfor its nuts and wood. Asia, where Casta-nea crenata Sieb. and Zucc. are grown inJapan and C. mollissima BL. in China andKorea. North America from where Casta-nea dentata Borkh originates, a giantwood tree, which dominated in thebroadleaf forests to the east of the conti-nent, until the middle of the last centuryand which is now almost extinct due tothe spread of canker blight.
According to FAO statistics (1999) theworld production of chestnuts, concentra-ted mainly in China and Korea, exceeds afigure of 500,000 t per year, Italy accoun-ting for over 14 % of the figure. In the Eu-ropean context, Italy is the leading chest-nut producing country, with a figure ofapprox. 78,000 t a year, which accountsfor 51 % of the European production.
The chestnut tree has been cultivated inItaly since ancient times, throughout theentire Alpine chain up to a height of 800-900 m a.s.l., along the Apennine ridgeright down to Calabria, on the slopes ofMount Etna in Sicily and in Sardinia, andhas served to etch the landscape, testi-mony of the particular national vocationfor this tree variety.
The province of Cuneo is undoubtedlyone of the most important chestnut culti-vation areas in Italy, where the tree en-joys particularly suitable climate and soilconditions, and where the chestnut hasdeep cultural roots.
After years of decline in the chestnut cul-tivation field, due to a series of unfavou-rable circumstances of a pathological na-ture (ink disease and canker blight) andof a social-economic nature (the depopu-lation of the hillside and mountain areas);there has been a marked turnaround sin-ce the beginning of the 80’s with a consi-derable return of demand both for thewood and the nuts of the tree itself, bothfor fresh consumption and for the trans-formation industry.
Chestnut cultivation in the Cuneo pro-vince from past to presentIt is possible that the presence of thechestnut in the province of Cuneo can bedated back to thousands of years ago,but it is very probable that it spread to theAlpine valleys during the Roman era. Du-ring the period between the XIII and XIVcenturies, this species increasingly cameto replace the oak tree, a naturallygrowing tree which accounted for much ofthe original woodland. As from the XIVcentury the chestnut tree came to have aparticular local importance as testified bynumerous Communal Statutes referringto this tree (Bounous, 1999), which hadfor centuries constituted an important ele-ment in the local economy.
The chestnut tree currently covers over14% of the agricultural and forest lands ofthe province, the annual production rateof chestnuts and “marroni” (top qualitychestnuts) is of approx. 8,400 t. The localarea features an abundance of differentchestnut varieties suited to a wide rangeof requirements more than 50 differentvarieties were recorded, including boththe more well known varieties as well astypes which are now days only present incertain valleys (Bounous et al., 1988).
Each cultivar has its own specific meansof use and processing. In those areaswhich favour the production of early pro-duce, the chestnut of the Madonna or Ca-nalina in the Roereo district, the Selvas-china and the various Tempurive or Tem-pestive varieties which are grown in thewarmer parts of the Alpine valleys, arehighly prized and fetch good prices on theearly produce market.
In the Mondovì and Ceva areas, the localsmall chestnut varieties that are sweetand easy to peel (‘Gabbiana’, ‘Frattona’,‘Siria’, ‘Ciapastra’, ‘Rossastra’, ‘Spinalun-ga’) are traditionally dried to producedried (or white) chestnuts and flour.
Out of the Marroni and Marron types,which are similar and which are larger si-zed chestnuts that are specially selectedand used for prestigious processing forthe traditional pastry making industry andfor the confectionery industry, the Marro-

45FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
ne di Chiusa Pesio, as well as the RedGarrone and Black Garrone varieties areof particular note.
The typical nature of the chestnuts of theCuneo area is acknowledged in regula-tion 2081/92 of the European Union,which calls for the institution of a Protec-ted Geographic Indication (IGP) and forthe setting up of a Denomination of Pro-tected Origin (DOP) in order to protectthe products of the specific chestnutgrowing areas, based on the assumptionthat these products have superior senso-rial (aspect, fragrance and taste) as wellas nutritional features, thanks to the parti-cular environmental cultivation conditionsand the transformation techniques used.
The particular aspects which differentiatethe different varieties of the Province isbased on a complex blend of natural andanthropic features relating to the genoty-pe, the effect of the pollinizers on the fruitand the cultivation and conservation tech-niques used, as well as the pedo-climaticenvironment.
The genetic features of the various culti-vars are very important as they are theresult of the painstaking work of man inthe selection of the best tree types.Thanks to modern day technology it isnow possible to make a molecular analy-sis in order to identify the specific geneticidentity, which forms the “finger print” (fig.1) of each individual chestnut type (Mari-noni et al., 2000), this is a study currentlybeing carried out by the Department ofArboriculture of the University of Turin incollaboration with the Piedmont Regionand the IPLA (Institute for Woody Trees
and Environment). On the identification ofthe genotype of each cultivar it is thenpossible to establish its IGP, defining thequality and sensorial aspects typical ofthe variety, in order to define its origin.
The chestnut as a food in the pastThe chestnut was for centuries a staplefood for generations of mountain peopleand also constituted the food of rural po-pulations who turned to it in times of fami-ne and poverty. Its wood was used toheat the country dwellings, it providedtannin, litter and leaves for the livestock,raw materials for buildings, pole produc-tion and items of daily use.
As it provided a food of such basic impor-tance it was for centuries known as the“tree of bread”, and it spread to the mostdensely populated areas, extending be-yond its natural cultivation area, where itgrew and bore fruit only thanks to pains-taking care taken in tending the trees. Asthe cultivation gradually extended, it pro-vided an alternative to cereals, as a foodfor the masses, thanks to the fact that itwas easily available and easy to store.Later on thanks to its low cost, and highnutritional content it later became knownas the “bread of the poor”, providing ener-gy and protein to the poorer people. Inthe daily struggle for survival the poorlearnt how to use chestnuts in a variety ofways. In this way they were able to meettheir nutritional needs, if only just, andavoid hunger.
In past times in many alpine valleys, notonly in the Cuneo province, the averagediet was based on chestnuts for at least 4- 6 months of the year, and according to
Merz (1919) the pro-capita consumptionwas of around 150 kg/year. In a self-suffi-ciency economy chestnut growers oftenplanted different varieties of chestnuttrees to meet various requirements (fordrying, for flour making and for fresh con-sumption) (Conedera, 1996). Everythingplaced at disposal by nature, was trans-formed in a highly practical manner intohighly creative dishes, which formed thebasis of a subsistence diet. Great creati-vity was used in inventing various ways ofpreparing the chestnuts: roasted or boiledin water or milk, and eaten, especially inthe mountains, as a substitute for bread;served hot with wine or milk in the form ofa soup; they were grounded and wereused as substitutes for more costly cerealflours for the preparation of polenta, por-ridge, flat bread, chestnut breads andthick soups.
Nutritional valueChestnuts meet the current demand ofconsumers, who are increasingly tendingtowards actively organic foods, which areessential for healthy eating. Nutritionaland digestible foods which satisfy the de-mand for natural and wholesome pro-ducts, coming from areas which, espe-cially in the most favourable areas, do notrequire the use of any pesticides productsand in full accordance with organic far-ming principles. From a nutritional pointof view they are similar to rice or wheatand have therefore come to be termed“the grain that grows on a tree” (Burnett,1988).
The remarks on the dietary and nutritionalvalue which follow are based both on theinformation obtained from the available li-terature (tables 1 and 2) and on the basisof the results obtained from bio-chemicalanalysis undertaken on samples of twolocal cultivars which are clearly diversi-fied in terms of technological and marke-ting features but which are highly repre-sentative of the chestnut varieties in Cu-neo: the ‘Marrone’ (frozen chestnut sam-ples) traditionally used for the fine con-fectionery sector and for roasting, and the‘Gabbiana’ variety (dried chestnut sam-ple) a classic variety for drying and flourmaking (tables 3 and 4).
The starch was analysed by enzymaticmeans (Rasmussen et al., 1990; Boehrin-ger, 1983) while the sugars were ana-lysed for HPLC and the food fibre by en-zymatic means according to the AOAC985,29 method. In order to determine theprotein content, the nitrogen value wasanalysed according to Kjeldhal and multi-plied by 5.3 (McCarthy et al., 1988). Thefat content was calculated by gravimetryand the individual fatty acids by means ofgas-chromatography (Senter et al.,1994). The calcium, magnesium, potas-
Fig. 1 – The DNA profiles of the Red Garrone and the Marrone di ChiusaPesio varieties analyzed by micro-satellite loci ssrQpZAG15 (blue peaks)and ssrQpZAG110 (black peaks) by means of sequencer ABI-PRISM 377
(PE-Bio-systems). The two cultivars may be distinguished in terms of the molecularweight 120 bp and 220 bp, present only in the Red Garrone
(the molecular weights are expressed in pairs of bases).
100 120 140 160 180 200 220
220 bp
210 bpGARRONE ROSSO
MARRONE DI CHIUSA PESIO
210 bp
128 bp
120 bp
0
800
1600
2400
3200
4000
4800
5600
0400800
120016002000240028003200360040004400
128 bp

46 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
sium, phosphorus and iron content havebeen evaluated by means of plasmasource emission spectrometry.
Unlike the majority of pulp fruits (apples,peaches) the water content is quite mo-dest, and is of around 50% in the freshproduct, while in the dried chestnut it is ofaround 10%. Fresh chestnuts have a highcalorie content (160 Kcal for every 100gof edible product), a good fibre content(7-8%) an excellent sugary and amyl car-bohydrate content (about 35%), a fairquality protein content, a low percentageof fats and a moderate percentage of mi-neral salts (especially potassium, and toa less degree calcium, sodium and phos-phorus) and phytic acid, it also has a mo-dest content of hydrosoluble vitamins (B1and B2 in particular).
CarbohydratesChestnuts have a high carbohydrate con-tent (sugars and starch) which, as in allfruit in general, prevails over that of othernutrients (34 g average and 36.9 in the‘Marrone’ for each 100 g of fresh edibleproduct (tables 1 and 3). Starch is alsopresent in considerable quantity (24.4 per100 g average, 28.3 g per 100 g in the‘Marrone’), as are the sugars (8.1 g per100 g in the ‘Marrone’).
Sucrose (6.7 g per 100 g) is present ingreater concentration than in wheat, wal-nuts and potatoes, this is the main sugar,while glucose, fructose and maltose arepresent only in small quantities. The pre-sence of soluble carbohydrates makes
extensive storing difficult due to the pos-sibility of the growth of micro-organisms(fungi in particular), but curing the nuts inwater is a way of solving the problem(Giacalone et al., 1993).
Carbohydrates are an immediate andeconomical source of energy for the bodyand are converted into energy before pro-tein. The sucrose is hydrolysed into glu-cose and fructose, the latter being meta-bolised in the liver while glucose is onlypartly (10-30%). It is important to pointout that the energy consumption by thenervous system is based on glucose(120-150 g per day in an adult) (Fidanza,Liguori, 1981) conveyed by the blood andthat the glucose and the hepatic glycogenare not sufficient to meet the daily requi-rement (falls in glycemia levels are com-mon after strenuous physical activity).
Although glucose may be produced bythe body in other forms, it is recommen-ded that carbohydrates be assimilated as55-65% of the total energy intake, whilethe protein and lipid content should becontained to values of between 10-15%and 25-30% respectively (Fidanza, Liguo-ri, l.c.).
The carbohydrate content should mainlybe made up of polysaccharides (starch,which is digested in the form of glucosebefore being assimilated), thereby limitingthe consumption of simple sugars, sucro-se in particular, to not over 7-8% of thetotal energy input. Being very rich in car-bohydrates the chestnut is an excellent
energy source and is therefore ideal inconditions of physical and intellectualstrain, for those subject to stress and forthose involved in sporting activities whileit is not recommended for diabetics.
The attention given to the sugar contentof chestnuts has increased, because ithas come to be considered a valid alter-native food for children allergic to cow’smilk or lactose (Grassi et al., 1997).Chestnut flour is an ideal alternative inthe preparation of sweet products andsoups, providing the required carbohydra-te content for those individuals with ce-real intolerance.
The simple sugar content give the chest-nuts their sweet flavour, which is the pre-dominant organoleptic feature of freshfruit and which can also be appreciated inthe chestnut. Fruit with a predominanceof starch usually needs to be cooked tobring out the best in terms of taste andtexture.
FibreAs concerns the fibre content (7.3 g per100 g of fresh product), the insoluble part(6.7 g, Table1) prevails over the solublepart (0.6 g). It is responsible for the struc-ture of the proper seed, and therefore de-termines the consistency of the chestnut,which is important in assessing the accep-tability of the product.
Fibre is mainly made up of polysacchari-des that cannot be assimilated in thebody and considered as playing an impor-tant role on the function of the intestinesand on the micro-flora whilst also helpingto lower cholesterol. It helps to preventgastrointestinal problems by acceleratingthe transit of waste substances from theintestine and therefore it is recommendedin diets.
ProteinThe protein content (3.2 g per 100 g, 3 gper 100 g in the ‘Marrone’, Tables 1 and3) is equivalent to that of milk, althoughprolamin and glutenin (gluten progeni-tors) are absent; due to the absence ofthese substances it can only be madeinto bread if mixed together with cereal orrye flour to make mixed cereal and chest-nut bread. The protein contained is of ahigh quality as it contains amino-acids es-sential for the human body (tryptophan,lysine and the sulfurated amino-acids,methionine and cystine), and it is compa-rable to the protein content of eggs consi-dered ideal for amino-acid balancing(Burnett, l.c). In order to be used by thehuman body, a protein must contain allthe necessary amino acids in specificproportion; it is for this reason that chest-nuts excellently complete what is missingin vegetables.Range of hazelnut derived products
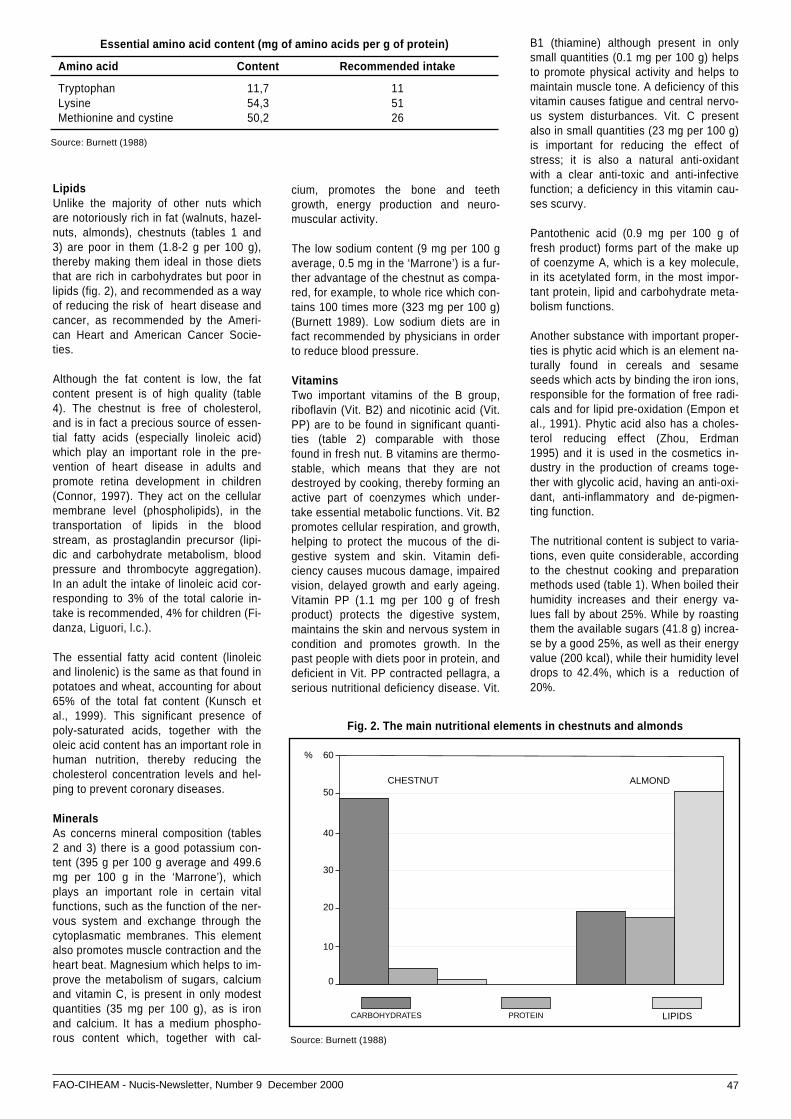
47FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
LipidsUnlike the majority of other nuts whichare notoriously rich in fat (walnuts, hazel-nuts, almonds), chestnuts (tables 1 and3) are poor in them (1.8-2 g per 100 g),thereby making them ideal in those dietsthat are rich in carbohydrates but poor inlipids (fig. 2), and recommended as a wayof reducing the risk of heart disease andcancer, as recommended by the Ameri-can Heart and American Cancer Socie-ties.
Although the fat content is low, the fatcontent present is of high quality (table4). The chestnut is free of cholesterol,and is in fact a precious source of essen-tial fatty acids (especially linoleic acid)which play an important role in the pre-vention of heart disease in adults andpromote retina development in children(Connor, 1997). They act on the cellularmembrane level (phospholipids), in thetransportation of lipids in the bloodstream, as prostaglandin precursor (lipi-dic and carbohydrate metabolism, bloodpressure and thrombocyte aggregation).In an adult the intake of linoleic acid cor-responding to 3% of the total calorie in-take is recommended, 4% for children (Fi-danza, Liguori, l.c.).
The essential fatty acid content (linoleicand linolenic) is the same as that found inpotatoes and wheat, accounting for about65% of the total fat content (Kunsch etal., 1999). This significant presence ofpoly-saturated acids, together with theoleic acid content has an important role inhuman nutrition, thereby reducing thecholesterol concentration levels and hel-ping to prevent coronary diseases.
MineralsAs concerns mineral composition (tables2 and 3) there is a good potassium con-tent (395 g per 100 g average and 499.6mg per 100 g in the ‘Marrone’), whichplays an important role in certain vitalfunctions, such as the function of the ner-vous system and exchange through thecytoplasmatic membranes. This elementalso promotes muscle contraction and theheart beat. Magnesium which helps to im-prove the metabolism of sugars, calciumand vitamin C, is present in only modestquantities (35 mg per 100 g), as is ironand calcium. It has a medium phospho-rous content which, together with cal-
cium, promotes the bone and teethgrowth, energy production and neuro-muscular activity.
The low sodium content (9 mg per 100 gaverage, 0.5 mg in the ‘Marrone’) is a fur-ther advantage of the chestnut as compa-red, for example, to whole rice which con-tains 100 times more (323 mg per 100 g)(Burnett 1989). Low sodium diets are infact recommended by physicians in orderto reduce blood pressure.
VitaminsTwo important vitamins of the B group,riboflavin (Vit. B2) and nicotinic acid (Vit.PP) are to be found in significant quanti-ties (table 2) comparable with thosefound in fresh nut. B vitamins are thermo-stable, which means that they are notdestroyed by cooking, thereby forming anactive part of coenzymes which under-take essential metabolic functions. Vit. B2promotes cellular respiration, and growth,helping to protect the mucous of the di-gestive system and skin. Vitamin defi-ciency causes mucous damage, impairedvision, delayed growth and early ageing.Vitamin PP (1.1 mg per 100 g of freshproduct) protects the digestive system,maintains the skin and nervous system incondition and promotes growth. In thepast people with diets poor in protein, anddeficient in Vit. PP contracted pellagra, aserious nutritional deficiency disease. Vit.
B1 (thiamine) although present in onlysmall quantities (0.1 mg per 100 g) helpsto promote physical activity and helps tomaintain muscle tone. A deficiency of thisvitamin causes fatigue and central nervo-us system disturbances. Vit. C presentalso in small quantities (23 mg per 100 g)is important for reducing the effect ofstress; it is also a natural anti-oxidantwith a clear anti-toxic and anti-infectivefunction; a deficiency in this vitamin cau-ses scurvy.
Pantothenic acid (0.9 mg per 100 g offresh product) forms part of the make upof coenzyme A, which is a key molecule,in its acetylated form, in the most impor-tant protein, lipid and carbohydrate meta-bolism functions.
Another substance with important proper-ties is phytic acid which is an element na-turally found in cereals and sesameseeds which acts by binding the iron ions,responsible for the formation of free radi-cals and for lipid pre-oxidation (Empon etal., 1991). Phytic acid also has a choles-terol reducing effect (Zhou, Erdman1995) and it is used in the cosmetics in-dustry in the production of creams toge-ther with glycolic acid, having an anti-oxi-dant, anti-inflammatory and de-pigmen-ting function.
The nutritional content is subject to varia-tions, even quite considerable, accordingto the chestnut cooking and preparationmethods used (table 1). When boiled theirhumidity increases and their energy va-lues fall by about 25%. While by roastingthem the available sugars (41.8 g) increa-se by a good 25%, as well as their energyvalue (200 kcal), while their humidity leveldrops to 42.4%, which is a reduction of20%.
Essential amino acid content (mg of amino acids per g of protein)
Amino acid Content Recommended intake
Tryptophan 11,7 11Lysine 54,3 51Methionine and cystine 50,2 26
Source: Burnett (1988)
Source: Burnett (1988)
Fig. 2. The main nutritional elements in chestnuts and almonds
0
PROTEIN LIPIDS
10
20
30
40
50
% 60
CARBOHYDRATES
CHESTNUT ALMOND
48 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Table 3. Nutritional features of two Cuneo chestnut cultivars
Marron ‘Gabbiana’(fresh frozen chestnut) (dried chestnut)
Water % 47.1 10.3Carbohydrates (g/100 g) 36.9 61.4Sugars (g/100 g): 8.6 22.4
Sucrose (g/100g) 6.7 21.0Glucose (g/100g) 0.9 0.5Fructose (g/100g) 0.6 0.6Maltose (g/100g) 0.4 0.3
Starch (g/100 g) 28.3 39.0Food fibre (g/100 g) 8.2 13.6Proteins (g/100 g) 3.0 5.3Lipids (g/100 g) 2.1 3.5Potassium (mg/100 g) 499.6 1022Phosphorus (mg/100 g) 72.8 137Calcium (mg/100 g) 23.2 58Iron (mg/100 g) 0.9 1.5Sodium (mg/100 g) 0.5 1
Table 1. Composition and nutritional value of chestnuts (per 100 g of product) according to the means of storing and use
fresh dry roasted boiled flour LARN
EDIBLE PART % 81 100 82 88 100WATER % 52,9 10,1 42,4 63,3 11,4CALORIES (kcal) 160 287 200 120 343 2900 (2150)NUTRITIONAL ELEMENTScarbohydrates (g/100 g) 34,0 57,8 39 24,4 63,6 522 (413) gsugars (sucrose, glucose, fructose, maltose) (g/100 g) 9,6 16,1 10,7 7,5 23,6starch (g/100 g) 24,4 41,7 28,3 16,9 40food fibre (g/100 g) 7,3 13,8 8,3 5,4 14,2*soluble (g/100 g) 0,6 1,1 0,7 0,6 1*insoluble (g/100 g) 6,7 12,7 7,6 4,8 13,2*proteins (g/100 g) 3,2 6 3,7 2,5 6,1 62 (53) glipids (g/100 g) 1,8 3,4 2,4 1,3 3,7 95 (73) g
Table 2. Mineral and vitamin composition of the chestnut according to the means of storing and use
MINERAL ELEMENTS fresh dry flour LARN
Potassium (mg/100 g) 395 738 847 3100 mgPhosphorous (mg/100 g) 70 131 164 800 mgSulphur (mg/100 g) 48** 126** 126Magnesium (mg/100 g) 35 74 350 mgCalcium (mg/100 g) 30 56 50 800 mgChloride (mg/100 g) 10** 18.6** 18Sodium (mg/100 g) 9 17 11Iron (mg/100 g) 1 1.9 3,2 10 (18) mgManganese (mg/100g) 0.7** 1.3** 1,3 4 mgCopper (mg/100g) 0.6** 0.6** 0,6 1,2 mgZinc (mg/100g) - 0.3** 0,3 10 (7) mgVITAMINSVit. B1 thiamine (mg/100g) 0.1 0.2 0,2 1,2 (0,9) mgVit. B2 riboflavin (mg/100g) 0.3 0.4 0,4 1,6 (1,3) mgVit. PP nicotinic acid (mg/100g) 1.1 2.1 1 18 (14) mgVit. C ascorbic acid (mg/100g) 23** - - 60 mgPantothenic acid (mg/100 g) 0.9** - - 5 mgPhytic acid (mg/100 g) 50 - -
Sources: taken from the Food Composition tables INN 1997, integrated with the data* of Brighenti et al., 1998, and data* from the Institut Scientifique d’HygièneAlimentaire, Paris 1974; Panatta, 1999.LARN = Recommended Daily Nutrient Intake for the Italian Population – referring to men or women (in brackets) of between 30-49 years of age, with a bodyweight of 65 and 56 kg, respectively (Human Nutrition Association, 1996).
Cooking alters the starch content, whichis reduced on boiling with a reduction inthe potassium and magnesium content,but not in calcium, while the sucrose, lipidand protein content are scarcely altered(Kunsch et al., 1999).
On drying (Tables 1-3) the protein con-tent further increases (5-6%) which ismore than that found in the potato (2%) orin other starch based products, but muchless than that present in cereals (10-12%)or in dried vegetables (20-25%), (Pa-natta, 1999), the carbohydrate contentalso increases reaching a value ofapproximately 60 g per 100 g of ediblematter. Dried chestnuts have a modestsodium content (17 mg per 100 g of edi-ble matter, 1 mg per 100 g in the ‘Gabbia-na’ variety), with a modest content of vita-mins of the B group, iron (1.5-1.9 mg per100 g), calcium (a little under 60 mg per

49FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Table 4. Fatty acid content of two Cuneo chestnut cultivars
Marron ‘Gabbiana’
% %
Myristic acid C14:0 0,2 0,1Palmitic acid.C16:0 19,5 15,7Palmitoleic acid C16:1 0,5 0,3n-heptadecanoic acid C17:0 0,1 0,1Stearic acid C18:0 1,4 1Oleic acid C18:1 44 45Linoleic acid C18:2 28,2 32,5Linolenic acid C18:3 2,6 3,1Eicosanoic acid C20:0 0,5 0,4Eicosanoic acid C20:1 0,8 0,8Docosanoic acid C22:0 0,6 0,3Lignoceric acid C24:0 0,2 -
100 g) and potassium (738 mg per 100 gaverage and 1022 mg per 100 g in the‘Gabbiana’ variety) which are clearly res-pectable values. While in chestnut flour(tables 1 and 2) the fibre content is verysignificant (14.2%), and clearly prevails(over 90%) over the insoluble portion. To-gether with the increased sugar and pro-tein content, the energy value also in-creases, which in fact reaches 343 kcal.
Advantages in modern day eatinghabitsDue to the organic value of the chestnutand the dietary-nutritional characteristicsmentioned, today it can once again makea comeback in our daily eating habits,free of connotations of poverty with whichit has been linked for centuries. Thechestnut has acquired a new standingdue to the need to restore traditional va-lues and the demand for truly wholesomefoods. This true gastronomic food reservetherefore has two faces, on the one side ithas strong links with past traditions, onthe other it takes the form of a food idealfor today’s healthy eating trends.
Although the chestnut has tended to befeatured as an ingredient for sweets anddesserts in the cookery books, the chest-nut can also be used in the preparation ofstarters, first and second course dishes.
The centuries of experience in the Cuneoarea in the preparation of dried chestnutand chestnut flour, used for all kinds ofsoups and polenta makes them traditionaldishes which are highly appreciated to-day. Its use in the preparation of tagliate-lle, gnocchi and ravioli is becoming in-creasingly common. Dried chestnuts areboiled in little water and served hot withlocal cured pork meats. Whether they areserved whole, boiled, stewed and espe-cially roasted, they make an excellentside dish or delicious ingredients for sa-lads. They are also delicious served withvarious types of meat (capon, turkey,pork, goose, rabbit) and often used forstuffing.
Thanks to the abundant sugar content ofthe nut, for centuries it has been used inthe preparation of refined desserts andsweets such as marron glacés, mousse,soufflé, creams “Bavaresi” (specialitycream based pastries) and ice-creams.The taste however is such to make themideal for use in less elaborated dessertssuch as “castagnacci” (chestnut flourbread), “necci” (type of savoury chestnutflour bread), fritters and milk- basedpuddings. The special type of dried chest-nut known as “biscotti” and “viette” is alsoworth a mention, being typical local pre-parations of the Cuneo area, mentionedby Remondino in his historical-statisticalstudy on chestnut cultivation in the pro-
vince of Cuneo at the start of the last cen-tury, which make a wholesome delicacyavailable throughout the year.
Just as in the past, roast chestnuts or“ballotte” (chestnuts cooked in water aro-matised with fennel seeds), washed downwith a glass of good red wine (whetheryoung or aged) create a great convivialatmosphere during autumn afternoons inthe open air or on cold winter eveningsaround the fire.
CONCLUSIONSBesides having a delicious taste, and be-ing so versatile as to find use in a varietyof gastronomic preparations (rangingfrom first to main course dishes as well asvegetable dishes, desserts and pastries),chestnuts also are a very healthy andhigh energetic food.
Thanks to their nutritional composition:with a low fat content, completely free ofcholesterol, with a low sodium and highpotassium content, and a moderate, buthigh quality protein content (thanks to thefavourable amino-acid make up), chest-nuts are a balanced and quality food.
The high carbohydrate and potassiumcontent make it a high energy food, withmineral restoring, anti-anaemic and anti-septic properties, being highly valuable insituations of physical or intellectual strain,excellent for people under stress and forthose who do sports. A delicious energybooster.
BIBLIOGRAPHY
Boehringer 1983. Starke (UV test). In:Methoden der enzymatischen Lebens-mittelanalytic, Boehringer MannheimGmbH, Deutschland, 75-77.
Bounous G., Barrel A., Pividori M., 1997.Stato attuale della castanicoltura nelle re-
gioni dell’Arco Alpino ed in Liguria. Attidel Convegno nazionale sul castagno, Ci-son di Valmarino (TV): 41-56.
Bounous G., Giacalone G., 1992. Ches-tnut storage, processing and usage inItaly. World Chestnut Conference, Mor-gantown, WV, (USA): 105-131.
Bounous G., 1999. Tra i castagni delcuneese. Ed. Metafore, Cuneo, pp. 80.
Bounous G., Giacalone G., MondoM.,1998. Situazione attuale e problema-tiche della castanicoltura da frutto cu-neese. Monti e boschi, 14:5-12.
Brighenti F., M. Campagnolo, D. Bassi,1998. Biochemical characterization ofthe seed in instinct chestnut genotypes(C. sativa). Abstracts 2nd Int. Symp. onchestnut, Bordeaux.
Burnett M.S., 1988. The grain thatgrows on a tree. Reprinted from “Chest-nutworks”. Portland, Oregon:12-15.
Conedera, 1996. Die Kastanie, derBrotbaum. Bundnerwald 49 (6): 28-46.
Connor W.E., 1997. The beneficial effectsof omega-3 fatty acids: cardiovascular di-sease and neuro-development. Currentopinion in Lipidology, 8:1-3.
Empson K.L., Labuza T.P., Graf E.,1991. Phytic acid as food antioxidant.Journal of Food Science; 56: 560-563.
F.A.O. Sito web: http://fao.org
Fidanza F., Liguori G.,1981. Nutrizioneumana. Ed. Idelson, Napoli.
Giacalone G., Bounous G., 1993. Tradi-zioni ed innovazioni nella trasformazio-ne e nell’utilizzo delle castagne. Monti eboschi, 5:33-41.
Grassi G., Mastronicola M., Parente A.,1997. Atti del Convegno nazionale sulcastagno, Cison di Valmarino (TV): 575.Il tempo delle castagne; www. marrone.net/iltempodellecastagne
INN, 1997. Tabelle di composizione de-gli alimenti.

50 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Kunsch U., Scharer H., Patrian B., Hur-ter J., Conedera M., Sassella A., Jermi-ni M., Jelmini J., 1999. Quality assess-ment of chestnut fruits. Proc. 2nd Int.Symp. on chestnut. Acta Hort. 494: 119-127.
Marinoni D., Boccacci P., Akkak A.,Bounous G., Ferrero A., Botta R., 2000.Impiego di marcatori microsatelliti diquercia per lo studio del germoplasmapiemontese di castagno (C. sativa Mill.).Atti giornate scientifiche SOI, Sirmione,Brescia.
Mc.Carthy M.A., Meredith F.I., 1988.Nutrient data on chestnut consumed inthe United States. Economic botany,42: 29-36.
Merz F., 1919. Die Edelkastanie: Ihrevolkswirtschaftliche Bedeutung, ihrAnbau und ihre Bewirtschaftung. VerlagSchw. Departement des Innern, Bern.71 S.
Muti E., 2000. Il cibo e il nostro corpo eil cibo e il nostro peso, ovvero la Bibbiadelle calorie, Demetra, pp. 495
Panatta G.B., 1999. Un frutto energeti-co e gustoso. Il Divulgatore, Bologna,XXII, 10: 73-76.
Rasmussen T.S., Henry R.J., 1990.Starch Determination in Horticulturalplant Material by an Enzymatic-Colori-metric procedure. J. Sci. Food Agric.52:159-170.
Senter S.D., Payne J.A., Miller G.,Anagnostakis S.L., 1994. Comparisonof total lipids, fatty acids, sugars andnonvolatile organic acids in nuts fromfour Castanea species. J. Sci. FoodAgric. 65:223-227.
Zhou J.R., Erdman J.W., 1995. Phyticacid in health and disease. critical re-views in food. science and Nutri-tion.35.495-508.
G. BounousR. Botta, G. Beccaro
Department of Arboreal CultivationUniversity of Turin
Via Leonardo da Vinci, 4410095 Grugliasco (TO)
ItalyTel. ++39 011 6708657 - Fax ++39 011
6708658E-mail: [email protected]
THE STONEPINE(Pinus pinea L.) BREEDING
PROGRAMME INCASTILE-LEON
(CENTRAL SPAIN)
THE STONEPINE IN CASTILE-LEONThe stonepine, Pinus pinea L., is one ofthe most characteristic tree species of theMediterranean Basin due to its singularumbrella shape. Since the antiquity, it has
been part of the landscape and was usedas nut tree, since its cones produce largeedible kernels, known in Spanish as “pi-ñones”.
In Spain, the stonepine forests coversome 400,000 ha, two thirds of theworld’s total stonepine area; 176,000 haof them have been deemed as naturalstands. Main growing areas are theSouthwest (Western Andalusia) and the“Meseta Norte” (central highland plateauin Castile-Leon), but pure or mixed stone-pine stands spread also over other areasin Catalonia, Castile La Mancha, Madridor Extremadura. In order to regulate theuse of forest reproductive material, sevenprovenance regions and four limited localareas have been defined in Spain, basedon geographic and ecological criteria(Prada et al., 1997).
In Castile-Leon there are two provenanceregions. Table 1 exposes a short ecologi-cal characterization of the two provenan-ce regions and Figure 1 shows their sto-nepine areas. “Meseta Norte” (numberES-23/01) includes the 50,000 ha in thecentral Douro Basin, the main part beingnatural stands. They are part of the “Tie-rra de Pinares” (Pine Country) betweenValladolid, Segovia and Avila. This coun-try is a vast, sandy plain covered with170,000 ha of mixed or pure maritimepine (Pinus pinaster AIT.) and stonepineforests. Stonepine dominates in the pro-vince of Valladolid, where it is the mainforest tree species and occupies 42% of
the total forest area of the province (Spa-nish Forest Resource Survey 1995). Thegeographic region “Meseta Norte” is a se-dimentary plain surrounded by highmountain ranges, which separate it fromthe mild oceanic influence. Therefore theclimate is continental with marked hot drysummers and long winters. Occasionallate frosts can occur up until May or Juneand early frosts from October. The distri-bution of rain is irregular as much as be-tween years as between seasons, exceptthe three months of summer drought.
Since pre-Roman ages, the predominantland use has been for dry farming agricul-ture, with grain as main crops. In the cen-tral province of Valladolid, 80% of theland surface is cultivated, and only 13%remains as forests which have not beenbroken up. This is mainly because of itsminimal agronomic aptitude. Specifically,the pine forests are relegated to sandyareas in the south of the Douro valley -authentic dune areas -, to calcaric litho-sols and to clayey or gypseous marls inthe easily eroded slopes.
The second provenance region in Castile-Leon (ES-23/02) Tietar and Alberche Val-leys, spreads over the limit areas of theprovinces of Avila, Madrid and Toledo.They are situated in the southern foothillsof Gredos and Guadarrama mountainranges in a genuine Mediterranean clima-te. The main function of these forests issoil, wildlife and landscape conservation.This is due to their situation on hills and
Figure 1. Stonepine areas in Castile-Leon.ES-23/01: Meseta Norte, ES-23/02: Tietar and Alberche Valleys

51FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
mountain slopes and the presence of en-dangered or endemic species such asIberian lynx (Lynx pardina), Iberian eagle(Aquila adalberti) or black vulture (Aegy-pius monachus).
In former times, the pine forests providedthe local population with a large numberof resources, documented since the 12th
century (Gil, 1999). Nowadays, the mainfunctions of the remaining forests are de-fined as environment protecting and con-servation of the ecosystem, its biodiversi-ty and its underlying genetic diversity. Atthe same time, the open and luminousstonepine forests are the preferred outingand picnic areas for the urban population.The cone yield is currently the most im-portant commercial production of the sto-nepine forests and provides averageyearly incomes of 24 euros per hectare1
to the owner (in the most productivestands up to 96 euros/ha yearly), whe-reas the timber sale comes only in se-cond place (15.5 euros/ha annual). Othermarketable products and uses, as gra-zing, fuel wood or hunting, which are mi-nor.
The local pine kernel industryThe increasing economic importance ofthe pine kernel production results in sus-tained incomes for the pine forest ow-ners. Also the regional kernel market hasconsolidated in the village of Pedrajas deSan Esteban (Valladolid). The averagevalue of stonepine cones sold in tree1 ismore than 400,000 euros only for the pu-blic forests of Valladolid province, thethird part of “Meseta Norte” stonepinestands. The pine-kernel shelling and manu-facturing industry in the “Tierra de Pinares”area includes about seventy companies,with a turnover of 12,000,000 euros andprocessing 1,500 to 2,500 t of unshelledkernels originated from Castile-Leon, Anda-lusia, Castile La Mancha and Portugal.
Grafted stonepine plantationsAmong the nut trees, the stonepine re-mains as being one of the most genuineforest species, as it was never domesti-cated in cultivated plantations like othercrops. At present the cones are still har-vested in forest stands. The difficulties ofits cultivation are one of the possible rea-sons of this lack of Neolithic revolution.
The stonepine does not root as cutting,nor does it allow woody grafting. Scionsshould be obtained from long shoot termi-nal buds at the very moment of initiatingits spring flush, or from other soft tissueslike needled dwarf shoots. Furthermore,the pruning possibilities are limited, asthe pine does not form long shoots fromaxilar buds, but only from the terminalbuds whirl at the top of the branches.Another point against the stonepine culti-vation is the three-year-long wait for ma-turation of the cones. This implies thatthree consecutive yields coincide in thetree, so a very irregular fruitfulness be-tween years results. Finally, female cone-lets and male catkins are situated separa-tely in different parts of the tree’s crown.During the first 10-15 years the graftedstonepines behave like female individualswithout male flowers, and need to grownear to mature stands that act as pollina-
tors. In conclusion, the stonepine is notan easy crop.
But perhaps the main reason to discardstonepine as cultivated crop has been thereduced kernel output. Some 200 kg ofcones, average harvest per hectare of na-tural stands, will give some 40 kg of shel-led kernels, and only 10 kg of unshelledkernels. Furthermore, until the first steamdriven shelling machine was designed atthe end of 19th century, kernel shelling one-by-one was a manual labour of women andchildren. So the costs of kernel harvesting,transport, extraction and shelling made thepine kernel more a pastries ingredient orluxury speciality than a genuine food thatcould match up with other crops’ efficiencyas interesting farmland use.
Actually, there is not any report about theuse of grafted stonepines before the
Table 1. Provenance region of Pinus pinea L. in Castile-Leon (Prada et al., 1997)
Provenance Region Altitude (m) Climate variables (range) Lithology Stonepine AreaT(ºC) P(mm) sp rock pH (approx. Ha)
1. Meseta Norte 650 - 900 10,5-13,5 351-610 2 - 16 50.000
2. Tietar and Alberche Valleys 600-1000 13,2-15,1 648-1007 7 - 12 s 5,7-7,0 3.500
T = temperature (ºC); P = rainfall (mm); sp = minimal monthly rainfall in summer (mm); s = siliceous parental rock; k = limestone rock.
s 6,4-7,0k 7,6-8,8
Stonepine clone growing at the “Meseta Norte” genebank
52 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
1960th, apart from local use in the villageof Biar (Alicante), where grafting stonepi-ne on wild grown P. halepensis roots-tocks (Cavanilles, 1797, cit. in Prada etal., 1997). Since the 60th, Italian breedershave carried out grafting experiments, topropagate selected stonepine clones, andfew years later the experimentation star-ted in Spain too.
Thirty years of experiences later, Pradaet al. (1997) and Catalán (1998) expo-sed the currently known advantages,possibilities and yields of grafted stone-pine plantations. Here we only emphasi-ze as the main advantage of grafting innew stonepine plantations, the early co-ming in production at few years2. Oppo-sited to it, traditional stonepine standsdo not start to yield cones until abouttwenty or thirty years later. The econo-mic viability and profitability of theseplantations is analysed by Abellanas etal. (2000) and Mutke et al. (2000). Ba-sed on data of experimental plantation
(Figure 2), the results of these papersshow that the potential use of graftedstonepine can be a very interesting al-ternative in farmland afforestations.
General lines of the stonepine bree-ding programme in Castile-LeonAs a consequence of the change in pro-fitability of the different products of pineforest, currently timber yield comes intosecond place. Cone yield is the mainmarketable production of the forest, ma-naged under the paradigm of protec-tions and multiple use. Therefore, theimprovement of the stonepine will haveto combine three objectives. Genetic re-source conservation in the silvicultureand management of the existing naturalregenerated forests, specific treatmentsfor cone yield improvement in the mostproductive stands, and a defined bree-ding line orientated to obtain and propa-gate high productive genotypes for theiruse in new plantations and farmlandafforestations.
The first two items result in sustainablesilviculture and management guidelines.The third is the aim of the StonepineBreeding Programme in Castile-Leon.The programme is the frame of variousworks started in the 90th as cooperationbetween the regional forest administra-tion of Castile-Leon and the ForestrySchool (Polytechnic University of Ma-drid). The main topics of this programmewere exposed by Catalán in the Nucis-Newsletter number 7 (1998); the current re-sults and prospects are presented below.
The genetic improvement of stonepinepoints at two different purposes. Firstly,to meet the seed demand for afforesta-tions with the selection and delimitation ofseed stands to yield selected seeds, cer-tified under EU and OECD regulation. Se-condly, to search plus trees for vegetativepropagation in specific plantations.
The necessity of seed stand delimitationis based on the traditional importance ofstonepine, used in the afforestations per-formed during the 20th century, on degra-ded and eroded soils. The plantations ofstonepine are justified by its low ecologi-cal requirements, easy nursery growing,high drought resistance and the commer-cial value of the expected cone yields. Atpresent, since European reforestationprogrammes of farmland started in the90th, stonepine has become one of themost widely used species. It is the firstone in the “Meseta Norte” region andfourth in the whole of Castile-Leon, with11.836 new hectares planted between1993 and 1997, a 20% increment of itsformer area. The aim is to achieve thebest future forests in every site, includingeroded or gypseous soils. This requiresthe use of certified seeds obtained in se-lect stands with good phenotypes, adap-ted to each specific soil class and with ahigh cone production.
As basic short-term strategy, ten standswere identified and delimited since 1993(Table 2) in order to obtain enough seedquantity, guarantying a broad genetic ran-
2500
2000
1500
1000
500
0
0 5 1510
a b c d e f g
Con
e yi
eld
(Kg/
ha)
Figure 2. Review of cone yield reports in grafted plantations (calculated for 278 trees per hectare)
a - Monte Tudia y sus Faldas, Badajoz (Garrote, 1994, cit. in Mutke et al., 2000)b - Clone bank, Italy (Magini & Giannini, 1971, cit. in Mutke et al., 2000)
c - Pezuela de las Torres, Madrid (Catalán & Catalán, 1996, cit. in Mutke et al., 2000)d - El Carrascal, Valladolid (Gordo et al., 2000)e - Valdegalindo, Valladolid (Gordo et al., 2000)f - Puerta de Hierro, Madrid (Sada et al., 2000)
g - Cabeza Aguda, Cordoba (Abellanas et al., 2000)
Table 2. Proposal of Selected Stonepine Seed Stands in Castile-Leon
Provenance Region Code Township (province) Elevation (m) Longitude Latitude Area (ha)
1 Meseta Norte ES-23/01/001 Iscar (Valladolid) 750 4º31’W 41º21’N 360ES-23/01/002 Tordesillas (Valladolid) 680 4º57’W 41º30’N 106ES-23/01/004 La Parrilla (Valladolid) 855 4º34’W 41º33’N 20ES-23/01/005 Portillo (Valladolid) 850 4º32’W 41º28’N 59ES-23/01/006 Cogeces de Iscar (Valladolid) 800 4º57’W 41º25’N 22ES-23/01/007 Toro (Zamora) 680 5º27’W 41º31’N 45ES-23/01/008 Quintanilla de O. (Valladolid) 780 4º18’W 41º37’N 50
2 Tietar and Alberche ES-23/02/001 El Hoyo de Pinares (Avila) 900-1000 4º21’W 40º31’N 26Valleys ES-23/02/002 El Hoyo de Pinares (Avila) 720-750 4º20’W 40º28’N 8
ES-23/02/003 Cebreros (Avila) 600 4º22’W 40º25’N 75

53FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
ge in the seed sources, although withoutan experimental evaluation of its superio-rity.
The second line of the breeding program-me started in 1989 with the fieldwork ofplus tree selection, centred on high pro-ductive phenotypes, by annual measureof the individual trees’ kernel yield insampled stands. From the 135 sampledtrees, grafted scions of 116 are includedin clone banks. Of these, 35 are selectedas plus trees by their cone yield superio-rity. At present, studies in the grafted clo-ne banks are performed to characterizeindividual kernel yield, flowering and ma-turing phenology and cone and kernelbiometry.
Comparative testsIn order to analyse the variability betweendifferent stonepine provenances or seedsources in their performance in afforesta-tions, various field trials have been esta-blished in different Mediterranean coun-tries. In 1995, the FAO Silva Mediterra-nea Network established an internationalfield test with 35 provenances from sevenMediterranean countries, 17 of them fromSpain. Four trial plots where planted inSpain. Castile-Leon took part with one ofthe plots in Tordesillas (Valladolid) andhas seven seed sources, six of them pro-posed as selected seed stands. In the fu-ture, the information obtained in this trialis expected to allow the comparison ofthe genetic variability between provenan-ces. Also it will allow to recommend thebest seed sources for afforestations ineach site (Martín & Prada, 1995).
Within the “Meseta Norte” provenance re-gion, the evaluation of genetic variabilityin reproductive material of seed sourcesis performed in another trial. Seven expe-rimental plots were established, whichrange all potential stonepine afforestationsite types -from genuine sandy pìne-fo-rest soil to clayey farmland and erodedmarls-. In this test nine seed sourceswere represented, including the proposedselected seed stands. The objective ofthis comparative trial is to define testedseed stands in accordance with European(EU, OECD) and National legal standard.It should take into account the fitness(quantitative and adaptive traits, as survi-val, vigour, growth, adaptation to specialsoil conditions, biotic and abiotic risk re-sistance) and cone yield superiority.
Finally, the grafted plantations are usedas test plots to estimate the genetic andenvironment control of flowering, fruitful-ness and kernel yield. The objective wasto test the pre-selected plus trees in thehomogenous environment of cultivatingand to select the best genotypes for con-trolled pollination.
View of the stonepine clonal bank “Meseta Norte”
Close up of a stonepine clone showing cones
Summary of performed laboursand results1. Seed stand selection(a) Ecological and phenotype characteri-zation of stonepine forests in Castile-Leon.(b) Delimitation and characterization ofrepresentative stands of each definedtype of stonepine forest.
(c) Based on this study, seven selectedseed stands in the Meseta Norte regionand three selected seed stands in theTietar and Alberche Valleys region havebeen proposed to provide selected seedfor afforestations. Currently, the catalo-gue of ten defined seed stands in Castile-Leon is in the approval phase in the Mi-nistry of Environment.

54 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
(d) Publication and popularisation of the-se topics to raise the conscience of po-tential users (foresters, nurserymen)about the advantage to use the best avai-lable seed sources.
2. Plus tree selection for vegetative re-productive material (graft scions)(a) Preselection of good cone producingphenotypes (candidates) in natural stone-pine stands in the Meseta Norte pine fo-rests and evaluation by their kernel yield.
• 1989: selection of 100 candidate treesin 20 sample plots, with kernel yield mea-surement during a period of 8 years(1990-1997).
• 1991: installation of the first clonal bank“El Carrascal” with all 100 candidates.This grafted plantation is a clone archiveto produce scions for future grafting.
• 1995-1996: selection of other 35 candida-tes in 7 new sample plots, with the objecti-
ve to range the sample population over allstonepine growing sites in the region.
• In the results of the yield sampling, adependence of the individual kernel pro-duction on the size and the growing spa-ce of the tree are observed, thus the se-lection criterion was the unshelled kernelyield referred to these two control para-meters. Finally, 60 of the 135 pre-selec-ted trees were selected to be propagatedin the second clone bank. Actually, twotwin plots were installed, one in Tordesi-llas, (Valladolid), the other in Puerta deHierro, (Madrid).
(b) Studies in grafted plots.
• Individual yield measurement in “El Ca-rrascal” clonal bank. On account of initialcultivation problems, a delay of growingand production was observed (Table 3).
• Growing and flowering phenology of sto-nepine in the “Meseta Norte” as main sto-
nepine region in inland Spain has beenstudied in various plots.
• Controlled pollinations were started toobtain full-sib progenies for further trials,such as progeny tests. The disadvantageof stonepine is the three-year wait for ma-ture seed.
PROSPECTSIn the seed stand selection line, the resultsof the current comparative trial will offer thepossibility to select the best for each affo-restation site in the Douro Basin. Its seedswill be included in the official category oftested forest reproduction material.
In the clone selection line, the aim to testthe grafted genotypes in homogenous con-ditions makes standarization of rootstocks,grafting and cultivation necessary. Current-ly, microcuttings are being developed fromjuvenile shoots to obtain clonal rootstockswhich would allow “self-grafting” of the tes-ted clones or the use of standard roots-tocks in clone trials. The disadvantage ofthe cutting technique is the need of rejuve-nation because of the rooting incapability ofmature tissues in stonepine. Therefore it isnot possible to obtain mature cuttings fit-to-test with early cone yields.
On the contrary, an accelerated matura-tion of propagules derived from cotyle-dons via tissue culture has been reportedin plantlets of various pines and otherspecies (M.S. Greenwood, personal com-munication). The organogenesis from co-tyledons of half-sib or full-sib seedlingswould allow premature progenies test andbreeding value estimation of the testedprogenitors. Furthermore, if the fructifica-tion of premature plants performs proper-ly, it opens a way to obtain breeding ge-nerations in shorter period of time.
NOTES1 Values (without VAT) for nearly 16,000ha of public stonepine forests in the pro-vince of Valladolid 1990-1998. The yearlycone yield rights of every pine stand is
Table 3. Cone yields in grafted stonepine plantations in Castile-Leon
Years after grafting “El Carrascal” (278 trees/ha) “Valdegalindo” (600 trees/ha)
kg/ha Average (kg/tree) kg/ha Average (kg/tree) Average of 20 plus tree
4 58 0.215 183 0.666 149 0.537 157 0.578 136 0.499 460 * 1.65 *
10 483 * 1.74 * 1,596 2,6 11,411 728 * 2.62 * 1,892 2,9 4,312 211 0,8 0,9
(*) Predicted values of flowering and maturing cones.First 9 years in the older “Valdegalindo” plantation without yield information.
Stonepine needle grafting

55FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
sold at auction, based in appraisals of thecones still on the trees. The harvesting la-bours are performed by the purchaser, sothe auction price is a direct forest owner’sincome.
2 In the presence of nearby grown-up sto-nepines as pollinators.
REFERENCES
Abellanas, B., Butler, I., Monteagudo,F.J., 2000: Estudio de la rentabilidadeconómica de una parcela de injertos depino piñonero a los nueve años de su ins-talación. Actas (II) del Primer Simposiodel Pino Piñonero (Pinus pinea L.) febre-ro 2000, Valladolid: 101-110.
Catalán, G., 1998: Current Situation andProspects of the Stonepines as Nut Pro-ducer. FAO - Nucis-Newsletter, number 7December1998. IRTA, Reus: 28-32.
Gil, L., 1999: La transformación históri-ca del paisaje: la permanencia y la ex-tinción local del pino piñonero. En: Ma-rín, F.; Domingo, J. & Calzado, A. (edi-tors): Los montes y su historia – unaperspectiva política, económica y so-cial. Universidad de Huelva: 151-185.
Gordo, J.; Mutke, S., Gil L., 2000: Lamejora genética de Pinus pinea L. enCastilla y León.. Actas (II) del PrimerSimposio del Pino Piñonero (Pinus pi-nea L.) febrero 2000, Valladolid: 21-32.
Martin S., Prada A., 1995. Ensayo deprocedencias de pino piñonero (Pinuspinea L.) en España en: El pino piñone-ro como árbol productor de frutos secosen los países mediterráneos. I Reuniónde la Red de Frutos Secos de la FAOsobre el Pino Piñonero. INIA- CIFOR.Madrid: 117-121.
Mutke, S.; Díaz-Balteiro, L., Gordo, J.,2000: Análisis comparativo de la renta-bilidad comercial privada de plantacio-nes de Pinus pinea L. en tierras agra-rias de la provincia de Valladolid. Inves-tigación Agraria- Sistemas y RecursosForestales Vol. 9(2) 2000 (in press).
Prada M.A., Gordo J., De Miguel J., Mut-ke S., Catalán G., Iglesias S., Gil L.,1997. Las regiones de procedencia de Pi-nus pinea L. en España. Organismo Au-tónomo de Parques Naturales. Madrid.
Sada, B.; Iglesias, S., Gil, L., 2000: Estu-dio de producción en un Banco Clonal dePinus pinea L. de procedencia CataluñaLitoral para la selección de grandes pro-ductores de fruto. Actas (II) del PrimerSimposio del Pino Piñonero (Pinus pineaL.) febrero 2000, Valladolid: 65-73.
S. Mutke*, J. Gordo** and L. Gil** Unidad de Anatomía, Fisiología y GenéticaForestal. ETS Ingenieros de Montes (UPM)
28040 Madrid.**Servicio Territorial de Medio Ambiente. Junta
de Castilla y León. Duque de la Victoria 5.47001 Valladolid
E-mail: [email protected]
POLLEN TUBE GROWTH ANDFERTILIZATION IN CAROB
(Ceratonia siliqua L.)
INTRODUCTIONThe present contribution is part of a four-year study that focused on flowering andfruiting of carob trees at different sites inPortugal (Haselberg 2000). Among thefindings was the fact that only less than60 % of the ovules in the pistils develo-ped into seeds as a result of either failureof seed set or malformation respectivelyabortion of seeds during development.
High seed contents is essential for goodcrop quality in carob: seeds (kernels)constitute 10 % of the pod weight butmake up more than 50 % of its value.Reasons for the sometimes high rate ofseed failure are not yet exactly known,since the problem has not been paid ade-quate attention. Its economic relevance isemphasized by the fact that seed yield ispositively correlated with the number ofseeds per pod (Haselberg 2000). The ob-jective of this study was to brighten theknowledge on the processes involved inpollen tube growth and fertilization andthus to contribute in finding strategies toimprove the ovule-to-seed ratio.
MATERIALS AND METHODSExperiments were carried out in SouthPortugal with trees of the female variety‘Mulata’. Flowering branches (3 to 5-years-old wood) were isolated with densegaze cages. Inflorescences with flowersat anthesis were tagged and all but 5 to 7synchronously developing flowers remo-ved. Mature anthers were collected fromvarious male trees and then stored in awarm environment until dehiscence.Hand pollination was carried out using abrush. Flowers were harvested between 6hours and 11 days after hand pollination.For measurements of ovary and stigmalongevity, flowers were hand pollinated 4,6, 8, and 10 days after anthesis, maintai-ned isolated and sampled after another 4days. Sampled flowers were fixed imme-diately in ethanol, formalin, glacial aceticacid solution (9:0,5:0,5). For further pre-paration, the samples were rinsed withdestillated water and then soaked in 8 NNaOH during 18 h for macerization. Aftercomplete removal of NaOH, flowers werefixed and stained in 0,1 N K3PO4 withalkaline aniline blue (0,1 %). Microscopicexaminations included 11 to 15 flowersper treatment. Flowers were cut by handinto vertical and/or longitudinal transsec-tions and surveyed for pollen tubegrowth; ovules and nucelli were measu-red and checked for pollen tubes. Anilinblue-induced fluorescence of callose si-tes was observed using a Zeiss standardmicroscope with Plan (2,5) and Neofluar
(16, 25) objectives. An HG-lamp lightsource, filter combination 48-77-04 (pro-viding blue-violet excitation), an excita-tion filter G 405 and colour partitioner 460with longpass barrier 495 were applied.
RESULTS AND DISCUSSIONPollen germination and tube growthcharacteristicsNon-germinated pollen were ellipsoid-for-med, in contrast, germinated pollen gra-ins were of approximately round shapeand had a diameter of 31,3 µm (± 2,0).Findings are in accordance with formerstudies on carob pollen by several au-thors (Ferguson 1980, Ciampolini et al.1988). In the hand-pollinated flowers anaverage number of 187 germinated pollengrains per stigma was assessed. Non-vi-tal (unstained and tubeless) pollen grainswere found on all stigmas. The pollenwas not subjected to vitality tests, but alarge variability in pollen germinability of4 to 69 % has been reported for both wildand cultivated male genotypes (Sfakiota-kis 1978, Ciampolini et al. 1988).
Pollen tubes penetrated between the pa-pillae, passed the stigma tissue, andorientated towards the dorsal side of thebent pistil, where they formed dense tubebundles in the stylar transmitting tissue.Tubes that had entered the ovule cavitygrew dorsally along the placenta surfaceand passed the ovules or entered themvia micropyle in order to penetrate the nu-cellus. No specific tissue functioning aspollen tube pathway through the placentawas identified. Pollen tubes often displa-yed irregularities like fringes, knots, andcallose plugs. In the ovule, penetratingtubes often formed knots in front of thenucellus (Fig. 1).
Fig. 1. Pollen tube growth in style and ovary:Pollen tube forming a knot in front of the
penetrated nucellus. Bars represent 200 µm.

56 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
The tubes began to decline five days aftergermination. They displayed decreasingfluorescence and irregular consistence.Tubes separated into swollen and fluo-rescing parts with massive callose plugsand callose free parts of thin transparentsegments.
Callose plugs are common in pollen tu-bes: they are considered to continuouslypartition older parts of the pollen fromyounger actively growing parts (Cresti etal. 1992). Large irregular plugs have alsobeen related to incompatibility conditions,which have not been reported for carobso far, but the observed knots in tubes infront of penetrated ovules might corres-pond to such irregularities.
Pollen tube growth in style and ovaryAir temperature greatly affected the velo-city of pollen tube growth through thestyle. After 6 h, tubes developing at20,1°C had grown through the upper partof the style and had often reached thecentre part, while at 32,3°C tubes hadonly extended to the upper style region.
Similar growth relations were observedafter 12 h for the respective temperatures(Fig. 2). No differences were found for pol-len tube growth in samples taken 24 and 48h after pollination at mean temperatures of19,7 to 26,4 °C. It is therefore presumedthat temperatures within the range of 20 to26°C offered favourable conditions for po-llen tube growth, whereas at 31 to 33°C(temporarily reaching 35,5°C) growth wasslowed but not inhibited.
These findings mostly correspond withobservations by Sfakiotakis (1978) in ca-rob, who found that pollen tube growth invitro thrived between 15 and 30°C withthe optimum at 25°C and was considera-bly reduced at 33°C. However, tubes ob-served in the present study (in vivo) rea-ched more than twice the length of thosegrowing in vitro.
Disregarding temperature effects, velocityof pollen tube growth always was highestduring the first hours after germination:Within 12 to 24 h tubes passed the trans-mitting tissue and extended to the vergeof the locule corresponding to a distanceof 2,8 mm. Subsequently the tube growthslowed. First penetration of proximal ovu-les was observed 48 h after pollination,and single tube tips had advanced to themost distal ovule, corresponding to a dis-tance of 5,2 mm, 120 h (5 d) after pollina-tion (Fig. 2). Density of pollen tubes alsodecreased between stigma surface andthe base of the ovary. A large proportionof tubes (87,1 %) ceased growth withouthaving reached the locule. Only 1,5 % ofthe tubes extended to the distal ovules.Furthermore, growth potential and veloci-ty showed considerable variation be-tween individual tubes.
A slow-down in growth velocity before tu-bes reach the ovule is a common pheno-menon in tree crops and has been repor-ted in almond (Pimienta et al. 1983), avo-cado (Sedgley 1979a), and cherry (Bartzand Stösser 1989). Sedgley (1979a) sug-gested immaturity of embryo sacs at an-
thesis and a resultant lack in attractionpower by the ovules to be the reason forthe growth delay. Early differences intube growth may also be related to thestart-position of pollen grains: Those ger-minating on the edge of the large stigmasurface have a longer way through the stylecompared to grains germinating in the cen-ter. Differences in development between in-dividual pollen tubes may as well resultfrom varying starch reserves, as it was re-ported for the in vitro germination of pollengrains in other fruit crops such as Prunusavium (Bartz and Stösser 1989). There areintensive interactions between the growingpollen tubes and the surrounding stylar tis-sue, which comprise the enzymatic degra-dation of stylar components by the pollentubes and their use as nutrition (Cresti et al.1992). In the present study on carob, thenumber of pollen tubes diminished increa-singly towards the stylar base, as has beenequally reported for several other tree spe-cies. According to Bartz and Stösser (1989)a reduction of tubes in the style occurs dueto their competition for the nutrient resour-ces from the surrounding tissue.
The basipetal gradient in number andgrowth velocity of pollen tubes has a directeffect on seed set. With increasing distanceof ovules from the stigma the fertilizationrate is reduced, which is indicated by thedistribution pattern of seeds in carob podsassessed by the author (Fig. 3).
Occasionally a single nucellus was pene-trated by more than one tube (Fig. 4). It isgenerally assumed that the final stage of
Time afterpollination
h d
6 T+
6 T -
12 T+
12 T -
24 1
36
48 2
3
4
5
Styleregion
top centre base locule 1 2 3 4 5 6 7 8 9 10 11 12
OvaryPosition of ovules
Stigma
Fig. 2. Scheme of pollen tube growth in carob flowers: Velocity of pollen tube growth in the pistil.Thickness of lines indicates the frequency of tubes at the respective position. T+, T- correspond to high (31,3 - 32,9°C)
and moderate (21,3 - 21,5°C) temperatures.
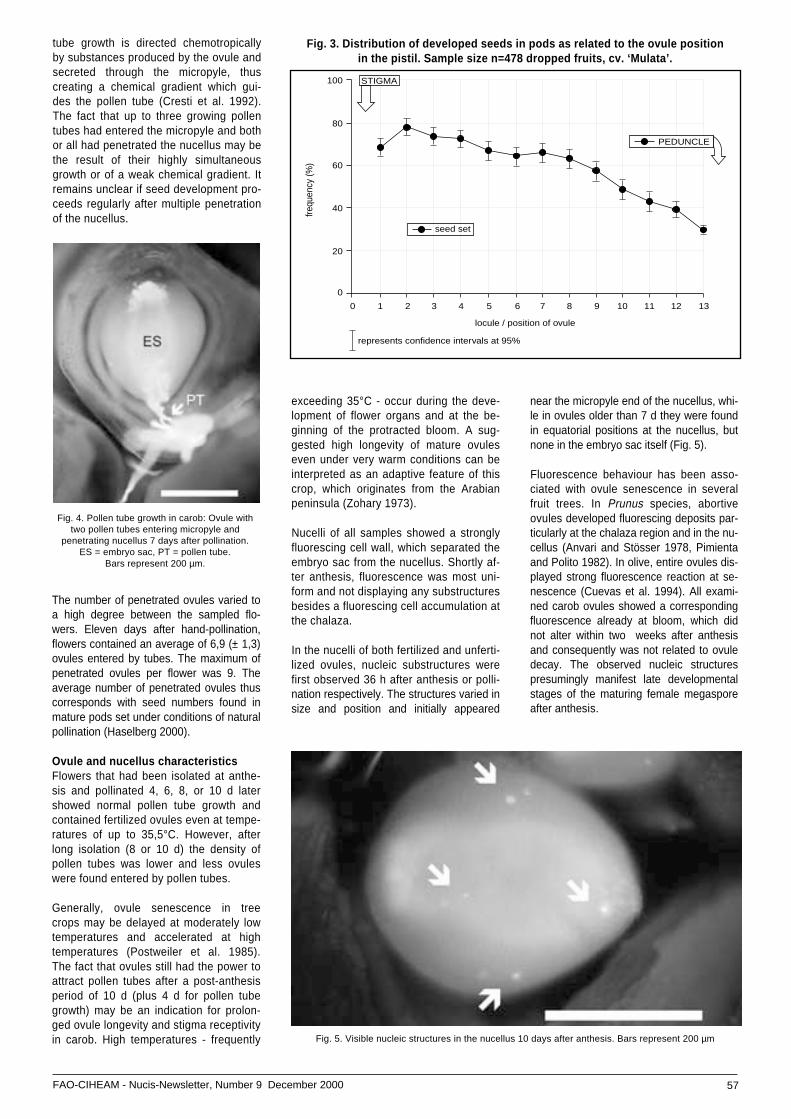
57FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
tube growth is directed chemotropicallyby substances produced by the ovule andsecreted through the micropyle, thuscreating a chemical gradient which gui-des the pollen tube (Cresti et al. 1992).The fact that up to three growing pollentubes had entered the micropyle and bothor all had penetrated the nucellus may bethe result of their highly simultaneousgrowth or of a weak chemical gradient. Itremains unclear if seed development pro-ceeds regularly after multiple penetrationof the nucellus.
exceeding 35°C - occur during the deve-lopment of flower organs and at the be-ginning of the protracted bloom. A sug-gested high longevity of mature ovuleseven under very warm conditions can beinterpreted as an adaptive feature of thiscrop, which originates from the Arabianpeninsula (Zohary 1973).
Nucelli of all samples showed a stronglyfluorescing cell wall, which separated theembryo sac from the nucellus. Shortly af-ter anthesis, fluorescence was most uni-form and not displaying any substructuresbesides a fluorescing cell accumulation atthe chalaza.
In the nucelli of both fertilized and unferti-lized ovules, nucleic substructures werefirst observed 36 h after anthesis or polli-nation respectively. The structures varied insize and position and initially appeared
near the micropyle end of the nucellus, whi-le in ovules older than 7 d they were foundin equatorial positions at the nucellus, butnone in the embryo sac itself (Fig. 5).
Fluorescence behaviour has been asso-ciated with ovule senescence in severalfruit trees. In Prunus species, abortiveovules developed fluorescing deposits par-ticularly at the chalaza region and in the nu-cellus (Anvari and Stösser 1978, Pimientaand Polito 1982). In olive, entire ovules dis-played strong fluorescence reaction at se-nescence (Cuevas et al. 1994). All exami-ned carob ovules showed a correspondingfluorescence already at bloom, which didnot alter within two weeks after anthesisand consequently was not related to ovuledecay. The observed nucleic structurespresumingly manifest late developmentalstages of the maturing female megasporeafter anthesis.
100
80
60
40
20
0
0 1 2 3 4 5 6 7 8 9 10 11 12 13
STIGMA
seed set
PEDUNCLE
locule / position of ovule
frequ
ency
(%)
represents confidence intervals at 95%
The number of penetrated ovules varied toa high degree between the sampled flo-wers. Eleven days after hand-pollination,flowers contained an average of 6,9 (± 1,3)ovules entered by tubes. The maximum ofpenetrated ovules per flower was 9. Theaverage number of penetrated ovules thuscorresponds with seed numbers found inmature pods set under conditions of naturalpollination (Haselberg 2000).
Ovule and nucellus characteristicsFlowers that had been isolated at anthe-sis and pollinated 4, 6, 8, or 10 d latershowed normal pollen tube growth andcontained fertilized ovules even at tempe-ratures of up to 35,5°C. However, afterlong isolation (8 or 10 d) the density ofpollen tubes was lower and less ovuleswere found entered by pollen tubes.
Generally, ovule senescence in treecrops may be delayed at moderately lowtemperatures and accelerated at hightemperatures (Postweiler et al. 1985).The fact that ovules still had the power toattract pollen tubes after a post-anthesisperiod of 10 d (plus 4 d for pollen tubegrowth) may be an indication for prolon-ged ovule longevity and stigma receptivityin carob. High temperatures - frequently
Fig. 3. Distribution of developed seeds in pods as related to the ovule positionin the pistil. Sample size n=478 dropped fruits, cv. ‘Mulata’.
Fig. 4. Pollen tube growth in carob: Ovule withtwo pollen tubes entering micropyle and
penetrating nucellus 7 days after pollination.ES = embryo sac, PT = pollen tube.
Bars represent 200 µm.
Fig. 5. Visible nucleic structures in the nucellus 10 days after anthesis. Bars represent 200 µm

58 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Initiation of fruit growthThe beginning of fruit development in ca-rob is characterized by the elongation ofthe pistil, which stretches, straightens,and by loosing its bent form appearsslightly flattened. Four days after pollina-tion, i.e. 24 to 48 h after the first penetra-tion of an ovule, flowers respectively fruit-lets exhibited initial signs of elongation.Seven days after pollination, the loculewas filling with a glibbery substance em-bedding the ovules, and after eleven dayspistil length had increased by 100 %. Un-like the stigmas in flowers with fertilizedovules, which subsequently displayed acolour change from yellow to brown, stig-matic papillae in non-pollinated flowersmaintained their yellow colour and nectarsecretion throughout the whole observa-tion period. Prolonged receptivity of thestigma tissue in isolated flowers, as ob-served here, was also mentioned by ILAHI
(1979), however, maximum period for lon-gevity remains unknown. It is suggested thatthe stigma performance is maintained longerthan ovule longevity as an adaptive strategyto uphold attractivity towards insects.
CONCLUSIONSVarious causes may account for low seedset in carob pods. Seed failure occursmore frequently in ovules located in distalpositions to the stigma, since these aremost affected by the observed basipetalgradient in growth velocity and density ofpollen tubes. This emphasizes the neces-sity for the presence of large numbers offit pollen grains on the stigma at anthesisin order to provide optimum conditions formaximum seed set. For the carob farmerit brings up the question of optimizing thedensity and quality of pollinators.
Phenomenons like knots and plugs in thepollen tubes possibly indicate pollen in-compatibility, which inhibits normal seeddevelopment. Furthermore, poor seed setmay also result from low pollen germina-tion rates.
The complex of pollination and fertiliza-tion in carob deserves further attention. Itis proposed to focus future research acti-vities on the effect of quantity and qualityof pollen, respectively the effect of polli-nator genotype on seed set, seed abor-tion, and seed quality, taking into accountpossible incompatibilities or xenia.
REFERENCES
Anvari, S.F. and R. Stösser. 1978. Fluo-reszenzmikroskopische Untersuchun-gen des Pollenschlauchwachstums unddes Zustandes der Samenanlagen beiSauerkirschen. Mitt. Klosterneuburg28:23-30.
Bartz, M. and R. Stösser. 1989. Quantita-tive Auswertung der Pollenschläuche imGriffel von Sauerkirschen (Prunus cera-sus L.) in Beziehung zum Fruchtansatz.Gartenbauwiss. 54:132-137.
Ciampolini, F., Cresti, M., Crescimanno,F.G., Di Lorenzo, R. and A. Raimondo.1988. Osservazioni morfo-fisiologiche supiante maschili ed ermafrodite di carrubo(Ceratonia siliqua L.). Proceedings II Int.Carob Symp., Valencia, Spain. 29 Sept.-1Oct. 1987: 153-167.
Cresti, M., Blackmore, S. and J.L. vanWent. 1992. Atlas of sexual reproductionin flowering plants. Springer-Verlag, Ber-lin, Heidelberg.
Cuevas, J., Rallo, L. and H.F. Rapoport.1994. Initial fruit set at high temperaturesin olive, Olea europea L. J. Hort. Sci. 69:665-672.
Ferguson, I.K. 1980. The pollen morpho-logy of Ceratonia (Leguminosae: Caesal-pinioideae). Kew Bull. 35:271-277.
Haselberg, C. von. 2000. Vegetativegrowth and flower and fruit developmentin carob trees (Ceratonia siliqua L.) withspecial emphasis on environmental con-ditions at marginal sites in South Portu-gal. Humboldt-Universität zu Berlin, Ger-many, 178 pages. (in press)
Ilahi, I. 1979a. The nature and balance ofgrowth regulators in the reproductive or-gans of Ceratonia siliqua L. Phyton (Ar-gentina) 37: 29-34.
O’Neill, S. 1997. Pollination regulation offlower development. Ann. Rev. Plant Phy-siol. 48: 547-574.
Pimienta, E. and V.S. Polito. 1982. Ovuleabortion in ‘Nonpareil’ almond [Prunusdulcis (Mill.) D.A. Webb]. Amer. J. Bot.69: 913-920.
Pimienta, E. and V.S. Polito. 1983. Embryosac development in almond [Prunus dulcis(Mill.) D.A. Webb] as affected by cross-,self- and non-pollination. Ann. Bot. 51:469-479.
Pimienta, E., Polito, V.S. and D.E. Kester.1983. Pollen tube growth in cross- andself-pollinated ‘Nonpareil’ almond. J. Am.Soc. Hort. Sci. 108: 643-647.
Postweiler, K., Stösser, R. and S.F. An-vari. 1985. The effect of different tempe-ratures on the viability of ovules in cher-ries. Scientia Hortic. 16: 29-38.
Sedgley, M. 1979a. Light microscope stu-dy of pollen tube growth, fertilization andearly embryo and endosperm develop-ment in avocado varieties ‘Fuerte’ and‘Hass’. Ann. Bot. 44: 353-359.
Sfakiotakis, E.M. 1978. Germination in vi-tro of carob (Ceratonia siliqua L.) Pollen.Z. Pflanzenphysiol. 89: 443-447.
Ch. von HaselbergFachgebiet Obstbau
Humboldt-University of BerlinAlbrecht-Thaer-Weg 314129 Berlin, Germany
E-mail: [email protected]
NOTES AND NEWS
OPPORTUNITIES FOR WALNUTGROWING AND RESEARCH IN
THE UK
Cultivation of walnut in Britain is minimal.Although odd walnut trees are not uncom-mon in farms and larger gardens thereare very few commercial orchards. Howe-ver, in the absence of late frosts, reason-able fruit set can be achieved, at least insouthern Britain, although squirrels mayconsume most of the nuts before harvest.
CONSUMER AWARENESS
The consumer increasingly demands toknow exactly what he is eating. Origin,type of cultivation, treatments applied,storing and processing are matters ofgreat importance in today’s world of banaland insipid foods.
This is why the return to traditional andgenuine food and tastes is becoming in-creasingly essential.
The chestnut therefore represents one ofthe most natural of foods. The significantrise in demand in recent years thereforemakes it essential to understand its intrin-sic characteristics, its beneficial qualitiesand methods of use.
The interesting study undertaken by Prof.Bounous of Turin University for the Ches-tnut Appreciation Association aims to pro-vide the consumer with an answer toeverything that he wants to know.
The conclusion is clear: the chestnut na-turally supplies an abundance of energywhich can help to face the ever increa-singly stressful pace of today’s lifestyle,being especially valuable for those invol-ved in sports activities whatever the age.
This is the true message that our Asso-ciation wishes to pass on to the consu-mer; that the tried and tested data provi-ded by the present study, can therebyguarantee that the “Chestnut season”lasts throughout the year.
E. MagnanoChairman of the “Chestnut Season” Association
Via Vittorio Amedeo, 1312100 Cuneo, ItalyFax: 0171 602773

59FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
The fruit may be used green, for pickling,or ripe, for fresh consumption. And, thetimber from suitable trees can be valua-ble for cabinet making. Research at HRIEast Malling over 50 years ago short-lis-ted several UK and continental varietiesthen considered promising. More recent-ly, some enthusiastic growers have star-ted to plant varieties such as ‘Broadview’,‘Buccaneer’ and ‘Lara’.
With renewed interest in walnut, that maylead to an expansion of acreage, stimulatedin part by the need for diversification andprospects of climate change, HRI East Mal-ling has recently undertaken three initiati-ves. A collection of some 40 fruit varieties(and another 40 timber clones of J. regia, J.nigra and hybrids of J. nigra x J. regia) wasestablished in 1999; by monitoring e.g. timeof leafing and yield, we plan to identify can-didates for trial. A ‘Walnut Day’ was held inOctober 2000 to alert interested parties toopportunities for walnut cultivation. The daycovered fruit and timber aspects of walnutgrowing, uses and research, and was wellattended; a report of the day is availablefrom the authors. As a result of the WalnutDay, a Walnut Club has been establishedto bring together growers, marketers andresearch workers to exchange information,consider research priorities and raisefunds. The Walnut Club will be working inconjunction with the British and Irish Hard-woods Improvement Programme, WalnutGroup in relation to timber aspects.
HRI East Malling would be very interestedto take part in international collaborativeprojects such as Shared Cost projectsfunded by the European Union. Althoughour renewed experience with walnuts isrelatively recent, we can offer expertise inbreeding and genetics, for example thedevelopment of microsatellites. We wouldalso be interested to acquire and exchan-ge germplasm.
K. Russell and K. TobuttHRI East Malling
Plant Breeding and Biotechnology Dept.West Malling, Kent ME19 6BJ, UK.
e-mail: [email protected]
RESPONSETO NUCIS 8 QUESTIONAIRE
Attached to NUCIS 8 issue there was asimple form requesting basic information toreaders to update our FAO-CIHEAM Inter-regional Research Network on Nuts. Inaddition there was a short questionaire onNUCIS to assess the level of interest raisedby each individual section including a placefor making comments. A summary of theanswers provided is presented here.
The number of NUCIS 8 issue distributedwas 1200. Regarding the level of respon-se to the questionaire included in the lastNUCIS issue was extremely poor as onlyabout 7% of the readers have filled thequestionaire and sent it back to us. It isknown that some 50 issues are sent to li-braries and thus the answer is not expec-ted. This is worrying as it was clearly sta-ted in the form that to carry on receivingthe Newsletter it was necessary to fill thecorresponding box (are you still interes-ted to receive NUCIS?). Perhaps readersare only focussed on their field of interestand do not bother about filling forms andsend them back or maybe they are notreally interested. As Coordination Centreand responsible of NUCIS mailing anddistribution, our position is difficult, shall
we still send the bulletin to the completeNUCIS mailing list or only to the respon-ders? The Network Coordinator has deci-ded to carry on sending it to all the peopleincluded in the mailing list.
In relation to information on species and/or fields of interest of readers the respon-se is summarised on Table 1. Informationregarding interest on the five differentNUCIS sections is compiled on Table 2.
The Editor thanks all readers who havecared filling the questionaire and sent itback to him. The information gatheredhas been extremely useful for an overallassessment of this Newsletter.
Table 2. Rating of the level of interest raised by the different NUCIS sections
Sections Interest Average (%) Evaluation Average(from 1 to 10)
Editorial 51.8 7.5
Articles and Reports 78.3 8.7
Notes and News 71.1 7.8
Congresses and Meetings 72.3 8.2
To be held 48.2 8.1
Bibliography 65.0 8.4
Table 1. Number of interested readers per species and/or field of interest
Species and/or fields of interest Number of interested people
Almond 42
Hazelnut 27
Walnut 48
Pistachio 41
Pecan 15
Chestnut 30
Stonepine 11
Genetic Resources 45
Economics 19
Others 26

60 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
CONGRESSES ANDMEETINGS
I INTERNATIONAL SYMPOSIUMON STONEPINE HELD IN
VALLADOLID, SPAIN
The First International Symposium onStonepine was held in Valladolid, Spainfrom 22nd to 24th February 2000 and orga-nized by the autochthonous regional go-vernment of Castile and León (Junta deCastilla y León) together with the SpanishInstituto Nacional de Investigación y Tec-nología Agraria y Alimentaria (INIA) andthe Confederación de Organizaciones deSilvicultores de España (COSE). In addi-tion some 10 different institutions, amongthem the FAO-CIHEAM Research NutNetwork, took part in its organization. Thevenue was the magnificent Palace ofCongresses “Conde Ansúrez” of Vallado-lid. The convener was G. Catalán fromINIA, CIFOR, Madrid, who is also the Liai-son Officer of the FAO-CIHEAM Subnet-work on Stonepine.
The Symposium was attended by some350 participants (producers, managers,researchers, students, processors anddealers) from 5 countries: Italy, Morocco,Portugal, Spain and Tunisia. Apart fromthe large Spanish representation the Por-tuguese group (some 30 people) was par-ticularly numerous.
The scientific programme included sevensessions related to the following topics:ecology and exploitation improvement ofadult stonepine stands; phytopathology,pests, sprays and fire; producing plan-tings, selection, breeding and biotechno-logy; the pinion pine in the diet; nut, woodand other products: characteristics andproduction; nut, wood and other products:industry, market and commerce; generalaspects, history and legality. Thesescientific sessions took up two days.Thanks to the simultaneous translationthis trilingual symposium (English, Spa-nish and Portuguese) promoted unders-tanding and useful discussions. As part ofthe programme and to present the activi-ties undertaken by the FAO-CIHEAM Re-search Nut Network and, in particular,within the Subnetwork on stonepine an in-vited paper was delivered by F.J. Vargasand G. Catalán. A technical visit was or-ganized during the third and last day tothe main Spanish stonepine producingarea and the village of Pedrajas de SanEsteban, where the major world stonepine industry is placed. Visits were alsopaid to adult stonepine stands, traditional
and mechanical harvesting exhibitionswere performed, also demonstrations ofneedle grafting technique on pine used toimprove production with clonal plant ma-terial were practiced by G. Catalan andcollaborators. Also the stonepine proces-sing industry was visited.
The second day the “Valladolid declara-tion” was signed to rise political aware-ness regarding the stonepine industry inEurope, as the EU is the main world pro-ducing area. The declaration includes sixissues. Among the six issues there is theestablishment of a clear and distinct pro-duct for the pinion produced by stonepine(Pinus pinea) and to organize and sup-port a campaign for the promotion of thisnut based on this differentiation. Current-ly there is an unfair competition with theimport of nuts from some other pine spe-cies produced in Asia as their nut qualityis lower. The “Valladolid declaration” wassigned by the main processors and whole-salers associations.
The proceedings of the I Symposiumwere published only in Spanish in two vo-lums I (340 pages) and II (437 pages).
V INTERNATIONAL CONGRESSON HAZELNUT HELD
IN OREGON, USA
The Fifth International Congress on Ha-zelnut was held August 27-31, 2000 onthe campus of Oregon State University(OSU) in Corvallis, Oregon, USA. TheCongress was sponsored by ISHS, OSUCollege of Agricultural Sciences, and theOregon hazelnut industry.
Seventy-one scientists from 19 countriesattended the Congress. They were joinedby an additional 24 people from Oregonand 18 accompanying persons. The re-presented countries included the four ma-jor producers (Turkey, Italy, USA, andSpain), several additional countries withimportant genetic resources, and severalcountries interested in establishing hazel-nut industries. Financial assistance wasprovided to scientists in developing coun-tries who would otherwise not have beenable to attend. Twenty-five new membersjoined ISHS.
A total of 86 papers were presented (37as oral presentations and 47 as posters).Status reports were presented for severalcountries. Other topics included breeding,genetics, genetic resources, cultivar eva-luation, kernel quality, climatic adapta-tion, floral biology, insect pests, diseases,propagation, mineral nutrition, training
systems, cultural practices, and econo-mics. Two talks were on related topics:truffle fungi and the Chilean hazel (Gevui-na avellana).
Following a four-day pre-Congress tour,the Congress started with tours of OSUhazelnut research plots, HarryLagerstedt’s “The Peach Place” orchard,and the USDA-ARS National ClonalGermplasm Repository, and a welcomingreception. The programme allowed for ex-tensive formal and informal exchangesamong the participants. An industry tourexposed participants to several aspectsof Oregon’s hazelnut industry, includingcommercial orchards, insect pest mana-gement practices, a processing plant, anursery, orchard equipment, an on-farmdryer, and an orchard devastated by eas-tern filbert blight.
At the Monday evening banquet, specialawards were presented to Drs. Harry B. La-gerstedt and Maxine M. Thompson to re-cognize their many contributions to hazel-nut research during their careers.Researchers from Reus, Spain offered tohost the Sixth Congress in late June or ear-ly July, 2004, and the invitation was accep-ted unanimously. For details, please con-tact Dr. Mercè Rovira ([email protected]) or Dr. Joan Tous ([email protected]) by e-mail or by Fax (977-344-055). Dr. Shawn A. Mehlenbacher, Conve-ner. Dept. of Horticulture, Oregon StateUniversity. Corvallis, OR 97331 USA.
Stonepine growing in Castile, Spain

61FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
III INTERNATIONAL ALMONDWORKSHOP HELD
IN TRAS-OS-MONTES,PORTUGAL
The Portuguese almond sector: produ-cers, researchers, advisers and proces-sors met in the region of Tras-os-Montes-Alto Douro (north-east of Portugal) duringtwo days, 7 and 8 of July 2000. The DirecçâoRegional de Agricultura de Tras-os-Mon-tes (DRATM), Divisâo de Produçâo Agrí-cola was the organizer and A. Monteirofrom DRATM was the convener. Themain aim was to asses the almond indus-try in Portugal and to plan some actionsfor the future of this tree crop in Portugal.
Two meetings were held; the first in Al-fândega da Fé the 7th of July. Some 25people gathered to discuss about the cur-rent situation of the almond crop in Portu-gal and future prospects. Four talks weredelivered during the morning sessionabout: Current situation of almond pro-duction in the Tras-os-Montes region byA. Monteiro (DRATM); Lines of almondresearch and development carried out inPortugal by Instituto de Biología Experi-mental e Tecnológica (BET), M. Oliveira;Lines of research and development carriedout in Spain by Institut de Recerca i Tecno-logia Agroalimentàries (IRTA) by F.J. Var-gas (IRTA); Research on almond in themain producing countries by I. Batlle(IRTA). After the talks an open discussionwas raised regarding almond versus olive,dry conditions versus irrigation, yields andcrop profitability. The main issues of the al-mond current situation in Portugal were rai-sed. The three main producing regions are:Tras-os-Montes (48%), Algarve (35%)and Beira Interior (13%). The traditionalplantings were made with early floweringand hard shelled local cultivars. Later thelate blooming French cultivars obtained atINRA namely ‘Ferragnes’, ‘Ferrastar’ and‘Ferraduel’ were used together with theItalian ‘Cristomorto’. Recently, the late flo-wering and vigorous cultivars released byIRTA: ‘Masbovera’, ‘Glorieta’ and ‘Francolí’are being planted. Almond orchards areusually placed on poor and acid soils,without giving much fertilization and oftenon too close tree spacing. The presence ofnatural honey bee populations in mostgrowing areas seems to be sufficient to en-sure cross pollination. After a superb lunchin the Estalagem da Senhora das Neves atthe Serra de Bornes, where the meetingwas held, F.J. Vargas gave a talk aboutmain aspects of almond cultivar choice fornew plantings and later the exchange ofopinions on varietal choice continued. Af-terwords a visit to the main almond crack-ing and processing plant in Portugal,«Amendouro», sited in Alfândega da Féwas paid.
The following day, the 8th of July, a se-cond meeting with growers was held inTorres de Moncorno in the venue of the50 years old Theater-cinema of the villa-ge. Two sessions chaired by J. Azevedo,DRATM’s Regional Director, were heldon: Almond growing in the Mediterraneancountries and on Development of the al-mond crop in Portugal in which 5 talkswere delivered by F.J. Vargas, V. Cordei-ro (DRATM), A. Pires (AREVDN), M. Pi-mentel (UTAD) and A. Martins (DRA-ALG). At the end of the session, an opendiscussion was held between growersand lecturers. The issue of aids to nutgrowers from the European Union andthe growers association were raised. Thefuture prospect of agricultural supportprogrammes for the socioeconomical de-velopment of European rural regionswere outlined by J. Azevedo.
SPANISH WORKSHOPON ALMOND TECHNOLOGYAND ECONOMICS HELD IN
ALMERÍA, SPAIN
The Institute of Economic Studies and Pro-jects of the “Caja Rural de Almería” organi-zed a two days II Spanish Workshop on al-mond technology and economics, in Alme-ría on the 27th and 28th of June, 2000.
Some 80 participants from most Spanishalmond producing regions, representing thewhole almond industry gathered in Almería(AEOFRUSE, ALMENDRAVE, CooperativeFederation, MAPA, Nut APOs, researchersetc.) The only missing were the representa-tives of the marzipan and “turrón” industry.
The programme included four invited pa-pers: 1. Almond economic importance.The impact of the support policies; 2Commerce and market policies regula-tion. Support policies to the producingsector and interprofessional needs. 3Contribution of research to almond deve-lopment. Book on almond by A. Felipepresentation and, 4 Assessment and ba-lance of the impact of the improvementplans on the almond crop. Current situa-tion and prospects. Acting proposals.
The main issue raised was the prospects ofthe EU support to the Nut producing sector.It is much needed as some other sectorsreceive support like cereals, forestation, oli-ve etc. There is uncertainty about futuresupport policies to nut and carob produc-tion in Europe (there are hopeful and pessi-mistic people). Apart from this main subjectthere was a feeling from the Spanish al-mond industry to support public R+D pro-grammes.
Almond orchard in Tras-os-Montes, Portugal
Some participants of the III International Almond Workshop

62 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
WORKSHOP ON SOILMANAGEMENT AND EROSION
CONTROL IN ALMONDORCHARDS HELD
IN CÓRDOBA, SPAIN
The 19th and 20th of December 2000 a Spa-nish Workshop on Soil management anderosion control in almond orchards washeld at CIFA “Alameda del Obispo”, Anda-lusia, Spain organised by the Dirección Ge-neral de Investigación y Formación Agrariay Pesquera of the Consejería de Agricultu-ra y Pesca, Junta of Andalucía.
The Workshop was focussed on privateand public managers and advisers of thealmond producing industry. The aim wasto rise the issue of soil erosion under Me-diterranean conditions (climatic andlandscaping) and try to find solutionswhich could alleviate this problem. Duringthe Workshop the high level of soil ero-sion generally shown by almond orchardswas clear, mainly due to:
• High sloping soils, most orchards morethan 20%
• Soil with low vegetative covering owingto this crop
• Unsuitable soil management, the mainsystem used is tillage which maintainsthe soil bare and disintegrated the wholeyear
Different techniques were analysed to re-duce the erosive process, like:• Orchard design, planting at contour line,making terraces and patches
• Making works to collect run-off water
• Soil management systems: tillage, nontillage, minimum tillage, green vegetationor inert covering, etc.
Last day a show and display of equip-ment used on soil management was per-formed: new ground sprayers technology,weed movers and mincing of prunedwood. The local organiser was O. Arque-ro, Department of Olive and Fruit treesfrom CIFA, Córdoba, E-mail: [email protected].
WORKSHOP ON TREE PHYSIO-LOGY AND PATHOGENS AND
PESTS OF CHESTNUT HELD INSOPRON, HUNGARY
The annual management committee mee-ting and scientific workshop of the workgroups Tree Physiology and Pathogensand Pests of COST G4 “Multidisciplinary
Chestnut Research” was held in Hotel Ma-roni, Sopron (Hungary), 5-9 May 1999. Thelocal organization was done by L. Radocz(Agricultural University of Debrecen) and F.Varga, M. Varga, I. Szabo and H. Vidoczi(Forestry University of Sopron). 41 resear-chers from 13 different European countriestook part in the meeting.
Invited keynote speakers were L. Gómez(Departamento de Biotecnología, ETS In-genieros de Montes Madrid, Spain) with alecture on Molecular aspects of stress tole-rance in Castanea sativa and B. Mal-Lushaj(Forest and Pasture Research Institute, Ti-rana) with a conference on Chestnut blightand its biological control in Albania.
Presentations to the work group of TreePhysiology dealt with: Incidence of agarbrand and riboflavin on the “in vitro” pro-duction of Marigoule (L. Bourrain and J.C.Navatel); “in vitro” multiplication and roo-ting of clone Marigoule (C.L.Le); Induc-tion of somatic embriogenesys in Casta-nea sativa (U. Sauer); Markers of juvenileand adult phases of chestnut shoots cul-tured “in vitro” (A. Ballester, J.L. Fernan-dez-Lorenzo, N. Vidal, M.C. San-Joseand A.M. Vieitez); Isolation of wound-in-ducible genes from Castanea sativa bythe differential mRNA display method (R.Schafleitner, K. Hohl and E. Wilhelm);Effect of Cryphonectria parasitica culturefiltrate on the growing of micropropagatedchestnut (H. Vidoczi); Effects of elevatedCO2 on acclimatization of “in vitro” rege-nerated chestnuts (J.C. Goncalves, G.Diogo and T. Coelho).
Presentations to the work group of Patho-gens and Pests dealt with: Phytophaguspest species of the zoocoenosy of chest-nut in Hungary (G. Burges); Molecularmethods for description of Cryphonectriaparasitica (T. Mayerhofer); Variability ofCHV1 type Hypoviruses associated withhypovirulent isolates of Cryphonectria pa-rasitica in Italy, assessed by sequenceanalysis of ORF-A regions (A. Mazzaglia,A. Gasbarri, V. Ponzio, N. Anselmi and A.Vannini); Seasonal changes is susceptibi-lity of Castanea sativa to virulent andhypovirulent isolates of Cryphonectria pa-rasitica (L. Guerin, S. Bastein, G. Capronand C. Robin); Virulence tests for C. pa-rasitica “in vivo” and “in vitro” (U. Heini-ger, V. Roethlisberger, P. Hoegger and D.Rigling); “in vivo” testing of pathogenicityof several Greek isolates of Cryphonec-tria parasitica on chestnut (C. Perlerouand S. Diamandis); Species of Phyto-phthora associated with chestnut areasaffected by ink disease in Italy (A.M. Vet-traino, G. Natili, N. Anselmi and A. Vanni-ni); Results on testing of virulent andhypovirulent isolates of the chestnutblight fungus with European testers (G.Juhasova, K. Adamcikova and L. Rado-
IX MEETING OF THEESCORENA COORDINATORSHELD AT GRIGNON, FRANCE
Every four years the coordinators of allEuropean System of Cooperative Rese-arch Networks on Agriculture (ESCORE-NA) Networks meet to review activitiesand jointly review the future orientation oftheir activities and those of the whole ES-CORENA system. This year the NinthMeeting of the ESCORENA Coordinatorswas held at the Institut National Agrono-mique Paris-Grignon (INA P-G), Grignon,France, from 23rd to 25th of November2000. This occasion was useful to dis-cuss and plan future Network activities,particularly for the Nut Network. This wasalso an opportunity to brief and update allCoordinators on new developmentswithin FAO, of particular importance. TheMeeting was organized by P. Morand-Fehr, Coordinator of the Sheep and GoatNetwork. C. Forthomme from FAO/REUled the Meeting which was conducted by
cz); Results of field inoculations of chest-nut blight fungus with converted hypoviru-lent strains in the surrounding of Sopron(M. Varga, H. Vidoczi, L. Radocz and I.Szabo); Progression of chestnut blighttoward the north in France (D. de Ville-bonne, J.C. Streito, A. Guye and C. Ro-bin); VC-type diversity of C. parasitica inforests and plantations of Sopron, Hunga-ry (H. Vidoczi, M. Varga and I. Szabo); “invitro” growing comparison of Cryphonec-tria parasitica and some other Ascomyce-tes isolated from chestnut bark (I. Szabo);Occurence and incidence of C. parasiticain Turkey (M. Gurer); Comparison of VC-types in Hungarian subpopulations ofCryphonectria parasitica (L. Radocz, E.Szabo and Zs. Szabo); Research concer-ning the chestnut blight in Romania (S.Florea and F. Dumitrescu).
The scientific excursion was organized onSaturday (8th of May 1999) in the KoszegMountain National Park area. There areclose to thirty landscape-protection areasin Hungary. Koszeg occupies a specialplace among them according to its beauti-ful chestnut stands and protected rows ofold cellars made of chestnut wood.
The Book of Abstracts of this workshop isavailable from the organizer: L. Radocz.University of Debrecen. Dep. of PlantProtection, Faculty of Agronomy. Univer-sity of Debrecen. P.O.Box:36 H-4015,Debrecen, Hungary. Phone: 36 52347888, fax: 36 52 413385. E-mail:[email protected]

63FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Chesnut orchard at Viterbo, Italy
TO BE HELD:
Almond and Pistachio
III ISHS International Symposium onPistachios and Almonds and XIIGREMPA - Pistachio and Almond MeetingDate: 20-24 May 2001Place: Zaragoza (Spain)Conveners: R. Socias i CompanyUnidad de Fruticultura SIA-DGAApartado 727, 50080 Zaragoza, SpainMontañana 176, 50016 Zaragoza, SpainTel: +34-976-576436Fax: +34-976-575501E-mail: [email protected]
Chestnut
International Symposium on ChestnutDate: October 2003Place: Vila Real, PortugalConvener: C. AbreuAddress: University of Tras-Os-Montese Alto DouroP.O. Box 2025001 Vila Real Codex - PortugalTel: + 351 59 323 688Fax: + 351 59 325 058E-mail: [email protected]
Hazelnut
VI ISHS International Symposiumon HazelnutDate: June-July 2004Place: Reus, SpainConvener: J. TousIRTA – Centre de Mas BovéDepartament d’Arboricultura MediterràniaP.O. Box 415, 43280 Reus, SpainTel: 33 977 34 32 52Fax: 33 977 34 40 55E-mail: [email protected]
R. Krell from FAO/REU. All four membersof ERNAC (European Research NetworkAdvisory Committee) attended. Also D.Gabiña, Representative of CIHEAM, a co-sponsor of three networks, participated.Most coordinators of the 13 ESCORENANetworks attended the Meeting.
Network reports were presented and dis-cussions from the beginning were very le-velly and constructive. Regarding theFAO-CIHEAM Interregional Nut Networkon Nuts, its Coordinator F.J. Vargas pre-sented a progress report of our Networks.In this report, a summary account of ob-jectives, organisation and activities, car-ried out after the last ESCORENACoordinator’s Meeting held in 1996 wasdelivered. During the meeting the suitabi-lity of simplifying the Nut Network structu-re making it more compact was raised.Largely the process agreed would be areduction in the number of sub-networksand joint management.
In relation to the whole ESCORENA Sys-tem and after extensive discussions onnetwork modalities, structure, fundingand the challenge of integrating researchand its application, the main issues consi-dered for improvement were:
- network communication (within andbetween the networks and to the outside),including network visibility, efficiency andknowledge transfer- geographic coverage and intensifica-tion of activities in selected areas- financial resources (availability, trans-parency and records/statistics, sponsors-hip, project approaches); and- improved participation by members,by the networks in other mechanisms andby outsiders in the network (industry, co-sponsors, etc.)
Almond orchard in Tras-os-Montes, PortugalPistachio orchard at Maials,
Lleida, Spain
Pistachio harvest in Ceylanpinar farmin Anatolia, Turkey

64 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
BIBLIOGRAPHY
ALMOND
Acosta, M.R.; Roux, K.H.; Teuber, S.S.; Sa-the, S.K., 1999. Production and characteriza-tion of rabbit polyclonal antibodies to almond(Prunus dulcis L.) major storage protein. Jour-nal of Agricultural and Food Chemistry, 47(10): 4053-4059.
Bilgrami, Z.; Ghaffar, A., 1999. Detection ofaflatoxin in almond seed. Pakistan Journal ofBotany, 31 (1): 227-230.
Browne, G.T.; Viveros, M.A., 1999. Lethalcankers caused by Phytophthora spp. In al-mond scions: Specific etiology and potentialinoculum sources. Plant Disease, 83 (8):739-745.
Cañizares, M.C.; Marcos, J.F.; Pallas, V.,1999. Molecular characterization of an al-mond isolate of hop stunt viroid (HSVd) andconditions for eliminating spurious hybridi-zation in its diagnosis in almond samples.European Journal of Plant Pathology, 105(6): 553-558.
Chandrababu, R.J.; Sharma, R.K., 1999. He-ritability estimates in almond [Prunus dulcis(Miller) D.A. Webb]. Scientia Horticulturae, 79(3-4): 237-243.
Channuntapipat, C.; Collins, G.; Bertozzi, T.;Sedgley, M., 2000. Cryopreservation of in vi-tro almond shoot tips by vitrification. Journalof Horticultural Science & Biotechnology, 75(2): 228-232.
Dicenta, F.; Martínez, P.; Ortega, E.; Duval,H., 2000. Cultivar pollinizer does not affect al-mond flavor. HortScience, 35 (6): 1153-1154.
Duval, H.; Froment, P., 2000. Amande. Qua-tre nouvelles variétés (in French). RéussirFruits & Légumes, (183): 65-67.
Egea, J.; Burgos, L., 2000. Ovule differencesbetween single-kernelled and double-kernel-led fruits in almond (Prunus dulcis). Annals ofApplied Biology, 136 (3): 291-295.
Estaun, V.; Calvet, C.; Camprubí, A.; Pino-chet, J., 1999. Long-term effects of nurserystarter substrate and AM inoculation of micropro-pagated peach x almond hybrid rootstockGF677. Agronomie, 19 (6): 483-489.
Felipe, A.J.; Socias i Company, R.; GómezAparisi, J., 1999. El concepto del ideotipo enla mejora de patrones para el almendro (inSpanish). ITEA, 95 (3): 209-217.
Franco, J.A.; Abrisqueta, J.M.; Hernansaez,A.; Moreno, F., 2000. Water balance in ayoung almond orchard under drip irrigationwith water of low quality. Agricultural WaterManagement, 43 (1): 75-98.
Goldhamer, D.A.; Viveros, M., 2000. Effectsof preharvest irrigation cutoff durations andpostharvest water deprivation on almondtree performance. Irrigation Science, 19 (3):125-131.
Gou, P.; Díaz, I.; Guerrero, L.; Valero, A.; Arn-au, J.; Romero, A., 2000. Physico-chemicaland sensory property changes in almonds ofDesmayo Largueta variety during toasting.Food Sci Tech Int, 6 (1): 1-7.
Gradziel, T.M.; Mahoney, N.; Abdallah, A.,2000. Aflatoxin production among almond ge-notypes is not related to either kernel oil com-position or Aspergillus flavus growth rate.HortScience, 35 (5): 937-939.
Gradziel, T.M.; Weinbaum, S.A., 1999. Highrelative humidity reduces anther dehiscencein apricot, peach and almond. HortScience,34 (2): 322-325.
Grane, N.; Luna, M.C.; Prats, M.S.; Beren-guer, V.; Martin, M.L., 1999. Statistical com-parative study of free acid HPLC data froma selected almond set. Food Chemistry, 65(1): 23-28.
Joobeur, T.; Periam, N.; Vicente, M.C.;King, G.J.; Arús, P., 2000. Development ofa second generation linkage map for al-mond using RAPD and SSR markers. Geno-me, 43 (4): 649-655.
Kumar, K.; Kumar, U., 2000. Evidence ofSelf-Compatibility in Indigenous Almond[Prunus dulcis (Miller) D.A. Webb.] Selectio-ns from India. Journal American Pomologi-cal Society, 54 (2): 68-71.
Ladizinsky, G., 1999. On the origin of al-mond. Genetic Resources and Crop Evolu-tion, 46: 143-147.
La Rosa, R.; Russo, A.; Verdone, A.; Alos-chi, S., 2000. Investigation on hygienic-sa-nitary quality of almond paste produced inSicily. Industrie Alimentari, 39 (389): 137+.
Martin, M.L.; Llorens, C.; Berenguer, V.;Grane, N., 1999. Comparative study on thetriglyceride composition of almond kerneloil. A new basis for cultivar chemometriccharacterization. Journal of Agricultural andFood Chemistry, 47 (9): 3688-3692.
Martínez, P.; Gradziel, T.M., Ortega, E.; Di-centa, F., 2000. Short-term storage of al-mond pollen. HortScience, 35 (6): 1151-1152.
Muncharaz, M., 2000. Afecciones no parasi-tarias en almendro. Fruticultura Profesional,(113): 36-42.
Nyomora, A.M.S.; Brown, P.H.; Krueger, B.,1999. Rate and time of boron application in-crease almond productivity and tissue boronconcentration. HortScience, 34 (2): 242-245.
Pearson, T.C., 1999. Spectral propertiesand effect of drying temperature on almondswith concealed damage. Food Science andTechnology, 32 (2): 67-72.
Prats, M.S.; Grane, N.; Berenguer, V.; Mar-tin, M.L., 1999. A chemometric study of ge-notypic variation in triacylglycerol composi-tion among selected almond cultivars. Jour-nal of the American Oil Chemists Society,76 (2): 267-272.
Ruiz, F.; Fernanez, J.; Gomez, A.; Torregro-sa, E., 1999. Determination of sorption and
diffusion properties of peeled almond nuts.Journal of Food Engineering, 41 (3-4): 209-214.
Shibli, R.A.; Shatnawi, M.A.; Ajlouni, M.M.;Jaradat, A.; Adham, Y., 1999. Slow growthin vitro conservation of bitter almond (Amyg-dalus communis L.). Adv. Hort. Sci., 13:133-134.
Sze Tao, K.W.C.; Sathe, S.K., 2000. Func-tional properties and in vitro digestibility ofalmond (Prunus dulcis L.) protein isolate.Food Chemistry, 69 (2): 153-160.
Takeoka, G.; Dao, L.; Teranishi, R.; Wong,R.; Flessa, S.; Harden, L.; Edwards, R.,2000. Identification of three triterpenoids inalmond hulls. Journal of Agricultural andFood Chemistry, 48 (8): 3437-3439.
Tamura, M.; Ushjima, K.; Sassa, H.; Hirano,H.; Tao, R.; Gradziel, T.M.; Dandekar, A.M.,2000. Identification of self-incompatibilitygenotypes of almond by allele-specific PCRanalysis. Theoretical and Applied Genetics,101 (3): 344-349.
Toles, C.A.; Marshall, W.E.; Wartelle, L.H.;McAloon, A., 2000. Steam-or carbon dioxi-de-activated carbons from almond shells:physical, chemical and adsorptive proper-ties and estimated cost of production. Biore-source Technology, 75 (3): 197-203.
Van Nerum, I.; Certal, A.C.; Oliveira, M.M.;Keulemans, J.; Broothaerts, W., 2000. PD1,an S-like Rnase gene from a self-incompati-ble cultivar of almond. Plant Cell Reports,19 (11): 1108-1114.
Woolley, F.M.; Collins, G.G.; Sedgley, M.,2000. Application of DNA fingerprinting forthe classification of selected almond [Pru-nus dulcis (Miller) D.A. Webb] cultivars.Australian Journal of Experimental Agricul-ture, 40 (7): 995-1001.
CAROB
Arista, M.; Ortiz, P.L.; Talavera, S., 2000. Apicalpattern of fruit production in the racemes of Ce-ratonia siliqua (Leguminosae: Caesalpinioi-deae): Role of pollinators. American Journal ofBotany, 86 (12): 1708-1716.
Ortiz, P.L.; Arista, M.; Talavera, S., 1999. Distan-ce-independent fruit-set pattern in a dioeciouspopulation of Ceratonia siliqua (Caesalpinioi-deae). Flora, 194 (3): 277-280.
Pérez, L.; García, M.; Ruiz, B., 2000. Note. In-fluence of pulp and natural carob fiber onsome aspects of nutritional utilization andcholesterolemia in rats. Food Science and Te-chnology International, 5 (5): 425-430.
Priolo, A.; Waghorn, G.C.; Lanza, M.; Biondi, L.;Pennisi, P., 2000. Polyethylene glycol as ameans for reducing the impact of condensedtannins in carob pulp: Effects on lamb growthperformance and meat quality. Journal of AnimalScience, 78 (4): 810-816.
Vinterhalter, D.V.; Vinterhalter, B.S., 1999.Hormone-like effects of sucrose in plant in vi-tro cultures. Phyton - Annales Rei Botanicae,39 (3): 57-60.

65FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Yousif, A.K.; Alghzawi, H.M., 2000. Proces-sing and characterization of carob powder.Food Chemistry, 69: 283-287.
CHESTNUT
Berrocal del Brío, M.; Carnero Villar, A.B.,1999. Primeros pasos en la lucha biológicacontra el Chancro del Castaño (in Spanish).Agricultura, LXVIII (809): 1036-1038.
Blanco, A.; Rubio, A.; Sánchez, O.; Elena, R.;Gómez, V.; Graña, D., 2000. Autoecología delos castañares de Galicia (España). Invest. Agr.:Sist. Recur. For., 9 (2): 337-361.
Craddock, J.H., 1998. Chestnut Resources inNorth America. 89th Annual Report of the Nor-thern Nut Growers Association: 19-30.
Mancuso, S.; Ferrini, F.; Nicese, F.P., 1999.Chestnut (Castanea sativa Mill.) genotypeidentification: An artificial neural networkapproach. Journal of Horticultural Science &Biotechnology, 74 (6): 777-784.
Nave, J.M., 1998. Large Fruited ChestnutsGrown in North America. 89th Annual Reportof the Northern Nut Growers Association: 42-82.
Nave, J.M., 1998. Some Thoughts on the ‘Colo-ssal’ Chestnut. 89th Annual Report of the Nor-thern Nut Growers Association: 83-96.
Rubio, A.; Elena, R.; Sanchez, O.; Blanco, A.;Sanchez, F.; Gomez, V., 1999. Autoecologíade los castañares catalanes (in Spanish). In-vest. Agri: Prod. Prot. Veg., 8 (2): 387-405.
HAZELNUT
Ackurt, F.; Ozdemir, M.; Biringen, G.; Loker,M., 1999. Effects of geographical origin andvariety on vitamin and mineral composition ofhazelnut (Corylus avellana L.) varieties culti-vated in Turkey. Food Chemistry, 65 (3): 309-313.
Aramburu, J.; Rovira, M., 2000. Incidence andnatural spread of apple mosaic ilarvirus in ha-zel in north-east Spain. Plant Pathology, 49:423-427.
Badalov, P.P., 1998. Mass Selection as a Me-thod for Developing Winter Hardy Filberts.89th. Annual Report of the Northern NutGrowers Association: 153-154.
Begeot, C., 1999. Pollen behaviour of Hazel-nut (Corylus avellana), an indicator or humanimpacts? One example along a transect inSouthern French Jura. Acta Botanica Gallica,145 (4): 271-279.
Cimino, G.; Passerini, A.; Toscano, G., 2000.Removal of toxic cations and Cr(Vi) fromaqueous solution by hazelnut shell. Water Re-search, 34 (11): 2955-2962.
Coyne, C.J.; Mehlenbacher, S.A.; Johnson,K.B.; Pinkerton, J.N.; Smith, D.C., 2000. Com-parison of Two Methods to Evaluate Quantita-tive Resistance to Eastern Filbert Blight in Eu-ropean Hazelnut. J.Amer.Soc.Hort.Sci, 125(5): 603-608.
Erdogan, V.; Mehlenbacher, S.A., 2000. Inters-pecific Hybridization in Hazelnut (Corylus). J.Amer. Hort. Sci., 125 (4): 489-497.
Frei, T., 1998. The effects of climate changein Switzerland 1969-1996 on airborne pollenquantities from hazel, birch and grass. Grana,37 (3): 172-179.
Galderisi, U.; Cipollaro, M.; Di Bernardo, G.;De Masi, L.; Galano, G.; Cascino, A., 1999.Identification of hazelnut (Corylus avellana)cultivars by RAPD analysis. Plant Cell Repor-ts, 18 (7-8): 652-655.
Holzhauser, T.; Vieths, S., 1999. Quantitativesandwich ELISA for determination of traces ofhazelnut (Corylus avellana) protein in complexfood matrixes. Journal of Agricultural and FoodChemistry, 47 (10): 4209-4218.
Kask, K., 1998. European Filbert in Estonia.89th. Annual Report of the Northern NutGrowers Association: 155-156.
Lunde, C.F.; Mehlenbacher, S.A.; Smith,D.C., 2000. Survey of hazelnut cultivars forresponse to eastern filbert blight inoculation.HortScience, 35 (4): 729-731.
Mannina, L.; Patumi, M.; Fiordiponti, P.; Ema-nuele, M.C.; Segre, A.L., 1999. Olive and ha-zelnut oils: A study by high-field H-1 NMR andgas chromatography. Journal of Food Scien-ce, 11 (2): 139-149.
Martinez, N.; Chiralt, A., 1999. Water diffusivi-ty and mechanical changes during hazelnuthydration. Food Research International, 32(6): 447-452.
Mehlenbacher, S.A.; Azarenko, A.N.; Smith,D.C.; McCluskey, R., 2000. ‘Lewis’ Hazelnut.HortScience, 35 (2): 314-315.
Ozdemir, M.; Devres, O., 2000. Analysis ofcolor development during roasting of hazel-nuts using response surface methodology.Journal of Food Engineering, 45 (1): 17-24.
Scortichini, M.; Marchesi, U.; Rossi, M.P.; An-gelucci, L.; Dettori, M.T., 2000. Rapid identifi-cation of Pseudomonas avellanae field isola-tes, causing hazelnut decline in central Italy,by repetitive PCR genomic fingerprinting. Jo-urnal of Phytopathology, 148 (3): 153-159.
Sezen, K.; Demirbag, Z., 1999. Isolation andinsecticidal activity of some bacteria from thehazelnut beetle (Balaninus nucum L.). AppliedEntomology and Zoology, 34 (1): 85-89.
Volovich, P.I.; Chripack, P.I., 1998. Diversityand Abundance of Cultivated Varieties of Fil-berts. 89th. Annual Report of the Northern NutGrowers Association: 157-158.
PECAN
Conner, P.J.; Worley, R.E., 2000. Alternatebearing intensity of pecan cultivars. HortS-cience, 35 (6): 1067-1069.
Dutcher, J.D.; Estes, P.M.; Dutcher, M.J.,1999. Interactions in entomology: Aphids,aphidophaga and ants in pecan orchards.Journal of Entomological Science, 34 (1):40-56.
Grauke, L.J., 1998. Dream Tree: Pecan. Cha-racteristics of the Ideal Tree. 89th. Annual Re-port of the Northern Nut Growers Association:113-120.
Grauke, L.J.; Thompson, T.E., 1998. Cata-log of Pecan Cultivars in the National ClonalGermplasm Repository. 89th Annual Reportof the Northern Nut Growers Association:108-112.
Hall, G.D., 1999. Pecan food potential inprehistoric North America. Economic Bo-tany, 54 (1): 103-112.
Herrera, E.A.; Esparza, V., 2000. Influenceof pruning methods on tree growth and earlynut production of mature transplanted pe-can trees [Carya illinoinensis (Wangenh.) K.Koch]. Phyton – International Journal of –Experimental Botany, 66: 119-127.
Herrera, E.A.; Duarte, E., 2000. Nitrogen ra-tes and time of application effect on growthand nut production of mature transplantedpecan trees [Carya illinoinensis (Wangenh.)K. Koch]. Phyton – International Journal ofExperimental Botany, 66: 129-135.
Li, M.; Bellmer, D.D.; Brusewitz, G.H., 1999.Pecan kernel breakage and oil extracted bysupercritical CO2 as affected by moisturecontent. Journal of Food Science, 64 (6):1084-1088.
Sanderlin, S., 2000. Pecan scion cultivareffects on freeze susceptibility of the roots-tock. Journal American Pomological Socie-ty, 54 (4): 188-193.
Sekita, K.; Saito, M.; Uchida, O.; Ono, A.;Ogawa, Y.; Kaneko, T.; Furuya, T.;Kurokawa, Y.; Inoue, T., 1998. Pecan nutcolor: 90-days dietary toxicity study in F344rats. Journal of the Food Hygienic Societyof Japan, 39 (6): 375-382.
Smith, M.W., 2000. Cultivar and mulchaffect cold injury of young pecan trees. Jo-urnal of American Pomological Society,Fruit Varieties Journal, 54 (1): 29-33.
Smith, M.W.; Carroll, B.L.; Cheary, B.S.,2000. Mulch improves pecan tree growthduring orchard establishment. HortScience,35 (2): 192-195.
Sorensen, R.B.; Jones, T.L.; Campbell, G.S.;Montes, M., 2000. Heat pulse needles tomeasure pecan tree transpiration. Applied En-gineering in Agriculture, 15 (6): 651-657.
Thompson, T.E.; Grauke, L.J., 2000. ‘Hopi’pecan. HortScience, 35 (2): 308-309.
Thompson, T.E.; Grauke, L.J., 2000. ‘Paw-nee’ pecan. Journal American PomologicalSociety, 54 (3): 110-113.
Thompson, T.E.; Grauke, L.J.; Sibbett,G.S., 2000. Host Plant Resistance to Black-margined Aphids on Pecan. Journal Ameri-can Pomological Society, 54 (4): 193-198.
Vendrame, W. A.; Kochert, G.D.; Sparks,D.; Wetzstein, H.Y., 2000 Field Performan-ce and Molecular Evaluations of PecanTrees Regenerated from Somatic Embryo-genic Cultures. J.Amer.Soc.Hort.Sci., 125(5): 542-546.

66 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Wood, B.W.; Reilly, C.C., 1999. Factors in-fluencing water split of pecan fruit. Hortscien-ce, 34 (2): 215-217.
Wood, B.W.; Reilly, C.C., 2000. Pest damageto pecan is affected by irrigation, nitrogenapplication, and fruit load. HortScience, 35(4): 669-672.
Zhang, Q.L.; Brown, P.H., 1999. The mecha-nism of foliar zinc absorption in pistachio andwalnut. Journal of the American Society forHorticultural Science, 124 (3): 312-317.
PISTACHIO
Castola, V.; Bighelli, A.; Casanova, J., 2000.Intraspecific chemical variability of the essen-tial oil of Pistacia lentiscus L. from Corsica.Biochemical Systematics and Ecology, 28 (1):79-88.
Fernández, A.; Camacho, A.; Fernández, C.;Altarejos, A.; Pérez, P., 2000. Composition ofthe essential oils from galls and aerial parts ofPistacia lentiscus L. Journal of Essential OilResearch, 12 (1): 19-23.
Golan, A., 1999. Developmental proteins ofPistacia vera L. bark and bud and their biote-chnological properties: A review. Journal ofFood Biochemistry, 22 (5): 375-382.
Maskan, M.; Karatas, S., 1999. Storage stabi-lity of whole-split pistachio nuts (Pistachiavera L.) at various conditions. Food Che-mistry, 66 (2): 227-233.
Mederos, S.; Trujillo, M.I., 1999. Eliminationof browning exudate and in vitro develop-ment of shoots in Pistacia vera L. Cv. Ma-teur and Pistacia atlantica Desf. Culture.Acta Societatis Botanicorum Poloniae, 68(1): 21-24.
Molina, S.M.; Trujillo, M.I., 1999. Techni-ques for in vitro seed germination in Pista-cia species. South African Journal of Bo-tany, 65 (2): 149-152.
Nzima, M.D.S.; Martin, G.C.; Nishijima, C.,1999. Effect of fall defoliation and springshading on shoot carbohydrate and growthparameters among individual branches ofalternate bearing ‘Kerman’ pistachio trees.Journal of the American Society for Horticul-tural Science, 124 (1): 52-60.
Onay, A., 2000. Micropropagation of Pista-chio from mature trees. Plant Cell Tissueand Organ Culture, 60 (2): 159-163.
Pearson, S.M.; Candlish, A.A.G.; Aidoo,K.E.; Smith, J.E., 1999. Determination ofaflatoxin levels in pistachio and cashewnuts using immunoaffinity column clean-upwith HPLC and fluorescence detection. Bio-technology Techniques, 13 (2): 97-99.
Pearson, T.; Toyofuku, N., 2000. Automatedsorting of pistachio nuts with closed shells.Applied Engineering in Agriculture, 16 (1):91-94.
Polito, V.S.; Pinney, K., 1999. Endocarp de-hiscence in pistachio (Pistacia vera L.). In-ternational Journal of Plant Sciences, 160(5): 827-835.
Rahemi, M.; Baninasab, B., 2000. Effect of gi-bberellic acid on seedling growth in two wildspecies of pistachio. Journal of HorticulturalSciene & Biotechnology, 75 (3): 336-339.
Vaknin, Y.; Eisikowitch, D., 2000. Effects ofshort-term storage on germinability of pista-chio pollen. Plant Breeding, 119 (4): 347-350.
Vemmos, S.N., 1999. Mineral composition ofleaves and flower buds in fruiting and non-fruiting pistachio trees. Journal of Plant Nutri-tion, 22 (8): 1291-1301.
Yildiz, M.; Gurcan, S.T.; Ozdemir, M., 1998.Oil composition of pistachio nuts (Pistaciavera L.) from Turkey. Fett – Lipid, 100 (3): 84-86.
Zhang, Q.L.; Brown, P.H., 1999. Distributionand transport of foliar applied zinc in pista-chio. Journal of the American Society forHorticultural Science, 124 (4): 433-436.
Zwarts, L.; Savage, G.P.; McNeil, D.L., 1999.Fatty acid content of New Zealand-grown wal-nuts (Juglans regia L.). International Journal ofFood Sciences and Nutrition, 50 (3): 189-194.
STONEPINE
Cañellas, I.; Finat, L.; Bachiller, A.; Montero,G., 1999. Comportamiento de planta de Pinuspinea en vivero y campo: Ensayos de técni-cas de cultivo de planta, fertilización y aplica-ción de herbicidas (in Spanish). Invest. Agri:Prod. Prot. Veg., 8 (2): 335-359.
Mutke, S.; Díaz, L.; Gordo, J., 2000. Análisiscomparativo de la rentabilidad comercial pri-vada de plantaciones de Pinus pinea L. entierras agrarias de la provincia de Valladolid.Invest. Agr.: Sist. Recur. For., 9 (2): 269-297.
WALNUT
Alonso, H.; Spangler, H.G.; Rogers, R.; Pa-paj, D.R., 2000. Acoustic component and so-cial context of the wing display of the walnutfly Rhagoletis juglandis. Journal of Insect Be-havior, 13 (4): 511-524.
Badalov, P.P.; Badalov, K.P., 1998. Selectionof Early-Ripening Varieties of Walnut for Win-ter-Hardiness and Nut Quality. 89th. AnnualReport of the Northern Nut Growers Associa-tion: 145-146.
Belisario, A.; Zoina, A.; Pezza, L.; Luongo, L.,1999. Susceptibility of species of Juglans topathovars of Xanthomonas campestris.
Cambardella, C.A.; Moorman, T.B.; Jaynes,D.B.; Hatfield, J.L.; Parkin, T.B.; Simpkins,W.W.; Karlen, D.L., 1999. Water quality inWalnut Creek watershed: Nitrate-nitrogen insoils, subsurface drainage water, and shallowgroundwater. Journal of Environmental Quali-ty, 28 (1): 25-34.
Dupraz, C.; Simorte, V.; Dauzat, M.; Bertoni,G.; Bernadac, A.; Masson, P., 1998. Growthand nitrogen status of young walnuts as affec-ted by intercropped legumes in a Mediterra-nean climate. Agroforestry Systems, 43 (1-3):71-80.
Eidem, J.M.; Simpkins, W.W.; Burkart, M.R.,1999. Geology, groundwater flow, and waterquality in the Walnut Creek watershed. Jour-nal of Environmental Quality, 28 (1): 60-69.
Eriksson, G., 2000. Red europea de conser-vación de recursos genéticos de frondosasnobles (in Spanish). Invest. Agr: Sist. Recur.For.: Fuera de Serie nº2: 59-69.
Escobar, M.A.; Park, J.I.; Polito, V.S.; Leslie,C.A.; Uratsu, S.L.; McGranahan, G.H.; Dan-dekar, A.M., 2000. Using GFP as a scorablemarker in walnut somatic embryo transforma-tion. Annals of Botany, 85 (6): 831-835.
Fernández, J.; Díaz, R.; Cogolludo, M.A.;Pereira, S., 2000. Conservación de recur-sos genéticos de las frondosas nobles enEspaña (in Spanish). Invest. Agr: Sist. Re-cur. For.: Fuera de Serie nº2: 71-93.
Gray, D.; Garrett, H.E.G., 1999. Nitrogenfertilization and aspects of fruit yield in aMissouri black walnut alley cropping practi-ce. Agroforestry Systems, 44 (2-3): 333-344.
Hatfield, J.L.; Jaynes, D.B.; Burkart, M.R.;Cambardella, C.A.; Moorman, T.B.; Prue-ger, J.H.; Smith, M.A., 1999. Water qualityin Walnut Creek watershed: Setting and far-ming practices. Journal of EnvironmentalQuality, 28 (1): 11-24.
Hussendorfer, E., 1999. Identification of na-tural hybrids Juglans x intermedia CARR-using isoenzyme gene markers. Silvae Ge-netica, 48 (1): 50-52.
Lehmkuhler, J.W.; Kerley, M.S.; Garrett,H.E.; Cutter, B.E.; McGraw, R.L., 1999.Comparison of continuous and rotational sil-vopastoral systems for established walnutplantations in southwest Missouri, USA.
LeRoux, X.; Grand, S.; Dreyer, E.; Daudet,F.A., 1999. Parameterization and testing ofa biochemically based photosynthesis mo-del for walnut (Juglans regia) trees and see-dlings. Tree Physiology, 19 (8): 481-492.
Loiko, R.E., 1998. Perspectives on WalnutCulture in Belarus. 89th. Annual Report of theNorthern Nut Growers Association: 147-148.
Loiko, R.E.; But-Gusiam, A.V., 1998. Suita-ble Varieties of Walnuts for Belarus. 89th.Annual Report of the Northern Nut GrowersAssociation: 149-152.
Mary, F.; Dupraz, C.; Delannoy, E.; Liagre,F., 1998. Incorporating agroforestry practi-ces in the management of walnut plantatio-ns in Dauphine, France: an analysis of far-mers’ motivations. Agroforestry Systems, 43(1-3): 243-256.
Moorman, T.B.; Jaynes, D.B.; Cambardella,C.A.; Hatfield, J.L.; Pfeiffer, R.L.; Morrow,A.J., 1999. Water quality in Walnut Creekwatershed: Herbicides in soils, subsurfacedrainage, and groundwater. Journal of Envi-ronmental Quality, 28 (1): 35-45.
Morone, C.; Janse, J.D.; Scortichini, M.,1998. Bark canker of Persian walnut (Ju-glans regia) trees incited by Erwinia nigri-fluens in Italy. Journal of Phytopathology,146 (11-12): 637-639.

67FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
Muncharaz, M., 2000. La recolección delnogal (in Spanish). Fruticultura Profesional,(108): 32-43.
Savage, G.P.; Dutta, P.C.; McNeil, D.L.,1999. Fatty acid and tocopherol contentsand oxidative stability of walnut oils. Journalof the American Oil Chemists Society, 76(9): 1059-1063.
Shirnina, L.V.; Kotliarova, T.I., 1998. Biolo-gy of walnut anthracnosis pathogen in Kras-nodar region. Mikologiya I Fitopatologiya,32 (6): 65-71.
Sze Tao, K.W.C.; Sathe, S.K., 2000. Wal-nuts (Juglans regia L.) proximate composi-tion, protein solubility, protein amino acidcomposition and protein in vitro digestibility.Journal of the Science of Food and Agricul-ture, 80 (9): 1393-1401.
Tobutt, K.R.; Russell, K., 2000. Report ofHRI walnut day – 26 October 2000. TheWalnut Club, HRI – East Malling West Ma-lling, Kent ME19 6BJ England.
Tokar, F., 1999. Leaf area index and dendro-mass production in red oak (Quercus rubra L.)and black walnut (Juglans nigra L.) stands.Ekologia – Bratislava, 18 (3): 246-254.
Zwarts, L.; Savage, G.P.; McNeil, D.L.,1999. Fatty acid content of New Zealand-grown walnuts (Juglans regia L.). Internatio-nal Journal of Food Sciences and Nutrition,50 (3): 189-194.
Thevathasan, N.V.; Gordon, A.M.; Voroney,R.P., 1999. Juglone (5-hydroxy-1,4 naptho-quinone) and soil nitrogen transformationinteractions under a walnut plantation insouthern Ontario, Canada. AgroforestrySystems, 44 (2-3): 151-162.
Willis, R.J., 2000. Juglans spp, juglone andallelopathy. Allelopathy Journal, 7 (1): 1-55.
ECONOMICS
Onunkwo, I.M.; Epperson, J.E., 2000. Exportdemand for U.S. pecans: impacts of U.S. ex-port promotion programs. Agribusiness, 16(2): 253-265.
Anonimous, 2000. Tree nut situation updates.World Horticultural Trade and US Export Op-portunities. (2): 5-9.
Anonimous, 2000. Situation and outlook formacadamias. World Horticultural Trade andUS Export Opportunities. (3): 10-14.
NUTS
Demir, C.; Cetin, M., 1999. Determination oftocopherols, fatty acids and oxidative stabi-lity of pecan, walnut and sunflower oils.Deutsche Lebensmittel – Rundschau, 95(7): 278-282.
Plocher, T., 1998. Nut Explorations in the Bal-tic Countries and Belarus. 89th. Annual Reportof the Northern Nut Growers Association:138-144.
BOOKS
Couceiro, J.F.; Coronado, J.M.; Menchén,M.T.; Mendiola, M.A., 2000. El cultivo delPistachero (in Spanish). Agro Latino, S.L.112 pages.
Köksal, A.Y. (editor), 2000. Inventory ofhazelnut research, germplasm and refe-rences. FAO - REU Technical series 56:129 pages.
Ridley, D.; Beaumont, J., 1999. The Austra-lian Chestnut Growers’ Resource Manual.Department of Natural Resources and Envi-ronment, PO Box 235, Myrtleford, Victoria,Australia 3737. 152 pages. Email:Dan.Ridley @nre.vic.gov.au
PROCEEDINGS
Albisu, L.M. (scientific editor), 1999. Econo-mics of nuts in the Mediterranean basin. Op-tions Méditerranéennes. CIHEAM-FAO - SE-RIE A: Séminaires Méditerranéennes, núm.37: 135 pages.
Junta de Castilla y León, 2000. 1er Simposiodel Pino Piñonero (Pinus pinea L.). Tomo 1:340 pages. Tomo II: 438 pages. Valladolid,Spain. February 2000. (in Spanish)
THESES
Silva, A.P., 1999. Estudos bioclimáticos naaveleira (Corylus avellana, L.) Efeitos poten-ciais na occorência de frutos ocos (in Portu-guese). Universidade de Trás-os-Montes eAlto Douro, Vila Real (Portugal): 211 pp.
de Herralde, F., 2000. Estudio integral de lasrespuestas ecofisiológicas al estrés hídrico:caracterización de variedades de almendro.(in Spanish). Universidad de Barcelona(Spain): 140 pp.
Isfendiyaroglu, M., 1999. Investigations on cu-tting propagation of the mastic tree (Pistacialentiscus var. Chia Duham.) and anatomical-physiological study of root formation. Ege Uni-versity, Faculty of Agriculture, Department ofHorticulture, Bornova-Izmir (Turkey): 123 pp.
Saadat, Y.A., 1998. In vitro culture of Persianwalnut (Juglans regia L.). University CollegeDublin, Dublin (Ireland): 176 pp.
Carob orchard at Ibiza, Spain
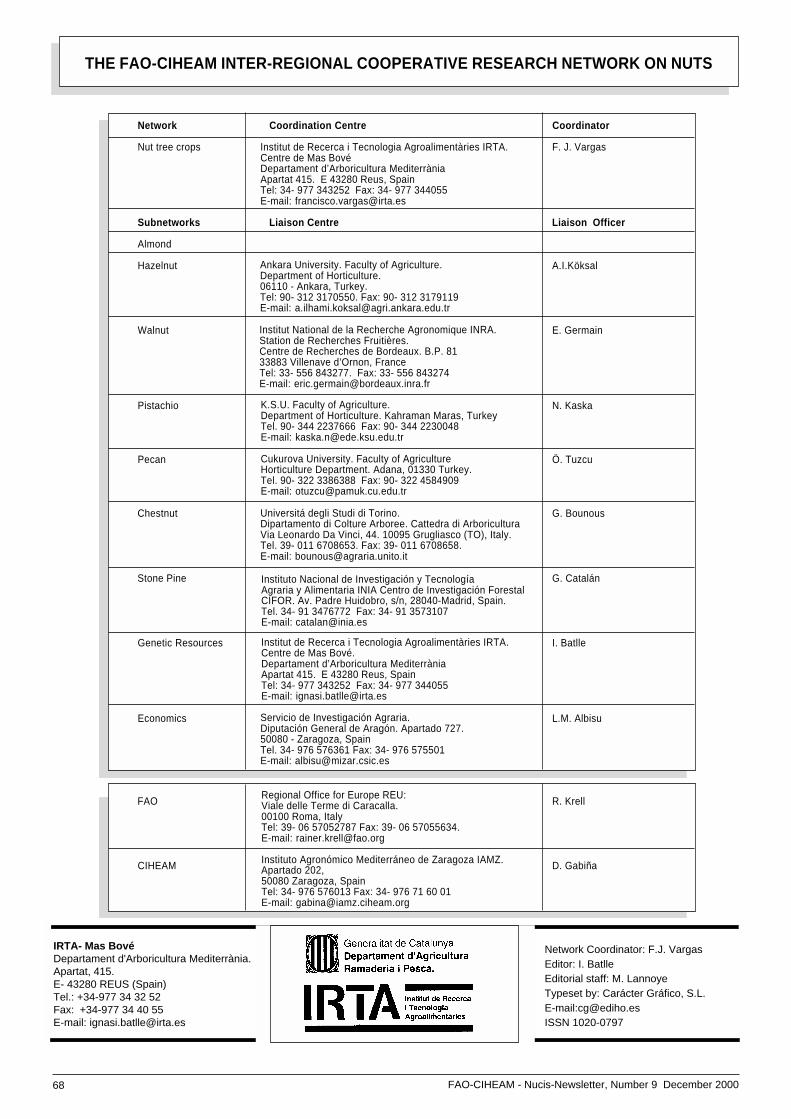
68 FAO-CIHEAM - Nucis-Newsletter, Number 9 December 2000
THE FAO-CIHEAM INTER-REGIONAL COOPERATIVE RESEARCH NETWORK ON NUTS
Network Coordinator: F.J. VargasEditor: I. BatlleEditorial staff: M. LannoyeTypeset by: Carácter Gráfico, S.L.E-mail:[email protected] 1020-0797
Network Coordination Centre Coordinator
Nut tree crops F. J. Vargas
Subnetworks Liaison Centre Liaison Officer
Almond
Hazelnut A.I.Köksal
Walnut E. Germain
Pistachio N. Kaska
Pecan Ö. Tuzcu
Chestnut G. Bounous
Stone Pine G. Catalán
Genetic Resources I. Batlle
Economics L.M. Albisu
FAO R. Krell
CIHEAM D. Gabiña
IRTA- Mas BovéDepartament d'Arboricultura Mediterrània.Apartat, 415.E- 43280 REUS (Spain)Tel.: +34-977 34 32 52Fax: +34-977 34 40 55E-mail: [email protected]
Institut de Recerca i Tecnologia Agroalimentàries IRTA.Centre de Mas BovéDepartament d’Arboricultura MediterràniaApartat 415. E 43280 Reus, SpainTel: 34- 977 343252 Fax: 34- 977 344055E-mail: [email protected]
Ankara University. Faculty of Agriculture.Department of Horticulture.06110 - Ankara, Turkey.Tel: 90- 312 3170550. Fax: 90- 312 3179119E-mail: [email protected]
Institut National de la Recherche Agronomique INRA.Station de Recherches Fruitières.Centre de Recherches de Bordeaux. B.P. 8133883 Villenave d’Ornon, FranceTel: 33- 556 843277. Fax: 33- 556 843274E-mail: [email protected]
K.S.U. Faculty of Agriculture.Department of Horticulture. Kahraman Maras, TurkeyTel. 90- 344 2237666 Fax: 90- 344 2230048E-mail: [email protected]
Cukurova University. Faculty of AgricultureHorticulture Department. Adana, 01330 Turkey.Tel. 90- 322 3386388 Fax: 90- 322 4584909E-mail: [email protected]
Universitá degli Studi di Torino.Dipartamento di Colture Arboree. Cattedra di ArboriculturaVia Leonardo Da Vinci, 44. 10095 Grugliasco (TO), Italy.Tel. 39- 011 6708653. Fax: 39- 011 6708658.E-mail: [email protected]
Instituto Nacional de Investigación y TecnologíaAgraria y Alimentaria INIA Centro de Investigación ForestalCIFOR. Av. Padre Huidobro, s/n, 28040-Madrid, Spain.Tel. 34- 91 3476772 Fax: 34- 91 3573107E-mail: [email protected]
Institut de Recerca i Tecnologia Agroalimentàries IRTA.Centre de Mas Bové.Departament d’Arboricultura MediterràniaApartat 415. E 43280 Reus, SpainTel: 34- 977 343252 Fax: 34- 977 344055E-mail: [email protected]
Servicio de Investigación Agraria.Diputación General de Aragón. Apartado 727.50080 - Zaragoza, SpainTel. 34- 976 576361 Fax: 34- 976 575501E-mail: [email protected]
Regional Office for Europe REU:Viale delle Terme di Caracalla.00100 Roma, ItalyTel: 39- 06 57052787 Fax: 39- 06 57055634.E-mail: [email protected]
Instituto Agronómico Mediterráneo de Zaragoza IAMZ.Apartado 202,50080 Zaragoza, SpainTel: 34- 976 576013 Fax: 34- 976 71 60 01E-mail: [email protected]





























