Embed Size (px)
Citation preview

American Fisheries Society Symposium 39:67–79, 2004© Copyright by the American Fisheries Society 2004
67
Ecological Segregation of Native and AlienLarval Fish Assemblages in the Southern
Sacramento–San Joaquin Delta
FREDERICK FEYRER*
Aquatic Ecology Section, California Department of Water Resources3251 S Street, Sacramento, California 95816-7017, USA
Abstract.—Fish larvae were sampled at multiple fixed sites from late winter to earlysummer over 6 years (1990–1995) in the southern Sacramento–San Joaquin Delta. Atotal of 394,797 fish larvae representing 15 species or taxonomic groups was collected.The assemblage was numerically dominated by three species that represented 98% ofthe total catch: alien shimofuri goby Tridentiger bifasciatus (71%), threadfin shad Dorosomapetenense (15%), and native prickly sculpin Cottus asper (12%). The abundance of nativeand alien species differentially clustered along environmental gradients of water tem-perature and river flow. Each native species (prickly sculpin, splittail Pogonichthysmacrolepidotus, delta smelt Hypomesus transpacificus, longfin smelt Spirinchus thaleichthys,and Sacramento sucker Catostomus occidentalis) and one alien species (bigscale logperchPercina macrolepida) were associated with the early season conditions of cool watertemperature and high river flow. Alien species (especially shimofuri goby, threadfinshad, and ictalurid catfishes) were associated with late season conditions of relativelywarm water temperature and low river flow. Accordingly, native species dominated theassemblage February–March, while alien species dominated May–July. However, peakseasonal abundance of alien species was typically five times greater than that of nativespecies. Seasonal succession of assemblage structure was persistent among years andwas highly correlated with water temperature, a likely result of the differential spawn-ing requirements of adult fishes. Interannually, the assemblage remained consistentover the study period despite considerable variability in delta inflow. I hypothesize,given the consistent temporal segregation between native and alien larval fish assem-blages, that direct interactions such as competition between the two groups may not bea major factor influencing poor native fish recruitment in the south delta.
Introduction
Larval fish assemblages often exhibit tempo-ral stability (i.e., consistent phenological ap-pearances of species) typically because of dif-fering environmental requirements for thespawning of adult fishes or due to environ-mental conditions that influence recruitmentof larvae in certain geographic locations(Powles et al. 1984; Yoklavich et al. 1992; Kelleret al. 1999; Witting et al. 1999; Lazzari 2001).Such chronology in appearance of larval fishes
can influence the partitioning of resources andinteractions among species. Given the threatsassociated with alien species and the desire tomaintain native fish assemblages, understand-ing the ecological interactions between nativeand alien fishes is a rapidly growing area ofinterest (Moyle 1986; Ross 1991). Within theSan Francisco Estuary, recent studies in anupstream tributary (Marchetti and Moyle 2000)and a downstream marsh (Meng and Matern2001) have demonstrated a phenological sepa-ration between assemblages of native andalien larval fishes; native larvae typically ap-pear earlier in the year than aliens. The princi-* [email protected]

68 FEYRER
pal factors believed to be driving these obser-vations were the influence of water tempera-ture and river flow on timing of adult fishspawning (Marchetti and Moyle 2000; Mengand Matern 2001).
Poor recruitment of native fishes duringearly life stages has been identified as one po-tential mechanism limiting their abundance inthe Sacramento–San Joaquin Delta (Bennettand Moyle 1996). Recruitment success is de-pendent upon survival rates, which can be in-fluenced by biotic and abiotic factors (Houde1987; Harvey 1991; Letcher et al. 1996). If theseasonal succession of native and alien fishlarvae in the south delta is similarly segregated,as has been observed in other regions of thesystem, then potential for direct interactions,such as competition between the two groups,may be limited in time and thus may notstrongly influence recruitment of native fishes.
In this paper, I present results of a 6-yearichthyoplankton study conducted in thesouthern delta. The study was initiated to de-scribe larval fish assemblages in the southdelta and to evaluate impacts of large-scalewater diversions by the State Water Project(SWP) and Central Valley Project (CVP), stateand federal facilities that divert water from thesystem for agricultural and municipal con-sumption. The present analysis focused ondescribing larval fish assemblages of the southdelta with an emphasis on the ecological as-sociations of native and alien species. My goalswere to evaluate the status of native fish re-production in the south delta, as determinedby the presence of larvae, and to evaluate theextent of spatial and temporal co-occurrencebetween native and alien species. I addressedthe following specific questions: (1) what spe-cies comprise the larval fish assemblage in thesouth delta? (2) are there associations amongspecies and environmental variables? and (3)are there persistent temporal and spatial pat-terns of assemblage structure?
Study Area
The Sacramento–San Joaquin Delta (Figure 1),located upstream of the confluence of the Sac-ramento and San Joaquin rivers, consists ofmore than 1,000 km of waterways with adrainage area encompassing approximately
40% of California’s surface area (Nichols et al.1986). During peak diversions, up to 65% ofthe delta’s natural discharge is diverted foragricultural and municipal consumption(Nichols et al. 1986). The majority of this water(up to 73 × 108 m3/year) is diverted at largepumping facilities in the south delta by the SWPand CVP. The south delta is arguably the mostdegraded region of the system due to alteredphysical habitat, hydrodynamics, and waterquality associated with the water project facili-ties and flood control projects (Arthur et al.1996). Similar to other locations with highlyaltered habitats (Ross 1991), the adult fishfauna of the south delta is numerically domi-nated by alien species (Feyrer and Healey 2003).
Fish larvae were sampled at seven sites inthe south delta: four sites in Old River and onesite each in Grant Line Canal, Salmon Slough,and Victoria Canal (Figure 1). These waterways,referred to locally as sloughs, are approximately1–5 m in depth, tidally influenced, and are con-stricted within flood control levees. Rock-rein-forced banks (riprap) dominate riparian habi-tats, and alien submerged aquatic vegetation,primarily Brazilian waterweed Egeria densa, isprevalent in the shallow littoral zone; only rem-nants of natural wetland riparian habitats ex-ist. The primary source of freshwater for thesouth delta is the San Joaquin River. Agricul-ture is the dominant land use activity beyondchannel levees, and hundreds of small localagricultural diversion facilities are scatteredthroughout the region.
Methods
Data collection
Fish larvae were sampled during daylighthours approximately 3 d per week from Aprilto July 1990 and 5 d per week from Februaryto July 1991–1995. A 10-min oblique tow witha 505-µm-mesh net mounted on a towing sledwas conducted at each site. The net measured1.5 m in length and had a 0.4-m mouth diam-eter. A 1,000-mL plastic collecting jar, screenedwith 470-µm-mesh bolting cloth, was attachedto the net to collect samples. A flowmeter wasattached to the net frame to measure the vol-ume of water filtered during a tow. After eachtow, samples were rinsed into the collecting

ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 69
jar, preserved in 5% formalin, and stained withrose-bengal dye. In the laboratory, sampleswere rinsed with water through a 300-mm-mesh sieve and sorted under a magnifying il-luminator, and all fish larvae were identifiedto species or species groupings (Table 1) fol-lowing the keys in Wang (1986).
Three environmental variables were mea-sured at the time of sampling: specific con-
ductance (µS) was estimated using a portableconductivity meter, water temperature (°C) wasmeasured with a mercury thermometer, andwater transparency (cm) was measured witha Secchi disk. Estimates of tidally averageddaily flow (m3/s) at each site were obtainedfrom the California Department of Water Re-sources’ CALSIM Hydrology Model. Raw datafrom this study are available online from the
FIGURE 1. Larval fish sampling sites in the southern Sacramento–San Joaquin Delta.

70 FEYRER
Interagency Ecological Program website athttp://iep.water.ca.gov.
Data analysis
I used canonical correspondence analysis(CCA) to examine the association of larval fishdensities (log10 [x/m3 + 1]) with environmentalvariables (Legendre and Legendre 2000). CCAis a multivariate direct ordination techniquethat extracts synthetic environmental gradientsthat maximize niche separation within speciesassemblages, thereby facilitating the interpre-tation of how species abundances relate to en-vironmental variables. CCA was conductedwith CANOCO software program (ter Braakand Smilauer 1998). I excluded two species,American shad and yellowfin goby, from thisanalysis because they occurred in less than 1%of the total samples (N = 1,436 samples withfish larvae). The final model was constrainedto only include environmental variables sig-nificant at P < 0.05, as estimated by the forwardselection procedure with Monte Carlo simula-tions (199 permutations) provided byCANOCO (ter Braak and Smilauer 1998).
I used detrended correspondence analy-sis (DCA) to investigate spatial and temporalvariation in assemblage structure, with thesame species matrix used in the CCA. DCA isan indirect eigenvector ordination technique
based upon reciprocal averaging that correctsfor the “arch effect” observed in reciprocal av-eraging (Gauch 1982; Legendre and Le-gendre2000). Inferred primary gradients within as-semblages are effectively displayed by DCA(Gauch 1982). Sample and species scores weregenerated with CANOCO software program(ter Braak and Smilauer 1998). DCA samplescores were subjected to an analysis of vari-ance (ANOVA) procedure with site, month,year, and their interactions as factors. Thisprocedure was used to partition the amountof variance accounted for by each of these vari-ables, not to test for significant differences be-cause replicate samples were not obtained ateach site. Factor variance components werederived from expected mean squares based ona fixed effects ANOVA. This procedure al-lowed me to evaluate whether the spatial (site)or temporal (month and year) effects on as-semblage structure were important alone or ifthey interacted. DCA sample scores were alsosorted in numerous ways, and resulting plotswere visually inspected for patterns. Detrend-ing by segments was incorporated, and there-fore only scores along axis I of the plots wereinterpreted as ecologically meaningful (Le-gendre and Legendre 2000). I used Pearsonproduct-moment correlations to examine therelationship between DCA axis I sample scoresand environmental variables.
TABLE 1. Species (asterisk indicates native status) or taxonomic groupings, code, total number, per-cent number (if greater than 1%), and percent occurrence in samples of larval fishes captured in thesouth delta.
Taxa Code Number (%) % samples
Shimofuri goby Tridentiger bifasciatus SG 280,461 (71) 65Threadfin shad Dorosoma petenense TS 60,994 (15) 54Prickly sculpin Cottus asper* PS 46,926 (12) 53Striped bass Morone saxatilis SB 3,153 (1) 36Centrarchidae CNT 979 18Cyprinidae (other than splittail) CYP 703 16Bigscale logperch Percina macrolepida BL 568 17Ictaluridae ICT 320 5Inland silverside Menidia beryllina IS 260 7Splittail Pogonichthys macrolepidotus* ST 201 5Delta smelt Hypomesus transpacificus* DS 74 4American shad Alosa sapidissima AS 65 <1Sacramento sucker Catostomus occidentalis* SS 47 2Longfin smelt Spirinchus thaleichthys* LS 31 2Yellowfin goby Acanthogobius flavimanus YG 15 <1

ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 71
Results
Environmental conditions
Among the environmental variables, riverflow varied most among sites (Figure 2). Low-est flows occurred at the southernmost OldRiver site, Middle River, and Salmon Slough,while highest flows occurred at Grant LineCanal and the other Old River sites. Season-ally, river flows were highest February–Apriland lowest May–July. Only the southernmostOld River site exhibited almost no seasonalvariation in river flow. Specific conductancewas seasonally consistent, but slightly higherat the southernmost Old River, Grant Line Ca-nal, and Salmon Slough sites (800–1,000 mS)
than at Victoria Canal and the other Old Riversites (400–600mS). Water temperature in-creased with month and was very consistentamong sites, ranging from approximately 12°Cin February to 22°C in July. Secchi depth con-sistently ranged between 40 and 80 cm acrossmonths and sites.
Catch summary
Over the study period, 394,797 fish larvae rep-resenting 15 species or taxonomic groups werecollected (Table 1). The assemblage was nu-merically dominated by three species that rep-resented 98% of the total catch: alien shimofurigoby (71%), threadfin shad (15%), and nativeprickly sculpin (12%). Alien striped bass was
FIGURE 2. Mean monthly values (±SE) for environmental variables at each site in the south delta, 1990–1995. Old River sites are are numbered from north to south.
Old River 1 Old River 2 Old River 3 Old River 4
Grant Line Canal Grant Line Canal Salmon Slough
0
500
1000
1500
2000
2500
3000
Feb Mar Apr May Jun Jul
Month
Flo
w (
m3/s
)
0
200
400
600
800
1000
1200
Feb Mar Apr May Jun Jul
Month
Sp
ec.
co
nd
. (µ
S)
0
10
20
30
40
50
60
70
80
Feb Mar Apr May Jun Jul
Month
Secch
i d
ep
th (
cm
)
0
5
10
15
20
25
30
Feb Mar Apr May Jun Jul
Month
Tem
pera
ture
(°C
)

72 FEYRER
the only other species that represented at least1% of the total catch. Seasonally, native spe-cies dominated the assemblage early in theyear (February–April) while alien speciesdominated late in the year (May–July; Figure3). However, peak seasonal abundance of na-tive species was typically five times lower thanthat of alien species.
Associations among species andenvironmental variables
The forward selection procedure retained allfour environmental variables in the final CCA
model. The first two CCA axes explained 20%of the variation in species abundance (19%and 1%, respectively; Table 2). The influenceof all four environmental variables on speciesabundances was depicted in a CCA ordina-tion diagram (Figure 4). Native and alien spe-cies differentially clustered along environmen-tal gradients of water temperature and riverflow. Each native species (prickly sculpin,splittail, delta smelt, longfin smelt, and Sacra-mento sucker) and one alien species (bigscalelogperch) were associated with the early sea-son conditions of cool water temperature andhigh river flow. Alien species (especially
FIGURE 3. Mean monthly densities (±SE) of native and alien fish larvae in the south delta, 1990–1995.
TABLE 2. Results of a canonical correspondence analysis relating larval fish densitities to environmen-tal variables in the south delta. Total inertia = 3.07.
Axis 1 Axis 2
Eigenvalue 0.594 0.033Species-environment correlation 0.838 0.308Cumulative percentage of variance explained
Species 19.4 20.4Species-environment relation 91.1 96.2
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Feb Mar Apr May Jun Jul
Mea
n n
o.
larv
ae/
m3
Natives
Aliens

ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 73
shimofuri goby, threadfin shad, and ictaluridcatfishes) were associated with late seasonconditions of relatively warm water tempera-ture and low river flow.
Spatial and temporal variation inassemblage structure
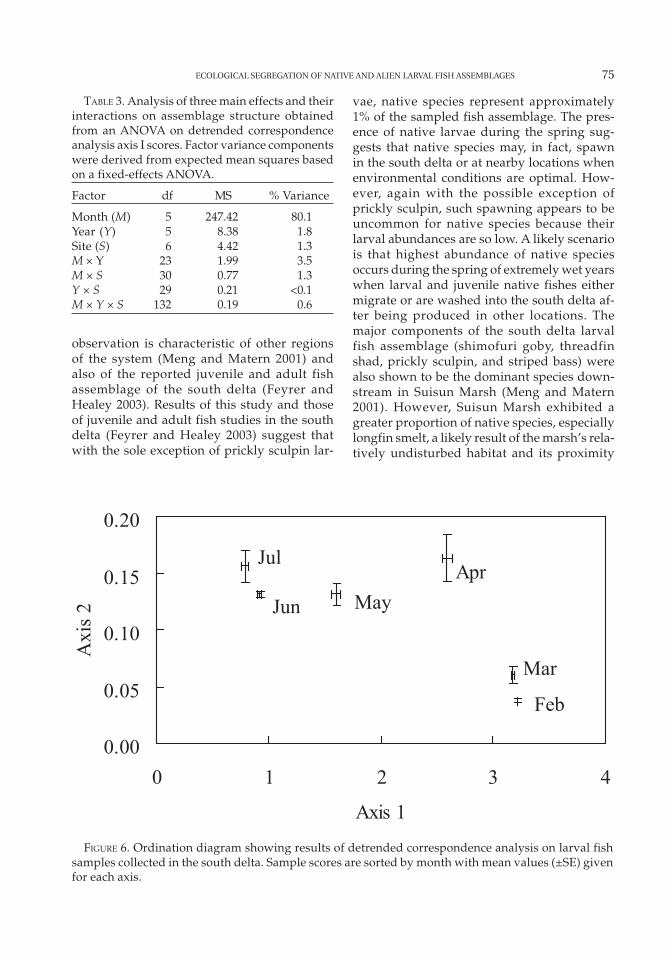
The first DCA axis explained 27.8% of the vari-ance (eigenvalue = 0.85) in the larval fish as-semblage matrix (Figure 5). Results of theANOVA variance partitioning technique sug-gested that month accounted for the greatestproportion of variance (80%) in assemblagestructure (Table 3). None of the other combi-nations of variables accounted for greater than3.5% of the variance. This result was clearlydemonstrated when DCA sample scores weresorted by month in an ordination diagram (Fig-ure 6). Sample scores clustered tightly bymonth with a clear phenological progressionfrom February to July. DCA axis I scores were
highly correlated with water temperature (r =0.82; P = 0.002), suggesting it was the most im-portant environmental variable structuring thelarval fish assemblage (river flow, r = –0.73, P =0.37; Secchi depth, r = –0.36, P = 0.022; specificconductance, r = 0.11, P = 0.017). This resultwas consistent with the CCA, which indicatedthat species abundances clustered primarilyalong a water temperature gradient. Nativespecies were associated with samples takenduring early months (high axis I scores), whilethe alien species were associated with samplestaken during later months (low axis I scores;Figure 5). These results also were consistentwith the CCA, which demonstrated an ecologi-cal segregation between native and alien spe-cies based primarily on water temperature.
Discussion
The south delta larval fish assemblage is nu-merically dominated by alien species. This
Axis 1
Axis
2
-1
0
1
2
3
4
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5 3.0
SC
TEMP
SECCHI
FLOW
TS CYPSB
BL
CNT
IS
LS
PS
DS
SS
ST
ICT SG
FIGURE 4. Ordination diagram showing results of canonical correspondence analysis depicting howlarval fish abundances related to environmental variables in the south delta. Species codes are pre-sented in Table 1. Environmental variables are given in bold: TEMP = water temperature, SC = specificconductance, FLOW = river flow, SECCHI = Secchi depth. Native species are enclosed by the polygon.

74 FEYRERA
xis
2
-0.4
0.0
0.4
0.8
1.2
1.6
2.0
2.4
Axis 1
Ax
is 2
TS
ICT
SG CNTCYP
IS
SB
SS*
BL
ST*
DS*PS* LS*
-0.4
0.0
0.4
0.8
1.2
1.6
2.0
2.4
-0.5 0.5 1.5 2.5 3.5 4.5
FIGURE 5. Ordination diagrams showing sample scores (top) and species scores (bottom) of a detrendedcorrespondence analysis on larval fish samples collected in the south delta. Axis I is scaled the same oneach chart and native species are indicated by an asterisk.
ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 75
observation is characteristic of other regionsof the system (Meng and Matern 2001) andalso of the reported juvenile and adult fishassemblage of the south delta (Feyrer andHealey 2003). Results of this study and thoseof juvenile and adult fish studies in the southdelta (Feyrer and Healey 2003) suggest thatwith the sole exception of prickly sculpin lar-
vae, native species represent approximately1% of the sampled fish assemblage. The pres-ence of native larvae during the spring sug-gests that native species may, in fact, spawnin the south delta or at nearby locations whenenvironmental conditions are optimal. How-ever, again with the possible exception ofprickly sculpin, such spawning appears to beuncommon for native species because theirlarval abundances are so low. A likely scenariois that highest abundance of native speciesoccurs during the spring of extremely wet yearswhen larval and juvenile native fishes eithermigrate or are washed into the south delta af-ter being produced in other locations. Themajor components of the south delta larvalfish assemblage (shimofuri goby, threadfinshad, prickly sculpin, and striped bass) werealso shown to be the dominant species down-stream in Suisun Marsh (Meng and Matern2001). However, Suisun Marsh exhibited agreater proportion of native species, especiallylongfin smelt, a likely result of the marsh’s rela-tively undisturbed habitat and its proximity
TABLE 3. Analysis of three main effects and theirinteractions on assemblage structure obtainedfrom an ANOVA on detrended correspondenceanalysis axis I scores. Factor variance componentswere derived from expected mean squares basedon a fixed-effects ANOVA.
Factor df MS % Variance
Month (M) 5 247.42 80.1Year (Y) 5 8.38 1.8Site (S) 6 4.42 1.3M × Y 23 1.99 3.5M × S 30 0.77 1.3Y × S 29 0.21 <0.1M × Y × S 132 0.19 0.6
0.00
0.05
0.10
0.15
0.20
0 1 2 3 4
Axis 1
Axis
2
Feb
Jul
Jun May
Apr
Mar
FIGURE 6. Ordination diagram showing results of detrended correspondence analysis on larval fishsamples collected in the south delta. Sample scores are sorted by month with mean values (±SE) givenfor each axis.

76 FEYRER
to brackish water (Meng and Matern 2001).Gadomski and Barfoot (1998) documented asimilar regional separation of native and alienlarval fish assemblages in relatively undis-turbed and disturbed habitats in the Colum-bia River basin. Together, these observationssupport the hypothesis that alien species mostcommonly become established in highly al-tered habitats (Ross 1991).
There was a striking temporal separationbetween the occurrence of native and alien fishlarvae in the south delta. Similar to what hasbeen reported in other regions of the system(Marchetti and Moyle 2001; Meng and Matern2001; Grimaldo et al. 2004; Sommer et al. 2004;both this volume), abundance of native spe-cies peaked early in the season, while abun-dance of alien species peaked later. Results ofboth multivariate methods were consistentwith this observation. CCA showed that na-tive species were associated with cool watertemperatures and high river flows, while DCAshowed that native species were associatedwith samples taken early in the year, the timeperiod when such environmental conditionsexist. The same methods showed an oppositepattern for the majority of alien species. Thetemporal segregation of native and alien fishreproductive periods that exists in the Sacra-mento–San Joaquin Delta system is similar tothat observed in the San Juan River, NewMexico and Utah (Gido et al. 1997)
Larval fish assemblages in the south deltaexhibited substantial monthly variationwithin years, but little variation was exhib-ited spatially among sampling sites. Similar-ity of assemblages among sites suggests thateither the available spawning habitat wasequally favorable among sites or they wereclose enough together in space to allow tidalexcursion flows to distribute larvae equallyamong sites. This observation is consistentwith results of Meng and Matern (2001) whofound similar assemblages among most siteswithin Suisun Marsh. Seasonal variation inassemblage structure was persistent amongyears and was primarily associated with theseasonal occurrence of native and alien spe-cies. Water temperature and river flow werethe most important environmental variablesinfluencing this pattern. Low water tempera-tures and high river flows early in the season
provide spawning cues for native adult fishes,while high water temperatures and low riverflows late in the season favor alien species(Moyle 2002). Of the two variables, water tem-perature was the most important explainingfish catches. It exhibited the highest impor-tance on the first CCA axis and the strongestcorrelation with the DCA axis I scores. Addi-tionally, temporal patterns of assemblagestructure were persistent over the study pe-riod despite the fact that total delta inflowamong years differed by as much as 600%, orput in a different context, nearly five standarddeviations of the 40-year running average from1955 to 1995 (not shown; flow data are avail-able online from the California Department ofWater Resources’ Dayflow Database at http://iep.water.ca.gov/dayflow). This suggeststhat the majority of spawning by both fishgroups will take place only when optimal tem-peratures are achieved regardless of flow con-ditions. However, the overall importance ofriver flow, especially total delta inflow, shouldnot be underemphasized because it is knownto influence the availability of spawning andrearing habitat and is positively correlatedwith the abundance of numerous native andcommercially important species (Jassby et al.1995; Sommer et al. 1997; Kimmerer 2002). Itseems likely that there is a certain interactionbetween temperature, river flow, and otherassociated variables that produce optimal con-ditions that drive recruitment of native fishesin the delta.
Interactions with alien species are mecha-nism that may influence the abundance of na-tive species in the delta (Bennett and Moyle1996). Given the consistent temporal segrega-tion between the native and alien larval fishassemblages, the opportunity for direct inter-actions, such as competition, between the twogroups is probably relatively small and maynot significantly influence native fish recruit-ment. Other factors that may be important in-clude limited suitable habitat for spawningor rearing caused by anthropogenic distur-bances, altered flow regimes or hydrodynam-ics caused by water exports and flood controlfacilities, limited food supply caused by foodweb changes, and predation by other lifestages of fishes (Kimmerer et al. 1994; Bennettand Moyle 1996). Other studies have found

ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 77
that predation or competition by alien specieson natives can be significant (Lemly 1985;Meffe 1985; Scoppettone 1993; Marsh andDouglas 1997). A combination of these factorsare probably responsible for the poor recruit-ment of native fishes in the south delta.
The results of this study provide usefuland challenging information for resourcemanagers and water project operations in thedelta. Because the native larvae are consis-tently present early in the year and their abun-dance is tightly correlated with water tempera-ture, water project exports can be scheduledaround this time period to minimize directimpacts such as entrainment. The environ-mental associations among native species andenvironmental variables described here areconsistent with results of other studies onnative fishes (Marchetti and Moyle 2000, 2001;Meng and Matern 2001; Feyrer and Healey2003; Grimaldo et al. 2004). These observa-tions highlight the importance of maintain-ing flow regimes that resemble natural condi-tions. Proper flow regimes in California’sCentral Valley streams benefit native speciesand probably keep alien species at bay(Marchetti and Moyle 2001; Brown and Ford2002). Natural flow regimes have also beendemonstrated to benefit native fish assem-blages in other regions. Travnichek et al. (1995)observed an increase in fish abundance anddiversity following enhanced flow regimes inthe Tallapoosa River, Alabama. Meffe (1991)noted that a natural flow regime prevented theestablishment of alien bluegill Lepomis macro-chirus in a South Carolina blackwater stream.Additionally, natural flow regimes, such asseasonal flooding, are important in maintain-ing native fish assemblages in the southwest-ern United States (Minckley and Meffe 1987;Gido et al. 1997). Although these examplessuggest that there is promise in restoring na-tive fishes with natural flow regimes, resultsof this study suggest that manipulations inflow regimes alone will probably not result insubstantial changes in the overall south deltafish assemblage. This conclusion stems fromthe fact that high assemblage similarity wasobserved among years in which inflow variedtremendously. Although a natural flow regimewill probably enhance the production of na-tive fishes during the spring flow pulse, the
production of alien species will also be main-tained during summer once the natural springflow pulse has receded. Therefore, althoughthere may be management options which havethe opportunity to enhance production of na-tive species, decreasing the production of alienspecies in the delta remains a considerableproblem.
Acknowledgments
This study was implemented in the fieldthrough the direction of S. Spaar with the as-sistance of many dedicated seasonal techni-cians. I am grateful to J. Wang for identifyingthe fish larvae, to P. Nader for laboringthrough the CALSIM hydrology modeling, toW. Kimmerer and T. Robinson for statisticaladvice, and to H. Rooks for the original sup-port to conduct this work. Comments by Z.Hymanson, M. Nobriga, S. Matern, J. Orsi, T.Sommer, and two anonymous reviewers im-proved the manuscript.
References
Arthur, J. F., M. D. Ball, and M. Y. Baughman. 1996.Summary of federal and state water project envi-ronmental impacts in the San Francisco Bay-DeltaEstuary, California. Pages 445–495 in J. T.Hollibaugh, editor. San Francisco Bay: the ecosys-tem. American Association for the Advancementof Science, San Francisco.
Bennett, W. A., and P. B. Moyle. 1996. Where have all thefishes gone? Interactive factors producing fish de-clines in the Sacramento-San Joaquin Estuary.Pages 519–542 in J. T. Hollibaugh, editor. San Fran-cisco Bay: the ecosystem. American Associationfor the Advancement of Science, San Francisco.
Brown, L. R., and T. J. Ford. 2002. Effects of flow on thefish communities of a regulated California river:implications for managing native fishes. River Re-search and Applications 18:331–342.
Feyrer, F., and M. P. Healey. 2003. Fish community struc-ture and environmental correlates in the highly al-tered southern Sacramento-San Joaquin Delta. En-vironmental Biology of Fishes 66:123–132.
Gadomski, D. M., and C. A. Barfoot. 1998. Diel anddistributional patterns of fish embryos and larvaein the lower Columbia and Deschutes rivers. Envi-ronmental Biology of Fishes 51:353–368.
Gauch, H. G. 1982. Multivariate analysis in communityecology. Cambridge University Press, Cambridge.
Gido, K. B., D. L. Propst, and M. C. Molles, Jr. 1997.Spatial and temporal variation of fish communi-

78 FEYRER
ties in secondary channels of the San Juan River,New Mexico and Utah. Environmental Biology ofFishes 49:417–434.
Grimaldo, L. F., R. E. Miller, C. M. Peregrin, and Z. P.Hymanson. 2004. Spatial and temporal distribu-tion of native and alien ichthyoplankton in threehabitat types of the Sacramento–San Joaquin Delta.Pages 81–96 in F. Feyrer, L. R. Brown, R. L. Brown,and J. J. Orsi, editors. American Fisheries Society,Symposium 39, Bethesda, Maryland.
Harvey, B. C. 1991. Interaction of abiotic and biotic fac-tors influences larval fish survival in an Oklahomastream. Canadian Journal of Fisheries and AquaticSciences 48:1476–1480.
Houde, E. D. 1987. Fish early life history dynamics andrecruitment variability. Pages 17–29 in R. D. Hoyt,editor. 10th annual larval fish conference. Ameri-can Fisheries Society, Symposium 2, Bethesda,Maryland.
Jassby, A. D., W. J. Kimmerer, S. G. Monismith, C. Ar-mor, J. E. Cloern, T. M. Powell, J. R. Schubel, and T.J. Vendlinski. 1995. Isohaline position as a habitatindicator for estuarine populations. EcologicalApplications 5:272–289.
Keller, A. A., G. Klein-MacPhee, and J. St. Onge Burns.1999. Abundance and distribution of ichthyoplank-ton in Narragansett Bay, Rhode Island, 1989–1990.Estuaries 22:149–163.
Kimmerer, W. J., E. Gartside, and J. J. Orsi. 1994. Preda-tion by an introduced clam as the likely cause ofsubstantial declines in zooplankton of San FranciscoBay. Marine Ecology Progress Series 113:81–93.
Kimmerer, W. J. 2002. Effects of freshwater flow on abun-dance of estuarine organisms: physical effects ortrophic linkages. Marine Ecology Progress Series243:39–55.
Lazzari, M. A. 2001. Dynmanics of larval fish abun-dance in Penobscot Bay, Maine. Fisheries Bulletin99:81–93.
Legendre, P., and L. Legendre. 2000. Numerical ecology.Developments in Environmental Modeling 20.Elsevier Science BV, Amsterdam.
Lemly, A. D. 1985. Suppression of native fish popula-tions by green sunfish in first-order streams of Pied-mont, North Carolina. Transactions of the Ameri-can Fisheries Society 114:705–712.
Letcher, B. H., J. A. Rice, L. B. Crowder, and K. A. Rose.1996. Variability in survival of larval fish: disen-tangling components with a generalized individual-based model. Canadian Journal of Fisheries andAquatic Sciences 53:787–801.
Marchetti, M. P., and P. B. Moyle. 2000. Spatial andtemporal ecology of native and introduced fish lar-vae in lower Putah Creek, California. Environmen-tal Biology of Fishes 58:75–87.
Marchetti, M. P., and P. B. Moyle. 2001. Effects of flowregime on fish assemblages in a regulated Califor-nia stream. Ecological Applications 11:530–539.
Marsh, P. C., and M. E. Douglas. 1997. Predation byintroduced fishes on endangered humpback chub
and other native species in the Little Colorado River,Arizona. Transactions of the American FisheriesSociety 126:343–346.
Meffe, G. K. 1985. Predation and species replacement inAmerican southwestern fishes: a case study. South-western Naturalist 30:173–187.
Meffe, G. K. 1991. Failed invasion of a southeasternblackwater stream by bluegills: implications forconservation of native communities. Transactionsof the American Fisheries Society 120:333–338.
Meng, L., and S. A. Matern. 2001. Native and intro-duced larval fishes of Suisun Marsh, California:the effects of freshwater flow. Transactions of theAmerican Fisheries Society 130:750–765.
Minckley, W. L., and G. K. Meffe. 1987. Differential selec-tion by flooding in stream fish communities of thearid American southwest. Pages 93–104 in W. J.Matthews and D. C. Heins, editors. Communityand evolutionary ecology of North American streamfishes. University of Oklahoma Press, Norman.
Moyle, P. B. 2002. Inland fishes of California. Revisedand expanded. University of California Press, Ber-keley.
Moyle, P. B. 1986. Fish introductions into North America:patterns and ecological impact. Pages 27–43 in H.A. Mooney and J. A. Drake, editors. Ecology ofbiological invasions of North America and Hawaii.Springer-Verlag, New York.
Nichols, F. H., J. E. Cloern, S. N. Luoma, and D. H.Peterson. 1986. The modification of an estuary. Sci-ence 231:567–573.
Powles, H., F. Auger, and G. J. Fitzgerald. 1984.Nearshore ichthyoplankton of a north temperateestuary. Canadian Journal of Fisheries and AquaticSciences 41:1653–1663.
Ross, S. T. 1991. Mechanisms structuring stream fish as-semblages: are there lessons from introduced spe-cies? Environmental Biology of Fishes 30:359–368.
Scoppettone, G. G. 1993. Interactions between nativeand nonnative fishes of the upper Muddy River,Nevada. Transactions of the American FisheriesSociety 122:599–608.
Sommer, T. R., W. C. Harrell, R. Kurth, F. Feyrer, S. C.Zeug, and G. O’Leary. 2004. Ecological patterns ofearly life stages of fishes in a large river-floodplainof the San Francisco Estuary. Pages 111–123 in F.Feyrer, L. R. Brown, R. L. Brown, J. J. Orsi. Early lifehistory of fishes in the San Francisco Estuary andwatershed. American Fisheries Society, Symposium39, Bethesda, Maryland.
Sommer, T., R. Baxter, and B. Herbold. 1997. Resilienceof splittail in the Sacramento–San Joaquin Estu-ary. Transactions of the American Fisheries Society126:961–976.
ter Braak, C. J. F., and P. Smilauer. 1998. CANOCOreference manual and user’s guide to CANOCOfor Windows: software for canonical communityordination (version 4). Microcomputer Power,Ithaca, New York.
Travnichek, V. H., M. B. Bain, and M. J. Maceina. 1995.

ECOLOGICAL SEGREGATION OF NATIVE AND ALIEN LARVAL FISH ASSEMBLAGES 79
Recovery of a warmwater fish assemblage after theinitiation of a minimum-flow release downstreamfrom a hydroelectric dam. Transactions of theAmerican Fisheries Society 124:836–844.
Wang, J. C. S. 1986. Fishes of the Sacramento-San JoaquinEstuary and adjacent waters, California: a guideto their early life histories. California Departmentof Water Resources, Interagency Ecological Pro-gram, Technical Report 9, Sacramento, California.
Witting, D. A., K. W. Able, and M. P. Fahay. 1999. Lar-val fishes of a middle Atlantic Bight estuary: as-semblage structure and temporal stability. Cana-dian Journal of Fisheries and Aquatic Sciences56:222–230.
Yoklavich, M. M., M. Stevenson, and G. M. Calliet. 1992.Seasonal and spatial patterns of ichthyoplanktonabundance in Elkhorn Slough, California. Estua-rine, Coastal, and Shelf Science 34:109–126.






























