Embed Size (px)
Citation preview

1

2
Ecology of the Amphibians and Reptiles at Organ Pipe Cactus National Monument, Arizona
Philip C. Rosen and Charles H. Lowe
Technical Report No. 53
April 1996
National Biological Service Cooperative Park Studies Unit
School of Renewable Natural Resources 125 Biological Sciences East
The University of Arizona Tucson, Arizona 85721
National Park Service Organ Pipe Cactus National Monument
Route 1, Box 100 Ajo, Arizona 85321

3

iii
Contents
List of Figures ................................................................................................................................ iv List of Tables ................................................................................................................................. vi
Acknowledgements ..................................................................................................................... viii Abstract ......................................................................................................................................... ix Introduction .....................................................................................................................................1
General Introduction to the Project ........................................................................................1 Overview of Herpetofauna on the Monument .......................................................................2
Ecology and Conservation of Amphibians and Reptiles at Organ Pipe Cactus National Monument .........................................................................................3
Criteria for the Red List .........................................................................................................3 Toads(Anurans) ......................................................................................................................3 Turtles and Tortoises (Chelonians) ........................................................................................7 Lizards and Snakes (Squamates) .........................................................................................10
Lizard Monitoring......................................................................................................................... 45
Data Handling, Analytical Procedures, and Baseline Results ...............................................45 Site-specific Results on Intensive Study Areas .....................................................................66
Recommendations .........................................................................................................................99
Toads(Anurans) .....................................................................................................................99 Turtles and Tortoises (Chelonians).........................................................................................99 Lizards and Snakes (Squamates) .........................................................................................100
Literature Cited ...........................................................................................................................103 Appendices
Appendix 1. Summary of Species Occurrence and Frequency ..........................................106 Appendix 2. Checklist of Amphibians and Reptiles ..........................................................133 Appendix 3. Red List for Amphibians and Reptiles ..........................................................136

iv
Figures
Figure 1. Three Major Underlying Features of Desert-landscape Structure ..............................13 Figure 2. Predominating Soil Textures and Particle-size Distribution in the
Subsystems of a Desert Ecosystem Unit .....................................................................13 Figure 3. Ecological Pattern in Changes of the Ophiofauna ......................................................19 Figure 4. Histographics of Time of Snake Activity on Route 85 ...............................................33
Figure 5. Two Graphical Models of Snake Highway-mortality on Route 85 ............................41
Figure 6. Annual Means of the Peak Values for Each Lizard Line Transect .............................49
Figure 7. Regression of Species Diversity (H') on Species Richness (N) for Lizard Population Assemblages ..............................................................................................50
Figure 8. Lizard Line-transect Results by Habitat Type ............................................................53
Figure 9. Short-term Effect of Reduced Rainfall on Lizard Activity .........................................54
Figure 10. Time-constrained Search Results in Macrohabitats along the Desert Slope Gradient .......................................................................................................................55
Figure 11. Time-constrained Search Results for All Diurnal Lizard Species over the Duration of a Drought on a Lower Bajada Site ..........................................................60
Figure 12. Population Age Structure of the Indicator (Index) Lizard Species, Western Whiptail on Valley Floor Floodplain and on Bajada ..................................................61
Figure 13. Changes in Population Age-structure of Western Whiptail Lizards over the Duration of a Drought at a Lower Bajada Site ...........................................................62
Figure 14. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Aguajita Wash Site .....................................................................................71
Figure 15. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Alamo Canyon Site .......................................................................................... 73
Figure 16. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Armenta Ranch Site ....................................................................................75

v
Figure 17. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Burn Site ......................................................................................................77 Figure 18. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Creosotebush Site ........................................................................................79
Figure 19. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Dos Lomitas Site .......................................................................................81
Figure 20. Herpetology Project Results from Intensive Lizard Monitoring Study
Areas for East Armenta Site .......................................................................................83
Figure 21. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Growler Canyon Site ....................................................................................85
Figure 22. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Lizard Grid Site .........................................................................................87
Figure 23. Trend of Lizard Abundance over 6 Years at Pozo Nuevo Site ...................................90
Figure 24. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Pozo Nuevo Site ..........................................................................................91
Figure 25. Herpetology Project Results from Intensive Lizard Monitoring Study
Areas for Salsola Site ..................................................................................................93
Figure 26. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Senita Basin Site .......................................................................................95
Figure 27. Herpetology Project Results from Intensive Lizard Monitoring Study Areas for Vulture Site ..............................................................................................97

vi
Tables Table 1. Anuran Breeding Localities Identified at or Adjacent to Organ Pipe Cactus
National Monument ................................................................................................6
Table 2. Lizard and Snake Species Turnover at the Rockslope-bajada Interface ...............11
Table 3. Lizard and Snake Species Turnover Between Valley-bottom Lowermost-bajadas and Upper-middle Bajadas ....................................................14 Table 4. Habitat Segregation within Snake Communities ..................................................16 Table 5. Highway Mortality in Snakes on Route 85............................................................27
Table 6. Distances Covered During Quantitative Road-cruising for Snake Mortality on Route 85 ............................................................................................................30
Table 7. Data for Snakes Found "Live on Road" During Quantitative Road-cruising for Snake Mortality on Route 85 ..........................................................................36
Table 8. Data for Snakes Found "Dead on Road" During Quantitative
Road-cruising for Snake Mortality on Route 85 ...................................................38
Table 9. Summary and Analysis of Data Collected During Quantitative Road-cruising for Snake Mortality on Route 85....................................................40
Table 10. Peak Values and Mean Measured Abundances of Lizards on the
Standardized Lizard Line Transects, Averaged over All Transect Runs ..............48
Table 11. Habitat Effects by Comparison of Mean Measured Abundances of Lizards on the Standardized Lizard Line Transects, Averaged over All Transect Runs .................................................................................................51
Table 12. Comparison of Years for Valley Floor Floodplain Communities by Mean Measured Abundances of Lizards on the Standardized Lizard Line Transects .......................................................................................................57
Table 13. Comparison of Years for Bajada Communities by Mean Measured Abundances of Lizards on the Standardized Lizard Line Transects .....................58
Table 14. Comparison of Probability Values, Probability Test Used, and
Trend-Direction of Population Change for Lizards on the Standardized Lizard Line Transects ............................................................................................59

vii
Table 15. Preliminary Figures for Lizard Population Density and Biomass on the Lizard Grid ............................................................................................................64
Table 16. Statistics for Diurnal Lizards Observed During Time-Constrained
Searches on Intensive Monitoring Sites ................................................................67

viii
Acknowledgements
Since the inception of this project we have become indebted to many. Major field assistance was provided by James Barnett, Steve Booth, Julia Coffman, Charles Conner, Daniel Mello, Peter Holm, Michael Lee, Charles A. (Cal) Lowe, Margaret Marshall, David Parizek, and Elizabeth Wirt. Many others participated importantly in the field work, including Jonathan Arnold, Daniel Beck, Lisa Beres, William Buskirk, Victor Carasco, Mark and Ronda Daniels, Roger Eagan, Philip Fernandez, Jane Hartline, Katherine Hiett, John Iverson, R. Roy Johnson, Travis Laduc, Brent Martin, Robert McCord, Ami Pate, Trevor Persons, John Rozum, Julie Schultz, David Spiteri, Laura Thompson-Olais, Mercy Vaughn, Caroline Wilson, and Wendy Woodward. Many additional people independently provided us with verifiable species observations and specimens, including Mark Baker, James Barnett, Peter Bennett, Steve Booth, George Bradley, Charles Conner, Mark Daniels, Peter Holm, George and Christina Huey, Roy Irving, Thomas R. Jones, James Gould, Robert Johnson, Sheron Julander, Michael Kunzmann, Brent Martin, William Mikus, Alex Mintzer, Yar Petryszyn, George Ruffner, Philip Selleck, Harold Smith, Cheto and Laura Thompson-Olais, Thomas Van Devender, Peter Warren, and Elizabeth Wirt. For other assistance and input we thank Jonathan Arnold, James Barnett, Peter Bennett, Al Black, George Bradley, Raymond and Frances Camper, John Cole, Charles Conner, Gina Dupuy, Joan Ford, James Gould, George and Christina Huey, Roy Irving, Sheron Julander, Michael Kunzmann, Charles (Cal) Lowe, Will Martinez, Shawn McLaughlan, William Mikus, Lin Moore, Julie Schultz, Cecil Schwalbe, Robert Stimson, Tamara Taylor, Yolanda Wigham, Elizabeth Wirt, Wendy Woodward, and Joan Zovik. Max Hensley discussed with us his early herpetological and ecological experience at Organ Pipe Cactus National Monument (ORPI). David Parizek provided important computer assistance. For administrative assistance we thank superintendent Harold Smith and staff at ORPI, chiefs of resources management William Mikus and James Barnett, Dennis Fenn, SEP program directors Charles Stone and Eugene Wehunt, and SEP coordinator R. Roy Johnson. This work was supported by the National Park Service and The University of Arizona.

ix
Abstract We report here on 4 years of herpetological study (1987-1991) in the Sensitive Ecosystems Program (SEP; now the Ecological Monitoring Program, EMP). Our goals were to (1) conduct an extensive, monument-wide survey of the amphibians and reptiles (the herpetofauna), (2) develop baseline datasets on an intensive basis at specific study areas (EMP sites, and others) that will stand for comparison in the future, and (3) develop a system for long-range EMP monitoring of ecosystem health by the Organ Pipe Cactus National Monument (ORPI) resource management team. The project objectives were:
1. Obtain species-specific information on occurrence, distribution, and abundance of the herpetofaunal species on the monument.
2. Produce an annotated checklist of the amphibians and reptiles at ORPI, with
consideration of species of hypothetical occurrence.
3. Prepare voucher specimens required to complete documentation at ORPI of species occurrence for the monument herpetofauna.
4. Evaluate and revise the ORPI herpetofauna "Red List." This list is used to identify
special-status species, and to describe in detail threats and potential threats facing the monument herpetofauna.
5. Obtain data to describe herpetofaunal interrelationships with other ecosystem
components.
6. Design a system for monitoring herpetofaunal elements of the ORPI ecosystem.
7. Prepare documents with step-by-step instructions for long-range EMP monitoring by ORPI resource management field workers, including discussion of (a) key ecological parameters, (b) specific methodologies, (c) protocols, and (d) tools.
Over the 4-year project duration, work on the monument totalled 489 days (634 person-days) of field work. The project objectives were achieved, and substantial additional work related to conservation and ecology has been carried out for the herpetofauna of ORPI and of surrounding areas. Much of this additional work is presented herein, and some is ongoing research that will continue to assist synthesis and integration within the overall EMP. During the project, we developed and refined methodologies and analytical procedures for both research and management. Important management-oriented developments include:
1. Refinement of standardized line-transect techniques for desert lizard monitoring.

x
2. A new analytical approach for measuring and estimating road mortality in snakes on a highway transect.
Research-oriented work includes:
1. Direct population-density estimation for Sonoran Desert reptiles, including snakes.
2. Design of new live-trapping techniques for safe capture of reptiles in the desert. We covered a substantial portion of the monument wilderness area during the extensive phase of study. Monument areas deserving further survey attention have been pinpointed. Quantitative listings of species for each of the 16 EMP sites and several additional sites have been prepared. In addition, we have curated and reviewed the ORPI-housed collection of amphibians and reptiles, and have reviewed records pertaining to ORPI in the resource center museum and in the literature. Key findings presented in sections of the final report:
l. The herpetofauna at ORPI is in excellent condition throughout the overwhelming majority of the monument.
2. Only 1 species appears to be in serious jeopardy. The Sonoran mud turtle population
(Kinosternon sonoriense) at Quitobaquito appears to be continuing an earlier-documented decline, and may soon approach extirpation, as earlier projected.
3. Four additional species (2 snakes and 2 lizards) have been verified for the known
herpetofauna of ORPI, and 1 species that was previously of questionable occurrence (the Sonoran green toad [Bufo retiformis]) has also been verified. The local and ecological distributions for each of these have been documented. One species, the yellow mud turtle (Kinosternon flavescens) has been removed from the list of naturally occurring breeding species for the ORPI herpetofauna.
4. These additions to the ORPI species list represent a Sonoran Desert faunal element of the
Lower Colorado Valley. This element is strongly represented on the valley-bottom fill in the north and northwest sectors of the monument.
5. Valley-bottom fill habitats supported drought-refugium populations of several
widespread, abundant species in xeroriparian-floodplain habitats during 1989-1990, and it is the valley bottom that supports the Lower Colorado Valley faunal element. These habitats, relatively well-represented at ORPI but in few other protected areas, are a primary and integral part of the Biosphere Reserve.
6. Many of these valley floor habitats are degraded by erosion on-monument, as is well
known; and degradation in large, adjoining off-monument areas is even more severe and ongoing. Organ Pipe Cactus National Monument resource management efforts to restore

xi
this habitat at Armenta Ranch, though difficult, have been successful from the herpetological standpoint: reptile abundance is great, and population structure in rattlesnakes matches the rehabilitation history.
7. Highway mortality along State Route 85 is severe in the monument, with a minimal
estimate approaching 500 snakes killed per year; conservatively, we estimate a minimum of 39,000 snakes killed by vehicles on Route 85 since establishment of the monument in 1937. This mortality-increasing as automobile traffic increases-appears to have important negative effects on at least 2 species: the Organ Pipe shovelnosed snake (Chionactis palarostris organica) and the rosy boa (Lichanura trivirgata). These are unfortunately also of special interest, both scientifically and for many park visitors.
8. High population densities of lizards and snakes at ORPI indicate without question that
these are important taxa in the dynamics of the desert ecosystems, as predators, competitors, and prey.
Full and elaborated listings of management and conservation recommendations are given later within this document, and a summarized listing is presented as a separate section. The key recommendations are as follows:
1. Recensus the turtle population at Quitobaquito, and use telemetry to identify nesting sites and patterns. This will establish the severity of the problem there, and provide the ecological knowledge to preserve this unique desert-outpost population of an aquatic reptile.
2. Maintain the rehabilitation project at Armenta Ranch, which is a valley-bottom floodplain
community surrounded by relatively undisturbed habitat. This project has been succeeding and should be continued and expanded as a model for restoration of valley-bottom habitats.
3. Consider options for incorporation into ORPI of unprotected, federal lands north of ORPI
and within the Valley of the Ajo. We recommend an attempt to restore normal runoff to the Armenta Ranch floodplain by removal of the upstream dams on the Kuakatch Wash branch just north of the monument.
4. Establish 2 or 3 new EMP sites that will represent, and be used to monitor, the valley-
bottom fill habitats. Cuerda de Lena and Growler Valley areas should be included. Upland desertscrub (creosotebush consociation) should be included, as well as xeroriparian desertscrub (mesquite and blue paloverde habitats). A nondegraded floodplain should be included in the design.
5. Attempt to minimize the traffic on State Route 85 that is killing thousands of snakes,
rodents, anurans, birds, and large mammals. Minimize additional paving of monument roads that would promote high-speed travel and poaching of special-status snake species.

xii
6. Determine the distribution, abundance, and ecology of the 2 highway-impacted, special-status species, the Organ Pipe shovelnosed-snake and the rosy boa.

1
Introduction
General Introduction to the Project The field-oriented study design carried forward over 4 years (1987-1991) was both an extensive and intensive ecological investigation of the herpetofauna at Organ Pipe Cactus National Monument (ORPI), a United Nations Educational, Scientific, and Cultural Organization (UNESCO) Biosphere Reserve. The extensive survey phase was conducted on a monument-wide basis. The intensive phase was concentrated at 16 sites in the Ecological Monitoring Program (EMP). An important aspect of the intensive phase conducted at EMP sites was refinement of earlier, originally developed, methods for measuring lizard abundance. This provided baseline species and population information for index species and other readily observed taxa. These data are for long-range monitoring of populations and communities directed toward estimating natural-resource health on the monument. Monitoring instructions are given in a separately published document (Rosen and Lowe 1995). Within the extensive monument-wide survey phase, we made an intensive analysis of road-killed snakes on paved Route 85, the high-speed highway that transects the monument north to south. The results of that study are of particular interest to long-term monument health, conservation, and resource management. Habitat selection was investigated and conclusions are given for the diverse lizard fauna (saurofauna), snake fauna (ophiofauna), and for the frogs and toads (anurans). The current status of the Sonoran mud turtle (Kinosternon sonoriense) population at Quitobaquito and of the desert tortoise (Gopherus agassizi) monument-wide were investigated. Management and conservation recommendations are provided for several aspects of the overall work. A discussion is offered on species of the monument Red List that is relevant to the question, "to list or not to list." This discussion includes the element of habitat structure at ORPI, across the desert gradient from desert mountain to valley-bottom fill. In particular, discussion is focused on the importance of the valley bottom-normally (but not at ORPI) usurped by human development-in the structure of herpetological communities. We further carried out mark-recapture experiments at certain monument sites for lizards and snakes. Some of the preliminary results from these studies, especially related to the population density and population dynamics of reptiles, are included in various sections of this report. Voucher specimens were collected for deposit in the herpetology resource collection at ORPI and at The University of Arizona. The overwhelming majority of these were collected dead on Route 85. Additional specimens document 3 species in this study that are additions to the known herpetofauna at ORPI, and the confirmation of 2 others thought to be monument residents but not previously verified.

2
Our work employed a diversity of methodologies, some of which were original. Standard methods included (1) on-foot searching for reptiles (time-constrained search); (2) using visual and auditory cues, as well as investigating burrows and looking under surface objects such as rocks; (3) turtle trapping with baited hoop-nets; (4) sign-tracking for snakes, tortoises, and certain lizard species; (5) sound transects for anuran breeding choruses; and (6) road cruising for snakes. We directed special field trips and techniques toward survey of several species of special interest. In addition, we developed methods for (1) measuring lizard abundance using standardized, permanently established line-transects; (2) catching hatchling turtles; and (3) live-trapping desert lizards and snakes. Our investigation of the herpetofauna involved the first-ever detailed effort for a monument-wide survey of the amphibians and reptiles. The report brings together our knowledge gained during that 4-year period of intensive fieldwork (totalling 489 days on-site and 634 person-days) and earlier ORPI experience of on-site fieldwork by Philip Rosen (since 1983) and Charles Lowe (since 1951). Overview of Herpetofauna on the Monument The ORPI herpetofauna is a rich, Sonoran Desert herpetofauna that is strongly dominated by lizard and snake species. There are 16 lizard species (12 genera in 4 families), 25 snakes species (17 genera in 5 families), and 2 species of turtles (2 genera in 2 families). There are 5 amphibian species, all toads (2 genera in 2 families). This fauna includes (1) a number of true (or obligate) desert species, (2) many desert-included species with broad ecological distributions both in and outside the desert, and (3) 2 riparian species-a garter snake and a mud turtle-that are widespread in mesic Sonoran Desert riparian habitats. The preponderance of species at ORPI, as elsewhere in the Sonoran Desert, are evolutionally derived from ancestors in thornscrub and thornforest to the south, as is true for the flora (Axelrod 1979). A smaller faunal element is of more northerly grassland and woodland derivation. Many of the ORPI desert species are desert-included species that occur today as important components of thornscrub and thornforest. Those in ORPI are especially prominent in xeroriparian and rocky canyon habitats in Arizona Upland Sonoran Desertscrub, and on the higher rockslopes. A smaller herpetofaunal element of true desert species has affinities with Lower Colorado Sonoran Desertscrub entering ORPI from the west and north, and with the still more westerly Mohave Desert. These species are characteristic of the valley bottoms and of relatively dry desert rockpiles at ORPI. The ORPI herpetofauna is an exciting North American herpetofauna with several heroic species including the desert tortoise, gila monster (Heloderma suspectum), western diamondback rattlesnake (Crotalus atrox), and western coralsnake (Micruroides euryxanthus). For the warm-season visitor, the lizards and snakes are a conspicuous element of the ORPI experience, both day and night. The species are widely distributed throughout the monument, and many are abundant. As we describe in parts of this report, the lizards and snakes are so abundant that they form critical elements in the integrated dynamics of ORPI ecosystems.

3
Ecology and Conservation of Amphibians and Reptiles at
Organ Pipe Cactus National Monument Criteria for the Red List The Red List is intended to highlight species that are subject to or face unusual and serious threats at the population level. The highest form of threat in this context is the range-wide threat to the species ecology, as with the desert tortoise. A species may be placed on the Red List when specific negative impacts have been identified for it at ORPI, such as potential poaching for the rosy boa (Lichanura trivirgata) and highway mortality in the Organ Pipe shovelnosed-snake (Chionactis palarostris organica). Another appropriate criterion for red listing is low, unstable population size, as with the Sonoran mud turtle at ORPI. Based on historical data, we might also include species appearing to show long-term population declines at ORPI that may be natural fluctuations, or not, and are not currently understood. Finally, the Red List will include species peripheral in distribution at the monument, as certain snakes and lizards that occur primarily or only in relatively mesic canyons and slopes of the Ajo Mountains, and others that are restricted to valley-bottom habitats. Our conclusions for certain species with respect to this last element are based principally on considerations of the role of climate and microclimate in relationship to potential global climate change, as well as the possibility that ongoing human habitat degradation outside the monument may foreseeably threaten populations on the monument or species-wide. Toads (Anurans) Introduction Four of 5 anuran species that occur at ORPI are abundant, breed successfully, and have probably benefitted indirectly from human economic activities on monument lands. A fifth species, the Sonoran green toad (Bufo retiformis), occurs locally, primarily to the north of the monument, and may not breed on-monument. It requires further study of its local distribution and breeding success. The anuran fauna of ORPI comprises 5 species of what are commonly called toads. The "true" toads (Family Bufonidae) are the Sonoran Desert toad (formerly Colorado River toad) (Bufo alvarius), the red-spotted toad (B. punctatus), the Great Plains toad (B. cognatus), and the Sonoran green toad. There is 1 spadefoot toad (Family Pelobatidae), the desert spadefoot (Scaphiopus couchi). With 1 exception, these species occur in numbers and are known to breed successfully on the monument. The exception, the attractive Sonoran green toad, has been recorded singly as isolated individuals at Why, monument headquarters, and in the vicinity of Lukeville. By our accounting, there are only 8 records since 1960, all of which are questionable in one way or another regarding the existence of natural breeding populations on or near the monument. The type locality of this species is imprecise in the literature but appears to be close to Why, well west of the now known species population center in Arizona (Sanders and Smith 1951; Savage 1954; Williams and Chrapliwy 1958; Hulse 1978).

4
We have obtained new records for the Sonoran green toad from near the East Armenta EMP site, Gunsight Wash, Why, and north into the Valley of the Ajo to 6 mi (9.7 km) by road north of Why. We have also verified its occurrence in very small numbers near monument headquarters and near Lukeville. The new records from the north monument boundary and northward in the valley demonstrate that there are breeding populations near Why and in the federal lands north of the monument on floodplains of the valley-bottom fill. Similar habitat is rare on-monument and should be searched for along Cuerda de Lena Wash near the north monument boundary and in Kuakatch Wash in the northeast corner of ORPI. The Sonoran green toad has no legally protected refuges within its known geographic range and is thus a key, albeit peripherally distributed, member of the ORPI herpetofauna. We recommend this species for the ORPI Red List. We will need to identify its breeding sites on and near the monument. It is likely that local conservation efforts may focus on the grazed federal lands adjoining ORPI to the north. These rich, valley-bottom environments are currently being degraded by off-road vehicle use and by grazing and should be included for possible purchase in long-term resource management plans. Species Not Found on the Monument There are 3 native anuran species that occur in south central Arizona on the Tohono O'odham Reservation, but have never been recorded at ORPI. These are the southern spadefoot (Scaphiopus multiplicatus), the Great Plains narrow-mouthed toad (Gastrophryne olivacea), and the burrowing treefrog (Pternohyla fodiens). These species are supported by strong summer rains and long-lasting summer pools, and may require year-round wetter conditions than exist at ORPI. Hence, they may be absent from the monument. We heard I narrow-mouthed toad calling at Lukeville, and 1 burrowing treefrog calling briefly at Why. These, and the Sonoran green toad, should be searched for in the extreme northeastern comer of ORPI, in the Kuakatch Wash system near the village of Kuakatch, and near Dos Lomitas, especially after a series of wet years. Habitats The anurans are closely tied to permanent or temporary surface water that is long-lasting enough to allow their eggs to hatch and produce term larvae (tadpoles); as little as 10 days may be required for the desert spadefoot, with a longer period required for the true toads (genus Bufo). Each of the 4 anurans known to breed at ORPI has a substantial geographic range that extends to the north, south, east, and west of the monument area. Each anuran species breeding at ORPI has a unique pattern of habitat occurrence. Widest habitat selection is seen in the largest species, the large Sonoran Desert toad; it is the most widespread of the 4 species, occurring in mountain canyons, on the bajadas and on the valley-bottom floodplains. The desert spadefoot is also widespread and abundant on bajadas and on the valley bottom, especially in the eastern half of the monument. The red-spotted toad is abundant on rockslopes and upper bajadas throughout the eastern half of the monument and around Quitobaquito and Bates Well. The Great Plains toad occurs in modest or low abundance at localities in the eastern half of the monument and is known from Quitobaquito.

5
The distributions of anurans in the Growler Valley, Cipriano Hills, and Bates Mountains areas, and other western parts of ORPI, remain poorly known. Each of the 4 known to breed elsewhere at ORPI are expected from these areas. Breeding Sites The anurans, as a whole, are not abundant at ORPI, in comparison to the larger numbers observed to the east in areas of the Arizona Upland Sonoran Desert that receive more summer rainfall. Natural charcos that originally would have supported anurans in the valleys are rare at ORPI; and some of those that existed were augmented by ranchers (at Dos Lomitas and along Route 85 between Alamo Canyon and the monument north boundary). Anuran breeding sites that we have identified are listed in Table 1. A striking number of these sites, especially the important sites with large breeding choruses, are man-made or man-modified. The major breeding areas in both the Sonoyta Valley (Mexican agricultural fields, abandoned tank at Dos Lomitas, Lukeville, Tiger Cage, headquarters residence area, campground sewage lagoon) and the Valley of the Ajo (abandoned tanks along Route 85, North Boundary Tank, tanks at Why) are all man-made habitats. Thus, with the exception of Growler Canyon, all of the known or suspected primary anuran breeding sites in valley habitats at ORPI have human origin or modification. Some of the tanks where anurans breed at ORPI have been constructed at areas that were natural depressions on the desert floor, usually associated with major washes. Undoubtedly, some were important anuran breeding sites originally. The data in Table 1 indicate that anuran populations of the monument valleys have been enhanced, even expanded, as a result of recent human agency. Prior to these unintended improvements of anuran breeding sites, the monument species bred successfully in smaller numbers at temporary pools along major washways, as they do today at one that is easily reached at about 150 m (500 ft) east of Route 85 on Cherioni Wash. Prior to modification, the largest and most consistent breeding sites in ORPI valley habitats were undoubtedly at Growler Canyon, Aguajita Spring, and the spring system along the west face of the Quitobaquito Hills. Quitobaquito Springs was previously a major anuran center at ORPI. In 1937, Huey (1942) collected 4 Great Plains toads there in irrigated fields, the last reports for this species at Quitobaquito. During the 1950s, but not since, we observed large numbers of red-spotted and Sonoran Desert toads breeding successfully in the pond margins and environs, and the desert spadefoot also bred in numbers in the area. Successive alterations of the aquatic habitat at Quitobaquito have worsened conditions for anurans, until today there are essentially no fish-free areas at Quitobaquito where anuran eggs and larvae can survive. Small scoured pools such as those found on the unincised floodplain at Armenta Ranch are widespread at ORPI, and support small numbers of breeders. These sites dry more quickly than the deeper tanks, including the abandoned tanks, and are vulnerable to potentially destructive flash flooding. They undoubtedly supported modest numbers of anurans, with successful breeding in years with frequent and evenly distributed summer rains.

6

7
Natural breeding areas of anurans at ORPI today are also located in the mountain canyons, where tinajas and ephemeral springs exist. While our observations are for the North and South forks of Alamo Canyon, and for Bull Pasture, it is certain that anurans breed successfully in natural tinajas elsewhere in the Ajo Mountains, in Diablo Canyon, near Pinckley Peak, and in Kino Pass, and it is very likely that they breed successfully at all major tinajas. There would be considerable scientific interest in study of anuran breeding patterns in regions of the monument that are largely unaffected by human alterations to the surface water. Such studies would have ecological interest in assisting an understanding of original conditions for anurans at ORPI, as a model of anuran communities in the precaucasian Sonoran Desert. It would be of evolutionary-ecological interest to learn whether egg-laying patterns and lengths of larval stages of anurans in natural ORPI areas differ from those found in the same species (1) at adjoining areas with stock tanks, and (2) in natural Sonoran Desert areas (with similarly undisturbed surface water resources) that have greater or more consistent summer rainfall. Turtles and Tortoises (Chelonians) Desert Tortoise The desert tortoise is a polytypic species with undescribed subspecies, one of which is the Sonoran Desert tortoise that occurs at ORPI. It was first recorded at ORPI by Huey (1942) during the first vertebrate survey of the monument. Huey's field investigation was conducted at the behest of William Supernaugh, the first superintendent (custodian) of the monument. Lawrence Huey was a mammalogist undertaking a field survey focused primarily on the birds and mammals. With the principal focus on other vertebrate groups, relatively few observations were made on the amphibians and reptiles. Huey (1942) reported "a number of empty shells," and 1 live tortoise, involving 3 localities during the 3-month field survey of 1939. The relatively few observations on tortoises and other reptiles and amphibians made by Huey and his field assistants are also explained in part by their planned avoidance of the summer-season heat in southwestern Arizona, and thus the inadvertent avoidance of the dominating summer rains in the Sonoran Desert. The Sonoran Desert tortoise is primarily surface-active during the period of summer rains from June through July into October. The Sonoran Desert tortoise occupies generally rocky environments that characterize its rockpile, upper bajada, and arroyo habitats at ORPI. Unlike the Mohave Desert tortoise, it does not occur in valley-bottom and lower-bajada positions on the desertscrub gradient; it extends downward into some middle bajadas along rockier washes with hard sheltersites such as small "caliche" caves in the xeroriparian habitat. Arroyo bank sheltersites, at distance from the foot of the rockslope, are a characteristic feature of tortoise ecological distribution at ORPI that is seen throughout the range of the desert tortoise. The desert tortoise occurs on all of the major rockpiles at the monument. During 1987-1990, 49 live tortoises were located in the monument-wide survey; the tortoise fieldwork was conducted primarily by Elizabeth Wirt. An additional 11 tortoise shells and 16 shell fragment sets, plus 1 bone record and 1 scute record from individual, dead tortoises were located, totalling 29 tortoise shells and parts (remains). Based on live tortoises (49) and tortoise shells (11), percent dead is

8
18.3. Based on live tortoises (49) and all remains (29), percent dead is 37.2. In this case, the usage of all sets of dead tortoise sign, disregarding the length of time since the tortoise died and the speed of decomposition of remains, doubles the mortality estimate for the period of study monument-wide. This mortality at ORPI, as indicated by the remains located, is relatively high but not the highest (for this ratio) in Arizona populations studied during approximately the same time frame (1987-1990). The year 1987 saw a late arrival of the summer rains, and with the very hot and dry year 1989 through June 1990, the period 1987-1990 was one of relative drought throughout the North American Desert. The failure of rains culminating in the drought and heat of 1989 apparently stressed tortoise populations in many parts of the desert, including ORPI. Our assessment is that this is a natural process, not a cause for concern at ORPI, and that the effect of this drought on tortoises was more severe elsewhere. The monument supports healthy, though not remarkably large or dense, populations of the desert tortoise, including juveniles. While the desert tortoise at ORPI is not endangered by natural events, it is especially vulnerable to being collected, picked up, or otherwise mistreated by people. We recommend it as a special-status species for inclusion on the Red List; it is a true heroic species in the NPS concept, and is periodically in trouble from both natural and man-made causes. Continuing Population Decline of Sonoran Mud Turtles at Quitobaquito Springs Introduction. Data gathered in 1989 during EMP research, while not conclusive, strongly suggest that the earlier-reported population decline of the Sonoran mud turtle at Quitobaquito is continuing. If so, the situation is critical. The primary recommendations are to (1) obtain current data to verify the trend, (2) prevent public access to exposed canal banks where turtles may be collected, and (3) develop a management plan for the Sonoran mud turtle at Quitobaquito. Quitobaquito Springs is the only locality at ORPI supporting aquatic turtles. The single native species present is the Sonoran mud turtle. The distinctive Rio Sonoyta mud turtle (Kinosternon sonoriense longifemorale) (Iverson 1976, 1981) occurs at Quitobaquito and in the Rio Sonoyta south of ORPI, and nowhere else. This subspecies population is isolated in the Rio Sonoyta-Quitobaquito system, with the nearest known species populations in the Avra Valley and in the Gila River; both of these sets of populations of this species have declined and may be threatened or already extirpated. Thus, desert populations of the Sonoran mud turtle are threatened in the United States. The Sonoran mud turtle retains its strongholds in the basins and ranges below and along the Mogollon Rim and White Mountains of Arizona and New Mexico, and it remains abundant in perennial waters of the grasslands and woodlands of southeastern Arizona and northeastern Sonora. A single, mating pair of a second species, the yellow mud turtle (Kinosternon flavescens) was collected at Quitobaquito in 1955 (Smith and Hensley 1957). This species has not been reported before or since at Quitobaquito, may not have occurred there naturally, and the 1955 records may

9
represent introduction (Lowe 1987). Elsewhere, it typically occurs in seasonally dry ponds, in the central Tohono O'odham Reservation, south into Mexico, and disjunctly in southeastern Arizona east and northward in the Great Plains (pers. obs.; Iverson 1979, 1986, 1989). The Sonoran mud turtle was once abundant at Quitobaquito (Lowe 1987; Rosen 1986) but had entered a period of marked population decline beginning in the 1970s at the latest, which was documented through 1985 (Rosen 1986). We conducted additional live-trapping with mark and recapture during October and November of 1989. Our results, though meager, fit most discouragingly with the earlier-reported trend of decline. Findings. During very intensive trapping at Quitobaquito in October and again in early November of 1989, we made 63 captures of 25 adults and 20 juveniles at Quitobaquito-the most intensive short-term trapping effort to date at the site. The large numbers caught reflect only this intensive effort and seasonally high capture rates in autumn. As in 1983-1985 (Rosen 1986), we found numerous hatchlings, juveniles, and small adult males. The 1989 sample again revealed that there is little, if any, recruitment of adult females or older adults of either sex into the Quitobaquito population. The Lincoln Index estimate for the adult population is 33 individuals for 1989, compared to 68 for 1983-1985. Our fall 1989 estimate for survivors from the 68 adults that were present in the 1983-1985 cohort is only 21-far too few to indicate population stability in these turtles. Although this is a small sample, our November re-sample contained 15 recaptures among 19 total captures, suggesting that we had registered most of the individuals present in the pond in the fall of 1989. It is possible that during the fall of 1989, only a fraction of Quitobaquito turtles were active, or active in the pond. However, sampling in October and again in early November of 1982 by Peter Bennett and Michael Kunzmann yielded an estimate of 77 adults (pers. obs.). Thus, our 1989 findings suggest (but by no means conclusively) that the population decline documented in the early 1980s is continuing unabated into the 1990s. During our 1989 trapping, we were very disturbed to locate 3 dead turtles with no evident cause of mortality. One was found immediately before traps were put out; one that had been marked in the early 1980s died several days after it was trapped, measured, and released in 1989; and one died at some point during our trapping but was never trapped by us. Thus, the trapping activities do not appear to be causing this mortality, as we should expect. We had 1 similar report from 1984, which we were unable to verify, of 2 dead, adult turtles floating in Quitobaquito Pond. At present, there is nothing but conjecture for this-disease, poor water quality, or malnutrition are conceivable causes. We also received a disturbing report that at least 1 person collected a turtle from the new canal system between the spring and pond, and transported it permanently away from ORPI. We are forced to assume that our chance learning of this event signifies a high probability that it has happened several times and will continue. The shallow waters and bare banks of this new system are unacceptable for the survival of this threatened population.

10
This is a unique and isolated population of the Sonoran mud turtle, that in 1983-1985 was already in jeopardy. The only other populations of this subspecies, the Rio Sonoyta mud turtle, are or were in the Rio Sonoyta and are threatened, some possibly already extirpated. The largest known concentration of turtles in Rio Sonoyta-at and near the highway bridge in the town of Sonoyta-was reduced and may have been eliminated between 1987 and 1989. We have no information on status in Rio Sonoyta since 1983, when mud turtles were moderately abundant in the perennial reach south of Quitobaquito; 1986, when they were abundant on the eastern edge of Sonoyta; and 1990, by which time they had disappeared from the highway bridge pool in Sonoyta. Population status in Rio Sonoyta must be considered insecure at this time, and a high priority given to saving the Quitobaquito population. Lizards and Snakes (Squamates) Introduction This section of the final report discusses habitat patterns, potential poaching threats, and new distributional findings obtained under EMP that are the basis for the conclusion that the ORPI lizard and snake faunas appear to be in excellent condition. The most serious impact on the snake fauna-highway mortality on Route 85-is also analyzed and discussed in detail under a separate heading. Further, in this section we reevaluate the Red List for species of lizards and snakes (classified together in the Order Squamata) at ORPI. Vegetation and Habitats of Squamates on the Monument Patterns of habitat use for squamates at ORPI are the basis for evaluating the distribution and abundance of species in and near the monument. Warren et al. (1981) provided identification and description of 29 ORPI vegetation associations digitized to the third and fourth decimal in the classification system of Brown et al. (1979). The vegetation associations described by Warren et al. correspond to habitat-use patterns of the lizards and snakes. Each lizard and snake species will use 1 or, in almost every case, 2 or more of these vegetation associations. Although it is possible to discuss each reptile species in relation to each vegetation association, such exercises tend to obscure the more meaningful, general, habitat patterns inclusive of substratum and other edaphic characteristics.
Habitat Patterns Here, we present the basic outline and preliminary analysis of squamate habitat divisions within ORPI, based on substratum and vegetation, and with reference to the more finely discriminated vegetation subdivisions of Warren et al. (1981). The most important habitat boundary for lizard and snake species (as for plants) at ORPI is at the rockslope-bajada interface, with major species and genus turnover (Table 2). Table 2 shows that species richness for lizards and snakes is higher in the valleys than in the rocky and montane areas at ORPI. Despite the presence of several relatively specialized desert species in valley habitats, there is a clear and very strong parallel in community structure for both lizards and snakes across this interface.

11

12
The rockslope-bajada interface is illustrated in Figure 1. Deserts (arid lands) and semi-arid lands, as represented in the Basin and Range Province in which ORPI is located, are composed of 3 underlying landscape features: (1) rockpiles, (2) bajadas, and (3) valley-bottom fill. The bajada is the flanking detrital skirt that surrounds the rockpile (mountain, rocky butte, and so forth) that is its source; the familiar alluvial fan is a portion of a bajada. The 3 components in Figure 1 are contiguous subsystems of a collective ecological unit. Superimposed on these 3 primary topographic features is a pattern of drainage lines produced by water, and sandpiles that are produced primarily by wind (Fig. 2). Most of the true (or obligate), desert species of lizards and snakes at ORPI are characteristic of fine-textured soils in valley-bottom fill and dune field environments that are found within the Lower Colorado Valley Sonoran Desert (Table 3). Only 2 true desert species at ORPI, the common chuckwalla (Sauromalus obesus) and speckled rattlesnake (Crotalus mitchelli), are obligate rock-dwellers. In contrast, many desert-included desert species are obligate rock or canyon dwellers at ORPI, such as common collared lizard (Crotaphytus collaris), Clark spiny lizard (Sceloporus clarki), blacktailed rattlesnake (Crotalus molossus), lyre snake (Trimorphodon biscutatus), and others (Table 2). Across the bajada/valley bottom ecotone, species transitions are strongly characterized by intrageneric replacements (Table 3). Species characteristic of valley bottom, Lower Colorado Valley Sonoran Desert habitats, are also characteristic of the Mohave Desert. In contrast, species transitions at the rockslope/bajada interface are marked by ecological equivalency between congeners or between phyllogenetically distant genera (Table 2). Many of the rockslope and mountain canyon species are thornforest and thornscrub derived species, as are those bajada species that are prominent high on the bajada, such as longnosed snake (Rhinocheilus lecontei), Sonoran shovelnosed snake (Chionactis palarostris), and saddled leafnosed snake (Phyllorhynchus browni). Habitat Pattems within Communities Strong within-community structuring processes occur in the context of the geographically visible pattern of the lizard and snake community boundaries between rockslope, bajada, and valley-bottom environments. Lizards. Compared to lizard communities in other subdivisions of the North American Desert (Chihuahuas, Mohave, Great Basin), the lizard assemblages in the Sonoran Desert are species rich, with as many as 14 species per site. At ORPI, the Sonoran Desert lizard assemblage is fully developed, where up to 14 species may be found at a single site. On the other hand, compared to other subtropical deserts in Africa and Australia that may have 42 or more species per site (Pianka 1986), the Sonoran Desert is species poor. Lizard communities in the arid Southwest have been thought to be structured primarily on the basis of foraging microhabitat and foraging mode summarized by Vitt (1991). It has been suggested that inter-specific exploitative competition for food may be the primary structuring process within desert lizard communities (Pianka 1986). However, field experiments designed to test these ideas (Dunham 1980; Smith 1981; Tinkle 1982), widely cited as "demonstrating" competitive effects, are inconclusive: weak or variable levels of competition were detected. The

13

14
experimental evidence is not sufficient to demonstrate competition as the primary or exclusive community-organizing force. Comparative studies (Pianka 1986; Vitt 1991), however, provide strong evidence that species interactions within lizard assemblages do play an important role in community organization. Within Sonoran Desert lizard communities, macrohabitat partitioning between upland and xeroriparian desertscrub communities is not pronounced. The differences within communities are on a smaller scale, and can be categorized according to the substratum on which the lizards forage, bask, avoid predators, sleep, and carry out reproductive activities. The extensive body of work referred to above on exploitative competition has focused almost exclusively on foraging. Lizards may forage on trees or shrubs, as in the size-segregated desert spiny lizard (Sceloporus magister) and tree lizard (Urosaurus ornatus), while others forage terrestrially either on open ground or in dense cover. Some of the terrestrial foragers are active foragers, such as the western whiptail (Cnemidophorus tigris); others are wait-and-strike foragers, such as the size-segregated zebratailed lizard (Callisaurus draconoides) and sideblotched lizard (Uta stansburiana); still others are sedentary, sit-and-wait foragers, such as the regal horned lizard (Phrysonoma solare). Size differences among species foraging on the same substratum have been thought to reflect prey-size differences and, as ever, interspecific food competition. Consideration of this problem of community structure in the context of interaction of animal form with substratum allows us to integrate processes separating lizard communities at the rockslope-bajada interface with those within lizard communities. Intense predation from snakes, birds, and other vertebrates, and potential for freezing or overheating in the desert place strong restrictions on safe areas that lizard species can inhabit. Lizard home ranges-including foraging areas-must include burrows or other refugia from predators, and shelters buffered against thermal extremes. Further, aggression between lizards-both intra- and interspecific-is strong, leading to niche preemption by dominating species. Interspecific aggression in lizards includes the constant

15
threat that larger taxa (spiny lizards, whiptails) will eat smaller species (sideblotched and tree lizards). The sum of these numerous threatening features of a lizard's environment may outweigh the effects of interspecific food competition in the desert. The desert environment places a premium on lizard species that can efficiently avoid predators and survive climatic extremes on a substratum where they can also find food. We argue that lizard communities are structured according to survival success on varied substrata, with interspecific food competition being secondary, and with dietary differences being primarily a consequence of varying availabilities of food on varied survival-safe substrata. If so, the root of niche partitioning and community structure in these lizards is in the realm of efficiency and ability to operate on different substrata. An integrated understanding of survival and reproduction processes in lizard community structure remains unavailable. Here is a major research problem that has been, and continues to be, largely ignored. It is the key problem for community ecology of these animals, and a key to understanding the integrated ecosystem processes active in the desert. The long-range EMP at ORPI offers the potential to contribute to an integrated understanding of lizard community dynamics, and of the autecology of the individual lizard species. The breadth of monitoring-including our ongoing studies of snakes that are key predators on lizards provides outstanding research potential. The network of automated weather-monitoring stations at ORPI is also key to results for the desired integrated view of community dynamics at ORPI. The monitoring program at ORPI-in its final, integrated format-will be exciting in this research potential. For these reasons, we suggest that the yet-to-come EMP integration is a key aspect of the overall, long-range EMP.
Snakes. The Sonoran Desert snake assemblage is unusually rich judged against other subtropical-desert snake assemblages (Pianka 1986). Snakes are a major component of the vertebrate-eating guild at ORPI; it is likely that their impact on prey populations is substantial enough to produce intraspecific competition among snake species, as well as between snakes and larger endothermic predators such as raptors and canids. Within major community types at ORPI, snake assemblages are structured by contrasting patterns of use of xeroriparian desertscrub vs upland-desertscrub habitats (Table 4), although no species is entirely excluded from one or the other environment. This macrohabitat partitioning may be related to substratum, on and in which snakes are concealed or seek refuge from predator attack and climatic extremes. Several of the species are specialized for burrowing and excavating in soft soils, and some-but not all-of these specialize in eating excavated foods such as reptile eggs. The snake species with habitat preference for the relatively more open upland-desertscrub are typically blotched, sand-colored animals with good substratum-matching, such as the sidewinder (Crotalus cerastes), Mohave rattlesnake (Crotalus scutulatus), glossy snake (Arizona elegans), spotted leafnosed snake (Phyllorhynchus decurtatus), and night snake (Hypsiglena torquata). Many of the species with xeroriparian habitat preferences are brightly marked with red and contrasting patterns of black and white, such as the various sand snakes, western coralsnake, longnosed snake, and common kingsnake (Lampropeltis getula). These species use aposematism

16

17
("warning signs") and mimicry for protection, and quickly escape into the dense vegetation and strong shadowing of their denser thornscrub-like xeroriparian habitat. There is a greater range of specialized feeding guilds in Sonoran Desert snakes than in carnivorous lizards. There are species specialized on ants and termites, on larger arthropods, on reptile eggs, and on other snakes. Most of these species of snakes existed at low population densities during our study, and may not have been competing for food. In contrast, there are many species of medium- to large-sized snakes with generalized diets of lizards and rodents that probably were competing for food during the drought years of our work. Interspecific size-structuring within this vertebrate-eating snake assemblage allows larger species to feed primarily on rodents and the larger lizards, while certain smaller snake species feed primarily on lizards and smaller rodents. The full extent of interspecific partitioning of snake diets according to species of prey is not yet known for this community. It is clear that abundant prey species, including the Merriam kangaroo rat (Dipodomys merriami), desert pocket-mouse (Chaetodipus penicillatus), desert spiny lizard, and western whiptail, are each major dietary items for several snake species. During the drought years of the late 1980s, substratum-based macrohabitat partitioning collapsed as almost all snake species converged on the denser xeroriparian habitats, characterized strongly by imported water; and both interspecific predation and food competition were intensified as a consequence. Species with the greatest change in substratum preference-from upland to xeroriparian-showed the greatest population declines during the drought.
Conservation The large-scale habitat patterns that define lizard and snake communities at ORPI form the primary framework for evaluating conservation needs and Red List status for each species. Species primarily occurring in temperate woodland formations in the Ajo Mountains (Associations 122.4151-Juniperus monosperma-Vauquelinia californica mixed scrub, and 123.319R-Quercus ajoensis mixed scrub [Warren et al. 1981]) are likely to require the greatest attention from conservation and management standpoints. Similarly, species of the valley bottom, at ORPI in the Lower Colorado Valley Sonoran Desert (Associations 154.1115R- Larrea tridentata-Prosopis glandulosa floodplain, 154.1114-Larrea tridentata with annuals, 154.1111-Larrea tridentate-Ambrosia dumosa, 154.1112 - Larrea tridentate-Ambrosia spp. [Warren et al. 1981]), are also likely to be of special interest. The widespread generalist species within the desert, such as the sideblotched lizard, western whiptail, gopher snake (Pituophis melanoleucus), and western diamondback rattlesnake, are not foreseen to face major threats to population persistence. The structuring of snake communities within the major habitat types has further conservation importance. Mark-recapture studies at ORPI showed that snake population sizes declined during the drought of the late 1980s for most, and probably all, species. During the drought peak in 1989-1990, almost every snake species used primarily xeroriparian desertscrub associations (154.1215R-Cercidium floridum-Prosopis glandulosa-Ambrosia ambrosioides, 154.1214R- Acacia gregg-Acacia constricta-Ambrosia ambrosioides, 154.1115R Larrea tridentata-Prosopis glandulosa floodplain, and 124.711R Prosopis glandulosa riparian woodland [Warren et al. 1981]) which thus served as refugia against the drought, while the upland desertscrub

18
associations were almost void of snakes. Species with xeroriparian habitat preference declined less or more slowly than those normally using upland desertscrub. The conservation implications of this within-community structure determined by habitat preference are manifest. Xeroriparian preference by snakes in the desert, regularly or periodically, may be determined by both high prey abundance and optimal anti-predator cover in the relatively lush vegetation along washes. In addition, higher subsurface relative humidity may bring snakes to the xeroriparian habitat, although this remains to be documented. With grazing and its sequelae (particularly arroyo cutting), benefits of the xeroriparian lifestyle may be sharply restricted. During a drought, such as occurred during our study, grazing would intensify along the washes, destroying the principal drought refugium for virtually every species of snake. This suggests that grazing and arroyo-cutting might severely reduce snake populations in a synergistic process with drought. Our results in the late 1980s, after more than a decade without grazing at ORPI, show a different snake assemblage than was observed by Hensley (1950, and pers. comm.) in the early years of the monument under grazing (Fig. 3). The tendency shown in Figure 3 for xeroriparian-preference species to increase in recent decades is consistent with the hypotheses on drought effects and grazing. During the recent drought, populations of several abundant snake species at ORPI declined substantially. The declines in upland-desertscrub species (glossy snake, night snake, and spotted leafnosed snake) were so severe that the combined population density of these species dropped from > 2/ha to only a few individuals per square kilometer. For the longnosed snake, in which we achieved a 100% marked population on our primary study area, population declined by at least 67% over 2 years. In this species, habitat use first collapsed from primarily xeroriparian to exclusively so. At the drought peak, the known population of a square kilometer was reduced to 5 adult males and 3 adult females living in an 8-ha (19.8-a.) core area of optimal habitat. We have documented drought-related population declines in lizards at ORPI (Rosen and Lowe 1995). While population decline in lizards was not as precipitous as in snakes, the overall pattern of decline was similar. Populations in upland desertscrub declined most strongly, followed by bajada xeroriparian populations. We found stable population numbers only in the mesquite-dominated communities of the valley-bottom floodplains (Associations 124.711R-Prosopis glandulosa riparian woodland, 154.1115R-Larrea tridentata-Prosopis glandulosa floodplain, and 154.1763 Atriplex polycarpa-A. linearis-Prosopis glandulosa [Warren et al. 1981]). In both lizard and snake populations, failure of reproduction and recruitment was the primary evident cause of decline. Successful reproduction carried further into the drought (and population decline was least) in the most productive environments-rich xeroriparian communities. What do such drought-related population declines mean? The species studied intensively are certainly in no danger of extirpation from ORPI or even from major regions of ORPI, at least not without a severe deterioration of the Sonoran Desert climatic regime. These are widely distributed species within desert valleys that will survive through the worst naturally occurring drought with which we are familiar. Nor are they highly pursued by collectors. Their habitats are, for the most part, ignored by park visitors.

19

20
The conservation implications of strong population and community-dynamic findings apply principally to species that are uncommon and are highly restricted to the most mesic areas, and to those that are pursued by collectors. The remarkably low drought minimum population size demonstrated for the longnosed snake (one of the most abundant snake species at ORPI) is a clear signal that extraneous impacts such as collecting and highway mortality can potentially pose a magnified population threat. Poaching There is substantial commercial interest in illegally collected gila monsters and chuckwallas. Both species are available at ORPI, and the latter is conspicuous in its abundance in the rocky areas along Route 85. It is illegal to collect for sale these reptiles anywhere in Arizona, and therefore NPS poses no unique legal challenge to commercial collectors. Rather, it is the much more rigorous enforcement and intensive patrolling at ORPI that discourages this illegal activity. Poaching is more convenient elsewhere, where populations of desirable reptiles of most species still persist.
Commercial poaching could and may be carried out at the monument, particularly along the less-travelled roads, such as the north leg of Puerto Blanco Drive. Access to the rock habitat of desirable lizards and snakes is easy, and monitoring the road for NPS enforcement staff is feasible at most or all points. Apprehension of any commercial poacher at ORPI would send a very strong signal through the black market in reptiles, reinforcing avoidance of NPS lands. Small-time poaching by reptile enthusiasts is an ongoing problem that is difficult to control and of relatively minor consequence. The most vulnerable species at ORPI is the sidewinder, which uses dirt roads for travel and could be seriously impacted by collectors. It is not, however, highly pursued. The most sought-after species are primarily rock-dwelling reptiles-the rosy boa, gila monster, tiger rattlesnake (Crotalus tigris), common chuckwalla, and desert tortoise. All of these occur at low population densities, at which removal of a single or a few individuals may represent a substantial (though not irreversible) impact. The greatest hazard from poaching (and the easiest way to detect its occurrence) is habitat destruction. Poachers seeking the desirable rock-dwelling reptiles will take crowbars and other implements into the rocks to pry and shatter essential rock retreats used by reptiles and a host of other animals. The damage is lasting-the equivalent of thousands of years of weathering and erosion in each stroke. The preferred collecting technique of serious reptile poachers is "caprocking." This involves removing (by any means available) thin slabs of rock that have fractured from the surfaces of boulders and outcrops but remain in place. These "caprocks" typically match the contours and irregularities of the underlying rock. We have observed clear evidence of caprocking in Alamo Canyon and at Eagle Pass ("66 Hills"), and at scattered locations elsewhere in ORPI. Evidence of caprocking is generally clear and unequivocal. Naturally exposed rock surfaces have a darkened, weathered appearance that contrasts sharply against the bright, raw scar of the underside of a removed caprock, as well as against the freshly exposed rock surface from which it was removed. Frequently, the caprock itself will be smashed into several pieces as it is moved

21
or thrown aside. Eventually, all caprocks along major trails will be destroyed by casual or incidental collectors. However, any evidence of caprocking activity away from trails is a sign that serious poaching effort could be ongoing. It should go without saying, but is stated here for completeness, that anyone carrying the following items is probably intent on poaching reptiles: cloth sacks, snake hooks, crowbars or other prying devices, welder's gloves, and thread nooses on flexible poles. Poachers also carry out their illegal activity by driving slowly on roads at night. Their most likely target is the rosy boa. It is likely that every snaker in Arizona has considered visiting ORPI to look for this species, and it is certain that many snake-fanciers from around the world have visited, fully intent on collecting this species. The principal time for such activity is April and May, from about sunset to 2200. Those seeking rosy boas-and it should be recognized that much of this activity is carried out within legal bounds at ORPI-are likely to embark at Ajo Mountain Drive or Puerto Blanco Drive in early evening. Some of those seeking rosy boas will search the roads and rocky hills around the campground and headquarters area. A second, prized species is the Organ Pipe shovelnosed snake, found frequently on Route 85. Those collecting on this highway will ordinarily drive at < 80 kph (50 mph), and often < 50 kph (30 mph), will brake frequently, and stop or turn around occasionally. This snake is also sought primarily during April or May in the early evening, but many poachers will be inclined to arrive during the summer rainy season. An automobile that is seen 3 or more times travelling up and down Route 85 may be a sign of road collecting. Snake fanciers willing to poach a desired snake are also likely to collect gila monsters, tiger rattlesnakes, or other reptiles that they happen across. Red List Evaluations Lizards. Lizards are abundant almost everywhere at ORPI. We have presented evidence that the biomass of lizard assemblages can exceed that of other vertebrate assemblages (e.g., passerine or raptorial birds, rodents, and canids) in many areas, and perhaps monument-wide at ORPI (Rosen and Lowe 1995). Most of these lizards are primarily insectivorous and may play an important role in the community as regulators of secondary production. They are also important prey for several snake species. It is possible to observe all of the 16 lizard species at ORPI in a single, well-planned, warm, spring day. The most difficult lizard to find is the gila monster, which exists normally at low population densities and spends most of its life below ground. Also relatively difficult to observe are the desert horned lizard (Phrysonoma platyrhinos), with restricted distribution at ORPI, and the regal horned lizard. Both of these are quite abundant in places, but are cryptically colored and inconspicuous by behavior. It is evident that each of the 16 lizard species occurring at ORPI is thriving, and none appears clearly to have suffered human-caused population reductions. Highway mortality, although numerically substantial, is far too low to pose a threat to ORPI lizards at this time. Collecting for personal possession occurs, but is also of little population consequence on the monument. We

22
currently have no evidence detailing commercial collecting at ORPI, but potential for such activity exists and we identify it as the major concern for lizards. It is, of course, highly likely that populations of long-lived species such as the gila monster and common chuckwalla, as well as many of the snakes, are still recovering from the long history of heavy grazing at ORPI, just as it is likely that the overall ecosystem is still not at a natural state as the return of the natural herbivory regime settles in. This aside, we see no immediate cause for concern in any lizard species at ORPI. None of the lizard species at ORPI are threatened or seriously impacted at this time. Thus, our Red List recommendations for lizard species at ORPI are based on (1) the peripheral distribution of some on the monument and (2) the potential sensitivity of these species to climate change and to off-monument habitat degradation. Several lizard taxa at ORPI enter the monument peripherally and are largely confined to rocky or valley-bottom habitats. These less well-known species are discussed here, first in terms of habitat patterns, and then in context of the Red List. We have documented the occurrence at ORPI of 2 lizard species that were not previously verified for the monument. Both of these are characteristic of the Lower Colorado Valley Sonoran Desert, and both are restricted at ORPI to the lowermost bajada and valley-bottom fill. The longtailed brush lizard (Urosaurus graciosus) was not previously known at ORPI. We directed our work for this species, and 3 others, in the extreme northwest sector of the monument. In doing so, we located abundant populations on the lower bajada and valley-bottom fill of the Growler Valley in northwestern ORPI, and in smaller populations in the westernmost ORPI areas of the Valley of the Ajo. This small, agile shrub- and tree-dweller also occurs at low densities along the lowermost bajada edge of the Valley of the Ajo. It is characteristic of the Lower Colorado Valley Sonoran Desert, and the Mohave Desert, westward and northward. The desert horned lizard has a distribution at ORPI and through the Mohave and Sonoran deserts that is closely similar to that of the longtailed brush lizard. At ORPI, it penetrates somewhat further and in greater numbers up the lower bajada and sparingly into middle bajada of the Valley of the Ajo. Both of these new-to-ORPI species have population centers off-monument. They are protected to the north and west on Fish and Wildlife Service lands, and they are vulnerable to habitat degradation to the north in the Valley of the Ajo. We recommend these less abundant and more peripherally distributed ORPI representatives of the genera Urosaurus and Phrysonoma for the ORPI Red List. Both the longtailed brush lizard and the desert horned lizard are replaced ecologically by congeners (the tree lizard and the regal horned lizard) that are characteristic of Arizona Upland Sonoran Desert, as outlined above. The transition of these species exemplifies an important herpetofaunal ecotone at ORPI-one that exists between the valley bottom and the middle bajada. This transition is important also in certain snakes.

23
The 2 species of Urosaurus appear to shift in relative abundance at and near Cuerda de Lena Wash, while the 2 species of Phrysonoma reach a transition area near the lowermost edge of foothill paloverde (Cercidium microphyllum) on the bajada gradient, in the Valley of the Ajo. All 4 species are abundant in the Growler Valley, with a more regular pattern of species replacement along the bajada gradient than the more complicated distributional pattern in the Valley of the Ajo. We have previously discussed the herpetofaunal ecotone that exists at the junction between bajadas and rockslopes. Specific aspects of species distributional patterns in relation to this ecotone are relevant to determination of Red List status.
The desert spiny lizard abundantly occupies all valley habitats at ORPI that contain desert trees, with the exception of edificarian habitats in the monument headquarters residence area. Its congener, Clark spiny lizard, appears to occupy the habitats with desert trees where the desert spiny lizard is absent. It is possible that the balance in a competitive ecological interaction between these species is tipped by differing physiological capacities or adaptations related to water balance and/or thermoregulation. Clark spiny lizard occupies relatively dry, rocky hills at ORPI, although, as expected, it is more conspicuous in mesic canyons and in small mesic pockets in drier mountain areas. It apparently is widespread and is, in places, moderately abundant in the desert mountains of the northern Sonoran Desert, and on this basis would not merit ORPI Red List status. Another species-pair partitioning habitat at the bajada-rockslope ecotone at ORPI comprises dominant terrestrial insectivores-the western whiptail and its congener, the red-backed whiptail (Cnemidophorus burti xanthonotus). Within the Ajo Mountains there is a sharp pattern of species replacement, with red-backed whiptails occupying the rockslopes, and western whiptails occupying the bajadas. In the major canyons of the Ajo Mountains, these species overlap over a vertically narrow transitional ecotone, which is approximately 1 km (0.6 mi) long in Alamo Canyon. Here, the red-backed whiptail gradually predominates, with major pockets of the other whiptail in pockets of desert valley vegetation (creosotebush and mesquite). In ecotonal vegetation zones, however, both species are found. At ORPI, the habitat situation for these whiptail lizards is different outside the Ajo Range. In the Sonoyta Hills, Puerto Blanco and Bates mountains, and the lava-rock hills in the monument headquarters area, western whiptails occur alone or are predominant on dry slopes and low foothills, in habitats ranging from rock- to boulder- and outcrop-dominated. On larger mountains, and in more mesic locations, the red-backed whiptail is found alone; like the Clark spiny lizard, it is far less abundant in these mountains than in the Ajos, and it is relatively rare in the western monument. In the Bates Mountains in 1990-1991, after the severe drought of 1989, both the red-backed whiptail and the Clark spiny lizard were only found in the most mesic habitats on mountain slopes, at the base of large rock faces. The Ajo Mountains support, by far, the largest known population of the red-backed whiptail in the United States. This alone qualifies it for the Red List. Its sensitivity to climate-related factors

24
(especially aridity) appears to be greater than for other lizards, further indicating Red List inclusion. The 2 pairs of congeners, in the genera Sceloporus and Cnemidophorus, appear to be distributed in relation to relatively subtle, climate-related factors of moisture and temperature. Clark spiny lizard and the red-backed whiptail express the important rockslope-bajada herpetofaunal ecotone at ORPI that is ultimately climatically determined. That is, it is probable that the intrageneric replacements within these genera at ORPI are modulated primarily by water-loss processes acting as functions of both water availability and temperature. They should be focal points for study of potential impacts of climate change in the Sonoran Desert, and for this reason both species are included on the Red List. Two additional lizard species pairs show the bajada-rockslope split. These are the herbivorous lizards, common chuckwalla and desert iguana (Dipsosaurus dorsalis), and the carnivorous species pair, longnosed leopard lizard (Gambelia wislizeni) and common collared lizard; in these, macrohabitat partitioning is virtually identical to that seen in the spiny lizards. They differ from the spiny lizard pair only in that the valley taxa-longnosed leopard lizard and desert iguana-are relatively uncommon on the upper bajadas and most abundant on lower bajadas and the valley floor. Ecological separation within these 2 species pairs is strongly dependant on differences in ability or tendency to use rocks as perches and shelters. Thus, this habitat partitioning at ORPI and elsewhere is a good example of direct substratum-based ecological distribution, in contrast to the water-balance-based control described for the 2 preceding species pairs. The strongly substratum-limited species face little direct or immediate threats from poaching, highway mortality, or climate change, and hence are not included on the Red List. Snakes. As in lizards, the snake populations at ORPI appear to be healthy. Snakes occur at population densities less than an order of magnitude below those of most lizards. Therefore, small losses that are insignificant for lizard populations may be serious for snakes. This will be analyzed below in relation to the ORPI Red List status for snake species. The large-scale habitat separation among snake species within ORPI is similar to the habitat partitioning along the slope gradient for lizards. In snakes, however, the species that are characteristic of rockslopes are frequently observed on, and actively living within, upper-bajada situations. This results, at least in part, from the large home ranges of individual snakes, and the tendency to move across expanses of marginal habitat and to include varied areas in relatively extensive movements within the home range over the course of a year. Only 2 species, the blacknecked garter snake (Thamnophis cyrtopsis) and southwestern blackheaded snake (Tantilla hobartsmithi), appear to be absent from all valley habitats: both are known at ORPI only from productive canyon bottoms in the Ajo Mountains. The restricted distribution of these species, both on the monument and in the surrounding region, indicates that they require the relatively mesic canyon habitats present in the Ajos. They can be expected to be sensitive to changes in moisture regime; and the largely diurnal garter snake could be collected occasionally by visitors. We recommend that these species be included on the ORPI Red List.

25
The 6 snake species predominantly inhabiting the rockslopes are (1) tiger rattlesnake, (2) blacktailed rattlesnake, (3) speckled rattlesnake, (4) Sonoran whipsnake (Masticophis bilineatus), (5) lyre snake, and (6) rosy boa. We have recorded tiger rattlesnakes in each major area of the monument corresponding to the principal rockpiles-the Ajos, Diablos, Sonoyta Hills, Puerto Blancos, the Quitobaquito and Cipriano hills, and the Bates-Growler Mountain area. The west and northwest boundaries of ORPI are near the northwestern range limit of this rattlesnake.
The speckled rattlesnake has been confirmed at ORPI for the first time during the EMP herpetology project. It occurs at its southeastern range limit in the Growler Mountains, where it coexists with the relatively similar tiger rattlesnake. Neither of these snakes are uncommon where they are known to occur at ORPI, and the tiger rattlesnake is one of the dominant vertebrate species on many of the monument rockpiles. The highly restricted distribution of the speckled rattlesnake at ORPI unquestionably marks it for the Red List. The tiger rattlesnake, like the Clark spiny lizard, is to be included because it is likely to be affected by climate change, and additionally because collectors seek it. The blacktailed rattlesnake is known from all monument rockpiles except the Bates-Growler complex, where it probably also occurs, and it is at least moderately abundant throughout large areas of rock habitat at ORPI. In major canyons of the Ajo Mountains, and probably also on the Ajo Mountain rockslopes, it is the most abundant rattlesnake. This species ranges widely in Sonoran Desert mountains that are more mesic or are more arid than the mountains at ORPI. It is less likely to be affected by climate change than the tiger and speckled rattlesnakes, and it is not highly sought by collectors. We recommend that this species be removed from the ORPI Red List. The distribution of the lyre snake in the monument area remains poorly known. We have recorded it only on Routes 85 and 86, where it has been observed at each rockpile transected by these highways-the lava-rock hills at monument headquarters, Eagle Pass, Pozo Redondo Mountains, the Gu Oidak Hills near Sells, the Baboquivari Mountains, and the Tucson Mountains. It has also been credibly reported at the Sonoyta Hills on Puerto Blanco Drive (Philip Selleck, NPS ranger, pers. comm.). We are unable to definitively evaluate the current abundance or distribution of the lyre snake at ORPI, but the several on-monument records and its wide geographic distribution through the northern Sonoran Desert lead us to expect that it is moderately common at ORPI, as elsewhere, and occurs on rockpiles throughout. It is clear that in decades past, this snake was seen much more frequently along Route 85 in all seasons than it is today. However, lyre snakes appeared on both Routes 85 and 86 in substantial numbers in early 1990 at the drought peak (8 of 11 total records for the EMP herpetology project), suggesting that its apparent abundance may be influenced markedly by movement patterns that vary from year to year. The several uncertainties concerning the distribution and abundance (as well as the possible decline in abundance) of the lyre snake at ORPI are not enough to warrant Red List inclusion. The Sonoran whipsnake is represented at ORPI by the subspecies, Ajo Mountain whipsnake (Masticophis bilineatus lineolatus). This form has recently been reported from near Ajo (Boundy and Ford 1989), and we have found it on Route 86 in the Quijotoa Hills. It is by no means

26
restricted to the Ajo Mountains: we have recorded it in the Diablos, throughout the lava-rock hills near headquarters, at Eagle Pass, and we have credible records for Dripping Springs (Brent Martin, pers. comm.) and Quitobaquito (George Bradley, pers. comm.). Nonetheless, this subspecies appears to be uncommon in much of its desert range, and is at the arid western edge Of the species range. Its apparent desert stronghold in the Ajo Mountains qualifies it for the Red List. The rosy boa and Sonoran shovelnosed snake have been recommended for the Red List under a separate heading, as a result of road mortality and desirability to collectors. The western shovelnosed snake (Chionactis occipitalis) has been recorded at ORPI for the first time during the EMP herpetology project. It is known from Pozo Nuevo and at the west monument boundary near Jose Juan Tank. This species undoubtedly occurs at other localities in the Growler Valley at ORPI, but is not known to be abundant. We expect it along the north-central monument boundary and south in the central valley of the Ajo. It enters the monument from a larger, peripheral geographic range, and is thus recommended for the Red List. Four species of snakes appear to have experienced population declines at ORPI over the past 5 decades: (1) spotted leafnosed snake, (2) saddled leafnosed snake, (3) glossy snake, and (4) sidewinder (Table 5). These are widespread and abundant species at ORPI, deserving continued population ecology research and monitoring, but they do not need to be included on the Red List at this time. The relatively long inventory of snakes recommended for the Red List stems from several factors: (1) the ORPI snake fauna, at 25 verified species, is relatively species-rich, (2) several of the taxa are at or near edges of their geographic range at ORPI, (3) certain heroic species are especially sought by collectors, and (4) there are considerable gaps in our knowledge of snake distributions at ORPI that can only be filled by long-term observations on the species discussed. The recommendations for snakes on the ORPI Red List are summarized as follows. Two species, the rosy boa and Sonoran shovelnosed snake, are recommended because they suffer significant highway mortality and are continuously sought by private collectors. Another species, the tiger rattlesnake, is also recommended, in part because it is sought by out-of-state collectors, and in part because it is near its northwestern range limit at ORPI. Two species, the speckled rattlesnake and western shovelnosed snake are recommended because they enter ORPI only peripherally and occur in small numbers within the monument. Three species, the blacknecked garter snake, southwestern blackheaded snake, and Sonoran whipsnake, are recommended because it appears that their ecological stronghold in the northwestern Sonoran Desert may be primarily in the relatively mesic canyons of the Ajo Mountains. Species Removed from the Red List Lizards. Two lizard species that are hunted by collectors are the gila monster and the common chuckwalla. These are abundant and widely distributed on the monument, and poaching at a level sufficient to impact the population would be conspicuous and should be controllable. Therefore, we do not consider that the threat of poaching warrants inclusion of these species on the Red List.

27

28

29
Snakes. Two snake species previously on the ORPI Red List are here recommended for removal: the western coralsnake and blacktailed rattlesnake. The abundance and wide distribution of this latter species, as described above, indicates that it is not appropriately included on the Red List. The western coralsnake has been found in substantial numbers on the monument, with 26 records during the EMP herpetology project (1987-1991) alone. This is a large sample size for a secretive species. The only record from the western half of the monument is on the road at Aguajita Wash, near the Mexican border. At monument headquarters residence area, and near the East Armenta EMP site, we have found this species in abundance, with individuals ranging from juveniles to large adults more than 550 mm (21.7 in.) total length. The habitat requirements of this snake at ORPI appear to be moderately narrow, with a preference for localities that support denser, more productive vegetation, as along natural wash banks. In this part of the desert it is a xeroriparian species. Highway Mortality of Snakes on State Route 85 at the Monument Introduction. A total of 368 snakes (104 live, 264 dead) (Table 5) were observed on Route 85 from Why to Lukeville, Arizona, during 15,527 km (9,649 mi) of quantitative road-cruising, August 1987-July 1991 (Table 6). We computed a strongly conservative estimate of 1,971 snakes killed during the 4 years on this stretch of pavement, and estimate (still conservatively) that nearly 3,300 were killed. One species, the rosy boa, appears to be virtually extirpated within its cruising range of Route 85, while another, the Organ Pipe shovelnosed snake, suffers very high road mortality in its natural range within the United States-astride Route 85. The overall Highway 85 impact is equivalent to killing all snakes in 2 or more square miles of wilderness area at ORPI. The ecological significance of this mortality on snake populations deep within the ORPI wilderness is unknown. Here, we offer several possible alternatives that would mitigate, but not properly eliminate, the highway mortality of snakes at ORPI. Road driving for snakes (road-cruising), and night driving in particular, became a standard field procedure following the work of several southern California herpetologists in the 1920s and 1930s (Klauber 1931, 1939). It is well known that snakes are thigmotherms that appreciate warm road surfaces, and may rest, even coil and sit at night, during certain seasons and primarily during spring. This behavior is a contributing factor to the high road-mortality of snakes in Southwest deserts and grasslands, where most are killed when simply crossing the road. Highway mortality, as automotive traffic increases, has therefore an increasingly severe impact on resident snake populations. It is likely that snakes are more severely affected by highway mortality than is any other animal group. They are mostly slow-moving animals that can present a large target as they move across roadways. Certain species of snakes are most active when automotive traffic may be heaviest, while others may occur by chance primarily in habitats within close distance from highways. Unfortunately, 2 snake species of great interest on our study area, the rosy boa and the Sonoran shovelnosed snake, fall into this category of species populations that are most heavily impacted by highway mortality.
During the 4 years of work on the EMP herpetology, we observed and studied large numbers of live and dead snakes on the main highway, State Route 85. We used the Quantitative Road
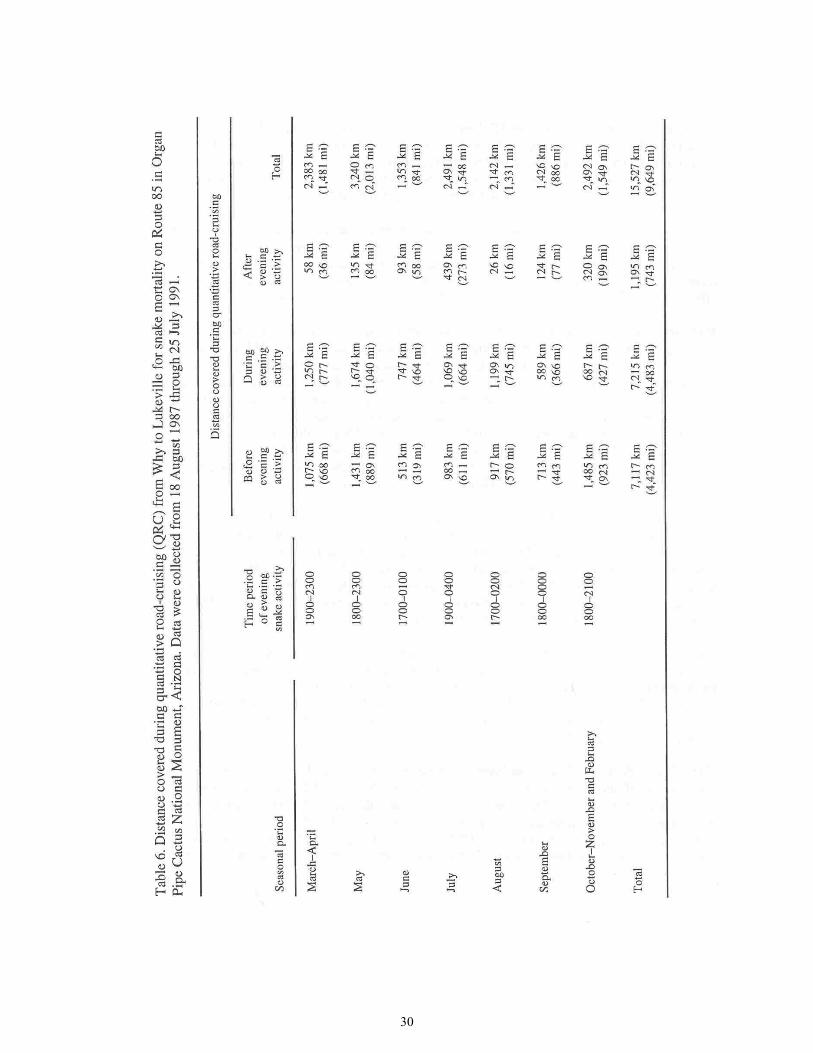
30

31
Cruising (QRC) technique, with careful record keeping of our time and place of travel, to record every snake observed, live or dead, during our EMP work. We are thus able to compute a quantitative estimate for highway mortality of snakes at ORPI for the years of study. We have additional information on the movement ecology and actual abundance of snakes on the monument, allowing us to evaluate area of impact, and severity, of highway mortality at ORPI. Both the number of dead snakes observed and the computed estimate for highway mortality are large. The impact of mortality on snake populations is substantial; so substantial, in fact, that its effect may extend away from the road into wilderness areas of the monument. Methods. Quantitative Road Cruising consists of driving an automobile slowly and recording all snakes observed on a road surface. Nighttime QRC was conducted at 15-23 kph (9-14 mph) on unpaved roads, and 35-48 kph (22-30 mph) on paved roads. During daylight, we drove at < 72 kph (45 mph). At sunset, QRC requires speed at or below 56 kph (35 mph), and during twilight hours-before automobile headlights are fully effective-the speed should be at or below 40 kph (25 mph). Constant attention is paid to the road surface at all times. Automobile windshields are kept clean. We used high-beam headlights whenever there was no oncoming traffic; these headlights were adjusted to give an optimal illumination of the full road, including 2 m (7 ft) out onto the road shoulders, from 2 m (7 ft) to 75 m (246 ft) ahead of the automobile. Most of our work was done in a 1966 General Motors Corporation pickup truck. Time, precise location, and mileage on a calibrated odometer, were recorded at the starting point of each QRC, and at each point where we turned around or moved onto a new roadway. Odometer mileage was recorded also for each observed snake, and at various, convenient, intermediate landmarks over the length of a QRC transect. The time of observation of each live snake was recorded. All snakes observed on roadways were collected, identified to species and sex, and measured and weighed. Live snakes were immediately released off the road. Dead snakes were placed on ice and held for further study. The transect we consider in this report is on Arizona State Route 85-from the town of Why (formerly Rocky Point Junction), at the junction of Routes 85 and 86, to the town of Lukeville at the international boundary. The highway distance covered is approximately 44.1 km (27.4 mi), approximately 82% of which is within the boundary of ORPI. The stretch of road most frequently covered was from monument headquarters to the north monument boundary, a distance of 28 km (17.4 mi). Data Reduction and Computations. We prepared a 38-page table of all QRC activity during our EMP work, 18 August 1987 through 26 July 1991. Each QRC transect was entered with its date, starting and ending times and locations, and all snakes observed on the transect were listed with categorization by species, sex, and size. Each snake was also categorized as "live on road" (LOR) or "dead on road" (DOR). We found many live snakes that had recently been run over by automobiles, but only 3 of these recovered; all of these snakes were categorized as DOR for this analysis because all of them would certainly have been run over by additional vehicles, and destroyed, had we not picked them up.

32
The sample of transects was subdivided into 7 seasonal periods: (1) March-April, (2) May, (3) June, (4) July, (5) August, (6) September, and (7) October-February. The data were subdivided in this way to account for seasonal differences in the intensity and timing of snake activity, and of automobile traffic in the study area. The time of observation for all LOR snakes was plotted on a histogram for each of the 7 periods. We utilized these snake-activity histograms to define peak periods of snake activity, when highway mortality was occurring. Each QRC transect was rated according to the distance covered (D), and to the time-correspondence to the observed peaks of snake activity. The time midpoint of the QRC transect was identified on the snake-activity histograms (Fig. 4). This median midpoint is a first estimate of the proportion of nightly snake activity that was encompassed by the QRC transect. It is hence the baseline estimate of the observed proportion of mortality (OPM) occurring on that night. For the latest QRC transect on each night, the proportion of the total snake mortality occurring on that night that we were able to observe for that night is given by the product:
The units of this product are completely studied miles, or CSM. The CSM is a proportion of total available miles (TAM = 27.4 mi [44.1 km]) over the entire study area from Why to Lukeville. Hence, for each of the 7 seasonal periods, the estimated observed proportion of total mortality (OPTM) is given by:
with the transect length, TAM = 27.4 mi (44.1 km). For the 24-hr period beginning at 0800, we compiled the total number of dead snakes recorded (totDOR) on each QRC transect; for each of the 7 seasonal periods, the estimated total highway mortality was computed for the 4 years of study as:
The assumptions involved in the use of equations (1)-(3) to compute total highway mortality of snakes, which are conservative, are described in the subsequent discussion section.
We have attempted also to be conservative in estimating the OPTM. (1) All QRC transects covered after 2400 were assigned OPM = 1.00; thus, we assumed that all highway mortality occurred before midnight. This was justified because the international port-of-entry at Lukeville was closed at 2400, and nearly all late-night traffic on Route 85 was cross-border activity. (2) For each QRC transect with a midpoint time at or before 2100, we added 0.05 to our baseline

33

34
estimate of the observed proportion of mortality, and for each transect with a midpoint time after 2100, we added 0.10 to the baseline estimate. Our arrival at these figures (to correct the OPM) are not entirely arbitrary, for they are based on the 2 essentials over time during a night: (a) increasing proportions of LOR:DOR snakes, associated with (b) decreasing utomobile traffic. Results. We observed 368 snakes (104 LOR, 264 DOR) of 20 species on the Route 85 transect during our 4 years of work, 18 August 1987 through 26 July 1991 (Table 5). During this work, we drove 15,527 km (9,649 mi), including 7,215 km (4,483 mi) during the peak times of evening snake activity (Table 6). The percentage LOR (28%) is identical to that reported by Fitch (1949) for a smaller road survey in Louisiana. The 3 most frequently observed species were the western diamondback rattlesnake, longnosed snake, and Sonoran shovelnosed snake; these account for 40.5% of the total sample of 368 snakes. Species characteristic of desert rockpile habitats (tiger rattlesnake, blacktailed rattlesnake, Sonoran whipsnake, lyre snake, and rosy boa) are not commonly observed on the transects because most of the transect is in middle-bajada habitat. Most species were observed alive, for an average of approximately 28% for all snakes combined. The striking exceptions are the sidewinder and the night snake (which are as expected because they are active relatively late at night) and the gopher snake, which may be explained here as a probable anomaly of small sample size. The 20 species observed on the Route 85 transect (Table 5) included all but 2 snake species that have been recorded within desert bajada and rockpile habitats in this area of Pima County, Arizona. One abundant species, the western blind snake (Leptotyphlops humilis), was unobserved; it is virtually confined to a subterranean microhabitat, and further, the few aboveground observations we have made were of small individuals that may be unobserved when they are on a highway surface. We have live-trapped several fully adult blind snakes near the road transects, but have not yet encountered adults crawling the desert floors during night foot-search with lights. The second expected snake species that we failed to observe on the highway is the rosy boa. In earlier decades this species was regularly, though infrequently, observed on our study-transect highway, with confirmed records by one of us (Lowe, pers. obs.), and in the ORPI records and preserved collection. The last-observed boa on Route 85 was 1 September 1983 (Thomas R. Jones, pers. comm.; photo voucher, The University of Arizona Herpetology Collection). Our failure to observe this species is highly significant because we made special efforts to time some of our QRC transect runs to correspond with peak boa-activity periods. Furthermore, several live boas were located at ORPI by us, other EMP researchers, and monument staff during our 4 years of study, all at distance from the highway. The rosy boa is a slow-crawling snake that is most active on the surface during cool and mild seasons-the same time that vehicular traffic is overwhelmingly heaviest at ORPI. We believe this species has been extirpated already from some parts of the monument that are near the main highway-as soon will be other snake species-by the steadily burgeoning automobile traffic.

35
The Organ Pipe shovelnosed snake is known to occur in the United States only in ORPI. We have a single record of this snake that was trapped within 3.2 km (2 mi) of Route 85. All of our other observations of it, and all other confirmed records are from the road surface of Route 85. Although this snake is often confused with local banded sand snakes (Chilomeniscus cinctus), there are several reports of it in the monument headquarters area that are probably valid. Elsewhere at ORPI and to the north, we have recorded only the western shovelnosed snake, with the 2 species not yet recorded in sympatry. The information presented here, and in Table 5, shows that the Organ Pipe shovelnosed snake has a very limited known range in the United States, and is strongly affected by highway mortality in much of this small area. The seasonal distribution of snakes observed on the Route 85 study area is presented in Table 7 for LOR snakes and in Table 8 for DOR snakes. The snakes included in Table 7, plus 4 LOR snakes that we observed being converted by vehicles into DORs, are the basis for the snake-activity histogram (Fig. 4) that we used to estimate the observed proportion of mortality (OPM), and to compute the observed proportion of total mortality (OPTM). Figure 4 shows, as expected, that the great majority of snakes are observed in the evening, and that during the summer the evening snake activity begins later and continues later into the night than during fall through spring. The low snake activity suggested by Figure 4 for 2100-2400 during July and August is an artifact of our driving schedule, which for those months was dictated by our travel to and from mark-recapture study areas. At a finer scale of resolution, we observed that peak time of snake activity on the road grew later from spring (2000) to late summer (2100), as expected, and then was earlier again in fall and winter (1900-2000). The seasonal distribution of snake observations on Route 85 (Tables 7 and 8) differs somewhat from the actual snake-activity patterns. As expected, snake activity was highest in the warm season. However, the high values (April, May) compared to lower summer values (July-September), are in part a result of our attempts to locate certain species on the highway during spring, and of our preoccupation with mark-recapture work away from the highway during peak snake activity in July-September. It is therefore not possible to infer the seasonality of snake activity from the data presented in Tables 7 and 8. The computational results show that snake highway mortality is at a maximum in spring (Table 9), when snake activity is moderate and automobile traffic has not yet reached its summer minimum. Mortality is also high during the monsoonal rainy season of July to early September, with reduced traffic and late-night snake activity partially compensating for the annual peak of overall snake activity. The computed total for snake highway mortality along the Why to Lukeville stretch of Route 85, over the 4-year period of August 1987-July 1991, is 2,383 DOR, or 596/year. For only the ORPI portion of the study area, the computed total is 1,954, or 488/year. We emphasize that these are conservative estimates, as described in the discussion, that represent minimal values for mortality. Figure 5 shows 2 models for hypothetical reconstructions of Route 85 road mortality of snakes since the monument was established in 1937. For the models figured, it is assumed that highway

36

37

38

39

40

41

42
mortality in 1940 was 33% of that computed here for 1987-1991, but it is obvious that this assumption makes little difference to the outcome estimates, unless highway mortality was much higher than assumed. The adjusted figure for number of snakes killed per year (see Discussion) in the lower graph is regarded as the most reasonable of these modelled estimates. This indicates that 32,000 or more snakes have been killed in ORPI since the monument was established, and 39,000 killed for the full study area from Why to Lukeville. Discussion. While there are, of course, several major assumptions involved in the computation of the estimates for total snake road-mortality at ORPI (Equations 1-3), the number of DOR snakes actually collected there over a 4-year period is itself a clear demonstration that highway mortality is substantial. The assumptions required for the computation presented are as follows:
1. We observed all DOR snakes that we passed on the highway. Clearly, despite the low road-cruising speeds we employed, this assumption cannot be true. Some snakes are too small to be observed, without fail, at these speeds under certain light and other conditions. Further, there was traffic each night on Route 85, and many snakes were certainly unobserved due to glare from oncoming automobile headlights.
2. All DOR snakes remained on the highway until we found and collected them. To varying
degrees this assumption is strongly violated. During daylight hours, vultures and ravens patrol Route 85 and quickly remove carcasses. During the 4 years of research, we observed only 2 fully stripped carcasses on the highway.
At night, when the majority of highway mortality occurs, dead snakes may lie on the road for hours, or even until morning. However, during certain seasons and primarily during July and August, coyotes regularly patrol the highway, eating anurans, snakes and other road-killed animals. Finally, people occasionally throw DOR snakes off the highway, or illegally collect them. We have observed numerous dead rattlesnakes from which the rattle had been removed.
3. We randomly selected nights to look for snakes on the highway. This assumption is
violated in that we were somewhat more likely to road-cruise on auspicious nights. However, most of our driving was incidental to travels to and from non-highway study areas that were visited at all times, good and bad, within each of the 7 seasonal periods defined herein.
This assumption is also violated because we were unable to conduct QRC activities regularly during the peak times of automobile traffic (Friday and Sunday nights), when snake highway-mortality was also at a maximum. This is because: (a) to do so would endanger the researchers or other motorists, and (b) the steady stream of oncoming headlights made it difficult to observe snakes on the
road. We occasionally succeeded in conducting transect activity at these times, when 2 trained herpetologists could be present in the vehicle together.

43
The violations of assumptions (1) and (2) clearly cause us to underestimate total highway-mortality, while the various violations of assumption (3) will tend to cancel each other out. We currently have no quantitative information on the effects of the violations of assumptions (1) and (2), but the violations of assumption (2) are likely to produce serious underestimation. If we arbitrarily assign a value of 40% to account for the number of road-killed snakes that are removed (by any means) from the highway before we find them, we can adjust the computed total highway mortality for the study area to a more realistic estimate of 3,972 over 4 years (993 DOR/yr), and for the monument stretch only, we would estimate 3,257 DOR snakes for the 4 yr (814 DOR/yr). Comparison to Other Studies. Only 1 other study has attempted to compute total road-mortality in snakes (Campbell 1953, 1956). Campbell (1956) estimated a total of only 0.26 DOR snakes/mi/yr (0.42/km/yr) for New Mexico highways in 1951-1953. This estimate, 100 times less than ours, may reflect vastly lower road mortality in the early 1950s in New Mexico, but more likely is a gross underestimation based on high speed driving and assumptions used for computation. Price (1983) in New Mexico, reported values for LOR and DOR per 100 mi (161 km) nighttime-driving (3.36 LOR/100 mi, 2.64 DOR/100 mi) comparable to those we recorded (2.16 LOR/100 mi, 5.18 DOR/100 mi). Price's results from Chihuahuan desertscrub are probably somewhat inflated by association with the Rio Grande riparian environment, and are thus not directly comparable to ours. Klauber (1939) presented quantitative data of 9.3 LOR/100 mi for a Sonoran Desertscrub road-cruising transect in California habitat. His transect, in the Borego Desert area, has lower primary productivity than along our transect at ORPI. Klauber's results suggest that in the early years of Route 85, prior to heavy traffic, there were many more live snakes than are seen today. The Borego sample was overwhelmingly dominated by 2 small, substratum-limited species-western shovelnosed snake and spotted leafnosed snake-that are relatively uncommon on Route 85 today, and thus might introduce bias to the comparison of Klauber's data with ours. Excluding snakes of these 2 genera, Klauber recorded 3.2 LOR/100 mi, whereas we observed 1.6 LOR/100 mi. Our observation of half of Klauber's rate, in the more productive Arizona Upland desertscrub at ORPI, supports the hypothesis that road mortality has had a substantial impact on snake populations near the highway at ORPI. ' Hensley (1950, and pers. comm.) collected 73 DOR snakes during a single year, 1949, road-cruising on the same highway transect we used. In 1987-1991, we recorded an average 66 DOR/yr covering at least as many miles as Hensley. He collected all DOR snakes that were in good condition for preservation (pers. comm.) and left the rest unrecorded, whereas we recorded all snakes. Hensley also found that live snakes strongly outnumbered dead ones; and he saw up to 40-45 snakes in a single night (pers. comm.). Hensley's results confirm that there were many more snakes prior to the increased Route 85 traffic of recent decades, and they also support our contention that the estimate of 39,000 snakes killed on Route 85 since the creation of the monument (Fig. 5) is a conservative one.

44
Ecological Impact of the Mortality. Our mark-recapture results have yielded preliminary estimates for snake abundance at 1,800 individuals per square mile for 1 study area. The study area is in a habitat supporting moderately above-average snake-population density. Thus, over the 4 years, our estimate for snake highway mortality is equivalent to approximately 5 km' (2 mi) of snake population, or to the natural snake density along the highway at ORPI to a distance of about 75 m (246 ft) on either side of Route 85. Our recapture data show also that individual snakes of most species have a home-range diameter > 150 m (492 ft), and that over several years, individuals will sometimes move farther than the home range that we observe within a year. Thus, despite the heavy mortality, the movement of individual snakes within the home range, and shifting of the home range, maintains a continuing supply of fodder for the highway meat-grinder. It is clear from observations along even more heavily travelled roads that devastating automobile traffic in the desert can, and ultimately will at ORPI, overwhelm all but the relatively thin supply of snakes that are moving beyond established home ranges. Penetration of the effects of highway mortality into monument wilderness areas cannot be assessed with any reliability at this time. While various other animals are killed on Route 85, many of these are species preyed upon by snakes or may compete with them for food. We don't know whether snakes may be drawn into the highway margins by low snake-density and/or high prey-availability. Should such a process operate, the highway would be creating a significant population sink that could reduce snake population-density deep within the wilderness. It is not fruitful to pursue further speculation along these lines until additional basic data become available or are better quantified. For the snakes, the basic data required are (1) additional and more precise estimates of natural population-density, (2) longer-term radiotelemetry data on the movements of individual snakes, and (3) information on the natural mortality and population-turnover rates in the absence of highway mortality. It would be useful to learn how other ecosystem components, especially rodents and lizards, are affected in proximity to the highway, and to the extent that they are, we should learn whether snakes respond to the highway-induced changes.

45
Lizard Monitoring
Data Handling, Analytical Procedures, and Baseline Results Introduction The objective of the Lizard Monitoring Protocol (Rosen and Lowe 1995) is to measure population changes in lizards that can be correlated with natural and human-caused environmental changes at ORPI. The lizards and the findings for these ectothermic vertebrates have intrinsic biological importance. They also form one component within the broader EMP study that is planned to be capable of detecting biotic effects of global climate change, of local human-caused disturbance, and of natural environmental fluctuation. The potential exists to document the immediate effects of environmental fluctuations on lizards, and to use this information to predict and/or illustrate the consequences of human-caused environmental change at ORPI. This section contains a synthesis and overview of the 1987-1990 lizard monitoring results, analyses and conclusions. We review data handling, storage, and evaluation, and discuss appropriate statistical methods that may be employed. The findings will guide the ORPI resource management staff in decisions regarding what data to collect, and how to evaluate data reliability. The analysis presented here may serve as a model for future workers to use in analyzing future data that are to be collected over many years at the intensive-monitoring study areas discussed herein. All of the analyses and syntheses presented in this section are based on the data in Site-specific Results on Intensive Study Areas. Included there is additional information on habitat type sampled by each transect; the section serves as a reference to Data Handling, Analytical Procedures, and Baseline Results (for the supervising resource manager). Overview of Methods The intensive methodology used for this report consists of standardized lizard line (SLL) transects, and time-standardized sampling (time-constrained search [TCS]). Both methods are detailed in Rosen and Lowe (1995); only summary descriptions are given here. The lizard lines are permanently located, walk-line transects standardized for recording all observed lizards in a 15-m-wide (49-ft) belt, 7.5 m (24.6 ft) on either side of the walked midline. The line is walked several times during a day beginning at or prior to the emergence of diurnal lizard species. The walks during the course of 1 day comprise a run. The primary data points generated during a run are the maximum number of lizards observed during any 1 walk, for each species. These species maxima are termed the peak values: they represent the best estimate for abundance of each species on the line at run-time. The sum of peaks over all species is the total estimated lizard abundance. We refer to the maximum peak for a species over a series of runs, over seasons or years, as the maxpeak.
The TCS method is similar to general herpetological field surveying. The investigator walks through the habitat observing lizards and snakes, and investigates potential shelter sites (within vegetation, under rocks, logs, etc.). The observed number for each species is recorded, and, as in the lizard line procedure, size (age) class, sex, and natural history observations are recorded. Like

46
SLL, TCS may be carried out within a single habitat type (i.e., wash xeroriparian, open desertscrub), or it may include an entire canyon or EMP site. Statistical Methods. The peak and maxpeak data points obtained for each run or for a series of runs at each site are used to compute species richness (S, the number of species observed) and species diversity (H', H-prime):
H'= -SUM (pi LOG pi)
where pi is the peak for species i, divided by the total peak over all species; and LOG is in base 10. The analytic focus of the monitoring program is to obtain an index of abundance that will document the status of, and the temporal changes in, lizard assemblages. The peak values and diversity indices are compared across sites, macrohabitats, habitat types, and-most importantly-years. Preliminary data examination has revealed interesting patterns for the different species and among seasons and years. Observation of these patterns served as a guide for the analyses performed. The number of sites studied was too small to permit quantitative tests for normality, so a conservative approach was adopted. Graphical examination of the data distributions showed moderate deviations from the normal distribution, with marked deviations from normality in sideblotched lizards. One-way analysis of variance (ANOVA, with a single classification variable testing for differences in the dependent variable, "peak value") is not highly sensitive to deviations from assumed normal, homoscedastic data distributions, and could therefore generally be applied to the lizard line data, with the exception of the data from sideblotched lizards. The second step in analysis proceeded with one-way ANOVA for the following categorical variables: (1) EMP site (= locality), (2) macrohabitat (montane, bajada, valley floor, and floodplain), (3) habitat (upland desertscrub vs xeroriparian desertscrub), (4) season (spring vs summer), and (5) year (1989 vs 1990). These ANOVAs were completed for each species and also for the combined results (= sum of the peak values for each species for a run). Two-way ANOVAs were also run to scan for statistically significant interactions between the classification variables. Each of the 5 categorical variables listed above was found to be highly significant in some, though more usually, most comparisons. The third step of analysis was to use non-parametric methods to estimate the statistical significance of the several classification variables affecting lizard abundance (peak value). The strongest categorical variable discovered by ANOVA was EMP site. Therefore, for confirmatory tests, paired-sample t-tests were chosen, and the non-parametric Wilcoxon Signed Rank test and Kruskal-Wallis test (all with pairing within site across years), whenever possible. The sign test was applied to highly skewed data recorded from sideblotched lizards. Finally, the results of analysis are presented in the format of (1) descriptive statistics (tabular or graphed) and (2) t-tests and non-parametric tests.

47
Baseline Results The peak numbers of observed lizards per 200 m (656 ft) of lizard line are presented in Table 10. There was significant variation over the 2 years of baseline study, with all species showing decreases from spring 1989 to summer 1990, except with the sideblotched lizard species, which increased. The most abundant lizards on the lines were the sideblotched lizard, the western whiptail, and the tree lizard. Variation among Sites. The variation among sites for 1989 and 1990 is shown in Figure 6. While there were substantial declines in lizard abundance from 1989 to 1990 at several sites, there is nonetheless a clear pattern for consistent results within sites. The consistent aspects among sites are described below under separate headings. Patterns of species diversity (H') and richness (S) are illustrated in Figure 7. It is apparent that species richness is correlated with species diversity. The effects of excessive disturbance are apparent in the low species richness and diversity at Burn Site and at Armenta Ranch Site. Both of these plots are on valley floor supporting mesquite-dominated communities that elsewhere have higher lizard diversity and abundance. Habitat Effects. The general habitat categories we have used for lizards are (1) xeroriparian (tree-form) desertscrub and thornscrub, (2) mesquite-dominated desertscrub, (3) paloverde-ironwood dominated desertscrub, and (4) creosotebush-saltbush-bursage desertscrub. For certain ANOVA models, categories (2) and (3) were combined into a composite category, "tree-form upland desertscrub"; this gave roughly equal cell sizes as demanded by the ANOVA model. The observed abundance of lizards in the habitat categories is shown in Table 11. Habitat was a significant factor for the western whiptail, the tree lizard, the desert spiny lizard, and for total lizards (Kruskal-Wallis tests, all P < 0.04), but not for the sideblotched lizard or the zebra-tailed lizard (Callisaurus draconoides). The higher value for abundance of sideblotched lizards in mesquite-dominated desertscrub (Table 11) was produced by results from a single, atypical, burned area (Burn Site). This species appeared to be the most successful colonizer after the fire, and achieved its highest density there. The only other transect with similarly high density was on the rocky hill at Pozo Nuevo Site. It appears that the sideblotched lizard reaches its greatest abundance where other lizard species have relatively low population density.
The habitat preferences suggested by Table 11 generally correspond to other, extensive observations for these lizard species; however, drought-induced lizard population declines that occurred in all paloverde and ironwood communities, as well as in most of the creosotebush, saltbush, and bursage communities during the period of study (below), have exaggerated some aspects of habitat suitability suggested by the static view given in Table 11. Observations indicate that, consistent with Table 11, both the sideblotched lizard and the zebra-tailed lizard were habitat generalists, whereas spiny-tail species and the tree lizard were habitat specialists on large woody shrubs or small trees.

48

49

50
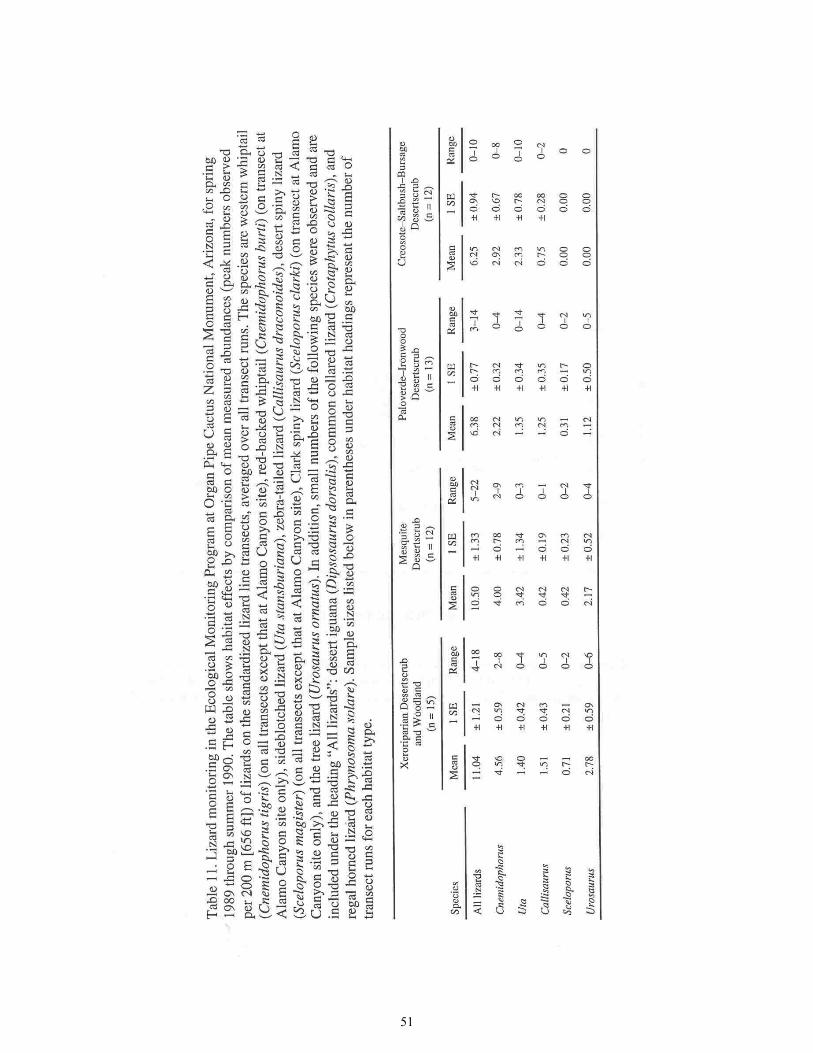
51

52
On a finer scale, within the 4 defined habitat categories, the baseline data (Fig. 6) suggest what is observed elsewhere as well, that abundances of all of the lizards except the sideblotched lizards are (1) highest in mesquite bosque communities (Growler Canyon Site, Aguajita Wash Site) and (2) lowest in open desertscrub communities dominated by creosotebush (Larrea divaricata) and white bursage (Ambrosia dumosa) (Pozo Nuevo dumosa Bursage Site Lizard Line, Creosotebush Site Lizard Line) and on open, rocky slopes (Pozo Nuevo Hill-Base Site Lizard Line). This is illustrated in Figure 8. Decreased Activity Levels at the Drought Maximum. The deepest period in the drought of the late 1980s occurred at ORPI during summer 1989. On much of the southern third and western two-thirds of the monument, rainfall was so poor that no primary production occurred. This followed an unusually hot foresummer dry season, and was within a longer period of poor rainfall. The peak values recorded for lizards on SLL transects were markedly low during summer 1989, particularly at sites receiving the least rain (Fig. 9). This effect was at least partially an activity reduction by individuals rather than population size reduction, as shown by rebounds in 1990 in spite of poor reproduction and recruitment of new individuals into the populations (see below). This indicates that over short time, caution is required for interpretation of changing peak values on standard lizard line transects. Lizards are less active during nonreproductive seasons, translating to lower peak values on the lizard lines. Year and Macrohabitat Effects. Macrohabitats are defined as (1) montane, (2) bajada, and (3) valley floor floodplain (Fig. 10). There is a clear gradient of increasing lizard abundance from rocky slopes across the bajada to a maximum on the valley floor. Figure 10 also illustrates that, orthogonal to the slope gradient of abundance, lizard abundance at ORPI is higher in xeroriparian communities than in upland communities.

53

54

55
It should be clear from Figure 10 that additional EMP sites are required to sample adequately the major herpetofaunal habitat axes at ORPI; certainly the same is true for other ecosystem components. Most importantly, upland desertscrub communities on strictly valley floor are not represented by EMP sites; that is, especially needed is a site representing the Larrea consociation on level ground of the valley floor, where Lower Colorado Valley herpetofaunal elements (longtailed brush lizard and desert horned lizard) are best represented. Additionally, only a small portion of 1 EMP site (Pozo Nuevo Hill-Base; not studied by all EMP component projects) includes a lizard line in the widely occurring upland desertscrub community on rock slopes at ORPI-on the rockpiles in Shreve's (1951) Arizona Upland subdivision of the Sonoran Desert. The lizard lines in desert mountain environments are Alamo Canyon South Fork Site (xeroriparian habitat), and Pozo Nuevo Hill-Base Site (rocky upland desertscrub habitat). Bajada study areas are East Armenta Kuakatch Wash Site, Lizard Grid Site, Creosotebush Site, Pozo Nuevo Wash Site, Pozo Nuevo dumosa Bursage Site, Pozo Nuevo deltoidea Bursage Site, Vulture Site, and Senita Basin Site. These sites are on sandy or gravelly loam, and are in paloverde-ironwood dominated desertscrub or Ceosotebush-bursage desertscrub.
Valley floor floodplain study areas were Armenta Ranch Site, Growler Canyon Site, Aguajita Wash Site, Burn Site, Dos Lomitas Site, and Salsola Site. With the exception of wash channels, silty soils and flat terrain predominate at these sites. All of these study areas are in communities

56
where mesquite is dominant or is significantly present. Burn Site and the Aguajita Wash Saltbush Site Lizard Line are in saltbush-dominated vegetation communities. Macrohabitat was a significant factor in lizard abundance for all species except the sideblotched lizard (Kruskal-Wallis tests, all P < 0.03). The major finding was the consistent decrease in lizard abundances from spring 1989 to summer 1990, on bajada sites, in contrast to the statistically nonsignificant increase in lizard abundance on valley floor floodplain sites (Tables 12 and 13). With the exception of the sideblotched lizard, all of the 5 most abundant lizard species showed statistically significant declines in bajada macrohabitat (Table 14). The observed increases in the sideblotched lizard species are probably real for both the bajada and valley floor floodplain communities, but the data distributions for this species were so skewed that only the relatively weak Sign Test could be applied. The lizard population declines observed on the lizard lines in 1989-1990 are part of a longerterm pattern initiated in 1985 with the onset of drought and the end of a relatively wet period that began in 1977-1978 (Fig. 11). Figure 11, for the lower bajada, Pozo Nuevo Site, shows that lizard population levels in 1990 decreased to half, or less, of their levels at the onset of drought 5 years earlier. The summer rains of 1990, initiated in the first week of July, dramatically broke the 5-year drought. The lizard line results show lizard population stability in valley floor and valley floor floodplain communities despite the drought, in contrast to sharp declines observed in the bajada communities. This may result from greater summer and winter annual plant (ephemeral) productivity and greater moisture-holding capacity of soils of valley floor and valley floor floodplain communities. An additional factor involved in the population stability appears to be runoff from a relatively small number of intense rainstorms that occurred during the drought, particularly during the winters of 1987-1988 and 1988-1989. These storms produced little response in ephemeral vegetation on bajada areas, whereas floodplain communities (notably Armenta Ranch Site, Dos Lomitas Site, and Salsola Site) apparently received sheet-flooding, and had substantial annual productivity in the late winter and early spring of both 1988 and 1989. Studies of reproduction and age-structure in the indicator species, western whiptail and red-backed whiptail shed some light on the relationship of these enhanced moisture levels to lizard population size.
. Age-structure Effects and Mechanisms of Population Change. Intensive observations on the Lizard Grid Site indicated that poor rains in summer 1989 produced mass mortality of juvenile western whiptails during fall 1989. No growth was recorded among the few hatchlings recaptured during this time, and we never found juveniles larger than hatchling size. Several of the hatchlings captured were weak and emaciated; 1 extremely emaciated hatchling expired during handling. During the subsequent spring, April-May 1990, no observations were made of gravid or pre-gravid female western whiptails, in contrast to pronounced peaks of gravid females observed in spring of 1988 and 1989. Further, there were no hatchlings in July 1990, again in contrast to the previous 2 years. Throughout spring 1990, adults of both sexes were underweight, although not emaciated. Clearly there was little, if any, egg production by the resident western whiptails on

57

58

59

60
the Lizard Grid Site in April-May 1990. Elsewhere on the monument, we observed a few hatchlings and juveniles, at scattered localities, produced as eggs in spring 1990.
During the early-arriving and strong rains of summer 1990, all adult female western whiptails were found to be gravid or immediately post-depositional during 2 time periods, mid-July and early mid-August. In contrast, during 1988 and 1989, there was a single reproductive peak during summer, and thus apparently produced only a single clutch during the summer rains in these years. Population declines observed in the western whiptail during the 1989-1990 drought were accompanied by decreases in proportions of young individuals (Fig. 12). Figure 13 shows that the pattern in Figure 12 holds over a longer time period at the site sampled from the early onset of drought in 1985. In Figure 12, it can be seen that an abundance of juveniles in one season is followed by a large cohort of subadults in the next. On the valley floor and valley floor floodplains, there were high proportions of nonadult lizards well into spring 1990, with drought effects appearing in the age- structure only in summer 1990. This confirms that reproduction occurred-and sustained the populations-on these sites until summer 1989 and spring 1990. Bajada populations had markedly decreased reproductive success as early in the drought as spring 1988, more than 2 years before these effects were observed on the valley floor.

61

62
Further, on the bajada, adult-only population structure occurred in spring 1990, a full reproductive season earlier than on the valley floor (Fig. 12). The hatchling die-off observed on the bajada at Lizard Grid Site in 1989 was less evident on the valley floor. Age-structure data in Figures 12 and 13 confirm that reproductive failure was the prime cause of the observed population decline for western whiptails. It is remarkable that, despite severe drought, whiptail populations did not decline on the valley floor sites. This is even more surprising in view of the low proportions of non-adult lizards (< 40%, Figs. 12 and 13) observed in valley floor communities during 1987-1990, and the clear evidence that reproductive failure does occur there (in spring 1990, especially) during dry periods. It is entirely possible that emigration of lizards into the valley floor floodplain communities, from nearby surrounding upland desertscrub communities, may have been the main factor producing the appearance of population stability. This hypothesis cannot be confirmed because, as noted above, there are no EMP sites in the upland desertscrub communities of the valley floor. However, there are 2 pieces of evidence from the middle bajada sites at Kuakatch Wash (Lizard Grid Site, East Armenta Site) that support an emigration hypothesis. First, SLL transects showed that upland desertscrub lizard populations decreased earlier than xeroriparian populations, although both ultimately declined to low levels. Second, we recorded long distance home-range shifts (up to 200 m [656 ft]) by marked whiptails. These home-range shifts were from areas of lower to higher quality habitat types (see Fig. 6 and Table 11). Study results indicate that valley floor floodplains are refugia from drought at the species population level. This underscores the need to consider the valley

63
floors, so routinely usurped by man, as critical ecosystem components; it reinforces the need for additional EMP monitoring in the upland desertscrub communities of the valley floor. In the zebra-tailed lizard, variation in the abundance and distribution of hatchlings also strongly suggests that reproductive success is closely related to moisture availability. Zebra-tailed lizard hatchlings were never observed after dry seasons, except for a very few individuals at one locality-Cuerda de Lena Wash, along the north monument boundary-that is a highly productive, mature valley floor floodplain. This suggests that egg production was more sharply affected by drought in the zebra-tailed lizard than in the western whiptail. In zebra-tails, limited evidence confirms that valley floor floodplain communities serve as refugia during drought. Faunal surveying and SLL transect work should be conducted, not only at disturbed floodplain sites (Salsola Site, Dos Lomitas Site, Burn Site, and Armenta Ranch Site), but also within relatively healthy examples of climax communities in the northwestern area of ORPI (for example, south of Armenta Ranch Site, at Cuerda de Lena, and in the Growler Wash system and Paloverde Camp).
For other lizard species, we do not yet understand the specific mechanism for drought-induced decline (low egg production, low egg survival, low juvenile survival, and low adult survival). We suspect that increased abundance of sideblotched lizards during the 1989-1990 drought resulted from population declines in several predators, especially lizards and snakes, that prey on this short-lived species. Important known predators on sideblotched lizards for which we observed population declines are the night snake, the leafnosed snake, and the lizards: western whiptail, desert spiny lizard, and longnosed leopard lizard. It is probable that several bird species preying on the sideblotched lizard also had reduced population sizes during the drought. These observations of extreme reproductive failure at the population level suggest that unusually low rainfall reduces lizard populations by reducing reproductive success. Elevated clutch frequency during the strong rains of summer 1990 also confirm that the ultimate source of population fluctuations in western whiptails was drought, and the proximate mechanism was, in general terms, nutritional deficiency. However, the specific nutritional problem(s)-water balance, energy shortage, and nutrient deficiency-and/or factor interactions remain unstudied. The important contribution of predation in modulating lizard population fluctuation, as well as in density-dependent damping of lizard population fluctuations, also remains largely unknown at ORPI. The effect of predation on lizard populations has rarely been estimated (see Holm 1988). Lizard Populations and Biomass at ORPI. The combination of (1) mark-recapture estimates of population size with (2) SLL transect techniques, both conducted on the Lizard Grid Site, allows preliminary computation of estimates of lizard population densities on the SLL transects throughout the monument. The study estimated the population density, excluding hatchlings and small juveniles, for western whiptails, desert spiny lizards, and sideblotched lizards on the Lizard Grid Site in 1989 and 1990 using mark-recapture methods (Table 15). The total of 154 individuals/ha estimated for strictly diurnal species only, would be increased to about 200 individuals/ha if the study included the

64

65
desert banded gecko (Coleonyx variegatus variegatus) and gila monster. During seasonal periods of hatchling and juvenile abundance, the number of lizards can be expected to approach 700/ha, and to exceed this in more productive habitats (Table 11) and during wetter periods (Fig. 11). It is appears that lizards may be the most abundant vertebrates at ORPI. The density of western whiptails was estimated in 1990 on the Lizard Grid Site, yielding a 30% decrease from 1989. This was similar to the decline observed elsewhere on the bajada for this species (see Table 13, 37%). In 1990, at the Lizard Grid Site, 7.5 lizards/200 m (656 ft) of SLL transect were observed-close to the average of 7.61 lizards/200 m (656 ft) for all ORPI transects the same year. In 1989, the average lizard count for all ORPI transects was 9.54, or 25% greater than in 1990. Using these figures, and the estimated 154 diurnal lizards/ha on the Lizard Grid in 1989-1990, it is possible to calculate an estimated 196 lizards/ha at ORPI on the area sampled by SLL transects in 1989-1990, translating to 26 million diurnal lizards on the monument in 1989. This gives a rough idea of the magnitude of the monument lizard population. A preliminary lizard biomass estimate of 1.44 kg/ha on the Lizard Grid Site, exclusive of hatchlings and small juveniles, similarly yields an estimate of more than a quarter-million kg (551,150 lb) of lizards for ORPI, or 276 tons as a standing crop. (This is roughly equivalent to the mass of 5,000 people, or 29,000 coyotes.) At nearly 1.5 kg/ha, the biomass of lizards may be greater than that of rodents, and nearly twice that of snakes. Lizards may comprise the largest part of vertebrate biomass in the Sonoran Desert ecosystem. Summary and Conclusions Methods The principal monitoring methodology for lizards is the standardized lizard line transect. The data objective of the transects is to obtain the peak value-the highest number observed in any walk during a daily run-for each lizard species. The secondary methodology is time-constrained search-general herpetological field activity during appropriate times of day and year, with careful record-keeping for time, activity, location, and lizards and snakes observed. With experience, data on age-structure in lizard populations can be obtained both on transects and during time-constrained search. Transect data should be photocopied and stored. The peak values are to be read from the data forms and stored in the dataset. The supervising resource manager is to scan the field data forms to ensure that proper technique is followed in the field. Improperly collected data are to be excluded from the dataset. Statistical analysis can be separated into (1) preliminary examination of data, (2) exploratory analysis using ANOVA and other appropriate techniques, and (3) confirmatory analysis using non-parametric tests and, whenever appropriate, parametric tests. Results Previous transect data have shown strong trends among EMP sites, as well as among habitats, macrohabitats, seasons, and years. During 1989-1990, lizards were most abundant in mesquite

66
bosques and other xeroriparian communities, and least abundant in shrub-dominated upland desertscrub; abundances increased down the gradient from rock slope to valley floor. Seasonality was most apparent in the transect data when juvenile recruitment was high, but adult activity levels also appeared to influence results. During the drought of the late 1980s, lizard population sizes decreased to half or less of pre-drought levels, at least on some sites. Very low rainfall observed at several sites during summer 1989 produced decreased activity of lizards, and yielded low peak values on the lizard lines. On the transects, lizard abundance declined significantly from 1989 (a year of severe drought at ORPI) to 1990 on bajada sites but not in valley floor floodplain environments. Consistent decreases were also observed on the 2 desert mountain (rockpile) transects. Population decline appears to be caused proximally by reproductive failure-both as reduced egg production, and as high juvenile mortality, as especially well seen in the species with the most robust data sets. Populations of one species, the sideblotched lizard, increased during the drought. Valley floor floodplain environments, represented by 8 transects, appeared to be refugia from drought for lizard species. Population trends in surrounding upland desertscrub, primarily in Larrea-dominated communities, were not studied, and emigration from these to the floodplains may help explain population stability in the floodplains. Site-specific Results on Intensive Study Areas Introduction We present here the baseline database from our 1987-1990 work, in both summary and illustrated form. Location, length, and habitat of all standardized lizard line transects established by the herpetology research team are described. Each EMP study site-and each additional site where lizard lines were established-is presented separately. This section also reports and illustrates the baseline monitoring results obtained by the research team. All lizard line results are presented under the heading of each intensive study site, while time-constrained search results are combined into a single table at the end (Table 16). Standardized Lizard Line-transect Results The following 13 sites were treated with lizard line-transects. An asterisk (*) indicates a transect site near, but not on, an EMT site.
Aguajita Wash Site Dos Lomitas Site Pozo Nuevo Site Alamo Canyon Site East Armenta Site Salsola Site Armenta Ranch Site Growler Canyon Site Senita Basin Site Burn Site Lizard Grid Site* Vulture Site Creosotebush Site*
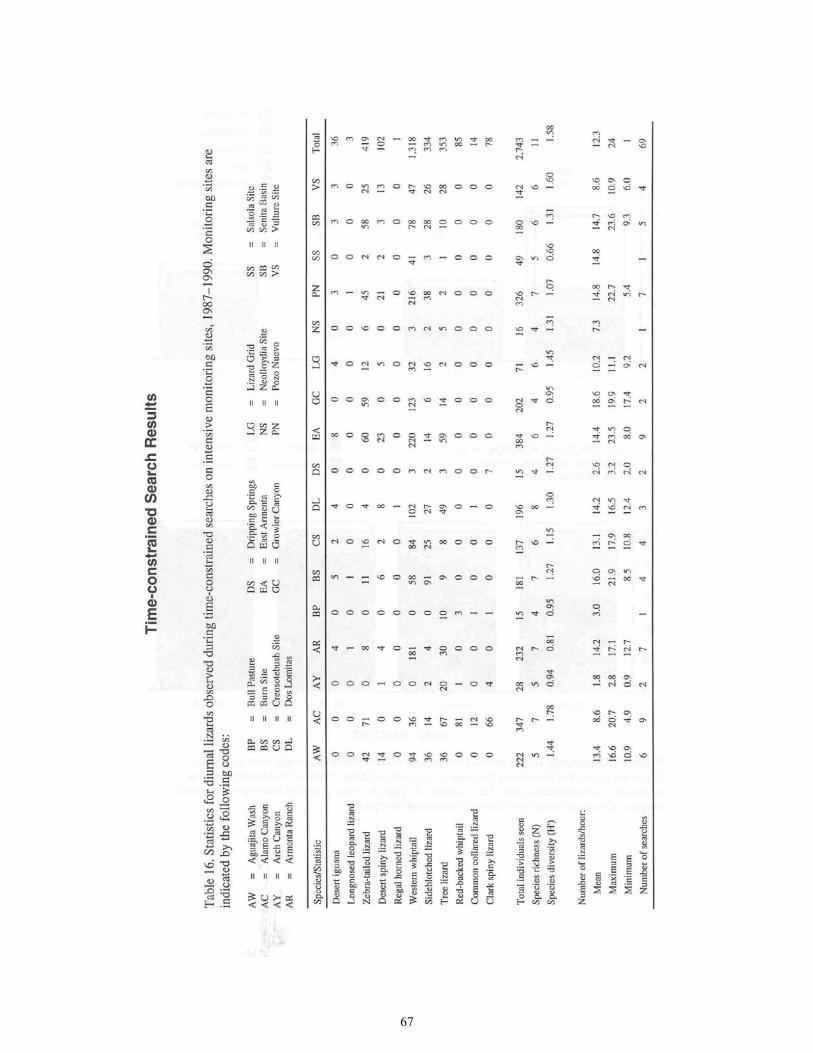
67

68
Data were obtained at the following EMP sites by time-constrained search (only), rather than by standardized lizard line transects:
Arch Canyon Site Dripping Springs Site Bull Pasture Site Neolloydia Site

69
Lizard Study Sites

70
Aguajita Wash Site
Number of standardized lizard line transects: 2 Length of the transects: 100 m (328 ft) each Habitat type on each transect: Transect 1 (Saltbush Line) is on the upper floodplain of Aguajita Wash in a dense stand of allscale (Atriplex polycarpa) with scattered, large honey mesquites (Prosopis juliflora). Transect 2 (Bosque Line) is on the floodplain of Aguajita Wash near the main wash channel, in open and dense mesquite bosque with mixed shrubs: honey mesquite, catclaw (Acacia greggi), allscale, creosotebush, and others. H-prime (H') for lines combined: 1.47 Baseline data: Peak number of individuals for each lizard species during each run, as follows:
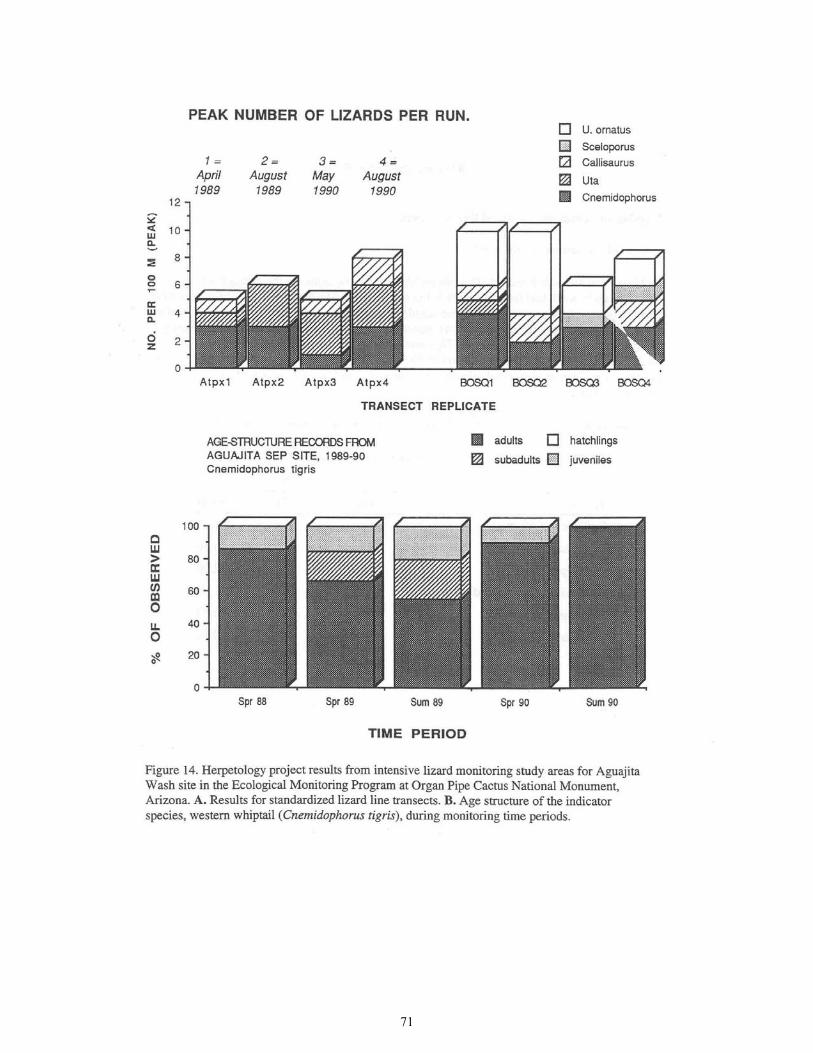
71

72
Alamo Canyon Site
Number of standardized lizard line transects: 1 Length of the transect: 300 m (984 ft) Habitat type on each transect: The transect runs down the center of the South Fork rocky-gravelly wash bed from boulders at the mouth of the Middle Fork. The upper 66% of the transect passes through sparse and dense stands of mature Ajo oak (Quercus ajoensis), and also has net-leaf hackberry (Celtis reticulata), one-seed juniper (Juniperus monosperma), bursage (Ambrosia ambrosioides), graythorn (Zizyphus obtusifolia), desert broom (Baccharis sarathroides), and many others. The lower 80 m (263 ft) of the transect is dominated by Lower Sonoran xeroriparian plants including honey mesquite, catclaw, with hop-bush (Dodonea viscosa) and others. Baseline data: Peak number of individuals for each lizard species during each run, as follows:
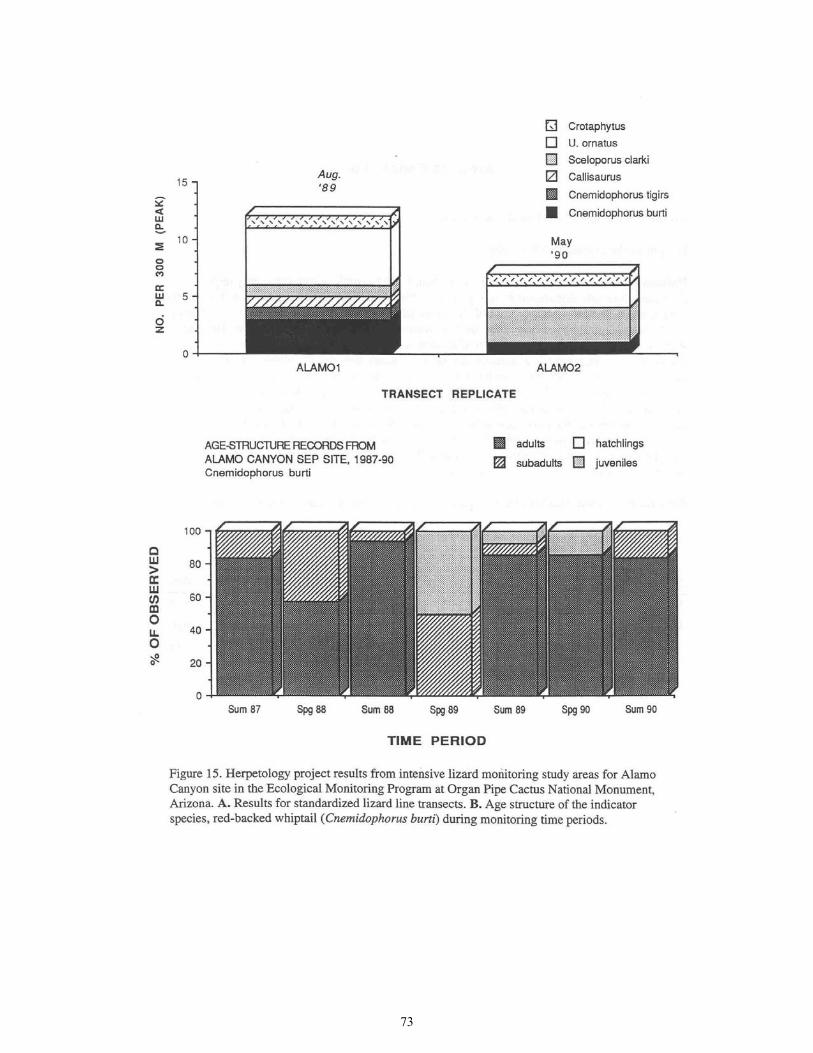
73

74
Armenta Ranch Site
Number of standardized lizard line transects: 1 Length of the transect: 200 m (656 ft) Habitat type on transect: The transect runs from north to south, across the highly degraded floodplain formerly dominated by mesquite and others that were more than likely in bosque-like community structure. Essentially all of the perennials except large mesquites were removed in the Armenta farming operation; there are few mature plants on the open floodplain. The transect starts at a small sandy wash braid (part of Kuakatch Wash) in dense honey mesquite with large individuals of creosotebush, progresses across the virtually denuded floodplain among 4 large, isolated and some scattered successful recruits of honey mesquite and several successful recruits of blue paloverde (Cercidium floridum). The southernmost 20-25 m (66-82 ft) of the transect is on open upland of creosotebush. A major thicket of diverse annuals, dominated in summer by quelite (Amaranthus palmeri), lies astride the central 50 m (164 ft) of the transect, and in wet times the annuals expand and cover the entire transect with at least low growth. This transect is situated clearly in a disclimax that is in cyclical flux; it appears to be changing directionally toward re-establishment of natural floodplain vegetation. Baseline data: Peak number of individuals for each lizard species during each run, as follows:
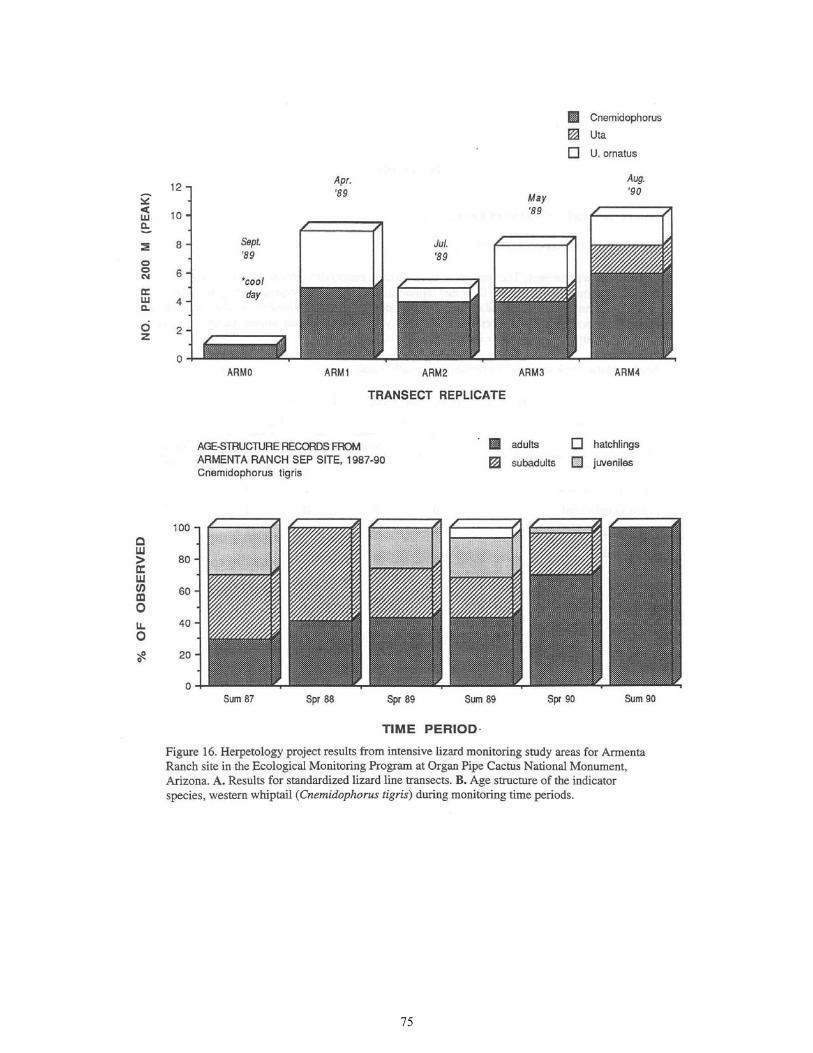
75
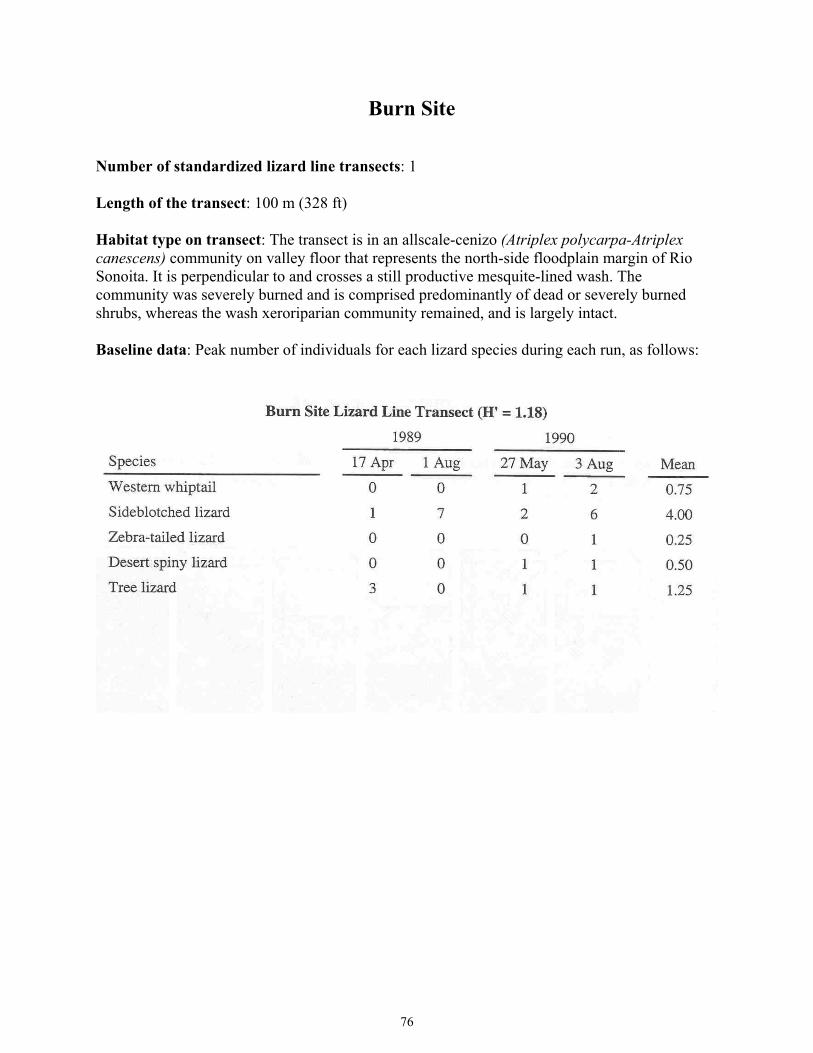
76
Burn Site Number of standardized lizard line transects: 1 Length of the transect: 100 m (328 ft) Habitat type on transect: The transect is in an allscale-cenizo (Atriplex polycarpa-Atriplex canescens) community on valley floor that represents the north-side floodplain margin of Rio Sonoita. It is perpendicular to and crosses a still productive mesquite-lined wash. The community was severely burned and is comprised predominantly of dead or severely burned shrubs, whereas the wash xeroriparian community remained, and is largely intact.
Baseline data: Peak number of individuals for each lizard species during each run, as follows:

77

78
Creosotebush Site
Number of standardized lizard line transects: 1 Length of the transect: 200 m (656 ft) Habitat type on transect: The transect runs from north to south and is bisected by Armenta Road (= North Boundary Road) at 1.45 km (0.90 mi) (rd) east of Armenta Ranch house. It is at the lowest bajada edge close to valley floor, in a creosotebush-white bursage (Larrea divaricata Ambrosia dumosa) community that crosses a minor washlet. Baseline data: Peak number of individuals for each lizard species during each run, as follows:

79

80
Dos Lomitas Site
Number of standardized lizard line transects: 2 Length of the transects: 100 m (328 ft) each Habitat type on each transect: Transect 1 (Inside Exclosure) is on the north-side floodplain margin of Rio Sonoita. It is in an allscale-cenizo (Atriplex polycarpa-Atriplex canescens) community also dominated by an open stand of honey-mesquite. Annual plants, especially quelite, grow profusely on the line during wet summers. Transect 2 (Outside Exclosure) is similar to Transect 1, but has less mesquite and annual growth, and in part traverses an open stand of Atriplex. H-prime (H') for lines combined: 1.35 Baseline data: Peak number of individuals for each lizard species during each run, as follows:

81

82
East Armenta Site
Number of standardized lizard line transects: 2 Length of the transects: 200 m (656 ft) each Habitat type on each transect: Transect 1 (desertscrub) is on essentially flat middle bajada, in foothill paloverde (Cercidium microphyllum), white bursage, desert-ironwood (Olneya tesota), creosotebush, saguaro (Camegiea gigantea), and others. Transect 2 (Kuakatch Wash) is in adjacent xeroriparian scrub dominated by large desert-ironwoods with catclaw, desert-thorn, (Lycium andersoni), cheese-bush (Hymenoclea salsola), and others. H-prime (H') for lines combined: 1.45 Baseline data: Peak number of individuals for each lizard species during each run, as follows:

83

84
Growler Canyon Site
Number of standardized lizard line transects: 2 Length of the transects: 100 m (328 ft) each Habitat type on each transect: Transect 1 (Wash Bed) starts at the point of the confluence of Cherioni and Cuerda de Lena Washes, goes across the sand diagonally and parallels the north bank of Growler Wash. The scanty perennial flora on the wash bed is principally cheese-bush. Transect 2 (Bosque) parallels the north bank of Growler Wash, near Transect 1. It is in dense bosque dominated by honey mesquite, catclaw, crucifixion-thorn (Castela emoryi), creosotebush, and others. H-prime (H') for lines combined: 1.09 Baseline data: Peak number of individuals for each lizard species during each run, as follows:

85

86
Lizard Grid Site
Number of standardized lizard line transects: 2 Length of the transects: 100 m (328 ft) each Habitat type on each transect: Both transects are in a middle bajada ecotone. Transect 1 (North) is amid foothill paloverde, saguaro, white bursage, creosotebush, jumping cholla (Opuntia fulgida), with others. Transect 2 (South) is in similar habitat but has much more big-galleta (Hilaria rigida), desert-thorn, and barrel cactus (Ferocactus wislizeni). H-prime (H') for lines combined: 1.42 Baseline data: Peak number of individuals for each lizard species during each run, as follows:

87

88
Pozo Nuevo Site
Number of standardized lizard line transects: 4 Length of the transects: 100 m (328 ft) each Habitat type on each transect: Transect 1 (Hill Base) is on the lower slope of the large hills just north of the camp at Pozo Nuevo. It has medium-sized rocks throughout, and in some places there are rockpiles. The vegetation is dominated by burrobush (Ambrosia deltoidea), foothill paloverde, brittlebush (Encelia farinosa), organ pipe cactus (Stenocereus thurberi), ocotillo (Fouquieria splendens), and others. Transect 2 (Wash) is in the xeroriparian and washbed strand of "Pozo" Wash near the camping area that is 0.16 km (0.10 mi) west of the corral. Dominant plants include honey mesquite, blue paloverde, cheese-bush, burrobush, and creosotebush. Transect 3 (dumosa Bursage) is on slightly rolling upland with widely scattered volcanic rocks. The vegetation is creosotebush-white bursage (Larrea divaricata Ambrosia dumosa). Transect 4 (deltoidea Bursage) is on flat upland along a small drainage. The vegetation is dominated by creosotebush, burrobush, and honey mesquite. Each of the transects lies within 1 of 4 long-term rodent monitoring grids previously established by Yar Petryszyn. H-prime (H') for lines combined: 1.09 Baseline data: Peak number of individuals for each lizard species during each run, as follows:

89

90

91

92
Salsola Site
Number of standardized lizard line transects: 1 Length of the transect: 200 m (656 ft) Habitat type on transect: The transect runs through a productive mesquite-creosotebush (Prosopis-Larrea) floodplain habitat on the perimeter of the north-side floodplain of Rio Sonoita. Annual plants, especially Russian-thistle (Salsola), combine with the diverse perennial floodplain flora to form a substantial thicket, and in many areas globe-mallow (Sphaeralcea) and perennial grasses form lower continuous ground cover. Baseline data: Peak number of individuals for each lizard species during each run, as follows:

93

94
Senita Basin Site
Number of standardized lizard line transects: 1 Length of the transect: 250 m (820 ft) Habitat type on transect: The transect runs roughly south to north across the open xeroriparian scrub characteristic of the main drainage on the lower portion of the EMP site. Important plants of this diverse community are senita cactus (Lophocereus schotti), creosotebush, burrobush, and desert-ironwood, with many others present. The southernmost 35 m (115 ft) of the transect are on a rocky hill slope dominated by foothill paloverde, jatropha (Jatropha cuneata), and burrobush. The northernmost 40 m (131 ft) of the transect are on flat, rich desertscrub that grades away from the somewhat more diverse and luxurient xeroriparian. Vegetation here is dominated by burrobush, foothill paloverde, creosotebush, organ pipe cactus, ocotillo, and others. Baseline data: Peak number of individuals for each lizard species during each run, as follows:
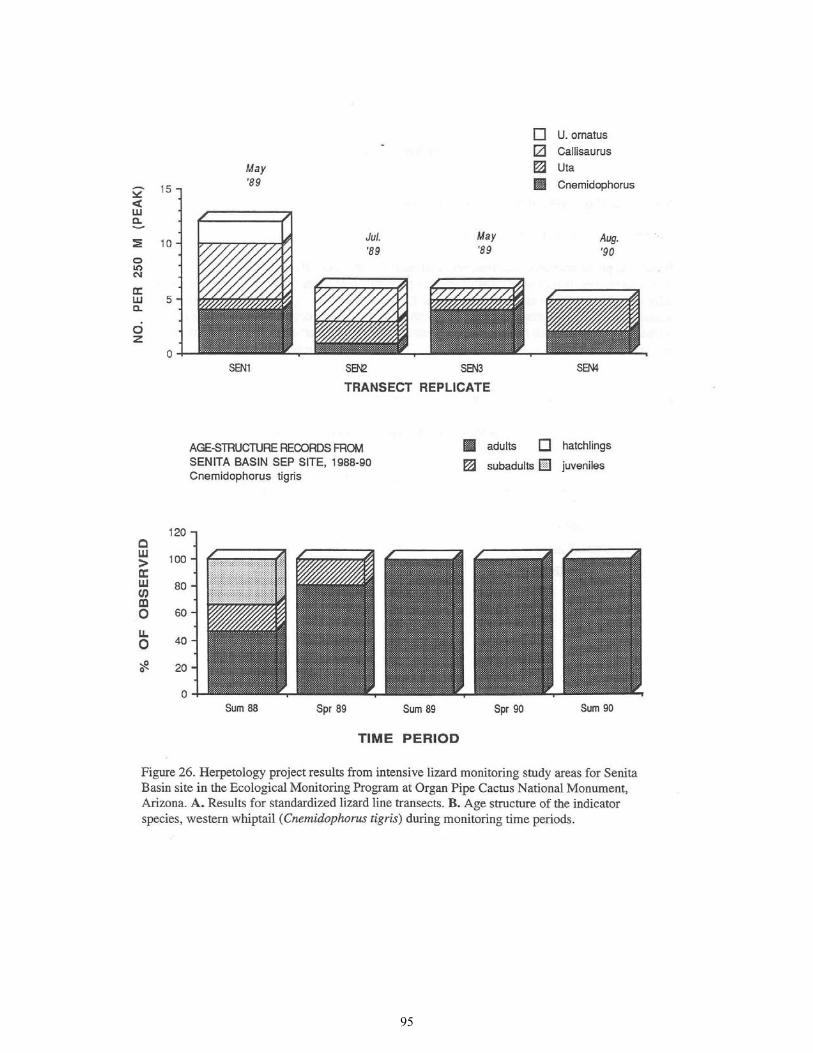
95

96
Vulture Site
Number of standardized lizard line transects: 1 Length of the transect: 200 m (656 ft) Habitat type on transect: The transect is on hard, slightly gravelly soil on the bajada. It runs from east to west from a small wash supporting foothill paloverde, desert-thorn, creosotebush, ashy jatropha (Jatropha cinerea), and others. It progresses about 120 m (394 ft) across gravelly upland dominated by creosotebush and burrobush. It then goes perpendicularly across the main Vulture Site wash, which has a diverse vegetation including desert-ironwood, blue paloverde, burrobush, bitter condalia (Condalia globosa), bursage, honey mesquite, cheese-bush, and many others. Baseline data: Peak number of individuals for each lizard species during each run, as follows:

97
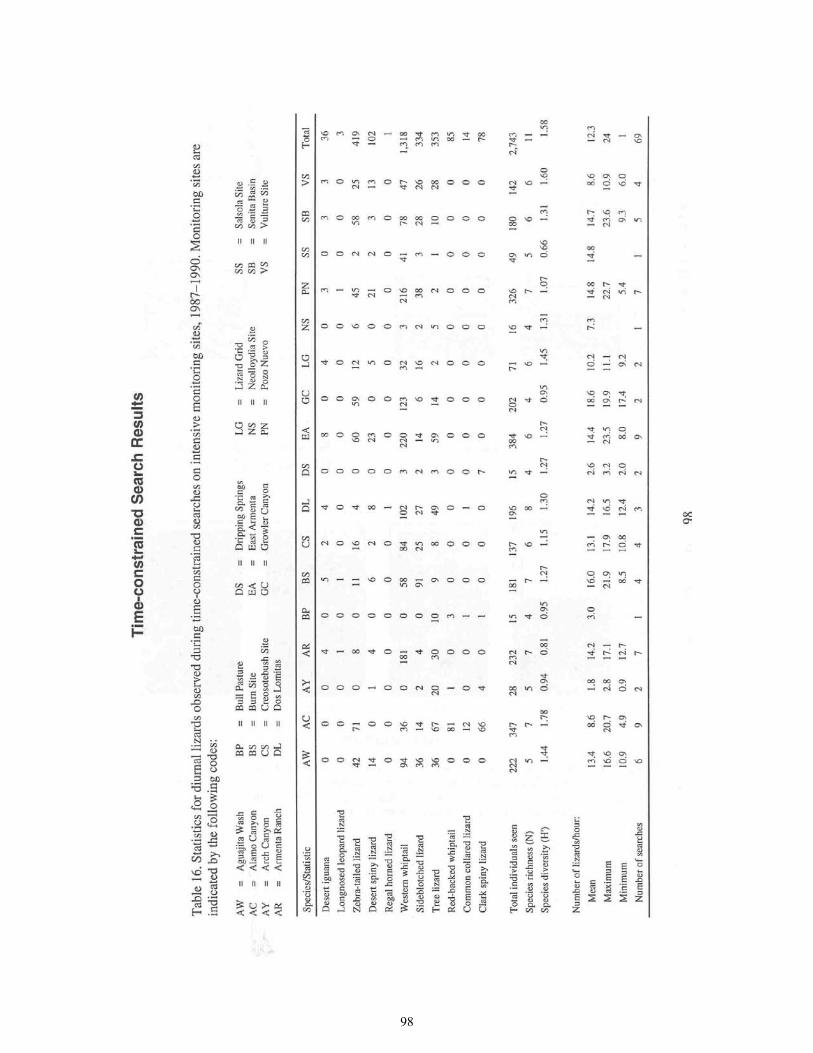
98

99
Recommendations
Toads (Anurans) Overall, the anuran fauna appears to be healthy, and the anurans are probably more abundant in the monument today than prior to the man-made landscape modifications during the last 100 years. The known local-distribution of the Sonoran green toad in areas that are off-monument and subject to heavy grazing pressure calls for distributional study, and may have resource management implications. The recommendations are as follows:
1. Maintain the Sonoran green toad on the ORPI Red List, and document each observed occurrence of it on and within several miles of the monument.
(a) Search for breeding populations of the Sonoran green toad at Cuerda de Lena Wash,
near the north boundary of the monument.
(b) Search for the Sonoran green toad and additional species of anurans in the extreme northeastern corner of ORPI, at Kuakatch Wash.
(c) Determine the full breeding-distribution of the Sonoran green toad in the Valley of
the Ajo, north of the monument.
2. Consider options for incorporation into ORPI of federal lands north of ORPI and within the Valley of the Ajo.
3. Determine the anurans present and breeding sites in the Growler Valley and environs.
4. Continue the policy of maintaining surface-water availability in a human-unaltered state.
5. Support scientific research efforts that may arise in connection with ecological interest in
the ecology and evolution of desert amphibians. Turtles and Tortoises (Chelonians) Our recommendations for conservation and management of the turtles at Quitobaquito include the following:
1. Researchers, if possible the authors, should recensus the turtle population at Quitobaquito to obtain more certain estimates for adult survivorship and current population size.
2. Researchers should use mini-transmitters to track adult females to their nest sites.

100
3. During the regular resource-management monitoring and maintenance work at Quitobaquito, conduct systematic searches within 5 m (16.4 ft) of the ponds, springs, canals, and the terrestrial habitat, to look for dead turtles. All dead turtles should:
(a) be placed immediately on ice in a cooler;
(b) be frozen in heavy-duty Ziplock freezer bags;
(c) be identified with permanent-ink pen or with pencilled labels in the bag.
4. Increase bank-side vegetation along canals. Dense stands of bullrush (Scirpus olneyi) must be immediately grown along all the cement canals connecting the spring and ponds.
5. Eliminate exposed banks. Visitor access to the exposed banks of these canals and to any
other exposed banks from which turtles may be easily collected must be prevented.
6. The Sonoran mud turtle should be retained on the ORPI Red List.
7. The Rio Sonoyta must be surveyed to determine whether viable turtle populations persist there.
Lizards and Snakes (Squamates) Lizards The valley-bottom fill regions of the monument require additional monitoring for lizards. The reasons are as follows:
1. The floodplain (xeroriparian) areas of the valley bottom were refugia from
drought-induced lizard population declines.
2. These areas normally support the greatest lizard densities of any habitat.
3. The valley-bottom fill at ORPI supports most of the Lower Colorado Valley herpetofaunal element of the Sonoran Desert, where it is the finest, and almost only, representation within the national parks system.
4. The upland desertscrub communities-Larrea consociations-surrounding the floodplains
have not yet been appropriately included in the monitoring program. 5. Valley bottoms typically receive heavy, and often destructive use from humans,
throughout the Sonoran Desert and in most other ecosystems.
6. Much of the valley-bottom fill at ORPI, especially on floodplains, has already been severely impacted by man, and is still degrading by erosion.
7. The currently established EMP sites of the valley bottom are primarily in (human-caused)
disturbed environments.

101
Accordingly, the following improvements are suggested:
1. Paired upland-xeroriparian EMP study areas should be established at and in the vicinity of Cuerda de Lena Wash, Growler Wash, near the west monument boundary, and near Paloverde Camp or similar site.
2. A relatively undisturbed site for pair-wise comparison to Armenta Ranch should be
established on a floodplain in the region south of Armenta Ranch.
3. Priority should go to erosion control at Armenta Ranch, which is a floodplain community surrounded by relatively undisturbed habitat, in contrast to the north side floodplain margin of Rio Sonoyta on the monument south boundary. The gabion check dam at Armenta Ranch, which washed out in 1991, should be repaired quickly to forestall headward erosion into the regenerating floodplain community. This gabion structure appears to have successfully halted erosion and produced natural community regeneration during the 1980s. Two or more additional, similar, structures could be placed downstream in the approximate mile reach of deeply incised floodplain.
4. The monument should attempt to restore normal runoff to the Armenta Ranch floodplain
by seeking to remove the upstream dams on the Kuakatch Wash branch that is just north of the monument.
Upland desertscrub communities on the rock slopes are a major habitat at ORPI that has not been adequately monitored for lizards. We therefore further recommend:
5. Additional monitoring site(s) be established on rock slopes to represent the diversity of
upland desertscrub communities present on the monument. Snakes Recommendations for snake species at ORPI are as follows:
1. The severe impacts that appear for 2 species of special importance at ORPI, the rosy boa
and Sonoran shovelnosed snake, indicate that additional research should be directed toward their ecology at ORPI. The focus of such research should be to learn:
(a) the details of their distribution monument-wide, especially for the Sonoran shovelnosed snake, and
(b) their mortality rates and population densities under natural conditions.
2. From the aesthetic perspective, we would recommend closing the highway to through
traffic at ORPI, and seriously reducing speed limits for monument-visitor traffic. Such a proposal would have the unfortunate consequence of exporting highway mortality to adjacent lands that now support healthy snake populations-perhaps including the species most directly impacted by Route 85.

102
3. An alternative to highway closure would be to close the port-of-entry at Lukeville at an early hour each day (1700 during October-April, 1900 during May-September).
4. Another solution, though very costly, would be to install a causeway to replace Route 85
for through travellers or those going directly to the campground. Such a project would clearly require detailed consultation with biologists competent to evaluate the impact of such a structure on organisms other than snakes-particularly large mammals.
5. An alternative solution to (4) would be to construct a drift-fence barrier to snake
movement onto the highway. This is feasible; however, unattended drift fences will have major ecological impacts of their own, including, but not restricted to, disruption of gene flow. Experimental models would have to be tried, over a period of several years, at some appreciable cost to NPS, and in consultation with biologists.
There are several obvious problems with this approach, including overall cost for purchase, installation and maintenance of the fence, and design and installation problems of passageways under the highway. We have observed many snakes at metal culverts that had crawled up embankments onto roadways rather than pass under the highway in the culvert. We are not certain at this juncture what sort of passageway would be required, nor its cost or other ecological impacts.
6. At a minimum, NPS should discourage further opening of the port-of-entry at Lukeville, and
encourage reinstatement of the earlier, more-restricted hours of its operation. Simultaneously, appropriate research should be carried out to determine whether more drastic measures are fully warranted by conservation needs for species protected at ORPI.
7. Paving of unpaved monument-roads should be minimized or avoided.

103
Literature Cited
Axelrod, D.1. 1979. Age and origin of Sonoran Desert vegetation. Occasional Papers of the California Academy of Sciences 132:1-74.
Boundy, J., and A. W. Ford. 1989. Masticophis bilineatus (Sonoran Whipsnake). Geographic
Distribution. Herpetological Review 20:75-76. Brown, D. E., C. H. Lowe, and C. P. Pase. 1979. A digitized classification system for the biotic
communities of North America, with community (series) and association examples for the Southwest. Journal of the Arizona-Nevada Academy of Sciences 14 (Supplement 1):1-16.
Campbell, H. 1953. Observations on snakes DOR in New Mexico. Herpetologica 9:157-160.
Campbell, H. 1956. Snakes found dead on the roads of New Mexico. Copeia 1956:124-125.
Dunham, A. E. 1980. An experimental study of interspecific competition between the iguanid
lizards Sceloporus merriami and Urosaurus ornatus. Ecological Monographs 50:309-330. Fitch, H. S. 1949. Road counts of snakes in western Louisiana. Herpetologica 5:87-90. Fouquette, M. J., Jr. 1970. Bufo alvarius Girard. Colorado River toad. Catalogue of American
Amphibians and Reptiles 93:1-4. Gloyd, H. K. 1937. A herpetological consideration of faunal areas in southern Arizona. Bulletin
of the Chicago Academy of Sciences 5:79-139. Hensley, M. M. 1950. Results of a herpetological reconnaissance in extreme southwestern
Arizona and adjacent Sonora, with a description of new subspecies of the Sonoran whipsnake, Masticophis bilineatus. Transactions of the Kansas Academy of Science 53:270-288.
Holm, P. H. 1988. Two populations of the tree lizard (Urosaurus ornatus) in southern Arizona.
Unpubl. Masters Thesis, The University of Arizona, Tucson. Huey, L. H. 1942. A vertebrate faunal survey of the Organ Pipe Cactus National Monument,
Arizona. Transactions of the San Diego Society of Natural History 9(32):353-376. Hulse, A. E. 1978. Bufo retiformis Sanders and Smith. Sonoran green toad. Catalogue of
American Amphibians and Reptiles 207:1-2. Iverson, J. B. 1976. Kinosternon sonoriense Sonoran mud turtle. Catalogue of American
Amphibians and Reptiles 176:1-3.

104
Iverson, J. B. 1979. A taxonomic reappraisal of the yellow mud turtles, Kinosternon flavescens (Testudines: Kinosternidae). Copeia 1979:212-225.
Iverson, J. B. 1981. Biosystematics of the Kinosternon hirtipes species group (Testudines:
Kinosternidae). Tulane Studies in Zoology and Botany 23:1-74. Iverson, J. B. 1986. A checklist with distribution maps of the turtles of the world. Paust Printing,
Richmond, Indiana. Iverson, J. B. 1989. The Arizona mud turtle, Kinosternon flavescens arizonense (Kinosternidae),
in Arizona and Sonora. Southwestern Naturalist 34:356-368. Klauber, L. M. 1931. A statistical survey of the snakes of the southern border of California.
Bulletin of the Zoological Society of San Diego 8:1-93. Klauber, L. M. 1939. Studies of reptile life in the arid Southwest. Part I. Night collecting on the
desert with ecological statistics. Bulletin of the Zoological Society of San Diego 14:2-64. Lowe, C. H. 1987. The amphibians and reptiles at Quitobaquito, Organ Pipe Cactus National
Monument, Arizona. Report to National Park Service, Cooperative Park Studies Unit, The University of Arizona, Tucson.
Lowe, C. H. 1990 (1984). A 50-year summary for the herpetofauna of Organ Pipe Cactus
National Monument, Arizona (1932-1982). Cooperative Park Studies Unit, The University of Arizona. P. 65-72 in M. M. McCarthy and R. R. Johnson, Organ Pipe Cactus National Monument Biosphere Reserve: Assessment of Scientific Information and Activities. Part I: Current and Required Research.
Lowe, C. H., and P. C. Rosen. 1991. Checklist of the amphibians and reptiles at Organ Pipe
Cactus National Monument. Southwest Parks and Monuments Association, Tucson, Arizona.
Lowe, C. H., and W. R. Supernaugh. 1953. Checklist for the amphibians and reptiles at Organ Pipe Cactus National Monument. Organ Pipe Cactus National Monument, Thermofax.
Pianka, E. R. 1986. Ecology and natural history of desert lizards. Princeton University Press,
Princeton, New Jersey. 216 p. Price, A. H. 1983. Roadriding as a herpetofaunal collecting technique and its impact upon the
herpetofauna of New Mexico. Unpublished report to Endangered Species Program, New Mexican Department of Game and Fish. 27 p.
Rosen, P. C. 1986. Population decline of Sonoran mud turtles at Quitobaquito Springs. Report to
National Park Service, Cooperative Park Studies Unit, The University of Arizona, Tucson. 39 p.

105
Rosen, P. C., and C. H. Lowe. 1995. Section 4 in Organ Pipe Cactus National Monument, Organ Pipe Cactus National Monument Natural Resources Management Ecological Monitoring Program: Monitoring Protocol Manual. Special Report 11, Cooperative Park Studies Unit, The University of Arizona, Tucson.
Sanders, O., and H. M. Smith. 1951. Geographic variation in toads of the debilis group of Bufo.
Field and Lab 19:141-160.
Savage, J. M. 1954. A revision of the Bufo debilis complex. Texas Journal of Science 6:83-112. Shreve, F. 1951. Vegetation and flora of the Sonoran Desert. Vol. I, Vegetation. Carnegie
Institute, Washington, Publication 591:1-192. Smith, D. C. 1981. Competitive interactions of the striped plateau lizard (Sceloporus virgatus)
and the tree lizard (Urosaurus ornatus). Ecology 63:679-687. Smith, P. W., and M. M. Hensley. 1957. The mud turtle Kinosternon flavescens stejnegeri
Hartweg, in the United States. Proceedings of the Biological Society of Washington 70:201-204.
Tinkle, D. W. 1982. Results of experimental density manipulations in an Arizona lizard
community. Ecology 63:57-65. Van Devender, T. R. 1987. Holocene vegetation and climate of the Puerto Blanco Mountains,
southwestern Arizona. Quaternary Research 27:51-72. Van Devender, T. R., A. M. Rea, and W. E. Hall. 1991. Faunal analysis of late quaternary
vertebrates from Organ Pipe Cactus National Monument. Southwestern Naturalist 36:94-106.
Vitt, J. 1991. Desert reptile communities. P. 249-277 in G. A. Polis (ed.), The Ecology of Desert
Communities. The University of Arizona Press, Tucson. Warren, P. L., B. K. Mortenson, B. D. Treadwell, J. E. Bowers, and K. L. Reichardt. 1981.
Vegetation of Organ Pipe Cactus National Monument. Technical Report 8:1-79. Cooperative Park Studies Unit, The University of Arizona, Tucson.
Wasserman, A. O. 1970. Scaphiopus couchii Baird. Couch's spadefoot toad. Catalogue of
American Amphibians and Reptiles 85:1-4. Williams, K. L., and P. S. Chrapliwy. 1958. Selected records of amphibians and reptiles from
Arizona. Transactions of the Kansas Academy of Science 61:299-301.

106
Appendix 1 Summary of Species Occurrence and Frequency on
Ecological Monitoring Program Sites and Associated Habitats in Organ Pipe Cactus National Monument, Arizona
Introduction This appendix comprises a series of 22 tables. Table A1-1 provides a numerical summary of all reptiles and amphibians recorded on Ecological Monitoring Program sites and associated habitats at and in the region of Organ Pipe Cactus National Monument, Arizona (ORPI). Tables Al-2-Al-22 list, for each EMP site or associated habitat, the number of individuals of each species recorded. Subtotals by taxonomic order and a grand total for each site/area are also provided. Data for table A1-1 include animals observed off-site during the course of the 1987-1991 work, including 3 species-the lesser earless lizard (Holbrookia maculata), ground snake (Sonora semiannulata), and yellow mud turtle (Kinosternon flavescens)-not currently known from the monument. The data in this appendix are based on the dataset assembled during the ORPI EMP Herpetology Project, 18 August 1987 through 1 January 1992, and were collected by University of Arizona herpetologists, with few exceptions. All of the data are direct observations based on standard field techniques. These include moving variously through the habitat observing active animals, turning loose rocks and other debris, and driving on established roads within the sites. Additional records obtained from knowledgeable scientists, National Park Service (NPS) rangers, and by the authors earlier in the 1980s are also included. The full dataset is housed at the University of Arizona Herpetology Laboratory. For the tortoise, sign records (skeletal material, scat, tracks, and so forth) are included as site records. The only trap data included are at Snake Study Area, and for turtles at Quitobaquito. This listing is a baseline dataset that should be added to only by knowledgeable persons fully conversant with sight-identification of Sonoran Desert reptiles in general, and ORPI reptiles in particular. It is not, of course, an exhaustive herpetofaunal listing for the sites covered. Several of the listed species have, in the past, been misidentified from these and other sites at ORPI. We cannot caution too strongly against adding to this list without full verification, preferably as photo vouchers and always with full accompanying data (date, time, size, habitat, activity, and observer). The most frequently and easily misidentified and misreported species have been (1) Mohave rattlesnake (Crotalus scutulatus), mistaken for western diamondback rattlesnake (C. atrox); (2) tiger rattlesnake (Crotalus tigris), mistaken for speckled rattlesnake (C. mitchelli); (3) western whiptail (Cnemidophorus tigris), mistaken for canyon spotted whiptail (C. burti); (4) banded sand snake (Chilomeniscus cinctus), mistaken for Sonoran shovelnosed snake (Chionactis palarostris); (5) regal horned lizard (Phrysonoma solare), mistaken for desert horned lizard (P. platyrhinos); (6) longtailed brush lizard (Urosaurus graciosus), mistaken for tree lizard (U. ornatus); and (7) desert spiny lizard (Sceloporus magister), mistaken for Clark spiny lizard (S. clarki). Additionally, the 2 shovelnosed snake species, Sonoran shovelnosed

107
snake (Chionactis palarostris) and western shovelnosed snake (C. occipitalis), can be easily confused.

108

109

110

111

112

113

114

115

116

117

118

119

120

121

122

123

124

125

126

127

128

129

130

131

132
Appendix 2 Checklist of Amphibians and Reptiles at
Organ Pipe Cactus National Monument, Arizona
Introduction There are 48 known amphibian and reptile species in the herpetofauna of ORPI. The herpetofauna includes toads (5 species), turtles (2 species), lizards (16 species), and snakes (25 species) living within the total monument area of 517 mi2 (1,339 km2). In the first field reconnaissance for amphibians and reptiles at ORPI, Gloyd (1937) recorded 13 species. The first all-vertebrates field survey at ORPI was conducted in 1939 by Lawrence Huey, and assistants Philip Richty and Charles Harbeson (Huey 1942); that survey recorded 4 amphibian and 21 reptilian species, although the work focused primarily on avifauna and mammal fauna. The first checklist for amphibians and reptiles at ORPI (Lowe and Supernaugh 1953) reported 45 taxa (4 amphibians and 41 reptiles). The presently known total reported in the monument (as of December 30, 1991) is 48. A recent checklist (Lowe and Rosen 1991) reports a total of 47 species (4 amphibians and 43 reptiles). The final checklist presented here adds 1 amphibian species (for 48 total species), and includes a few other minor changes. A brief review of the first 50 years of herpetology at ORPI (1932-1982) is given in Lowe (1990). We were unable to locate populations of the ground snake (Sonora semiannulata) at ORPI. Although a single specimen was earlier reported from ORPI by W. R. Supernaugh, we have been unable to verify or locate the specimen or to locate references to it in the ORPI resource collection records. We were also unable to locate the desert night lizard (Xantusia vigilis), which also may occur in the region. The relatively high richness of herpetological species at ORPI includes a diverse array of specialized life-forms. Such richness and diversity in the life-styles of the herpetofauna is related directly to the high diversity of the desert landscapes and habitats in which they live.

133
Checklist of Amphibians and Reptiles Voucher specimens document the species occurrence at the monument. The 8 dangerously venomous species are indicated with an asterisk (*). Amphibians (Class Amphibia)
Frogs and Toads (Order Anura)
Spadefoot toads (Family Pelobatidae)
Desert spadefoot (Scaphiopus couchi)
Toads (Family Bufonidae)
Sonoran Desert toad (Bufo alvarius) Great Plains toad (Bufo cognatus) Red-spotted toad (Bufo punctatus) Sonoran green toad (Bufo retiformis)
Reptiles (Class Reptilia)
Turtles (Order Testudines)
Tortoises (Family Testudinidae)
Desert tortoise (Gopherus agassizi)
Mud and Musk turtles (Family Kinostemidae)
Sonoran mud turtle (Kinosternon sonoriense)
Lizards and Snakes (Order Squamata)
Lizards (Suborder Sauria)
Beaded lizards (Family Helodermatidae)
* Gila monster (Heloderma suspectum)
Geckos (Family Gekkonidae)
Western banded gecko (Coleonyx variegatus)
Whiptails and allies (Family Teiidae)
Canyon spotted whiptail (Cnemidophorus burti) Western whiptail (Cnemidophorus tigris)
Iguanids (Family Iguanidae)
Zebra-tailed lizard (Callisaurus draconoides) Common collared lizard (Crotaphytus collaris) Desert iguana (Dipsosaurus dorsalis)

134
Checklist of Amphibians and Reptiles-continued.
Iguanids (continued)
Longnosed leopard lizard (Gambelia wislizeni) Desert homed lizard (Phrysonoma platyrhinos) Regal horned lizard (Phrysonoma solare)
Common chuckwalla (Sauromalus obesus) Clark spiny lizard (Sceloporus clarki) Desert spiny lizard (Sceloporus magister)
Longtailed brush lizard (Urosaurus graciosus) Tree lizard (Urosaurus ornatus) Sideblotched lizard (Uta stansburiana)
Snakes (Suborder Serpentes)
Blind snakes (Family Leptotyphlopidae)
Western blind snake (Leptotyphlops humilis)
Boas (Family: Boidae)
Rosy boa (Lichanura trivirgata)
Colubrids (Family Colubridae)
Banded sand snake (Chilomeniscus cinctus) Western shovelnosed snake (Chionactis occipitalis) Sonoran shovelnosed snake (Chionactis palarostris) Southwestern blackheaded snake (Tantilla hobartsmithi) Saddled leafnosed snake (Phyllorhynchus browni) Spotted leafnosed snake (Phyllorhynchus decurtatus) Sonoran whipsnake (Masticophis bilineatus) Coachwhip (Masticophis flagellum) Western patchnosed snake (Salvadora hexalepis) Glossy snake (Arizona elegans) Common kingsnake (Lampropeltis getula) Bullsnake (Gopher snake) (Pituophis melanoleucus) Longnosed snake (Rhinocheilus lecontei) Lyre snake (Trimorphodon biscutatus) Night snake (Hypsiglena torquata) Blacknecked garter snake (Thamnophis cyrtopsis)
Coralsnakes (Family Elapidae)
* Western coralsnake (Micruroides euryxanthus)

135
Checklist of Amphibians and Reptiles-continued.
Vipers (Family Viperidae)
* Western diamondback rattlesnake (Crotalus atrox) * Sidewinder (Crotalus cerastes) * Speckled rattlesnake (Crotalus mitchelli) * Blacktailed rattlesnake (Crotalus molossus) * Mohave rattlesnake (Crotalus scutulatus) * Tiger rattlesnake (Crotalus tigris)
Species Reported from the Monument that Lack Verified On-site Populations Potential Species on the Monument We regard the following 3 taxa to be of potential occurrence on the monument. They are listed in order of decreasing probability of occurrence at ORPI.
Burrowing treefrog (Pternohyla fodiens) Plains narrow-mouthed toad (Gastrophryne olivacea) Ground snake (Sonora semiannulata) Desert night lizard (Xantusia vigilis) Madrean alligator lizard (Gerrhonotus Elgaria kingi)
Other Reported Species Some species have been variously observed and/or reported from ORPI. These are treated below under native species and nonnative species. While it is not inconceivable that 1 or more of these species may have a breeding population represented on the monument, it seems now virtually certain that they do not.
Native Species of Sonoran Desert Region
Southern spadefoot (Scaphiopus multiplicatus) Yellow mud turtle (Kinosternon flavescens)
Nonnative Species
Tiger salamander (Ambystoma tigrinum) Bullfrog (Rana catesbieana) Painted turtle (Chrysemys picta) Slider turtle (Pseudemys scripta)

136

137

138





























