Embed Size (px)
Citation preview

oaeagtdtTtpactot1wCsACtriwpc
htr(tcis
Clinical ImmunologyVol. 90, No. 1, January, pp. 47–54, 1999Article ID clim.1998.4628, available online at http://www.idealibrary.com on
Early IL-4 Production Driving Th2 Differentiation in a Human in VivoAllergic Model Is Mast Cell Derived
Margaret Wang,* Andrew Saxon,*,†,‡ and David Diaz-Sanchez*
*The Hart and Louise Lyon Laboratory, Division of Clinical Immunology/Allergy, Department of Medicine, †The Jonsson Comprehensive
Cancer Center Institute, and ‡The Molecular Biology Institute, UCLA School of Medicine, Los Angeles, California 90095-1680itt(
rdocsrpswdcar
ua(ppbmoDtiTao
tsdsti
IL-4 is central to the formation of IgE and the devel-pment of Th2 effector cells, both key features of anllergic response. We have examined IL-4 productionarly in the formation of an allergic response by usingpreviously established human in vivo model of aller-
ic rhinitis where allergic subjects are challenged in-ernasally with allergen and the particulate pollutantiesel exhaust particles (DEP). This model is charac-erized by enhanced IgE production and deviation to ah2-type cytokine profile in nasal lavage fluid from
hese subjects. In this model, IL-4 protein and IL-4-ositive cells could first be detected 4 h after challengend maximal production was observed after 18 h. Two-olor flow cytometric analysis for the detection of in-racellular IL-4 and surface markers was performedn nasal cells recovered 4 h after challenge. At thisime, CD1171 (c-kit1) cells constituted between 65 and00% of the IL-41 cells, while 0–12% of the IL-41 cellsere CD3 positive. No IL-41 CD19/CD201 or IL-41
D561 cells were detected at 4 h. As the allergic re-ponse progressed the primary source of IL-4 changed.t the peak of IL-4 production, 18 h after challenge,D31 comprised the majority of cells staining for in-
racellular IL-4 (73 to 100%). Thus we show an initialole for cells of the mast cell/basophil lineage residingn the nasal mucosa in the initial production of IL-4,hich frames the subsequent immune response by ex-anding the repertoire of TH2 cytokine-producingells in the local microvicinity. © 1999 Academic Press
Key Words: IL-4; Th2 cells; mast cells; allergy.
INTRODUCTION
Since 1986 when Mosmann and Coffman divided Telper cells on the basis of their cytokine profiles (1),he Th1/Th2 paradigm has been attacked, revised, andefined but has generally been thought to hold truereviewed in 2). Humoral immunity involves recogni-ion of antigen leading to the clonal expansion of spe-ific Th cells into effector Th2. Thus, production of IgEn allergic disease relies on the ability of the immune
ystem to produce Th2-type cytokines. Of particular a47
mportance in this disease are IL-4 and IL-13 sincehey can initiate epsilon germline transcription andhereby in association with CD40 or CD58 ligation3–7) direct Ig class switching of B cells to IgE.
Several studies have focused on the factors that di-ect differentiation and have shown that IL-12 pro-uced by macrophages is the key factor in Th1 devel-pment (8, 9). In contrast, Th2 differentiation isritically dependent on the presence of IL-4 (10, 11), ashown by IL-4 2/2 mice, which can not mount Th2esponses to Nippostrongylus brasiliensis. A potentialaradox therefore arises: since autocrine IL-4 has beenhown to be unable to drive CD41 cell differentiation,here does IL-4 derive from, if it is itself required torive production of IL-4-producing T cells? Althoughells other than CD41 T cells can make IL-4, their roles a cellular source of early IL-4 during an allergicesponse is unclear.In order to address this question directly we have
sed our previously established human in vivo model oflocal allergic response. Diesel exhaust particles
DEP) are a major air component of inhaled particulateollution in the industrialized world. DEP and theirolycyclic aromatic hydrocarbon components haveeen demonstrated to enhance IgE production in ani-als and humans both in vivo and in vitro (12–17). In
ur human model, combined intranasal challenge withEP plus low-dose allergen induce a large increase in
he local production of allergen-specific IgE. Moreover,n allergic subjects this increase is associated with aH2-type cytokine profile, including enhanced IL-4nd IL-13 production (15, 16) and decreased productionf IFN-g.In this study we make use of this model to establish
he onset of IL-4 production during an allergic re-ponse localized to the upper respiratory system. Weemonstrate that while CD31 cells are the majorource of IL-4 during the height of cytokine production,hey are not the primary producers during the keynitial stages. We identify the cells that first make IL-4
1
s CD117 cells, cells of the basophil/mast cell lineage.1521-6616/99 $30.00Copyright © 1999 by Academic Press
All rights of reproduction in any form reserved.

S
aiastntsetv
N
foswptalD0lvitmpAstcsatwawj
m
gC(C
(DrCC
I
pbctt(PMm0aiw2PaeSssfearrc1ora
C
nbpap2sbvDc
48 WANG, SAXON, AND DIAZ-SANCHEZ
MATERIAL AND METHODS
ubjects
Twenty-seven, nonsmoking volunteers (11 malesnd 16 females) aged 20 to 53 years old were recruitedn Los Angeles, California. All had been shown to have
positive intradermal skin test (.4 mm) to eitherhort ragweed or house dust mite and an allergy his-ory consistent with allergic rhinitis. The subjects didot take any medication for the 3 days prior to or at anyime during the duration of the study. None of theubjects had any known extensive or extraordinaryxposure to pollutants. All studies were approved byhe Human Subject Protection Committee of the Uni-ersity of California at Los Angeles.
asal Lavage and Nasal Provocation with DEP andAllergen
Nasal lavages and provocation challenges were per-ormed as previously described (13, 16). Briefly, 10 mLf saline solution was delivered into the nostrils of eachubject and recovered after 10 s. A total of five lavagesere collected from each subject. The lavages wereooled and centrifuged at 1500g for 20 min to separatehe cells from the aqueous supernatant. Ten minutesfter the last lavage, subjects were internasally chal-enged with DEP, allergen, or a combination of both.EP was administered intranasally in suspension of.3 mg in 200 mL of saline. DEP were obtained from aight-duty diesel passenger car and processed as pre-iously described (13, 14). Three of the subjects werentranasally challenged with Amb a I, while the rest ofhe subjects received the house dust mite allergen Der-atophagoides pteronyssinus. Allergen challenge was
erformed as previously reported (16). Briefly, eithermb a I or Der.pteronyssinus was sprayed intranasallytarting at 10 AU in 10-fold increments until symp-oms (sneezing/irritation) appeared. No subject re-eived more than 10,000 or less than 100 AU. Allubjects displayed immediate allergic symptoms suchs sneezing, runny nose, and ocular itching. In addi-ion, 11 of the subjects reported symptoms consistentith late phase reactions (such as recurrent congestionnd rhinorrhea) some 5 to 12 h later. Nasal lavagesere then repeated exactly as above, on the same sub-
ects at several set times after challenge.
Abs
The following antibodies were used: Simultest g1/2a control (X40/X39), CD45-FITC, CD45-PE (2D1),D3-FITC, CD3-PE (SK7), CD19-FITC, CD19-PE
4G7), CD20-FITC, CD20-PE (L27), CD56-FITC,
D56-PE (NCAM16.2), IL-6-PE (AS12), IL-4-PE I3010.211), IFN-g-FITC (25723.11) from Becton–ickinson Immunocytometry Systems (San Jose, CA),
at IgG1 control (R3-34), IL-4-FITC (MP4-25D2), andD117-PE (YB5.B8) from PharMingen (San Diego,A).
ntracellular Staining and Flow Cytometry
Cells were recovered from nasal lavages, resus-ended in 50 mL PBS/10% human serum, and incu-ated at 100,00 cells/tube with the appropriate mAb forell surface markers (Becton–Dickinson Immunocy-ometry Systems) for 30 min at 4°C. Intracellular cy-okine staining was performed according to Jung et al.18), with minor modifications. After washing thrice inBS/0.1% Saponin (Sigma Chemical Co. St. Louis,O), the cells were then fixed at 4°C with 4% parafor-aldehyde for 10 min and incubated in 30 mL PBS/
.1% Saponin/0.01 M Hepes and 10% human AB serumt 25°C for 10 min. The cells were then washed againn PBS/0.1% Saponin/0.01 M Hepes and were stainedith 5 mL of excess intracellular cytokine antibodies at5°C for 30 min. The cells were then washed twice inBS, finally resuspended in 500 mL PBS, and immedi-tely collected and analyzed on a FACScan flow cytom-ter (Becton–Dickinson) using LYSIS II software.taining specificity was ensured by the use of a controlample with excess recombinant IL-4. A control sampletained with propidium iodide was used to set side andorward scatter gates so as to exclude dead cells. Tonsure that intracellular IL-4 signals were not due ton uptake of secreted proteins binding at their surfaceeceptors, we blocked the cells in 10% human AB se-um prior to the intracellular staining procedure. Aomparison of cells stained with IL-4 after blocking in0% human AB serum to cells stained with IL-4 with-ut serum blocking demonstrated that human AB se-um effectively prevented secreted IL-4 from bindingt its receptor sites (data not shown).
ytokine and IgE Measurements
IL-4 and IL-6 were measured in nasal lavage super-atants using commercial ELISA kits (Endogen, Cam-ridge, MA). The sensitivity of these assays was 0.2g/mL. To ensure the efficacy of our model, total andntibody-specific IgE levels were measured in the su-ernatant of nasal lavages performed on our subjects4 h and 4 days after allergen 6 DEP challenge. Mea-urements of IgE levels were done by ELISAs and haveeen previously described in detail (13–16, 19). As pre-iously seen (13, 16), in all subjects, challenge withEP plus allergen resulted in a very significant in-
rease over baseline of both total and antigen-specific
gE production, which was apparent at 24 h and more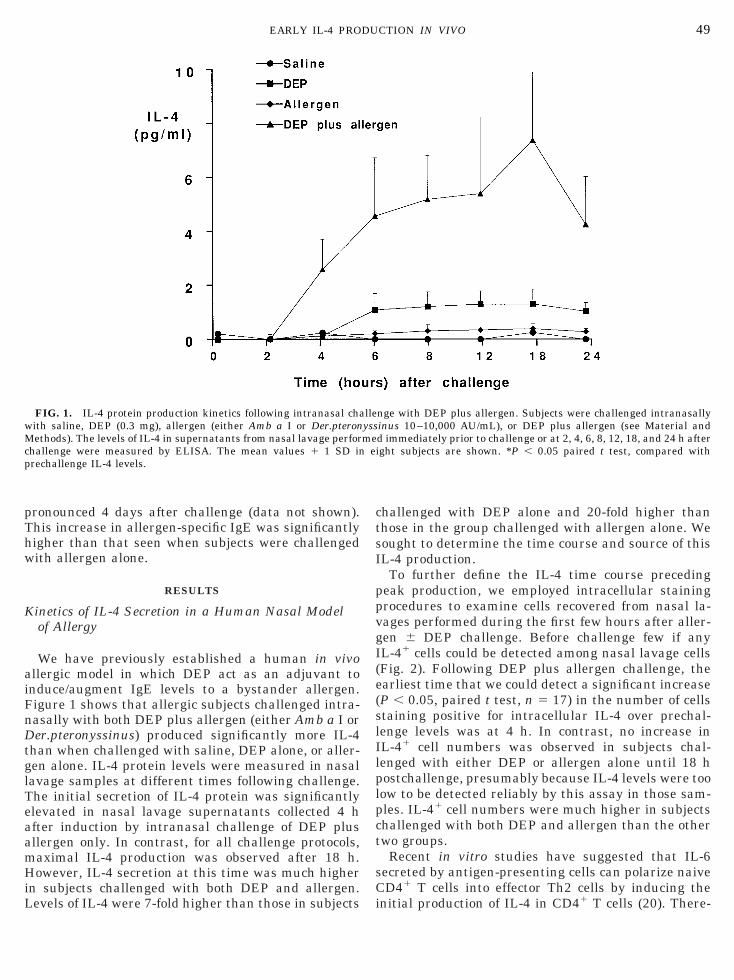
pThw
K
aiFnDtglTeaamHiL
ctsI
ppvgI(e(slIlplpct
sC
wMcp
49EARLY IL-4 PRODUCTION IN VIVO
ronounced 4 days after challenge (data not shown).his increase in allergen-specific IgE was significantlyigher than that seen when subjects were challengedith allergen alone.
RESULTS
inetics of IL-4 Secretion in a Human Nasal Modelof Allergy
We have previously established a human in vivollergic model in which DEP act as an adjuvant tonduce/augment IgE levels to a bystander allergen.igure 1 shows that allergic subjects challenged intra-asally with both DEP plus allergen (either Amb a I orer.pteronyssinus) produced significantly more IL-4
han when challenged with saline, DEP alone, or aller-en alone. IL-4 protein levels were measured in nasalavage samples at different times following challenge.he initial secretion of IL-4 protein was significantlylevated in nasal lavage supernatants collected 4 hfter induction by intranasal challenge of DEP plusllergen only. In contrast, for all challenge protocols,aximal IL-4 production was observed after 18 h.owever, IL-4 secretion at this time was much higher
n subjects challenged with both DEP and allergen.
FIG. 1. IL-4 protein production kinetics following intranasal chaith saline, DEP (0.3 mg), allergen (either Amb a I or Der.pteronethods). The levels of IL-4 in supernatants from nasal lavage perfor
hallenge were measured by ELISA. The mean values 1 1 SD inrechallenge IL-4 levels.
evels of IL-4 were 7-fold higher than those in subjects i
hallenged with DEP alone and 20-fold higher thanhose in the group challenged with allergen alone. Weought to determine the time course and source of thisL-4 production.
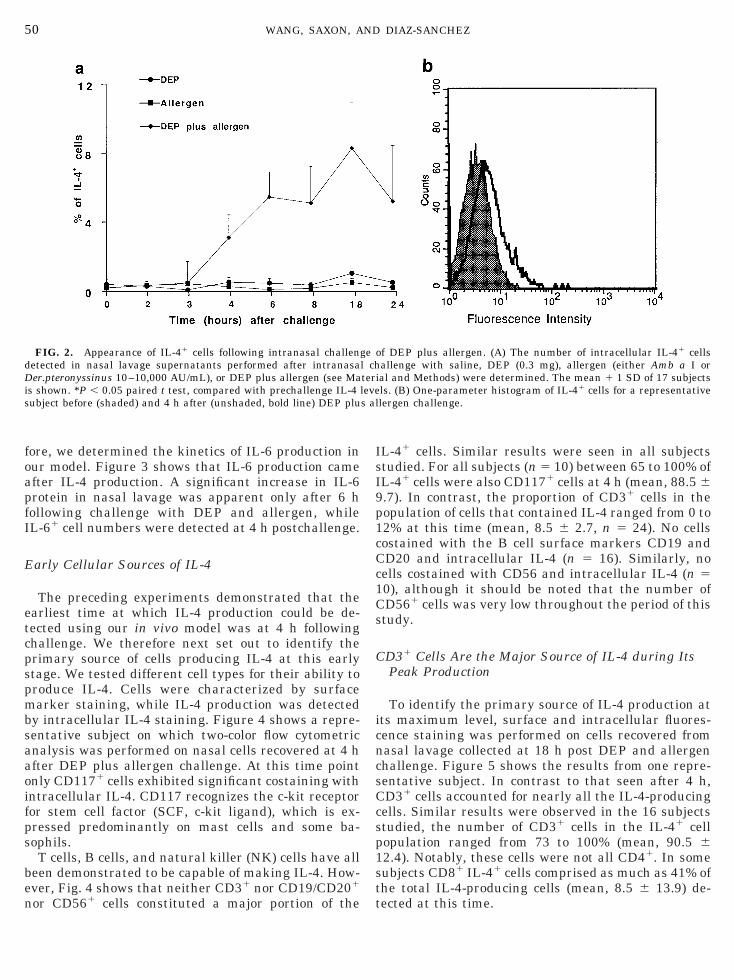
To further define the IL-4 time course precedingeak production, we employed intracellular stainingrocedures to examine cells recovered from nasal la-ages performed during the first few hours after aller-en 6 DEP challenge. Before challenge few if anyL-41 cells could be detected among nasal lavage cellsFig. 2). Following DEP plus allergen challenge, thearliest time that we could detect a significant increaseP , 0.05, paired t test, n 5 17) in the number of cellstaining positive for intracellular IL-4 over prechal-enge levels was at 4 h. In contrast, no increase inL-41 cell numbers was observed in subjects chal-enged with either DEP or allergen alone until 18 hostchallenge, presumably because IL-4 levels were tooow to be detected reliably by this assay in those sam-les. IL-41 cell numbers were much higher in subjectshallenged with both DEP and allergen than the otherwo groups.
Recent in vitro studies have suggested that IL-6ecreted by antigen-presenting cells can polarize naiveD41 T cells into effector Th2 cells by inducing the
nge with DEP plus allergen. Subjects were challenged intranasallyinus 10–10,000 AU/mL), or DEP plus allergen (see Material andd immediately prior to challenge or at 2, 4, 6, 8, 12, 18, and 24 h afterght subjects are shown. *P , 0.05 paired t test, compared with
lleyssme
ei
nitial production of IL-4 in CD41 T cells (20). There-
foapfI
E
etcpspmbsaaoifps
ben
IsI9p1cCc1Cs
C
icncsCcsp1st
dDis s al
50 WANG, SAXON, AND DIAZ-SANCHEZ
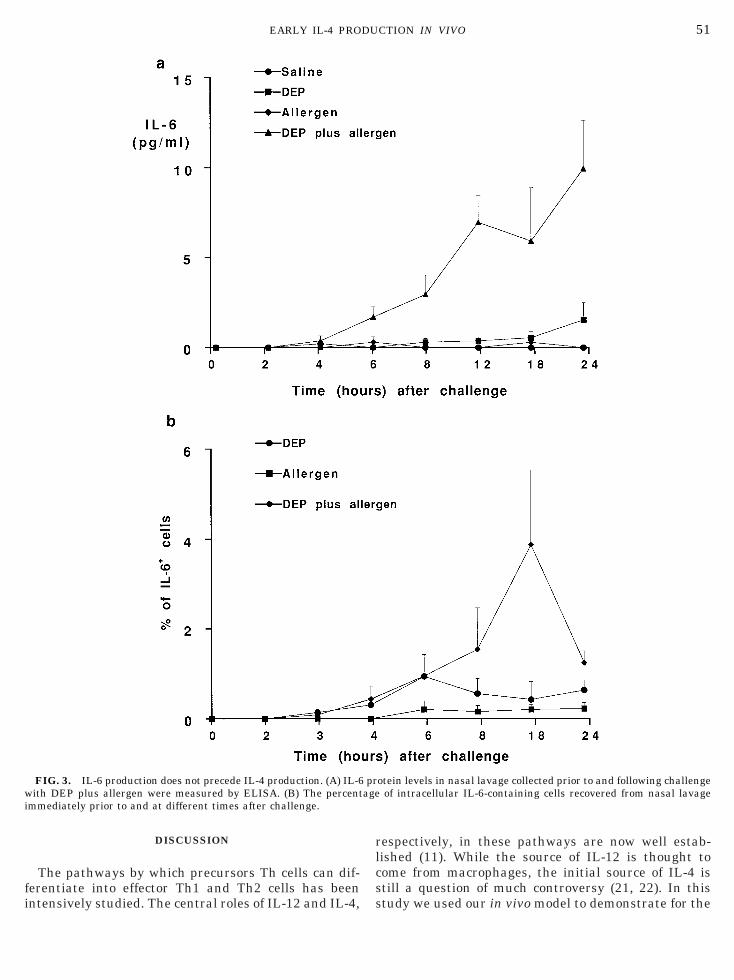
ore, we determined the kinetics of IL-6 production inur model. Figure 3 shows that IL-6 production camefter IL-4 production. A significant increase in IL-6rotein in nasal lavage was apparent only after 6 hollowing challenge with DEP and allergen, whileL-61 cell numbers were detected at 4 h postchallenge.
arly Cellular Sources of IL-4
The preceding experiments demonstrated that thearliest time at which IL-4 production could be de-ected using our in vivo model was at 4 h followinghallenge. We therefore next set out to identify therimary source of cells producing IL-4 at this earlytage. We tested different cell types for their ability toroduce IL-4. Cells were characterized by surfacearker staining, while IL-4 production was detected
y intracellular IL-4 staining. Figure 4 shows a repre-entative subject on which two-color flow cytometricnalysis was performed on nasal cells recovered at 4 hfter DEP plus allergen challenge. At this time pointnly CD1171 cells exhibited significant costaining withntracellular IL-4. CD117 recognizes the c-kit receptoror stem cell factor (SCF, c-kit ligand), which is ex-ressed predominantly on mast cells and some ba-ophils.T cells, B cells, and natural killer (NK) cells have all
een demonstrated to be capable of making IL-4. How-ver, Fig. 4 shows that neither CD31 nor CD19/CD201
FIG. 2. Appearance of IL-41 cells following intranasal challengetected in nasal lavage supernatants performed after intranasaer.pteronyssinus 10–10,000 AU/mL), or DEP plus allergen (see Ma
s shown. *P , 0.05 paired t test, compared with prechallenge IL-4 lubject before (shaded) and 4 h after (unshaded, bold line) DEP plu
or CD561 cells constituted a major portion of the t
L-41 cells. Similar results were seen in all subjectstudied. For all subjects (n 5 10) between 65 to 100% ofL-41 cells were also CD1171 cells at 4 h (mean, 88.5 6.7). In contrast, the proportion of CD31 cells in theopulation of cells that contained IL-4 ranged from 0 to2% at this time (mean, 8.5 6 2.7, n 5 24). No cellsostained with the B cell surface markers CD19 andD20 and intracellular IL-4 (n 5 16). Similarly, noells costained with CD56 and intracellular IL-4 (n 50), although it should be noted that the number ofD561 cells was very low throughout the period of thistudy.
D31 Cells Are the Major Source of IL-4 during ItsPeak Production
To identify the primary source of IL-4 production atts maximum level, surface and intracellular fluores-ence staining was performed on cells recovered fromasal lavage collected at 18 h post DEP and allergenhallenge. Figure 5 shows the results from one repre-entative subject. In contrast to that seen after 4 h,D31 cells accounted for nearly all the IL-4-producingells. Similar results were observed in the 16 subjectstudied, the number of CD31 cells in the IL-41 cellopulation ranged from 73 to 100% (mean, 90.5 62.4). Notably, these cells were not all CD41. In someubjects CD81 IL-41 cells comprised as much as 41% ofhe total IL-4-producing cells (mean, 8.5 6 13.9) de-
f DEP plus allergen. (A) The number of intracellular IL-41 cellsallenge with saline, DEP (0.3 mg), allergen (either Amb a I oral and Methods) were determined. The mean 1 1 SD of 17 subjectsls. (B) One-parameter histogram of IL-41 cells for a representativelergen challenge.
e ol chterieve
ected at this time.
fi
rlcs
w agei
51EARLY IL-4 PRODUCTION IN VIVO
DISCUSSION
The pathways by which precursors Th cells can dif-erentiate into effector Th1 and Th2 cells has been
FIG. 3. IL-6 production does not precede IL-4 production. (A) IL-6ith DEP plus allergen were measured by ELISA. (B) The percent
mmediately prior to and at different times after challenge.
ntensively studied. The central roles of IL-12 and IL-4, s
espectively, in these pathways are now well estab-ished (11). While the source of IL-12 is thought toome from macrophages, the initial source of IL-4 istill a question of much controversy (21, 22). In this
otein levels in nasal lavage collected prior to and following challengeof intracellular IL-6-containing cells recovered from nasal lavage
pr
tudy we used our in vivo model to demonstrate for the

fipaicttm
ivdoswamoiegcCfce
(ssdclmtp
datcoNwawbNTontcl3ssvewmtSoikCo
iWr(dlcmIclti
pal(s
52 WANG, SAXON, AND DIAZ-SANCHEZ
rst time the cellular sources and the changes in IL-4roduction that occur in humans during the onset of anllergic response. We show that the early source of IL-4s non-T-cell derived and is predominantly CD1171
ells and that as the response develops, T cells becomehe predominant producers of IL-4. CD117 recognizeshe c-kit receptor for SCF and can be present on bothast cells and, to a lesser degree, on basophils.Although basophils and mast cells have been shown
n vitro to produce large quantities of IL-4 upon acti-ation (23, 24), their role in Th2 development has beenownplayed by some since it is argued that they areften not present at sites of primary T cell activationuch as the lymphoid organs. However, in this studye have used a model for allergic rhinitis in whichllergen stimulation occurs at mucosal sites whereast cells and basophils abound. Indeed, many pieces
f evidence point to a potential function for these cellsn the clonal expansion of Th2 cells: Mast cells canxpress MHC class II molecules and present immuno-enic peptides to CD41 T cells (25). The priming of Tells for IL-4 production in vivo generally requiresD28-B7 interactions, while activation of already dif-
erentiated Th2 cells is independent of B7 (26). Mastells can provide this costimulation since they may
FIG. 4. Early production of IL-4 during an allergic response isredominantly from CD1171 cells. Nasal cells were collected prior tond 4 h post DEP plus allergen challenge and stained for intracel-ular IL-4 and the surface markers (A) CD117 (B) CD19 and CD20,C) CD3, and (D) CD56. The result from one representative subject ishown.
xpress both CD80 (B7-1) and CD86 (B7-2) molecules f
27). The finding that these cells are also the earliestource of IL-4 which can be visualized in our modeluggests an important role for them in initiating Th2ifferentiation. Of course we can only state that c-kit1
ells are the earliest detectable source of IL-4 since lowevels of IL-4 below the limit of detection of our assays
ay be produced earlier. However, it does seem clearhat in this model, IL-4 production from CD1171 cellsrecedes that from CD31 cells.Several other cell types have been offered as candi-
ates for inducers for T-cell-derived IL-4 and it is prob-ble that the route of immunization and the nature ofhe antigen or infection will determine which is the keyell in Th2 induction (21). Much attention has focusedn subpopulations of CD41 cells themselves. In mice,K1.11 CD41 T cells can transcribe and secrete IL-4ithin 90 min upon activation in vivo with anti-CD3ntibodies or superantigen (28). However, stimulationith anti-CD3 presumably restricts the cells that maye activated. Indeed, other studies have shown thatK11 T cells are not essential for IL-4 production orh2 responses following challenge with antigen (29). Inur study, NK cells as identified by the CD56 mAb didot have any apparent role in IL-4 production, al-hough it should be noted that the number of CD561
ells detected was very low. Different T cell subpopu-ations have also been proposed as IL-4 sources (30,1), while other studies have suggested non-T-cellources. Intraperitoneal injection of Schistosoma man-oni eggs into certain strains of mice will lead to aigorous Th2 response to helminth antigen in which anarly burst of IL-4 (2 to 12 h after antigen exposure)ill occur. This burst originates from eosinophils, withast cells producing IL-5 and thereby recruiting addi-
ional eosinophils into the local environment (32, 33).till others have demonstrated that either naive T cellsr APCs can induce Th2 responses indirectly by induc-ng IL-4 production through secretion of other cyto-ines. Both IL-6 and IL-7 have been shown to activateD41 cells to produce IL-4 in vitro (20, 34), although inur model IL-6 production did not precede that of IL-4.The in vivo human model used in our study mimics
nduction of the expression of a local allergic response.e have shown that DEP exposure in vivo alters the
egulation of local IgE production in the human nose13, 15–17) and results in increased IgE antibody pro-uction. Combined nasal challenge with DEP plus al-ergen has been shown to result in a local Th2-typeytokine pattern characterized by an increased level ofRNA for IL-4, IL-5, IL-10, and IL-13 and decreased
NF-g (16). One limitation of this model is that weannot be sure that the cells recovered in the nasalavage are truly representative of the cells present inhe nasal environment. However, it is noteworthy thatn a fashion analogous to our model, nasal mast cells
rom subjects with perennial allergic rhinitis, a disease
cc
bcIpatdI
tPlNI
1
1
1
1
1
1
1
po
53EARLY IL-4 PRODUCTION IN VIVO
haracterized by elevated IgE production, show in-reased expression of IL-4 (35).
In conclusion, we propose that cells of the mast cell/asophil lineages residing in the nasal mucosa areritical contributors in the early or initial production ofL-4 during a local mucosal allergic response. This IL-4roduction drives the response toward Th2 polaritynd expands the repertoire of IL-4-producing cells inhe local microvicinity. Subsequently, other cells, pre-ominantly CD41 cells, become the major source ofL-4 as the polar Th2 response is established.
ACKNOWLEDGMENTS
We thank Ms. Minna Jyrala and Birgitte Keld for their excellentechnical assistance. This work was supported by United Statesublic Health Service Grant AI-15251 and the UCLA Asthma, Al-
ergy, and Immunologic Disease Center (AI-34567) funded by theational Institute of Allergy and Infectious Disease and the National
nstitute of Environmental Health Science.
REFERENCES
1. Mosmann, T. R., Cherwinski, H., Bond, M. W., Giedlin, M. A.,and Coffman, R. L., Two types of murine helper T cell clone. I.Definition according to profiles of lymphokine activities and se-creted proteins. J. Immunol. 136, 2348–2357, 1986.
2. Del Prete, G., The concept of type-1 and type-2 helper T cells andtheir cytokines in humans. Int. Rev. Immunol. 16, 427–455,1998.
3. Jabara, H. H., Fu, S. M., Geha, R. S., and Vercelli, D., CD40 andIgE: Synergism between anti-CD40 monoclonal antibody andinterleukin 4 in the induction of IgE synthesis by highly purifiedhuman B cells. J. Exp. Med. 172, 1861–1864, 1990.
4. Zhang, K., Clark, E. A., and Saxon, A., CD40 stimulation pro-vides an IFN-g-independent and IL-4-dependent differentiationsignal directly to human B cells for IgE production. J. Immunol.146, 1836–1842, 1991.
5. Punnonen, J., Aversa, G., Cocks, B. G., Mckenzie, A. N. J.,Menon, S., Zurawski, G., Malefyt, R. W., and de Vries, J. E.,Interleukin 13 induces interleukin 4-independent IgG4 and IgEsynthesis and CD23 expression by human B cells. Proc. Natl.
FIG. 5. Production of IL-4 at the peak of the allergic response isost DEP plus allergen challenge and stained for intracellular IL-4 anne representative subject is shown.
Acad. Sci. USA 90, 3730–3734, 1993.
6. Diaz-Sanchez, D., Chegini, S., Zhang, K., and Saxon, A., CD58(LFA-3) stimulation provides a signal for human isotype switch-ing and IgE production distinct from CD40. J. Immunol. 153,10–20, 1994.
7. Kimata, H., Fujimoto, M., Ishioka, C., and Yoshida, A., Hista-mine selectively enhances human immunoglobulin E (IgE) andIgG4 production induced by anti-CD58 monoclonal antibody. J.Exp. Med. 184, 357–364, 1996.
8. Hsieh, C. S., Macatonia, S. E., Tripp, C. S., Wolf, S. F., O’Garra,A., and Murphy, K. M., Development of TH1 CD41 T cellsthrough IL-12 produced by Listeria-induced macrophages. Sci-ence 260, 547–549, 1993.
9. D’Andrea, A., Rengaraju, M., Valiante, N. M., Chehimi, J., Ku-bin, M., Aste, M., Chan, S. H., Kobayashi, M., Young, D., andNickbarg, E., Production of natural killer cell stimulatory factor(interleukin 12) by peripheral blood mononuclear cells. J. Exp.Med. 184, 1387–1398, 1992.
0. Swain, S. L., Bradley, L. M., Croft, M., Tonkonogy, S., Atkins, G.,Weinberg, A. D., Duncan, D. D., Hedrick, S. M., Dutton, R. W.,and Huston, G., Helper T-cell subsets: Phenotype, function andthe role of lymphokines in regulating their development. Immu-nol. Rev. 123, 115–144, 1991.
1. Seder, R. A., and Paul, W. E., Acquisition of lymphokine-produc-ing phenotype by CD41 T cells. Annu. Rev. Immunol. 12, 635–673, 1994.
2. Muranaka, M., Suzuki, S., Koizumi, K., Takafuji, S., Miyamoto,T., Ikemori, R., and Tokiwa, H., Adjuvant activity of diesel-exhaust particulates for the production of IgE antibody in mice.J. Allergy Clin. Immunol. 77, 616–623, 1986.
3. Diaz-Sanchez, D., Dotson, A. R., Tekenaka, H., and Saxon, A.,Diesel exhaust particles induce local IgE production in vivo inhuman and alter the pattern of IgE mRNA isoforms. J. Clin.Invest. 94, 1417–1425, 1994.
4. Takenaka, H., Zhang, Z., Diaz-Sanchez, D., Tsein, A., and Saxon,A., Enhanced hman IgE results from exposure to the aromatichydrocarbons in diesel exhaust: Direct effects on B cells IgEproduction. J. Allergy Clin. Immunol. 95, 103–115, 1995.
5. Diaz-Sanchez, D., Tsien, A., Casillas, A., Dotson, A. R., andSaxon, A., Enhanced nasal cytokine production in human beingsafter in vivo challenge with diesel exhaust particles. J. AllergyClin. Immunol. 98, 114–123, 1996.
6. Diaz-Sanchez, D., Tsien, A., Fleming, J., and Saxon, A., Com-bined diesel exhaust particulate and ragweed allergen challengemarkedly enhanced in vivo nasal ragweed-specific IgE andskews cytokine production to a T helper 2-type pattern. J. Im-
dominantly from CD31 cells. Cells were collected prior to and 18 hhe surface markers (A) CD3, (B) CD4, and (C) CD8. The results from
pred t
munol. 158, 2406–2413, 1997.

1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
R
54 WANG, SAXON, AND DIAZ-SANCHEZ
7. Fujieda, S., Diaz-Sanchez, D., and Saxon, A., Combined nasal chal-lenge with diesel exhaust particles and allergen induces in vivo IgEisotypr switching. Am. J. Respir. Mol. Biol. 19, 507–512, 1998.
8. Jung, T., Schauer, U., Heusser, C., Neumann, C., and Rieger, C.,Detection of intracellular cytokines by flow cytometry. J. Immu-nol. Methods. 159, 197–207, 1993.
9. Macy, E., Kemeny, D. M., and Saxon, A., Enhanced ELISA: Howto measure less than 10 picograms of a specific protein (immu-noglobulin) in less than 8 hours. FASEB J. 2, 3003–3010, 1988.
0. Rincon, M., Anguita, J., Nakamura, T., Fikrig, E., and Flavell,R. A., Interleukin (IL)-6 directs the differentiation of IL-4-pro-ducing CD41 T cells. J. Exp. Med. 185, 461–469, 1997.
1. Coffman, R. L., and von der Weid, T., Multiple pathways for theinitiation of T helper 2 (Th2) responses. J. Exp. Med. 185, 373–375, 1997.
2. Pearce, E. J., and Reiner, S. L., Induction of Th2 responses ininfectious diseases. Curr. Opin. Immunol. 7, 497–504, 1995.
3. Plaut, M., Pierce, J. H., Watson, C. J., Hanley-Hyde, J., Nordan,R. P., and Paul, W. E., Mast cell lines produce lymphokines inresponse to cross-linkage of Fc epsilon RI or to calcium iono-phores. Nature 339, 64–67, 1989.
4. Paul, W. E., Seder, R. A., and Plaut, M., Lymphokine and cyto-kine production by Fc epsilon RI1 cells. Adv. Immunol. 53, 1–29,1993.
5. Frandji, P., Tkaczyk, C., Oskeritzian, C., Lapeyre, J., Peronet,R., David, B., Guillet, J. G., and Mecheri, S., Presentation ofsoluble antigens by mast cells: Upregulation by interleukin-4and granulocyte/macrophage colony-stimulating factor anddownregulation by interferon-gamma. Cell. Immunol. 163, 37–46, 1995.
6. Gause, W. C., Halvorson, M. J., Lu, P., Greenwald, R., Linsley,P., Urban, J. F., and Finkelman, F. D., The function of costimu-latory molecules and the development of IL-4-producing T cells.
Immunol. Today 18, 115–120, 1997.7. Mecheri, S., and David, B., Unravelling the mast cell dilemma:Culprit or victim of its generosity? Immunol. Today 18, 212–215,1997.
8. Yoshimoto, T., Bendelac, A., Watson, C., Hu-Li, J., and Paul,W. E., Role of NK1.11 T cells in a TH2 response and in immu-noglobulin E production. Science 270, 1845–1847, 1995.
9. Brown, D. R., Fowell, D. J., Corry, D. B., Wynn, T. A., andMoskowitz, N. H., Cheever, A. W., Locksley, R. M., and Reiner,S. L., Beta 2-microglobulin-dependent NK1.11 T cells are notessential for T helper cell 2 immune responses. J. Exp. Med. 184,1295–1304, 1996.
0. Gollob, K. J., and Coffman, R. L., A minority subpopulation ofCD41 T cells directs the development of naive CD41 T cells intoIL-4-secreting cells. J. Immunol. 152, 5180–5188, 1994.
1. Kamogawa, Y., Minasi, L. A., Carding, S. R., Bottomly, K., andFlavell, R. A., The relationship of IL-4- and IFN gamma-produc-ing T cells studied by lineage ablation of IL-4-producing cells.Cell 75, 985–95, 1993.
2. Sabin, E. A., and Pearce, E. J., Early IL-4 production by non-CD41 cells at the site of antigen deposition predicts the devel-opment of a T helper 2 cell response to Schistosoma mansonieggs. J. Immunol. 155, 4844–53, 1995.
3. Sabin, E. A., Kopf, M. A., and Pearce, E. J., Schistosoma mansoniegg-induced early IL-4 production is dependent upon IL-5 andeosinophils. J. Exp. Med. 184, 1871–1978, 1992.
4. Webb, L. M., Foxwell, B. M., and Feldmann, M., Interleukin-7activates human naive CD41 cells and primes for interleukin-4production. Eur. J. Immunol. 27, 633–40, 1997.
5. Pawankar, R., Okuda, M., Yssel, H., Okumura, K., and Ra, C.,Nasal mast cells in perennial allergic rhinitics exhibit increasedexpression of the Fc epsilonRI, CD40L, IL-4, and IL-13, and caninduce IgE synthesis in B cells. J. Clin. Invest. 99, 1492–1499,
1997.eceived August 26, 1998; accepted September 28, 1998

![The prevention of 2,4-dinitrochlorobenzene-induced ...€¦ · [5, 6] and T-helper 2 (Th2)-dependent cells [7, 8]. Mast * Correspondence: pm.thehoo@gmail.com; epiko@khu.ac.kr 2Department](https://img.pdfslide.us/doc/110x75/60a198517546aa433c0bf138/the-prevention-of-24-dinitrochlorobenzene-induced-5-6-and-t-helper-2-th2-dependent.jpg)



























