Embed Size (px)
Citation preview

Early Retinal Developmentin the Zebrafish, Danio rerio:
Light and Electron Microscopic Analyses
ELLEN A. SCHMITT AND JOHN E. DOWLING*Department of Molecular and Cellular Biology, Harvard University,
Cambridge, Massachusetts 02138
The morphological differentiation of the zebrafish retina was analyzed by using light (LM)and transmission electron (TEM) microscopy between the time of initial ganglion celldifferentiation (<32 hours postfertilization; hpf) and shortly after the point when the retinaappears functional (<74 hpf), i.e., when all major cell types and basic synaptic connections arein place. The results show that the inner retinal neurons, like the photoreceptor and ganglioncells, differentiate first within the ventronasal region, and differentiation subsequentlyspreads asymmetrically into the nasal and dorsal regions before reaching the ventrotemporalretina. In addition, we show that the attenuation of the optic stalk occurs in parallel withganglion cell differentiation between 32 and 40 hpf. The first conventional synapses appearwithin the inner plexiform layer simultaneously with the first photoreceptor outer segmentdiscs at 60 hpf; functional ribbon triads arise within photoreceptor synaptic terminals at 65hpf; and synaptic ribbons occur within bipolar cell axon terminals at the time larvae exhibittheir first visual responses (<70 hpf). Although development is initially more advanced withinthe ventronasal region between 50 and 60 hpf, development across the retina rapidlyequilibrates such that it is relatively comparable within all quadrants of the central retina by70 hpf. An area within the temporal retina characterized by tightly packed and highly tieredcones emerges with subsequent development. Retinal differentiation in the zebrafish corre-sponds with that generally described in other vertebrates and can be correlated with thedevelopment of visual and electroretinographic responses in the animal. J. Comp. Neurol.404:515–536, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: optic stalk; retinal lamination; photoreceptor development; synaptogenesis;
teleost retina
The retina is an advantageous part of the vertebratecentral nervous system (CNS) in which to analyze neuro-genesis and other aspects of development (for reviews, seeCepko et al., 1996; Graw, 1996; Fini et al., 1997; Oliver andGruss, 1997). Recently, much attention has focused specifi-cally on the development of the zebrafish eye as a modelsystem (Burrill and Easter, 1994, 1995; Schmitt andDowling, 1994, 1996; Ekker et al., 1995; Raymond et al.,1995; Macdonald and Wilson 1996; Hyatt et al., 1997a,b).However, with the exception of the development of gamma-aminobutyric acid (GABA) and tyrosine receptor kinase(trk) immunoreactivity in the inner plexiform layer andoptic nerve (Sandell et al., 1994a,b), reports dealing withthe developing zebrafish retina have focused exclusivelyon photoreceptors and ganglion cells (Branchek andBremiller 1984; Kljavin, 1987; Larison and Bremiller,1990; Burrill and Easter, 1994, 1995; Raymond et al.,1995; Schmitt and Dowling, 1996). As a result, relativelylittle is known regarding the events involving the differen-
tiation of the cell types and structures within the innernuclear and outer and inner plexiform layers.
Unlike most vertebrates, in which retinal developmentcommences within the central region of the retina, differen-tiation of photoreceptors and ganglion cells in the ze-brafish is initiated within the ventral retina (Kljavin 1987;Burrill and Easter, 1995; Raymond et al., 1995; Schmittand Dowling, 1994, 1996). This was first recognized forphotoreceptors by Kljavin (1987). Subsequent investiga-tions have reported that ganglion cells are initially foundwithin the ventral region, nasal to the optic nerve (Schmittand Dowling, 1994, 1996; Burrill and Easter, 1995), and
Grant sponsor: NIH; Grant number: EY-00811.*Correspondence to: John E. Dowling, Department of Molecular and
Cellular Biology, Harvard University, 16 Divinity Avenue, Cambridge, MA02138.
Received 2 September 1997; Revised 14 September 1998; Accepted 17September 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 404:515–536 (1999)
r 1999 WILEY-LISS, INC.

that the initial expression of opsin and other cell markerswithin both rods and cones also occurs in the ventral retinaalong the nasal edge of the choroid fissure (Raymond et al.,1995; Schmitt and Dowling, 1996). Cell labeling of bothganglion cells and cones have demonstrated that differen-tiation of these cell types subsequently sweeps from theventronasal patch into the nasal and dorsal regions beforereaching the temporal retina (Burrill and Easter, 1995;Raymond et al., 1995; Schmitt and Dowling, 1996). Theprogressive spread of rod photoreceptors, however, dis-plays a pattern independent from that of the other celltypes; after the rapid generation of an initial patch of rodsnasal to the choroid fissure (<50–55 hours postfertiliza-tion; hpf), rods quickly begin to differentiate on the tempo-ral side of the fissure (<55–57 hpf). Finally, rods arisedorsally in a relatively slow and sporadic fashion after 60hpf (Raymond et al., 1995; Schmitt and Dowling, 1996).
This article provides a description of eye growth, retinallamination, and the differentiation of the cells within theinner nuclear layer in the zebrafish between 32 and 74 hpfby using light (LM) and transmission electron (TEM)microscopy. This time period delineates the interval be-tween the first sign of ganglion cell differentiation and thepoint when the retina appears functional in possessing allthe major cell types and basic synaptic connections. Animportant reason for a comprehensive description of thedeveloping zebrafish retina is to provide a basis for theanalysis of the numerous eye and retinal mutations nowisolated from mutagenesis screens (Brockerhoff et al.,1995, 1997; Driever et al., 1996; Haftner et al., 1996;Heisenberg et al., 1996; Malicki et al., 1996; Fadool et al.,1997; Lall et al., 1997). Retinal mutations reported thusfar range from perturbations affecting eye size and retinallamination to much more subtle effects involving a singlecell type or changes detected only at the behavioral level(Brockerhoff et al., 1995, 1997; Malicki et al., 1996; Fadoolet al., 1997; Lall et al., 1997).
MATERIALS AND METHODS
Zebrafish embryos were collected and reared in 50-mlPetri dishes at 28.5°C. Groups of 60–70 embryos (orlarvae) between 32 and 74 hpf were dechorionated andstaged by criteria outlined by Westerfield (1994). Larvaewere also sampled on day 5 after fertilization.
LM
Prior to fixation, embryos and larvae were anesthetizedon ice at 4°C in accordance with standards for animal careoutlined by Harvard University and the NIH. In themajority of cases, groups of 15 specimens were immedi-ately fixed with 1% paraformaldehyde, 2.5% glutaralde-hyde made in 0.06 M phosphate buffer with 3% sucrose, pH7.4, for 1.5 hours (4°C). Embryos at 40 hpf were fixed in thesame fixative made in 0.08 M sodium cacodylate bufferwith 0.0075 M CaCl, 3% sucrose, pH 7.4, which enhancedthe visibility of dividing cells in the retina. All specimenswere postfixed in 1% osmium tetroxide for 1 hour at 4°C.Primary fixation was omitted for 5-day-old larvae, whichwere processed in 1% osmium for 2 hours at 4°C. Afterrinsing, specimens were dehydrated in a graded series ofethanol-water mixtures and infiltrated with epon/aralditeresin overnight (20°C). A total of ten specimens from eachstage were oriented and embedded for transverse, sagittal,or horizontal sectioning. Serial sections (0.5–1.0 µm) werecut and stained with an aqueous solution containing 1%methylene blue, 1% azure II made with 1% borax andphotographed under brightfield. Measurements of the eyecups, retinas, retinal laminae, and so forth were generallytaken from transverse sections by using a 103 objectivewith a gradicule. Mean values were calculated for thedifferent parameters from at least three specimens of eachof the major time points (32, 36, 40, 50, 60, 70, and 74 hpf).
Abbreviations
A amacrine cellB bipolar cellbb basal bodyC conec centriolecc connecting ciliumCN cone nucleiDP dorsal peripheryDR dorsal retinacf choroid fissurecos cone outer segmente photoreceptor ellipsoidFb forebrainG ganglion cellg glycogen granulesGCL ganglion cell layergz germinal zoneH horizontal cellHN horizontal cell nucleush horizontal sectionHa hyaloid arteryINL inner nuclear layerIPL inner plexiform layerips interphotoreceptor spaceis inner segmentL lens, lipid dropletLM light microscopylss long single coneM Muller cell fiberm mesenchymemi mitochondria
my photoreceptor myoidnb neuroblastNR nasal retinan nucleusNe neuroepitheliumnp neural processoc optic chiasmOL optic lumenON optic nerveONL outer nuclear layerOPL outer plexiform layerOS optic stalkos outer segmentsPe pigment epitheliumpg pigment granuleR retina, rodRN rod nucleiros rod outer segments sagittal sectionssc short single conesr synaptic ribbonst synaptic terminalsv synaptic vesiclest transverse sectionTR temporal retinaV ventricleVP ventral patchVR ventral retinay yolkZA zonulae adherentes
516 E.A. SCHMITT AND J.E. DOWLING

TEM
Embryos between 50 and 74 hpf were staged, fixed, andembedded as described above. Ultrathin transverse sec-tions of the retina through the region of the optic nervewere stained with uranyl acetate and lead citrate. Sectionswere viewed and photographed with a JOEL 101B trans-mission electron microscope.
In situ hybridization
The wholemount in situ hybridization protocol used wassimilar to that described by Oxtoby and Jowett (1993).Embryos were prehybridized for 3–5 hours at 65°C. La-beled probe at a final contration of 0.5 ng/µl was added toprehybridized embryos, and the hybridization reactionwas allowed to proceed at 65°C for 20 hours. Riboprobeswere generated as previously described (Oxtoby and Jowett,1993) from plasmids containing cDNA of zebrafish rhodop-sin. After in situ wholemount staining, embryos werecleared in a glycerol-water series (25, 50, 70, 95, and 100%)and mounted for photography in glycerol within deep-wellslides without coverslips.
RESULTS
Eye morphogenesis in the zebrafish commences at 11.5hpf and proceeds rapidly such that the eye cup is wellformed by 24 hpf (Schmitt and Dowling, 1994). Between 24and 36 hpf, the retina undergoes rapid proliferation.Clusters of mitotic profiles are found along the opticlumen, and the width of the retina almost doubles, increas-ing from <35 µm to <55 µm. We previously reported thatthe eye cups rotate in the anterior direction after 24 hpfsuch that the position of the choroid fissure graduallyshifts from an anterior position to its final ventral locationby 36 hpf (Schmitt and Dowling, 1994). Axons of ganglioncells exit the eyes between 32 and 34 hpf (Stuermer, 1988;Burrill and Easter, 1994, 1995), and the optic stalksconstrict and are gradually replaced by the optic nerves(Schmitt and Dowling, 1994). To examine morphologicalchanges associated with these events, horizontal andtransverse sections were taken from embryos at 32, 36,and 40 hpf (lines h and t in Fig. 1A). Subsequent stages ofretinal development were examined between 50 and 74 hpfin transverse and sagittal sections generally taken throughthe eye at the level of the optic nerve (lines t and s,respectively, in Fig. 1A,B).
32–40 hpf: Ganglion cell differentiation andthe optic stalk
Between 32 and 34 hpf, the retina is an undifferentiatedpsuedostratified epithelium that is approximately three tofour cell layers wide and from 36 to 40 µm across. Clustersof mitotic profiles are frequently observed along the opticlumen, and pigment granules are present within theelongate and flattened cells of the pigment epithelium (Pe)(arrowheads in Fig. 2a). As noted in previous investiga-tions, the first signs of ganglion cell differentiation areseen in horizontal sections taken ventral to the midline atthe level of the optic stalks (G in Fig. 2a). A small numberof ganglion cells identified by oval, lightly staining nucleiwith prominent nucleoli are positioned within the nasalregion of the ventral retina, i.e., in the ventronasal patch.In contrast, neighboring neuroblasts are darkly stainingand more densely packed due to a lack of perinuclear
cytoplasm. Despite the presence of somata, ganglion cellaxons are not yet visible by standard LM. Studies usinglipophilic fluorescent dyes, however, have shown the trajec-tories of ganglion cell axons as they exit the eye and reachthe optic chiasm at 32–36 hpf (Stuermer, 1988; Burrill andEaster, 1994, 1995; Table 1).
Horizontal sections taken ventral to the vitreal cavityshow the optic stalks extending from the eye cups across tothe anterior region of the forebrain (OS in Fig. 3a). At thislevel, the choroid fissures of each eye are positioned suchthat they divide the retina into a one-third nasal compart-ment and a two-thirds temporal compartment (see Fig. 2a;open arrow in Fig. 3a). Where the optic stalks extend fromthe eye cups, flattened cells extend along both sides of thechoroid fissure (asterisk in Fig. 2a). Although these cellsare continuous with the Pe, they are devoid of pigmentgranules. Unlike mammalian species, melanin is alsoabsent within the optic stalks during the early outgrowthof ganglion cell axons (Silver and Sapiro, 1981; Stronginand Guillery, 1981).
The walls of the optic stalks are composed of one to twolayers of elongated cells (OS in Fig. 3a). Although thesecells are segregated from those within the retina, theyappear similar morphologically, with the exception ofvacuoles occasionally found in cells located along theanterior surface of the forebrain (asterisk in Fig. 3a). Asmall number of mitotic profiles are observed within theoptic stalks adjacent to the optic lumen (OL), which existswithin the walls of the optic stalks and extends across theanterior surface of the forebrain (arrows and OL, respec-tively, in Fig. 3a). The optic lumen is narrow as it exits theeye cup but broadens centrally, where it measures <8 µmacross. Serial sections taken through more dorsal regionsof the optic stalks show that the lumen is also continuouswith the ventricle within the anterior region of the fore-brain (not shown). Numerous fusiform mesenchymal cellsare observed both anterior and posterior to the optic stalksand along the proximal surface of the eye cups adjacent tothe forebrain (m in Fig. 3a).
Zebrafish embryos increase by <0.2 mm in length at30–36 hpf and measure <2.7 mm in length at 36 hpf
Fig. 1. Diagrams of the zebrafish eye that illustrate the planes ofsections shown in Figures 2–10. A: Lateral view. Horizontal sections(h) were taken ventral to the midline at the level of the optic stalks inembryos 32–40 hpf and are shown in Figures 2 and 3. Transversesections (t) were taken along the dorsal–ventral axis through theapproximate location of the ventronasal patch (arrow) in embryos50–74 hpf and are shown in Figures 4, 6, and 8. B: Dorsal view.Sagittal sections (s) shown in Figures 5, 7, 9, and 10 were takenthrough the eye cup at different planes parallel to the anterior–posterior axis (A= P).
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 517

(Kimmel et al., 1995). Externally, pigmentation of the eyeappears prominent because the density of pigment gran-ules has increased within the Pe (arrowheads in Fig. 2b).The psuedostratified retina has increased to <55 µm wideand is now more than six cell layers across (R in Fig. 2b).The retinal neuroblasts are smaller and more denselypacked as compared with those observed at 32 hpf (com-pare Fig. 2a and b). In addition, the nuclei are highlyheterochromatic. As noted previously, clusters of dividingcells are observed within the retina along the optic lumen(short arrows in Fig. 2b). In horizontal sections takenventral to the midline of the eye cup, a patch of lightlystaining ganglion cells is now present within the nasal
region of the ventral retina (G in Fig. 2b). Lightly stainingaxons are now visible that extend from the ganglion cellsnear the vitreal retinal border and converge to exit theretina (ON in Fig. 2b). Clusters of mitotic profiles areparticularly abundant in the retina where these earlyaxons exit. The differentiation of ganglion cells is not yetobserved temporal or dorsal to the optic nerve.
As noted earlier, the optic stalks are best observed inhorizontal sections taken ventral to the vitreal cavity atthe level of the choroid fissure (OS in Fig. 3b). Flattenedcells, which are continuous with the Pe, line both sides ofthe fissures. The walls of the optic stalks are reduced to asingle cell layer, where they exit the eye cups and the cellshave become more elongated (white arrows in Fig. 3b). Inthe central region of the optic stalks (i.e., anterior to theforebrain), this attenuation is less extensive and the cellsremain cubodial and include mitotic profiles (black arrowsin Fig. 3b). Although the lumen within the optic stalks hasbecome more constricted, axons can now be seen within it(Fig. 3b). The mesenchymal cells noted at 32 hpf have begun togenerate a meshwork at the back of the eye cups, wherethe choroidal vascularization is presumably forming.
Between 36 and 40 hpf, ganglion cell differentiationspreads rapidly into the temporal region of the retina (G inFig. 2c). In addition to the large patch of cells nasal to theoptic nerve, two layers of ganglion cells are observed alongthe vitreal retinal border within the temporal retina.Concomitant with the differentiation of a large number ofganglion cells, the optic nerves become much broader (<7µm in diameter), exiting the eye cups at the apex of thechoroid fissures (arrows in Fig. 3c). As noted earlier, thebroad separation of the Pe occurs at the scleral surface,where the optic nerves exit, and patches of mitotic profilesare still found in the retina adjacent to the optic stalks(asterisk in Fig. 2c). The walls of the optic stalks aregreatly reduced, and only a small number of elongatedstalk cells extend from the eye cup in association with theoptic nerves on each side (short arrows in Fig. 3c). Theoptic stalks have become essentially obliterated at thisstage. The optic nerves (long arrows) extend centrallyalong the track previously traversed by the optic stalks,and these cross at the optic chiasm located at the anterior/ventral surface of the forebrain (oc in Fig. 3c). The mesen-chymal meshwork now completely surrounds the optic
Fig. 2. Horizontal sections taken along the nasal–temporal axis(N = T for all panels) show the initial differentiation of ganglion cells.(An alternative fixation protocol was used for the section shown in c;see Materials and Methods for details.) a: At 32 hours postfertilization(hpf), the first ganglion cells (G) are found within the ventronasalregion at the level of the optic stalk (OS) and choroid fissure (cf). Thesecells possess large, lightly staining nuclei and are distinct from thedarkly staining and tightly packed cells of the retinal neuroepithe-lium. The optic stalk (OS) extends from the eye, where the choroidfissure separates the retina. Elongated cells continuous with thepigment epithelium (arrowheads) but devoid of pigment lie within thechoroid fissure (asterisk). Mesenchymal cells (m) surround the opticstalk adjacent to the eye cup. b: At 36 hpf, the retina (R) has increasedin thickness and cell density. A patch of lightly staining ganglion cells(G) lies within the ventronasal retina adjacent to the optic nerve (ON).Mitotic profiles (arrows) lie adjacent to the pigment epithelium(arrowheads). c: By 40 hpf, the retina has continued to increase in boththickness and cell density. In addition to the large patch of ganglioncells within the nasal retina, two layers of ganglion cells (G) extendtemporal to the optic nerve. A patch of mitotic profiles (asterisk) liesadjacent to the optic nerve (ON). Pigmentation has increased withinthe pigment epithelium (arrowheads). L, lens. Scale bar 5 25 µm.
518 E.A. SCHMITT AND J.E. DOWLING

nerves and extends across the proximal surface of the eye(m in Fig. 3c).
2 days postfertilization (50 hpf)
Lamination. At 50 hpf, embryos measure <3 mm inlength. Externally, patches of scattered iridophores areobserved across the surface of the eye, some of which forma discontinuous ring around the lens (Kimmel et al., 1995).The eye cup measures <200 µm along the dorsal–ventralaxis in transverse sections (Fig. 4a). The optic nervedivides the retina into a roughly two-thirds dorsal compart-ment and a one-third ventral compartment. Transversesections show that extensive lamination has occurredsince 40 hpf within two regions of the retina. One lami-
nated region extends over 80 µm dorsal to the optic nervewithin the central retina (Fig. 4b), and a second regionextends across 50 µm of the ventral retina distal to theoptic nerve (Fig. 4c). Although the cells within the lami-nated region of the ventral retina are presumably closelyassociated with the ventronasal patch, the maturation ofthe neurons observed in both regions appears roughlycomparable by LM (compare Fig. 4b and c); three nuclearlayers (ganglion cell, inner nuclear, and outer nuclearlayers) are separated by the inner and outer plexiformlayers. However, pseudostradified neuroepithelium is ob-served directly ventral to the optic nerve and appears lessdifferentiated (Ne in Fig. 4b), and no sign of lamination isyet seen near the dorsal periphery (asterisk in Fig. 4a).
Inner retina. Within the dorsal and ventral lami-nated regions, the ganglion cell layer is <12 µm wide andincludes two to three layers of ganglion cells that aresituated along the vitreal retinal border (Fig. 4b,c). Theinner plexiform layer (IPL) averages 6 µm across andincludes a population of darkly staining nuclei (smallarrows in Fig. 4b). These nuclei are regularly spacedwithin the IPL and are presumed to be displaced amacrinecells, which subsequently relocate from the IPL to theganglion cell layer (Genis Galvez et al., 1977; Hinds andHinds, 1983). Displaced amacrine cells within the IPL areespecially evident in more peripheral regions where differ-entiation is not as advanced (black arrowheads in Fig. 5a).TEM shows that their nuclei are highly heterochromaticand that they possess very little cytoplasm while extend-ing across the IPL (arrows in Fig. 5c).
The IPL is composed of a homogeneous population ofneural processes interspersed by numerous intercellularspaces (not shown). Occasionally, small vesicles, <45 nmin diameter, are seen within the neural processes. Al-though no mature synapses are observed by TEM, a sparsenumber of electron-dense zones, 100–150 nm in length,occur along membranes between neighboring processesand may represent the initial stages of conventionalsynapse formation.
Fig. 3. Horizontal sections taken along the temporal–nasal axis(T = N for all panels) ventral to the midline show the optic stalks (OS)extending from the retina (R) across the anterior surface of theforebrain (Fb). (An alternative fixation protocol was used for thesection shown in c; see Materials and Methods for details.) a: At 32hours postfertilization (hpf), the walls of the optic stalk are one to twocell layers thick and mitotic cells are often observed (black arrows).Vacuoles (white asterisk) are occasionally found within the stalk cellsat the anterior side of the optic lumen (OL). Ventral to the optic nerve,the choroid fissure (open arrow) separates the retina (R) into a nasaland temporal compartment, and elongated cells extend within thefissure, which are continuous with the pigment epithelium (arrow-heads). Mesenchymal cells (m) are scattered at the back of the eye cupand across the anterior surface of the optic stalks. b: At 36 hpf, theoptic stalks have attenuated, particularly where they exit the eye cups(white arrows) adjacent to the choroid fissure (open arrow). Morecentrally, the stalk (OS) remains broad and mitotic cells are observed(black arrows). Lightly staining material, which resembles the axonsof ganglion cells, can be observed centrally within the optic lumen(OL). Mesenchyme (m) has begun to formed a meshwork at the back ofthe eye cup and anterior to the optic stalks. c: At 40 hpf, the opticstalks have been virtually obliterated and replaced by the optic nerve(long arrows). A small number of elongated stalk cells are associatedwith axons as they exit the eye (short arrows) and lie at the anterior ofthe optic chiasm (OC). The mesenchymal meshwork (m) has becomeextensive, particularly at the anterior of the optic stalks (OS). V,ventricle; arrowheads, pigment epithelium. Scale bar 5 30 µm.
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 519

The inner nuclear layer (INL) averages <25 µm wide at50 hpf. Two to three layers of lightly staining, ovalamacrine cell nuclei lie at the vitreal border, adjacent tothe IPL (A in Fig. 4b,c). Horizontal cells that lie along thescleral INL border, adjacent to the outer plexiform layer,are small and irregularly shaped (H in Fig. 4b). Within themid-region of the INL, three layers of spindle-shapedneuroblasts occur. These cells, which are darkly stainingand highly heterochromatic, are presumed to be immaturebipolar cells. Thus, similar to other vertebrates, differentia-tion of amacrine and horizontal cells precedes that ofbipolar cells within the INL (reviewed in Grun, 1982).
Outer retina. The outer plexiform layer (OPL) is lessthan 2 µm wide on day 2 and appears as an irregularanuclear border within the central and ventral retina(thick arrow in Fig. 4b,c). The OPL is not yet evidentwithin the dorsal periphery (Figs. 4a, 5a). TEM showsnumerous dendritic processes in the OPL despite theabsence of photoreceptor synaptic terminals (OPL in Fig.4f). Centrioles and diplosomes are occasionally foundwithin the OPL processes (thick arrows in Fig. 4f). In thecase of diplosomes, the two centrioles are oriented at rightangles (arrow in Fig. 4e). Usually, centrioles and diplo-somes are noted within large expanded processes. Themajority of neural processes in the OPL at 50 hpf arepresumed to be those of horizontal cells because of thepresence of glycogen granules (30 nm in diameter), whichare also found in the perinuclear cytoplasm of horizontalcells (g in Fig. 4f).
In transverse sections, large oval nuclei presumed to bethose of cones extend within the outer nuclear layer(ONL), which is <8 µm wide both dorsal and ventral to theoptic nerve (Fig. 4b,c). Ventral to the optic nerve, thesenuclei are interspersed by smaller round nuclei generallypositioned near the OPL (thin arrow in Fig. 4c). As notedby Kljavin (1987), clear differences are not yet detectablebetween the nuclei of rods and cones. This observation isunlike descriptions of embryonic retinas from other tele-osts, in which developing rods and cones are clearlydistinguishable at initial stages of differentiation (Kunz etal., 1983; Schmitt and Kunz, 1989). A small number ofneuroblasts, which resemble the rod progenitor cells com-
monly described in teleost retinas (Raymond, 1985; Ray-mond and Rivlin, 1987), are seen within the OPL of theventral retina. Where differentiation is most advanced(i.e., in the ventronasal patch and more dorsocentralregion), photoreceptor inner segments extend <2 µmscleral to the outer limiting membrane (OLM; curvedarrows in Fig. 4b,c). Inner segments contain mitochondria,rough endoplasmic reticulum, and abundant free ribo-somes (is in Fig. 4d). Muller cell differentiation has alsocommenced, and zonulae adherentes are seen betweenphotoreceptor inner segments and the Muller cell apicalfibers to form the OLM (arrows in Fig. 4d). Patches ofelectron-lucent Muller cell apical fibers (M in Fig. 4d)frequently extend between developing photoreceptors andcontain extremely large mitochondria. Although no outersegment discs are yet observed, the connecting cilia extendfrom the apical ends of the inner segments (cc in Fig. 4dinset). These cilia measure <1 µm in length 3 <0.4 µm inwidth. Microtubules extend from the basal body of thecilium into the axoneme, which extends within the photo-receptor space. The associated centriole is often observedoriented at a right angle to the basal body of the cilium (notshown).
The pigment epithelium is <2.5 µm wide and possesseslarge irregular nuclei oriented laterally to the retina. Ahigh density of both spherical and spindle-shaped pigmentgranules is present throughout the cytoplasm (Pe in Fig.4d). Although Pe processes have not yet formed, largespaces (interphotoreceptor space) occur between the Peand developing inner segments in regions where retinaldifferentiation is most advanced (ips in Fig. 4d). In con-trast, the Pe directly contacts the retina at the level of theOLM in less mature regions where photoreceptor innersegments have just begun to form (asterisks in Fig. 5d).
Ventronasal patch. The location of the ventronasalpatch is best observed in sagittal sections (line s in Fig. 1).This patch lies nasal to the optic nerve and is particularlynoticeable because of the precocious differentiation of thephotoreceptors (right of dashed line in Fig. 5a,b,d). How-ever, careful examination shows that the differentiation ofother major cell types, i.e., amacrine, bipolar, and horizon-tal cells, is also more advanced within this region as
Fig. 4. Transverse sections taken along the dorsal–ventral axis(D = V for all panels except (b), where the orientation of the photomicrograph is shifted 90° in relation to that shown in (a). Dorsal is tothe right.) at 50 hours postfertilization (hpf). a: Retinal lamination isobserved within the ventral retina (VR) distal to the optic nerve (ON)and extends from the optic nerve into the dorsal retina (DR). Mitoticprofiles (arrow) occur along the outer plexiform layer, where lamina-tion is observed, and at the optic lumen (large arrow) adjacent to thepigment epithelium, where lamination is not yet observed in thedorsal periphery (asterisk). The dorsal (b5) and ventral region (c5) areshown at higher magnification in Figures 4b,c, respectively. b,c:Retinal laminae dorsal and ventral to the optic nerve include theganglion cell layer (GCL), inner plexiform layer (IPL), inner nuclearlayer (INL), and outer nuclear layer (ONL). The outer plexiform layer(broad arrow) is somewhat discontinuous. The differentiation ofganglion cells (G), amacrine cells (A), and horizontal cells (H) is noted.Lamination is not observed immediately ventral to the optic nerve,where undifferentiated neuroepithelial cells (Ne) are located. Nucleilocated within the IPL are presumed to be displaced amacrine cells(small arrows in b), and spindle-shaped neuroblasts (nb) in themid-region of the INL are presumptive bipolar cells. The innersegments of photoreceptors (curved arrows) appear as a lightlystaining amorphous layer adjacent to the pigment epithelium (arrow-heads). c: The optic nerve (ON) has a forked appearance (asterisk) as
bundles of axons from the dorsal and ventral retina merge at thevitreal border. Small round nuclei lie adjacent to the outer plexiformlayer within the ONL of the ventral retina (thin arrow). d: Electronmicrograph shows the initial differentiation of photoreceptor innersegments (is) possessing large mitochondria (asterisks) extendingtoward the pigment epithelium into the interphotoreceptor space (ips).The lightly staining cytoplasm of Muller cells (M) project between conenuclei (CN) and form zonulae adherentes (arrows) with the developinginner segments. Inset: Microtubules (small arrows) extend from thebasal body (bb) into the axoneme of a photoreceptor connecting cilium(CC). The cilium extends from inner segments toward the pigmentepithelium prior to the formation of outer segment discs. e: Adiplosome (large arrow) comprised of two centrioles oriented at rightangles occurs within a neural process located in the outer plexiformlayer (OPL), adjacent to the nucleus of a horizontal cell (HN).f: Although there is no sign of photoreceptor synaptic terminals,numerous lightly staining neural processes (thin arrows) extendwithin the OPL. Processes that contain glycogen granules (g) arebelieved to be those of horizontal cells because glycogen is abundantwithin the cell cytoplasm adjacent to horizontal cell nuclei (HN).Centrioles (thick arrows) are frequently found within expanded neuralprofiles within the OPL. Scale bars 5 40 µm in a, 12 µm in b and 14 µmin c, 1 µm in d and 0.5 µm in inset, 0.25 µm in e, 0.45 µm in f.
520 E.A. SCHMITT AND J.E. DOWLING

Figure 4
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 521

compared with regions temporal to the midline (right ofdashed line in Fig. 5b). Temporally, undifferentiated neuro-blasts occur within the INL, the OPL is not yet formed, andmitotic profiles (arrows) sit along the optic lumen (Fig. 5b).
At <55 hpf, a small number of photoreceptor outersegments ,1 µm in length are observed concomitant witha slight expansion of the interphotoreceptor space (Fig.5d). The OPL (black arrows) has become more distincttemporal to the ventronasal patch but remains poorlyformed within the dorsal periphery (Fig. 5a). Where theOPL is lacking, mitotic cells are found along the opticlumen adjacent to the Pe (white arrowheads in Fig. 5a).However, mitotic profiles are frequently observed directlyvitreal to the OPL once the OPL has formed (white arrowsin Fig. 5a). Although the OPL and ONL are well formedmore dorsally within the nasal retina (small black arrowsin Fig. 5a), they are, surprisingly, not distinct within thenasal retina immediately adjacent to the ventronasalpatch, and numerous mitotic cells are still observed alongthe optic lumen (asterisk in Fig. 5a).
2.5 days postfertilization (60 hpf)
Lamination. The growth rate of embryos slows to 20µm per hour during the second day of development (Kim-mel et al., 1995). By 60 hpf, they measure <3.3 mm inlength, and the head has lifted considerably from the yolksuch that the body axis is relatively straight. Transversesections show that the eye cup continues to enlarge rapidlyand now averages <220 µm along the dorsal–ventral axis(Fig. 6a). Unlike the period between 32 and 40 hpf, thewidth of the retina increases at a more moderate rate andnow averages <70 µm. The width of the Pe has increasedto <3 µm and the optic nerve is now <12 µm in diameter.Extensive differentiation has occurred between 50 and 60hpf. Transverse sections through the central region showretinal laminae extending across more than 80% of theretina, and the undifferentiated peripheral margins (germi-nal zones) are now shorter than 40 µm (asterisks in Fig.6a). Although transverse sections suggest that overalldifferentiation is roughly comparable within the ventrona-sal patch and more central retina dorsal to the optic nerve(compare Fig. 6c and b), sagittal sections show thatdevelopment is still more advanced within the ventronasalpatch (VP in Fig. 7a). Unlike earlier stages, mitotic profiles
are very rarely observed, and pyknotic cells are nowoccasionally found within the ganglion cell and innernuclear layers (short arrows in Fig. 6a,b).
Inner retina. Ganglion cells have increased in sizeand appear less heterochromatic than do those observed at50 hpf. Most of the darkly staining nuclei presumed to bedisplaced amacrine cells are more oval in shape and arenow positioned along the scleral border of the ganglion celllayer, which is <14 µm wide (thin arrows Fig. 6a,c). Moreperipherally, these cells are still located within the IPLand often appear fusiform as a small amount of peri-nuclear cytoplasm tapers in the direction of the innernuclear layer (not shown).
The IPL is <8 µm wide, and TEM shows the appearanceof a new population of darkly staining neural processesthat contain a homogeneous distribution of synapticvesicles characteristic of bipolar cell axons and axonterminals (reviewed in Grun, 1982; Dowling, 1987). Thesepresumptive bipolar cell axons are easily distinguishedfrom smaller, electron-lucent processes of amacrine andganglion cells. Similar to descriptions in other vertebrates,amacrine cell processes are identified by an irregulardistribution of synaptic vesicles, whereas those of ganglioncells generally lack these vesicles and possess ribosomes(A, G in Fig. 6e). In addition to the electron-dense contactzones noted first at 50 hpf, conventional synapses are nowoccasionally present in the IPL both ventral and dorsal tothe optic nerve (curved arrow Fig. 6e). These synapsesrange between 100 and 350 nm in length and are foundprimarily on processes, which are characteristic of ama-crine and ganglion cells. The number of intercellularspaces within the IPL has decreased, and fine branches ofMuller cell fibers containing abundant glycogen are nowinterspersed between the neural processes.
Simultaneous to the appearance of bipolar cell axonterminals in the IPL, two to three layers of bipolar cellssomata are now distinguishable within the mid-region ofthe INL (B in Fig. 6b,c). Bipolar cells have round anddarkly staining somata that are distinct from the lightlystaining somata of horizontal and amacrine cells. Horizon-tal cells also appear more mature because they are nowelongate and laterally oriented along the OPL (H in Fig.6b,c). Muller cell differentiation has progressed such thatlarge, lightly staining fibers are now seen intermittently
Fig. 5. Sagittal sections taken along the temporal–nasal axis (T =N for all panels) at 50 hours postfertilization (hpf) (b–c) and 54 hpf(a,d). a: Sagittal section through the mid-peripheral region showsnuclei presumed to be displaced amacrine cells (black arrowheads)within the inner plexiform layer. Lamination appears most advancedwithin the ventronasal patch (VP), which lies nasal to the midline(dashed line). Although the ganglion cell layer (G) and inner nuclearlayer (INL) are found in all regions, the outer plexiform layer (smallarrows) is not yet evident within some regions (e.g., the dorsalperiphery), and numerous mitotic profiles (white arrowheads) liealong the pigment epithelium, including the region nasal to theventronasal patch (asterisk). Mitotic profiles also occur along the outerplexiform layer (white arrows). L, lens. b: Sagittal section through thecentral retina shows the ventral region at high magnification. Theventronasal patch (VP) lies on the nasal side of the midline (dashedline). Ganglion cells (G) appear comparable across the ventral region.The neurons within the inner and outer nuclear layers of the ventrona-sal patch show signs of advanced development relative to the undiffer-entiated neuroblasts (Nb) positioned temporal to the midline. Ama-crine (A), bipolar (B), and horizontal (H) cells are distinct, as is theouter plexiform layer (arrow). The inner segments of photoreceptors(is) are forming adjacent to the pigment epithelium, which is broad
relative to that located temporal to the ventral midline. Mitoticprofiles (arrows), indicative of less advanced development, frequentlyoccur in the ventrotemporal region. Black arrowheads, displacedamacrine cells. c: Electron micrograph shows nuclei (arrows) withinthe inner plexiform layer presumed to be displaced amacrine cells,which move from the inner plexiform layer to the ganglion cell layer(GCL) between 50 and 60 hpf. d: The first outer segments (os) arevisible within the ventronasal patch by 54 hpf with electron micros-copy. A distinct boundary (dashed line) lies between the nasal region(NR) of precocious development and the immature differentiationtemporal (TR) to the midline. Unlike the temporal region, the OPL(bracket) is characterized by a lightly staining anuclear region adja-cent to the ONL. The pigment epithelium (Pe) is broad and possessesspindle-shaped and round pigment granules, mitochondria (smallarrows), and nuclei (n). Also, the development of inner (is) and outer(os) segments is associated with an expansion of the interphotorecep-tor space (ips). (This expansion may be artifactually exaggerated dueto slight tissue shrinkage.) Although small inner segments are presenttemporal to the midline, the interphotoreceptor space and outersegments are not evident, and the Pe is much thinner (asterisks). Ha,hyaloid artery. Scale bars 5 45 µm in a, 9.5 µm in b, 6 µm in c, 5 µm in d.
522 E.A. SCHMITT AND J.E. DOWLING

Figure 5
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 523

along the midline of the INL extending from fusiformnuclei (not shown). Concomitant with cell differentiation,the INL has increased by <8 µm and now averages<28–30 µm in width.
Outer retina. Within the OPL, TEM shows the firstsigns of the photoreceptor synaptic terminals within boththe ventronasal patch and the region dorsal to the opticnerve (st in Fig. 6f). Photoreceptor synaptic terminals arecharacterized by relatively darkly staining cytoplasm dueto an extremely high-density of ribosomes and polyribo-somes. Although synaptic ribbons are not present at 60hpf, they do appear within a small number of terminals inthe ventronasal patch by 62 hpf. Unlike other teleosts (e.g.,guppy, salmonids), the synaptic terminals of rods in thezebrafish are not easily distinguished from those of cones,where they develop together in the ventral patch (Kunz etal., 1983; Schmitt and Kunz, 1989). During this period, thenumber of neural processes in the OPL continues toincrease (np in Fig. 6f). Centrioles and diplosomes arefrequently observed within the processes. Outside of theventronasal patch, only a small number of lightly stainingpostsynaptic processes extend into the photoreceptor syn-aptic terminals. Electron-dense material is frequentlyfound along the membranes of the processes. As notedpreviously, neural processes that contain glycogen gran-ules (g) are believed to be those of horizontal cells (g andarrowheads, respectively, in Fig. 6f). The migratory neuro-blasts noted within the OPL of the ventronasal patch at 50hpf are not observed where photoreceptor terminals arepresent but are found in peripheral regions where retinaldevelopment is less advanced.
Dorsal to the optic nerve, photoreceptor outer segmentsare detected only by TEM, whereas those in the ventralpatch are now visible by LM at 60 hpf (compare Fig. 6b andc). This difference, reported by Kljavin (1987), reflects inpart the rapid development of rod outer segments withinthe ventral patch, where they now average <3 µm inlength (ros in Fig. 6c,d). However, the length of cone outer
segments is no more than 1.8 µm both within the ventralpatch and the central retina and are generally only visibleby TEM (cos in Fig. 6d). Photoreceptor inner segmentshave increased in size and extend 3–3.5 µm beyond theouter limiting membrane. The ellipsoids are now distinctfrom the myoids; large mitochondria are localized withinthe apical ellipsoid region, and elaborate Golgi and smoothand rough endoplasmic reticula are found in the myoidregion adjacent to the nucleus (Fig. 6d, inset). Smallprocesses of the pigment epithelium have begun to extendbetween the photoreceptors, and an extensive array ofintercellular organelles is present within the Pe, includingrough and smooth endoplasmic reticula, and free ribo-somes. Many of the pigment granules are patchy andappear as membrane-bound clusters of miniature gran-ules (arrows in Fig. 6d). These are similar to thosereported previously in developing teleosts and are pre-sumed to represent melanin granules in early stages ofmaturation (Schmitt and Kunz, 1989).
Ventronasal patch. In sagittal section, the ventralpatch continues to remain somewhat conspicuous as com-pared with other retinal regions, primarily because of thepresence of photoreceptor outer segments (Fig. 7a,c). How-ever, the precocious development noted previously withinthe ventronasal patch is less marked because the ONL andOPL are now well formed throughout the central retina,including in the dorsal region (compare Figs. 5a and 7a).These layers measure <8 and <2.5 µm, respectively.Although careful LM inspection shows the advanced devel-opment of horizontal cells within the ventronasal patch(i.e., more elongated and less heterochromatic; arrows inFig. 7c), additional differences are detectable only by TEM.For example, in sagittal sections of the central retina,photoreceptor synaptic terminals in the ventral patch arelarger and possess a greater number of postsynapticprocesses, whereas the development of those within thedorsal and temporal retina is significantly delayed. Synap-tic terminals in the nasal region show an intermediate
Fig. 6. Light and electron micrographs of transverse sections alongthe dorsal–ventral axis (D = V) at 60 hours postfertilization (hpf).a: Lamination has spread into the dorsal and ventral peripheries(asterisks) and into the region directly ventral to the optic nerve.Unlike at 50 hpf, pyknotic nuclei (short arrows) are found within thenuclear layers, and mitotic profiles are rarely observed. The opticnerve has retained a ‘‘forked’’ appearance as axons from the dorsal andventral regions converge at the vitreal surface (thin arrows). Round,darkly staining nuclei of displaced amacrine cells are regularly spacedalong the scleral edge of the ganglion cell layer (small arrows). Thedorsal (b5) and ventral region (c5) regions are shown at a highermagnification in Figures 6b,c, respectively. DP, dorsal periphery.b: The orientation of the photomicrograph is shifted 90° in relation tothat shown in a. Dorsal is to the right. Lamination now extends withinthe retina directly ventral to the optic nerve (small arrows). Darklystaining bipolar cells (B) are now evident within the mid-region of theinner nuclear layer (INL). Ganglion (G) and amacrine (A) cells arelarger and possess a greater amount of perinuclear cytoplasm. Thenuclei of horizontal cells (H) now appear elongated. The outer plexi-form layer (broad arrow) is now continuous, and nuclei are moreorganized within the outer nuclear layer (ONL). Although photorecep-tor outer segments are not observed, inner segments (curved arrows)appear as a lightly staining boundary adjacent to the pigmentepithelium (arrowheads). Arrow, pyknotic nucleus. c: In the ventralregion, all retinal laminae now extend dorsal, up to the optic nerve(ON). Small outer segments of both rods (ros) and cones (cos) can beseen. The pigment epithelium is broader where outer segments havebegun to develop (arrowheads). A single displaced amacrine cell (smallarrow) is observed at the scleral edge of the ganglion cell layer (G).
d: Developing inner and outer segments within the ventronasal patchshown by trasmission electron microscopy. Outer segments of rods(ros) are best discerned from those of cones (cos) by discontinuousouter segment discs (arrowhead). Zonulae adherentes (white arrows)between Muller cells (M) and ellipsoids (e) form the outer limitingmembrane. Pigment granules (black arrows) of the Pe appear to becomposed of numerous miniature granules. RN, rod nucleus. Inset:Ellipsoids (e) possess numerous mitochondria, whereas myoids (my)contain elaborate endoplasmic reticulium adjacent to the nucleus (n).The connecting cilium with an associated centriole (arrow) is presentprior to the development of outer segment discs. e: A conventionalsynapse (curved arrow) within the inner plexiform layer exhibits asmall cluster of synaptic vesicles (sv) at the presynaptic membraneand electron-dense material coats the postsynaptic membrane (aster-isk). The presynaptic profile is presumed to be that of an amacrine cell(A) because it possesses synaptic vesicles. Small profiles devoid ofvesicles are likely those of ganglion cells (G). f: The synaptic terminals(st) of rods and cones, identified by electron-dense cytoplasm, rich inribosomes and synaptic vesicles now extend within the outer plexiformlayer (OPL; arrows). Electron-lucent neural processes (np) extendfrom the OPL to invaginate within the terminals. Those that containglycogen (arrowheads) are believed to be those associated with horizon-tal cells (H) because glycogen (g) is abundant within the perinuclearcytoplasm. Neural processes (np) adjacent to the horizontal cellsomata are expanded in size and often contain centrioles (c). The firstsynaptic ribbons (sr) appear within the ventronasal patch after 62 hpf.CN, cone nucleus. Scale bars 5 48 µm in a, 15 µm in b,c, 1.5 µm in dand 4 µm in inset, 0.5 µm in e, 5 µm in f.
524 E.A. SCHMITT AND J.E. DOWLING

Figure 6
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 525

stage of development. In addition, TEM observations ofsagittal sections show that, although outer segments areup to 3 µm in length within the ventronasal patch (Fig. 6d),outer segments are shorter than 1 µm within the nasalretina and are not yet observed in the dorsal and temporalretina (not shown).
3 days postfertilization (70–74 hpf)
The majority of embryos hatch by 72 hpf; they are nowreferred to as larvae (Kimmel et al., 1995). At this time,they begin to display visual and electroretinographic re-sponses (see Discussion and Table 1). TEM observationssuggest that the basic vertical synaptic pathways betweenphotoreceptors and ganglion cells are completed between70 and 74 hpf. Ribbon synapses between bipolar andganglion cells are occasionally found within the IPL, andthe bipolar cell dendrites, in combination with horizontalcell processes, form triads at photoreceptor synaptic rib-bons in the OPL. Although these synaptic developmentsare observed within the ventral patch before 70 hpf,sagittal sections at 74 hpf show that ribbon synapses haveformed within both the OPL and IPL of all regions,including the dorsal and temporal retina.
70 hours postfertilization
Inner retina. Mature ribbon synapses within theaxon terminals of bipolar cells in the ventronasal patch arestill rare at 70 hpf (Fig. 8g). These ribbons generallymeasure less than 100 nm in length and are opposed byeither one or two postsynaptic processes. Despite the lackof mature ribbon synapses, small electron-dense profiles,presumed to be ribbons in early stages of formation, occurmore frequently. In contrast, the number of conventionalsynapses has greatly increased throughout the retina by70 hpf, including both the dorsal and temporal regions,where development is less advanced.
Outer retina. Synaptic ribbons within the synapticterminals of photoreceptors are observed outside the ven-tronasal patch by 70 hpf. Surprisingly, these ribbons arealso found within the nasal and temporal regions, whereoverall photoreceptor differentiation is much less ad-vanced. Many of the ribbons do not appear functionalbecause they appear to ‘‘float’’ within the cytoplasm, unas-sociated with postsynaptic processes. However, as thenumbers of postsynaptic processes increase and penetratemore deeply within the synaptic terminals, ribbons are
occasionally opposed by one or more postsynaptic pro-cesses. A small percentage of the synapses appears func-tional because synaptic vesicles (45 nm) surround theribbon and electron-dense material coats the postsynapticdendrites (Fig. 8f).
Within the temporal and dorsal retina, the appearanceof photoreceptor synaptic ribbons does not appear to bestrictly tied to the development of either outer segments orthe number of invaginating postsynaptic processes. Forexample, although synaptic ribbons are present, outersegments are rarely observed, and those that have formedare shorter than 1 µm. Furthermore, synaptic terminalsare smaller and possess a meager number of postsynapticprocesses as compared with those in the ventronasal patchor nasal retina. The presence of ribbon synapses within theOPL and IPL at 70 hpf implies that the basic ‘‘vertical’’pathways among photoreceptors, bipolar, and ganglioncells have been established. Nevertheless, such synapsesare still relatively rare at this time.
Ventronasal patch. Sagittal sections show that theeye cup rapidly expands between 60 and 70 hpf andaverages <235 µm along the dorsal–ventral axis (compareFig. 7a and b). The ventral patch continues to be lessconspicuous as differentiation across the retina becomesmore comparable (VP in Fig. 7b). Unlike the previousstages, a widened interphotoreceptor space is no longerevident, and the Pe has become much broader within theventral patch (compare Fig. 7c and f). This change is likelythe result of the extension of Pe cytoplasmic processesbetween the photoreceptor inner segments. Rod outersegments are longer than 4 µm in the ventronasal patchand are now observed extending from the patch into thetemporal region (arrows in Fig. 7f). Unlike the topographi-cal spread in the differentiation of cones and ganglion cells,rods are very rarely observed extending from the ventrona-sal patch in the nasal direction at this time. This temporalspread of rod photoreceptors corresponds to data obtainedby in situ hybridization with a riboprobe against rhodopsinmRNA at 60 hpf when rhodopsin expression extends fromthe ventral patch into the temporal retina in wholemountembryos (small arrows in Fig. 7e).
74 hours postfertilization
Photoreceptor differentiation progresses rapidly suchthat by 74 hpf outer segments are observed in transversesections across the retina near to the dorsal and ventral
Fig. 7. Sagittal sections taken along the temporal–nasal axis(T = N for all panels) through the central retina at the level of theoptic nerve (ON) at 60 hours postfertilization (hpf) (a,c,d) and 70 hpf(b,f). a: At 60 hpf, the ventronasal patch (VP) remains distinctiveprimarily due to the precocious development of photoreceptor innerand outer segments and the expansion of the interphotoreceptorspace. The outer plexiform layer (OPL) has formed across all regions ofthe central retina, including the dorsal region (DR). The inner regionof the inner nuclear layer (INL) is distinct from the outer regionbecause of more lightly staining amacrine cells (A) and the darklystaining bipolar cells (B), respectively. The ventral region (c5) is shownat higher magnification in c; arrowheads, pigment epithelium. b: By70 hpf, the eye cup has increased significantly in size. The ventronasalpatch (VP) is less conspicuous as differentiation has become morecomparable across the retina. The pigment epithelium has becomebroader (arrowheads) and extends within the interphotoreceptorspace. Darkly staining bipolar cells (B) are distinct from more lightlystaining amacrine cells (A). The ventral region (f5) is shown at highermagnification in f. c: The precocious development of photoreceptor
outer segments (os) and horizontal cells (arrows) within the ventrona-sal patch is observed at 60 hpf. With the exception of photoreceptordevelopment, cell differentiation temporal (TR) to the midline (dashedline) is now more comparable to that within the ventronasal patchcompared with 50 hpf. NR, nasal region. d: The outer plexiform layer(thin arrows) is well formed within the dorsal region, and the firstindications of photoreceptor inner segments (curved arrows) areobserved at 60 hpf. Pyknotic cells (arrowheads) are found in the INL.ONL, outer nuclear layer. e: In situ hybridization of rhodopsin mRNAin an embryonic wholemount at 60 hpf. Lateral view shows robust rodstaining within the ventronasal patch (VP) and a spread of rhodopsinstaining across the choroid fissure (dotted line) into the ventrotempo-ral region (small arrows). L, lens; y, yolk. f: The ventronasal patch (VP)is not as conspicuous at 70 hpf and is identified primarily by thepresence of outer segments (white arrows). Rod outer segments (blackarrows) also extend across the midline (dashed line) into the temporalregion (TR). NR, nasal region; G, ganglion cells; D = V, dorsal–ventralaxis for all panels. Scale bars 5 50 µm in a,b, 20 µm in c,d, 100 µm in e,25 µm in f.
526 E.A. SCHMITT AND J.E. DOWLING

peripheral margins (Fig. 8a). Also, the eye cup substan-tially increases in size and now measures <240 µm alongthe dorsal–ventral axis. The retina averages <80 µm inwidth, and a germinal zone (gz) is now seen, which persists
for life in teleosts (Raymond, 1985). At this stage, thedorsal germinal zone generally appears slightly broader(<30–40 µm) than the ventral germinal zone (<25 µm; gzin Fig. 8a). This is especially noticeable because photorecep-
Figure 7
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 527

tor outer segments extend much closer to the ventralgerminal zone than to the dorsal germinal zone (arrow-heads in Fig. 8a). In addition, preliminary observationssuggest that the size of the germinal zones is variableduring the first month of development. They are particu-larly narrow between 7 and 14 days postfertilizationbefore becoming broader by day 21 postfertilization (unpub-lished results).
The optic nerve has enlarged to <14 µm in diameter by74 hpf and has retained the forked appearance first notedon day 2. Axons within the ventral half of the optic nerveappear more tightly packed than those positioned dorsally,and the hyaloid artery containing nucleated blood cells isobserved along the ventral border of the optic nerve(asterisks and small arrows, respectively, in Fig. 8a).
Although rod outer segments are maximally 5 µm inlength within the ventronasal patch (arrows in Fig. 8c),the outer segments of cones are maximally 3 µm in lengthboth within the ventronasal patch and the region dorsal tothe optic nerve (arrows in Fig. 8d). In addition, transversesections show large outer segments of 4–5 µm near thedorsal periphery (ros in Fig. 8b). TEM confirms that theseare outer segments of developing rods present within asomewhat restricted region near the dorsal periphery (Fig.8e with inset). Unlike the single layer of nuclei within theONL of the central retina, a layer of staggered photorecep-tor nuclei is present in association with these dorsal rods(compare Fig. 8b and d). Rod outer segments are notevident at the extreme dorsal periphery. However, smallcone outer segments accompanied by a single layer ofdensely packed oval nuclei in the ONL are observed (os inFig. 8b). A large number of heterochromatic neuroblastsare detected within the INL in this peripheral region(thick arrow). These neuroblasts appear to be particularlyabundant in regions where rod differentiation is takingplace and possibly include rod progenitor cells characteris-tic of the teleost retina (thick arrows in Fig. 8b,c). UnderTEM, a darkly staining neuroblast is seen occasionally toextend across the OPL in these regions, suggesting thatprogenitor cells migrate from the INL to the ONL (notshown). However, these darkly staining neuroblasts arerarely observed in the central retina, where overt rodgenesis has not yet commenced (Fig. 8d). The Pe ranges
between 5 and 7 µm in width and is especially broad inregions associated with developing rods, e.g., ventronasalpatch and dorsal periphery. Small lipid droplets andphagosomes are also prominent in association with roddevelopment (L in Fig. 8e). The extremely slender nuclei ofthe Pe measure less than 1 µm wide but up to 12 µm longin sections (n in Fig. 8e).
Photoreceptor synaptic terminals have enlarged, andribbon triads are more common as the number of invaginat-ing postsynaptic processes has increased (Fig. 9e). Synap-tic ribbons are 150–400 nm in length but can extend up to500 nm and are accompanied by numerous synapticvesicles. Although infrequent, two ribbons are observedwithin cone pedicles, especially within the nasal region ofthe retina, suggesting cone differentiation is slightly moreadvanced.
The inner plexiform layer, 12–14 µm in width, hasdeveloped considerably since 60 hpf. Axon terminals ofbipolar cells appear to be organized within four distinctstrata or sublaminae (not shown). Two strata lie on eitherside of the IPL midline, whereas the third and fourthstrata lie <4–5 µm from the INL and ganglion cell layer(GCL), respectively. The bipolar axon terminals are largeand irregularly shaped and are 2–2.5 µm across. Someprofiles, however, are up to 3.5 µm across. These processesare distinctive because they contain a homogeneous distri-bution of synaptic vesicles and large mitochondria, up to<800 nm in diameter. Small axons <0.5 µm in diameter,possessing a high density of microtubules, extend betweenthe strata and are generally presumed to be those ofbipolar cells. Muller cell fibers, which also traverse theIPL, are distinct from these axons because they are morelightly staining, devoid of synaptic vesicles, and contain ahigh density of glycogen granules and ribosomes. Conven-tional synapses have greatly increased in number and nowaverage 150 nm in length. They are most frequently foundin association with the processes of amacrine and ganglioncells. Synaptic ribbons are still seen only occasionallywithin the terminals of bipolar cells and average <150 nmin length (Fig. 8g). However, because processes of bothganglion and amacrine cells oppose these ribbons andclusters of synaptic vesicles are present around them, theyappear functional.
Fig. 8. Transverse sections along the dorsal–ventral axis (D = V)at 74 hours postfertilization (hpf). a: The eye cup has becomeelongated along the dorsal–ventral axis, and the total width of theretina has increased as the layers have broadened. Outer segments(arrowheads) extend to the germinal zone (gz) in the ventral retina.The hyaloid artery (triple arrows) extends parallel to the ventralborder of the optic nerve. Axons within the ventral region of the opticnerve (asterisk) appear more densely packed as compared with thosepositioned more dorsally. The dorsal periphery (b5), ventral region (c5)and region directly doral to the optic nerve (d5) are shown in b–d,respectively. b: Rod outer segments (ros) are clearly visible within thedorsal periphery. Differentiating horizontal (H), bipolar (B), andamacrine (A) cells lie within the inner nuclear layer. More peripher-ally, small outer segments (os) of less mature photoreceptors areobserved, and a large number of darkly staining neuroblasts (thickarrow) lie within the inner nuclear layer. ONL, outer nuclear layer. c:Densely packed axons lie within the ventral region of the optic nerve(asterisk). Numerous rods (arrows) are present within the ventronasalpatch (VP). Photoreceptors on either side of the patch appear lessdeveloped, and darkly staining neuroblasts (thick arrows) lie withinthe adjacent inner nuclear layer. G, ganglion cells; ONL, outer nuclearlayer. d: The orientation of the photomicrograph is shifted 90° inrelation to that shown in a. Dorsal is to the right. Dorsal to the optic
nerve (ON), a single layer of cone nuclei (CN) lies within the ONL andis associated with small outer segments (long arrows). Horizontal cells(H), bipolar (B), and amacrine (A) cells are observed within the innernuclear layer (INL). Darkly staining neuroblasts are not observed.e: Electron microscopy shows rods (R) and cones (C) develop togetherwithin the dorsal periphery. Rods are identified by discontinuous outersegment discs (white arrow in inset). Rod nuclei (RN) are situatedvitreal to those of cones (CN) within the outer nuclear layer. Roddevelopment is generally accompanied by the presence of lipid drop-lets (L) within the pigment epithelium. The nuclei (n) of the pigmentepithelium have become elongated. Inset: Discontinuous discs (whitearrow) of the rod outer segment (R) with connecting cilium (cc) andassociated lipid droplet (L). f: Photoreceptor ribbon synapse forms atriad: two lateral horizontal cell processes (1,2) and a third centralbipolar cell process (3) oppose the ribbon. Synaptic vesicles (sv) andelectron-dense postsynaptic zones (arrows) are observed adjacent tothe ribbon. g: Presumptive synaptic ribbon (sr) within a bipolar cellaxon terminal in the IPL is opposed by two postsynaptic processes(1,2). Synaptic vesicles (arrowhead) are present, and electron-densematerial (arrow) is associated with the postsynaptic membranes.Scale bars 5 26 µm in a, 12 µm in b–d, 7.5 µm in e and 2 µm in inset,120 nm in f and 90 nm in g.
528 E.A. SCHMITT AND J.E. DOWLING

Figure 8
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 529

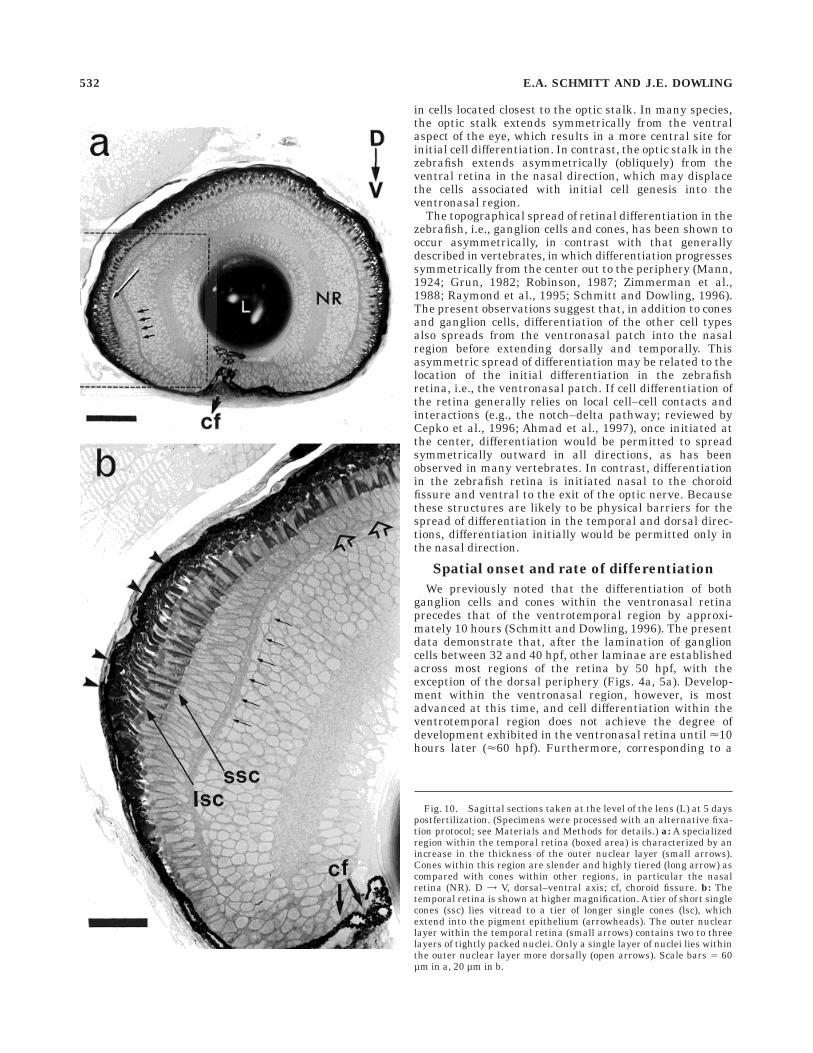
Observations of sagittal sections at 74 hpf show somedifferences among the various regions of the central retina(Fig. 9a–d). In the temporal region, for example, a promi-nent bulge is exhibited by the photoreceptor layer at thetemporal pole (Fig. 9a). The bulge is primarily the result ofan increased number of photoreceptor nuclei within theONL, which measures 18 µm across. In the other regions,the ONL is only 10–12 µm wide and possesses fewer thanthree layers of nuclei (Fig. 9b–d). Cone nuclei are predomi-nately observed in this temporal location and, althoughthey are of a length (<3–5 µm) similar to those in otherregions, these nuclei are <0.5 µm thinner (<2–2.6 µmwide), thereby affording a tighter packing of the nucleiwithin the bulge. Unlike photoreceptors in other regions,cones in the temporal region exhibit the first clear evi-dence of tiering because their outer segments lie in threelayers (compare Fig. 9a and b–d). The outer segmentslying nearest to the ONL appear to be the longest, <3.5µm. Outer segments within the second layer average 2.6µm in length, whereas those located most sclerad withinthe Pe are the shortest, averaging <2.0 µm in length. Inaddition, both the Pe and OPL are relatively broad at thetemporal pole (<8.5 and 3.0 µm wide, respectively). Rodsare rarely found within the bulge but are observed inconsiderable numbers more ventrally. These featureswithin the temporal retina are first observed by LM insagittal sections on day 4 but become more pronounced byday 5 postfertilization (Fig. 10a,b). These characteristicswithin the temporal retina persist at least through 21 dayspostfertilization (Schmitt and Dowling, in preparation).
In the ventral retina, rod nuclei, unlike other regions,are regularly spaced within the ONL at 74 hpf (arrows inFig. 9c). A layer of rod nuclei is distinctive because thenuclei are more darkly staining and lie vitread to approxi-mately two layers of cone nuclei. Rod nuclei are seldomseen within the nasal retina at this time but are observedoccasionally in the dorsal region (arrows in Figs. 9b,d,respectively). As noted earlier, the Pe in the ventral retinais broad (<8 µm wide) and contains numerous lipiddroplets <2 µm in diameter. The outer segments of bothrods and cones extend deep into the Pe (Fig. 9c). Incontrast, the Pe is only 5 µm wide in the dorsal retina anddoes not possess lipid droplets (Fig. 9d). Also, rod inner andouter segments are frequently oriented sideways withinthe interphotoreceptor space rather than extending upinto the Pe. Although this is seen to a lesser extent withcones, their outer segments also can lie at an angle suchthat well-formed calycal processes can be observed aroundthe circumference of these dorsal cones (arrows in Fig. 9f).
DISCUSSION
Retinal differentiation in the zebrafish generally followsthe inner-to-outer- (vitreoscleral) directed sequence charac-teristic of all vertebrate species (for review, see Grun,1982). It commences with the differentiation of ganglioncells along the vitreal border followed by the appearance ofthe IPL and amacrine cells. Development continues in thescleral direction, with a band of photoreceptor nucleiappearing within the ONL and the formation of the OPL.Similar to other vertebrates, the notable exception to theoverall vitreoscleral directed sequence is the late differen-tiation of bipolar cells within the INL, which occurs afterthat of horizontal cells and photoreceptors (Table 1).
Synaptogenesis
In addition to cell differentiation, the fundamental eventsof synaptogenesis also progress in a scleral direction, withthe exception of bipolar cell synapses (Grun, 1982). Thedevelopment of the IPL occurs before that of the OPL, andthe outgrowth of vitreally directed axons of ganglion andbipolar cells occurs before that of their sclerally directeddendrites. Within the OPL, the differentiation of vitreallypositioned horizontal cell dendrites precedes that of thesclerally positioned synaptic terminals of the photorecep-tors (Table 1). The formation of the first photoreceptorouter segments roughly coincides with the development ofthe first conventional synapses within the IPL (<60 hpf),which is followed by the appearance of the first photorecep-tor synaptic ribbons (65 hpf). Corresponding to the latedevelopment of bipolar cells, the formation of synapticribbons within bipolar cell axon terminals in the IPL is thelast stage to be noted (<70–72 hpf).
Ventronasal patch
In most vertebrates, retinal development is initiated in acentral location, roughly dorsal to the optic nerve head(Mann, 1928; Grun, 1982; Robinson, 1987; Straznicky andChehade, 1987; LaVail et al., 1991; Hendrickson, 1995). Incontrast, the initiation of ganglion cell and cone photorecep-tor differentiation in the zebrafish occurs within theventronasal retina, adjacent to the edge of the choroidfissure (Burrill and Easter, 1994, 1995; Schmitt and Dowl-ing, 1994, 1996; Raymond et al., 1995). The present dataindicate that the differentiation of the other major celltypes including amacrine, bipolar, and horizontal cells alsocommences from this ventronasal location (see Fig. 5b).Burrill and Easter (1995) suggested that, in both zebrafishand higher vertebrates, retinal differentiation is initiated
Fig. 9. Electron micrographs of photoreceptors in a sagittal sectionat 74 hours postfertilization (hpf). a: In the temporal region (TR), abulge (small arrows) occurs within the photoreceptor cell layer andpigment epithelium. Cone nuclei (CN) are tightly packed within theouter nuclear layer (ONL). Tiers of cones have begun to emergebecause outer segments are organized within three rows (1–3). Rodsare not observed. b: Cones are primarily found within the nasal region(NR). A rod nucleus (arrows), identified by dark heterochromaticstaining, is located vitread to those of cones (CN). Cone outer segments(C) are not well organized. Synaptic terminals (arrowheads) extendwithin the outer plexiform layer. c: In the ventral region (VR), largeouter segments of rods (R) extend within the pigment epitheliumsclerad to those of cones (C). Lipid droplets (L) are observed. Inaddition to a staggered row of cone nuclei (CN), a layer of rod nuclei(arrows) are found along the outer plexiform layer. Synaptic terminals
are observed (arrowheads). d: In the dorsal region (DR), photorecep-tors are occasionally oriented obliquely within the interphotoreceptorspace. Numerous outer segments of cones (C) and a presumptive rod(R) outer segments are observed. Two rod nuclei (arrows) lie vitread totwo rows of cone nuclei (CN), and a number of well-formed synapticterminals are observed (arrowheads). e: Both the nucleus (RN) andsynaptic terminal (arrows) of a rod are darkly stained compared withthose of cones (CN). Synaptic ribbons (sr) lie adjacent to the conenuclei. Lightly staining cytoplasm of a neural process (np) lies betweentwo horizontal cell somata (H) and includes a centriole (c). f: In thedorsal retina, cone outer segments (cos) are shown in cross sectionbecause they are oriented sideways within the interphotoreceptorspace (asterisks). Well-formed calycal processes (arrows) are observedaround the circumference of the outer segment. Pg, pigment granules.Scale bars 5 13 µm in a–d, 1.5 µm in e, 2.5 µm in f.
530 E.A. SCHMITT AND J.E. DOWLING

Figure 9
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 531
in cells located closest to the optic stalk. In many species,the optic stalk extends symmetrically from the ventralaspect of the eye, which results in a more central site forinitial cell differentiation. In contrast, the optic stalk in thezebrafish extends asymmetrically (obliquely) from theventral retina in the nasal direction, which may displacethe cells associated with initial cell genesis into theventronasal region.
The topographical spread of retinal differentiation in thezebrafish, i.e., ganglion cells and cones, has been shown tooccur asymmetrically, in contrast with that generallydescribed in vertebrates, in which differentiation progressessymmetrically from the center out to the periphery (Mann,1924; Grun, 1982; Robinson, 1987; Zimmerman et al.,1988; Raymond et al., 1995; Schmitt and Dowling, 1996).The present observations suggest that, in addition to conesand ganglion cells, differentiation of the other cell typesalso spreads from the ventronasal patch into the nasalregion before extending dorsally and temporally. Thisasymmetric spread of differentiation may be related to thelocation of the initial differentiation in the zebrafishretina, i.e., the ventronasal patch. If cell differentiation ofthe retina generally relies on local cell–cell contacts andinteractions (e.g., the notch–delta pathway; reviewed byCepko et al., 1996; Ahmad et al., 1997), once initiated atthe center, differentiation would be permitted to spreadsymmetrically outward in all directions, as has beenobserved in many vertebrates. In contrast, differentiationin the zebrafish retina is initiated nasal to the choroidfissure and ventral to the exit of the optic nerve. Becausethese structures are likely to be physical barriers for thespread of differentiation in the temporal and dorsal direc-tions, differentiation initially would be permitted only inthe nasal direction.
Spatial onset and rate of differentiation
We previously noted that the differentiation of bothganglion cells and cones within the ventronasal retinaprecedes that of the ventrotemporal region by approxi-mately 10 hours (Schmitt and Dowling, 1996). The presentdata demonstrate that, after the lamination of ganglioncells between 32 and 40 hpf, other laminae are establishedacross most regions of the retina by 50 hpf, with theexception of the dorsal periphery (Figs. 4a, 5a). Develop-ment within the ventronasal region, however, is mostadvanced at this time, and cell differentiation within theventrotemporal region does not achieve the degree ofdevelopment exhibited in the ventronasal retina until <10hours later (<60 hpf). Furthermore, corresponding to a
Fig. 10. Sagittal sections taken at the level of the lens (L) at 5 dayspostfertilization. (Specimens were processed with an alternative fixa-tion protocol; see Materials and Methods for details.) a: A specializedregion within the temporal retina (boxed area) is characterized by anincrease in the thickness of the outer nuclear layer (small arrows).Cones within this region are slender and highly tiered (long arrow) ascompared with cones within other regions, in particular the nasalretina (NR). D = V, dorsal–ventral axis; cf, choroid fissure. b: Thetemporal retina is shown at higher magnification. A tier of short singlecones (ssc) lies vitread to a tier of longer single cones (lsc), whichextend into the pigment epithelium (arrowheads). The outer nuclearlayer within the temporal retina (small arrows) contains two to threelayers of tightly packed nuclei. Only a single layer of nuclei lies withinthe outer nuclear layer more dorsally (open arrows). Scale bars 5 60µm in a, 20 µm in b.
532 E.A. SCHMITT AND J.E. DOWLING

previous report, full lamination within the dorsal periph-ery is the last to occur, taking place between 50 and 60 hpf(Schmitt and Dowling, 1996). The present observationssuggest that the onset of differentiation is stringentlycontrolled because the differentiation of cells differs signifi-cantly even within a few microns, e.g., cells immediatelynasal and temporal to the ventronasal patch (see Fig.5a,b,d).
Although the initial onset of cell differentiation is notsynchronous across the retina, the rate of differentiationappears to differ between different regions in such a waythat development equilibrates to some degree between 60and 70 hpf. Sagittal sections clearly show that, althoughdifferentiation within the ventronasal region is still moreadvanced at 60 hpf, development within the ventral, nasal,dorsal, and temporal retina appears relatively comparableby 70 hpf (compare Fig. 7b and f). However, a slight delayin the development of ultrastructural features, e.g., synap-tic terminals, can still be detected in the ventrotemporalretina by using TEM.
By 74 hpf, however, a specialized area characterized by ahigh density and tiering of cones arises in the temporalretina, suggesting that cone differentiation has becomeaccelerated in this region (compare Fig. 9a–d). However,cone development within most of the ventral retina ap-pears to be less advanced despite the presence of a largenumber of well developed rods. This difference in the rateof rod and cone development may be attributed to therelatively high levels of retinoic acid (RA) present within
the zebrafish ventral retina, which retards the develop-ment of cones while accelerating that of rods (Marsh-Armstrong et al., 1994; Hyatt et al., 1996b). If this is true,low levels of RA may exist in the temporal retina, resultingin the opposite effect, i.e., an acceleration in cone develop-ment and a simultaneous inhibition of rod development.These possibilities concur with the fact that differentlevels of RA exist across the retina, which result from thedifferential distribution of the synthetic enzymes (re-viewed by Drager and McCaffery, 1997).
Rod differentiation
Unlike the other cell types, the topographical spread ofrod differentiation in the zebrafish does not proceed as acontinuous wave across the retina, despite being initiatedalong the nasal edge of the choroid fissure (Raymond et al.,1995; Schmitt and Dowling, 1996). After the formation ofan initial patch of rods nasal to the fissure, both in situhybridization and LM demonstrate that rods extend intothe ventrotemporal region before they are observed acrossthe nasal and dorsal regions (Fig. 7e,f vs. Fig. 8b,e). (Weconsistently find that developing rods are detected <10hours earlier when using in situ hybridization, or immuno-labeling, for rhodopsin vs. conventional LM.) In the chick,a marked difference in the initial topographical spread ofrods versus that of cones is also observed, which appears tobe related to the final topography of these cell types in theadult (Bruhn and Cepko, 1996). This possibility may
TABLE 1. Comparison of Retinal Development in the Zebrafish Between 32 and 74 hpf With That of Retinotectal Projections and BehaviorBased on the Present Data or Those by Other Investigators1
Hourspost-fertil-ization
Outer retina Inner retinaGanglioncell layer
Otherfeatures
Retinotectalprojections
Behavior/retinal
responsesPhotoreceptor OPL INL IPL
32
36
40
50
55
60
65
70
72
74
Inner segment for-mation/opsinexpression inventronasalpatch
Outer segments inventronasalpatch
Outer segmentsdorsal/nasal tooptic nerve
Rhodsopsin expres-sion in dorsalretina
Specialized area intemporal retinafirst evident
Horizontal cell pro-cesses withzonulae adher-entes
Rod and cone syn-aptic terminals
Synaptic ribbons(dyads andtriads) in ventro-nasal patch
Synaptic ribbons inother regions
Differentiation ofamacrine andhorizontal cells
Differentiation ofbipolar cells
Amacrine and gan-glion cell pro-cesses/presump-tive synapticzones
Bipolar cell pro-cesses/conven-tional synapses
Bipolar cell ribbonsynapses in ven-tronasal patch
Bipolar cell ribbonsynapses inother regions/four sublaminae
Differentiation inventronasalpatch
Ganglion cell birth-days complete/differentiation inventrotemporalregion
Displaced ama-crine cells inintraplexiformlocation
Displaced ama-crine cells inganglion celllayer
Pigmentationwithin pigmentepithelium
Optic stalk attenu-ation commences
Optic stalk attenu-ation complete
GABA and trkimmunoreac-tivity in opticnerve
GABA and trkimmunoreac-tivity in OPL,INL, IPL
Extraocular eyemuscle matura-tion
Emmetropizationof eye and lens
Optic axons exitthe retina
Optic axons reachoptic chiasm
Optic axons reachtectum
Arborizationwithin ninefields in tectum
Arborizationwithin tenthfield in tectum
Weak touchresponse
Vigorous touchresponses
Hatching begins
First visual-evokedresponses
First electroretino-graphicresponses
Optokineticresponses
1Branchek (1984), Nawrocki (1985), Stuermer (1988), Burrill and Easter (1994), Sandell et al. (1994a,b), Kimmel et al. (1995), Raymond et al. (1995), and Easter and Nicola (1996).OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GABA, gamma-aminobytyric acid; trk, tyrosine receptor kinase.
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 533

explain the different topographical patterns of rod differen-tiation observed in the zebrafish versus that in the goldfish(Stenkamp et al., 1996). The more random spread ofdeveloping rods corresponds to investigations that haveindicated that diffusible factors (e.g., retinoic acid, ciliaryneurotrophic factor, sonic hedgehog) rather than direct cellcontacts influence rod differentiation (Stenkamp et al.,1993; Kelley et al., 1994; Fuhrman et al., 1995; Hyatt etal., 1996b; Ezzeddine et al., 1997; Levine et al., 1997).
Temporal region specialization
In a large number of vertebrates, retinal differentiationis initiated in the vicinity of a region that in the adultprovides high-acuity vision (e.g., area centralis in cat,fovea in primates; Robinson, 1987; Zimmerman et al.,1988; LaVail et al., 1991; Henrickson, 1995). Surprisingly,such a specialized retinal region in zebrafish does not arisewithin the ventronasal region but rather within the tempo-ral retina. However, the early differentiation of the ventralretina in zebrafish would be of great advantage to newlyhatched larvae between days 3 and 4, when they predomi-nately lie at the bottom of a stream or tank. Light fromobjects above falls on the ventral retina during this period.After day 4, larvae fill their swim bladders and becomeneutrally buoyant, which positions them throughout thewater column. Corresponding to this expansion of thevisual field, a distinct ‘‘area’’ exhibiting a high density ofcones (and a paucity of rods) arises in the temporal retinaat this time (Fig. 10a,b). The presence of this ‘‘areatemporalis’’ suggests that zebrafish possess better visualacuity in the anterior visual field. Such specialized areashave been described in numerous teleosts, in particularmarine species (Walls, 1942; Ito and Murakami, 1984;Collin and Pettigrew, 1988a,b). They are located predomi-nantly within the temporal retina and are seen especiallyin species that exhibit voluntary eye movements (Walls,1942). In zebrafish larvae, this specialized area is firstdetected at 74 hpf by TEM (Fig. 9a) and persists at leastthrough day 21 postfertilization (unpublished results).
Displaced amacrine cells
Between 50 and 60 hpf, a number of nuclei, presumed tobe displaced amacrine cells, is found throughout the IPL.In zebrafish, these cells are regularly spaced and are easilydistinguished by darkly staining cytoplasm. Althoughdisplaced amacrine cells were first identified by Cajal(1911) in fish, amphibians, reptiles, and birds, they haveonly recently been recognized as a standard feature of thevertebrate retina (Vaney, 1984; for review, see Maslandand Tauchi, 1987). These cells represent a sizable popula-tion composing as much as 38%, 50%, and 80% of theneuronal profiles within the ganglion cell layer in therabbit, rat, and cat, respectively (Hughes, 1985; Perry,1981; Wassle et al., 1987). Only a few investigators havecommented on the development of these cells (GenisGalvez et al., 1977; Hinds and Hinds, 1983; Spira et al.,1987). The present observations suggest that these cellsmigrate together from the INL to the GCL during a specifictime of development. These preliminary findings indicatethat the zebrafish retina may provide an advantageoussystem in which to examine the differentiation of thisparticular cell type and to identify the events that lead totheir final positioning within the ganglion cell layer.
Development of visual behavior
As in other teleosts (e.g., salmonids, sole, mackerel,whiting, Fundulus, cichlids, goldfish), zebrafish are with-out visual abilities at hatching, <60 hpf (2.5 days postfer-tilization; reviewed by Blaxter, 1975; Grun, 1982). Thefirst visually evoked responses are present in zebrafishlarvae between 68 and 72 hpf, and electroretinographicresponses can initially be generated at 72 hpf (Branchek,1984; Easter and Nicola, 1996; Table 1). Emmetropizationand functioning of extraocular muscles also occur by 72hpf, and the first optokinetic reflex responses are observedslightly later, between 73 and 80 hpf (Easter and Nicola,1996). The rate of retinogenesis is tightly coordinated withlarval development, so that the retina becomes functionalby 72 hpf (day 3), shortly before larvae become freeswimming and begin feeding (day 4; Kimmel et al., 1995).The development of initial photoreceptor outer segments(<60 hpf) and their synapses within the OPL (<65 hpf) arein place just before the initial appearance of visuallyevoked responses. Synaptic ribbons within the axon termi-nals of bipolar cells arise later (<70 hpf) and closelycoincide with the onset of visually evoked responses (Table1). Easter and Nicola (1996) noted that the maturation ofboth the retinal arborizations within the tectum and themotor abilities for the visually evoked response occurs wellbefore the onset of the response and concluded that thedevelopment of the connections between sensory andmotor systems in the brain may be the limiting factor forthe onset of visual responses. The present data suggestthat the relatively late development of synaptic ribbons inthe IPL may also be a key limiting factor for the onset ofvisually evoked responses in larvae. These synapses repre-sent the point when signals from the photoreceptors arerelayed to the ganglion cells before they can reach thevisual centers in the brain.
The rapid temporal synchrony among early retinogen-esis, functional ocular musculature, emmetropization, andinnervation of central targets is highly impressive (Table1). However, important synaptic features such as gapjunctions between horizontal cells, interreceptor contacts,spinules within the cone pedicles, serial and reciprocalsynapses within the IPL, etc., are still not obvious at theonset of visual behavior. Improved visual acuity, freeswimming, and the onset of photopositive behavior coin-cide with the appearance of such synaptic features in therainbow trout (Schmitt and Kunz, 1989). Thus, it is likelythat the improved acuity in zebrafish larvae after day 3(Clark, 1981) can be attributed to the development of thesefeatures, which are generally associated with the integra-tion of color and spatial information.
ACKNOWLEDGMENTS
We thank Bill McCarthy and the staff of the HarvardUniversity zebrafish facility for providing zebrafish embryos.This study was supported by NIH grant EY-00811 to J.E.D.
LITERATURE CITED
Ahmad I, Dooley CM, Polk DL. 1997. Delta-1 is a regulator of neurogenesisin the vertebrate retina. Dev Biol 185:92–103.
Blaxter JHS. 1975. The eyes of larval fish. In: Ali MA, editor. Vision inFishes: New Approaches in Research. New York: Plenum Press. p427–444.
Branchek T. 1984. The development of photoreceptors in the zebrafish,Brachydanio rerio. II. Function. J Comp Neurol 224:116–122.
534 E.A. SCHMITT AND J.E. DOWLING

Branchek T, Bremiller R. 1984. The development of the photoreceptors inthe zebrafish, Brachydanio rerio. I. Structure. J Comp Neurol 224:107–115.
Brockerhoff SE, Hurley JB, Janssen-Bienhold U, Neuhauss SCF, DrieverW, Dowling JE. 1995. A behavioral screen for isolating zebrafishmutants with visual system defects. Proc NatlAcad Sci U.S.A. 92:10545–10549.
Brockerhoff SE, Hurley JB, Niemi GA, Dowling JE. 1997. A new form ofinherited red-blindness identified in zebrafish. J Neurosci 17:4236–4242.
Bruhn SL, Cepko CL. 1996. Development of the pattern of photoreceptorsin the chick retina. J Neurosci 15:1430–1439.
Burrill JD, Easter, Jr. SS. 1994. The development of the retinofugalprojections in the embryonic and larval zebrafish, Brachydanio rerio. JComp Neurol 346:583–600.
Burrill, J.D. and S.S. Easter, Jr. 1995. The first retinal axons and theirmicroenvironment in zebrafish: Cryptic pioneers and the pretract. JNeurosci 15:2935–2947.
Cajal SR. 1911. Histologie du Systeme Nerveux de l’Homme et desVertebres, Vol. II. Paris: Maloine.
Cepko CL, Austin CP, Yang X, Alexiader M, Ezzeddine D. 1996. Cell fatedetermination in the vertebrate retina. Proc Natl Acad Sci U.S.A.93:589–595.
Clark DT. 1981. Visual Responses in Developing Zebrafish. Ph.D. thesis.Eugene: University of Oregon.
Collin SP, Pettigrew JD. 1988a. Retinal topography in reef teleosts I. Somespecies with well-developed areae but poorly-developed streaks. BrainBehav Evol 31:269–282.
Collin SP, Pettigrew JD. 1988b. Retinal topography in reef teleosts II. Somespecies with prominent horizontal streaks and high-density areae.Brain Behav Evol 31:283–295.
Dowling JE. 1987. The Retina—An Approachable Part of the Brain.Cambridge: Harvard University Press.
Drager, U.C. and P. McCaffery (1997) Retinoic acid and the development ofthe retina. Prog Retinal Eye Res 16:323–351.
Driever W, Solnica-Krezel L, Schier AF, Neuhaus SCF, Malicki J, StempleDL, Stainier DYR, Zwartkruis F, Abdelilah S, Ranginin Z, Belak J,Boggs C. 1996. A genetic screen for mutations affecting embryogenesisin zebrafish. Development 123:37–46.
Easter SS, Jr., Nicola GN. 1996. The development of vision in the zebrafish,Danio rerio. Dev Biol 180:646–663.
Ekker SC, Ungar AR, Greenstein P, von Kessler DP, Porter JA, Moon RT,Beachy P. 1995. Patterning activities of vertebrate hedgehog proteins inthe developing eye and brain. Curr Biol 5:944–955.
Ezzeddine ZD, Yang X, DeChiara T, Yancopoulos G, Cepko CL. 1997.Postmitotic cells fated to become rod photoreceptors can be respecifiedby CNTF treatment of the retina. Development 124:1055–1067.
Fadool JM, Brockerhoff SE, Hyatt GA, Dowling JE. 1997. Mutationsaffecting eye morphology in the developing zebrafish, Danio rerio. DevGenet 20:288–295.
Fini ME, Strissel KJ, West-Mays JA. 1997. Perspectives on eye develop-ment. Dev Genet 20:175–185.
Fuhrmann S, Kirsch M, Hofmann H-D. 1995. Ciliary neurotrophic factorpromotes chick photoreceptor development in vitro. Development 121:2695–2706.
Genis Gelavez JM, Puelles L, Prada C. 1977. Inverted (displaced) retinalamacrine cells and their embryonic development in the chick. ExpNeurol 56:151–157.
Graw J. 1996. Genetic aspects of embryonic eye development in verte-brates. Dev Genet 18:181–197.
Grun GG. 1982. The development of the vertebrate retina: A comparativesurvey. Adv Anat Embryol Cell Biol 78:1–84.
Haftner P, Granato M, Brand M, Mullins MC, Hammerschmidt M, KaneDA, Odenthal J, van Eeden FJM, Jiang Y-J, Heisenberg C-P, Kelsh RN,Furutani-Seiki M, Vogelsang E, Beuchle D, Schach U, Fabian C,Nusslein-Volhard C. 1996. The identification of genes with unique andessential functions in the development of the zebrafish, Danio rerio.Development 123:1–36.
Heisenberg C-P, Brand M, Jiang Y-J, Warga RM, Beuchle D, van EedenFJM, Furutani-Seiki M, Granato M, Haftner P, Hammerschmidt M,Kane DA, Mullins MC, Odenthal J, Nusslein-Volhard C. 1996. Genesinvolved in forebrain development in the zebrafish, Danio rerio. Devel-opment 123:191–203.
Henrickson AE. 1995. Primate foveal development: A microcosm of currentquestions in neurobiology. Invest Ophthalmol Vis Sci 35:3129–3133.
Hinds JW, Hinds PL. 1983. Development of retinal amacrine cells in themouse embryo: Evidence for two modes of formation. J Comp Neurol213:1–23.
Hughes A. 1985. New perspective in retinal organization. Prog Retinal Res4:243–313.
Hyatt GA, , Schmitt EA, Marsh-Armstrong N, McCaffery P, Drager UC,Dowling JE. 1996a. Retinoic Acid establishes ventral retinal character-istics. Development 122:195–204.
Hyatt GA, Schmitt EA, Fadool JM, Dowling JE. 1996b. Retinoic acid altersphotoreceptor development in vivo. Proc Natl Acad Sci U.S.A. 93:13298–13303.
Ito H, and Murakami T. 1984. Retinal ganglion cells in two teleost species,Sebastiscus marmoratus and Navodon modestus J Comp Neurol 229:80–96.
Kelley MW, Turner JK, Reh TA. 1994. Retinoic acid promotes differentia-tion of photoreceptors in vitro. Development 120:2091–2102.
Kimmel CB, Ballard WW, Kimmel S, Ullmann B, Schilling TF. 1995. Stagesof embryonic development of the zebrafish. Dev Dynam 203:253–310.
Kljavin I. 1987. Early development of photoreceptors in the ventral retinaof the zebrafish embryo. J Comp Neurol 260:461–471.
Kunz YW, Ennis S, Wise C. 1983. Ontogeny of the photoreceptors in theembryonic retina of the viviparous guppy, Poecilia reticulata P. CellTissue Res 230:469–486.
Lall AB, Brockerhoff SE, Dowling JE. 1997. Electroretinographic character-ization of nrc mutant larvae of zebrafish. Invest. Ophthamol Vis Sci38:S619.
Larison K, Bremiller R. 1990. Early onset of phenotype and cell patterningin the embryonic zebrafish retina. Development 109:567–576.
LaVail MA, Rappaport DH, Rakic P. 1991. Cytogenesis in the monkeyretina. J Comp Neurol 309:86–114.
Levine EM, Roelink H, Turner J, Reh TA. 1997. Sonic hedgehog promotesrod photoreceptor differentiation in mammalian retinal cells in vitro. JNeurosci 17:6277–6288.
Macdonald R, Wilson SW. 1996. Pax proteins and eye development. CurrOpin Neurobiol 6:49–56.
Malicki J, Neuhauss SCF, Schier AF, Solnica-Krezel L, Stemple DL,Stainier DYR, Abdelilah S, Rangini Z, Zwartkruis F, Driever W. 1996.Mutations affecting development of the zebrafish retina. Development123:263–273.
Mann IC. 1928. The regional differentiation of the vertebrate retina. Am JOphthalmol 11:515–526.
Marsh-Armstrong N, McCaffery P, Dowling JE, Drager UC. 1994. Retinoicacid is necessary for development of the ventral retina in zebrafish. ProcNatl Acad Sci U.S.A. 91:7286–7290.
Masland RL, Tauchi M. 1987. The cholinergic amacrine cell. TrendsNeurosci 9:218–223.
Nawrocki L W. 1985. Development of the Neural Retina in the Zebrafish,Brachydanio rerio. Ph.D. dissertation. Eugene: University of Oregon.
Oliver G, Gruss P. 1997. Current views on eye development. TrendsNeurosci 20:415–421.
Oxtoby E, Jowett T. 1993. Cloning the zebrafish Krox-20 gene (krx-20) andits expression during hindbrain development. Nucl Acids Res 21:1087–1095.
Perry VH. 1981. Evidence for an amacrine cell system in the ganglion celllayer of the rat retina. Neuroscience 6:931–941.
Raymond PA. 1985. The unique origin of rod photoreceptors in the teleostretina. Trends Neurol 8:12–17.
Raymond PA, Rivlin PK. 1987. Germinal cells in the goldfish retina thatproduce rod photoreceptors. Dev Biol 122:120–138.
Raymond PA, Barthel LK, Curran GA. 1995. Developmental patterning ofrod and cone photoreceptors in embryonic zebrafish. J Comp Neurol359:537–550.
Robinson S. 1987. Ontogeny of the area centralis in the cat. J Comp Neurol255:50–67.
Sandell JH, Martin SC, Heinrich G. 1994a. The development of GABAimmunoreactivity in the retina of the zebrafish, Brachydanio rerio. JComp Neurol 345:596–601.
Sandell JH, Martin SC, Heinrich G. 1994b. The development of neuro-trophin receptor Trk immunoreactivity in the retina of the zebrafish,Brachydanio rerio. Dev Brain Res 81:192–200.
EARLY RETINAL DEVELOPMENT IN THE ZEBRAFISH 535

Schmitt EA, Dowling JE. 1994. Eye morphogenesis in the zebrafish,Brachydanio rerio. J Comp Neurol 344:532–542.
Schmitt EA, Dowling JE. 1996. Comparison of topographical patterns ofganglion and photoreceptor cell differentiation in the retina of thezebrafish, Danio rerio. J Comp Neurol 371:222–234.
Schmitt EA, Kunz YW. 1989. Retinal morphogenesis in the rainbow trout,Salmo gairdneri. Brain Behav Evol 34:48–64.
Silver J, Sapiro J. 1981. Axonal guidance during development of the opticnerve: The role of pigmented epithelia and other extrinsic factors. JComp Neurol 202:521–538.
Spira AW, Millar TJ, Ishimoto I, Epstein ML, Johnson CD, Dahl JL, MorganIG. 1987. Localization of choline acetytransferase-like immunoreactiv-ity in the embryonic chick retina. J Comp Neurol 260:526–538.
Stenkamp DL, Gregory JK, Adler R. 1993. Retinoid effects in purifiedcultures of chick embryo retina neurons and photoreceptors. Invest.Ophthalmol Vis Sci 34:2425–2436.
Stenkamp DL, Hisatomi O, Barthel LK, Tokunaga F, Raymond PA. 1996.Temporal expression of rod and cone opsin in embryonic goldfish retinapredicts the spatial organization of the cone mosaic. Invest OphthalmolVis Sci 37:363–376.
Straznicky C, Chehade M. 1987. The formation of the area centralis of theretinal ganglion cell layer in the chick. Development 100:411–420.
Strongin AC, Guillery RW. 1981. The distribution of melanin in thedeveloping optic cup and stalk and its relation to cellular degeneration.J Neurosci 1:1193–1204.
Stuermer CA. 1988. Retinotopic organization of the developing retinotectalprojections in the zebrafish embryo. J Neurosci 8:4513–4530.
Vaney DI. 1984. ‘‘Coronate’’ amacrine cells in the rabbit retina have the‘‘starburst’’ dendritic morphology. Proc R Soc Lond (Biol) 202:409–416.
Walls G. 1942. The Vertebrate Eye and its Adaptive Radiation. New York:Hafner.
Wassle H, Chun MH, Muller F. 1987. Amacrine cells in the ganglion celllayer of the cat retina. J Comp Neurol 265:391–408.
Westerfield M. 1994. The Zebrafish Book: A Guide for the Laboratory Use ofZebrafish (Brachydanio rerio). Eugene: Institute of Neuroscience, Uni-versity of Oregon.
Zimmerman RP, Polley EH, Fortney RL. 1988. Cell birthdays and rate ofdifferentiation of ganglion and horizontal cells of the developing catretina. J Comp Neurol 274:77–90.
536 E.A. SCHMITT AND J.E. DOWLING





























