Embed Size (px)
Citation preview

Development 109, 567-576 (1990)Printed in Great Britain © T h e Company of Biologists Limited 1990
567
Early onset of phenotype and cell patterning in the embryonic zebrafish
retina
KAREN D. LARISON and RUTH BREMILLER
Institutes of Molecular Biology and Neuroscience, University of Oregon, Eugene, OR 97403, USA
Summary
The regular arrangement of retinal cone cells in a mosaicpattern is a common feature of teleosts. In the zebrafish,Brachydanio rerio, the retinal cone mosaic comprisesparallel rows consisting of a repeating motif of four conetypes. In order to elucidate the temporal and spatialaspects of the genesis of the cone mosaic in the develop-ing retina, we generated a monoclonal antibody thatspecifically binds to the double cone photoreceptor of theadult. We first saw staining in the developing retina withthis antibody, FRet 43, at 48 hours postfertilization, thetime at which the first photoreceptor cells undergo theirfinal mitotic division. We then injected embryonic fishwith the thymidine analog, 5-bromo-2'-deoxyuridine(BrdU), confirming with a double-labeling experiment
that the onset of FRet 43 antigenicity occurs within threehours of the cellular division that generates the doublecone photoreceptors. Then we stained tangential sectionsof the 54-hour embryonic retina with FRet 43, furthershowing that cells devoid of staining alternate withstained pairs of cells in a pattern that is consistent withthe arrangement of photoreceptors in the adult conemosaic. These results indicate that a marker of thedouble cone phenotype is expressed at approximately thesame time as cellular birthday and that the mosaicpatterning is present within 6 hours of this expression.
Key words: retinal development, double cones, conemosaic, photoreceptors, cell patterning.
Introduction
In the retinas of many teleosts, the cone photoreceptorsare arranged in a simple, repeating pattern known asthe cone mosaic (Lyall, 1957; Engstrom, 1963a). Thezebrafish retina {Brachydanio rerio) is no exception.Engstrom (1960) was the first to describe the conemosaic in this species as a row pattern consisting of aninvariant array of four cone types (the short single, thelong single, and the unequal double cones); this mosaicwas found to extend throughout the photoreceptorlayer.
Using radioactive thymidine to mark actively dividingcells, Nawrocki (1985) had previously shown that zebra-fish photoreceptors first become postmitotic at 48 h(48h=48 hours postfertilization). Kljavin (1987) thenused electron microscopy to examine the subsequentdifferentiation of these postmitotic photoreceptors,demonstrating that outer segments of both rod andcone photoreceptors first appear in a localized patch onthe ventral aspect of the retina at 60 h. In Branchek andBreMillers' light microscopy studies (1984), however,the cone mosaic was not apparent until 8 days postferti-lization and some of the photoreceptor cell types werenot identifiable until 12 days. From Branchek andBreMillers' results, one might hypothesize that thenascent cone cells exist as a homogenous population ofunspecified cell types until very late in the development
of the retina. Alternatively, one might propose thatspecific cone types are present in a cryptic form prior tothe onset of their late overt morphological differen-tiation. To determine which of these alternativeschemes is correct, we developed a monoclonal anti-body, FRet 43, that specifically binds to the double conephotoreceptor. We hoped that the temporal and spatialpattern of the onset of FRet 43 antigenicity wouldreveal whether a diverse population of cone cells existsduring early stages of development.
FRet 43 staining confirmed that the double cones areindeed present prior to morphological differentiation.The time of onset of antigenicity at 48 h was indis-tinguishable from the time at which Nawrocki (1985)had shown that the first photoreceptor cells undergotheir last mitotic division. We further examined thetemporal relationship between cellular birthday andonset of double cone specificity by staining retinas ofembryonic fish that had previously been injected withthe thymidine analog, BrdU, with both anti-BrdU andFRet 43 antibodies. These double-label experimentsshowed that a marker for the double cone cell pheno-type is expressed within three hours of the final celldivision.
Because of its early expression, the FRet 43 antigenalso provided us with a marker to investigate photo-receptor pattern formation. The arrangement of stainedand unstained cells demonstrated that a pattern consist-

568 K. D. Larison and R. BreMiller
ent with the adult mosaic is present in the 54-h embryo.These results suggest that photoreceptor cell typesdifferentiate and form the mosaic pattern very early indevelopment, only shortly after their cellular birthdays.
Materials and methods
Generation of hybridomasMale Balb/c mice were immunized at multiple intraperitonealand subcutaneous sites with a preparation of whole adultretinal cells that had been fixed in 2 % paraformaldehyde for30 minutes at room temperature and resuspended in PBS andFreund's adjuvant. Each immunization consisted of retinalcells obtained from approximately fifteen adult wild-typezebrafish. Each mouse received at least nine immunizationsover the course of approximately one year.
The cell fusion and cloning of hybridomas were completedby the methods described by Ciment and Weston (1982) andmodified by Trevarrow et al. (1990). An immunohistochemi-cal screen was performed on eye-containing transverse sec-tions of lightly fixed adult zebrafish heads.
Antibody typing and subtyping were performed utilizing animmunodiffusion kit for mouse monoclonal antibodies(Miles).
AnimalsIn the wild-type zebrafish, the cellular details of the photo-receptor layer are obscured by the pigment epithelial pro-cesses that extend into this layer of the retina. Therefore, allanimals used were homozygous mutant at one or more of theloci affecting pigment expression as previously described byStreisinger et al. (1986). In the double-labeling experiments,alb-1 zebrafish {Brachydanio rerio) were used; in all otherexperiments, we used animals that were homogygous mutantat three pigment pattern loci: gol-1, alb-1, and spa-1. Thesefish provide a convenient tool for histological studies of theretina as they lack all but the reflective pigmentation. Inparticular, antibody staining of photoreceptor cells is notobscured by the closely associated pigment epithelium.
Embryos were obtained from spontaneous spawnings fromthis strain of zebrafish. Morphological stagings were per-formed within about 2 hours of spawning by a careful countingof cell numbers. Time of fertilization can be ascertained bythis procedure to a precision of ±15 minutes. Embryos wereincubated in a temperature-controlled, 28.5°C water bathuntil time of killing. Developmental stage is expressed here ashours postfertilization at the standard temperature (h). Atleast 14 embryos were examined at 44 h, 48 h, and 54 h; and 10embryos were examined at 96 h.
In the double-label experiments, embryos were injected at51 h (±lh) with 10mM 5-bromo-2'-deoxyuridine (BrdU)(Amersham, Arlington Heights, Illinois) and 1 mM5-fluoro-2'-deoxyuridine; the latter reagent was added toinhibit endogenous production of thymidine. To enable directvisualization of the injection process, we added 1 % neutralred to the solution. Approximately 5nL of this solution wasinjected into the yolk sac, and the embyos were then killed 3 h(±20min) following injection.
Light conditionsTo ascertain whether light affected the onset of antigenicity,embryos were raised in one of two light conditions. One groupof embryos was raised under conditions of a variable dark/light cycle. These embryos were subject to ambient laboratorylight conditions, with some portion of each day spent in
darkness. The second group of embryos was raised under aconstant 2 footcandles of mixed fluorescent and incandescentlight as measured by a Luna-Pro light meter. The lightconditions had no effect upon the onset of antigenicity (datanot shown).
Sample preparation for EM microscopyEyes were dissected from the heads of adults. The corneaswere opened and the lens extracted. The resulting eye cupswere fixed for 60min at 4°C in a trialdehyde fixative (Kimmelet al, 1981); postfixed in 1% buffered osmium tetroxide,pH7.3, for 60min at room temperature; washed; dehydratedthrough graded ethanols; embedded in Epon-Araldite; andsectioned at a thickness of 7/«m. Sections were chosen for re-embedding with use of Nomarski optics on a Zeiss Universalmicroscope (Schabtach and Parkening, 1974). Sections of70 nm were placed on formvar-coated grids, stained withuranyl acetate, carbon stabilized and examined with a PhilipsCM 12 electron microscope.
ImmunohistologySample preparation for sections labeled with FRet 43
Tissue for the cryostat sections was prepared by immersingeither whole embryos or adult heads in a 4°C fixative solutionfor 12 to 20 hours. This fixative solution was composed of 1 %paraformaldehyde in 0.1M phosphate buffer at a pH of7.2-7.4. After fixation, the tissue was washed and immersedin 30 % sucrose at 4°C until permeated. The adult heads werethen frozen directly onto the mounting platform for section-ing, whereas embryos were aligned and embedded in TissueTek O.C.T. compound prior to freezing. Sections of 16/«mwere cut and transferred to gelatin-coated slides and allowedto dry.
Tissue for the vibratome sections was subjected to the samefixation protocol as described above, washed, and immersedin 0.3 M sucrose at 4°C. Embryos were sectioned by themethod described by Hanneman et al. (1988). Vibratomesections of adult eyes were obtained in the following manner.Eyes were removed and the cornea was opened to extract thelens. The resulting eye cup was then gently aspirated toremove the vitreous fluid and then filled with a warmedsolution containing 17% gelatin in 10% Hank's saline. Theeye was then mounted in the gelatin solution; 30- to 60-jansections were cut and mounted onto a gelatin-coated slide.
Immunological labeling was performed as described byHanneman et al. (1988).
Sample preparation for sections labeled with bothFRet 43 and anti-BrdU antibody
Heads were removed from the fish and were fixed for 3 hoursat 4°C in fixative (4% paraformaldehyde, 4% sucrose, and0.15 mM CaCl2 in 0.1M phosphate buffer, pH7.3). The tissuewas washed in several changes of phosphate buffer andembedded in a solution of 5% sucrose and 1.5% agar,warmed to 40°C. The embedded heads were sunk in 30%sucrose overnight at 4°C.
Sections of 20 jan were incubated overnight with FRet 43antibody at 4°C. After washing in PBS, a biotinylatedsecondary antibody was added for 30 min. Sections were againwashed in PBS and then incubated in an avidin-biotinylatedcomplex for 60min. After several washes, the sections wereincubated in a solution containing 0.05 % diaminobenzidine(DAB) and 1% DMSO in 0.05 M phosphate buffer, pH7.4,for 10min. Hydrogen peroxide was added to the DAB sol-ution to give a final concentration of 0.003 %. The sectionswere monitored visually. After the brown reaction product
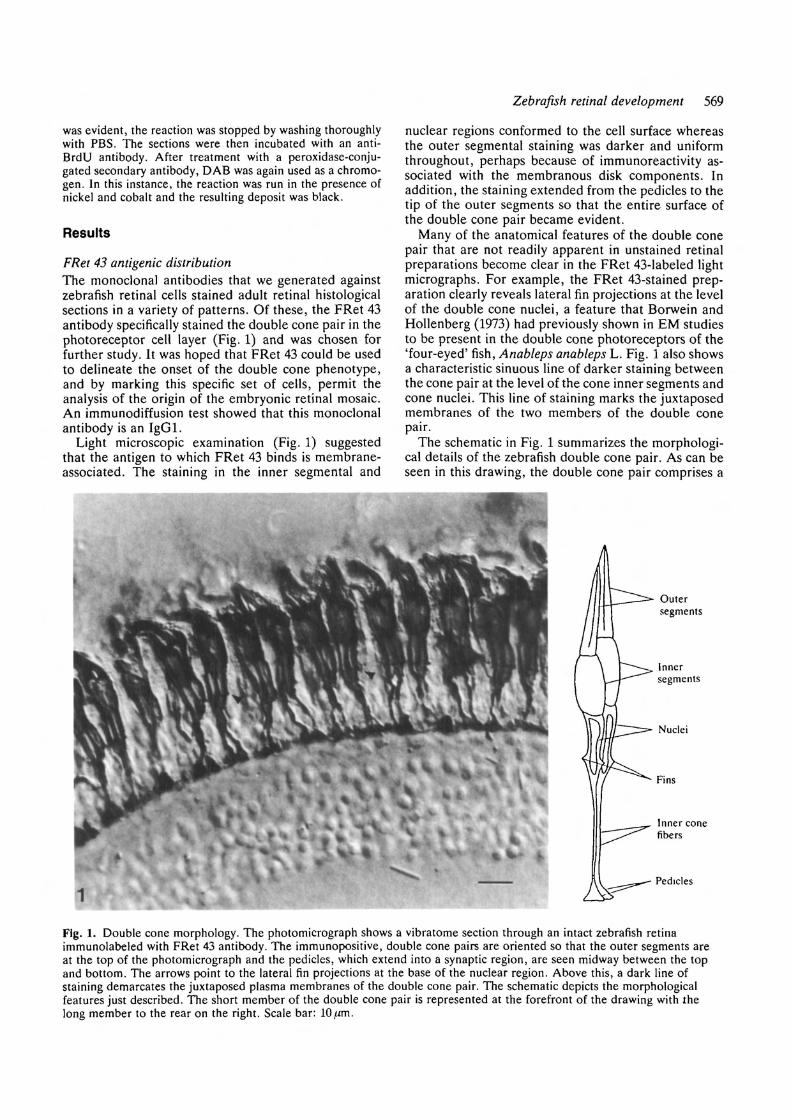
Zebrafish retinal development 569
was evident, the reaction was stopped by washing thoroughlywith PBS. The sections were then incubated with an anti-BrdU antibody. After treatment with a peroxidase-conju-gated secondary antibody, DAB was again used as a chromo-gen. In this instance, the reaction was run in the presence ofnickel and cobalt and the resulting deposit was black.
Results
FRet 43 antigenic distributionThe monoclonal antibodies that we generated againstzebrafish retinal cells stained adult retinal histologicalsections in a variety of patterns. Of these, the FRet 43antibody specifically stained the double cone pair in thephotoreceptor cell layer (Fig. 1) and was chosen forfurther study. It was hoped that FRet 43 could be usedto delineate the onset of the double cone phenotype,and by marking this specific set of cells, permit theanalysis of the origin of the embryonic retinal mosaic.An immunodiffusion test showed that this monoclonalantibody is an IgGl.
Light microscopic examination (Fig. 1) suggestedthat the antigen to which FRet 43 binds is membrane-associated. The staining in the inner segmental and
nuclear regions conformed to the cell surface whereasthe outer segmental staining was darker and uniformthroughout, perhaps because of immunoreactivity as-sociated with the membranous disk components. Inaddition, the staining extended from the pedicles to thetip of the outer segments so that the entire surface ofthe double cone pair became evident.
Many of the anatomical features of the double conepair that are not readily apparent in unstained retinalpreparations become clear in the FRet 43-labeled lightmicrographs. For example, the FRet 43-stained prep-aration clearly reveals lateral fin projections at the levelof the double cone nuclei, a feature that Borwein andHollenberg (1973) had previously shown in EM studiesto be present in the double cone photoreceptors of the'four-eyed' fish, Anableps anableps L. Fig. 1 also showsa characteristic sinuous line of darker staining betweenthe cone pair at the level of the cone inner segments andcone nuclei. This line of staining marks the juxtaposedmembranes of the two members of the double conepair.
The schematic in Fig. 1 summarizes the morphologi-cal details of the zebrafish double cone pair. As can beseen in this drawing, the double cone pair comprises a
Outersegments
Innersegments
Nuclei
Fins
Inner conefibers
Pedicles
Fig. 1. Double cone morphology. The photomicrograph shows a vibratome section through an intact zebrafish retinaimmunolabeled with FRet 43 antibody. The immunopositive, double cone pairs are oriented so that the outer segments areat the top of the photomicrograph and the pedicles, which extend into a synaptic region, are seen midway between the topand bottom. The arrows point to the lateral fin projections at the base of the nuclear region. Above this, a dark line ofstaining demarcates the juxtaposed plasma membranes of the double cone pair. The schematic depicts the morphologicalfeatures just described. The short member of the double cone pair is represented at the forefront of the drawing with thelong member to the rear on the right. Scale bar: lO t̂m.
570 K. D. Larison and R. BreMiller
long member and a short member. Close inspection ofthe photomicrograph in Fig. 1 confirms that the indi-vidual members of the double cone pair are bothasymmetrically positioned and unequal in size.
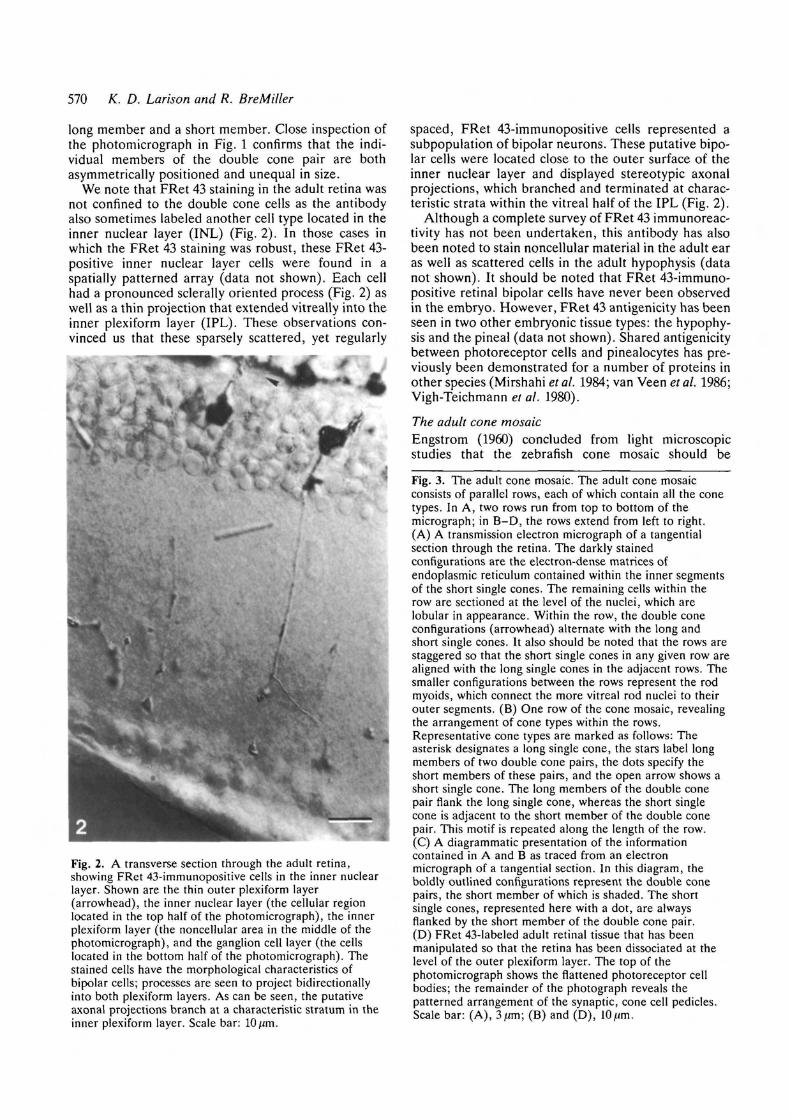
We note that FRet 43 staining in the adult retina wasnot confined to the double cone cells as the antibodyalso sometimes labeled another cell type located in theinner nuclear layer (INL) (Fig. 2). In those cases inwhich the FRet 43 staining was robust, these FRet 43-positive inner nuclear layer cells were found in aspatially patterned array (data not shown). Each cellhad a pronounced sclerally oriented process (Fig. 2) aswell as a thin projection that extended vitreally into theinner plexiform layer (IPL). These observations con-vinced us that these sparsely scattered, yet regularly
Fig. 2. A transverse section through the adult retina,showing FRet 43-immunopositive cells in the inner nuclearlayer. Shown are the thin outer plexiform layer(arrowhead), the inner nuclear layer (the cellular regionlocated in the top half of the photomicrograph), the innerplexiform layer (the noncellular area in the middle of thephotomicrograph), and the ganglion cell layer (the cellslocated in the bottom half of the photomicrograph). Thestained cells have the morphological characteristics ofbipolar cells; processes are seen to project bidirectionallyinto both plexiform layers. As can be seen, the putativeaxonal projections branch at a characteristic stratum in theinner plexiform layer. Scale bar: 10 /im.
spaced, FRet 43-immunopositive cells represented asubpopulation of bipolar neurons. These putative bipo-lar cells were located close to the outer surface of theinner nuclear layer and displayed stereotypic axonalprojections, which branched and terminated at charac-teristic strata within the vitreal half of the IPL (Fig. 2).
Although a complete survey of FRet 43 immunoreac-tivity has not been undertaken, this antibody has alsobeen noted to stain noncellular material in the adult earas well as scattered cells in the adult hypophysis (datanot shown). It should be noted that FRet 43-immuno-positive retinal bipolar cells have never been observedin the embryo. However, FRet 43 antigenicity has beenseen in two other embryonic tissue types: the hypophy-sis and the pineal (data not shown). Shared antigenicitybetween photoreceptor cells and pinealocytes has pre-viously been demonstrated for a number of proteins inother species (Mirshahi et al. 1984; van Veen et al. 1986;Vigh-Teichmann et al. 1980).
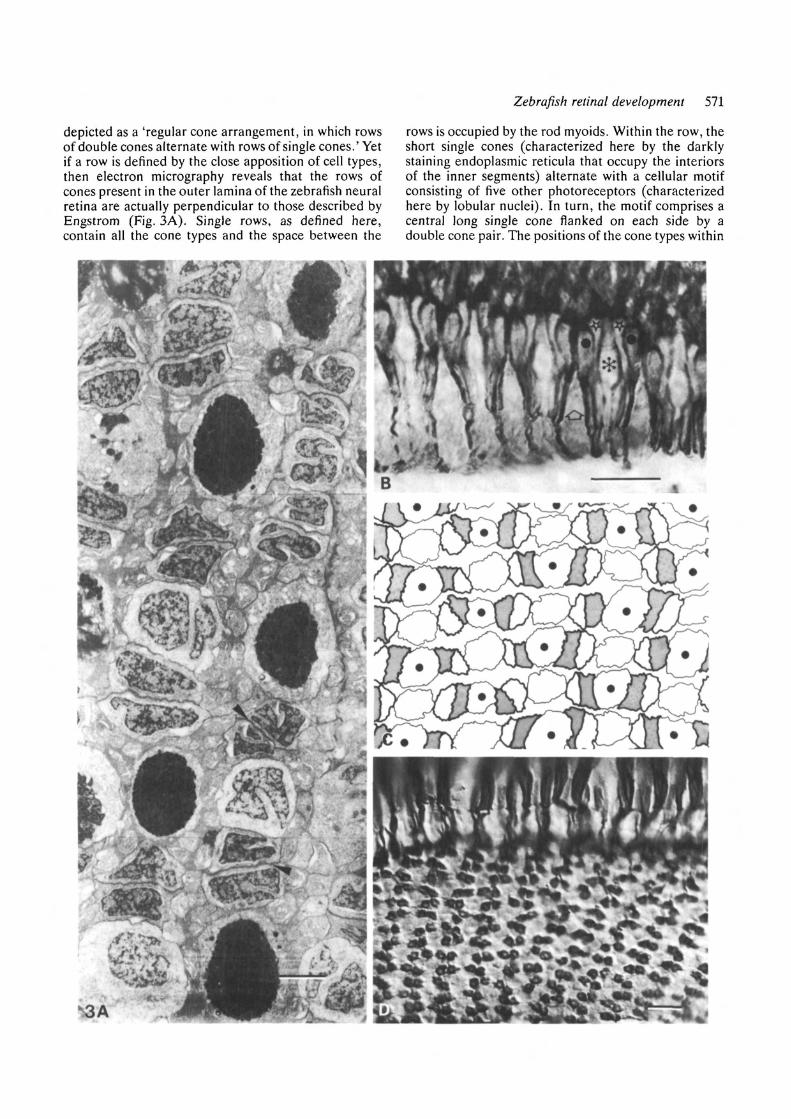
The adult cone mosaicEngstrom (1960) concluded from light microscopicstudies that the zebrafish cone mosaic should be
Fig. 3. The adult cone mosaic. The adult cone mosaicconsists of parallel rows, each of which contain all the conetypes. In A, two rows run from top to bottom of themicrograph; in B-D, the rows extend from left to right.(A) A transmission electron micrograph of a tangentialsection through the retina. The darkly stainedconfigurations are the electron-dense matrices ofendoplasmic reticulum contained within the inner segmentsof the short single cones. The remaining cells within therow are sectioned at the level of the nuclei, which arelobular in appearance. Within the row, the double coneconfigurations (arrowhead) alternate with the long andshort single cones. It also should be noted that the rows arestaggered so that the short single cones in any given row arealigned with the long single cones in the adjacent rows. Thesmaller configurations between the rows represent the rodmyoids, which connect the more vitreal rod nuclei to theirouter segments. (B) One row of the cone mosaic, revealingthe arrangement of cone types within the rows.Representative cone types are marked as follows: Theasterisk designates a long single cone, the stars label longmembers of two double cone pairs, the dots specify theshort members of these pairs, and the open arrow shows ashort single cone. The long members of the double conepair flank the long single cone, whereas the short singlecone is adjacent to the short member of the double conepair. This motif is repeated along the length of the row.
(C) A diagrammatic presentation of the informationcontained in A and B as traced from an electronmicrograph of a tangential section. In this diagram, theboldly outlined configurations represent the double conepairs, the short member of which is shaded. The shortsingle cones, represented here with a dot, are alwaysflanked by the short member of the double cone pair.(D) FRet 43-labeled adult retinal tissue that has beenmanipulated so that the retina has been dissociated at thelevel of the outer plexiform layer. The top of thephotomicrograph shows the flattened photoreceptor cellbodies; the remainder of the photograph reveals thepatterned arrangement of the synaptic, cone cell pedicles.Scale bar: (A), 3^m; (B) and (D), 10 jrni.
Zebrafish retinal development 571
depicted as a 'regular cone arrangement, in which rowsof double cones alternate with rows of single cones.' Yetif a row is defined by the close apposition of cell types,then electron micrography reveals that the rows ofcones present in the outer lamina of the zebrafish neuralretina are actually perpendicular to those described byEngstrom (Fig. 3A). Single rows, as defined here,contain all the cone types and the space between the
rows is occupied by the rod myoids. Within the row, theshort single cones (characterized here by the darklystaining endoplasmic reticula that occupy the interiorsof the inner segments) alternate with a cellular motifconsisting of five other photoreceptors (characterizedhere by lobular nuclei). In turn, the motif comprises acentral long single cone flanked on each side by adouble cone pair. The positions of the cone types within
572 K. D. Larison and R. BreMiller
a row have been further defined in Fig. 3B. Here, it canbe seen that the long single cones are always bound bythe long members of the double cone pair and the shortsingle cones by the short members. The pattern re-vealed in Figs 3A and 3B is schematically representedin the drawing contained in Fig. 3C.
Fig. 3D is a photomicrograph of a whole mount of thephotoreceptor layer that displays the double conepedicles stained with FRet 43 antibody. At the top ofthe photo, one observes the bent, flattened photorecep-tor cells. At the bottom, the double cone pedicles arealigned in rows of doubled configurations. This exten-
sion of the cone mosaic into the synaptic layer haspreviously been described in a number of other teleosts(Engstrom, 19636; Wagner, 1978).
Photoreceptors: cellular birthdays and onset of FRet 43antigenicityUsing radioactive thymidine to label actively dividingcells, Nawrocki (1985) had previously shown that thefirst postmitotic zebrafish photoreceptor cells arise atapproximately 48 h. Using FRet-43 staining as a markerfor double cone differentiation, we performed a num-ber of experiments to determine the temporal relation-
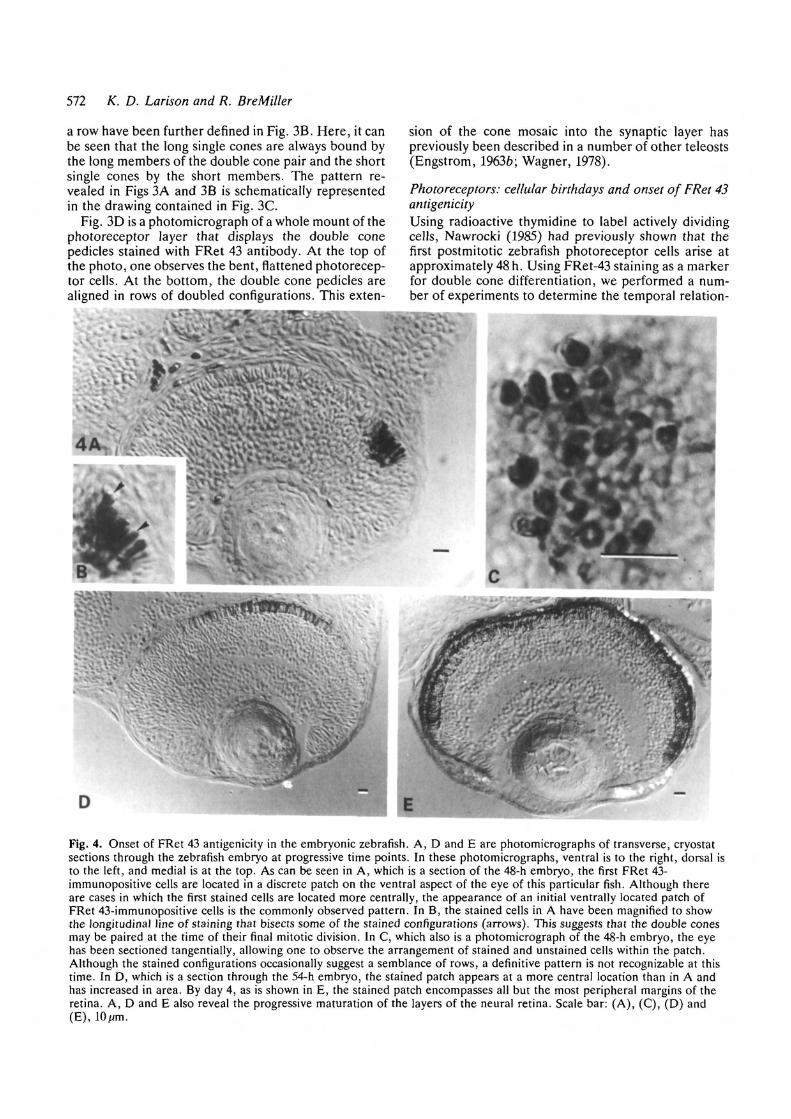
Fig. 4. Onset of FRet 43 antigenicity in the embryonic zebrafish. A, D and E are photomicrographs of transverse, cryostatsections through the zebrafish embryo at progressive time points. In these photomicrographs, ventral is to the right, dorsal isto the left, and medial is at the top. As can be seen in A, which is a section of the 48-h embryo, the first FRet 43-immunopositive cells are located in a discrete patch on the ventral aspect of the eye of this particular fish. Although thereare cases in which the first stained cells are located more centrally, the appearance of an initial ventrally located patch ofFRet 43-immunopositive cells is the commonly observed pattern. In B, the stained cells in A have been magnified to showthe longitudinal line of staining that bisects some of the stained configurations (arrows). This suggests that the double conesmay be paired at the time of their final mitotic division. In C, which also is a photomicrograph of the 48-h embryo, the eyehas been sectioned tangentially, allowing one to observe the arrangement of stained and unstained cells within the patch.Although the stained configurations occasionally suggest a semblance of rows, a definitive pattern is not recognizable at thistime. In D, which is a section through the 54-h embryo, the stained patch appears at a more central location than in A andhas increased in area. By day 4, as is shown in E, the stained patch encompasses all but the most peripheral margins of theretina. A, D and E also reveal the progressive maturation of the layers of the neural retina. Scale bar: (A), (C), (D) and(E), lO/m.
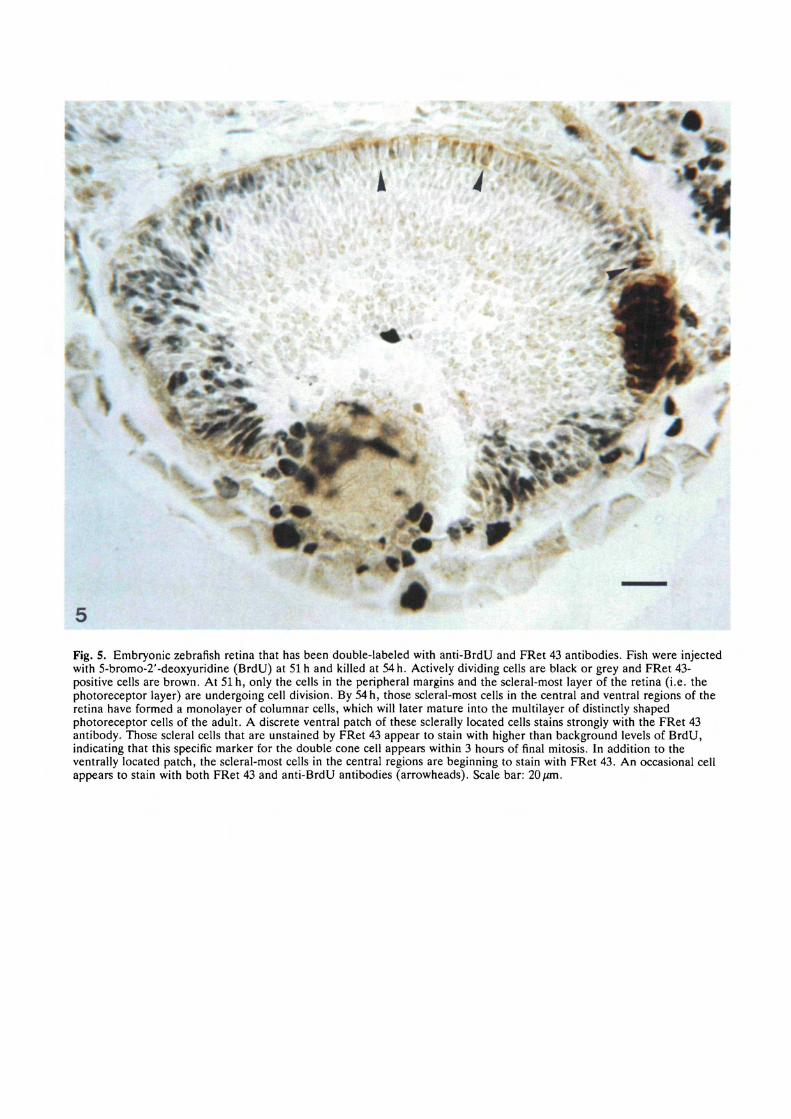
Fig. 5. Embryonic zebrafish retina that has been double-labeled with anti-BrdU and FRet 43 antibodies. Fish were injectedwith 5-bromo-2'-deoxyuridine (BrdU) at 51 h and killed at 54 h. Actively dividing cells are black or grey and FRet 43-positive cells are brown. At 51 h, only the cells in the peripheral margins and the scleral-most layer of the retina (i.e. thephotoreceptor layer) are undergoing cell division. By 54 h, those scleral-most cells in the central and ventral regions of theretina have formed a monolayer of columnar cells, which will later mature into the multilayer of distinctly shapedphotoreceptor cells of the adult. A discrete ventral patch of these sclerally located cells stains strongly with the FRet 43antibody. Those scleral cells that are unstained by FRet 43 appear to stain with higher than background levels of BrdU,indicating that this specific marker for the double cone cell appears within 3 hours of final mitosis. In addition to theventrally located patch, the scleral-most cells in the central regions are beginning to stain with FRet 43. An occasional cellappears to stain with both FRet 43 and anti-BrdU antibodies (arrowheads). Scale bar: 20/an.


Zebrafish retinal development 573
ship between cellular birthday and the specification ofdouble cone identity.
First, we screened fourteen 44-h embryos for FRet 43antigenicity. Of these, only one fish stained with theFRet 43 antibody in the photoreceptor cell layer (datanot shown). We attributed the presence of FRet 43staining in this particular fish to variation in the rate ofdevelopment, concluding that FRet 43 staining was notactually present four hours prior to cellular birthday. By48 h, all 38 embryos that were screened contained FRet43-immunopositive photoreceptor cells. In the embry-onic eye shown in Fig. 4A, FRet 43-immunoreactivecells appear in a discrete, ventrally located patch on thescleral surface of the neural retina. This location iswhere Kjlavin (1987) observed the first outgrowth ofphotoreceptor outer segments at 60 h, i.e. 12 hours afterwe demonstrated labeling. In our study, some periph-eral-to-central variability in the specific location of thispatch of initially stained cells has been seen (seeFig. 4D). In the higher magnification of the stainedarea, shown in Fig. 4B, the immunopositive configur-ations were characterized by a longitudinal line ofdarker staining (see arrows), which suggested that eachconfiguration represented a double cone pair. InFig. 4C, which is a tangential section through the patch,the FRet 43-immunoreactive cells are interspersed in asomewhat disordered fashion among those cells thatlack staining; a semblance of a row pattern may or maynot be present at 48 h. By 54 h, the area containing FRet43-positive cells has increased (see Fig. 4D). By 96h, atime at which the zebrafish first actively feeds usingvisual cues (Clark, 1981), only the peripheral margins ofthe photoreceptor cell layer remain unstained(Fig. 4E).
These data suggested to us that the onset of FRet 43antigenicity closely followed the spatial and temporalpattern of photoreceptor birthdays as seen in Naw-rocki's birthday studies (unpublished data). Thissuggested to us that the double cone phenotype mayappear as early as the final mitotic division. To test thishypothesis, we injected the yolk sacs of 51-h embryoswith a thymidine analog, BrdU, and then killed theanimals 3 hours later. The animals were then double-labeled with both anti-BrdU and FRet 43 antibodies.
In Fig. 5, BrdU-positive cells are black or grey,whereas Fret-43 positive cells are brown. At 51 h, mostretinal cells in the inner layers of the retina arepostmitotic; cell division is confined to the photorecep-tor layer and the peripheral regions of the retina. Adiscrete patch of FRet-43 positive cells can be seen onthe ventral aspect of the embryonic eye. Those photo-receptors found in regions not stained by FRet 43 showsome level of BrdU staining. In those regions in thecentral retina in which FRet-43 staining is just beginningto be apparent, cells appear to stain with anti-BrdU athigher than background levels. These data show thatthis marker for the double cone cell phenotype isexpressed within 3 hours of the final cell division.
Embryonic photoreceptor cell patterningExamination of the 54-h embryo strongly suggested to
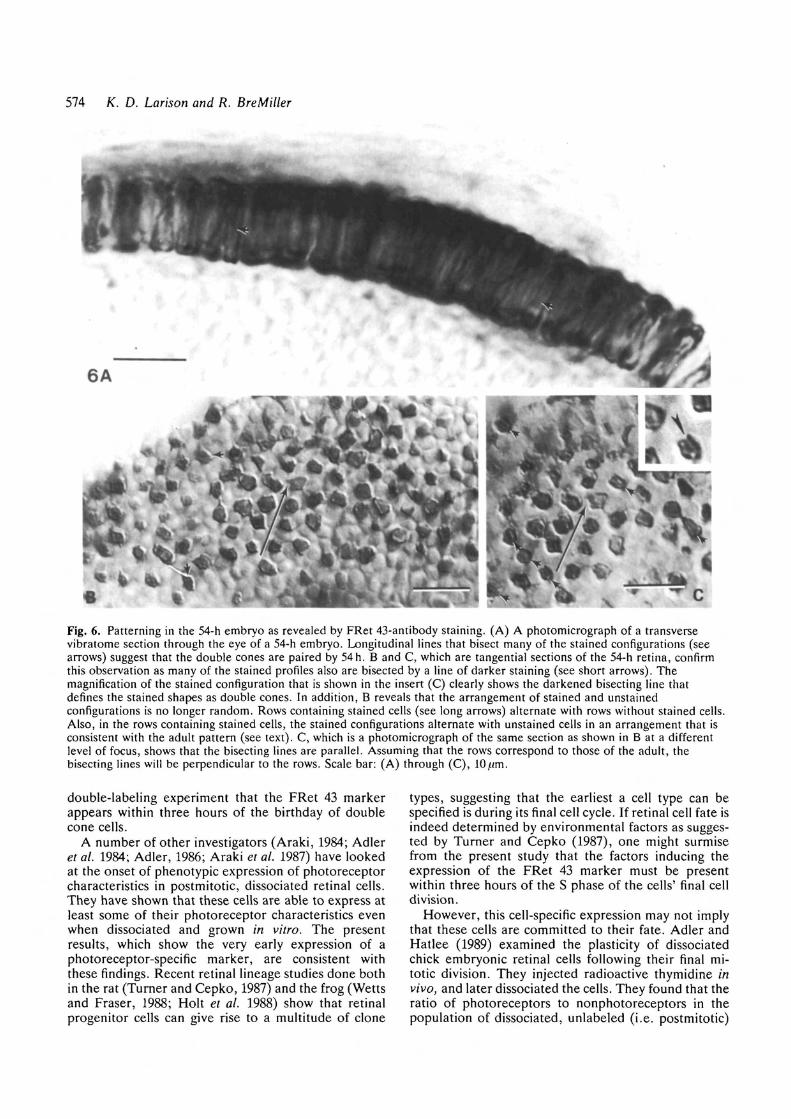
us that cell patterning had occurred in the photorecep-tor layer by this time. As previously seen in the 48-hembryo, close inspection of transverse sections of the54-h retina revealed that most (if not all) stainedconfigurations were characterized by a longitudinal lineof darker staining in the midregion (Fig. 6A). Intangential sections of the 54-h embryo (Figs 6B and6C), a darker line of staining bisected many of thestained configurations (see the short arrows and inset inFig. 6C), confirming the doubled nature of these em-bryonic cell types.
By far the most striking feature observed in thestained 54-h retina, though, was the appearance oforder in the arrangement of the stained and unstainedcells (Figs 6B and 6C). Not only were the stainedconfigurations isolated on all sides by unstained cells,but it also appeared that the FRet 43-immunopositivecells were arranged in rows. In addition, the lines thatbisected the individual stained configurations (Fig. 6C),and which presumably marked the position of theapposing membranes of the double cone pair, were allapproximately parallel. In the adult, the juxtaposedmembranes of the double cone pair are perpendicularto the row. Assuming that this is also true in theembryo, the long arrows in Figs 6B and 6C would definea row of cone cells.
As can be seen in Fig. 6C, rows of cells containingstained configurations alternate with rows with nostaining; and the stained configurations within a rowalternate with unstained cells. This pattern of staining isconsistent with the adult photoreceptor mosaic. Thecells in the unstained rows may represent developingrods, previously shown to be present by Nawrocki et al.(1985) at 6-8 days and by Kljavin (1987) at 60 hours.The cells contained in the rows with stained configur-ations are arranged in a manner that is consistent withthe adult mosaic in which the double cone pair alter-nates with one of the two single cone types (see Fig. 4).This evidence strongly suggests that the adult mosaic iscryptically present at a time when the photoreceptorsare morphologically indistinguishable from oneanother.
Discussion
We have developed a monoclonal antibody, FRet 43,that is specific for the double cone phenotype in thephotoreceptor layer of the zebrafish retina. We haveused this antibody to explore the relationship betweencellular birthday, onset of cellular phenotype, andmosaic pattern formation. Using morphological cri-teria, Branchek and BreMiller (1984) were unable todefinitively identify the double cone phenotype untilday 12 postfertilization and could not distinguish amosaic pattern until day 8. Using our marker, we couldidentify the double cone phenotype at48h,andby54h,these FRet 43-positive cells were arranged in a patternconsistent with that of the adult mosaic. We theninjected 51-h zebrafish with the thymidine analog,BrdU, and by killing at 54-h, were able to show with a
574 K. D. Larison and R. BreMiller
Fig. 6. Patterning in the 54-h embryo as revealed by FRet 43-antibody staining. (A) A photomicrograph of a transversevibratome section through the eye of a 54-h embryo. Longitudinal lines that bisect many of the stained configurations (seearrows) suggest that the double cones are paired by 54h. B and C, which are tangential sections of the 54-h retina, confirmthis observation as many of the stained profiles also are bisected by a line of darker staining (see short arrows). Themagnification of the stained configuration that is shown in the insert (C) clearly shows the darkened bisecting line thatdefines the stained shapes as double cones. In addition, B reveals that the arrangement of stained and unstainedconfigurations is no longer random. Rows containing stained cells (see long arrows) alternate with rows without stained cells.Also, in the rows containing stained cells, the stained configurations alternate with unstained cells in an arrangement that isconsistent with the adult pattern (see text). C, which is a photomicrograph of the same section as shown in B at a differentlevel of focus, shows that the bisecting lines are parallel. Assuming that the rows correspond to those of the adult, thebisecting lines will be perpendicular to the rows. Scale bar: (A) through (C),
double-labeling experiment that the FRet 43 markerappears within three hours of the birthday of doublecone cells.
A number of other investigators (Araki, 1984; Adleret al. 1984; Adler, 1986; Araki et al. 1987) have lookedat the onset of phenotypic expression of photoreceptorcharacteristics in postmitotic, dissociated retinal cells.They have shown that these cells are able to express atleast some of their photoreceptor characteristics evenwhen dissociated and grown in vitro. The presentresults, which show the very early expression of aphotoreceptor-specific marker, are consistent withthese findings. Recent retinal lineage studies done bothin the rat (Turner and Cepko, 1987) and the frog (Wettsand Fraser, 1988; Holt et al. 1988) show that retinalprogenitor cells can give rise to a multitude of clone
types, suggesting that the earliest a cell type can bespecified is during its final cell cycle. If retinal cell fate isindeed determined by environmental factors as sugges-ted by Turner and Cepko (1987), one might surmisefrom the present study that the factors inducing theexpression of the FRet 43 marker must be presentwithin three hours of the S phase of the cells' final celldivision.
However, this cell-specific expression may not implythat these cells are committed to their fate. Adler andHatlee (1989) examined the plasticity of dissociatedchick embryonic retinal cells following their final mi-totic division. They injected radioactive thymidine invivo, and later dissociated the cells. They found that theratio of photoreceptors to nonphotoreceptors in thepopulation of dissociated, unlabeled (i.e. postmitotic)

Zebrafish retinal development 575
cells depended on the time of dissociation. Their resultssuggest that nonphotoreceptor retinal cells are commit-ted only after an extended exposure to the in vivoenvironment. An immunocytochemical study byMcLoon and Barnes (1989), however, shows that amarker for the chick ganglion cell phenotype, like thedouble cone-specific FRet 43 marker, is expressed assoon as these cells become postmitotic.
Our results, along with those of McLoon and Barnes(1989), might at first seem to contradict those of Adlerand Hatlee (1989). However, at least two explanationscan be proffered that would reconcile these apparentlycontradictory findings. First, retinal cells that areexpressing these early cell-specific markers may remainlabile, so that they can switch cell fates when dissociatedand introduced into a new environment. In otherwords, perhaps embryonic cells can express cell-specificmarkers and yet remain uncommitted.
Second, there is nothing to suggest that all cellswithin the retina utilize the same mechanisms to arriveat cellular identity. Photoreceptors and ganglion cellsmay be determined early, whereas the cells in the innernuclear layer may require a longer exposure to themicroenvironment, perhaps even using the ganglioncells and photoreceptors as a template from which toderive environmental information.
Our double cone marker, FRet 43, has permitted usto study pattern formation in the photoreceptor layer ofthe embryonic retina. By 54h, rows containing bothstained and unstained cells alternate with rows contain-ing only unstained cells. It is likely, from comparisonwith the adult pattern, that the cells present in theunstained rows are rod photoreceptors. Rods have beenshown to be present in the one-cell thick photoreceptorlamina of the 60-h retina (Kljavin, 1987). Our study alsoclearly suggests that each of the stained configurationswithin the rows containing immunopositive cells com-prise the two members of the double cone pair. Thestained configurations alternate systematically withunstained cells, suggesting that the unstained cellswithin these rows are the two single cone types. Theseresults are compatible with EM studies of the guppyretina (Kunz el al. 1983) in which double cones can bedistinguished prior to the initiation of outgrowth of theouter segments. Using EM techniques, however, theseresearchers were unable to identify a cone mosaic untilafter the outgrowth of the outer segments. But weobserve a pattern in the 54-h embryo that is consistentwith the adult mosaic. This is six hours before theobserved outgrowth of outer segments (Kljavin, 1987),and many days before the mosaic was first detectedpreviously in the zebrafish (Branchek and BreMiller,1984).
This rapid sequence of events in the zebrafish retina,in which cell-specific markers and pattern arises withina few short hours of cellular birthday, is not dissimilarto that described in the Drosophila retina (Ready et al.1976; Tomlinson, 1985; and Tomlinson and Ready,1987). In Drosophila, cell-cell interactions appear toplay an important role in the developing retina (for areview, see Ready, 1989); proteins expressed in some
photoreceptor types appear to be required for thedifferentiation of other, neighboring photoreceptors(Reinke and Zipursky, 1988; Tomlinson et al. 1988). Ifother photoreceptor-specific markers in the zebrafishare developed, it can then be investigated whethersimilar mechanisms, in particular cell interactions thatgenerate patterning, exist in this vertebrate retina.
This work is dedicated to the memory of George Streis-inger. The authors wish to thank Davy Grunwald for hisenthusiasm and support; Bill Trevarrow, Deanna Frost,Tracie Bork, Tom Schilling, Harry Howard, and ReidaKimmel for their technical assistance; Chuck Kimmel for hismany helpful discussions; and John Dowling, Monte Wester-field, and Judith Eisen for reviewing the manuscript. Thiswork was supported by NIH Grants GM22731, RR07080,NS17963, and HD22486.
References
ADLER, R. (1986). Developmental predetermination of thestructural and molecular polarization of photoreceptor cells. DeviBiol. 117, 520-527.
ADLER, R. AND HATLEE, M. (1989). Plasticity and differentiation ofembryonic retinal cells after terminal mitosis. Science 243,391-393.
ADLER, R., LINDSEY, J. D. AND ELSNER, C. L. (1984). Expression
of cone-like properties by chick embryo neural retina cells inglial-free monolayer cultures. J. Cell Biol. 99, 1173-1178.
ARAKI, M. (1984). Immunocytochemical study on photoreceptorcell differentiation in the cultured retina of the chick. Devi Biol.103, 313-318.
ARAKI, M., IIDA, Y., TAKETANI, S., WATANABE, K., OHTA, T. AND
SAITO, T. (1987). Characterization of photoreceptor celldifferentiation in the rat retinal cell culture. Devi Biol. 124,239-247.
BORWEIN, B. AND HOLLENBERG, M. J. (1973). The photoreceptorsof the "Four-eyed" fish, Anableps anableps L. J. Morph. 140,405-442.
BRANCHEK, T. AND BREMILLER, R. (1984). The development ofphotoreceptors in the zebrafish, Brachydamo rerio. I. Structure.J. comp. Neurol. 224, 107-115.
CIMENT, G. AND WESTON, J. A. (1982). Early appearance in neuralcrest and crest-derived cells of an antigenic determinant presentin avian neurons. Devi Biol. 93, 355-367.
CLARK, D. T. (1981). Visual responses in developing zebrafish(Brachydanio rerio). Ph.D. Dissertation, Univ. of Oregon,Eugene, Oregon.
ENGSTROM, K. (1960). Cone types and cone arrangements in theretina of some Cyprinids. Acra Zool. 41, 277-295.
ENGSTROM, K. (1963a). Cone types and cone arrangements inteleost retinae. Ada Zool. 44, 179-243.
ENGSTROM, K. (19636). Structure, organization and ultrastructureof the visual cells in the teleost family Labridae. Acta Zool 44,1-41.
HANNEMAN, E., TREVARROW, B., METCALFE, W. K., KIMMEL, C. B.
AND WESTERFIELD, M. (1988). Segmental pattern of developmentof the hindbrain and spinal cord of the zebrafish embryo.Development 103, 49-58.
HOLT, C. E., BERTSCH, T. W., ELLIS, H. M. AND HARRIS, W. A.
(1988). Cellular determination in the Xenopus retina isindependent of lineage and birth date. Neuron 1, 15-26.
KIMMEL, C. B., SESSIONS, S. K. AND KIMMEL, R. J. (1981).Morphogenesis and synaptogenesis of the zebrafish Mauthnerneuron. J. comp. Neurol. 198, 101-120.
KLJAVIN, I. J. (1987). Early development of photoreceptors in theventral retina of the zebrafish embryo. J. comp. Neurol. 260.461-471.
KUNZ, Y. W., ENNIS, S. AND WISE, C. (1983). Ontogeny of the
photoreceptors in the embryonic retina of the viviparous guppy,Poecilia reticulata P. (Teleostei). Cell Tissue Res. 230, 469-486.

576 K. D. Larison and R. BreMiller
LYALL, A. H. (1957). Cone arrangements in teleost retinae. Q. J.Mic. Sci. 98, 189-201.
MCLOON, S. C. AND BARNES, R. B. (1989). Early differentiation ofretinal ganglion cells: an axonal protein expressed bypremigratory and migrating retinal ganglion cells. J. Neurosci. 9,1424-1432.
MIRSHAHI, M., FAURE, J.-P., BRISSON, P., FALCON, J., GUERLOTTE,
J. AND COLUN, J.-P. (1984). S-antigen immunoreactivity inretinal rods and cones and pineal photosensitive cells. Biol. Cell.52, 195-198.
NAWROCKI, L. W. (1985). Development of the neural retina in thezebrafish, (Brachydanio rerio). Ph.D. Dissertation, Univ. ofOregon, Eugene, Oregon.
NAWROCKI, L., BREMILLER, R., STREISINGER, G. AND KAPLAN, M.
(1985). Larval and adult visual pigments of the zebrafish,Brachydanio rerio. Vision Res. 25, 1569-1576.
READY, D. F. (1989). A multifaceted approach to neuraldevelopment. Trends Neurosci. 12, 102-110.
READY, D. F., HANSON, T. E. AND BENZER, S. (1976).
Development of the Drosophila retina, a neurocrystalline lattice.Devi Biol. 53, 217-240.
REINKE, R. AND ZIPURSKY, S. L. (1988). Cell-cell interaction in theDrosophila retina: the bride of sevenless gene is required inphotoreceptor cell R8 for R7 cell development. Cell 55, 321-330.
SCHABTACH, E. AND PARKENING, T. A. (1974). Method forsequential high-resolution light and electron-microscopy ofselected areas of same material. J. Cell Biol. 61, 261-264.
STREISINGER, G., SINGER, F., WALKER, C , KNAUBER, D.
ANDDOWER, N. (1986). Segregation analysis and gene-centromeredistances in zebrafish. Genetics 111, 311-319.
TOMLINSON, A. (1985). The cellular dynamics of pattern formationin the eye of Drosophila. J. Embryo!, exp. Morph. 89, 313-331.
TOMUNSON, A., KIMMEL, B. E. AND RUBIN, G. M. (1988). rough, a
Drosophila homeobox gene required in photoreceptors R2 andR5 for inductive interactions in the developing eye. Cell 55,771-784.
TOMLINSON, A. AND READY, D. F. (1987). Neuronal differentiationin the Drosophila ommatidium. Devi Biol. 120, 366-376.
TREVARROW, B., MARKS, D. L. AND KIMMEL, C. B. (1990).
Organization of hindbrain segments in the zebrafish embryo.Neuron (in press).
TURNER, D. L. AND CEPKO, C. L. (1987). A common progenitor forneurons and glia persists in rat retina late in development.Nature 328, 131-136.
VAN VEEN, T H . , OSTHOLM, T., GIERSCHIK, P., SPIEGEL, A., SOMERS,
R., KORF, H.-W. AND KLEIN, D. C. (1986). Alpha-transducinimmunoreactivity in retinae and sensory pineal organs of adultvertebrates. Proc. natn. Acad. Sci. U.S.A. 83, 912-916.
VIGH-TEICHMANN, I., ROHLICH, P., VIGH, B. AND AROS, B. (1980).
Comparison of the pineal complex, retina and cerebrospinal fluidcontacting neurons by immunocytochemical antirhodopsinreaction. Z. Mikrosk. Anal. Forsch. 94, 623-640.
WAGNER, H.-J. (1978). Cell types and connectivity patterns inmosaic retinas. Adv. Anal., Einbr. Cell Biol. 55, 9-79.
WETTS, R. AND FRASER, S. E. (1988). Multipotent precursors cangive rise to all major cell types of the frog retina. Science 239,1142-1145.
{Accepted 20 March 1990)





























