Embed Size (px)
Citation preview

Io
ASa
b
c
d
a
AA
KEEEEOSZ
1
t2tnaaaEi
UT
0d
Ecological Modelling 220 (2009) 3194–3209
Contents lists available at ScienceDirect
Ecological Modelling
journa l homepage: www.e lsev ier .com/ locate /eco lmodel
nvasive species impacts on ecosystem structure and function: A comparisonf Oneida Lake, New York, USA, before and after zebra mussel invasion
ndrea L. Jaeger Miehlsa,b,∗, Doran M. Masona,b, Kenneth A. Franka,c, Ann E. Kraused,cott D. Peacora, William W. Taylora
Department of Fisheries and Wildlife, Michigan State University, 13 Natural Resources Building, East Lansing, MI 48824, USANOAA Great Lakes Environmental Research Laboratory, 4840 S. State Road, Ann Arbor, MI 48108, USADepartment of Counseling, Educational Psychology, and Special Education, Michigan State University, 462 Erickson Hall, East Lansing, MI 48824, USADepartment of Environmental Sciences, The University of Toledo, 2801 Bancroft Street, Toledo, OH 43606, USA
r t i c l e i n f o
rticle history:vailable online 24 August 2009
eywords:cological network analysiscosystem functioncosystem structurexotic speciesneida Lakeocial network analysisebra mussels
a b s t r a c t
Exotic species invasion is widely considered to affect ecosystem structure and function. Yet, few con-temporary approaches can assess the effects of exotic species invasion at such an inclusive level. Ourresearch presents one of the first attempts to examine the effects of an exotic species at the ecosystemlevel in a quantifiable manner. We used ecological network analysis (ENA) and a social network analy-sis (SNA) method called cohesion analysis to examine the effect of zebra mussel (Dreissena polymorpha)invasion on the Oneida Lake, New York, USA, food web. We used ENA to quantify ecosystem functionthrough an analysis of food web carbon transfer that explicitly incorporated flow over all food web paths(direct and indirect). The cohesion analysis assessed ecosystem structure through an organization of foodweb members into subgroups of strongly interacting predators and prey. Our analysis detected effectsof zebra mussel invasion throughout the entire Oneida Lake food web, including changes in trophic flowefficiency (i.e., carbon flow among trophic levels) and alterations of food web organization (i.e., paths of
carbon flow) and ecosystem activity (i.e., total carbon flow). ENA indicated that zebra mussels alteredfood web function by shunting carbon from pelagic to benthic pathways, increasing dissipative flow loss,and decreasing ecosystem activity. SNA revealed the strength of zebra mussel perturbation as evidencedby a reorganization of food web subgroup structure, with a decrease in importance of pelagic pathways,a concomitant rise of benthic pathways, and a reorganization of interactions between top predator fish.Together, these analyses allowed for a holistic understanding of the effects of zebra mussel invasion on.
the Oneida Lake food web. Introduction
Globally, exotic species invasion has altered ecosystem struc-ure and function (Vitousek et al., 1987; Wilcove et al., 1998; Crooks,002). Non-native species impact ecosystems through competi-ion with and predation on native species, and by altering habitats,utrient cycles, and energy budgets (Baxter et al., 2004; Mack etl., 2000). Invasions occur across all trophic levels and potentially
ffect entire food webs (Crooks, 2002). In recent years, vertebratend invertebrate species from the Ponto-Caspian region of easternurope have exerted pronounced invasion pressure in North Amer-ca, Europe, and Russia (Drake and Bossenbroek, 2004). Among∗ Corresponding author at: Department of Fisheries and Wildlife, Michigan Stateniversity, 13 Natural Resources Building, East Lansing, MI 48824, USA.el.: +1 517 353 7981; fax: +1 517 432 1699.
E-mail addresses: [email protected], [email protected] (A.L.J. Miehls).
304-3800/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.ecolmodel.2009.07.020
© 2009 Elsevier B.V. All rights reserved.
the most recognized Ponto-Caspian invertebrate invaders in NorthAmerica are two mollusk species, the zebra (Dreissena polymorpha)and quagga (D. rostriformis bugensis) mussels.
Dreissenid mussel invasion can considerably affect aquatic envi-ronments (Jones et al., 1994; Vanderploeg et al., 2002). Collectively,Mills et al. (2003) summarize the effects of dreissenid invasion asthe process of “benthification”, which entails ecosystem changesthat facilitate growth of benthic algae and submersed vegetation,favor colonization of bottom-dwelling organisms, and promote fishcommunities that use benthic habitat. The process of benthificationencompasses many of the ecosystem changes observed subsequentto dreissenid invasion, yet determining causality in these changeshas proven difficult.
The most clearly attributable effects of dreissenid musselsoccur at lower trophic levels. Zebra mussels increase water clar-ity through filtration (Idrisi et al., 2001) and shunt energy frompelagic to benthic pathways, thereby promoting benthic sources ofprimary production (Rutherford et al., 1999; Mayer et al., 2002; Zhu

Mode
ezmaTpsasrvLwDa
dlDteGeals(ip2d(aeJrsan
ifsctieofeoTsfief
paq(upbfla
A.L.J. Miehls et al. / Ecological
t al., 2006). Additionally, dreissenid mussels can indirectly affectooplankton production and abundance by decreasing pelagic pri-ary production and changing nutrient cycling in lakes (Mellina et
l., 1995; Johannsson et al., 2000) and rivers (Strayer et al., 1999;horp and Casper, 2003), although in some lakes (e.g., Oneida Lake)elagic primary production did not change after zebra mussel inva-ion (Idrisi et al., 2001). Also, dreissenid mussels may increase thebundance of macroinvertebrates, such as amphipod and isopodpecies, through pseudofeces deposition which is used as a foodesource (Beekey et al., 2004). Yet, the abundance of some macroin-ertebrate species, such as Diporeia spp. (a native amphipod in theaurentian Great Lakes), has precipitously declined in correlationith the invasion and expansion of dreissenids (Nalepa et al., 2000).reissenid effects on intermediate trophic levels are, thus, morembiguous than for lower trophic levels.
Dreissenid invasion may also affect fish communities, butrawing inferences about dreissenid effects on fish is particu-
arly difficult due to their separation by multiple trophic levels.reissenid mussels may affect fish recruitment and production
hrough modification or degradation of spawning habitat (Stewartt al., 1999; Marsden and Chotkowski, 2001). Additionally, somereat Lakes fishes, such as lake whitefish (Coregonus clupeaformis),xhibit decreased body condition and growth potentially due todiet shift away from high-energy prey, such as Diporeia, to the
ower-energy dreissenid mussels (Pothoven et al., 2001). Dreis-enid effects on other important fish species, e.g., yellow perchPerca flavescens), vary by life stage and ecosystem. For examplen Lake Michigan, zebra mussels may adversely affect larval yellowerch by decreasing zooplankton food resources (Dettmers et al.,003). Conversely in some inland lakes, zebra mussels apparentlyo not affect young yellow perch growth, biomass, or productionMayer et al., 2000; Idrisi et al., 2001) and may indirectly benefitdult yellow perch and lake sturgeon (Acipenser fulvescens) throughnhanced benthic invertebrate production (Rutherford et al., 1999;ackson et al., 2002). Some Centrarchidae species also benefit indi-ectly by zebra mussel facilitation of macrophyte production whicherves as habitat (Strayer et al., 2004). Overall, although difficult tossess, this research suggests that dreissenids affect fish commu-ities, but these effects are variable.
Part of the difficulty in understanding the effects of dreissenidnvasion at all trophic levels is the complexity of linkages withinood webs. Due to this complexity, most research aimed at under-tanding dreissenid effects on higher trophic levels, including fishommunities, is largely based on correlational data and observa-ions of biomass change after dreissenid invasion. Yet, if food webnteractions are delineated, food webs offer the ability to traceffects of lower trophic level perturbation (i.e., dreissenid invasion)n higher trophic levels. Moreover, if energy and material flow inood web pathways are quantified (i.e., feeding relationships arestimated), then a more realistic depiction of food web processes isbtained, which allows for a better description of indirect effects.hus, quantifying food web pathways allows for a rigorous under-tanding of dreissenid effects on higher trophic levels, includingsh communities. A suite of integrated and quantitative approachesxist, known as network analyses, that can be used to analyze theseood webs once they are obtained.
Network analysis offers innovative methods to clarify the com-lex relationships in food webs of invaded ecosystems. For networknalysis, food webs are depicted as networks of exchange byuantifying (i.e., weighting) feeding interactions and energy flowBondavalli et al., 2000). Much of the early work on food webs used
nweighted flows, which can over-simplify or misrepresent fluxrocesses: the role of taxa involved in small magnitude flows maye exaggerated while the role of taxa involved in large magnitudeows may be diminished. By quantifying flows, researchers teasepart these unequal contributions to provide a clearer depiction oflling 220 (2009) 3194–3209 3195
food web energy and material transfer and allow a better under-standing of how interactions are transmitted through ecosystems.
One of the most valuable attributes of network analysis is itsapplicability at the whole-system level; few other contemporaryapproaches assess processes at such an inclusive level. By quantify-ing interactions throughout the food web, insight into processes notevident at small-scale resolutions is gained (Simon, 1962; Heymanset al., 2002). Bailey et al. (1999) suggest small-scale impacts aggre-gate synergistically and these overall effects are only visible whenviewed at the whole-system level. As such, network analysis provesuseful for examining the effects of fisheries harvest (Pauly et al.,1998), fisheries stocking (Fayram et al., 2006), and nutrient loading(Bondavalli et al., 2006) on ecosystems, while having implicationsfor managing carbon emissions (Bondavalli et al., 2000) and habi-tat for endangered species (Heymans et al., 2002). By accountingfor entire systems, the use of network analysis in these studiesallows an understanding – distinct from, but complementary to,single-species, species-pair or similar studies – of how changespercolate through food webs, including indirect pathways. More-over, due to the food web approach, network analysis is useful forunderstanding how changes in lower trophic levels affect highertrophic levels. Therefore, network analysis is a valuable tool forclarifying the effects of ecosystem perturbation, including exoticspecies invasion, at lower trophic levels. In light of the equivocalconsequences of dreissenid invasion described above, we elucidatedreissenid effects at the ecosystem scale using network analysis.
Ecosystem level change can be parsed into two categories:(1) effects on ecosystem structure, and (2) effects on ecosystemfunction. Ecosystem structure encompasses the components of afood web, whereas ecosystem function refers to the processeswithin food webs (Stevenson et al., 1996). Structural character-istics include measurements of system state, such as biomassand taxonomic composition; structural characteristics also includecompartmentalization of food webs into “subgroups” of stronglyinteracting taxa (e.g., Simon, 1962; Pimm, 1979; Pimm and Lawton,1980; Krause et al., 2003). Functional characteristics encompassrates of change in a system, including measurements of produc-tivity and respiration (Stevenson et al., 1996). Given the inclusiveand quantifiable nature of ecosystem structure and function, weuse these attributes to understand ecosystem response to zebramussel invasion at all trophic levels (sensu the concept of networkadaptation to changed conditions from Jørgensen, 2007).
The primary objective of this research is to quantify zebra mus-sel effects on ecosystem structure and function. We base thisanalysis on an extensive data set of food web linkages from OneidaLake, New York, USA, assembled to model ecosystem dynamics asaffected by several recent invaders (Irwin, 2006; Koops et al., 2006).We use network analyses to address structural and functional ques-tions jointly in Oneida Lake in order to understand how structuraland functional food web characteristics interact in response to per-turbation (Jørgensen, 2007). Specifically, we address the followingquestions:
1. Structure: Do zebra mussels alter the membership of food websand food web subgroups (defined as clusters of strongly inter-acting predators and prey)?
2. Function: Does the magnitude of carbon flow within food websand food web subgroups change as a result of zebra mussel inva-sion?
We emphasize changes in structure and function of higher
trophic level communities due to the difficulty in assessing thesechanges with traditional ecological approaches. Although otherecosystem changes concomitant with the increase in zebra mus-sels may affect the results, we emphasize zebra mussel effectsbecause this species is known to strongly affect invaded ecosystems
3 l Mode
aLao
2
2
Lm2scav(aefiF–fie
2
tt1sa1yctttd
fsmnaTiae(gsTwiFipdtwtp
196 A.L.J. Miehls et al. / Ecologica
nd has higher biomass than any other species present in Oneidaake (Koops et al., 2006). Finally, as Mayer et al. (2002) and Zhu etl. (2006) suggest, we look for further evidence of benthificationccurring in Oneida Lake.
. Methods
.1. Study site
Oneida Lake is a shallow, eutrophic, 207 km2 lake on the Ontarioake Plain in central New York with a mean depth of 6.8 m andaximum depth of 16.8 m (Jackson et al., 2002; Rudstam et al.,
004). The lake freezes in winter and mixes continuously duringummer except for brief periods of thermal stratification duringalm weather (Hansen and Hairston, 1998). Oneida Lake supports
valuable warmwater recreational fishery for walleye (Sanderitreus) and yellow perch, in addition to multiple other speciesVanDeValk et al., 2002). Zebra mussels were discovered in 1991nd were established throughout the ecosystem by 1992 (Mayert al., 2000). Because of the long-term history of limnological andshery research on Oneida Lake (e.g., Mills et al., 1978; Mills andorney, 1988), data exist throughout the various stages of invasionfrom pre-introduction, to invasion, through establishment, and
nally reorganization and accommodation of the ecosystem – toxamine the effects of zebra mussel invasion.
.2. Network construction
We constructed and compared weighted food web networks forime periods before and after zebra mussel invasion and analyzedhe networks using ecological network analysis (ENA) (Ulanowicz,986) to assess ecosystem function and a social network analy-is (SNA) method, cohesion analysis (CA) (Frank, 1995; Krause etl., 2003), to assess ecosystem structure. We defined the years986–1991 as the pre-zebra mussel invasion time stanza and theears 1992–2002 as the post-zebra mussel invasion time stanza. Toonstruct the networks, we first identified the taxa in the ecosys-em (including full-year resident taxa as well as transient taxa), andhen quantified carbon exchange (flows) between taxa and withhe environment (we adjusted flows for transient taxa based onuration in the ecosystem).
For taxa identification, we included all species from Oneida Lakeor which data were available. Taxonomic resolution affects thetudy of food webs and lumping organisms into aggregate groupsay decrease the ability to detect differences in strengths of con-
ections between functionally different organisms (Abarca-Arenasnd Ulanowicz, 2002; Krause et al., 2003; Pinnegar et al., 2005).herefore, wherever possible, we used species level data to avoidnformation loss and in some cases, e.g., walleye, yellow perch,nd seven other fish taxa, we divided species by life stage. How-ver, in some cases, data limitations forced aggregation of taxae.g., age-0 centrarchids). In some of these situations, we aggre-ated by “trophospecies”, a collection of species in a food web thathare similar predators and prey (Yodzis and Winemiller, 1999).his aggregation method is a fundamental unit of study in foodeb and ecosystem research (Yodzis and Winemiller, 1999) and
s widely used in food web analysis (e.g., Teal, 1962; Moloney andield, 1991; Bondavalli et al., 2000). Where high resolution trophicnformation was not available (such as for benthic invertebrate andhytoplankton species), clustering adhered closely to taxonomic
istinctions as opposed to trophic relationships. Finally, data onhe microbial food web were not available for Oneida Lake andere not included in our food web networks. A complete list ofhe seventy-seven species and aggregate groups for the pre- andost-zebra mussel invasion networks are listed in Table 1.
lling 220 (2009) 3194–3209
We next created the network connection topography, i.e., foodweb exchanges. Exchanges occur both within the food web throughpredator–prey interactions and between the food web and sur-rounding environment through migration, primary production,respiration, and harvest (Bondavalli et al., 2000). We used carbon asour network currency for biomass (gC m−2) and exchange betweentaxa (gC m−2 yr−1). To estimate carbon exchanges, we obtaineddata on biomass, production-, consumption-, and respiration-to-biomass ratios, diet proportions, migrations, and harvest for all taxafrom the primary literature, field studies, and expert researcherson Oneida Lake. These parameters along with their sources arelisted in Appendix 1. Exchanges either flow into a taxa as a car-bon input (e.g., consumption and immigration), or flow out ofthe taxa as a carbon output (e.g., production, respiration, harvest,and emigration). To calculate production, consumption, and res-piration for taxa, we multiplied the production-, consumption-,and respiration-to-biomass ratios by the taxa biomass. To quan-tify feeding relationships between predatory taxa and prey, weapportioned the predator’s consumption among its prey items bymultiplying the predator consumption estimate and predator dietvector (a list of predator diet items proportioned by weight inthe diet). This calculation yielded a vector of carbon exchanges(gC m−2 yr−1) between the predator and its prey. Aligning theexchange vectors for every predator in the system creates theexchange matrix. The exchange matrix quantifies all feeding rela-tionships within the food web network and is the foundation of ournetwork analysis calculations (see Ulanowicz, 1986 for details).
ENA routines require networks to be mass-balanced, i.e., a con-dition where the amount of medium entering any taxa equals theamount leaving (Allesina and Bondavalli, 2003). We assessed theflow balance of our networks by comparing inputs and outputs forall taxa. If a flow imbalance existed, we rectified the discrepancy byeither assigning a flow to detritus (if inputs exceeded outputs) orchanging one or more model inputs (Christensen and Pauly, 1992;Heymans and Baird, 2000). The detrital-balancing approach is par-tially drawn from standard mass-balance methods employed in thesoftware EcoPath (Christensen and Pauly, 1992) and was our meansto assign flow to detritus – an important component of networkmodels (Jørgensen, 2007) – in lieu of field estimates. When chang-ing a model input to obtain mass-balance, we used factors such asecological plausibility (i.e., the similarity of the Oneida Lake param-eter to other systems of similar trophic status) and confidence inaccuracy of parameter estimates as guides to determine whichparameter to change. Finally, to resolve any remaining imbalances,we used the DATBAL routine incorporated in the EcoNetwrk soft-ware (http://www.glerl.noaa.gov/EcoNetwrk/), a Windows-basedversion of the NETWRK software (Ulanowicz and Kay, 1991). Theseroutines resulted in balanced food web networks for pre- and post-zebra mussel invasion with as few changes to the original modelinputs as possible. It should be noted that balancing routines canintroduce uncertainty into model inputs; however, our routinesresulted in small percent changes to model inputs and all changeswere inspected for ecological plausibility. The balanced exchangematrices for the pre- and post-zebra mussel invasion networks arepresented in Appendix 2. As with any modeling study, the resultsof this analysis are based on the assembled model and the qualityof the results depend on the validity of the input data. Thus, wewere careful to build models as ecologically accurate as possiblefor Oneida Lake. We acknowledge that our models are not perfect,but believe our models represent a highly thorough account of foodweb dynamics in Oneida Lake.
2.3. Ecological network analysis
After we constructed the networks, we performed the ENA andCA routines to quantify Oneida Lake function (via ENA) and struc-

A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209 3197
Table 1List of food web taxa for Oneida Lake food web networksa.
No. Common Name Taxonomic Classification No. Common Name Taxonomic Classification
1 Cormorants Phalacrocorax auritus 41 Leeches Hirudinea2 Walleye Age >4 Sander vitreus 42 Oligochaetes Oligochaeta3 Walleye Age 1–3 Sander vitreus 43 Snails Gastropoda4 Walleye Age-0 Sander vitreus 44 Zebra Mussels Dreissena polymorpha5 Yellow Perch Age >3 Perca flavescens 45 Alona species Alona species6 Yellow Perch Age 1–2 Perca flavescens 46 Bosmina longirostris Bosmina longirostris7 Yellow Perch Age-0 Perca flavescens 47 Camptocercus harpae Camptocercus harpae8 White Perch Age >1 Morone americana 48 Ceriodaphnia quadrangular Ceriodaphnia quadrangular9 White Perch Age-0 Morone americana 49 Chydorus sphaericus Chydorus sphaericus
10 Black Crappie Age >1 Pomoxis nigromaculatus 50 Daphnia galeata mendotae Daphnia galeata mendotae11 Bluegill Age >1 Lepomis macrochirus 51 Daphnia pulicaria Daphnia pulicaria12 Pumpkinseed Age >1 Lepomis gibbosus 52 Daphnia retrocurva Daphnia retrocurva13 Rock Bass Age >1 Ambloplites rupestris 53 Diaphanosoma species Diaphanosoma species14 Centrarchids Age-0 Centrarchidae 54 Eubosmina coregoni Eubosmina coregoni15 Gizzard Shad Age >1 Dorosoma cepedianum 55 Sida crystallina Sida crystallina16 Gizzard Shad Age-0 Dorosoma cepedianum 56 Leptodora kindtii Leptodora kindtii17 Alewife Alosa pseudoharengus 57 Acanthocyclops vernalis Acanthocyclops vernalis18 Brown Bullhead Ameiurus nebulosus 58 Diacyclops thomasi Diacyclops thomasi19 Burbot Lota lota 59 Ergasilus species Ergasilus species20 Channel Catfish Ictalurus punctatus 60 Mesocyclops edax Mesocyclops edax21 Cisco Coregonus artedii 61 Epischura lacustris Epischura lacustris22 Common Carp Cyprinus carpio carpio 62 Leptodiaptomus minutes Leptodiaptomus minutus23 Darters Etheostoma species 63 Skistodiaptomus oregonensis Skistodiaptomus oregonensis24 Emerald Shiners Notropis athernoides 64 Nauplii Copepoda25 Freshwater Drum Aplodinotus grunniens 65 Rotifers Rotatoria26 Golden Shiners Notemigonus crysoleucas 66 Blue-green Algae Cyanophyceae27 Lake Sturgeon Acipenser fulvescens 67 Diatoms Bacillariophyceae28 Log Perch Percina caprodes 68 Euglena Euglenophyceae29 Mottled Sculpin Cottus bairdii 69 Flagellates Cryptophyceae and Dinophyceae30 Northern Pike Esox lucius 70 Golden Algae Chrysophyceae31 Red Horse Sucker Moxisoma species 71 Green Algae Chlorophyceae32 Smallmouth Bass Micropterus dolomieu 72 Epiphytes Epiphytes33 Trout Perch Percopsis omiscomaycus 73 Macrophytes Macrophytes34 White Bass Morone chrysops 74 Periphytes Periphytes35 White Sucker Catostomus commersonii 75 Pelagic Detritus Pelagic Detritus36 Amphipods Amphipoda 76 Sedimented Detritus Sedimented Detritus37 Chironomids Chironomidae 77 DOC DOC38 Clams Sphaeriidae39 Insects Arthropoda40 Isopods Isopoda
ussei 1–35a
tfpaen
TSp
a Cisco were present in the pre-zebra mussel time stanza but not the post-zebra mn the post-zebra mussel time stanza, but not the pre-zebra mussel time stanza. Taxand 75–77 are detrital groups.
ure (via CA) before and after zebra mussel invasion (see Table 2
or a summary of these routines and their relationship to ecosystemroperties). ENA is a method that evaluates the efficiency of energynd material flow (e.g., transfers, assimilations, and dissipations) incosystems (Ulanowicz, 1986). We used three types of ecologicaletwork analyses (Ulanowicz, 1986): (1) input/output analysis, (2)able 2ummary of network analysis routines and their relationships to ecosystemroperties.
Network analysis routine Ecosystem property
Ecological network analyses: Ecosystem functionInput/output analysis Direct and indirect effectsTrophic level analysis Trophic flow efficiencyEcosystem indices: Quantification of whole system
functionTotal system throughput (TST) Ecosystem size and growthAverage mutual information (AMI) Ecosystem flow organizationAscendency (A) Ecosystem growth and
developmentOverhead (O) Ecosystem inefficiencies and flow
redundancyDevelopment capacity (C) Upper bound on ecosystem growth
and development
Social network analysis: Ecosystem structureCohesion analysis Subgroup structure
l time stanza; lake sturgeon, zebra mussels, and Camptocercus harpae were presentare fish, 36–44 are benthic invertebrates, 45–65 are zooplankton, 66–74 are plants,
trophic level analysis, and (3) the calculation of ecosystem indices.All analyses were performed using EcoNetwrk software. We pro-vide a brief description of ENA methods below; for greater detailsee Ulanowicz (1986) and Ulanowicz (1997).
Input/output analysis (Hannon, 1973; Patten et al., 1976;Szyrmer and Ulanowicz, 1987) details the direct and indirect link-ages between any two taxa in a network, quantifying the requisitecarbon needs (or other element, e.g., nitrogen or phosphorus, orenergy, e.g., calories, depending on the goal of the analysis) of anyone taxon supplied by any other taxon (Bondavalli et al., 2000).The analysis includes a routine called IMPACTS that quantifies therelative effect of one taxon on another through direct and indirectpredatory interactions (Ulanowicz and Puccia, 1990; Heymans andBaird, 2000). The IMPACTS analysis is a rigorous way to trace trophiceffects of zebra mussels in the Oneida Lake food web since all feed-ing interactions, both direct and indirect, are incorporated in theanalysis. Indirect paths are especially important as they may over-whelm direct interactions and even yield an opposite effect thana direct exchange between two taxa (Patten, 1984). Importantly,the IMPACTS analysis is conducted only using one network; a com-
parison of networks is not done. Therefore, our IMPACTS analysisused only the post-zebra mussel network (as opposed to involv-ing a comparison between networks before and after invasion). Assuch, the IMPACTS analysis provides a snapshot of trophic effects(e.g., trophic cascades) involving zebra mussels.
3198 A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209
Table 3Association between common subgroup membership and the occurrence of ties between predators and prey (adapted from Frank, 1995). The odds ratio method maximizesthe ratio AD:BC.
Tie Occurring
No Yes
Intera
tdtoailWasc
st(((
assmttUocttAfcitatt
eiwsFbaflObttt(p(c
Using the software Kliquefinder (Frank, 1995), we identifiedsubgroups within the Oneida Lake food web networks before andafter zebra mussel invasion and compared the grouped webs toreveal changes in food web structure. Although our ENA required
Subgroup Membership Different A BSame C D
Unrealized Interactions Realized
Trophic level analysis condenses the web of predator-preyransfers into a linear trophic chain following the concept of Lin-eman (Lindeman, 1942; Ulanowicz, 1995). Using input/outputechniques, the trophic analysis apportions the feeding activitiesf taxa among a series of hypothetical integer trophic levels to cre-te a trophic chain called the Lindeman spine. The Lindeman spines used to evaluate the efficiency of carbon flow between trophicevels and at the whole-system level (Heymans and Baird, 2000).
e used the trophic level analysis to create Lindeman spines beforend after zebra mussel invasion and compared the spines to under-tand changes in flow efficiency (but note, as described below, weould only create spines at the subgroup level).
Ecosystem indices quantify abstract system level propertiesuch as growth and development, and provide insight into bothhe vulnerability and resilience of an ecosystem to perturbationUlanowicz, 1997). These indices are: total system throughputTST), average mutual information (AMI), ascendency (A), overheadO), and development capacity (C).
TST quantifies the activity level of the ecosystem as the sum ofll carbon (or other elemental or energetic currency) flows in theystem and has units of production (gC m−2 yr−1). TST relates to theize of an ecosystem gauged similarly to how economic activity iseasured (i.e., as the flow of some currency). AMI is an information
heoretic index (Shannon, 1948; McEliece, 1977) that quantifieshe organization of a network based on pathway flow constraints.sing conditional probabilities, AMI quantifies the average amountf information known about the pathways a unit of material (e.g.,arbon) can follow upon entering a system. The more constrainedhe system is (i.e., the fewer the flow paths), the more informa-ion there is on pathways and the higher the AMI. Likewise, theMI is higher when the amount of material (e.g., carbon) trans-
erred is concentrated on a few pathways, while other pathwaysarry comparatively little. In a food web context, a high AMI values found when the food web is actually a food chain, where eachaxon only has one predator and prey and flow is unidirectionalnd predictable. A low AMI value is found in a food web where allaxa are connected to each other and a unit of energy leaving oneaxon would travel to another taxon with equal probability.
Ascendency quantifies the growth and development of ancosystem (Ulanowicz, 1986) as well as the system’s performancen processing medium. Ascendency is calculated by scaling the AMI
ith TST: A = TST × AMI. As such, ascendency encompasses both theize (via TST scaling) and organization (via AMI) of an ecosystem.or example, a food web network with few flows between taxaut large amounts of carbon traveling over those flows would havehigher ascendency value than a food web network with many
ows between taxa, each of which carry a small amount of carbon.verhead quantifies the system’s inefficiencies at processing car-on flow (Heymans et al., 2002) as well as the degree of freedomhat the system has to reconfigure itself in the face of perturba-ion (Heymans and Baird, 2000). There are four main contributors
o ecosystem overhead: imports (i.e., immigrations) to and exportsi.e., emigrations and fisheries harvest) from the ecosystem, dissi-ative flow loss due to respiration, and redundant food web flowsi.e., multiple paths of carbon flow between taxa). Overhead is cal-ulated by scaling the system’s conditional entropy, a measurePossible relations between predators and prey in different subgroupsPossible relations between predators and prey in the same subgroup
ctions Total possible relations
of the disorganization in a network, by TST: O = TST × ConditionalEntropy. Conditional entropy is the uncertainty remaining afterthe flow structure is determined; conditional entropy is comple-mentary to AMI (Ulanowicz, 1986). A network that has manylow-magnitude carbon flows between taxa as well as flows orig-inating or terminating outside the system (e.g., immigrations andemigrations) would have a high overhead value, especially if theseflows are inefficient (e.g., involve high flow losses due to respira-tion).
Intuitively, for any system, an upper limit must exist on organi-zation and growth (or else organization and growth would continueindefinitely). The final index, development capacity, quantifies thisupper limit in ecosystems. Development capacity represents thetheoretical upper bound on system organization and growth andsubsumes both ascendency and overhead, such that: C = A + O. Wecalculated the indices TST, AMI, ascendency, overhead, and devel-opment capacity for our networks before and after zebra musselinvasion to understand functional changes in the Oneida Lake foodweb resulting from zebra mussel invasion.
2.4. Cohesion analysis
Cohesion analysis identifies subgroups in food webs basedon the strength of interactions (i.e., predator-prey relationships)within subgroups (Krause et al., 2003), where the maximization ofan odds ratio is used as a criterion to assign subgroup membership.1
CA uses an iterative algorithm that reassigns taxa to subgroups tomaximize the odds that interactions occur within subgroups, ver-sus between subgroups. The algorithm uses interaction weight topreferentially assign predators and prey with strong interactions tocommon subgroups. This algorithm is a robust method for assign-ing subgroups because it: (1) is an objective technique to assignsubgroups; (2) does not require pre-specification of the numberof subgroups; (3) identifies discrete subgroups (i.e., each taxon isassigned to a single subgroup); and (4) tests the statistical signif-icance of the results (Frank, 1995, 1996; Krause et al., 2003). Todetermine the optimized subgroups, the algorithm sums the weightof realized interactions (i.e., predator–prey exchanges) within asubgroup and evaluates that sum against realized interactions thatoccur between subgroups, in addition to unrealized interactions(i.e., taxa combinations that do not interact) within and betweensubgroups (see Table 3). The intent is to determine the networkstructure that maximizes realized interactions within subgroupsand unrealized interactions between subgroups while minimizingrealized interactions between subgroups and unrealized interac-tions within subgroups.
1 Our use of the term “subgroup” is analogous to the Pimm and Lawton (1980)definition of food web “compartment”. We use the term “subgroup” to avoid con-fusion between Pimm and Lawton (1980) and Ulanowicz (1986) uses of the term“compartment”.

A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209 3199
F he scai re plai Table
bawnedfliTA
to(nocsp
tfcsafslpgtiwr
tefs(m
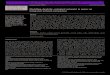
ig. 1. Zebra mussel impacts on the Oneida Lake food web, organized by subgroup. Tmpacts below the zero line are negative impacts of zebra mussels. Taxa numbers anvasion; taxa 75–77 (detrital groups) were not used in the subgroup analysis. See
alanced networks, our CA did not. Therefore, we used the unbal-nced networks for CA of both time stanzas. Furthermore, becausee derive detrital diet as a result of balancing procedures, we couldot include detrital groups in our CA. Finally, the range of our flowsxceeded the input range of Kliquefinder. To remedy the rangeifferences, we parsimoniously adjusted the smallest and largestows to fall within the input range of Kliquefinder. Statistical tests
ndicated that our range adjustments had no bearing on our results.he methods and statistical tests we used are described in detail inppendix 3.
Kliquefinder incorporates a Monte Carlo-like routine to testhe odds ratio of a subgrouped network against a distributionf odds ratios from randomly generated re-combinations of dataFrank, 1996). Using this method, we tested the statistical sig-ificance of our subgroups against 1000 randomized versions ofur networks. Additionally, using the Quadratic Assignment Pro-edure (QAP) method (Hubert, 1987) for significance testing, wetatistically compared subgroup membership between the pre- andost-zebra mussel invasion time stanzas.
We summarized results of the CA as “crystallized sociograms”hrough multidimensional scaling (Frank, 1996) in SAS Systemor Windows. With this scaling method, proximity of subgroupsorresponds to: (1) the strength of predator–prey relationshipspanning the subgroups (i.e., closely spaced subgroups have rel-tively stronger interactions connecting them than subgroupsarther apart); (2) connections to similar subgroups; and (3) theubgroup’s importance to overall food web structure (i.e., centrallyocated subgroups are more important to food web structure thaneripherally located subgroups). Also, taxon location within a sub-roup boundary indicates: (1) the strength of interactions betweenaxa; and (2) the importance of taxa to the subgroup (i.e., taxan the center of subgroups relate more strongly to the subgroup,
hile taxa near subgroup boundaries only experience peripheralelationships to the subgroup).
After we identified cohesive subgroups, we performed ENA onhe largest subgroup before and after zebra mussel invasion to
valuate functional linkages. The largest subgroup was chosen forurther analysis because it was the subgroup containing zebra mus-els and also was the most dominant subgroup in the food webin terms of flow and taxa membership). We maintained the sameass-balance in the subgroup analysis as the full food webs by
le is relative: impacts above the zero line are positive impacts of zebra mussels andced above or below each bar. Taxon 21 (cicso) was not present after zebra mussel
1 for taxa codes.
treating all flows to and from taxa outside the subgroup as importsand exports to subgroup taxa. Because detrital groups were notassigned to subgroups, detritus could not be explicitly incorporatedinto the subgroup ENA. Nevertheless, flow to detritus was implicitlyincluded in the subgroup ENA by treating detrital flow as an exportfrom the subgroup. This procedure had two ramifications: (1) wecould address detrital flow, albeit indirectly, in the subgroups; and(2) we maintained the same mass-balance used in the full food webENA. If zebra mussel invasion had significant effect on the OneidaLake food web, we expected to see changes in the structure andfunction of this subgroup.
3. Results
3.1. Ecological network analysis—entire network
3.1.1. Input/output analysisThe IMPACTS analysis investigates the effects of zebra mussels
expected from the feeding pathways in the food web. This analysisindicated that zebra mussels strongly affected species across mul-tiple trophic levels, with more negative effects across all taxa thanneutral or positive effects (Fig. 1). In general, zebra mussels posi-tively influenced benthic fish such as catfish and bullhead species(taxa 18 and 20 in Fig. 1; hereafter numbers in parentheses referto taxa numbers in Fig. 1), carp (22), and lake sturgeon (27), whilenegatively affecting pelagic fish, including walleye (2–4), yellowperch (5–7; pelagic for some life stages), white perch (8–9), alewife(17), and shiners (24 and 26). Some fish, such as gizzard shad(15–16) and sunfish species (10–14), showed mixed effects basedon life stage (e.g., juvenile gizzard shad were negatively affectedwhile adult gizzard shad were positively affected). Most likely thisis due to ontogenetic diet shifts between the juvenile and adultlife stages (Werner and Gilliam, 1984), although some fish thatundergo diet shifts, e.g., yellow perch, did not show a similar pat-tern. Benthic invertebrates (36–43) exhibited mixed effects, with
amphipods (36) and isopods (40) benefiting from zebra musselinteraction, while zebra mussels had a large negative influence onthemselves (44) due to heavy grazing on their food sources. Zebramussels also negatively influenced phytoplankton (66–71), likelydue to direct consumption, and zooplankton (45–65), likely due
3200 A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209
Table 4Ecosystem indices for the full Oneida Lake food web network before and after zebra mussel invasiona.
Index Pre-Zebra Mussels Value Post-Zebra Mussels Value % Difference Value
Total system throughput (gC m−2 yr−1) 2653.8 2143.6 −19.2Development capacity (gC bits m−2 yr−1) 13316.7 11037.2 −17.1Ascendency (gC bits m−2 yr−1) 3350.9 2926.3 −12.7Total overhead (gC bits m−2 yr−1) 9965.9 8110.9 −18.6
Overhead on imports (gC bits m−2 yr−1) 2598.4 1873.0 −27.9Overhead on exports (gC bits m−2 yr−1) 0.4 0.3 −35.0Dissipative overhead (gC bits m−2 yr−1) 3936.3 3203.3 −18.6Redundancy (gC bits m−2 yr−1) 3430.8 3034.3 −11.6
Unscaled capacity (bits) 5.0 5.1 2.0Ascendency/capacity (%) 25.2 26.5 5.4Total overhead/capacity (%) 74.8 73.5 −1.8
Overhead on imports/capacity (%) 19.5 17.0 −13.0Overhead on exports/capacity (%) 0.0 0.0 −21.6
tea4mmmssmAti
3
pfipea5wLtoswt
3
2et1s2o1cdceUf
Dissipative overhead/capacity (%) 29.6Redundancy/capacity (%) 25.8
a The percent difference was calculated as: %Difference = Post − Pre/Pre × 100%.
o competition for phytoplankton. Zebra mussels had little, if any,ffect on some fishes (e.g., smallmouth bass and white bass, 32nd 34, respectively), invertebrates (e.g., oligochaetes and snails,2 and 43, respectively), and periphyton (74), epiphyton (72), andacrophytes (73). Finally, zebra mussel influence on detritus wasixed: pelagic detritus (75) and DOC (77) were negatively affected,ost likely through direct consumption by zebra mussels, while
edimented detritus (76) was positively affected likely due to depo-ition of pseudofeces. The positive effect on sedimented detritusay explain why influence on benthic invertebrates was mixed.mphipods and isopods rely more heavily on sedimented detri-
us and less heavily on plankton resources than do other benthicnvertebrates such as chironomids and clams (Appendix 2).
.1.2. Lindeman trophic analysisZebra mussel invasion affected total flow loss due to res-
iration in the Oneida Lake food web with a 23% decreaserom 1015.6 gC m−2 yr−1 before invasion to 777.9 gC m−2 yr−1 afternvasion. Zebra mussel invasion also marginally decreased totalroduction (i.e., feeding exchanges between taxa plus the usablexports from the system) by 5% in the Oneida Lake food web withdrop from 624.1 gC m−2 yr−1 before zebra mussel invasion to
89.1 gC m−2 yr−1 after zebra mussel invasion. Unfortunately, weere unable to create the Lindeman trophic spine for the full Oneida
ake food web networks due to the complexity of trophic interac-ions in our networks; trophic analysis computations were run forver six months without determining a Lindeman spine (the analy-is was finally terminated by an extended power outage). Therefore,e could not analyze efficiency of flow between trophic levels for
he full food web networks.
.1.3. Ecosystem indicesTST decreased from 2653.8 before zebra mussel invasion to
143.6 gC m−2 yr−1 after invasion, an approximately 20% loss incosystem activity (Table 4). Additionally, the potential for sys-em development (i.e., development capacity) decreased from3 316.7 before invasion to 11 037.2 gC bits m−2 yr−1 after inva-ion. Likewise, ascendency dropped after invasion, from 3350.9 to926.3 gC bits m−2 yr−1 constituting a 13% decrease, as did totalverhead, from 9965.9 to 8110.9 gC bits m−2 yr−1 representing a9% decrease. However, these results were partially driven byhanges in TST (i.e., system growth), which scales capacity, ascen-
ency, and overhead. To remove the effects of TST scaling onapacity, we divided capacity by TST (the resultant quantity isquivalent to the Shannon flow diversity index; Ulanowicz, 1997).nscaled capacity showed a nominal, likely not significant, increaserom 5.0 to 5.1 bits, i.e., 2% increase in the potential for system
29.0 −1.827.5 6.7
development. To remove the effect of TST scaling on ascendencyand overhead, and look solely at the amount of organized anddisorganized ecosystem function, we divided both indices by thedevelopment capacity (Heymans and Baird, 2000). This calcula-tion removed the TST scaling through division and yielded theproportions that ascendency and overhead comprised of capacity(expressed as a percent of capacity in Table 4), which were termed“relative ascendency” and “relative overhead”, respectively. Ascen-dency constituted 25.2% of capacity before zebra mussel invasionand 26.5% of capacity after invasion. Overhead represented 74.8% ofcapacity before zebra mussel invasion and 73.5% after. These resultsindicate that the Oneida Lake food web is predominantly com-prised of disorganized flow (i.e., inefficient flow paths, as evidencedby a large overhead value), but may have become slightly moreorganized after zebra mussel invasion. Considering the ecosystemprocesses that contribute to system overhead (imports, exports,dissipations, and flow redundancies), these processes decreasedafter zebra mussel invasion between 1% and 22% except for flowredundancies, which increased almost 7%.
3.2. Cohesion analysis
Zebra mussel invasion altered food web structure in OneidaLake via a re-organization of interactions among taxa. The numberof structural subgroups increased from six significant subgroupsbefore zebra mussel invasion (p ≤ 0.001; odds ratio = 15.4; Table 5)to seven significant subgroups after invasion (p ≤ 0.001; oddsratio = 14.7; Table 6 ). Both networks contained three subgroups ofpredator–prey interactions: (1) a planktivorous food web (includ-ing planktivorous fish, zooplankton, and phytoplankton; subgroup1), (2) benthic-associated predators and prey (subgroup 2), and(3) planktivores and zooplankton prey (subgroup 3). Additionally,both networks contained three mostly fish subgroups: (1) centrar-chids (subgroup 4), (2) piscivores (subgroup 5), and (3) piscivoresand invertivores (subgroup 6). Finally, the seventh subgroup in thepost-invasion network contained mainly low biomass taxa frommultiple trophic levels (subgroup 7; post-invasion network only).We loosely named subgroups based on ecological descriptions thatrepresent the majority of taxa within each subgroup. The namesare meant as a tool to facilitate discussion of analyses; however,not all taxa fit the subgroup name.
Subgroup membership was statistically similar between the two
time stanzas (Z-score = 20.60, p ≤ 0.001) despite a large number ofchanges to subgroup membership. Of the seventy taxa present forboth time stanzas, twenty-three (33% of all taxa) changed sub-group membership after invasion: seventeen fish, five zooplankton,and one benthic invertebrate. The statistical test we used assessed
A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209 3201
Table 5Subgroups identified for the pre-zebra mussel invasion timestanza. We loosely named subgroups based on ecologicaldescriptions that represent the majority of taxa within eachsubgroup; however, not all taxa fit the subgroup name (e.g.,age-0 yellow perch and white perch were located in a gener-ally piscivorous subgroup, yet these taxa are not piscivores).
No. Subgroups
Subgroup 1: Planktivorous Food Web16 Gizzard Shad Age-037 Chironomids42 Oligochaetes46 Bosmina longirostris48 Ceriodaphnia quadrangular49 Chydorus sphaericus50 Daphnia galeata mendotae51 Daphnia pulicaria52 Daphnia retrocurva53 Diaphanosoma species54 Eubosmina coregoni58 Diacyclops thomasi60 Mesocyclops edax61 Epischura lacustris62 Leptodiaptomus minutes63 Skistodiaptomus oregonensis64 Nauplii65 Rotifers66 Blue-green Algae67 Diatoms68 Euglena69 Flagellates70 Golden Algae71 Green Algae
Subgroup 2: Benthic-associated5 Yellow Perch Age >38 White Perch Age >1
20 Channel Catfish22 Common Carp25 Freshwater Drum35 White Sucker36 Amphipods39 Insects40 Isopods41 Leeches43 Snails56 Leptodora kindtii72 Epiphytes73 Macrophytes74 Periphytes
Subgroup 3: Planktivores & Zooplankton17 Alewife21 aCisco26 Golden Shiners33 Trout Perch45 Alona species55 Sida crystalline57 Acanthocyclops vernalis
Subgroup 4: Centrarchids10 Black Crappie Age >111 Bluegill Age >112 Pumpkinseed Age >113 Rock Bass Age >123 Darters28 Log Perch29 Mottled Sculpin59 Ergasilus species
Subgroup 5: Piscivores2 Walleye Age >43 Walleye Age 1–34 Walleye Age-06 Yellow Perch Age 1–27 Yellow Perch Age-09 White Perch Age-0
19 Burbot30 Northern Pike
Table 5 (Continued)
No. Subgroups
32 Smallmouth Bass34 White Bass
Subgroup 6: Piscivores and Invertivores1 Cormorants
14 Centrarchids Age-015 Gizzard Shad Age >118 Brown Bullhead24 Emerald Shiners31 Red Horse Sucker
38 Clamsa Refers to taxa present in the pre-zebra mussel time stanza,but not the post-zebra mussel time stanza.
pair-wise (i.e., two taxa) co-membership in subgroups; i.e., whethertwo taxa present (or not present) in the same subgroup beforeinvasion were present (or not present) in the same subgroup afterinvasion. If pairs of taxa transferred subgroups together, or if pairsof taxa belonging to different subgroups remained in different sub-groups (even if one taxon changed to another subgroup), this testwould yield statistically similar membership, therefore potentiallyexplaining why the large percentage change we found in sub-group membership was not statistically significant. Zebra musselsinvaded the planktivorous food web subgroup (subgroup 1), thelargest subgroup (based on number of taxa) in both time stan-zas. Important subgroup shifts in the Oneida Lake fish communityinclude subadult (age 1–3 years) walleye, which moved to the sub-group (subgroup 6) with cormorants (Phalacrocorax auritus) afterinvasion and adult walleye (age 4 years and older), which shiftedto the subgroup containing zebra mussels (subgroup 1). Juvenile(age-0) yellow perch also transferred to the subgroup containingzebra mussels (subgroup 1) while subadult (age 1–2 years) yellowperch shifted to the benthic-associated subgroup (subgroup 2).
The cohesion analysis is visually summarized in crystallizedsociograms in Figs. 2 and 3. Before zebra mussel invasion, theplanktivorous food web (subgroup 1), benthic-associated (sub-group 2), and piscivore (subgroup 5) subgroups strongly interactedand played central roles within the food web (Fig. 2 Panel A).The remaining subgroups had more peripheral roles. After inva-sion, the subgroups became more closely related except for thesubgroup containing low biomass taxa (subgroup 7; Fig. 3 PanelA). The taxa within the planktivorous food web and benthic-associated subgroups strongly interact whereas the taxa in theremaining subgroups tend not to be as cohesive (Figs. 2 Panel Band 3 Panel B). Phytoplankton and zooplankton play central roleswithin the planktivorous food web subgroup (subgroup 1), whilebenthic invertebrates occupy central positions within the benthic-associated subgroup (subgroup 2).
3.3. Ecological network analysis—grouped network
Below, we analyze the functional characteristics of the subgroupthat zebra mussels invaded, the planktivorous food web subgroup(subgroup 1; hereafter referred to as the zebra mussel subgroup),which is the largest subgroup before and after invasion in terms ofnumber of taxa and TST (Table 7).
3.3.1. Input/output analysisWhen we quantified the direct and indirect effects of zebra
mussels on their subgroup (via the IMPACTS analysis), we found
that zebra mussels negatively affected all their subgroup members(Fig. 4). Our full food web analysis also indicated that zebra musselsnegatively affected these taxa (Fig. 1); however, the magnitudes ofthe effects differed slightly in the subgroup analysis. The largest dif-ference involved zebra mussel influence upon themselves (taxon
3202 A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209
Table 6Subgroups identified for the post-zebra mussel invasiontime stanza. We loosely named subgroups based on ecolog-ical descriptions that represent the majority of taxa withineach subgroup; however, not all taxa fit the subgroup name.
No. Subgroups
Subgroup 1: Planktivorous Food Web2 aWalleye Age >47 aYellow Perch Age-09 aWhite Perch Age-0
16 Gizzard Shad Age-024 aEmerald Shiners37 Chironomids42 Oligochaetes44 bZebra Mussels46 Bosmina longirostris49 Chydorus sphaericus50 Daphnia galeata mendotae51 Daphnia pulicaria53 Diaphanosoma species54 Eubosmina coregoni56 aLeptodora kindtii57 aAcanthocyclops vernalis58 Diacyclops thomasi60 Mesocyclops edax61 Epischura lacustris62 Leptodiaptomus minutus63 Skistodiaptomus oregonensis64 Nauplii65 Rotifers66 Blue-green Algae67 Diatoms68 Euglena69 Flagellates70 Golden Algae71 Green Algae
Subgroup 2: Benthic-associated5 Yellow Perch Age >36 aYellow Perch Age 1–28 White Perch Age >1
20 Channel Catfish22 Common Carp25 Freshwater Drum32 aSmallmouth Bass35 White Sucker36 Amphipods39 Insects40 Isopods41 Leeches43 Snails72 Epiphytes73 Macrophytes74 Periphytes
Subgroup 3: Planktivores & Zooplankton14 aCentrarchids Age-026 Golden Shiners28 aLog Perch38 aClams45 Alona species
Subgroup 4: Centrarchids11 aBluegill Age >112 Pumpkinseed Age >113 Rock Bass Age >119 aBurbot34 aWhite Bass59 Ergasilus species
Subgroup 5: Piscivores4 Walleye Age-0
23 aDarters29 aMottled Sculpin30 Northern Pike33 aTrout Perch
Subgroup 6: Piscivores & Invertivores1 Cormorants3 bWalleye Age 1–3
10 aBlack Crappie Age >1
Table 6 (Continued)
No. Subgroups
15 Gizzard Shad Age >118 Brown Bullhead31 Red Horse Sucker
Subgroup 7: Miscellaneous Low Biomass17 aAlewife27 bLake Sturgeon47 bCamptocercus harpae48 aCeriodaphnia quadrangular52 aDaphnia retrocurva55 aSida crystalline
a
Refers to taxa that changed subgroup membership afterzebra mussel invasion.b Refers to taxa present in the post-zebra mussel timestanza, but not the pre-zebra mussel time stanza.
44). The IMPACTS value changed from −0.33 in the full food web(Fig. 1) to −0.11 in the subgroup, representing a 67% change. Poten-tially we see a lessened effect of zebra mussels upon themselvesat the subgroup level because detrital groups, including pelagicdetritus and DOC, were not included in the analysis. Zebra musselsprey heavily on detrital groups, negatively affecting themselves viadepletion of their food supply.
Although we cannot directly conduct an IMPACTS analysis ofzebra mussels on the other six subgroups (to conduct the analy-sis, zebra mussels would need to be present in the subgroup) wecan indirectly detect zebra mussel influence in those subgroups byconsidering effects on individual taxa from our full food web anal-ysis (Fig. 1). Unlike the (wholly negative) zebra mussel influence ontaxa in their subgroup (subgroup 1), zebra mussel effects on taxa inother subgroups were mixed with no consistent pattern betweensubgroups. The majority of negative effects were confined to thezebra mussel subgroup while some subgroups, including subgroups2, 4, 5, and 6, exhibited net positive effects of zebra mussels
3.3.2. Lindeman trophic analysisThe Lindeman trophic analysis identified six trophic levels in the
zebra mussel subgroup before invasion and nine trophic levels afterinvasion (Figs. 5 and 6). Before invasion, age-0 gizzard shad werethe top predator in the subgroup and fed up to the sixth trophiclevel (the other subgroup members were benthic invertebrates,zooplankton, and phytoplankton and it was therefore possible fora young fish to be a top predator); after invasion and with the addi-tion of multiple fish to this subgroup, adult walleye were the toppredator and fed at the ninth trophic level. Zebra mussels fed atthe second trophic level; consequential to their invasion, trophicflow efficiency increased over 10% at trophic level II despite res-pirative losses more than tripling and flow loss to detritus almostdoubling. Flow efficiencies at trophic levels IV and higher increasedsubsequent to invasion.
Total respiration and production decreased in the zebra mus-sel subgroup after invasion. Flow loss due to respiration was424.1 gC m−2 yr−1 before invasion and 396.2 gC m−2 yr−1 after inva-sion (6% decrease), while production was 484.7 gC m−2 yr−1 beforeinvasion and 401.0 gC m−2 yr−1 after invasion (17% decrease). Sub-group production constituted 78% of the full food web productionbefore invasion, but only 68% after invasion. Furthermore, subgrouprespiration contributed 42% to the full food web before invasion and51% after invasion.
3.3.3. Ecosystem indicesSimilar to the analysis of the full food web, TST and development
capacity fell in the zebra mussel subgroup after invasion (Table 8).Before invasion, TST and capacity were 1705.2 gC m−2 yr−1 and7574.4 gC bits m−2 yr−1, respectively; after invasion, throughput

A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209 3203
F nza. UF ionshin B: Plar l subg
a6cUdsddfdfdFw6tto
4
iocObps
ig. 2. Panel A: Crystallized sociogram for the pre-zebra mussel invasion time starank, 1996). Subgroups 1 through 6 are plotted with the direction of feeding relatot vice versa); thickness of arrows indicates weight of feeding relationships. Panelepresent general groupings of taxa. Subgroup numbers are to the upper right of al
nd capacity declined to 1415.8 gC m−2 yr−1 (a 17% decrease) and651.5 gC bits m−2 yr−1 (a 12% decrease), respectively. Unscaledapacity increased after invasion, from 4.4 to 4.7 bits, a 7% increase.nlike the full food web ecosystem analysis, relative ascendencyecreased over 3% while relative overhead increased by 1% in theubgroup after invasion. Relative overhead on imports and exportsecreased by 4% and 28%, respectively, while relative flow redun-ancies increased by 54% after invasion. These results parallel theull food web analysis, although the increase in relative flow redun-ancies was substantially greater for the subgroup than the fullood web (7% increase). However, unlike the full food web, relativeissipative overhead increased by 4% in the subgroup after invasion.inally, the zebra mussel subgroup constituted 64% of the full foodeb TST and 57% of capacity before invasion, and 66% of TST and
0% of capacity after invasion. Essentially, subgroup size and poten-ial for development increased after zebra mussel invasion relativeo the full food web, making this subgroup a more important partf the food web.
. Discussion
Our results indicate that zebra mussels exert a far-reachingnfluence on the Oneida Lake food web, altering the membershipf food web subgroups (question 1) and changing the magnitude of
arbon flows within the full food web and subgroups (question 2).ur analyses support the observation that zebra mussels lead to theenthification of their invaded ecosystems by shunting carbon fromelagic to benthic pathways and promoting benthic-associatedpecies; moreover, this research is the first test of benthificationnits are relative distances based on the inverse of the density of interactions (seeps represented by arrows (e.g., subgroup 6 consumes members of subgroup 1, butcement of taxa within subgroups. Circles indicate subgroup boundaries and colors
roups.
using ecosystem level measures. Furthermore, this research is thefirst application of CA, a social network analysis, in conjunction withENA. Unions of methods from ecology and sociology (e.g., Krause etal., 2003; Zhao and Frank, 2003) reveal there is value found in theconvergence of these disciplines, especially when applied to foodwebs. While SNA largely focuses on structural analyses (Mayhew,1980), ENA is a functional analysis (Ulanowicz, 1986). Yet, Johnsonet al. (2001) affirm that ENA and SNA are markedly similar math-ematically and conceptually. The merging of these two methodsto understand the integration of ecosystem structure and function(as called for by Jørgensen, 2007) seems a natural extension. Webegin our discussion below with CA, and then place the structuralfindings within the context of the full food web and subgroup ENA,emphasizing the contribution to our knowledge of ecosystems.
4.1. Cohesion analysis—structure
Almost one-third of taxa changed subgroups after invasion.Although the difference in subgroup membership between thepre- and post-zebra mussel time stanzas is not statistically signifi-cant (despite large membership shifts), we believe these shifts areecologically significant based on the restructuring of carbon flowpaths within subgroups. The zebra mussel subgroup underwentthe greatest change, increasing membership from twenty-four
to twenty-nine taxa, including the addition of zebra mussels aswell as multiple fish and zooplankton. These shifts broadenedthe once pelagic-based subgroup by increasing food chain lengthand creating important links via benthic pathways. Although thissubgroup experienced the greatest change after invasion, the
3204 A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209
Fig. 3. Crystallized sociogram for the post-zebra mussel inv
Table 7Throughput of subgroups before and after zebra mussel invasion (only six subgroupswere identified before zebra mussel invasion).
Subgroup Pre-Zebra Mussels Throughput(gC m−2 yr−1)
Post-Zebra MusselsThroughput(gC m−2 yr−1)
1 1705.2 1415.82 462.9 452.13 0.1 0.04 0.1 0.25 3.9 0.26 0.2 0.57 Not Present 0.3
Fig. 4. Zebra mussel impacts on subgroup members. The scale is relative: impacts aboveare negative impacts of zebra mussels.
asion time stanza. See Fig. 2 for description of panels.
benthic-associated subgroup also increased membership by onefish group, while all other subgroups lost members. These subgroupshifts suggest that benthic pathways gained importance as a resultof zebra mussel invasion.
Zebra mussel invasion in Oneida Lake potentially affects impor-tant fishery related species via shifts in subgroup membershipresulting from invasion. Life stages of walleye and yellow perchcomprise a large proportion of the subgroup shifts resulting from
zebra mussel invasion and, consequently, the dynamics of subgroupinteractions suggest these species may be vulnerable to the effectsof zebra mussels. Theoretical work (Simon, 1962; McCann, 2000;McCann et al., 2005) suggests that within subgroups, the strongthe zero line are positive impacts of zebra mussels and impacts below the zero line

A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209 3205
F 1) int cy ofi s enteb elec
isaoegat
lppdc(ossmw
F
ig. 5. Lindeman trophic spine for the planktivorous food web subgroup (subgrouphe integer trophic levels; the number within each trophic box is the percent efficienn the grazer food chain, arrows leaving the top of trophic boxes are exports, arrowoxes are flows to detritus, represented by the detrital box. The ground symbol from
nteractions among members transfer the effect of a disturbance,uch as exotic species invasion, throughout the subgroup even if notll members are directly affected by the event. Conversely, taxa inther subgroups are either weakly impacted or not impacted as theffects must be transferred through the weak interactions betweenroups. Thus taxa within the subgroup invaded by zebra musselsre especially vulnerable to effects of this exotic species, whereasaxa in other subgroups are more sheltered from effects.
Common subgroup membership of zebra mussels and variousife stages of yellow perch and walleye offers structural under-innings for potential zebra mussel influence on juvenile yellowerch growth rate (Mayer et al., 2000) and adult walleye abun-ance (Rutherford et al., 1999). Additionally, due to the tightoupling of walleye and yellow perch populations in Oneida LakeRose et al., 1999; Rudstam et al., 2004), within-subgroup influence
f zebra mussels on these species can cascade to their corre-ponding life stages in other subgroups, despite the buffering thatubgroup structure offers. Yet, despite this mechanism for zebraussels to affect walleye and yellow perch life stages, subadultalleye moved to the subgroup containing piscivore and inverti-ig. 6. Lindeman trophic spine for the planktivorous food web subgroup (subgroup 1) in t
the pre-zebra mussel invasion time stanza. Boxes with Roman numerals representthat trophic level at processing carbon. Arrows between the trophic levels are flowsring the top of trophic boxes are imports, and arrows leaving the bottom of trophictronic circuitry represents flow loss due to respiration. All flows are in gC m−2 yr−1.
vore taxa (subgroup 6), including cormorants, after zebra musselinvasion. Common subgroup membership gives structural supportfor Oneida Lake diet studies that suggest cormorants exert strongpredatory pressure on subadult walleye (VanDeValk et al., 2002;Rudstam et al., 2004). Since subadult walleye compose one-quarterof cormorant diet by weight (Appendix 2) and cormorant biomassmore than quadrupled from the 1980s to the 1990s (Appendix 1),we feel it is likely that cormorants, rather than zebra mussels, influ-enced the shift of subadult walleye, corroborating Rudstam et al.(2004). For the same reasons given above for zebra mussels, com-mon subgroup membership of cormorants and subadult walleyecan lead to cascading effects of cormorants on other life stages ofwalleye and yellow perch that are found in other subgroups.
Structural analysis of the effects of zebra mussel invasionsuggests that the food web structure of Oneida Lake is potentially
determined by physical system structure. In both time stanzas,one large subgroup dominates food web structure while multiplesmaller subgroups play less significant roles. This result differsfrom CA findings on the Chesapeake Bay by Krause et al. (2003)who identify two subgroups (one benthic, the other pelagic)he post-zebra mussel invasion time stanza. See Fig. 5 for a description of the figure.

3206 A.L.J. Miehls et al. / Ecological Modelling 220 (2009) 3194–3209
Table 8Ecosystem indices for the zebra mussel subgroup before and after zebra mussel invasiona.
Index Pre-Zebra Mussels Value Post-Zebra Mussels Value % Difference Value
Total system throughput (gC m−2 yr−1) 1705.2 1415.8 −17.0Development capacity (gC bits m−2 yr−1) 7574.4 6651.5 −12.2Ascendency (gC bits m−2 yr−1) 1847.8 1567.0 −15.2Total overhead (gC bits m−2 yr−1) 5726.7 5084.5 −11.2
Overhead on imports (gC bits m−2 yr−1) 1836.3 1552.4 −15.5Overhead on exports (gC bits m−2 yr−1) 1410.4 890.8 −36.8Dissipative overhead (gC bits m−2 yr−1) 1628.1 1492.5 −8.3Redundancy (gC bits m−2 yr−1) 851.9 1148.8 34.8
Unscaled capacity (bits) 4.4 4.7 6.8Ascendency/capacity (%) 24.4 23.6 −3.4Total overhead/capacity (%) 75.6 76.4 1.1
Overhead on imports/capacity (%) 24.2 23.3 −3.7
cbmwlfssebicCLmiitps
4
tubplUtthtl2t
ztctacasdm
Overhead on exports/capacity (%) 18.6Dissipative overhead/capacity (%) 21.5Redundancy/capacity (%) 11.2
a The percent difference was calculated as: %Difference = Post − Pre/Pre × 100%.
o-dominating food web structure. Perhaps structure differsetween the systems because Oneida Lake is a shallow, well-ixed ecosystem with only brief periods of thermal stratification,hile the Chesapeake Bay maintains thermal stratification from
ate spring to early fall (Bidle and Fletcher, 1995), creating distinctood web structure between the thermal regions. These findingsuggest food web structure is, at least in part, related to physicalystem structure. Recently, the littoral zone of Oneida Lake hasxpanded farther offshore due to increased water clarity; it wille interesting if future subgroup structure reflects this change
n physical structure via an increase in importance of subgroupsontaining littoral taxa (e.g., subgroups 2 and 4). Similar to thehesapeake Bay analysis, bivalve taxa (zebra mussels in Oneidaake; clams and oysters in the Chesapeake Bay) share subgroupembership with their pelagic prey. The majority of carbon flow
n the zebra mussel subgroup originates in phytoplankton and ismmediately routed through zebra mussels, making these musselshe main flow nexus in the system linking pelagic and benthicathways. Our study suggests this coupling of pelagic and benthictructure is a central influence of zebra mussels in Oneida Lake.
.2. Ecological network analysis—function
Zebra mussel invasion has large effects on ecosystem func-ion in Oneida Lake. Analysis of these effects not only helps usnderstand the ecology of Oneida Lake post-zebra mussel invasion,ut also contributes to our general understanding of ecosystemerturbation by exotic species. Below, we discuss our results in
ight of previous theoretical research on ecosystem perturbation.lanowicz (1996) makes predictions for ecosystem level response
o perturbation by drawing from Odum’s (1969) theory of ecosys-em development. Ulanowicz suggests that perturbed ecosystemsave less efficient and shorter trophic chains, decreased systemhroughput, and decreased food web organization. Our analysisargely supports Ulanowicz (1996) and affirms research question: zebra mussels alter the magnitude of carbon flow within ecosys-ems.
IMPACTS analysis (i.e., input/output analysis) indicates thatebra mussels have an overwhelmingly negative effect on food webaxa, especially subgroup members. The majority of large effects areontained in the zebra mussel subgroup, validating the supposi-ion that subgroup members exert a stronger influence over onenother than non-subgroup members. Moreover, regarding fish
ommunities, our results (Fig. 1) corroborate findings of Irwin etl. (submitted for publication) who indicate that, after zebra mus-el invasion, biomass of walleye, yellow perch, and white perchecreased relative to long-term expectations (based on pre-zebraussel biomass) while biomass of gizzard shad, pumpkinseeds,13.4 −28.122.4 4.417.3 53.6
channel catfish, brown bullhead, freshwater drum, white sucker,and red horse sucker increased relative to long-term expectations.However, Irwin et al. (submitted for publication) find that small-mouth bass biomass is higher than expected after zebra musselinvasion given long-term trends whereas our analysis suggests asmall, but negative effect of zebra mussels on smallmouth bass.Potentially this discrepancy is due to non-trophic effects of zebramussels: zebra mussels increase water clarity in Oneida Lake (Idrisiet al., 2001), which may positively affect smallmouth bass throughincreased prey detection ability and an increase in littoral zonehabitat. Perhaps these positive, non-trophic effects out-weigh thenegative trophic effect of zebra mussels.
When interpreting results of the IMPACTS analysis, it is impor-tant to bear in mind that this analysis only requires use of thepost-zebra mussel invasion network (1992–2002). The IMPACTSresults explicate how direct and indirect trophic effects of zebramussels combine to influence other species in the food web. Neg-ative effects of zebra mussels do not necessarily imply realizedeffects on, e.g., biomass or production, of certain species in theOneida Lake food web. Instead, the IMPACTS analysis mechanis-tically describes paths of feeding interactions through which zebramussels affect other species. For example, we can consider thethroughput of subadult yellow perch, juvenile yellow perch, andjuvenile walleye before and after invasion. Although the IMPACTSanalysis indicates that zebra mussels negatively influence theselife stages, throughput of these life stages increased after inva-sion. Perhaps these opposing results are due to the relaxation ofother deleterious pressures on these taxa that were present in thepre-invasion time stanza (i.e., the food web reorganization thatoccurred after zebra mussel invasion could yield a net positiveor neutral effect on various walleye and yellow perch life stages,despite the IMPACTS analysis suggesting negative effects on thesetaxa). This conclusion is supported by other research investigatingthe effects of zebra mussels on walleye and yellow perch. Idrisi etal. (2001) does not find evidence that zebra mussels have a neg-ative effect on young-of-year yellow perch growth, biomass, orproduction and Mayer et al. (2000) likewise does not find nega-tive effects of zebra mussels on young-of-year and adult yellowperch in Oneida Lake. Moreover, Rudstam et al. (2004) suggests thatrecent declines in adult walleye and yellow perch are more likelydue to cormorant predation than zebra mussel invasion; Irwinet al. (2008) also conclude that cormorant predation was impor-tant for the observed walleye decline. Therefore, the results of the
IMPACTS analysis in general, and specifically in relation to walleyeand yellow perch dynamics in Oneida Lake, should be consideredin conjunction with other functional analyses to understand howfeeding interactions contribute to realized changes in the OneidaLake food web.
Mode
Ohitmwttiwwwtcc
siiTaWpbcciIdcwwiAmipi1sriz
pasTd(isavtdweolflc
nz
A.L.J. Miehls et al. / Ecological
The zebra mussel subgroup is the most important subgroup inneida Lake, dominating both food web structure and function andaving large effects on carbon flow. After invasion, the trophic chain
ncreased by three levels and the subgroup garnered more carbonhroughput relative to the full food web. Additionally, the develop-
ent capacity of the subgroup increased compared to the full foodeb. These results suggest this subgroup not only increased in func-
ional size, but also became a more complex and integrated part ofhe food web after invasion. However, despite gaining functionalmportance, the subgroup contributes proportionally more to food
eb flow loss after invasion. Respirative flow losses increased (9%)hile production decreased (almost 10%) relative to the full foodeb. Thus, not only do we see evidence of an energy shunt to ben-
hic pathways after zebra mussel invasion, but also indication ofarbon flow loss to dissipative paths and decreased food web effi-iency.
The Lindeman trophic analysis indicates that zebra mussel inva-ion changed the functional relationships between trophic levelsn Oneida Lake. After invasion, flow efficiency for trophic level Increased 5% and efficiency at trophic level II increased over 10%.he driver of increased efficiency at the first trophic level is prob-bly the decrease in phytoplankton biomass following invasion.ith less phytoplankton available for consumption, a greater pro-
ortion of biomass passes into the grazing food chain, with lessiomass loss to detrital pathways. Similar to trophic level I, the effi-iency increase at the second trophic level may be due to increasedonsumption of zooplankton proportional to their biomass. It isnteresting that there is an increase in efficiency at trophic levelI despite high respirative loss of zebra mussels relative to pro-uction (Appendix 2). In light of the subgroup results, we canonjecture trophic efficiency in the full food web even thoughe could not create the Lindeman trophic spine for the full foodeb. After invasion, phytoplankton biomass decreased, potentially
ncreasing flow efficiency at trophic level I, similar to the subgroup.dditionally, non-pelagic sources of primary production, such asacrophytes and detritus (considered primary sources of carbon
n ENA), gained functional importance. Before invasion, macro-hytes produced 8.0 gC m−2 yr−1, compared to 9.0 gC m−2 yr−1 after
nvasion, a 13% increase. Detrital groups combined produced1.6 gC m−2 yr−1 before invasion and 65.1 gC m−2 yr−1 after inva-ion. Despite a small absolute increase in carbon flow, this changeepresents an over 460% increase in detrital use, illustrating themportance of alternate sources of carbon, especially benthic, afterebra mussel invasion.
Considering the indices of ecosystem function, we were sur-rised that full food web organization, measured by relativescendency, increased and food web disorganization, as mea-ured by relative overhead, decreased after zebra mussel invasion.hese findings contradict food web theory that suggests ascen-ency decreases, and overhead increases, following perturbationUlanowicz, 1996). To understand why system overhead shouldncrease, it is helpful to consider the components of overhead: dis-ipative flow loss due to respiration, flow redundancies, imports to,nd exports from the system. Although these processes are disad-antageous during benign conditions, overhead is advantageous ifhe system is perturbed in a novel fashion. Bondavalli et al. (2000)escribe overhead as a “strength in reserve of degrees of freedomhich the system can call upon to adapt to a new threat.” In gen-
ral, overhead increases after perturbation due to reorganizationf flow structure. For instance, as an exotic species becomes estab-ished in a system, the ecosystem may shift from a stable state to a
ux system, with concomitant flow reorganization, interruption ofarbon cycling, and even loss of native species.Perhaps the Oneida Lake food web exhibits increased orga-ization due to the considerable flow asymmetry present afterebra mussel invasion. Zebra mussels conduct 10% of food web
lling 220 (2009) 3194–3209 3207
flow, potentially increasing organization by focusing flow (andincreasing AMI). However, Ulanowicz’s (1996) perturbation the-ory holds for the analysis of the zebra mussel subgroup, whichshould contain the most pronounced influence of zebra musselsin the food web (according to food web stability theory). Rela-tive ascendency decreased, while relative overhead increased inthe subgroup, suggesting that the subgroup underwent reorgani-zation of carbon flow as it accommodated zebra mussel presence(as Ulanowicz’s theory predicts). This finding presents compellingevidence for investigating food web function at multiple structurallevels. Despite differences in organization between the subgroupand full food web (unscaled) development capacity increased atboth levels after invasion. The rise in capacity indicates that zebramussels add a layer of complexity to food web function. Complexityimparts multiple benefits to ecosystems, including more diversityof interconnections which allows alternate routes for taxa to obtainenergy in the face of ecological change (Pérez-Espana and Arreguín-Sánchez, 1999). Consequently, despite resulting in several negativeeffects on Oneida Lake, including decreased food web efficiencyand production, zebra mussel invasion imparts some benefits tothe ecosystem.
A limitation of ENA is that variability in carbon flow estimatesis not incorporated into the analysis. As a result, we cannot discernstatistical significance in food web change with ENA. However, inthe case of the whole system analysis, ecosystem indices are basedon log-scaled measures (Ulanowicz, 1986) and thus even small per-cent changes represent larger ecological changes. Another potentiallimitation of our research is the scope of taxa represented in ourfood web network. Although we advocate the use of high reso-lution data for all food web taxa (including microbial pathways;Allesina et al., 2005; Jørgensen, 2007) to gain a holistic under-standing of ecosystem structure and function, these data were notalways available for Oneida Lake. Therefore, we acknowledge thatour aggregation scheme and choice of taxa may affect our results.However, our food web networks, with over seventy taxa and lifestages for each time stanza, represent one of the most in-depthtreatises of food webs to date. A final limitation in this research isthe need to mass balance our networks for ENA. After constructinga food web network, taxa rarely meet a condition of mass balance,as was the case in this analysis. Balancing introduces a degree ofuncertainty into the analysis, thus careful choice of methods isimperative. We acknowledge that our balancing routines may affectour results; however, we were careful to choose parsimonious bal-ancing methods that yielded ecologically realistic networks.
An assumption of our analysis is that any differences observedin food web structure and function between time stanzas are dueto zebra mussel invasion. Although Oneida Lake has been bufferedfrom most exotic species introductions that plague the Great Lakes(e.g., the spiny (Bythotrephes longimanus) and fish-hook (Cercopagispengoi) water fleas, the round goby (Neogobius melanostomus), and,until recently, quagga mussels), zebra mussel invasion is not theonly ecological change in recent years. For example, white perchincreased in abundance, a lake sturgeon stocking program began in1995, and gizzard shad declined in 1992. Additionally, as describedabove, cormorant biomass increased independent of zebra musselintroduction. Effects of direct predation by cormorants on wall-eye and yellow perch are strong in Oneida Lake, and therefore,our interpretation of zebra mussel effects on certain species isconfounded by other ecological changes. However, although theseevents may cause food web alteration, we believe the effect ofzebra mussel invasion, when assessed over the entire Oneida Lake
food web, is substantial even in the presence of these other eco-logical changes. Zebra mussels garner over 10% of total systemflow and constitute over two-thirds of Oneida Lake’s living biomass(gC m−2). These numbers alone suggest zebra mussel introductionis undoubtedly one of the most significant changes to Oneida Lake’s
3 l Mode
esLsoIae1
wacc(dittaLsa
zehgobdbtlaaiO
A
sWCRUachGoc
A
t
R
A
A
A
208 A.L.J. Miehls et al. / Ecologica
cology in recent years. Given the limitations of ENA, but con-idering the overwhelming presence of zebra mussels in Oneidaake, we are confident in the robustness of our results and conclu-ions. Moreover, our research corroborates other work conductedn Oneida Lake (e.g., Rutherford et al., 1999; Mayer et al., 2000;drisi et al., 2001; Rudstam et al., 2004; Zhu et al., 2006; Irwin etl., submitted for publication) as well as supports research on theffects of zebra mussel invasion in other systems (e.g., MacIsaac,996; Nalepa et al., 1999; Strayer et al., 2004).
A large body of research indicates that aquatic ecosystemsorldwide are in the midst of large-scale ecological alteration. To
ddress ecosystem level change, we need quantifiable, objectiveriteria at the whole-system level (Westra et al., 2000), espe-ially with the contemporary emphasis on ecosystem managementChristensen et al., 1996). Network analysis is a valuable tool foreciphering effects of perturbation at the ecosystem level because
t offers a quantitative depiction of food web structure and func-ion, inclusive of all food web taxa (barring data limitations). Inhis research, our use of network analyses from multiple disciplinesllows us to gain a holistic understanding of the ecology of Oneidaake, as well as a general understanding of exotic species inva-ion as an ecosystem perturbation, and of how ecological networksdapt to changed conditions (as called for by Jørgensen, 2007).
In conclusion, our study elucidates ecosystem level effects ofebra mussel invasion in Oneida Lake. Also, this research givescosystem-level support for the work of Mills et al. (2003) whoypothesize that zebra mussels lead to the benthification and reor-anization of their invaded ecosystems. Considering the attributesf benthification that Mills et al. (2003) suggest (i.e., growth ofenthic algae and submersed vegetation, colonization of bottom-welling organisms, and promotion of fish communities that useenthic habitat), we find that zebra mussels promote the use of ben-hic sources of production (including macrophytes and detritus),ink formerly pelagic subgroups to benthic pathways, and positivelyffect benthic fish species. We suggest network analysis methods bepplied to other ecosystems that have experienced exotic speciesnvasion to determine if the results presented here are unique toneida Lake, or ubiquitous across invaded ecosystems.
cknowledgements
We thank two anonymous reviewers, Edward Mills, Lars Rud-tam, Stefano Allesina, Andrew Fayram, Scott Miehls, and Tyler
agner for helpful comments, the Bay of Quinte–Oneida Lakeomparative Modelling Project Workgroup, Edward Mills, Larsudstam, Brian Irwin, Kristen Holeck and the staff at the Cornellniversity Biological Field Station for data contributions and guid-nce, Anne Clites, Timothy Hunter, and Marten Koops for theiromputational and programming assistance, and Cathy Darnell forer aid on graphics. This work was supported by a grant from thereat Lakes Fishery Commission. This is contribution number 1521f the NOAA Great Lakes Environmental Research Laboratory andontribution number 266 from the Cornell Biological Field Station.
ppendices. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.ecolmodel.2009.07.020.
eferences
barca-Arenas, L.G., Ulanowicz, R.E., 2002. The effects of taxonomic aggregation onnetwork analysis. Ecol. Model. 149, 285–296.
llesina, S., Bondavalli, C., 2003. Steady state of ecosystem flow networks: a com-parison between balancing procedures. Ecol. Model. 154, 221–229.
llesina, S., Bondavalli, C., Scharler, U.M., 2005. The consequences of the aggregationof detritus pools in ecological networks. Ecol. Model. 189, 221–232.
lling 220 (2009) 3194–3209
Bailey, R.C., Grapentine, L., Stewart, T.J., Schaner, T., Chase, M.E., Mitchell, J.S., Coulas,R.A., 1999. Dreissenidae in Lake Ontario: impact assessment at the whole lakeand Bay of Quinte spatial scales. J. Great Lakes Res. 25, 482–491.
Baxter, C.V., Fausch, K.D., Murakami, M., Chapman, P.L., 2004. Fish invasion restruc-tures stream and forest food webs by interrupting reciprocal prey subsidies.Ecology 85, 2656–2663.
Beekey, M.A., McCabe, D.J., Marsden, J.E., 2004. Zebra mussel colonisation of softsediments facilitates invertebrate communities. Fresh. Biol. 49, 535–545.
Bidle, K.D., Fletcher, M., 1995. Comparison of free-living and particle-associated bac-terial communities in the Chesapeake Bay by stable low-molecular-weight RNAanalysis. Appl. Environ. Microbiol. 61, 944–952.
Bondavalli, C., Bodini, A., Rossetti, G., Allesina, S., 2006. Detecting stress at the wholeecosystem level: The case of a mountain lake (Lake Santo Italy). Ecosystems 9,768–787.
Bondavalli, C., Ulanowicz, R.E., Bodini, A., 2000. Insights into the processing of car-bon in the South Florida Cypress Wetlands: a whole-ecosystem approach usingnetwork analysis. J. Biogeogr. 27, 697–710.
Christensen, N.L., Bartuska, A.M., Brown, J.H., Carpenter, S., D’Antonio, C., Francis,R., Franklin, J.F., MacMahon, J.A., Noss, R.F., Parsons, D.J., Peterson, C.H., Turner,M.G., Woodmansee, R.G., 1996. The report of the Ecological Society of Amer-ica Committee on the scientific basis for ecosystem management. Ecol. Appl. 6,665–691.
Christensen, V., Pauly, D., 1992. ECOPATH II—A software or balancing steady-stateecosystem models and calculating network characteristics. Ecol. Model. 61,169–185.
Crooks, J.A., 2002. Characterizing ecosystem-level consequences of biological inva-sions: The role of ecosystem engineers. Oikos 97, 153–166.
Dettmers, J.M., Raffenberg, M.J., Weis, A.K., 2003. Exploring zooplankton changesin southern Lake Michigan: Implications for yellow perch recruitment. J. GreatLakes Res. 29, 355–364.
Drake, J.M., Bossenbroek, J.M., 2004. The potential distribution of zebra mussels inthe United States. BioScience 54, 931–941.
Fayram, A.H., Hansen, M.J., Ehlinger, T.J., 2006. Characterizing changes in maturity oflakes resulting from supplementation of walleye populations. Ecol. Model. 197,103–115.
Frank, K.A., 1995. Identifying cohesive subgroups. Soc. Networks 17, 27–56.Frank, K.A., 1996. Mapping interactions within and between cohesive subgroups.
Soc. Networks 18, 93–119.Hannon, B., 1973. The structure of ecosystems. J. Theor. Biol. 41, 535–546.Hansen, A.-M., Hairston Jr., N.G., 1998. Food limitation in a wild cyclopoid cope-
pod population: Direct and indirect life history responses. Oecologia 115,320–330.
Heymans, J.J., Baird, D., 2000. Network analysis of the northern Benguela ecosystemby means of NETWRK and ECOPATH. Ecol. Model. 131, 97–119.
Heymans, J.J., Ulanowicz, R.E., Bondavalli, C., 2002. Network analysis of the SouthFlorida Everglades graminoid marshes and comparison with nearby cypressecosystems. Ecol. Model. 149, 5–23.
Hubert, L., 1987. Assignment Methods in Combinatorial Data Analysis. MarcelDekker, New York, NY, USA, p. 326.
Idrisi, N., Mills, E.L., Rudstam, L.G., Stewart, D.J., 2001. Impact of zebra mussels (Dreis-sena polymorpha) on the pelagic lower trophic levels of Oneida Lake New York.Can. J. Fish. Aquat. Sci. 58, 1430–1441.
Irwin, B.J., 2006. Evaluating effects of ecological change on important sport fishes inOneida Lake, New York. PhD Dissertation. Cornell University, Ithaca, NY, USA.
Irwin, B.J., Rudstam, L.G., Jackson, J.R., VanDeValk, A.J., Forney, J.L. Long-term trendsin the fish community of Oneida Lake: analysis of the effect of the zebra musselintroduction. In: Mills, E.L., Rudstam, L.G., Jackson, J.R., Stewart, D.J. (Eds.), OneidaLake: Long-term Dynamics of a Managed Ecosystem and its Fisheries, submittedfor publication.
Irwin, B.J., Treska, T.J., Rudstam, L.G., Sullivan, P.J., Jackson, J.R., VanDeValk, A.J., For-ney, J.L., 2008. Estimating walleye (Sander vitreus) density, gear catchability, andmortality using three fishery-independent data sets for Oneida Lake New York.Can. J. Fish. Aquat. Sci. 65, 1366–1378.
Jackson, J.R., VanDeValk, A.J., Brooking, T.E., vanKeeken, O.A., Rudstam, L.G., 2002.Growth and feeding dynamics of lake sturgeon, Acipenser fulvescens, in OneidaLake New York: Results from the first five years of a restoration program. J. Appl.Ichthyol. 18, 439–443.
Johannsson, O.E., Dermott, R., Graham, D.M., Dahl, J.A., Millard, E.S., Myles, D.D.,LeBlanc, J., 2000. Benthic and pelagic secondary production in Lake Erie after theinvasion of Dreissena spp. with implications for fish production. J. Great LakesRes. 26, 31–54.
Johnson, J.C., Borgatti, S.P., Luczkovich, J.J., Everett, M.G., 2001. Network role analysisin the study of food webs: An application of regular role coloration. J. Soc. Struct.2, 1–15.
Jones, C.G., Lawton, J.H., Shachak, M., 1994. Organisms as ecosystem engineers. Oikos69, 373–386.
Jørgensen, S.E., 2007. Ecological network theory. Ecol. Model. 208, 1–2.Koops, M.A., Irwin, B.J., MacNeil, J.E., Millard, E.S., Mills, E.L., 2006. Comparative
modelling of the ecosystem impacts of exotic invertebrates and productivitychanges on fisheries in the Bay of Quinte and Oneida Lake. Great Lakes Fishery
Commission Project Completion Report, Ann Arbor, MI, USA.Krause, A.E., Frank, K.A., Mason, D.M., Ulanowicz, R.E., Taylor, W.W., 2003. Compart-ments revealed in food-web structure. Nature 426, 282–285.
Lindeman, R.L., 1942. The trophic-dynamic aspect of ecology. Ecology 23, 399–418.MacIsaac, H.J., 1996. Potential abiotic and biotic impacts of zebra mussels on the
inland waters of North America. Am. Zool. 36, 287–299.

Mode
M
M
M
M
M
MM
M
M
M
M
M
M
N
N
OP
P
P
P
PP
P
P
R
R
aquatic food web. Oikos 87, 327–340.Zhao, Y., Frank, K.A., 2003. Factors affecting technology uses in schools: An ecological
A.L.J. Miehls et al. / Ecological
ack, R.N., Simberloff, D., Lonsdale, W.M., Evans, H., Clout, M., Bazzaz, F.A., 2000.Biotic invasions: causes, epidemiology, global consequences, and control. Ecol.Appl. 10, 689–710.
arsden, J.E., Chotkowski, M.A., 2001. Lake trout spawning on artificial reefs andthe effect of zebra mussels: Fatal attraction? J. Great Lakes Res. 27, 33–43.
ayer, C.M., Keats, R.A., Rudstam, L.G., Mills, E.L., 2002. Scale-dependent effectsof zebra mussels on benthic invertebrates in a large eutrophic lake. J. N. Am.Benthol. Soc. 21, 616–633.
ayer, C.M., VanDeValk, A.J., Forney, J.L., Rudstam, L.G., Mills, E.L., 2000. Response ofyellow perch (Perca flavescens) in Oneida Lake, New York, to the establishmentof zebra mussels (Dreissena polymorpha). Can. J. Fish. Aquat. Sci. 57, 742–754.
ayhew, B., 1980. Structuralism vs. individualism Part 1: Shadow boxing in the dark.Soc. Forces 59, 335–375.
cCann, K.S., 2000. The diversity-stability debate. Nature 405, 228–233.cCann, K., Rasmussen, J., Umbanhowar, J., Humphries, M., 2005. The role of space,
time, and variability in food web dynamics. In: deRuiter, P., Wolters, V., Moore, J.(Eds.), Dynamic Food Webs: Multispecies Assemblages Ecosystem Developmentand Environmental Change. Academic Press, San Diego, CA, USA, pp. 56–70.
cEliece, R.J., 1977. The Theory of Information and Coding. Addison-Wesley, Read-ing, MA, USA, p. 302.
ellina, E., Rasmussen, J.B., Mills, E.L., 1995. Impact of zebra mussel (Dreissena poly-morpha) on phosphorus cycling and chlorophyll in lakes. Can. J. Fish. Aquat. Sci.52, 2553–2573.
ills, E.L., Forney, J.L., 1988. Trophic dynamics and development of freshwaterpelagic food webs. In: Carpenter, S.R. (Ed.), Complex Interactions in Lake Com-munities. Springer-Verlag, New York, NY, USA, pp. 11–30.
ills, E.L., Casselman, J.M., Dermott, R., Fitzsimons, J.D., Gal, G., Holeck, K.T., Hoyle,J.A., Johannsson, O.E., Lantry, B.F., Makarewicz, J.C., Millard, E.S., Munawar, I.F.,Munawar, M., O’Gorman, R., Owens, R.W., Rudstam, L.G., Schaner, T., Stewart, T.J.,2003. Lake Ontario: Food web dynamics in a changing ecosystem (1970-2000).Can. J. Fish. Aquat. Sci. 60, 471–490.
ills, E.L., Forney, J.L., Clady, M.D., Schaffner, W.R., 1978. Oneida Lake. In: Bloomfield,J.A. (Ed.), Lakes of New York. Academic Press, New York, NY, USA, pp. 367–445.
oloney, C.L., Field, J.G., 1991. The size-based dynamics of plankton food webs. I: Asimulation model of carbon and nitrogen flows. J. Plankton Res. 13, 1003–1038.
alepa, T.F., Fahnenstiel, G.L., Johengen, T.H., 1999. Impacts of the zebra mus-sel (Dreissena polymorpha) on water quality: A case study in Saginaw Bay,Lake Huron. In: Claudi, R., Leach, J.H. (Eds.), Nonindigenous Freshwater Organ-isms: Vectors, Biology and Impacts. Lewis Publishers, Washington DC, USA, pp.255–271.
alepa, T.F., Lang, G.A., Fanslow, D.L., 2000. Trends in benthic macroinvertebratepopulations in southern Lake Michigan. Verh. Internat. Verein. Limnol. 27,2540–2545.
dum, E.P., 1969. The strategy of ecosystem development. Science 164, 262–270.auly, D., Christensen, V., Dalsgaard, J., Froese, R., Torres Jr., F., 1998. Fishing down
marine food webs. Science 279, 860–863.atten, B.C., 1984. Toward a theory of the quantitative dominance of indirect effects
in ecosystems. Verh. Ges. Ökol. 13, 271–284.atten, B.C., Bosserman, R.W., Finn, J.T., Cale, W.G., 1976. Propagation of cause in
ecosystems. In: Patten, B.C. (Ed.), Systems Analysis and Simulation in Ecology,vol. IV. Academic Press, New York, NY, USA, pp. 457–479.
érez-Espana, H., Arreguín-Sánchez, F., 1999. A measure of ecosystem maturity. Ecol.Model. 199, 79–85.
imm, S.L., 1979. The structure of food webs. Theor. Popul. Biol. 16, 144–158.imm, S.L., Lawton, J.H., 1980. Are food webs divided into compartments? J. Anim.
Ecol. 49, 879–898.innegar, J.K., Blanchard, J.L., Mackinson, S., Scott, R.D., Duplisea, D.E., 2005. Aggre-
gation and removal of weak-links in food-web models: System stability andrecovery from disturbance. Ecol. Model. 184, 229–248.
othoven, S.A., Nalepa, T.F., Schneeberger, P.J., Brandt, S.B., 2001. Changes in dietand body condition of lake whitefish in southern Lake Michigan associated withchanges in benthos. N. Am. J. Fish. Manage. 21, 876–883.
ose, K.A., Rutherford, E.S., McDermot, D.S., Forney, J.L., Mills, E.L., 1999. Individ-ual based model of yellow perch and walleye populations in Oneida Lake. Ecol.Monogr. 69, 127–154.
udstam, L.G., VanDeValk, A.J., Adams, C.M., Coleman, J.T.H., Forney, J.L., Richmond,M.E., 2004. Cormorant predation and the population dynamics of walleye andyellow perch in Oneida Lake. Ecol. Appl. 14, 149–163.
lling 220 (2009) 3194–3209 3209
Rutherford, E.S., Rose, K.A., Mills, E.L., Forney, J.L., Mayer, C.M., Rudstam, L.G., 1999.Individual-based model simulations of a zebra mussel (Dreissena polymorpha)induced energy shunt on walleye (Stizostedion vitreum) and yellow perch (Percaflavescens) populations in Oneida Lake New York. Can. J. Fish. Aquat. Sci. 56,2148–2160.
Shannon, C.E., 1948. A mathematical theory of communication. Bell System Technol.27, 379–423.
Simon, H.A., 1962. The architecture of complexity. Proc. Am. Philos. Soc. 106,467–482.
Stevenson, R.J., Bothwell, M.L., Lowe, R.L. (Eds.), 1996. Algal Ecology: FreshwaterBenthic Ecosystems. Academic Press, San Diego, CA, USA, p. 753.
Stewart, T.J., Lange, R.E., Orsatti, S.D., Schneider, C.P., Mathers, A., Daniels, M.E., 1999.Fish community objectives for Lake Ontario. Great Lakes Fishery CommissionSpecial Publication 99-1, Ann Arbor, MI, USA.
Strayer, D.L., Caraco, N.F., Cole, J.J., Findlay, S., Pace, M.L., 1999. Transformation offreshwater ecosystems by bivalves. BioScience 49, 19–27.
Strayer, D.L., Hattala, K.A., Kahnle, A.W., 2004. Effects of an invasive bivalve (Dreis-sena polymorpha) on fish in the Hudson River estuary. Can. J. Fish. Aquat. Sci. 61,924–941.
Szyrmer, J., Ulanowicz, R.E., 1987. Total flows in ecosystems. Ecol. Model. 35,123–136.
Teal, J.M., 1962. Energy flow in the salt marsh ecosystem of Georgia. Ecology 69,1647–1676.
Thorp, J.H., Casper, A.F., 2003. Importance of biotic interactions in large rivers: Anexperiment with planktivorous fish, dreissenid mussels and zooplankton in theSt Lawrence River. River Res. Appl. 19, 265–279.
Ulanowicz, R.E., 1986. Growth and Development: Ecosystems Phenomenology.Springer Verlag, New York, NY, USA, p. 203.
Ulanowicz, R.E., 1995. Ecosystem trophic foundations: Lindeman exonerate. In: Pat-ten, B.C., Jørgensen, S.E. (Eds.), Complex Ecology: The Part-Whole Relation inEcosystems. Prentice-Hall, Englewood Cliffs, NJ, USA, pp. 549–560.
Ulanowicz, R.E., 1996. Trophic flow networks as indicators of ecosystem stress.In: Polis, G.A., Winemiller, K.O. (Eds.), Food Webs: Integration of Patterns andDynamics. Chapman and Hall, New York, NY, USA, pp. 358–368.
Ulanowicz, R.E., 1997. Ecology The Ascendent Perspective. Columbia UniversityPress, New York, NY, USA, p. 224.
Ulanowicz, R.E., Kay, J.J., 1991. A package for the analysis of ecosystem flow networks.Environ. Soft. 6, 131–142.
Ulanowicz, R.E., Puccia, C.J., 1990. Mixed trophic impacts in ecosystems. Coenoses5, 7–16.
Vanderploeg, H.A., Nalepa, T.F., Jude, D.J., Mills, E.L., Holeck, K.T., Liebig, J.R., Grig-orovich, L.A., Ojaveer, H., 2002. Dispersal and emerging ecological impacts ofPonto Caspian species in the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci. 59,1209–1228.
VanDeValk, A.J., Adams, C.M., Rudstam, L.G., Forney, J.L., Brooking, T.E., Gerken, M.A.,Young, B.P., Hooper, J.T., 2002. Comparison of angler and cormorant harvest ofwalleye and yellow perch in Oneida Lake New York. Trans. Am. Fish. Soc. 131,27–39.
Vitousek, P.M., Walker, L.R., Whiteaker, L.D., Mueller-Dombois, D., Matson, P.A.,1987. Biological invasion by Myrica faya alters ecosystem development inHawaii. Science 238, 802–804.
Werner, E.E., Gilliam, J.F., 1984. The ontogenetic niche and species interactions insize structured populations. Annu. Rev. Ecol. Syst. 15, 393–425.
Westra, L., Miller, P., Karr, J.R., Rees, W.E., Ulanowicz, R.E., 2000. Ecological integrityand the aims of the Global Integrity Project. In: Pimentel, D., Westra, L., Noss, R.F.(Eds.), Ecological Integrity: Integrating Environment, Conservation and Health.Island Press, Washington, DC, USA, pp. 19–41.
Wilcove, D.S., Rothstein, D., Dubow, J., Phillips, A., Losos, E., 1998. Quantifying threatsto imperiled species in the United States. BioScience 48, 607–615.
Yodzis, P., Winemiller, K.O., 1999. In search of operational trophospecies in a tropical
perspective. AERJ 40, 807–840.Zhu, B., Fitzgerald, D.G., Mayer, C.M., Rudstam, Mills, E.L., 2006. Alteration of ecosys-
tem function by zebra mussels in Oneida Lake. Impacts Submerged Macrophytes9, 1017–1028.