Embed Size (px)
Citation preview

Kate Block
1
The Intermediate Disturbance Hypothesis: Theoretical Underpinnings, Validity and Applications to North American Grasslands
Introduction Coexistence between species occurs only under limited conditions and interspecific
competition generally reduces biodiversity unless a community is acted upon by an external force which reduces interspecific competition or its effects (Petraitis et al 1989, p. 394). Disturbance often provides this type of external intervention. The Intermediate Disturbance Hypothesis states that diversity (species richness) is maximized when disturbance occurs at an intermediate scale (Connell 1978, p. 1303) and attempts to describe the manner in which disturbance maintains biodiversity within ecological communities. This paper is an exploration of interspecific competition theory, the Intermediate Disturbance Hypothesis (IDH), the evidence that supports and refutes the IDH, and the applications to North American grasslands and their management. To conclude the paper, general recommendations regarding the use of prescribed disturbance for prairie preservation, restoration, and management are provided. Theoretical Background: Interspecific Competition All populations of organisms in nature demonstrate the inherent propensity for unlimited exponential growth. However, as modeled by the logistic equation, abiotic and biotic factors present in their environments limit their capacity to increase (Crombie 1947, p. 45). These factors can be described by three main categories (Crombie 1947, p. 45): (1) Factors arising from the organism itself – its adaptation to the habitat in which it lives, life history traits, and reproductive characteristics. (2) Abiotic factors – weather, climate, soil characteristics, and solar radiation. (3) Biotic factors – organism-‐organism interactions and alterations organisms make to the physical environment surrounding them. Biotic factors frequently influence populations in a density dependent manner. Depending upon their specific nature, disturbances, which will be discussed in detail later in this paper, may be categorized as either abiotic or biotic factors although they are most commonly abiotic. While density independent factors may dictate the presence and nature of interspecific competition as well as its eventual outcomes, only density dependent factors are directly involved in interspecific competition (Crombie 1947, p. 46). Factors that increase the ratio of organisms to environmental resources increase competition while factors that decrease the ratio of organisms to environmental resources decrease competition (Crombie 1947, p. 46). When two or more species with similar niche requirements are present in a given environment, interspecific competition occurs when the ratio of organisms to environmental resources increases above a certain critical level (Crombie 1947, p. 48). One species may be a superior competitor to another species by either possessing a superior rate of reproduction and survival or via a superior ability for interference (Crombie 1947, p. 49). Often a species’ superiority will depend upon the exact environmental conditions present and, thus, because environmental conditions often fluctuate, on species may be the superior competitor under one specific set of environmental conditions while another may be the superior competitor under a different set of environmental conditions (Crombie 1947, p. 49). In plants, the necessary factor – light, water, a particular nutrient – in shortest supply relative to total demand is generally the instigator of intraspecific competition (Rubel 1953 qtd. in Crombie 1947,

2
p. 51). Because an actual physical struggle between plants almost never occurs, intraspecific competition in plants almost solely “arises from the reaction of one plant upon the physical factors about it and the effect of these modified factors upon its competitors” (Rubel 1953 qtd. in Crombie 1947, pp. 50-‐51). As a superior competitor’s dominance within its community increases via the elimination of inferior competitors, intraspecific competition decreases and a climax community emerges (Crombie 1947, p. 51). The classical Lotka-‐Volterra model of interspecific competition is a logical extension of the logistic equation that describes the competitive interaction between two species whose growth rates are density dependent (Neuhauser & Pacala 1999, p. 1227). The two equations comprising the classical Lotka-‐Volterra model are as follows (Malcolm 3 February 2009, slide 8):
For species 1: dN1/dt = r1N1[K1-‐N1-‐(αN2/K1)] For species 2: dN2/dt = r2N2[K2-‐N2-‐(βN1/K2)]
In each of these equations, N represents the size of the population of species 1 or species 2, r represents the intrinsic rate of natural increase for species 1 or species 2, K represents the carrying capacity of species 1 or species 2, α represents the competition coefficient for species 1, and β represents the competition coefficient for species 2 (Malcolm 3 February 2009, slide 8). To clarify, the term αN2 represents the effect of species 2 on species 1 and the term βN1 represents the effect of species 1 on species 2 (Malcolm 3 February 2009, slide 9). If α or β is less than 1, intraspecific competition has a greater impact on the species’ (1 or 2, respectively) population dynamics than interspecific competition and if α or β is greater than 1, interspecific competition has a greater impact on the species’ (1 or 2, respectively) population dynamics than intraspecific competition (Malcolm 3 February 2009, slide 9).
Four different outcomes are possible in the Lotka-‐Volterra model of interspecific competition (Malcolm 3 February 2009): (1) Competitive exclusion in which species 1 prevails and species 2 is extirpated (or extinct) – occurs when K1 is greater than K2/β and K1/β is greater than K2. (2) Competitive exclusion in which species 2 prevails and species 1 is extirpated (or extinct) – occurs when K2 is greater than K1/α and K2/α is greater than K1. Competitive exclusion of one species by the other can occur even if the effect of intraspecific competition on the “winner” is stronger than its impact (via interspecific competition) on the “loser.” (3) Either species 1 or species 2 can “win” – the effect of interspecific competition is greater than the effect of intraspecific competition for both species and, therefore, the ultimate outcome is dependent upon the initial densities of each species. This outcome is frequently termed founder control (Neuhauser & Pacala 1999, p. 1227). (4) Coexistence of species 1 and species 2 – occurs when both species have less of a competitive effect on each other than they do on themselves. In other words, for both species 1 and species 2, the effects of intraspecific competition supersede the effects of interspecific competition (Neuhauser & Pacala 1999, p. 1227). Each of these possible outcomes is illustrated in the figure below (Malcolm 3 February 2009, slide 17).

3
Two species can compete for the same resources and still coexist as long as a sufficient degree
of niche displacement is present (Neuhauser & Pacala 1999, p. 1227-‐1228); the habitat in which species 1 and species 2 exist is such that species 1 and species 2 are not limited by the same resource (Begon et al 2006, p. 263; Neuhauser & Pacala 1999, p. 1227). The resulting outcomes of a particular interaction between two species can be determined by graphing the zero isoclines – a connected line of points at which population growth for species 1 or species 2 is zero (Malcolm 3 February 2009, slide 10) – of each species (Begon et al 2006, p. 236). Because coexistence is only possible in one of the four potential outcomes of interspecific competition, we can easily recognize that interspecific competition frequently reduces biodiversity unless a community is acted upon by an external force which reduces interspecific competition or its effects (Petraitis et al 1989, p. 394). Disturbance often provides this type of external intervention. Disturbance and the Maintenance of Biodiversity Disturbance and predation have generally been cited as the two primary mechanisms by which interspecific competition’s effects are muted (Petraitis et al 1989, p. 394). While predation – which acts as a form of biotic disturbance (Mackey & Currie 2001, p. 3480) – reduces interspecific competition via selective mortality (of prey species), disturbance reduces interspecific competition via the mass mortality that follows random, localized events (Petraitis et al 1989, p. 394). Disturbance is typically defined as “any relative discrete event in time that disrupts ecosystem, community, or population structure and changes resources, substrate availability, or the physical environment” (White & Pickett 1985 qtd. in Petraitis et al 1989, p. 395). Alternately, disturbance has been more explicitly conceptualized as “a temporally discrete even that abruptly kills or displaces individuals or that directly results in the loss of biomass” (Mackey & Currie 2001, p. 3480). Fire, intense storm events, landslides, flooding, and extreme drought are all examples of abiotic disturbance and directly or indirectly change resource availability (Mackey & Currie 2001, p. 3480). The Intermediate Disturbance Hypothesis
Figure 1: Outcomes generated by the classical Lotka-‐Volterra model of interspecific competition, as depicted by the zero isoclines for species 1 and species 2. In (a), species 1 prevails and species 2 is eliminated. In (b), species 2 prevails and species 1 is eliminated. In (c), an unstable equilibrium is generated – indicated by the arrows pointing away from the intersection of the two isoclines – and either species 1 or 2 will win, depending upon initial conditions. In (d), a stable equilibrium is generated – indicated by the arrows pointing toward the intersection of the two isoclines – and coexistence occurs.

4
The vast majority of ecology textbooks and research-‐based literature credit Joseph H. Connell with the formulation of the intermediate disturbance hypothesis (IDH) in his 1978 article “Diversity in Tropical Rain Forests and Coral Reefs,” published in Science (Wilkinson 1999, p. 145; Connell 1978). Although several scholars have questioned the legitimacy of recognizing Connell as the chief and original author of the IDH because several other authors – most notably Grime and Horn – had published similar speculations a few years prior to Connell’s paper (e.g. Wilkinson 1999, pp. 145-‐146; Fox 1979, p. 1345), in this paper, the account of the IDH that I give is Connell’s. In the early 1960s, Connell became curious about how the biodiversity in tropical ecosystems arose and how it was maintained (Connell 1987, p. 1). He had subscribed to popular theoretical position of the time: Ecological communities were conceived as highly regulated at a particular equilibrium state and their members (various species) were co-‐adapted such that each of their niches fit neatly among the others like an elaborate jigsaw puzzle (Connell 1987, p. 1). In this view, a range of “forces” existed within ecological communities and prevented any single species from becoming either too common (dominant) or too rare (Connell 1987, p. 1). In his own words, after witnessing a hurricane decimate first a coral reef and the his rain forest study plots, Connell decided that perhaps he “should entertain some alternative hypothesis about mechanisms maintaining tropical diversity instead of clinging to the one I had brought to the tropics from my temperate ivory tower” (Connell 1987, p. 1). Thus, the IDH was born. In his landmark 1978 paper in Science, Connell actually presented six different hypotheses – three equilibrium hypotheses and three non-‐equilibrium hypotheses – that attempt to explain the diversity of tropical rain forests and coral reefs (pp. 1302-‐1303). Connell clarifies that, while they may apply to a wide variety of ecosystems, he only intends to apply these hypotheses to tropical communities (1978, p. 1302). In reality, each of the six hypotheses may indeed contribute to the preservation of biodiversity in the tropics; however, Connell asserts that the relative importance of each may vary dramatically, with non-‐equilibrium processes likely to be the most vital (1978, pp. 1308-‐1309). Similarly, even though these hypotheses may indeed accurately account for the tropical communities’ biodiversity, biodiversity may also be generated and maintained by a variety of other mechanisms (Connell 1978, p. 1302). It is beyond the scope of this paper to discuss each of these six hypotheses in detail. I will instead focus on a single hypothesis which is arguably presently the most widely espoused; the IDH. Simply put, the IDH states that diversity (species richness) should be highest when disturbance occurs at an intermediate scale (Connell 1978, p. 1303). The rationale undergirding this claim is as follows: (1) A trade-‐off between competitive ability and resistant to or tolerant of disturbance exists (Petraitis 1989, p. 397; Mackey & Currie 2001, p. 3479; Connell 1978, p. 1303). This assumption presumes that a species cannot be both a superior competitor and superiorly adapted to disturbance (e.g., by being an especially good disperser/colonizer or unlikely to experience disturbance-‐induced mortality). (2) When disturbance is high – frequent (frequency is evaluated on the basis of the generation times of organisms within the community of interest), intense (conceptualized as total area affected by disturbance multiplied by frequency of disturbance), or severe – few species will be present (low species richness) because only those species that are sufficiently tolerant of or resistant to disturbance can persist (Connell 1978, p. 1303; Petraitis et al 1989, p. 397; Roxburgh et al 2004, p. 360). (3) When disturbance is low – infrequent, of low intensity, or mild – few species will be present (low species richness) because only those species that are superior competitors can persist (Connell 1978, p. 1303; Petraitis et al 1989, p. 397). Connell describes several types of superior competitors – those that are “most efficient in exploiting limited resources,” those that are “most efficient in

5
interfering with other species,” those that are “most resistant to damage or to death caused by physical extremes or natural enemies” (Connell 1978, p. 1303). (4) When disturbance is intermediate – moderate frequency, intensity, or severity – more species will be present (higher species richness) because a wider range of species are able to tolerate environmental and biological conditions (Connell 1978, p. 1303; Petraitis et al 1989, p. 397). “There is a balance between [the] competitive exclusion” that occurs when disturbance is low “and [the] loss of competitive dominants” that occurs when disturbance is high (Mackey & Currie 2001, p. 3479). The relationship between species richness and disturbance is depicted in the figure below.
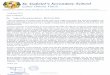
Figure 2: Graphical representations of the IDH. In (a), species diversity should be highest at intermediate levels of disturbance (Hobbs & Huenneke 1992, p. 326). In (b), the x-‐axis represents the mortality resulting from disturbance and the level species diversity is related to competitively dominant species. Species diversity is highest when disturbance-‐induced mortality is at intermediate levels (Hacker & Gaines 1997, p. 1992). Connell provided evidence supporting the IDH from several studies of tropical rain forests in Uganda, Nigeria, Australia, and Guyana (1978, pp. 1303-‐1305). Evidence from research conducted in coral reef communities was more limited. Connell only presented a detailed discussion of supporting evidence from studies conducted in Belize and Australia (1978, pp. 1304-‐1305). Other authors (e.g. Huston 1985) have specifically questioned the validity of the IDH in coral reef communities, concluding that a variety of factors interact to determine the community structure and species richness of coral reefs. In particular, depth and light gradients, the frequency and intensity of disturbance, and competitive displacement are all important factors that impact the coral reef ecosystem (Huston 1985, p. 172). The IDH has enjoyed widespread acceptance from the scientific community over the past three decades for good reason. The support for the IDH present in the literature is overwhelming if not unanimous. Several studies specifically evaluating the IDH’s relevance in North American grassland ecosystems will be reviewed later in this paper. For now, we will turn our attention to a paper published by Roxburgh, Shea, and Wilson in 2004. Using a multi-‐species reciprocal-‐yield law model, the authors obtained the following results (Roxburgh et al 2004, p. 368): When disturbance occurred less than once every fifteen years, competitive exclusion occurred, generating a monoculture. When disturbance occurred in alternate years, many of the highly competitive species could not persist, generating a five-‐species community. Diversity peaked at 25 species when disturbance occurred once every seven years.
6
Additionally, the findings reported by Roxburgh and his colleagues indicate that the coexistence intermediate disturbance generates arises via two primary mechanisms (2004, p. 361): (1) The storage effect – “the species’ attributes allow gains made during favorable growth periods to be ‘stored’ in the population for use during unfavorable periods … and/or periods when the impact of competition is increased.” (2) Relative nonlinearity – “differences in the responses of competitors to fluctuations in resource availability [are] relatively nonlinear.” The functionality of these mechanisms has important implications for species coexistence. First, true, stable coexistence can only occur when competing species’ responses to disturbance differ (Roxburgh et al 2004, p. 361). Therefore, if competing species differ only in their degree of competitive superiority, disturbance cannot generate coexistence (Roxburgh et al 2004, p. 361). Secondly, many of disturbance’s coexistence-‐inducing mechanisms operate at varying degrees, and sometimes in concert, depending upon the particular realities present in a given situation (Roxburgh et al 2004, p. 361).
Research has also suggested that the IDH may only accurately reflect specific situations, communities, or trophic levels or that it may require substantial revision. Wooten (1998) has suggested that the IDH is only valid for sessile species, which tend to comprise the basal trophic level of the community, collecting outside energy and converting it to a form usable by the remaining members of the community. If multi-‐trophic level interactions are considered, the IDH’s validity as an explanatory model is greatly reduced (Wooten 1998, p. 804).
Hacker and Gaines (1997) have suggested that “positive interactions” – commensalisms and mutualisms – between facilitator species may exert a substantial influence on community composition. Taking these positive interactions into account requires minor modifications of the IDH. Essentially, the authors claim that “if the presence of one or more facilitator species allows the persistence of others by ameliorating physical stress or by providing protection from disturbance or predation, the decline in species diversity” that the IDH predicts at high levels of disturbance “may occur under more intense conditions of mortality from physical disturbance, stress, or predation” (Hacker & Gaines 1997, p. 1993). Similarly, the reestablishment of certain species post-‐disturbance may be enhanced by facilitator species via increases in recruitment, colonization, and/or growth rates, increasing diversity across a range of disturbance levels after facilitator species are released from interspecific competition (Hacker & Gaines 1997, p. 1993). The figure below illustrates the potential impact of positive interactions on the diversity-‐disturbance relationship predicted by the IDH (Hacker & Gaines 1997, p. 1992).
Figure 3 (Roxburgh et al 2004, p. 370): Outcomes generated by a multi-‐species reciprocal yield model demonstrate maximum species richness at intermediate frequency of disturbance.

7
While Hacker and Gaines modified model of the IDH may prove an extreme example of the influence positive interactions have on biodiversity, it provides and important insight into one of the IDH’s shortcomings; namely, that when diversity-‐enhancing mechanisms involving facilitator species are not specifically recognized, the predictions made by the IDH may not reflect the true nature of diversity-‐disturbance relationship in the field (Hacker & Gaines 1997, p. 1997). Furthermore, if the potentially critical effects of positive interactions in determining biodiversity are considered and competition displacement (exclusion) is graphed against disturbance-‐induced mortality, a ridge of maximum diversity is revealed (Hacker & Gaines 1997, p. 1998), as depicted in the figure on the following page.
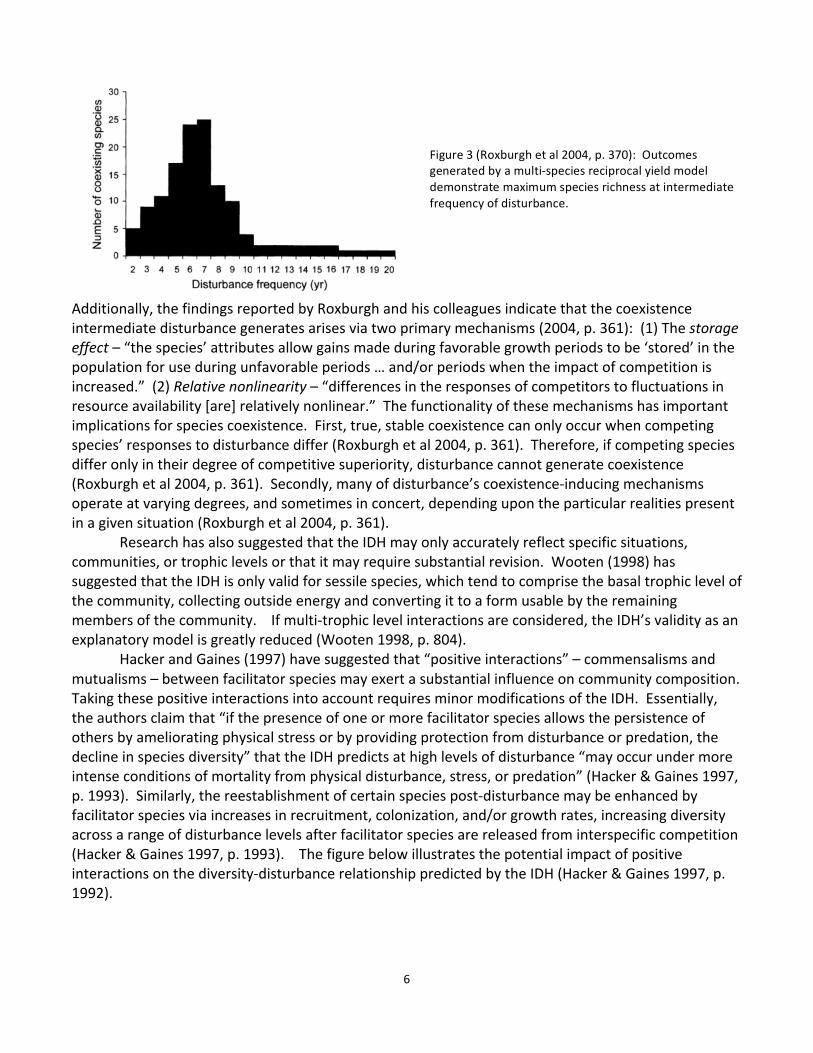
A 2001 review of the IDH conducted by Mackey and Currie reveals that although the contemporary literature accepts that the relationship between biodiversity and disturbance is strong and peaked, an examination of disturbance literature demonstrates that this wholesale acceptance is unjustified (Mackey & Currie 2001, p. 3479). While peaked responses characteristic of the IDH were reported in 11% of evenness studies, 16% of richness studies, and 19% of diversity studies, non-‐significant relationships between disturbance and evenness, richness, or diversity were most often
Figure 4: When released from interspecific competition, facilitator species can both heighten and extend the IDH’s species diversity curve under disturbance conditions generating moderate to high mortality levels by “preventing or ameliorating harsh conditions” and “[creating] new interaction webs” (Hacker & Gaines 1997, p. 1992).
Figure 5: Dynamic equilibrium model (Huston 1979, 1994 qtd. in Hacker & Gaines 1997). Keystone facilitator species are generally most important when the rate of competitive displacement is low but disturbance, stress, or predation (represented by the mortality that results) is high. Conversely, keystone predators are most important when competitive displacement is high and disturbance, stress, or predation is low. Maximum species richness is predicted under the conditions present (pairings of competitive displacement and mortality values) along the ridge shown in the figure.
8
reported (Mackey & Currie 2001, p. 3479 & 3483). Positive and negative monotonic and unimodal relationships between disturbance and evenness, richness, or diversity were also present in the literature Mackey and Currie reviewed. The table below provides data for the various types of relationships found in each of the 85 studies (of an initial sample of 1962 papers and abstracts) examined by the authors (Mackey & Currie 2001, p. 3483).
Mackey and Currie conclude that the substantial proportion of studies reporting a non-‐significant relationship between disturbance and evenness, richness, or diversity suggest that the widespread support IDH enjoys may not be justified as it does not appear to accurately reflect ecological realities in many cases (Mackey & Currie 2001, p. 3487). The median variation explained by disturbance was 53% for species richness and 60% for both evenness and diversity (Mackey & Currie 2001, p. 3488). However, in accordance with Wooten’s (1998) findings, explained variation was significantly higher in studies of sessile autotrophs than for heterotrophs or motile autotrophs (Mackey & Currie 2001, p. 3489), allowing one to speculate that if the IDH is a fairly realistic description of the diversity-‐disturbance relationship in any ecological community, perhaps it is the plant community. Based on their review, the authors argue that while published literature does provide some evidence that disturbance does influence spatial and temporal patterns of diversity to some extent, “diversity-‐disturbance relationships are neither consistently strong nor consistently peaked” and that “disturbance is probably not generally among the most important” factors determining species diversity (Mackey & Currie 2001, p. 3491). The Intermediate Disturbance Hypothesis and North American Grasslands The grassland-‐related literature both supports and refutes the appropriateness of the IDH as a model of the relationship between diversity and disturbance and, thus, offers little in the way of cut and dry conclusions. At times, evidence that corroborates the IDH and evidence that invalidates it are even presented within the same study (e.g. McIntyre & Lavorel 1994). Although the majority of studies that explore disturbance and diversity patterns in grasslands do not provide unambiguous evidence that the IDH accurately reflects the diversity-‐disturbance relationship in these ecosystems, most do
9
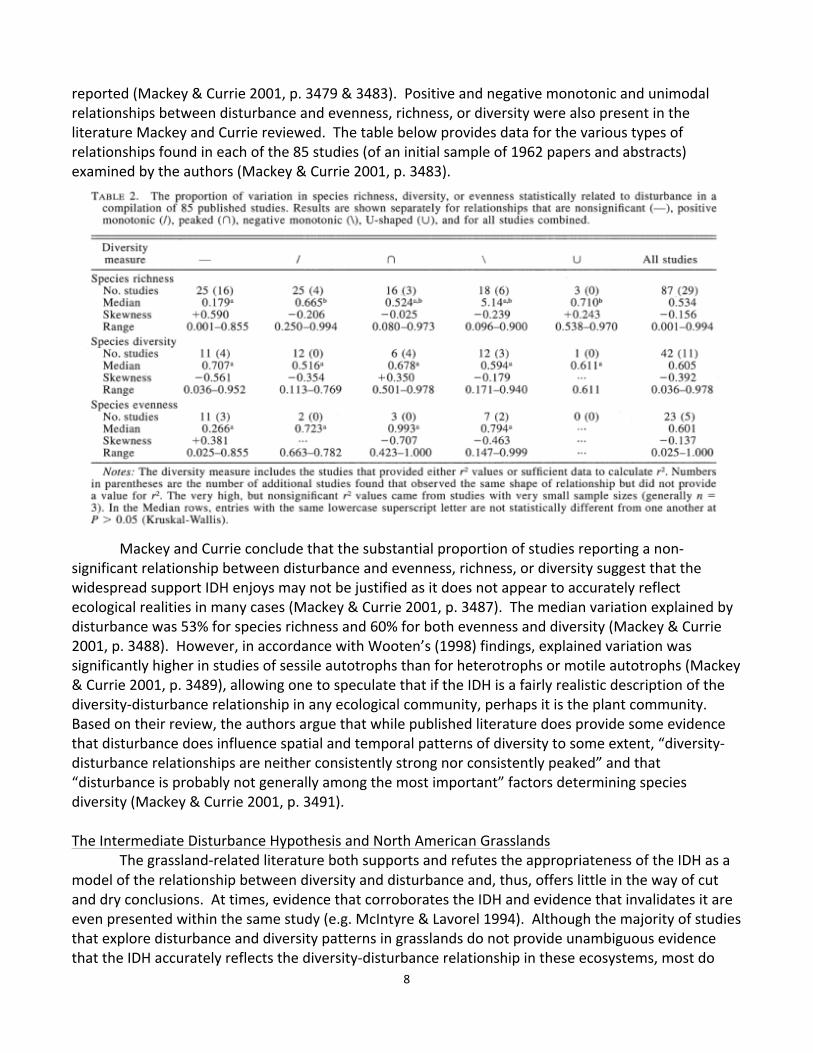
state that disturbance – particularly fire – has profound effects on community composition (Stohlgren et al 1999, Collins 1987, Collins 2000, Gibson 1988, McIntyre & Lavorel 1994, Vujnovic et al 2002, Collins et al 1995). First, let us examine the literature in support of the IDH’s relevance in grasslands. In a study examining the effects of water-‐enrichment (due to increased run off that was anthropogenic in origin), soil disturbance, and grazing by herbivores on total, native, rare, and exotic species richness, intermediate levels of grazing generated the highest total and native species richness (McIntyre & Lavorel 1994, pp. 524-‐525). This same study showed decreased total, native, and rare species richness with increasing soil disturbance and water-‐enrichment (McIntyre & Lavorel 1994). Contrastingly, exotic species richness increased with increasing soil disturbance and water-‐enrichment and did not vary significantly with grazing (McIntyre & Lavorel 1994, p. 525). In a 2002 paper, Vujnovic and his coauthors present the findings of a study conducted in grassland remnants in Alberta, Canada. The examined the total, native, and non-‐native species richness and evenness present in quadrats that had experienced various types and levels of disturbance, testing the following hypotheses (Vujnovic et al 2002, p. 505): (1) Within-‐patch species richness is at its highest at intermediate levels of disturbance – a direct test of the IDH. (2) Non-‐native vascular plant species richness increases will increasing disturbance. The results of their study are summarized in the figure below (Vujnovic et al 2002, p. 509).
As shown in the figure in the on the previous page, species evenness and Simpson’s diversity index for non-‐native species and total species did not show the relationship predicted by the IDH (Vujnovic et al 2002, p. 508-‐509). However, when disturbance occurred at intermediate levels, total
10
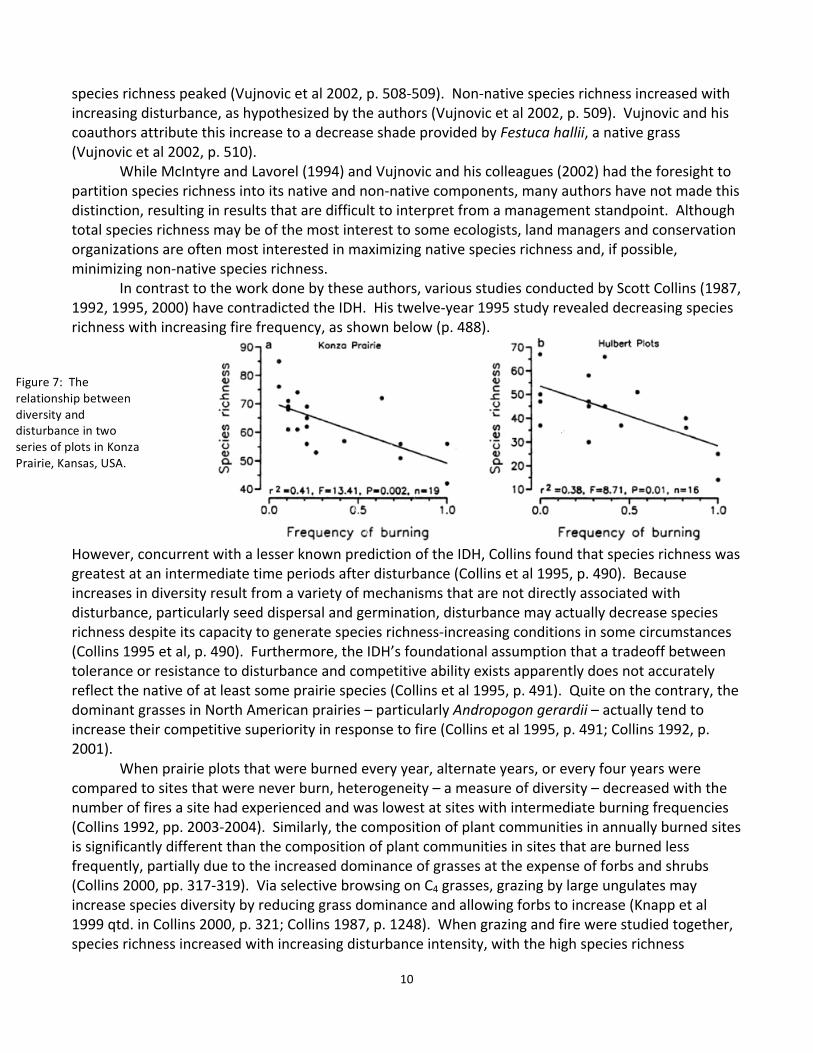
species richness peaked (Vujnovic et al 2002, p. 508-‐509). Non-‐native species richness increased with increasing disturbance, as hypothesized by the authors (Vujnovic et al 2002, p. 509). Vujnovic and his coauthors attribute this increase to a decrease shade provided by Festuca hallii, a native grass (Vujnovic et al 2002, p. 510). While McIntyre and Lavorel (1994) and Vujnovic and his colleagues (2002) had the foresight to partition species richness into its native and non-‐native components, many authors have not made this distinction, resulting in results that are difficult to interpret from a management standpoint. Although total species richness may be of the most interest to some ecologists, land managers and conservation organizations are often most interested in maximizing native species richness and, if possible, minimizing non-‐native species richness. In contrast to the work done by these authors, various studies conducted by Scott Collins (1987, 1992, 1995, 2000) have contradicted the IDH. His twelve-‐year 1995 study revealed decreasing species richness with increasing fire frequency, as shown below (p. 488).
However, concurrent with a lesser known prediction of the IDH, Collins found that species richness was greatest at an intermediate time periods after disturbance (Collins et al 1995, p. 490). Because increases in diversity result from a variety of mechanisms that are not directly associated with disturbance, particularly seed dispersal and germination, disturbance may actually decrease species richness despite its capacity to generate species richness-‐increasing conditions in some circumstances (Collins 1995 et al, p. 490). Furthermore, the IDH’s foundational assumption that a tradeoff between tolerance or resistance to disturbance and competitive ability exists apparently does not accurately reflect the native of at least some prairie species (Collins et al 1995, p. 491). Quite on the contrary, the dominant grasses in North American prairies – particularly Andropogon gerardii – actually tend to increase their competitive superiority in response to fire (Collins et al 1995, p. 491; Collins 1992, p. 2001). When prairie plots that were burned every year, alternate years, or every four years were compared to sites that were never burn, heterogeneity – a measure of diversity – decreased with the number of fires a site had experienced and was lowest at sites with intermediate burning frequencies (Collins 1992, pp. 2003-‐2004). Similarly, the composition of plant communities in annually burned sites is significantly different than the composition of plant communities in sites that are burned less frequently, partially due to the increased dominance of grasses at the expense of forbs and shrubs (Collins 2000, pp. 317-‐319). Via selective browsing on C4 grasses, grazing by large ungulates may increase species diversity by reducing grass dominance and allowing forbs to increase (Knapp et al 1999 qtd. in Collins 2000, p. 321; Collins 1987, p. 1248). When grazing and fire were studied together, species richness increased with increasing disturbance intensity, with the high species richness
Figure 7: The relationship between diversity and disturbance in two series of plots in Konza Prairie, Kansas, USA.

11
occurring in quadrats that were subjected to both fire and grazing (Collins 1987, p. 1247). Statistical analysis revealed a significant interaction between these two disturbances, resulting in increased species diversity (Collins 1987, p. 1247). Rocky Mountain grasslands may differ substantially from the Midwestern prairies studied extensively by Collins. Stohlgren and his colleagues collected data from nine areas in four states to determine whether grazing would increase native species richness via the reduction of competitive exclusion and whether the IDH holds true for this system (1999, p. 45). The authors claim (Stohlgren et al 1999, pp. 58-‐61): (1) “Grazing probably has little effect on native species richness at landscape levels in these Rocky Mountain grasslands” (emphasis mine). (2) “Grazing probably has little effect on the accelerate spread of most exotic plant species at landscape scales.” (3) “Grazing affects local plant species and life-‐form composition and cover, but spatial variation is considerable.” (4) “Soil fertility, climate, and other factors have a greater effect on plant species diversity than” grazing does. (5) “Few plant species show consistent, directional responses to grazing and cessation of grazing.” All conclusions regarding the validity of the IDH should be examined critically. Outcomes may depend upon the type of disturbance studied and the numbers and ranges of frequencies and intensities studied as well as whether or not several different disturbances are examined in concert (Grime 1973 and Martinsen 1990 qtd. in Vujnovic et al 2002, p. 505). Additionally, as noted by Mackey and Currie (2001), unless research protocols are appropriately designed, carefully sampling methodologies are used, and rigorous statistical analyses are performed, inferences that are drawn about the diversity-‐disturbance relationship and the relevance of the IDH to particular ecological communities of interest may not be valid. Implications for Management
The results of the studies reviewed in the paper specifically for their findings regarding the legitimacy of the IDH as well as other disturbance-‐related field studies conducted in grassland ecosystems have important applications for conservation. Individual and communal evolutionary histories should be considered when attempting to ascertain the appropriate management strategies for grassland ecosystems, especially when considering the use of prescribed disturbance. The types, frequencies, and intensities of disturbance that may achieve particular management objectives are likely to be site-‐specific and will often depend upon a variety of interacting factors.
Historically, grasslands have experienced relatively frequent fire (Leach & Givnish 1996). Presently, grasslands experience minimal disturbance by fire. A lack of ignition sources, the prevalence of barriers to fire (e.g. roads and urban areas), and fire suppression in grassland ecosystems composed predominantly of fire-‐adapted species, such as C4 grasses, may produce a significant disruption of ecosystem processes and function (Hobbs & Huenneke 1992; Leach & Givnish 1996). In order to increase their survivability in the presence of periodic fire, some prairie species may have developed specific physiological characteristics that actually amplify their flammability (Mutch 1970). For this reason, fire often functioned as both a disturbance and a regenerative agent in North American prairies (Gibson 1988) by removing litter and standing dead biomass (Hulbert 1988 & 1969, Wilson & Shay 1990, Knapp 1985), increasing light intensity near the soil surface and nitrogen availability early in the growing season.
When managing existing prairie remnants or newly restored prairies, the use of prescribed fire may be important to mimic the existence historical disturbance regimes (Mutch 1970, Leach &Givnish 1996). Although the benefits of prescribed fire are fairly well-‐established in the literature, consensus has not been reached regarding the ideal frequency at which prescribed burns should occur. Historical

12
data indicates that the frequency of prairie fires pre-‐settlement was once every three to ten years; however, research scientists and prairie property managers alike most commonly utilize an annual or “alternate springs” prescribed burn schedule (Gibson 1988). Similar inconsistencies regarding the appropriate seasonal timing of prescribed fire exist. Howe (1994) asserts that although many studies evaluate the effects of spring burns, based on historical fire regimes in which lightning from midsummer storms was the primary ignition source, the summer months are more appropriate for prescribed fire.
Grazing may also be an important disturbance for grassland ecosystems based on the community’s evolutionary history. The elimination of large ungulate from grassland food webs has likely affected the plant community composition by releasing the competitively superior C4 grasses from predation, reducing the community’s biodiversity (Knapp et al 1999 qtd. in Collins 2000, p. 321; Collins 1987, p. 1248). However, the work of Stohlgren and his colleagues (1999) indicates that although prairie managers may find exposing grasslands to grazing-‐induced disturbance with the goal of increasing species richness intuitively appealing, grazing probably does not exert significant effects on the plant community’s diversity at the landscape level. This finding does not preclude the use of grazing to increase species richness within small patches of prairie. Based on the results of Collins’ 1987 study of the combined effects of grazing and fire, management strategies that utilize both of these disturbances may be most successful in maximizing species richness.
In short, when attempting to manage grassland ecosystems, multiple factors must be considered: (1) Individual and communal evolutionary histories, including historical disturbances regimes, particularly fire and grazing. (2) The dispersal and colonization tendencies of the species present in the community. (3) The degree of spatial aggregation and landscape dependence exhibited by disturbances occurring within the community (Elkin & Possingham 2008). (4) Communal competitive hierarchies and whether or not a meaningful trade-‐off between competitive superiority and ability to tolerate or resist disturbance exists. (5) The specific management objectives for a given site, including, but not limited to, maximizing native species richness and/or abundance goals, minimization of non-‐native richness and/or abundance, or accurately recreating the historical prairie community. Therefore, it is imperative that those involved efforts to conserve and preserve prairie species and communities possess a deep, holistic understanding of the life histories and metapopulation dynamics of species of interest, the nature of significant disturbance regimes and their effects, and the characteristics of interspecific competition within grassland communities to best ensure that these communities are effectively preserved and restored.
References
Begon, Michael, Colin R. Townsend, and John L. Harper. Ecology: From Individuals to Ecosystems. 4th
ed. Oxford, United Kingdom: Blackwell Publishing, 2006. Benning, T. L., and Thomas B. Bragg. "Response of Big Bluestem (Andropogon gerardii Vitman) to
Timing of Spring Burning." American Midland Naturalist 130.1 (1993): 127-‐32. Connell, Joseph H. “Diversity in Tropical Rain Forests and Coral Reefs.” Science 199 (1978): 1302-‐1310. Connell, Joseph H. “Citation Classic.” ISI 46 (1987). Collins, Scott L. "Interaction of Disturbances in Tallgrass Prairie: A Field Experiment." Ecology 68.5
(1987): 1243-‐50.

13
Collins, Scott L. "Fire Frequency and Community Heterogeneity in Tallgrass Prairie Vegetation." Ecology 73.6 (1992): 2001-‐06.
Collins, Scott L., Susan M. Glenn, and David J. Gibson. "Experimental Analysis of Intermediate Disturbance and Initial Floristic Composition: Decoupling Cause and Effect." Ecology 76.2 (1995): 486-‐92.
Collins, Scott L. “Disturbance Frequency and Community Stability in Native Tallgrass Prairie.” The American Naturalist 155.3 (2000): 311-‐325.
Crombie, A.C. “Interspecific Competition.” The Journal of Animal Ecology 16.1 (1947): 44-‐73. Elkin, Che M., and Hugh Possingham. “The Role of Landscape-‐Dependent Disturbance and Dispersal in
Metapopulation Persistence.” The American Naturalist 172.4 (2008): 563-‐575. Fox, John F. “Intermediate-‐Disturbance Hypothesis.” Science 204 (1979): 1344-‐1345. Gibson, David J. "Regeneration and Fluctuation of Tallgrass Prairie Vegetation in Response to Burning
Frequency." Bulletin of the Torrey Botanical Club 115.1 (1988): 1-‐12. Hacker, Sally D., and Steven D. Gaines. “Some Implications of Direct Positive Interactions for
Community Species Diversity.” Ecology 78.7 (1997): 1990-‐2003. Hobbs, Richard J., and Laura F. Huenneke. "Disturbance, Diversity, and Invasion: Implications for
Conservation." Conservation Biology 6.3 (1992): 324-‐37. Howe, Henry F. "Managing Species Diversity in Tallgrass Prairie: Assumptions and Implications."
Conservation Biology 8.3 (1994): 691-‐704. Huston, M.A. “Patterns of Species Diversity on Coral Reefs.” Annual Review of Ecology and Systematics
16 (1985): 149-‐177. Hulbert, Lloyd C. "Fire and Litter Effects in Undisturbed Bluestem Prairie in Kansas." Ecology 50.5
(1969): 874-‐77. Hulbert, Lloyd C. "Causes of Fire Effects in Tallgrass Prairie." Ecology 69.1 (1988): 46-‐58. Knapp, Alan K. "Effect of Fire and Drought on the Ecophysiology of Andropogon gerardii and Panicum
virgatum in a Tallgrass Prairie." Ecology 66.4 (1985): 1309-‐20. Leach, Mark K., and Thomas J. Givnish. "Ecological determinants of species loss in remnant prairies."
Science 273.5281 (1996): 1555-‐59. Mackey, Robin L., and David J. Currie. “The Diversity-‐Disturbance Relationship: Is it Generally Strong
and Peaked?” Ecology 82.12 (2001): 3479-‐3492. Malcolm, Stephen. “Interspecific Competition.” Western Michigan University, Kalamazoo, MI. 3
February 2009. McIntyre, S., and Lavorel, S. “Predicting Richness of Native, Rare, and Exotic Plants in Response to
Habitat and Disturbance Variables across a Variegated Landscape.” Conservation Biology 8.2 (1994): 521-‐531.
Mutch, Robert W. "Wildland Fires and Ecosystems -‐-‐ A Hypothesis." Ecology 51.6 (1970): 1046-‐51. Neuhauser, Claudia, and Stephen W. Pacala. “An Explicitly Spatial Version of the Lotka-‐Volterra Model
with Interspecific Competition.” The Annals of Applied Probability 9.4 (1999): 1226-‐1259. Petraitis, Peter S., Roger E. Latham, and Richard A. Niesenbaum. “The Maintenance of Species Diversity
by Disturbance.” 64.4 (1989): 393-‐418. Roxburgh, Stephen H., Katriona Shea, and J. Bastow Wilson. “The Intermediate Disturbance
Hypothesis: Patch Dynamics and Mechanisms of Species Coexistence.” Ecology 85.2 (2004): 359-‐371.

14
Stohlgren, Thomas J., Lisa D. Schell, and Brian Vanden Heuvel. “How Grazing and Soil Quality Affect Native and Exotic Plant Diversity in Rocky Mountain Grasslands.” Ecological Applications 9.1 (1999): 45-‐64.
Vujnovic, K., R.W. Wein, and M.R.T. Dale. “Predicting Plant Species Diversity in Response to Disturbance Magnitude in Grassland Remnants of Central Alberta.” Canadian Journal of Botany 80.5 (2002): 504-‐511.
Wilkinson, David M. “The Disturbing History of Intermediate Disturbance.” Oikos 84.1 (1999): 145-‐147. Wooten, J. Timothy. “Effects of Disturbance on Species Diversity: A Multitrophic Perspective.” The
American Naturalist 152.6 (1998): 803-‐825.