Embed Size (px)
Citation preview

Comments
238
Conservation Biology, Pages 238–243Volume 16, No. 1, February 2002
Ecoregions in Ascendance: Reply to Jepson and Whittaker
ERIC WIKRAMANAYAKE,* ERIC DINERSTEIN,* COLBY LOUCKS,* DAVID OLSON,* JOHN MORRISON,* JOHN LAMOREUX,* MEGHAN M
C
KNIGHT,* ANDPRASHANT HEDAO†
*Conservation Science Program, World Wildlife Fund – United States, 1250 24th Street NW, Washington, D.C. 20037, U.S.A.†Environmental Systems Research Institute, Inc., Redlands, CA 92373, U.S.A.
Introduction
In their essay, Jepson and Whittaker (2002 [this issue])express three aims: (1) to place the World Wildlife Fund( WWF ) ecoregions (Olson et al. 2001; Wikramanayakeet al. 2001) in context with alternative systems for set-ting conservation priorities; (2) to express concern aboutthe explicitness, transparency, and repeatability of themethods employed to define and delineate ecoregions;and (3) to ask whether the WWF ecoregions improve uponexisting schemes of representing biodiversity (which theyattempt to investigate by performing a qualitative test withIndonesia as a case study).
Their essay contains a number of misrepresentationsof our ecoregion approach. In the interest of brevity, weaddress only the major issues in this reply. We begin byarguing for the value of ecoregions, outlining the ratio-nale behind the ecoregion framework and the delinea-tion process, and comparing these with other efforts. Theapproach is detailed explicitly by Wikramanayake et al.(2001) and others ( Ricketts et al. 1999; Abell et al. 2000;Dinerstein et al. 2000).
Why Ecoregions?
Ecoregions may be a relatively new concept in conserva-tion biology, but they are based on classical biogeogra-phy. Although a single institution like WWF may actuallyproduce an ecoregion map, the ecoregions themselves
are developed through extensive collaboration with bio-geographers, taxonomists, conservation biologists, andecologists from around the world. Since 1999, three ofthe leading international nongovernmental organizations—WWF, The Nature Conservancy (TNC), and ConservationInternational (CI )—and many other conservation partnersaround the world have identified ecoregions or larger bio-geographic units such as hotspots (CI ) as an appropriatescale for conservation priority setting and planning. Theseorganizations have found that ecoregions provide the crit-ical spatial link between global priority-setting efforts andsite-based assessments.
Perhaps the most attractive feature of ecoregions isthat they define biogeographic units that are most suit-able for meeting a fundamental goal of biodiversity con-servation: the representation of all habitats and distinctbiotas in networks of conservation areas. The use of eco-regions allows us to avoid redundancy and enhance com-plementarity in the design of reserve networks betterthan do approaches that rely on political units to deter-mine conservation priorities. Developing conservationstrategies within a framework of ecoregions also allowsus to address the ecological processes that maintain bio-diversity and to protect populations of species with largespatial needs that cannot be accommodated at the sitescale or that do not adhere to political boundaries ( Din-erstein et al. 2000; Groves et al. 2000; Mace et al. 2000).
The magnitude of the shift to ecoregions for conserva-tion planning is worth noting for the benefit of conser-vation biologists unaware of this trend. The WWF hastargeted 189 of the planet’s 867 terrestrial ecoregions, 15freshwater ecoregions, and 24 marine ecoregions for suchanalyses. Within a few years, TNC plans to finish ecoregionstrategies for all 62 of its U.S. ecoregions, and CI plans tocomplete similar analyses for all 25 hotspots ( Myers et al.2000), many of which overlap with the Global 200 Ecore-gions identified by WWF (Olson & Dinerstein 1998). The
Address correspondence to E. Dinerstein, email [email protected] submitted August 13, 2001; revised manuscript accepted Au-gust 20, 2001.

Conservation BiologyVolume 16, No. 1, February 2002
Wikramanayake et al. Ecoregions in Ascendance
239
WWF is collaborating with both organizations on several ofthese efforts. Regional analyses, such as those conductedfor the Nearctic, Neotropics, and Indo-Pacific ( Diner-stein et al. 1995; Ricketts et al. 1999; Abell et al. 2000;Wikramanayake et al. 2001), constitute the foundationfor the WWF’s efforts and have received favorable re-views in the conservation biology literature ( Pimm 1999;Terborgh 1999; Caicco 2000; Cassin 2000; Ebersole 2000;Ruhren 2000). Other regional assessments in this seriesare in press ( Burgess et al. 2002) or in preparation (Afro-tropics freshwater, Palearctic, Neotropics). Given thetrend among conservation nongovernmental organiza-tions, we predict that within 2 years, submissions on thepractice of ecoregion-based conservation will become in-creasingly prevalent in peer-reviewed journals such as
Conservation Biology.
Even Jepson (1999) acknowledges the usefulness ofour ecoregion framework to the conservation of biodi-versity. In a rather critical review of BirdLife International’slandmark publication,
Endemic Bird Areas of the World:Priorities for Biodiversity Conservation
, he writes: “Inshort, the meat in this book does not justify the 846 pages.WWF–US has incorporated endemic bird areas (EBAs) inthe ecoregion approach which combines a representa-tion and hotspot approach in the Global 200. Ecoregionsprobably represent the culmination of the biologist’sperspective on biodiversity priorities.”
Jepson confuses, as he has elsewhere ( Jepson & Can-ney 2001), representation with prioritization. The Das-mann-Udvardy and Omernik-Bailey assessments are rep-resentation systems, as are WWF’s ecoregions (Olson etal. 2001). In contrast, strategic prioritization highlightsareas essential for the conservation of irreplaceable andthreatened biodiversity (Margules & Pressey 2000). Con-servation International’s hotspots (Myers et al. 2000),WWF’s Global 200 Ecoregions (Olson & Dinerstein 1998),BirdLife International’s EBAs (Stattersfield et al. 1998),and the World Conservation Union ( IUCN) and WWF’scenters of plant diversity ( IUCN & WWF 1995) exem-plify prioritization systems that are highly congruent de-spite their differing scales of analysis ( da Fonseca et al.2000).
Ecoregions in Context
Here we address Jepson and Whittaker’s (2002 [this is-sue]) suggestion that WWF ecoregions are inferior to ex-isting systems of classification. Jepson’s specific criticismof WWF’s ecoregions is that they conflict with the Das-mann-Udvardy system of biogeographic representation.They do not. Rather, ecoregions represent continuousupdating and improvement of this system, with the goalof reconciling the numerous idiosyncratic approachesthat have developed over the last few decades into astandardized global scheme. They also introduce the con-
cept to the current generation of conservation plannersand incorporate aquatic systems. Where the implemen-tation of existing frameworks is weak, the delineation ofecoregions will strengthen it; where implementation isstrong, as in Indonesia, it will reinforce it and address re-maining gaps as demonstrated below.
Because of their biogeographic baseline, ecoregionsreflect the distribution of species and communities moreaccurately than do units derived from models based oncombinations of coarse-scale biophysical features suchas rainfall and temperature, which Jepson and Whittakerview as objective, transparent, and repeatable. Bailey’s(1998) terrestrial ecoregions are derived primarily fromsuch models and are based on global-scale biophysicaldata. The resulting ecoregion units are excessively largefor regional planning and do not effectively reflect bio-geographic patterns determined largely by historical pro-cesses, particularly in island systems such as in the Indo-Pacific. Bailey’s system also ignores many tropical islandswith extraordinary levels of endemism but too small insize to be classified in his approach.
Dasmann (1974) and Udvardy (1975) were the first tomap a global system of biogeographic units (referred toas biotic provinces) within a framework of realms andbiomes. Their biotic provinces are based on known bio-geographic patterns, rather than on biophysical models,and are thus comparable to ecoregions in their deriva-tion and intent (i.e., identifying distinct biotas). But thebiogeographic resolution of Dasmann and Udvardy’s bi-otic provinces is considerably coarser than our system ofterrestrial ecoregions. We have identified 867 terrestrialecoregions for the world (Olson et al. 2001), comparedwith the 198 and 193 units of Dasmann’s and Udvardy’ssystems, respectively. The increased resolution is mostapparent in the tropics (between the Tropics of Cancerand Capricorn), where Dasmann (1974) and Udvardy(1975) identified 115 and 117 units, respectively, com-pared with 463 identified in the WWF terrestrial ecore-gion map of the world. A finer-scale example that coverspart of Indonesia—Irian Jaya, the western part of the is-land of New Guinea, and the country of Papua NewGuinea—makes the point even more clearly. Dasmannand Udvardy treat the island of New Guinea as a singleunit, whereas we distinguish 12 ecoregions consisting offour lowland and four montane broadleaf forests, one al-pine scrub ecoregion along the central cordillera, onemangrove forest, one freshwater swamp forest, and onesavanna-grassland ecoregion.
In sum, the Bailey, Dasmann, and Udvardy models, how-ever useful as a broad framework for understanding bioge-ography, are far too coarse for either representation or pri-oritization analyses (sensu Jepson). Thus, MacKinnon andWind (1981) identified a third, finer-scale tier, biounits,which they nested within Udvardy’s biogeographic prov-inces. MacKinnon subsequently applied the biounit (andsubunit) concept to a broader regional area, from Pakistan

240
Ecoregions in Ascendance Wikramanayake et al.
Conservation BiologyVolume 16, No. 1, February 2002
to Papua New Guinea ( MacKinnon & MacKinnon 1986;MacKinnon 1997). Although this step facilitates regionalplanning, the biounit classification mixed distinct biomes,making representation analyses difficult. For instance, asingle biounit can be comprised of mangroves, tropicaldry deciduous forests, thorn scrub, tropical semi-ever-green forests, and tropical moist forests. These habitattypes differ not only in characteristic species, but also inecological dynamics such as species assemblages, natu-ral densities of large herbivores, vectors for seed dis-persal, types of natural-disturbance regimes, response todisturbance, and degree of resiliency. Mangroves are eco-logical systems vastly different from tropical dry thornscrub, and the conservation planning and representationof each is improved by recognizing them as separateecoregions (if, for example, the area of mangroves is suf-ficiently extensive to meet the spatial criterion of anecoregion). Thus, in our analysis of terrestrial ecoregionsof the Indo-Pacific region ( Wikramanayake et al. 2001),we used MacKinnon’s construct of the original habitatto replace biounits with ecoregions within the Udvardyframework.
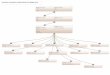
To subdivide Udvardy’s biogeographic realms ( Fig. 1),we identified bioregions. Bioregions are based on acceptedbiogeographic demarcations and have been described indetail, with maps, by Wikramanayake et al. (2001). Withina given bioregion, ecoregions can be nested under differ-ent biomes. In other words, bioregions allow us toachieve a further level of representation in a global or re-gional priority portfolio by capturing a minimum num-ber of ecoregions within each biome occurring in eachbioregion. For example, the Western Ghats Mountainsalong India’s western coast are clothed in rainforests, asare the mountains of central Borneo. But the flora andfauna of the Western Ghats are distinctly different fromthose in central Bornean forests: the former has Nilgiri
tahr (
Hemitragus hylocriusI
), lion-tailed macaques (
Ma-caca silenus
), Nilgiri langurs (
Presbytis johnii
), tigers, anda diverse forest association, whereas the latter has orangu-tans (
Pongo pygmaeus
), gibbons, Sumatran rhinoceros(
Dicerorhinus sumatrensis
), and forest associations domi-nated by
Dipterocarpaceae
spp. To conserve one or theother as a representative rainforest habitat of Asia in a glo-bal or regional conservation strategy would be insufficient.
Our analysis carefully considered previous approaches,such as EBAs (Stattersfield et al. 1998) and centers of plantdiversity ( IUCN & WWF 1995). Where these efforts dis-tinguished distinct biotas at a high level of resolution, webased our ecoregion delineation on them. For example, inwestern Indonesia we used the maps of the original vegeta-tion from MacKinnon (1997), but in eastern Indonesia weused the EBAs (Stattersfield et al. 1998) because we con-sidered them finer and more robust discriminators ofbiodiversity for this system of small island complexes.We did not use hotspots (Mittermeier et al. 1999) be-cause these units are regional in extent and are based ona combination of high biodiversity and threat, ratherthan strictly on biogeography.
Explicitness, Transparency, and Repeatability
We delineated ecoregions according to a set of rules basedon biogeography, habitat type, and elevation. Wikramanay-ake et al. (2001) explicitly describe these rules and definethe biogeographic units and all indices and thresholds thatwere used to delineate and assess ecoregions for the Indo-Pacific region. To make the process of ecoregion delinea-tion transparent and repeatable, we have made available allthe information we used from Wikramanayake et al.(2001). We have also provided lists of all contributors and
Figure 1. Hierarchy of spatial units used in the conservationassessment of the Indo-Pacific.

Conservation BiologyVolume 16, No. 1, February 2002
Wikramanayake et al. Ecoregions in Ascendance
241
reviewers for the respective ecoregions. As in any attemptto represent nature in spatial units, we do not expect unan-imous acceptance of the ecoregion boundaries as we havepresented them. We do hope that many regional biogeog-raphers and conservation biologists, once they understandhow the global map incorporates published, widely usedclassification schemes, will find it acceptable.
Ecoregions Misrepresented: Gestalt, Experts, and Indonesia
If nature could be fit into a one-size-fits-all framework,our task would be simple. Unfortunately, it does not. Amap of eastern Indonesia shows a plethora of lines thathave sprung up around Wallace’s now-famous line—allin attempt to refine Wallace’s original effort and observa-tions. This is because the biodiversity of archipelagos re-sults from a complex suite of biogeographic variablesincluding species vagility and origin, geologic history, geo-morphology, landform, size, and interisland distance. Com-pounding the problem of detecting patterns of biogeogra-phy is a lack of comprehensive and reliable biodiversity datafor many places on Earth. There are no biological databasesthat are uniform in detail and resolution for the entireplanet. Thus, developing a global framework of ecologicalunits necessitates working with uneven information. Attimes, this requires sifting through and combining the bestavailable information from several sources and using vari-ous biological and geophysical parameters. In his afore-mentioned review of EBAs, Jepson (1999) commends usfor incorporating them into the Global 200 Ecoregionanalysis. But in their current essay, Jepson and Whittaker(2002, this issue) criticize it as part of a gestalt approachto ecoregion delineation. Yet they defend the Dasman-Udvardy-MacKinnon-Wind EBA approach that was usedto guide planning for Indonesia’s current protected-areasystem—even though it uses different types of data anddata sources, and “zonal” and “azonal” features ( Jepsonand Whittaker’s words). It is unclear to us how such anapproach is less a gestalt synthesis than ours. We also be-lieve that expert review and input strengthen the valid-ity and accuracy of ecoregions and our approach, despiteJepson and Whittaker’s skepticism.
The WWF Ecoregion Approach and the Goal of Representation of Indonesia’s Protected-Areas System
Much of what has been described above can be viewedas a rather narrow set of academic disagreements bestsuited for a biogeography journal or a debate among par-ticipants at a workshop on ecoregion conservation plan-ning in Indonesia. But Jepson and Whittaker’s critiqueassumes real-world significance when the issue becomesthe relationship between designing biogeographic frame-works and establishing representative networks of pro-tected areas. They claim that “Given that the existingschemes, despite flaws, have achieved their purpose inIndonesia . . . we question the wisdom of introducingthe WWF ecoregion system.” This statement is the ex-tent of their “test” of the efficacy of ecoregions againstthe existing planning system. We strongly disagree withtheir opinion, believe that the ecoregion approach im-proves on the previous framework, and present a quanti-tative test of our own below.
Ecoregions are ultimately tools for conservation plan-ning. Although the ecoregion analyses point out many ofthe strengths of the existing design of a protected-areassystem for Indonesia, it also highlights some significantgaps. A recent assessment by Derek Holmes ( by Wikra-manayake et al. 2001) projects that, at current rates of de-struction, the remaining intact lowland forests ofSumatra and Kalimantan will disappear within the next 5and 9 years, respectively. Taking this analysis further, wecan compare current levels of protection in lowland andmontane forest ecoregions of Sumatra, Java, Borneo, andSulawesi. Almost 15% of the montane forests are con-tained within IUCN category I–IV protected areas, butless than 6% of the remaining lowland forests are pro-tected ( Table 1). Thus, protection is biased toward inclu-sion of montane forests, which are largely unproductivefor agriculture and too steep for logging, plantations, andsettlements, whereas the rich lowland rainforests of In-donesia remain underrepresented within the protected-areas system. An analysis based on ecoregions illustratesthis disparity in representation better than one based onclassification schemes promoted by Jepson and Whit-taker that do not effectively distinguish montane andlowland habitats.
Table 1. Comparison of degree of protection (PA, protected area) in lowland and montane forests of Java, Sumatra, Borneo, and Sulawesi.
Protected areas (km
2
)
*
Total area of lowland andmontane forests (km
2
)Forest IUCN I-VIII IUCN I-IV I–VIII in PA (%) I–IV in PA (%)
Lowland 80,029 68,519 1,221,400 6.6 5.6Montane 53,630 46,056 312,200 17.2 14.8
*
This analysis uses World Conservation Monitoring Centre and Asian Bureau for Conservation (1997 ) protected areas and does not includethose designated as proposed.

242
Ecoregions in Ascendance Wikramanayake et al.
Conservation BiologyVolume 16, No. 1, February 2002
Jepson and Whittaker also suggest that ecoregions candivert conservation efforts from biologically importantareas. They are concerned that treatment of the lowlandforests of Sulawesi as a single ecoregion may suggest toplanners that targeting resources to any peninsula in Su-lawesi is equally good, thereby detracting from impor-tant and high-priority conservation areas such as the north-ern Minahasa peninsula. In reality, any intelligent,competent conservation planner will look carefully at bio-geographic patterns to ensure adequate representationwithin a given ecoregion, typically by creating biogeo-graphic subdivisions, a method used by the WWF, CI, andTNC (see Dinerstein et al. 2000).
Jepson and his colleagues have over the past 2 yearscriticized the prioritization efforts of BirdLife Interna-tional’s EBAs, CI’s hotspots, and WWF’s terrestrial ecore-gion analysis without offering the scientific community acompelling alternative for representation of global biodi-versity or priority setting. In their critiques of all three ap-proaches they are literally and figuratively all over themap, offering conflicting recommendations. For example:in a critique of EBAs, Jepson (1999) proposes that WWFecoregions are better units for prioritization and represen-tation than are targets based on range-restricted birds asproposed by BirdLife; in a critique of hotspots, Jepsonand Canney (2001) evaluate hotspots using nonbiologicalcriteria and chastise CI for not doing a better job of repre-senting biodiversity, when hotspots is not a representa-tion scheme but a prioritization scheme. Finally, in thecurrent essay Jepson and Whittaker criticize the WWFecoregion approach in favor of more “transparent, less ge-stalt-like” approaches for setting priorities such as EBAs,when the WWF ecoregions approach is first and foremostan attempt to improve representation of distinct habitatsand communities.
Ecoregions have already helped improve conservationplanning by catalyzing debate on critical issues such asrepresentation of biodiversity and large-scale ecologicalprocesses. In the case of Indonesia, and we suspect inmany other biologically important places on Earth, theincreased use of ecoregions or other biogeographic unitswill assist conservation planners in making a compelling,rigorous case to decision-makers and donors for greatlyexpanded conservation efforts. The next phase of ourwork is to use ecoregions as the basis for a “grand syn-thesis” of global priority-setting approaches so that wecan speak with one voice to decision-makers and donorsabout the fate of the biota on our planet. We hope Jep-son and his colleagues will join us in this important stepto save life on Earth.
Literature Cited
Abell, R. A., D. M. Olson, E. Dinerstein, P. T. Hurley, J. T. Diggs, W. Eich-baum, S. Walters, W. Wettengel, T. Allnutt, C. J. Loucks, and P. Hedao.
2000. Freshwater ecoregions of North America: a conservation as-sessment. Island Press, Washington, D.C.
Bailey, R. G. 1998. Ecoregions: the ecosystem geography of the oceansand continents. Springer Verlag, New York.
Burgess, N., E. Underwood-Russell, J. D’Amico, D. M. Olson, E. Diner-stein, and I. Itoua. 2002. Terrestrial ecoregions of Africa: a conser-vation assessment. Island Press, Washington, D.C.
Caicco, S. 2000. Review of terrestrial ecoregions of North America: aconservation assessment. Natural Areas Journal
20(4):
390.Cassin, J. L. 2000. Filling the gap in conservation strategies: a meso-
scale tool for biodiversity assessment and conservation. Conserva-tion Biology
14:
1555–1556.da Fonseca, G. A. B., et al. 2000. Following Africa’s lead in setting pri-
orities. Nature
405:
393–394.Dasmann, R. F. 1974. Biotic provinces of the world: further develop-
ment of a system for defining and classifying natural regions forpurposes of conservation. Occasional paper 9. World ConservationUnion, Gland, Switzerland.
Dinerstein, E., D. M. Olson, D. Graham, A. Webster, S. Pimm, M. Book-binder, and G. Ledec. 1995. A conservation assessment of the ter-restrial ecoregions of Latin America and the Caribbean. WorldBank, New York.
Dinerstein, E., G. Powell, D. M. Olson, E. Wikramanayake, R. Abell,C. Loucks, E. Underwood, T. Allnutt, W. Wettengel, T. Ricketts,H. Strand, S. O’Connor, and N. Burgess. 2000. A workbook for con-ducting biological assessments and developing biodiversity visionsfor ecoregion-based conservation. World Wildlife Fund, Washing-ton, D.C.
Ebersole, J. P. 2000. Review of terrestrial ecoregions of North America:a conservation assessment. BioScience
50:
541–543.Groves, C., L. Valutis, D. Vosick, B. Neely, K. Wheaton, J. Touval, and
B. Runnels. 2000. Designing a geography of hope: a practitionershandbook for ecoregional conservation planning. The Nature Con-servancy, Washington, D.C.
Jepson, P. 1999. [Review of] Endemic bird areas of the world: priori-ties for biodiversity conservation. Ibis
141:
163.Jepson, P. 2001. Global biodiversity plan needs to convince local pol-
icy-makers. Nature
409:
12.Jepson, P., and S. Canney. 2001. Biodiversity hotspots: hot for what?
Global Ecology & Biogeography
10:
225–227.Jepson, P., and R. J. Whittaker. 2002. Ecoregions in context: a cri-
tique with special reference to Indonesia. Conservation Biology
16:
42–57.Mace, G. M., et al. 2000. It’s time to work together and stop duplicat-
ing conservation efforts. Nature
405:
393.MacKinnon, J. 1997. Protected areas systems review of the Indo-
Malayan realm. The Asian Bureau for Conservation and The WorldConservation Monitoring Centre/World Bank Publication, Canter-bury, United Kingdom.
MacKinnon, J., and K. MacKinnon. 1986. Review of the protected ar-eas system in the Indo-Malayan Realm. World Conservation Union,Gland, Switzerland, and United Nations Environmental Programme,Cambridge, United Kingdom.
MacKinnon, J., and J. Wind. 1981. Birds of Indonesia. Special reportF0/INS/78/061. Available from the Food and Agriculture Organiza-tion of the United Nations, Rome.
Margules, C. R., and R. L. Pressey. 2000. Systematic conservation plan-ning. Nature
405:
243–253.Mittermeier, R. A., N. Myers, P. Gil, and C. G. Mittermeier, editors.
1999. Hotspots, Earth’s biologically richest and most endangeredterrestrial ecosystems. CEMEX, Mexico City.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca,and J. Kent. 2000. Biodiversity hotspots for conservation priorities.Nature
403:
853–858.Olson, D. M., and E. Dinerstein. 1998. The Global 200: a representa-
tion approach to conserving the Earth’s distinctive ecoregions.Conservation Biology
12:
502–515.

Conservation BiologyVolume 16, No. 1, February 2002
Wikramanayake et al. Ecoregions in Ascendance
243
Olson, D. M., et al. 2001. Terrestrial ecoregions of the world: a newmap of life of Earth. BioScience
51:
933–938.Pimm, S. 1999. Seeing both the woods and the trees: a novel approach
to classifying biodiversity may aid attempts at conservation. Nature
402:
853–854.Ricketts, T. H., E. Dinerstein, D. M. Olson, C. J. Loucks, W. Eichbaum,
D. DellaSala, K. Kavanagh, P. Hedao, P. T. Hurley, K. M. Carney,R. Abell, and S. Walters. 1999. Terrestrial ecoregions of NorthAmerica: a conservation assessment. Island Press, Washington, D.C.
Ruhren, S. 2000. Review of terrestrial ecoregions of North America: aconservation assessment. World Birdwatch
22(4):
23–24.Stattersfield, A. J., M. J. Corsby, A. J. Long, and D. C. Wege. 1998. Glo-
bal directory of endemic bird areas. BirdLife International, Cam-bridge, United Kingdom.
Terborgh, J. 1999. Requiem for nature. Island Press, Washington, D.C.
Udvardy, M. D. F. 1975. A classification of the biogeographical prov-inces of the world. Occasional paper 18. World ConservationUnion, Gland, Switzerland.
Wikramanayake, E., E. Dinerstein, C. J. Loucks, D. M. Olson, J. Morri-son, J. Lamoreux, M. McKnight, and P. Hedao. 2001. Terrestrialecoregions of the Indo-Pacific: a conservation assessment. IslandPress, Washington, D.C.
World Conservation Monitoring Centre ( WCMC) and Asian Bureau forConservation. 1997. Protected areas system review of the Indo-Malayan realm. J. MacKinnon, editor. Asian Bureau for Conserva-tion Limited, Canterbury, England.
World Conservation Union ( IUCN ) and World Wildlife Fund ( WWF ).1995. Centres of plant diversity: a guide and strategy for their con-servation. 2. Asia, Australasia and the Pacific. IUCN, Cambridge,United Kingdom.