Embed Size (px)
Citation preview

Ecology of Vertebrate Animals in Relation to Chaparral Fire in the Sierra Nevada FoothillsAuthor(s): George E. LawrenceSource: Ecology, Vol. 47, No. 2 (Mar., 1966), pp. 278-291Published by: Ecological Society of AmericaStable URL: http://www.jstor.org/stable/1933775Accessed: 01/03/2010 10:46
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=esa.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
Ecological Society of America is collaborating with JSTOR to digitize, preserve and extend access to Ecology.
http://www.jstor.org

ECOLOGY OF VERTEBRATE ANIMALS IN RELATION TO CHAPARRAL FIRE IN THE SIERRA NEVADA FOOTHILLS
GEORGE E. LAWRENCE
Museum of Vertebrate Zoology, University of California, Berkeley, Californial
Abstract. Chaparral fire brings decided changes in the species composition and density of both plant and animal populations in the Sierra Nevada foothills. Some species decrease whereas others increase following a burn, but no species is totally eliminated, nor is there any apparent diminution of total life on a burn after plant growth resumes.
These conclusions were reached in the course of a 4-year study of adjoining burned and unburned areas near Glennville, Kern County, California. Field work began in 1953 at which time study plots were selected and plant and vertebrate population were censused. A year later part of the study area was burned, and ensuing investigation compared populations on the burned and check areas for a period of 3 years, terminating in 1957.
At the time of the fire, temperatures were recorded in sites both above and below ground, and the actions of animals were observed. There was very little evidence of direct mortality among any of the vertebrates, most of them escaping the heat in one way or another. The woodrat was perhaps the most vulnerable species because of its dependence on houses made of dry twigs. However, in the bare ash after the fire many species were severely exposed to predation, and populations of most small mammals and some brush-dwelling birds decreased rapidly. Predatory birds and mammals increased, as did some seed-eating birds that found good foraging on the exposed earth.
When the rains stimulated new plant growth, a very different habitat developed in the area of burned chaparral. Most of the original trees sustained little damage, although the pitchy digger pines were largely eliminated. However, the extensive brush stands were reduced by almost 90%, and there was a corresponding increase in invading grasses and forbs.
Birds and mammals that normally exhibit a strong preference for chaparral habitat were substantially reduced in numbers in the years following the burn. Conversely, some of the birds that normally prefer grassland or oak woodland increased in number. The fire resulted in an overall increase in densities of nesting birds. None of the small mammals increased in numbers but some of the larger predators, such as the coyote and badger, moved into the burn during the months following the fire.
INTRODUCTION
Chaparral covers over 7 per cent of the total area of California; it is distributed in the arid foot- hills of the coast ranges and the interior moun- tains. These lands are becoming increasingly important in watershed protection, livestock pro- duction, and wildlife management. Because chap- arral itself gives little nourishment to domestic animals, and crowds out valuable grasses, it is often removed by range managers. The method most frequently employed is controlled brush burning. For instance, in 1950 ranchers in 30 counties cleared 97,000 acres of chaparral (Love and Jones 1952; Wieslander and Gleason 1954). Little is known, however, about the impact of chaparral fires on the total biota. The present study was undertaken to determine the effect of controlled brush burning on the native animal populations- both by direct mortality and by indirect effects occasioned by changes in the plant cover.
STUDY AREA
A series of "before" and "after" field observa- tions on an area in the vicinity of Glennville, Kern
County, California, constitutes the bulk of this report. A private land owner, Henry Bowen, initiated experiments with chaparral burning as a means of improving forage production on an area of more than 1,200 acres. The present study began in 1953 with census and study of the resident plant and animal species. On July 28, 1954, the area was cleared of the dominant shrub species by the use of controlled fire. Subsequent field observa- tions to analyze the shifts in the flora and fauna caused by the fire then followed for 3 years, through September 1957.
Elevation of the study area ranges from 2,650 ft to 3,206 ft above sea level. The area is bounded on two sides by streams, Cedar Creek on the east and Poso Creek on the west. The geographical location is 3 miles south of Glennville and 33 miles northeast of Bakersfield, California. Eco- logically the plant associations typify the Upper Sotloran life zone, and the dominant woody forms include blue oak (Quercus douglasii), digger pine (Pinus sabiniana), interior live oak (Quercus wislizenii), and buckbrush (Ceanothus cuneatus).
The terrain in this area of the Sierra Nevada foothills slopes shelflike toward the west. Major stream drainage is generally westerly in direction,
1 Present address: Bakersfield College, Bakersfield, California.
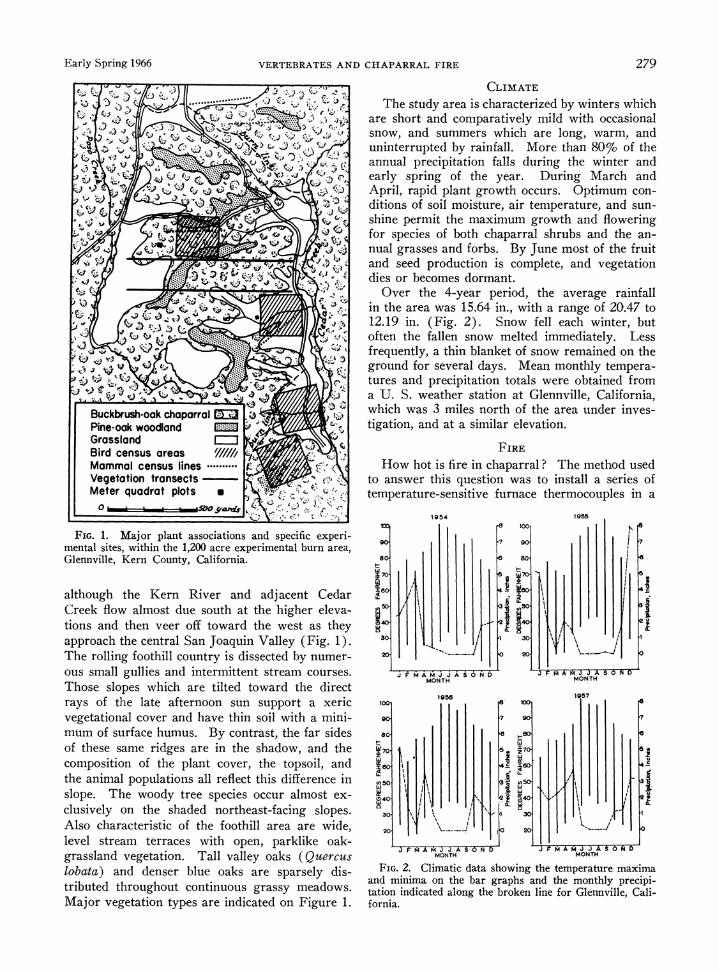
Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 279
) w
1~~~~~~~~~~~~~
k110J
Buckbrush-oak chaparral t5aZ3 ta i,
Pine-oak woodland Grassland I1> <,s\ Bird census areas 9/////fim4' Mammal census lines ........ff Sti
Vegetation transects-- ' Meter quadrant plots .
iJA (,-s U d.. ^s * .-
FIG. 1. Major plant associations and specific experi- mental sites, within the 1,200 acre experimental burn area, Glennville, Kern County, California.
although the Kern River and- adjacent Cedar Creek flow almost due south at the higher eleva- tions and then veer off toward the west as they approach the central San Joaquin Valley (Fig. 1). The rolling foothill country is dissected by numer- ous small gullies and intermittent stream courses. Those slopes which are tilted toward the direct
rays of the late afternoon sun support a xeric
vegetational cover and have thin soil with a mini- mum of surface humus. By contrast, the far sides of these same ridges are in the shadow, and the
composition of the plant cover, the topsoil, and the animal populations all reflect this difference in
slope. The woody tree species occur almost ex-
clusively on the shaded northeast-facing slopes. Also characteristic of the foothill area are wide, level stream terraces with open, parklike oak-
grassland vegetation. Tall valley oaks (Quwercus lob-ata) and denser blue oaks are sparsely dis- tributed throughout continuous grassy meadows.
Major vegetation types are indicated on Figure 1.
CLIMATE
The study area is characterized by winters which are short and comparatively mild with occasional snow, and summers which are long, warm, and uninterrupted by rainfall. More than 80% of the annual precipitation falls during the winter and early spring of the year. During March and April, rapid plant growth occurs. Optimum con- ditions of soil moisture, air temperature, and sun- shine permit the maximum growth and flowering for species of both chaparral shrubs and the an- nual grasses and forbs. By June most of the fruit and seed production is complete, and vegetation dies or becomes dormant.
Over the 4-year period, the average rainfall in the area was 15.64 in., with a range of 20.47 to 12.19 in. (Fig. 2). Snow fell each winter, but often the fallen snow melted immediately. Less frequently, a thin blanket of snow remained on the ground for several days. Mean monthly tempera- tures and precipitation totals were obtained from a U. S. weather station at Glennville, California, which was 3 miles north of the area under inves- tigation, and at a similar elevation.
FIRE
How hot is fire in chaparral? The method used to answer this question was to install a series of temperature-sensitive furnace thermocouples in a
1954 1955 100 ~ ~~~~~8 100.
90 -~~~~~7 97
8o. 80.
C~~~~~~~~~~~~~~~~L
4141~~~~3
J AM AM A S6NA MA TJJAS
FD MONTH MONTH
FIG. 2. W9 1957
and miim 9ntebrgah n temnhypeii
80 ~8 t W~~~~~~~~~II
170- 5 70
4 I j
tation indicated along the broken inr I ,i 4 LIr i I'~~~~~~~~~~ 240 1~~~~~~~~~~~~0
20 10 2
MONTH MONTH
FIG. 2. Climatic data showing the temperature maxima and minima on the bar graphs and the monthly precipi- tation indicated along the broken line for Glennville, Cali- fornia.
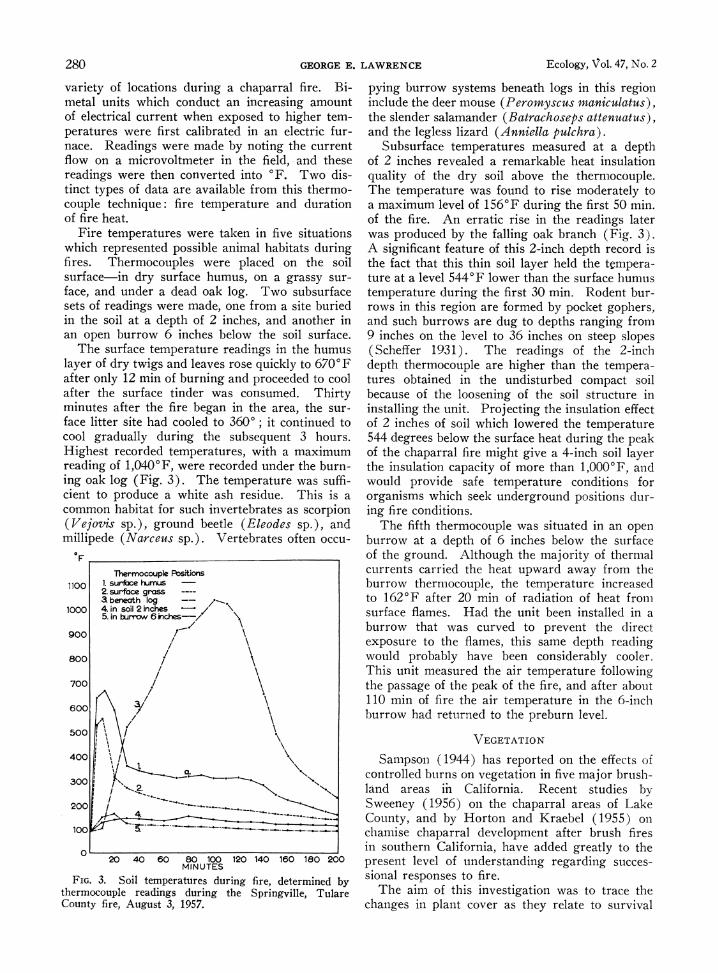
280 GEORGE E. LAWRENCE Ecology, Vol.. 47, No. 2
variety of locations during a chaparral fire. Bi- metal units which conduct an increasing amount of electrical current when exposed to higher tem- peratures were first calibrated in an electric fur- nace. Readings were made by noting the current flow on a microvoltmeter in the field, and these readings were then converted into 'F. Two dis- tinct types of data are available from this thermo- couple technique: fire temperature and duration of fire heat.
Fire temperatures were taken in five situations which represented possible animal habitats during fires. Thermocouples were placed on the soil surface-in dry surface humus, on a grassy sur- face, and under a dead oak log. Two subsurface sets of readings were made, one from a site buried in the soil at a depth of 2 inches, and another in an open burrow 6 inches below the soil surface.
The surface temperature readings in the humus layer of dry twigs and leaves rose quickly to 6700F after only 12 min of burning and proceeded to cool after the surface tinder was consumed. Thirty minutes after the fire began in the area, the sur- face litter site had cooled to 3600; it continued to cool gradually during the subsequent 3 hours. Highest recorded temperatures, with a maximum reading of 1,0400F, were recorded under the burn- ing oak log (Fig. 3). The temperature was suffi- cient to produce a white ash residue. This is a common habitat for such invertebrates as scorpion (Vejovis sp.), ground beetle (Eleodes sp.), and millipede (Narceus sp.). Vertebrates often occu-
F
Thermocouple Positions
10oo 1. suribce humus 2. surface grass a beneath log
1000 4. in soil 2 inches 5. in buryw 6 inches--//
900/
800 /
700 /
600 3/ A /
500: I
400!
300 Y
200 /-
100
0 20 40 60 80 100 120 140 160 180 200
MINUTES
FIG. 3. Soil temperatures during fire, determined by thermocouple readings during the Springville, Tulare County fire, August 3, 1957.
pying burrow systems beneath logs in this region include the deer mouse (Peromyscus maniculatus), the slender salamander (Batrachoseps attenuatus), and the legless lizard (Anniella pulchra).
Subsurface temperatures measured at a depth of 2 inches revealed a remarkable heat insulation quality of the dry soil above the thermocouple. The temperature was found to rise moderately to a maximum level of 1560F during the first 50 min. of the fire. An erratic rise in the readings later was produced by the falling oak branch (Fig. 3). A significant feature of this 2-inch depth record is the fact that this thin soil layer held the tempera- ture at a level 5440F lower than the surface humus temperature during the first 30 min. Rodent bur- rows in this region are formed by pocket gophers, and such burrows are dug to depths ranging from 9 inches on the level to 36 inches on steep slopes (Scheffer 1931). The readings of the 2-inch depth thermocouple are higher than the tempera- tures obtained in the undisturbed compact soil because of the loosening of the soil structure in installing the unit. Projecting the insulation effect of 2 inches of soil which lowered the temperature 544 degrees below the surface heat during the peak of the chaparral fire might give a 4-inch soil layer the insulation capacity of more than 1,0000F, and would provide safe temperature conditions for organisms which seek underground positions dur- ing fire conditions.
The fifth thermocouple was situated in an open burrow at a depth of 6 inches below the surface of the ground. Although the majority of thermal currents carried the heat upward away from the burrow thermocouple, the temperature increased to 1620F after 20 min of radiation of heat from surface flames. Had the unit been installed in a burrow that was curved to prevent the direct exposure to the flames, this same depth reading would probably have been considerably cooler. This unit measured the air temperature following the passage of the peak of the fire, and after about 110 min of fire the air temperature in the 6-inclh burrow had returned to the preburn level.
VEGETATION
Sampson (1944) has reported on the effects of controlled burns on vegetation in five major bruslh- land areas in California. Recent studies by Sweeney (1956) on the chaparral areas of Lake County, and by Horton and Kraebel (1955) on chamise chaparral development after brush fires in southern California, have added greatly to the present level of understanding regarding succes- sional responses to fire.
The aim of this investigation was to trace the changes in plant cover as they relate to survival

Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 281
of the resident animal population. Vegetational changes were evaluated by strip transects, m2 quadrat studies, and direct photographic records. The preburn dominant chaparral plant cover con- sisted of overaged buckbrush and gooseberry (Ribes quercetorumr) which had achieved a growth form of dense clumps with their centers composed largely of dead and dying old wood. New growth in such clumps was found at an ever-increasing distance from the original central stalks. Photo- graphs taken in June 1953 show this accumulation of dead wood in the Ceanothus chaparral, and later photo records taken from the same points serve to demonstrate the shift from chaparral toward an open oak-grassland association. The gentle slopes on the western section of the region under investigation are shown in Figure 4, where the Poso Creek drainage has been largely cleared of the dense chaparral, and grasses (Az'ena bar- bata and Brownms tectorumn) now form the domi- nant ground cover beneath the blue oaks.
Aerial photographs also were taken of the study area both before and after the fire. There is a remarkable constancy in the major vegetational
FIG. 4. Chaparral succession on the east slope of Hill 3206, showing the growth of annual and perennial grasses following the removal of the chaparral cover. The upper preburn view was taken June 12, 1953, and the postburn lower view July 2, 1956.
associations. Although the shift in chaparral can be clearly seen on the ground records, change in the cover of woody trees and larger shrubs is al- most indistinguishable from the air. The contrast in flora on opposite slope exposures is well shown in the aerial photographs.
Transect analysis of vegetation An objective measure of shift in dominant plant
species following controlled fire was obtained from the comparison of 2,000 m2 of individual plant coverage along two linear transects. The transect lines, marked out with metal markers, extended in an east-to-west direction, and included both the shaded and sunny slopes. The sampling technique consisted of walking along the line of the transect with a meter stick while recording the dominant plant species found in each m2 of the transect. The transects were recorded in June during the time when the maximum plant cover was developed, and fruiting structures were sufficiently mature to assure accurate identification of the grasses and herbaceous forms. The technique was used to compare per cent species composition during a 4-year period. Of course, the plant species as recorded in June might be reported quite differ- ently in September when such late-growing forms as vinegar weed (Trichostena lanceolatuni) or the composite (Hemizonia corymbosa) reach their peak of growth. Nonetheless, the transect sam- pling does permit an accurate year-to-year estimate of the shifts in vegetation.
One year's transect record, taken in June 1954, forms the single sample of the undisturbed chap- arral vegetation before controlled burn. Regrowth and change in plant cover was then recorded in the following 3 years of transect data. Plant species were recorded after pacing 5 m distance along the 1-m wide transect, noting all species that covered at least 1 m2.
Plant species with ability to reoccupy exposed areas are here designated as invader species. The tree species are those which normally reach heights above 15 ft, and shrubs reported are those which grow to heights of less than 12 ft. The surface area which each plant species occupied during the 4-year study is reported in numbers of m2 along the total 2,000 m linear transects.
Results Trees.-Trees were least influenced by the con-
trolled burning of the surface vegetation as indi- cated by a small decline in coverage from 278 to 251 m2 during the first postburn year; later insect damage further lowered the tree coverage to 233 in2
at the end of the period of study. The shrublike young blue oaks and the inflammable, pitchy dig-

282 GEORGE E. LAWRENCE Ecology, Vol. 47, No. 2
ger pines were the species most affected. The deciduous species-blue oak, elderberry, and buck- eye, were all found to be heat tolerant to a large degree. Their leaves were drying and the cork- forming abscission cells were active in this late- summer period of the fire. The largest trees, including valley oaks and digger pines, had grown to such size that thickness of bark, as well as elevation of lower branches above the chaparral, prevented serious scorching. The evergreen in- terior live oak revealed a remarkable response to the fire. This tree forms a thick rounded canopy which is virtually in contact with the ground sur- face; the fire failed to affect this species because of the absence of inflammable material near or under its branches and leathery leaves. The spe- cies most severely reduced in numbers during the controlled burn was the digger pine. That only about 17% of the original stand of digger pines survived the fire was indicated by the reduction of 83 m2 of transect surface to only 13 m2 of surface in the year after the burn. The pitch in the bark, in the cones, and at the base of the needle bundles was a unique physiological hazard in the burn. The cones themselves were a further hazard, as they frequently fell from the tree during the fire and carried the fire across the firebreak in the areas of steeper slopes.
Shrubs.-Chaparral was the most widespread plant association in the Glennville region before plant cover modification by man. Of the total 2,000 linear m of transect, 452 m were originally chaparral. This coverage was reduced to 41 m by controlled fire. The shrub species which in- creased following the fire was poison oak (Rhus diversiloba). The occurrence of this plant in granitic outcrops protected it from the blaze. Total coverage of two shrubs valuable as browse for the deer, mountain mahogany (Cercocarpus betu- loides) and redberry (Rhantnus crocea), was reduced only slightly, as the moist shaded slopes where they were most prevalent were least affected by burning.
Grasses.-Two different types of fire responses were noted in grass species. The perennial purple needlegrass (Stipa pulchra) was able to survive the heat beneath the soil surface. The first deli- cate shoots of green found emerging through the ash layer proved to be needlegrass leaves. The fact that the perennial rootstocks were in a viable state is further indication of the minimal depth of heat penetration. The ability of the postburn turf to resist erosion under the impact of late fall precipitation is enhanced by the presence of living root systems of such perennial plant species. The second type of grass response to the fire distur- bance was found in oats (Avena barbata) and
downy chess grass (Bromus tectorum). These species reseed more rapidly under conditions of exposed mineral soils than in the static habitat situation. The transect coverage of the wild oat nearly doubled in the 3 years of postburn re- growth, increasing from 231 to 411 m2 of the area sampled. Significant to resistance of sheet erosion in this region of the southern Sierra Ne- vada foothills is the extent of soil coverage by continuous plant cover. The chaparral species Ceanothus cuneatus and Ribes quercetorum give minimal protection to the ground surface beneath the spread of their branches. The leaf -form in this xeric habitat is too small to intercept many rain droplets. In addition, the surface of the soil beneath these shrub species is essentially bare except for a thin accumulation of litter from de- ciduous leaf fall. As a consequence of the increase in total grass turf coverage following the removal of dominant chaparral shrubs, the soil surface is somewhat better covered and more insulated from impact of rain droplets. The grass species which grows most actively in the winter and early spring, ripgut brome (Broamus rigidus), was a turf- forming soil binder in the rainy season. During the 4 years of sampling ripgut brome increased in total transect coverage from 76 to 164 m2 of surface area and was a significant part of the post- burn plant succession.
Postburn plant invader species.-The most important early invader species which reseeded naturally in denuded areas were redstem filaree (Erodium cicutarium), vinegar weed (Tricho- stems lanceolktum), popcorn flower (Plagioboth- rys nothofulvus), blue phacelia (Phacelia pur- pusii), miner's lettuce (Montia perfoliata), and golden brodiaea (Brodiaea ixoides). Denuded areas, particularly in the areas of hottest fire where white ash was deposited, were reseeded with a hardy annual grass, Lolium multiflorum. Some native species of plants more quickly occupied the patches of exposed soil than the artificially spread annual ryegrass.
The high seed-producing potential and the tol- erance of variable temperature conditions by seeds help these species reoccupy patches of exposed soil early. Seed production by miner's lettuce on northeast facing slopes not only accelerated regrowth of the general plant cover in these moist areas but also contributed to the increased food supply for flocking bird species such as Mourning Dove (Zenaidura macroura) and the Western Meadowlark (Sturnella neglecta). The earlier work of Biswell, Taber, Hedrick and Schultz (1952) reports the presence of Montia perfoliata in crops of both Mourning Doves and California Quail (Lophortyx californicus) examined from
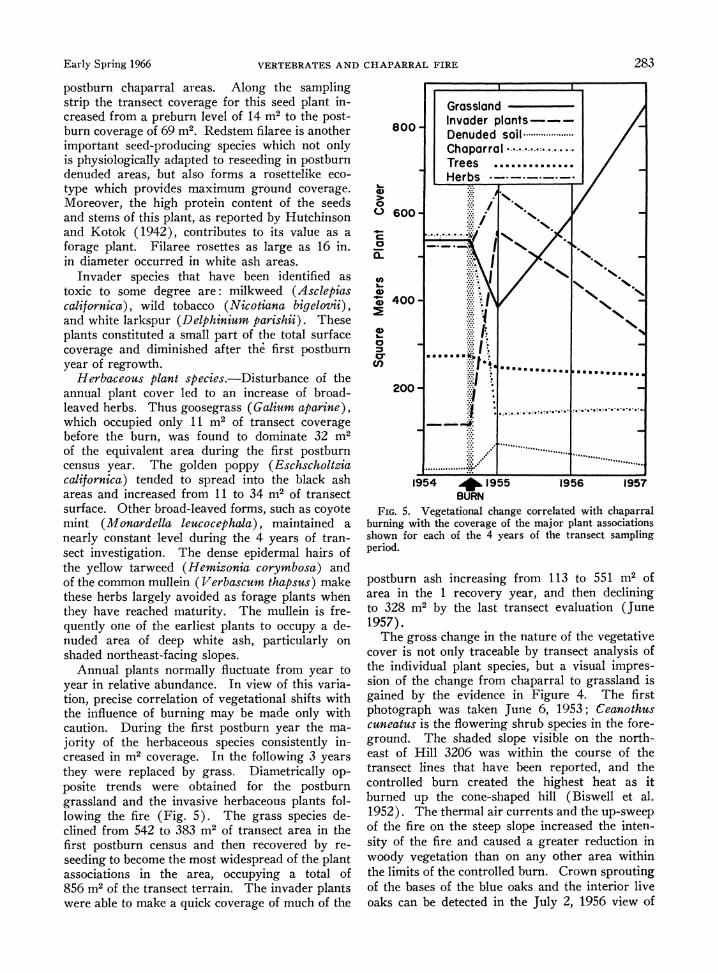
Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 283
postburn chaparral areas. Along the sampling strip the transect coverage for this seed plant in- creased from a preburn level of 14 m2 to the post- burn coverage of 69 M2. Redstem filaree is another important seed-producing species which not only is physiologically adapted to reseeding in postburn denuded areas, but also forms a rosettelike eco- type which provides maximum ground coverage. Moreover, the high protein content of the seeds and stems of this plant, as reported by Hutchinson and Kotok (1942), contributes to its value as a forage plant. Filaree rosettes as large as 16 in. in diameter occurred in white ash areas.
Invader species that have been identified as toxic to some degree are: milkweed (Asclepias californica), wild tobacco (Nicotiana bigelovii), and white larkspur (Delphinium parishii). These plants constituted a small part of the total surface coverage and diminished after the first postburn year of regrowth.
Herbaceous plant species.-Disturbance of the annual plant cover led to an increase of broad- leaved herbs. Thus goosegrass (Galium aparine), which occupied only 11 m2 of transect coverage before the burn, was found to dominate 32 m2 of the equivalent area during the first postburn census year. The golden poppy (Eschscholtzia californica) tended to spread into the black ash areas and increased from 11 to 34 m2 of transect surface. Other broad-leaved forms, such as coyote mint (Monardella leucocephala), maintained a nearly constant level during the 4 years of tran- sect investigation. The dense epidermal hairs of the yellow tarweed (Hemizonia corymbosa) and of the common mullein ( Verbascum thapsus) make these herbs largely avoided as forage plants when they have reached maturity. The mullein is fre- quently one of the earliest plants to occupy a de- nuded area of deep white ash, particularly on shaded northeast-facing slopes.
Annual plants normally fluctuate from year to year in relative abundance. In view of this varia- tion, precise correlation of vegetational shifts with the influence of burning may be made only with caution. During the first postburn year the ma- jority of the herbaceous species consistently in- creased in m2 coverage. In the following 3 years they were replaced by grass. Diametrically op- posite trends were obtained for the postburn grassland and the invasive herbaceous plants fol- lowing the fire (Fig. 5). The grass species de- clined from 542 to 383 m2 of transect area in the first postburn census and then recovered by re- seeding to become the most widespread of the plant associations in the area, occupying a total of 856 m2 of the transect terrain. The invader plants were able to make a quick coverage of much of the
I IL
Grassland Invader plants--- Denuded soil ................ Chaparral Trees .............. Herbs ___ __
0
o IN
0 6004 i;----""l@--ww NBUR
2400 .
aeinte195 1955er year, 1957e dcinn
BURN ~ ~
to 3 b Vetionla cange ctrelateith (Jun 197.
burnigrs wihanei the covera e of the maorplntasocations shownefr eaho ofl theac eable of transect samplyingo
psioun ashe icreasing from 113ara to grssan is o
areate in the flreoveryn yearubn theein thelining groud 38byThe shaded slope evisiluaton theunerh
1957.
Teas goss chang 326wswtin the nature of the vgttv covransecot linly tratchable bye trnepoted analsi the
19 954 T9h5 th9rm6 ai195etsadthp-we
ofthe iniireuon plan speepslope inrasedsual impres-
Food 5 Vegetationl chaneom chorrelted with capari
buringd wit the covieraecfe ima oigre planTh asoiastin
showen fo ecth of the 4ing yersboftec trawnse spring
gound The bsshadedh slope oaksil and the introrth-v
oast oas Hin etectasin frin the 1 ou551 mo of
tarsea intes 1harecover yeearepre, and then elnn
tor328 mupb the lanesthtansect evaluaio (Junetl 1957).Ththraaicurnsadteu-wp
The grsthneihe fronteseplpe inatreasof the veettien
cooyvergientatonl traceonablny thrasc anealysishof
sh iont of the changetromle chaprnCralwt gprasslandgi
gound.e Thseso shaed slope viksil ond the introrth-v
ofk the fie onethecsteepslp incrae the Jl2,15vinen-o
284 GEORGE E. LAWRENCE Ecology, Vol. 47, No. 2
the same area. The blue oaks, which are on more level ground, reveal a more columnar tree form after burning of the chaparral understory, since the fire tended to kill the tips of the branches which were farthest from the resistant central trunk parts of the oaks. The widely studied Tillamook burn in Oregon produced the same columnar effect on the postburn woody vegetation, and created a tree population consisting of older-age groupings (Nei- land 1958).
THE MAMMAL POPULATIONS
Live-trapping census methods The impact of the chaparral fire on the popula-
tions of the resident birds and mammals was in- vestigated. Although field records were made of the local amphibian and reptile species, no orga- nized attempt was made to census these groups. The larger mammals observed in the course of the fieldwork were not restricted to the specific study area, but moved in and out of the region in their foraging activity. Consequently, black-tailed deer (Odocoileus hemionus columbianus), coyote (Canis latrans), bobcat (Lynx rufux), badger (Taxvidea taxus), raccoon (Procyon lotor), and the California jackrabbit (Lepus californicus) are not considered here. Research in the brushland areas of northern California by Biswell et al. (1952) has carefully documented the changes in density of deer on postburn chaparral lands. Sub- stantial sampling of the fawn/doe ratio indicated increased ovulation and fawn production on those areas opened up by controlled brush fires. The present report contributes information on the re- sponse of nongame mammal populations.
Grid markers were established in 1953, and these served as permanent points of reference for the movements of mammals and wide-ranging bird species during the 4-year study. Seventy- two metal tags were attached to trees and rock outcroppings to delineate the grid. All predator activity and animal movements were noted in the field records in relation to the grid markers.
Relative density of small mammals was mea- sured by line-trapping. This method allows the comparison of relative numbers of preburn and postburn animals, but does not give an absolute measure of density. The Calhoun method of small mammal trapping has been widely adopted, and the results of many different investigations are published annually in the North American census of small mammals (Calhoun 1950). The type B line was adopted. This consists of a total of 60 live-traps set in groups of three at each site; trap- sites are spaced along the line at 50-ft intervals, and traps are set for three consecutive nights.
Each morning the traps are checked, and collected animals are weighed, measured, ear-marked, and released.
A total of four such Calhoun lines was set each 3-month period. Trapping results were reported in trap-night units. The location of the four trap lines is indicated in Figure 1. The lines were originally laid out to sample both the chap- arral area to be burned and terrain beyond the proposed fire. Sixty traps set per line for three consecutive nights totals 180 trap-nights per line; and four lines set gives 720 trap-night per 3-month period. Preburn censusing of the small mammal population was posisble for 1 year, and the post- burn census continued for 3 years. The total sampling between September 1953 and June 1957 included 11,520 trap-nights.
Habitat preference of resident mammals
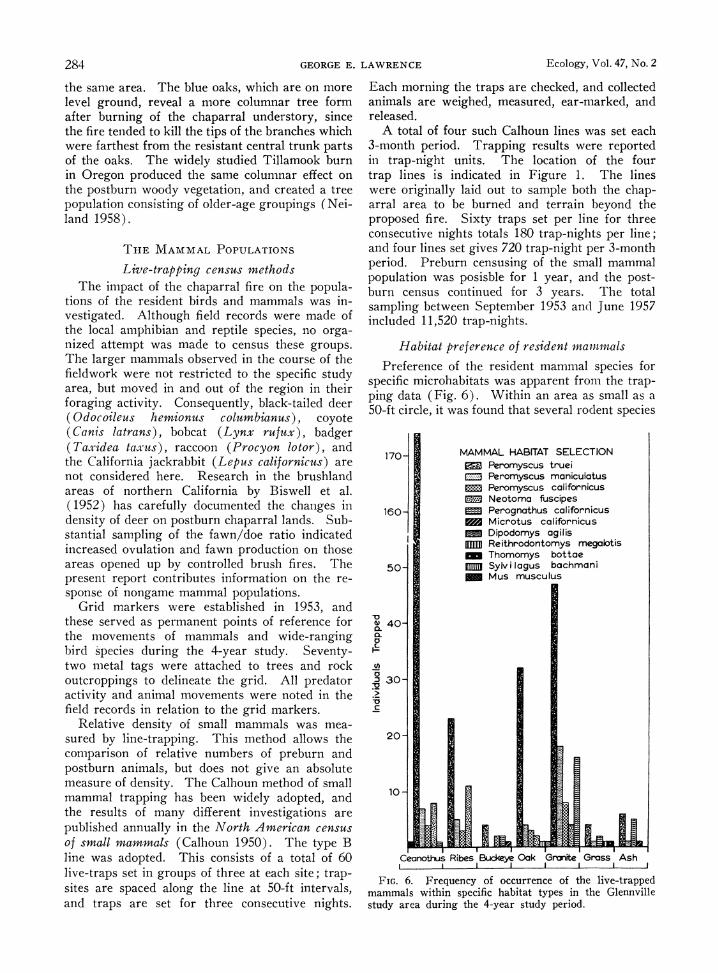
Preference of the resident mammal species for specific microhabitats was apparent from the trap- ping data (Fig. 6). Within an area as small as a 50-ft circle, it was found that several rodent species
170 MAMMAL HABITAT SELECTION - Peromyscus truei
Peromyscus maniculatus
Ei
Peromyscus californicus 1 Neotoma fuscipes 160 Perognathus californicus
Microtus californicus Dipodomys agilis
111D Reithrodontomys megalotis Thomomys bottae
50- MM Sylv i lagus bachmani Mus musculus
40-
0)
a 30-
2-o
20
Ceanothus Ribes Buckeye Oak Grnite Grass Ash
FIG. 6. Frequency of occurrence of the live-trapped mammals within specific habitat types in the Glennville study area during the 4-year study period.
Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 285
might be found living in ecologically distinct seg- ments of the habitat. The differentiation of grass- dominated, chaparral-covered, and granitic rocky terrain as mammal habitats has been noted in the central California foothills region by Fitch ( 1954) and Quast (1954).
The most frequently trapped species was the chaparral mouse (Peromyscus truei, with a total of 289 of the total 405 live-trapping records. The dominant shrub species, Ceanothus cuneatits, was the preferred habitat of the chaparral mouse; 174 of the 289 individuals were caught in live-traps set under the overhanging branches of Ceanothus shrubs (Fig. 6). At the same time, the chaparral mouse appeared to be the most eurytopic of the trapped mammals for it was found in all six of the habitat types sampled.
Other small mammals found in the wedgeleaf ceanothus chaparral were house mouse (Mus mus- culus), deer mouse (Peromyscus maniculatus), California mouse (Peromyscus californicus), and pocket mouse (Perognathus californicus).
Woodrats were most frequently trapped in the fringes of gooseberry patches, because they often built their mounded house of twigs in the center of a thorny stand of Ribes. Gooseberry thickets, which frequently encircle rocky outcrops or dead tree trunks, form an almost impregnable barrier to the approach of large predators. The advan- tage of the nest location in resisting predation is offset, however, by the vulnerability of the nest to fire.
Under the canopy of such trees as buckeye and blue oak, the mammal species most frequently trapped were the brush rabbit (Sylvilagus bach- mani) and the large California mouse. Chaparral mice, pocket mice, field mice and deer mice were all live-trapped in the ash-covered postburn region (Fig. 6). The field mouse (Microtus californi- cus) was trapped most frequently under the shade of blue oaks where the ripgut brome grass was more than 16 in. in height, providing coverage for the surface runways. However, Microtus is scarce in this xeric habitat; it prefers denser grass and continual summer growth of the grass shoots upon which it feeds (Pearson 1959).
The most unaffected mammal habitats were out- crops of fractured granite. Cracks and crannies in the rock are utilized by the deer mouse and the pocket mouse.
Several workers have successfully conducted live-trapping on burned-over forest land, showing that many individual animals survive the burning (Horn 1938; Tevis 1956; Gashwiler 1959; and Pruitt 1953). Much less is known about the ani- mals' responses to the postburn conditions of habitat in terms of the altered cover, food, and
nesting materials which must be available to sus- tain the animals. Williams (1955) carried out trapping studies on a burn in Colorado during the summer months 13 years following the fire, and reported that the food supply was substantial, and that the cover provided by the shrub and herba- ceous growth sustained even greater animal popu- lations on the burned-over lands than were present elsewhere.
Preburn and postburn mammal populations
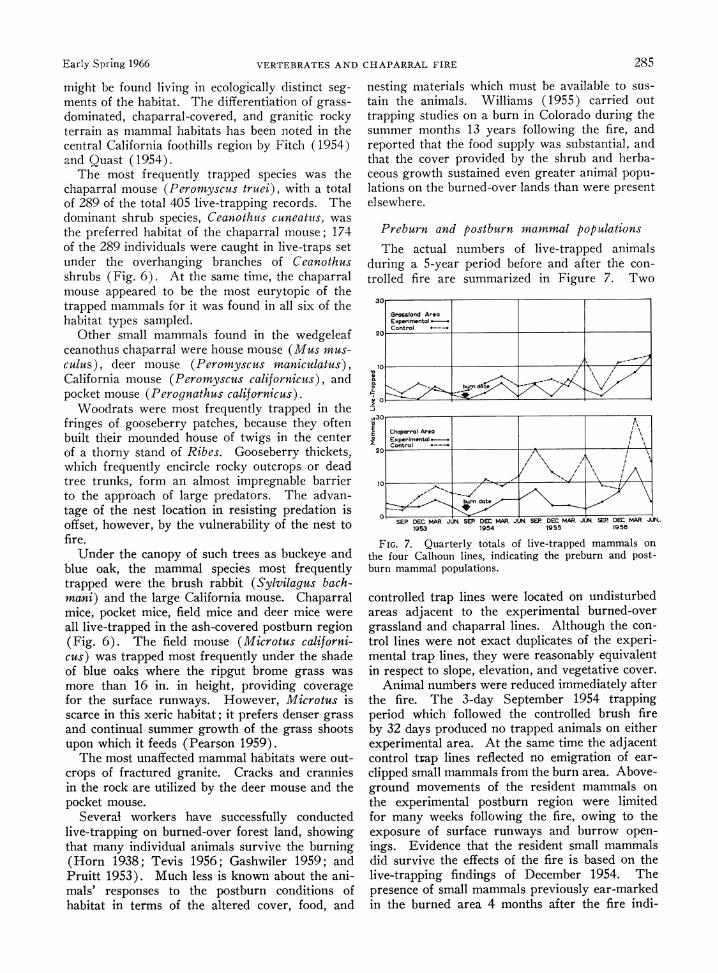
The actual numbers of live-trapped animals during a 5-year period before and after the con- trolled fire are summarized in Figure 7. Two
30.
Grassland Area Experim-ental Control
t10A burn~t V
u,30
E Chaparral Area 1 ExperimentalI
10 - \- -
SEP DEC MAR. JUN SEP DEM MAR JUN SEP DEC MAR. JLM SEP DM MAR. JUL. 1953 1954 1955 1956
FIG. 7. Quarterly totals of live-trapped mammals on the four Calhoun lines, indicating the preburn and post- burn mammal populations.
controlled trap lines were located on undisturbed areas adjacent to the experimental burned-over grassland and chaparral lines. Although the con- trol lines were not exact duplicates of the experi- mental trap lines, they were reasonably equivalent in respect to slope, elevation, and vegetative cover.
Animal numbers were reduced immediately after the fire. The 3-day September 1954 trapping period which followed the controlled brush fire by 32 days produced no trapped animals on either experimental area. At the same time the adjacent control trap lines reflected no emigration of ear- clipped small mammals from the burn area. Above- ground movements of the resident mammals on the experimental postburn region were limited for many weeks following the fire, owing to the exposure of surface runways and burrow open- ings. Evidence that the resident small mammals did survive the effects of the fire is based on the live-trapping findings of December 1954. The presence of small mammals previously ear-marked in the burned area 4 months after the fire indi-
286 GEORGE E. LAWRENCE Ecology, Vol. 47, No. 2
cates survival of a resident population. These findings are confirmed by results after a small wild- fire on the chaparral control area. Ear-marked, small mammals were recovered in the black ash area 3 days after this fire was extinguished September 26, 1956. The insulating effect of granite boulder outcroppings and blue oaks was sufficient to protect such dense shrubs as yellow gooseberry, Ribes quercetorum. Surface trapping of small mammals was most successful in such insulated habitats, following the disturbance of fire.
Movements of resident mammals
Of the 289 chaparral mice (Peromyscus truei) caught in the course of this study, 26% of the records were those of retrapped animals. Eleven per cent or 32 animals were dead when the traps were checked in the early morning. Home range activity was within a 50-ft radius, or within the interval of one Calhoun trap-line site. One ear- marked individual was taken in a trap five stations removed (250 ft) from the original trap site. Although most recatches occurred within 3 months of the time of original marking, one individual was recaptured as late as 9 months after marking.
The small mammal species which demonstrated the greatest tendency to be repeatedly trapped was the large Peromyscus californicus; 38% of the total trap occurrences were retrapped individuals. This trap hardiness was also more pronounced in the woodrat (Neotoma fuscipes) than in the smaller species of mammals caught.
Shift in weight of the Peromyscus truei population
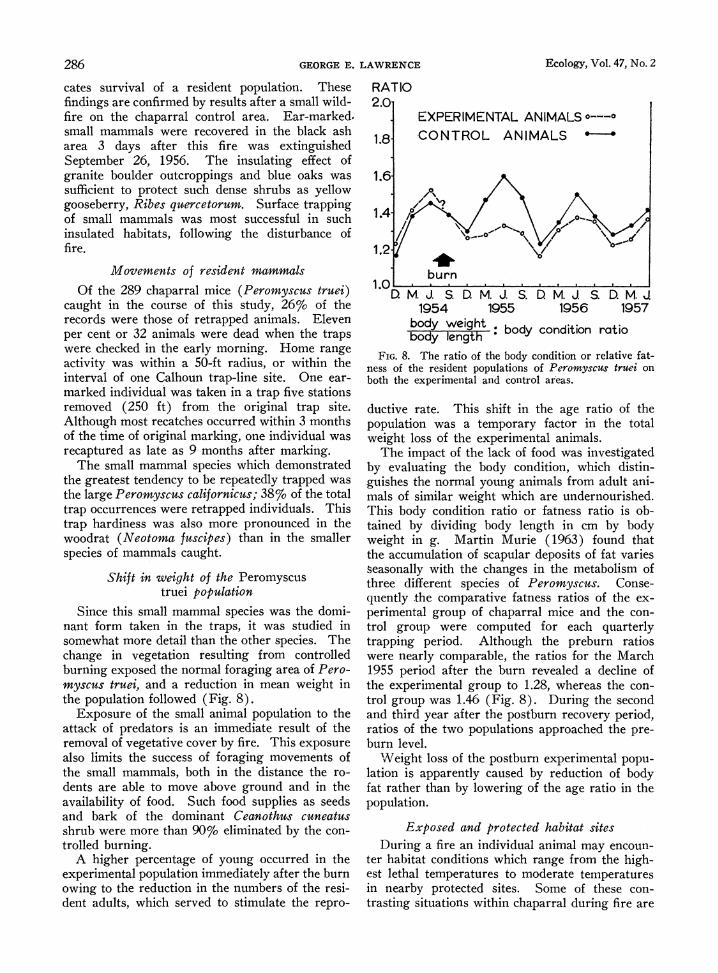
Since this small mammal species was the domi- nant form taken in the traps, it was studied in somewhat more detail than the other species. The change in vegetation resulting from controlled burning exposed the normal foraging area of Pero- myscus true, and a reduction in mean weight in the population followed (Fig. 8).
Exposure of the small animal population to the attack of predators is an immediate result of the removal of vegetative cover by fire. This exposure also limits the success of foraging movements of the small mammals, both in the distance the ro- dents are able to move above ground and in the availability of food. Such food supplies as seeds and bark of the dominant Ceanothus cuneatus shrub were more than 90% eliminated by the con- trolled burning.
A higher percentage of young occurred in the experimental population immediately after the burn owing to the reduction in the numbers of the resi- dent adults, which served to stimulate the repro-
RATIO 2.0
EXPERIMENTAL ANIMALS ?---?
1.8 CONTROL ANIMALS *
0\
1.4
1.2-
burn 1.0
D. M. J. S D. M. J. S. D M. J. S. D. M. J 1954 1955 1956 1957
body weight body condition ratio body length
FIG. 8. The ratio of the body condition or relative fat- ness of the resident populations of Peromyscus truei on both the experimental and control areas.
ductive rate. This shift in the age ratio of the population was a temporary factor in the total weight loss of the experimental animals.
The impact of the lack of food was investigated by evaluating the body condition, which distin- guishes the normal young animals from adult ani- mals of similar weight which are undernourished. This body condition ratio or fatness ratio is ob- tained by dividing body length in cm by body weight in g. Martin Murie (1963) found that the accumulation of scapular deposits of fat varies seasonally with the changes in the metabolism of three different species of Peromyscus. Conse- quently the comparative fatness ratios of the ex- perimental group of chaparral mice and the con- trol group were computed for each quarterly trapping period. Although the preburn ratios were nearly comparable, the ratios for the March 1955 period after the burn revealed a decline of the experimental group to 1.28, whereas the con- trol group was 1.46 (Fig. 8). During the second and third year after the postburn recovery period, ratios of the two populations approached the pre- burn level.
Weight loss of the postburn experimental popu- lation is apparently caused by reduction of body fat rather than by lowering of the age ratio in the population.
Exposed and protected habitat sites During a fire an individual animal may encoun-
ter habitat conditions which range from the high- est lethal temperatures to moderate temperatures in nearby protected sites. Some of these con- trasting situations within chaparral during fire are

Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 287
TABLE I. Comparison of the habitat areas which provided maximum survival conditions, with the areas of minimal survival conditions
Exposed habitat sites Protected habitat sites
1. Southwest-facing slopes with 1. Northeast-facing slopes dry vegetation, low soil moisture, which are shaded, moist, and exposure to winds and protected from prevailing
winds
2. Upper surfaces of granite outcrop- 2. Deep crevices in granite out- pings surrounded by chaparral croppings surrounded by
little inflammable material
3. Within hollow logs on the ground 3. Under logs or tree trunks or standing, with much exposed insulated by living bark wood unprotected by living bark
4. Shallow burrows less than 5 4. Burrow systems well below inches in depth and lacking cross the 5-inch depth with several ventilation surface openings
5. Lower branches of the woody trees 5. Lower branches of woody trees which are above dense dry grass which touch the ground and or dry chaparral have no inflammable material
below
6. Underground nesting areas which 6. Underground nesting sites are poorly insulated insulated by dry mineral soil
enumerated in Table I. Cleavage of the rock in this foothill region forms deep crevices which serve as refuges for the resident animals. Rodents, lizards, and even bats of the genus Pipistrella have been found to occupy such crevices. In investiga- tions on the San Joaquin Experimental Range by Howard, Fenner, and Childs (1959), caged ani- mals placed in rocky crevices and among large living roots during chaparral fire were found to survive.
Survival of mammals under high temperatures Fire does not produce a uniform heat through-
out the chaparral habitat. Entire islands of vege- tation are left unburned during the course of con- trolled burning, as shown in Table I, as well as in the Madera County report of Howard et al. (1959). Within such unburned islands the sur- vival of resident animals is determined by insula- tion from direct heat and by the availability of fresh air. The actual heat which results from burning of grass and humus reaches moderate temperatures of 5500F and less. The residue from moderate fire heat is characteristically a black ash. Dry chaparral and woody vegetation burns at temperatures above 9000 F as measured by thermo- couples (Fig. 3). Such high fire temperatures leave a deep residue of white ash, and the former dimensions of fallen logs can be detected by the nature of the ash deposit following the fire.
Temperature measurements below the ground surface indicate that animals probably experience
no ill effects of the fire heat if they occupy a bur- row system of at least 3 inches in depth. The theoretical insulation of the dry soil was investi- gated more realistically by testing live animals under simulated fire conditions. Much of the literature implies that temperature alone is re- sponsible for the death of animals under fire con- ditions. Actually, a more critical expression of the suitability of the air is vapor pressure, as emphasized by Thornthwaite (1940). The evapo- ration of water vapor from a unit area of lung surface into the air is a function of the difference in water vapor pressure of the lung surface and the atmospheric air. Consequently, subsurface temperature and relative humidity were both mea- sured in experiments involving live mice so that vapor pressure could be determined.
Since vapor pressure is proportional to air tem- perature, it is evident that fire temperatures raise the vapor pressure of water in a burrow system. To lower body temperature, mammals evaporate water vapor from the surface of the lungs and accomplish evaporative cooling and maintenance of a constant temperature in a heated environment.
An experiment carried out with chaparral mice in Contra Costa County demonstrated the neces- sity for air movement in the underground burrow system. All five experimental animals were found suffocated following a fire built on the surface of the soil above the burrow with only one opening. A subsequent experiment on June 14, 1959 was designed to allow underground air movement. A burrow was made with a soil sample borer which extended to a depth of 12 inches along a 10-foot passage. Five meadow mice were placed in non- conductive fiberboard and screen cages and located in the burrow at depths of 6, 9, and 12 inches. The relative humidity reached 82% in the burrow at the 12-inch depth, 75%o at the 9-inch depth, and nearly 60%o at the 6-inch burrow depth. At the surface of the soil the relative humidity was 48%o and the temperature was 81'F. All five animals survived the fire which warmed the soil to 1390F at a depth of 6 inches. In the deepest point in the burrow the temperature was raised only 2 degrees above the normal soil temperature.
The lethal limits of vapor pressure, at which point small mammals are no longer able to regu- late temperature and release body heat to the atmosphere, are not adequately published in the literature. At the lower limits of 22% relative humidity, the lethal temperatures range from 138 to 1450F. The vapor pressures in these conditions are from approximately 30 to 38 mm Hg. The lethal vapor pressure is reached at much higher temperatures under conditions of low relative humidity, and at lower temperatures when the
288 GEORGE E. LAWRENCE Ecology, Vol. 47, No. 2
160-
I 140/
E 1201 S
,W 100
80.
60. 0
40 \
20 We 0 N
040 60O 800 1600 12 14cy 1600 Temperature 0F
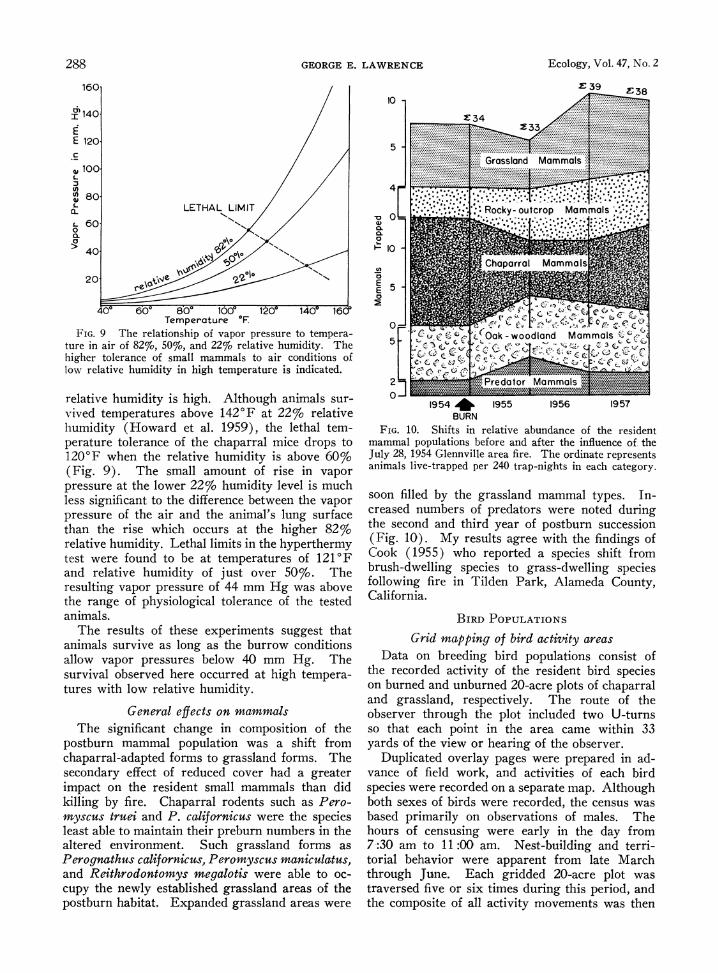
FIG. 9 The relationship of vapor pressure to tempera- ture in air of 82%, 50%, and 22% relative humidity. The higher tolerance of small mammals to air conditions of low relative humidity in high temperature is indicated.
relative humidity is high. Although animals sur- vived temperatures above 1420F at 22% relative humidity (Howard et al. 1959), the lethal tem- perature tolerance of the chaparral mice drops to 120'F when the relative humidity is above 60% (Fig. 9). The small amount of rise in vapor pressure at the lower 22% humidity level is much less significant to the difference between the vapor pressure of the air and the animal's lung surface than the rise which occurs at the higher 82% relative humidity. Lethal limits in the hyperthermy test were found to be at temperatures of 121'F and relative humidity of just over 50%. The resulting vapor pressure of 44 mm Hg was above the range of physiological tolerance of the tested animals.
The results of these experiments suggest that animals survive as long as the burrow conditions allow vapor pressures below 40 mm Hg. The survival observed here occurred at high tempera- tures with low relative humidity.
General effects on mammals The significant change in composition of the
postburn mammal population was a shift from chaparral-adapted forms to grassland forms. The secondary effect of reduced cover had a greater impact on the resident small mammals than did killing by fire. Chaparral rodents such as Pero- myscus truei and P. californicus were the species least able to maintain their preburn numbers in the altered environment. Such grassland forms as Perognathus californicus, Peromyscus maniculatus, and Reithrodontomys megalotis were able to oc- cupy the newly established grassland areas of the postburn habitat. Expanded grassland areas were
102 A.,g. X 34
233
Grassland Mammals.....:: =
4
.,.*Rocky- outcrop Mammals
I~ 0
Li) ~~~~~~~Chaparral Ma2mm~als
E5
E 5) C
L ? . {, e *;; X. loOak - woodland Mammals 5 - . c- !3<O
2 3 Predator Mammals
1954 * 1955 1956 1957 BURN
FIG. 10. Shifts in relative abundance of the resident mammal populations before and after the influence of. the July 28, 1954 Glennville area fire. The ordinate represents animals live-trapped per 240 trap-nights in each category.
soon filled by the grassland mammal types. In- creased numbers of predators were noted during the second and third year of postburn succession (Fig. 10). My results agree with the findings of Cook (1955) who reported a species shift from brush-dwelling species to grass-dwelling species following fire in Tilden Park, Alameda County, California.
BIRD POPULATIONS
Grid mapping of bird activity areas Data on breeding bird populations consist of
the recorded activity of the resident bird species on burned and unburned 20-acre plots of chaparral and grassland, respectively. The route of the observer through the plot included two U-turns so that each point in the area came within 33 yards of the view or hearing of the observer.
Duplicated overlay pages were prepared in ad- vance of field work, and activities of each bird species were recorded on a separate map. Although both sexes of birds were recorded, the census was based primarily on observations of males. The hours of censusing were early in the day from 7:30 am to 11:00 am. Nest-building and terri- torial behavior were apparent from late March through June. Each gridded 20-acre plot was traversed five or six times during this period, and the composite of all activity movements was then
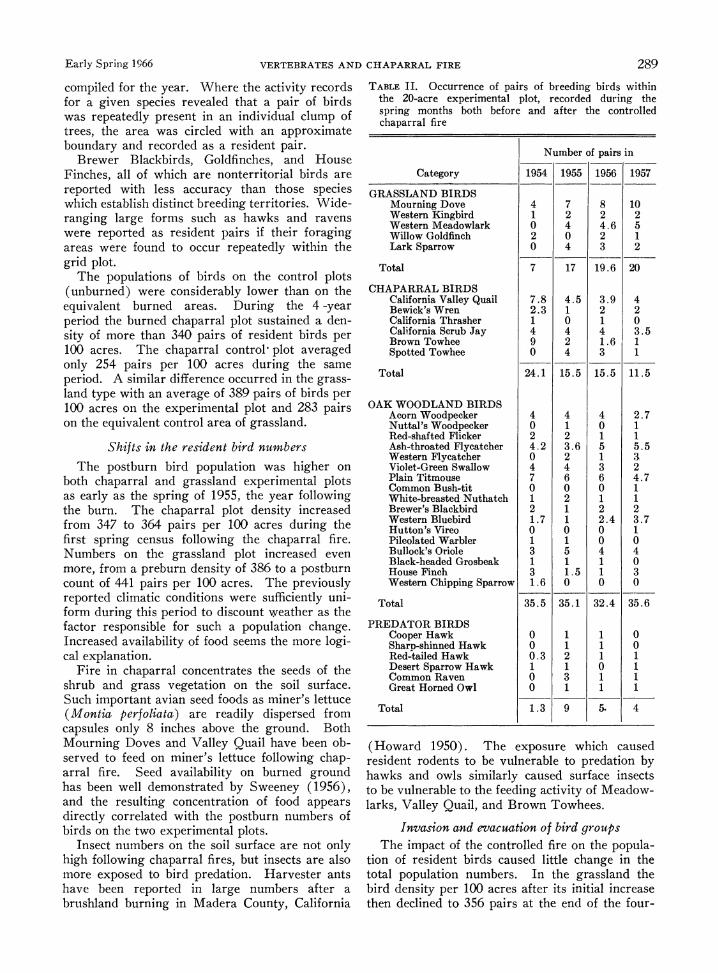
Early Spring 1966 VERTEBRATES AND CHAPARRAL FIRE 289
compiled for the year. Where the activity records for a given species revealed that a pair of birds was repeatedly present in an individual clump of trees, the area was circled with an approximate boundary and recorded as a resident pair.
Brewer Blackbirds, Goldfinches, and House Finches, all of which are nonterritorial birds are reported with less accuracy than those species which establish distinct breeding territories. Wide- ranging large forms such as hawks and ravens were reported as resident pairs if their foraging areas were found to occur repeatedly within the grid plot.
The populations of birds on the control plots (unburned) were considerably lower than on the equivalent burned areas. During the 4 -year period the burned chaparral plot sustained a den- sity of more than 340 pairs of resident birds per 100 acres. The chaparral control- plot averaged only 254 pairs per 100 acres during the same period. A similar difference occurred in the grass- land type with an average of 389 pairs of birds per 100 acres on the experimental plot and 283 pairs on the equivalent control area of grassland.
Shifts in the resident bird numbers
The postburn bird population was higher on both chaparral and grassland experimental plots as early as the spring of 1955, the year following the burn. The chaparral plot density increased from 347 to 364 pairs per 100 acres during the first spring census following the chaparral fire. Numbers on the grassland plot increased even more, from a preburn density of 386 to a postburn count of 441 pairs per 100 acres. The previously reported climatic conditions were sufficiently uni- form during this period to discount weather as the factor responsible for such a population change. Increased availability of food seems the more logi- cal explanation.
Fire in chaparral concentrates the seeds of the shrub and grass vegetation on the soil surface. Such important avian seed foods as miner's lettuce (Montia perfoliate) are readily dispersed from capsules only 8 inches above the ground. Both Mourning Doves and Valley Quail have been ob- served to feed on miner's lettuce following chap- arral fire. Seed availability on burned ground has been well demonstrated by Sweeney (1956), and the resulting concentration of food appears directly correlated with the postburn numbers of birds on the two experimental plots.
Insect numbers on the soil surface are not only high following chaparral fires, but insects are also more exposed to bird predation. Harvester ants have been reported in large numbers after a brushland burning in Madera County, California
TABLE II. Occurrence of pairs of breeding birds within the 20-acre experimental plot, recorded during the spring months both before and after the controlled chaparral fire
Number of pairs in
Category 1954 1955 1956 1957
GRASSLAND BIRDS Mourning Dove 4 7 8 10 Western Kingbird 1 2 2 2 Western Meadowlark 0 4 4.6 5 Willow Goldfinch 2 0 2 1 Lark Sparrow 0 4 3 2
Total 7 17 19.6 20
CHAPARRAL BIRDS California Valley Quail 7.8 4.5 3.9 4 Bewick's Wren 2.3 1 2 2 California Thrasher 1 0 1 0 California Scrub Jay 4 4 4 3.5 Brown Towhee 9 2 1.6 1 Spotted Towhee 0 4 3 1
Total 24.1 15.5 15.5 11.5
OAK WOODLAND BIRDS Acorn Woodpecker 4 4 4 2.7 Nuttal's Woodpecker 0 1 0 1 Red-shafted Flicker 2 2 1 1 Ash-throated Flycatcher 4.2 3.6 5 5.5 Western Flycatcher 0 2 1 3 Violet-Green Swallow 4 4 3 2 Plain Titmouse 7 6 6 4.7 Common Bush-tit 0 0 0 1 White-breasted Nuthatch 1 2 1 1 Brewer's Blackbird 2 1 2 2 Western Bluebird 1.7 1 2.4 3.7 Hutton's Vireo 0 0 0 1 Pileolated Warbler 1 1 0 0 Bullock's Oriole 3 5 4 4 Black-headed Grosbeak 1 1 1 0 House Finch 3 1.5 1 3 Western Chipping Sparrow 1.6 0 0 0
Total 35.5 35.1 32.4 35.6
PREDATOR BIRDS Cooper Hawk 0 1 1 0 Sharp-shinned Hawk 0 1 1 0 Red-tailed Hawk 0.3 2 1 1 Desert Sparrow Hawk 1 1 0 1 Common Raven 0 3 1 1 Great Horned Owl 0 1 1 1
Total 1.3 9 5- 4
(Howard 1950). The exposure which caused resident rodents to be vulnerable to predation by hawks and owls similarly caused surface insects to be vulnerable to the feeding activity of Meadow- larks, Valley Quail, and Brown Towhees.
Invasion and evacuation of bird groups The impact of the controlled fire on the popula-
tion of resident birds caused little change in the total population numbers. In the grassland the bird density per 100 acres after its initial increase then declined to 356 pairs at the end of the four-

290 GEORGE E. LAWRENCE Ecology, Vol. 47, No. 2
year period. Similarly in the chaparral, birds leveled off at 343 pairs per 100 acres. Although total numbers did not change materially, there was a shift in species composition which is shown in Table II.
Chaparral birds.-Analysis of the chaparral- adapted species from year to year shows that the original 26.6 pairs of nesting birds were largely displaced by the fire succession, and numbers dropped to 11.7 pairs per 20 acres during the post- burn spring. After 3 years of postburn census these chaparral birds were reduced to 10 pairs per 20 acres. Brown Towhees, California Thrash- ers, California Quail, California Jays, and Bewick Wrens represented the most typical chaparral adapted forms. Field observations during the sev- eral chaparral fires indicated that the shrub asso- ciation birds were not fire-killed, but rather sur- vived the flames to relocate in an appropriate adjacent habitat. The critical limiting factor in the habitat appeared to be the removal of the shrub cover which was required for these particular birds.
Grassland birds.-The original seven pairs of resident grassland bird species increased to 17 pairs in the first postburn year, then to 19.6 pairs and 20 pairs in the subsequent 2 years. The Western Meadowlark, Mourning Dove, Kingbird, Willow Goldfinch and the Lark Sparrow consti- tuted the principal species in the grassland associa- tion of birds. The influx of grassland species is more marked when the temporary feeding groups of grassland species are included. Only the resi- dent spring breeding numbers are reported in the bird population figures. The movement of the grassland-adapted bird species into the postburn area may be termed an invasive movement, but in the light of the permanent nature of the popula- tion, these birds might better be described as ex- panded resident population.
Oak-woodland birds.-The majority of the spe- cies of birds censused in this region were residents of the upper levels of the several species of oak trees. Eighteen of the reported species are in- cluded in this habitat association. The Plain Tit- mouse, Western Bluebird, California Acorn Woodpecker, Western Flycatcher, Bullock Oriole, and House Finch were the most abundant forms representative of the oak-woodland area. This population remained essentially stable during the chaparral succession over the 4-year period. In 1954, 33.5 pairs per 20 acres constituted the popu- lation. Following the fire, the oak-woodland birds were counted at 35.1 pairs in 1955, 32.4 pairs in 1956, and then to the approximate preburn level of 35.6 pairs in 1957 within the limits of the 20-acre
grid plot No. 1. Survival of the stands of oak trees is indicated by the stability of numbers of resident pairs of birds which used the oaks for foraging and nesting sites over the 4-year period.
Increased predation following fire
One of the immediate effects of the chaparral fire was to attract a large number of predators to the area. Chaparral removal increased the availa- bility of prey to the hawks and owls. Those predators which increased in number following the controlled burning of the vegetative cover in the Glennville area were the Red-tailed Hawk, Cooper Hawk, Sharp-shinned Hawk, Sparrow Hawk, Great Horned Owl, and Raven. The spring fol- lowing the fire, nine pairs of predatory birds were present in the 20-acre grid plot for foraging pur- poses. The peak population then declined to five pairs in 1956 and four pairs in 1957. Postburn predation probably was a more restrictive factor on the small animal and bird population than direct killing by the fire itself. The lack of cover pro- duces a more exposed environment during this immediate postburn period, and the surviving prey species are forced to forage in undesirable places lacking safe roosting sites, nesting sites, and escape cover. The degree of habitat recovery during the first spring following the chaparral fire was sufficient to support an accelerated reproduc- tive rate in some prey species including mice of the genus Peromyscus and such grassland birds as Meadowlarks and Mourning Doves.
ACKNOWLEDGMENTS
I should like to express my thanks to A. Starker Leo- pold for guidance of this study and for his help in the organization and revision of the manuscript; to 0. P. Pearson for his aid in the measurements of environmental factors in fire; to H. H. Biswell for his assistance in vegetation analysis; and to A. M. Schultz and R. D. Taber for their kind assistance in the field measurement of ani- mal and plant populations.
LITERATURE CITED
Biswell, H. H., R. D. Taber, D. W. Hedrick, and A. M. Schultz. 1952. Management of chamise brushlands for game in the north coast region of California. Calif. Fish and Game 38: 453-484.
Calhoun, J. B. 1951. North American census of small mammals, release No. 4. Roscoe B. Jackson Mem. Lab. Rept. 4: 1-136.
Cook, S. F., Jr. 1959. The effects of fire on a popu- lation of small rodents. Ecology 40: 102-108.
Fitch, H. S. 1954. Seasonal acceptance of bait by small animals. J. Mammal. 35: 39-49.
Gashwiler, J. S. 1959. Small mammal study in west- central Oregon. J. Mamm. 40: 128-138.
Horn, E. E. 1938. Some wildlife forest relationships. Third No. Amer. Wildlife Conf. Trans., 376-380.

Early Spring 1966 COMMUNITY EVOLUTION AND THE ORIGIN OF MAMMALS 291
Horton, J. S., and C. J. Kraebel. 1955. Development of vegetation after fire. Ecology 36: 244-260.
Howard, W. E., R. L. Fenner, and H. E. Childs, Jr. 1959. Wildlife survival on brush burns. J. Range Mgmt. 12: 230-234.
Hutchinson, C. B., and E. I. Kotok. 1942. The San Joaquin experimental range. Agric. Exp. St. Bull. 663: 3-145.
Love, R. M., and B. J. Jones. 1952. Improving Cali- fornia brush ranges. Calif. Agric. Exp. Sta. Circ. 371: 4-38.
Murie, M. 1963. Homing and orientation of deermice. J. Mamm. 44: 338-349.
Neiland, B. J. 1958. Forest and adjacent burn in the Tillamook burn area of northeastern Oregon. Ecol- ogy 39: 660-671.
Pearson, 0. P. 1959. A traffic survey of Microtus- Reithrodontomys runways. J. Mammal. 40: 169-180.
Pruitt, W. O., Jr. 1953. An analysis of some physical factors affecting the local distribution of the shorttail shrew Blarina brevicauda in the northern part of the lower peninsula of Michigan. Univ. Mich. Misc. Publ. Zool. No. 79, 39 p.
Quast, J. C. 1954. Rodent habitat preferences on foot- hill pastures in California. J. Mamm. 35: 515-521.
Sampson, A. W. 1944. Plant succession on burned chaparral lands. Univ. Calif. Agric. Exper. Sta. Bull. No. 685: 1-144.
Scheffer, T. H. 1931. Habits and economic status of pocket gopher. U.S.D.A. Tech. Bull. No. 224: 1-9.
Sweeney, J. R. 1956. Responses of vegetation to fire; a study of the herbaceous vegetation following chap- arral fires. Univ. Calif. Publ. Bot. 28: 143-250.
Tevis, L., Jr. 1956. Effect of slash burn on forest mice. J. Wildl. Mgmt. 20: 405-409.
Thornthwaite, C. W. 1940. Atmospheric moisture in relation to ecological problems. Ecology 21: 17-28.
Wieslander, E. E., and Clark H. Gleason. 1954. Major brushland areas of the coast ranges and Sierra Cas- cade foothills in California. Misc. paper No. 15. Southeast Forest and Range Exp. Sta., Berkeley, California.
Williams, 0. 1955. Distribution of mice and shrews in a Colorado montane forest. J. Mammal. 36: 221- 231.
COMMUNITY EVOLUTION AND THE ORIGIN OF MAMMALS'
EVERETT C. OLSON Department of Geophysical Sciences, University of Chicago, Chicago, Illinois
Abstract. The evolutionary course from primitive pelycosaurian reptiles through therapsids to mammals can be profitably studied in relationship to modifications of the structure of the communities in which these reptiles existed. For this purpose the community is defined in very broad terms. Three types of communities are recognized upon the basis of the nature of the food chain. Each has an important tetrapod component.
Early phases of the evolution that culminated in mammals took place in communities that were strongly tied to water by the structure of the food chain. The physiological bases of the development of mammals appear to have been related to this environmental restriction. In successive pulses, however, the pelycosaur-therapsid communities developed terrestrial reptilian herbivores and thereby broke with the water-based food chain. More strictly terrestrial com- munities developed concurrently, with the insects, which were a food source for the reptiles, as the principal herbivores. From this sort of community came the terrestrial lepidosaurian- archosaurian reptilian radiation.
The terrestrial communities so developed came into competition. In this competition the therapsid lines were temporarily unsuccessful, leaving only small, but very mammal-like, repre- sentatives in the late Triassic. After a long period with relatively little adaptive radiation, these remnants provided the basis for the radiations of mammals that led to the great successes of the Cenozoic era.
INTRODUCTORY EXPLANATIONS
Studies of vertebrate evolution centered around the concept of faunal modifications have consti- tuted one of the major fields of interest of the writer over the last decade and a half. A number of publications which have resulted from this in- terest, as cited specifically in the following text, have stressed the changes of communities with the passage of geological time. The present paper represents a continuation and extension of this kind of work. Most of the data upon which it is based have been included in the earlier studies,
but the synthesis is somewhat more general than any attempted previously, and the interpretations are more directly ecological.
Studies, such as this one, which involve broad areas of paleoecology are necessarily cast at rather different levels from those of most neoecological investigations. Naturally, as well, a strong ele- ment of speculation must enter in, for assumptions of a rather sweeping nature are necessary and conclusions often must be based on complexly interwoven threads of evidence. Yet the resulting insights into the relationships of ecology and evo- lutionary processes are such that, even though crude, the interpretations are stimulating in them-
1 The research leading to this paper was supported by NSF grants 19093 and 2543.