-
B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9 4
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Dystrophic serotonergic axons in neurodegenerative diseases
Efrain C. Azmitiaa,b,c,⁎, Ralph Nixonb,c,d
aDepartment of Biology and Center for Neural Science, 100
Washington Square East, New York, New York 10003, USAbDepartment of
Psychiatry, New York School of Medicine, New York, New York 10016,
USAcDepartment of Cell Biology, New York School of Medicine, New
York, New York 10016, USAdNathan Kline Institute, Orangeburg, New
York 10962, USA
A R T I C L E I N F O
⁎ Corresponding author. 10-09 Silver BuildingE-mail address:
[email protected] (E.C. Azmi
0006-8993/$ – see front matter © 2008
Elsevidoi:10.1016/j.brainres.2008.03.060
A B S T R A C T
Article history:Accepted 14 March 2008Available online 7 April
2008
Neurodegenerative diseases such as Parkinson's disease (PD),
frontal lobe dementia (FLD) anddiffuse Lewy-body dementia (DLBD)
have diverse neuropathologic features. Here we reportthat serotonin
fibers are dystrophic in the brains of individuals with these three
diseases. Inneuropathologically normal (control) brains (n=3),
serotonin axons immunoreactive (IR) withantibodies against the
serotonin transporter (5-HTT) protein were widely distributed in
cortex(entorhinal and dorsolateral prefrontal), hippocampus and
rostral brainstem. 5-HTT-IR fibers-of-passage appeared thick,
smooth, and unbranched in medial forebrain bundle, mediallemniscus
and cortex white matter. The terminal branches were fine, highly
branched andvaricose in substantia nigra, hippocampus and cortical
gray matter. In the diseased brains,however, 5-HTT-IR fibers in the
forebrain were reduced in number and were frequentlybulbous,
splayed, tightly clustered and enlarged. Morphometric analysis
revealed significantdifferences in the size distribution of the
5-HTT-IR profiles in dorsolateral prefrontal areabetween
neurodegenerative diseases and controls. Our observations provide
directmorphologic evidence for degeneration of human serotonergic
axons in the brains ofpatients with neurodegenerative diseases
despite the limited size (n=3 slices for each region(3) from each
brain (4), total sliceswas n=36) and the lack of extensive clinical
characterizationof the analyzed cohort. This is the first report of
dystrophic 5-HTT-IR axons in postmortemhuman tissue.
© 2008 Elsevier B.V. All rights reserved.
Keywords:DegenerationNeuropathologyParkinson'sLewy-bodyFrontal
lobe
dementiaAgingPostmortemHippocampusPrefrontalEntorhinalSubstantia
nigraHuman
1. Introduction
The anatomy and plasticity of 5-HT projecting axons
inexperimental animals has been extensively studied. The fine5-HT
axons were first visualized in the rat brain withhistochemical
fluorescence (Fuxe, 1965). Immunocytochem-ical analyses with
antibodies raised against 5-HT laterrevealed the full global
projections of the raphe neurons,showing also that 5-HT
fibers-of-passage are thick and un-branched while the terminal
fibers are fine, highly branched
, 100 Washington Squaretia).
er B.V. All rights reserved
and varicose (Steinbusch, 1981; Azmitia and Gannon, 1983).Dense
innervation is apparent throughout the neuroaxis andit has been
suggested that every cell in the rat cortex is near a5-HT
containing axon (Molliver, 1987).
It has been difficult to study the morphology of the 5-HTaxonal
system in the adult human brain. Postmortem auto-lysis and the
usual methods of postmortem brain fixationresult in the loss of
5-HT from axonal storage sites, whichmakes histochemical
fluorescence and 5-HT immunocyto-chemistry ineffective. Antibodies
raised against monoamine
East, New York, New York 10003-6688, USA. Fax: +1 212 995
4015.
.
mailto:[email protected]://dx.doi.org/10.1016/j.brainres.2008.03.060
-
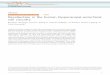
Fig. 1 – 5-HTT-IR axons are seen in brains from control
individuals. A. In the midbrain/substantia nigra sections heavy
fiberlabeling is seen in the medial lemniscal with the fibers
throughout the entire pathway. These fibers-of-passage have
smallvaricosities and often form tight bundles (arrows). Scale bar
is 200μm.B. In the hippocampus and entorhinal areas the
5-HTT-IRaxons are forming terminal boutons. In these sections
larger boutons can be seen (arrow). Scale bar is 50μm. C. In the
prefrontalcortex, the 5-HTT-IR fibers form clusters around cell
bodies in pyramidal layers (arrows). Scale bar is 200 μm.
186 B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
hydroxylase enzymes label 5-HT cell bodies and processes inthe
human brainstem, but fail to reveal the distal terminalfibers
(Craven et al., 2005; Underwood et al., 1999; Baker et al.,1991;
Haan et al., 1987).
A better approach may be to focus on the 5-HT
transporterprotein. The 5-HT transporter (5-HTT) protein is
specific to 5-HTaxons in the adult brain, although occasional
astrocytic stainingcan be observed. The axonal sitewas visualized
in experimentalanimals by autoradiography after uptake 3H-5-HT by
thetransporter protein in situ (Descarries et al., 1975) or in
vitro infresh brain slices (Azmitia, 1981). Antibodies to 5-HTT
allowedthemorphology of 5-HT terminal axons to be studied
immuno-cytochemically in brains of non-human animals (Qian et
al.,1995; Zhou et al., 1996). In humans, radioactive ligands
specificfor the5-HT transporter permit thegross regional
distributionof5-HT axons to be revealed in vivo by PET (Abi-Dargham
et al.,1966; Szabo et al., 1996) or in postmortem tissue by film
auto-radiography (Varnas et al., 2004; Chinaglia et al., 1993).
5-HTTimmunoreactive (IR) axons of normalmorphology were seen inthe
postmortem brainstem and prefrontal cortex of individualswithout
known neurological disorders (Qian et al., 1995; Austinet al.,
2002). The 5-HTT-IR axons were described as fine,
highlybranchedandvaricose, similar inappearance to thosedescribedin
non-human animals.
In animal studies, serotonin axons can be damaged under avariety
of conditions. Immunocytochemical analysis or immu-nofluoresence
labeling with antibodies raised against 5-HTdetects dystrophic
immunoreactive fibers in rats after neuro-toxic injections (Wiklund
and Bjorklund, 1980; Frankfurt andAzmitia, 1983); ingestion of
designer-drugs of abuse (Wilson
Fig. 2 – Dystrophic 5-HTT-IR axons are seen, but infrequently,
inentorhinal cortex is heavily innervated with 5-HTT-IR axons.
Amdense clusters, which can condense into a larger degenerating
prolarge varicosities can be found (arrow). C. In the dendritic
regionScale bar is 50 μm in all panels.
and Molliver, 1994); and in aged animals (van Luijtelaar et
al.,1989). Immunocytochemistry with antibodies to 5-HTT alsoshows
dystrophic 5-HTT immunoreactive axons after neuro-toxic injections
(Zhou et al., 1996). These dystrophic axonsare enlarged, bulbous or
splayed after acute damage to 5-HTfibers, suggesting an active
degeneration process. In aginganimals, 5-HT fibersmay appear
abnormally fine, varicose andtightly clustered suggesting atrophy
and withdrawal fromtheir forebrain targets (van Luijtelaar et al.,
1980).
Despite a preponderance of pharmacological, neurochem-ical,
andmolecular evidence that the 5-HT system is disruptedin many
clinical disorders, dystrophic serotonergic axons inthe human brain
have never been described. There is indirectevidence that 5-HT
axons are damaged in neurological dis-orders such as FLD (Sparks
andMarkesbery, 1991; Menza et al.,1999); PD (Halliday et al., 1990;
Marksteiner et al., 2003); Al-zheimer's disease and ischemic heart
disease (Stout et al,2003); and DLBD (Ballard et al., 2002). For
example in DLBD,Lewy bodies occur in the dorsal raphe nucleus, and
serotoninlevels are markedly reduced in the striatum, neocortex
andfrontal cortex (Langlais et al., 1993; Ohara et al., 1998;
Perryet al., 1993).
In this study, we obtained well-characterized postmor-tem
neurodegenerative brains from patients with PD, FLDand DLBD, as
well as from neuropathologically normal (con-trol) individuals. In
neuropathologically normal brains, 5-HTTimmunocytochemistry showed
typical 5-HT axonal morphol-ogy in abundant axons, including both
projecting fibersand terminating regions, and only rare dystrophic
fibers. Bycontrast, dystrophic 5-HTT-IR fibers were frequent and
wide-
the brain of a control 79-year-old-male. A. Layer III ofong the
normal fibers, abnormal fibers can be seen formingfile (arrows). B.
In the polymorphic area of the DG abnormallyof CA3 degenerating
profiles are seen among normal fibers.
-
Fig. 3 – 5-HTT-IR axons in brains from patients diagnosed with
diffuse Lewy-body dementia. A. This picture shows normalappearing
5-HTT-IR axonal bundles in the dorsal aspect of the section just
above the inferior colliculus (IC). There is evidence ofdystrophic
5-HTT-IR axons in the fiber bundles (arrows). Scale bar is 200μm.
B. Normal and abnormal (arrows) terminals in thesubstantia nigra.
Two pigmented nigra neurons can be seen. Scale bar is 50 μm. C.
Splayed 5-HTT-IR terminals seen in CA3dendritic region. Scale bar
is 50 μm. D. Tight clusters of 5-HTT-IR axons in layer III of
entorhinal cortex. The number of labeledfibers is reduced in this
region. Scale bar is 50 μm. E. Tight cluster of 5-HTT-IR fibers in
layer III of prefrontal cortex. Scale bar is50 μm. F. In deep
layers of cortex, aggregates of fibers are found among dystrophic
5-HTT-IR axons with enlarged varicosities.Scale bar is 200 μm.
187B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
spread in all brains from patients with degenerative
disease.Using amorphometric software system, we identified
significantalterations in thenumber and sizes of 5-HTT-IR axonal
structuresin the prefrontal terminal regions of all three
neurodegenerativediseases. To our knowledge, this study provides
the first directevidence that 5-HT axons in the human brain are
vulnerable todegeneration in neurodegenerative states.
2. Results
2.1. 5-HTT patterns in neuropathologically normal brains
5-HTT-IR fibers were seen in all examined regions of
neuro-pathologically normal brains. The 5-HTT-IR axons formedlarge,
dense bundles in the rostral midbrain especially withinthemedial
lemniscus (arrows, Fig. 1A). The 5-HTT-IR fibers-of-passage were
thick, unbranched and either long and straight(possibly myelinated
fibers) or wavy in appearance (Fig. 1A).
2.1.1. Ascending projectionsWe observed fine and varicose, or
coarse and non-varicose 5-HTT-IR axons in all regions examined. The
non-varicose fiberswere particularly abundant in brainstem areas
associatedwith major ascending pathways as well as in the forebrain
incorpus callosum, fornix, and perforant path fibers. The 5-HTT-IR
fibers in these forebrain pathways ascended into the grayterminal
areas of the cortex where the fibers became thin-ner and more
branched. In addition, a dense fiber plexus oftangential projecting
fibers was found in layer I of entorhinal(Fig. 1B) and prefrontal
cortical regions (Fig. 1C). These fiberswere in close proximity to
the pia layer and could be seenextending into layers II and
III.
2.1.2. Terminal distributionIn the terminal areas within the
midbrain (region of the Sub-stantia Nigra), the 5-HTT antibody
labeling resolved thin, va-ricose, and highly branched 5-HTT-IR
axonal fibers. Distinct 5-HTT-IR boutons were visualized at the
terminals of thesefibers. In entorhinal and prefrontal cortices,
the distribution of5-HTT-IR fibers extended throughout all cortical
layers, withextensive branching seen in deeper layers near large
pyra-midal neurons. The 5-HTT-IR axons were fine, highly bran-ched
and formed irregularly spaced varicosities (Figs. 1B-C).The
terminals were frequently seen closely surrounding largepyramidal
neurons in what can be described as pericellularplexuses.
Occasional dystrophic 5-HTT-IR axons (Fig. 2) were seen
intheparahippocampal terminal regions, especially obvious in
theoldest brain examined (B3573; male, 79 years of age, postmor-tem
interval of 15.3 h). The abnormal 5-HTT axons – tightgrouping of
small varicosities (clustering) (Fig. 2A, arrows) andenlarged
varicosities (Figs. 2B–C, arrows) – were scatteredamong typical
fibers, especially in the hilus of the dentategyrus (Fig. 2B) and
the deep entorhinal layers (layers V–VI)(Fig. 2C) and in the CA
layers of the hippocampus (not shown).However, therewasnoevidence
in controlmaterial of advancedneurodegeneration (splayed endings or
dark aggregates ofstained material) or a marked reduction in fiber
density.
2.2. 5-HTT patterns in the brain in
neurodegenerativediseases
5-HTT-IR axons were found in all regions of the brains
fromindividuals with any of the three neurodegenerative
diseasesexamined. In the brainstem from these cases, 5-HTIR
axonsappeared normal, although occasional abnormal fibers were
-
Fig. 4 – 5-HTT-IR axons in brains frompatients diagnosedwith
Parkinson's disease. A. 5-HTT-IR axons in the red nucleus showsome
abnormal fibers (arrows) among many normal fibers. Scale bar is 200
μm. B. In midline areas 5-HTT-IR fibers withenlarged varicosities
are found among normal fibers. Scale bar is 50μm. C. Dystrophic
5-HTT-IR axonswith enlarged varicosityin the dendritic regions of
CA1. Scale bar is 50 μm. D. Dystrophic 5-HTT-IR axons are splayed
and degenerating in the upperlayers of the entorhinal cortex. Scale
bar is 50 μm. E. Fine degenerating 5-HTT-IR axons in the upper
layer of prefrontal cortex.Scale bar is 50 μm. F. Fine, dense
clusters of 5-HTT-IR axons in the deeper layers of prefrontal
cortex. Scale bar is 50 μm.
188 B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
seen. Similarly, in the corpus callosum of the forebrain, long
5-HTT-IR axons could be followed with no apparent evidence
ofpathology, although in the gray matter from this brain regionand
other cortical gray areas the number of 5-HTT-IR axonsappeared
markedly reduced. Significant numbers of 5-HTT-IRaxons were
dystrophic in shape in prefrontal and temporalcortical areas. The
hippocampus and entorhinal and prefron-tal cortices showed four
main types of abnormal profiles: (1)enlarged, twisted and swollen
varicosities, sometimes appear-ing as ballooned profiles; (2) fine
fibers forming tight clus-
Fig. 5 – 5-HTT-IR axons in brains from patients diagnosed with
Frois seen in the substantia nigra nucleus. No dystrophic 5-HTT-IR
fibethe medial lemniscal tract. No dystrophic 5-HTT-IR fibers seen.
Scaldentategyrus.Scalebar is50μm.D.Several
splayedanddegeneratinNote reduced appearance of normal 5-HTT-IR
fibers. Scale bar is 50layers II–III of the prefrontal cortex. Note
normal tangential fibers in lthe deeper layers of prefrontal
cortex. 5-HTT-IR axonal innervation
ters; (3) isolated splayed fibers with an irregular shape;
and(4) densely labeled aggregates and degenerating profiles.
2.3. Diffuse Lewy-body dementia
In brainstem sections from cases of DLBD, the 5-HTT-IR
fibersappeared less dense than normal and occasionally the
pro-cesses were abnormal in appearance and darkened (arrows,Figs.
3A–B). In both the entorhinal cortex (Figs. 3B–C) andprefrontal
(Figs. 3E–F), 5-HTT-IR axons in layer I, close to the pia,
ntal Lobe Dementia. A. Dense innervation by 5-HTT-IR fibersrs
seen. Scale bar is 200 μm. B. Heavy labeling of the fibers ine bar
is 200 μm. C. Enlarged varicosities seen in hilus area ofg5-HTT-IR
terminals seen in theupper layerof entorhinal cortex.μm. E. Dense
clustering and aggregation of 5-HTT-IR axons inayer I. Scale bar is
50μm. F. Enlarged 5-HTT-IR axons are seen inappears reduced from
normal. Scale bar is 50 μm.
-
Fig. 6 – This figure shows the results from morphometricanalysis
of the labeled objects found in prefrontal cortex ofcontrol and
diseased brains using threshold setting ofimmunoreactive density.
A. The total number of particles(varicosities, fibers and
degenerating axons) was counted inan area of 0.5mm2 in layers III–V
of the cortex. Each bar is themean of three brains from each group
(see Experimentalprocedures for details). The number of particles
selected waslower for all diseased-groups compared to normal
prefrontalcortex, and was significant for the DLBD and PD.B. A
histogram of all particle areas was made form theparticles selected
for part A. The percentage of the totalnumber of particles which
had the smallest area (
-
Table 1 – The diagnosis, demographic, postmortem interval (PMI)
history of depression and antidepressant treatment
Brain # Diagnosis Gender Age PMI Depression Antidepressant
treatment
B2374 None M 72 8.5 – –B2469 None F 54 3.5 – –B3573 None M 79
15.3 – –B4426 DLBD M 75 23.5 + –B4866 DLBD M 67 14.85 + –B5085 DLBD
M 68 5.5 + +B4927 FLD F 68 14.33 + +B5007 FLD F 72 6.58 N/A
N/AB5035 FLD F 75 20 + +B4934 PD M 68 17.16 N/A N/AB5207 PD M 64
18.58 N/A N/AB5227 PD M 77 26 + +
190 B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
axons in postmortem brain sections from individuals withouta
diagnosed neurological or psychiatric diseasewere similar tothose
described previously in humans and animal studies. Wedetected
extensive neuropathology of 5-HTT-IR axons, how-ever, in brains of
patientswith Parkinson's disease, frontal lobedementia or diffuse
Lewy-body dementia. This is the firstreport of 5-HT axonal
pathology in humans and shows that the5-HT system in the human
brain is vulnerable to degenerationin various neurodegenerative
disorders, as shown previouslyin animal models of disease.
3.1. Serotonin system and neurodegeneration
Although numerous pathologies in late-age onset
neurodegen-erative diseases have been identified with markers of
specificpathologies (eg.β-amyloid, ubiquitin,
hyperphosphorylated-tauor α-synuclein reactive), there are
relatively few examples of atransmitter-specific neuronal
degeneration in neurodegenera-tive diseases (see reports on
acetylcholine: Bossy-Wetzel et al.(2004), Perry et al. (1993) and
norepinephrine: Haglund et al.(2006)). Even in Parkinson's
diseases, we could not find a des-cription of dystrophic
dopaminergic fibers (see Dickson et al.(1994)) although the normal
distribution of projections fromdopaminergic neurons to the human
caudate has been des-cribed (Kung et al., 1998).
Wenowreport that serotonin fibers, immunocytochemicallylabeled
with antibody against the 5-HTT, show extensive andwide-spread
pathology in the brains of patients with PD, FLD orDLBD. In animal
studies, serotonin fibers degenerate whenexposed to a variety of
environmental and neurotoxic factors(e.g. van Luijtelaar et al.
(1989)—aging; O'Hearn et al. (1988)—MDMA and related drugs; Zhou et
al. (1994)—alcohol; Liu andNakamura (2006), Aucoin et al.
(2005)—amyloid; Baumgartenand Bjorklund (1976), Frankfurt and
Azmitia (1984)—5,7-DHT).Our findings demonstrate that human
serotonin axons are alsovulnerable to degeneration in pathological
states.
3.2. Comparison of axonal pathology
Depletion of normal fibers and appearance of
degeneratingprofileswere evident in all three groups of brains
frompatientswith degenerative diseases. There was no evidence that
gen-der or postmortem interval contributed to these
observations.Recent unpublished work using 5-HTT immunostaining
(n=22
brains from neurologically typical normal subjects, range 32–85
years, average age 57.2 years and n=3 female) showed noevidence of
the frequent severe 5-HTT-specific axonal pathol-ogy reported here
in the brains from patients with neurode-generative diseases. The
5-HTT-IR dystrophic axonal profilesin this report are similar in
appearance to those describedafter systemic administration of
neurotoxin (Baumgarten andBjorklund, 1976) and designer-drug
(Molliver and Molliver,1990) induced degeneration of 5-HT fibers in
rat brain. Thedystrophic axons seen in the deep layers of FLD
prefrontalcortex exhibited increased caliber, reduced branching,
andswollen varicosities and resembled those seen after
5,7-DHTneurotoxin intracerebral injections into the 5-HT
fibers-of-passage in MFB (Frankfurt and Azmitia, 1984) or
cingulumbundle (Zhou andAzmitia, 1986). The 5-HTT fibers
inDLBDandPD brains show degenerating fibers characterized by
varicoseswelling and clustering of fine terminals. This pattern
ofdegeneration is seen inaged rats (vanLuijtelaar et al., 1989)
andS100B knockout animals (unpublished observation). Thedystrophic
axonal pattern suggests that these terminal fibersmaybe retracting
froma region inwhich levels of trophic factorare reduced.
3.3. Possible relevance of 5-HTT axon degeneration tosymptoms of
neurodegenerative diseases
Dystrophic degeneration of serotonergic axonsmay contributeto
the development of many of the symptoms of neurodegen-erative
diseases such as mood, motor, sensory, autonomic,cognitive, and
sleepdisorders (see SandykandFisher (1988)). InPD patients,
selective 5-HT reuptake inhibitors (SSRIs) improvebradykinesia
treatedwith L-dopa (Rampello et al., 2002). In ourstudy, six of
nine patientswith neurodegenerative disease hadhistories of
late-life depressive symptoms. Depression isbelieved to be a risk
factor for the onset of Alzheimer's disease(Green et al., 2003;
Chen et al., 1999; Kokmen et al., 1991; Kraland Emery, 1989).
Affective disorders are often comorbid withneurodegenerative
disorders that are associated with demen-tia (Allen and Burns,
1995; Schreinzer et al., 2005; Lauterbachet al., 2004). Affective
symptoms are frequently part of theinitial presentation in
neurodegenerative diseases (Ishiharaand Brayne, 2006; Kessing and
Andersen, 2004), and, in somecases, may be the dominant presenting
symptoms (Ballard etal., 2002). A history of major depression,
without specification
-
Fig. 7 – Dark field montage.
191B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
of episode-related cognitive impairment, appears to be arisk
factor for subsequent onset of dementia (Kessing andAndersen,
2004). This is supported by early work with twins,which found that
depression and psychiatric illness were riskfactors for developing
dementia (Wetherell et al., 1999). Moodmay be improved in depressed
cognitively impaired older pa-tients by treatment with
antidepressants including selectiveserotonin reuptake inhibitors
(SSRI) even in the absence ofimprovement in cognitive or motor
domains (Schrag, 2006;Ishihara and Brayne, 2006; Nebes et al.,
2003). The actions ofserotonin in the development and treatment of
impairmentsseen in neurodegenerative diseases require further
investiga-tion (Gruden et al., 2007; Schmitt et al., 2006; Truchot
et al.,2007).
3.4. Study limitations
Although our findings demonstrated consistent differences inthe
extent and character of 5-HT axonal pathology in the
nineneurodegenerative disease cases compared to the threecontrol
cases analyzed, the small size of the sample precludeda precise
matching of the cases for age and postmorteminterval. Recent
unpublishedwork using 5-HTT immunostain-ing now includes 22 brains
from neurologically typicalnormals, three of which are female, age
range is 32–85 andaverage age is 57.2. No evidence of the frequent
5-HTT-specificaxonal pathology reported here in the brains from
neurode-generative diseases is seen. The occasional
5-HTT-specificaxonal pathology in the temporal cortical regions of
neurolo-gically normal subjects is confirmed (unpublished
observa-tion). Moreover, in contrast to the extensive
neuropathologicalevaluation, the limited clinical characterization
of the subjectsin the study precluded a complete assessment of
psychiatrichistory andmedications in all cases. Nevertheless, these
initialfindings on 5HT fiber pathology in neurodegenerative
diseases
provide a strong rationale for more extensive analyses ofthe
relationship between cognitive or affective parameters and5-HT
pathology in a larger cohort of clinically well-character-ized
subjects.
4. Experimental procedures
4.1. Brains
Brains were obtained from the Harvard Brain Tissue
ResourceCenter at McLean Hospital in Belmont, Massachusetts.
Weobtained 36 tissue blocks for our study from 12 brains, whichmet
diagnostic criteria for the neurodegenerative diseases
ofParkinson's disease (PD, n=3), Frontal Lobe Dementia (FLD,n=3),
and Lewy-body dementia (DLBD, n=3). The patients allmet pre-mortem
and postmortem criteria for the diagnosismade at Harvard Brain
Tissue Resource Center. The durationof the illness or specific
treatments were not available fromthe records for all cases in this
study. A majority of thepatients with a neurodegenerative disorder
also had apsychiatric history of depression (6/9), and in three
cases,the information was unavailable and cannot be
considerednegative for depressive disorder. Neuropathologically
normalbrains (n=3) from donors with no medical or psychiatric
his-tory of illness were studied for comparison (Table 1).
4.2. Selection of regions for study
We used anterior brainstem blocks, which contained thesubstantia
nigra and red nucleus, and the fiber tracts ofthe medial forebrain
bundle, dorsal raphe cortical tract andthe medial lemniscus. The
temporal block we used containedthehippocampus (subiculum, CA
fields anddentate gyrus) andthe entorhinal cortex and associated
whitematter. The frontalblock we used contained the dorsolateral
prefrontal cortex(Brodmann layer 9) with all six cortical layers
and a significantamount of the corpus callosum. We examined all
layers ofentorhinal and prefrontal cortices. In the hippocampus,
welooked at the hilus of the dentate gyrus and the stratum
ra-diatum of CA3 (paying particular note to themossy fibers)
andstratum radiatum of CA1. We also examined the
fibers-of-theperforant path connecting the entorhinal cortex to CA1
andmolecular layer of the dentate gyrus.
4.3. Orientation
We developed a method to produce a full section
dark-fieldmontage (Fig. 7) using a prototype condenser (Leitz
Micro-scopic Company, Germany) for viewing with a 1.6×
objective.Pictures were taken with a Kodak-6.0 million pixel
camera(DCL 760). The montage of the various brain regions
requiredbetween 6–25 images to assemble and was used to
establishprecise locations within the brain section.
4.4. Immunocytochemistry
Immunocytochemical studies were performed as previouslydescribed
(Shiurba et al., 1998) using rabbit polyclonal anti-bodies raised
against serotonin transporter (synthetic peptide
-
192 B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
from the carboxy-terminus (aa 602–622: GTLKERIIKSITPETP-TEIPC)
of the cloned rat serotonin transporter (AB177L,Calbiochem.). This
site-specific antibody has been demon-strated to be highly
selective for the 5-HTT proteins inimmunocytochemistry and western
analysis (Zhou et al.,1996; Qian et al., 1995). Dilutions of
antibody were calculatedto be in the linear range of staining
intensity (1/1000–1/10,000ab5-HTT). Brain tissue used for
immunocytochemical analyseswas immersion-fixed in cold
10%phosphate-buffered formalin(0.15 mol/L), pH 7.4.
Immunocytochemistry was performedby the avidin-biotin complex (ABC)
method using Vectastainkits (Vector Labs, Inc., Burlingame, CA).
Free-floating vibra-tome 50 μm sections were treated for 30 min
with 3%methanolic hydrogen peroxide to block non-specific
endogen-ous peroxidase activity and rinsed in 0.05 M Tris-buffered
(pH7.4) saline (TBS) containing 0.4% Triton X-100. The sectionswere
treated for 30minwith 20% normal rabbit serum (NRS) toreduce
non-specific background staining. Sections were thenincubated in
TBS with 1% NRS and 0.4% Trition X-100 withappropriate dilutions of
primary antisera (AB-1, 1/5000) over-night at room temperature. The
tissue was incubated first inbiotinylated-secondary antibody (1:200
dilution) and subse-quently in preformed ABC (90/d/10 ml avidin and
90;d/mlbiotin). The final reactionwas achievedby treating the
sectionswith 0.02% hydrogen peroxide and DAB (0.5 mg/ml) in 0.1
MTBS, pH 7.4, for 5 min. Vibratome sections were mounted
ongel-coated slides and air-dried following any immunocyto-chemical
or histological procedures. All sections were dehy-drated in a
series of ethanol to xylene and coverslipped withPermount.
Immunocytochemical controls consisted of eitherincubating tissue in
non-immune sera or omitting incubationin primary antisera.
4.5. Morphometric methods
Non-stereologic, morphometric measures of particle numberand
area were performed on each brain (n=3) for statisticalanalysis.
Stereologic methods were not used because ofthe limited number of
brain sections available, and the highdensity of 5-HTT-IR axons.
The sections were viewed with aLeitz orthoplan microscope with
Kohler illumination andphotographed with a Nikon DCL760 digital
camera for theanalysis of 5-HTT-IR axonal number and area.
Morphometricanalysis was performed with the UTHSCSA Image Tool
forwindows (version 3.00) using pictures taken with a 25×PlFluotar
Objective with 8× column magnification. At leastthree pictures were
randomly taken in the prefrontal corticallayers (II–VI) for each
section from all groups. The photographswere untouched except for
conversion into Grayscale beforeperforming a threshold selection
for illumination densitymeasures of 5-HTT-IR labeling (setting of
150 on a 1–255scale). All 5-HTT-IR labeled structures were
automatically se-lected by the computer (10 pixel minimum cut-off)
for count-ing, and measures of area. In the morphometric analysis
ofparticles, normal varicosities that were assumed from theimages
obtained to have an area of
-
193B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
Baumgarten, H.G., Bjorklund, A., 1976. Neurotoxic
indoleaminesand monoamine neurons. Annu. Rev. Pharmacol. Toxicol.
16,101–111.
Bossy-Wetzel, E., Schwarzenbacher, R., Lipton, S.A., 2004.
Molecularpathways to neurodegeneration. Nat. Med. 10, S2–S9
Suppl.
Chen, P., Ganguli, M., Mulsant, B.H., DeKosky, S.T., 1999.The
temporal relationship between depressive symptoms anddementia: a
community-based prospective study. Arch. Gen.Psychiatry 56,
261–266.
Chinaglia, G., Landwehrmeyer, B., Probst, A., Palacios, J.M.,
1993.Serotoninergic terminal transporters are differentially
affectedin Parkinson's disease and progressive supranuclear
palsy:an autoradiographic study with [3H]citalopram.
Neuroscience54, 691–699.
Craven, R.M., Priddle, T.H., Cooper, S.J., Crow, T.J., Esiri,
M.M., 2005.The dorsal raphe nucleus in schizophrenia: a post
mortemstudy of 5-hydroxytryptamine neurones. Neuropathol.Appl.
Neurobiol. 31, 258–269.
Descarries, L., Beaudet, A., Watkins, K.C., 1975. Serotonin
nerveterminals in adult rat neocortex. Brain Res. 100, 563–588.
Dickson, D.W., Schmidt, M.L., Lee, V.M., Zhao, M.L., Yen,
S.H.,Trojanowski, J.Q., 1994. Immunoreactivity profile
ofhippocampal CA2/3 neurites in diffuse Lewy body disease.Acta
Neuropathol. (Berl) 87, 269–276.
Frankfurt, M., Azmitia, E.C., 1983. The effect of
intracerebralinjections of 5,7-dihydroxytryptamine and
6-hydroxydopamineon the serotonin-immunoreactive cell bodies and
fibers in theadult rat hypothalamus. Brain Res. 261, 91–99.
Frankfurt, M., Azmitia, E.C., 1984. Regeneration of
serotonergicfibers in the rat hypothalamus following
unilateral5,7-dihydroxytryptamine injection. Brain Res. 298,
273–282.
Fuxe, K., 1965. Evidence for the existence of monoamine
neuronsin the central nervous system. IV. The distribution
ofmonoamine terminals in the central nervous systemActa Physiol.
Stand. 64 (Suppl. 247), 41–85.
Green, R.C., Cupples, L.A., Kurz, A., Auerbach, S., Go, R.,
Sadovnick,D., Duara, R., Kukull, W.A., Chui, H., Edeki, T.,
Griffith, P.A.,Friedland, R.P., Bachman, D., Farrer, L., 2003.
Depression as arisk factor for Alzheimer disease: the MIRAGE
Study.Arch Neurol. 60, 753–759.
Gruden, M.A., Davidova, T.B., Malisauskas, M., Sewell,
R.D.,Voskresenskaya, N.I.,Wilhelm, K., Elistratova, E.I.,
Sherstnev, V.V.,Morozova-Roche, L.A., 2007.Differential
neuroimmunemarkers tothe onset of Alzheimer's disease
neurodegeneration anddementia: autoantibodies to Abeta((25–35))
oligomers,S100b and neurotransmitters. J. Neuroimmunol. 186,
181–192.
Halliday, G.M., Blumbergs, P.C., Cotton, R.G., Blessing,
W.W.,Geffen, L.B., 1990. Loss of brainstem serotonin- and
substanceP-containing neurons in Parkinson's disease. Brain Res.
510,104–107.
Haan, E.A., Jennings, I.G., Cuello, A.C., Nakata, H., Fujisawa,
H.,Chow, C.W., Kushinsky, R., Brittingham, J., Cotton, R.G.,
1987.Identification of serotonergic neurons in human brain by
amonoclonal antibody binding to all three aromatic amino
acidhydroxylases. Brain Res. 426, 19–27.
Haglund, M., Sjobeck, M., Englund, E., 2006. Locus
ceruleusdegeneration is ubiquitous in Alzheimer's disease:
possibleimplications for diagnosis and treatment. Neuropathology
26,528–532.
Ishihara, L., Brayne, C., 2006. What is the evidence for a
premorbidparkinsonian personality: a systematic review. Mov.
Disord. 21,1066–1072.
Kessing, L.V., Andersen, P.K., 2004. Does the risk of
developingdementia increase with the number of episodes in
patientswith depressive disorder and in patients with bipolar
disorder?J Neurol. Neurosurg. Psychiatry 75, 1662–1666.
Kokmen, E., Beard, C.M., Chandra, V., Offord, K.P., Schoenberg,
B.S.,Ballard, D.J., 1991. Clinical risk factors for Alzheimer's
disease: apopulation-based case-control study. Neurology 41,
1393–1397.
Kral, V.A., Emery, O.B., 1989. Long-term follow-up of
depressivepseudodementia of the aged. Can. J. Psychiatry 34,
445–446.
Kung, L., Force, M., Chute, D.J., Roberts, R.C.,
1998.Immunocytochemical localization of tyrosine hydroxylase inthe
human striatum: a postmortem ultrastructural study.J. Comp. Neurol.
390, 52–62.
Langlais, P.J., Thal, L., Hansen, L., Galasko, D., Alford, M.,
Masliah,E., 1993. Neurotransmitters in basal ganglia and cortex
ofAlzheimer's disease with and without Lewy bodies. Neurology43,
1927–1934.
Lauterbach, E.C., Freeman, A., Vogel, R.L., 2004. Differential
DSM-IIIpsychiatric disorder prevalence profiles in dystonia
andParkinson's disease. J. Neuropsychiatry Clin. Neurosci.
16,29–36.
Liu, Y., Nakamura, S., 2006. Stress-induced plasticity
ofmonoamine axons. Front. Biosci. 11, 1794–1801.
Marksteiner, J., Walch, T., Bodner, T., Gurka, P., Donnemiller,
E.,2003. Fluoxetine in Alzheimer's disease with severe
obsessivecompulsive symptoms and a low density of
serotonintransporter sites. Pharmacopsychiatry 36, 207–209.
Menza, M.A., Palermo, B., DiPaola, R., Sage, J.I., Ricketts,
M.H., 1999.Depression and anxiety in Parkinson's disease: possible
effectof genetic variation in the serotonin transporter. J.
Geriatr.Psychiatry Neurol. 12, 49–52.
Molliver, M.E., 1987. Serotonergic neuronal systems: what
theiranatomic organization tells us about function. J.
Clin.Psychopharmacol. 7 (6 Suppl), 3S–23S.
Molliver, D.C., Molliver, M.E., 1990. Anatomic evidence for
aneurotoxic effect of (+/−)-fenfluramine upon
serotonergicprojections in the rat. Brain Res. 511, 165–168.
Nebes, R.D., Pollock, B.G., Houck, P.R., Butters, M.A., Mulsant,
B.H.,Zmuda, M.D., Reynolds III, C.F., 2003. Persistence of
cognitiveimpairment in geriatric patients following
antidepressanttreatment: a randomized, double-blind clinical trial
withnortriptyline and paroxetine. J. Psychiatr. Res. 37,
99–108.
Ohara, K., Kondo, N., Ohara, K., 1998. Changes of monoamines
inpost-mortem brains from patients with diffuse Lewy bodydisease.
Prog. Neuro-psychopharmacol. Biol. Psychiatry 22,311–317.
O'Hearn, E., Battaglia, G., De Souza, E.B., Kuhar, M.J.,
Molliver, M.,1988. Methylenedioxy amphetamine (MDA)
andmethylenedioxymethamphetamine (MDMA) cause selectiveablation of
serotonergic axon terminals in forebrain:immunocytochemical
evidence for neurotoxicity.J. Neurosci. 8, 2788–2803.
Perry, E.K., Irving, D., Kerwin, J.M., McKeith, I.G., Thompson,
P.,Collerton, D., Fairbairn, A.F., Ince, P.G., Morris, C.M., Cheng,
A.V.,et al., 1993. Cholinergic transmitter and neurotrophic
activitiesin Lewy body dementia: similarity to Parkinson's and
distinctionfrom Alzheimer disease. Alzheimer Dis. Assoc. Disord.
7,69–79.
Qian, Y., Melikian, H.E., Rye, D.B., Levey, A.I., Blakely, R.D.,
1995.Identification and characterization of
antidepressant-sensitiveserotonin transporter proteins using
site-specific antibodies.J. Neurosci. 15 (2), 1261–1274.
Rampello, L., Chiechio, S., Raffaele, R., Vecchio, I.,
Nicoletti, F.,2002. The SSRI, citalopram, improves bradykinesia in
patientswith Parkinson's disease treated with L-dopa.Clin.
Neuropharmacol. 25 (1), 21–24.
Sandyk, R., Fisher, H., 1988. Serotonin in involuntary
movementdisorders. Int. J. Neurosci. 42, 185–208.
Schmitt, J.A., Wingen, M., Ramaekers, J.G., Evers, E.A., Riedel,
W.J.,2006. Serotonin and human cognitive performance.Curr. Pharm.
Des. 12, 2473–2486.
Schreinzer, D., Ballaban, T., Brannath, W., Lang, T., Hilger,
E.,Fasching, P., Fischer, P., 2005. Components of
behavioralpathology in dementia. Int. J. Geriatr. Psychiatry 20,
137–145.
Schrag, A., 2006. Quality of life and depression in
Parkinson'sdisease. J. Neurol. Sci. 248, 151–157.
-
194 B R A I N R E S E A R C H 1 2 1 7 ( 2 0 0 8 ) 1 8 5 – 1 9
4
Shiurba, R.A., Spooner, E.T., Ishiguro, K., Takahashi, M.,
Yoshida,R., Wheelock, T.R., Imahori, K., Cataldo, A.M., Nixon,
R.A., 1998.Immunocytochemistry of formalin-fixed human brain
tissues:microwave irradiation of free-floating sections. Brain
Res.Brain Res. Protoc. 2, 109–119.
Sparks, D.L., Markesbery, W.R., 1991. Altered serotonergic
andcholinergic synaptic markers in Pick's disease. Arch. Neurol.48,
796–799.
Steinbusch, H.W., 1981. Distribution of
serotonin-immunoreactivityin the central nervous system of the
rat-cell bodies andterminals. Neuroscience 6, 557–618.
Stout, S.S., Somerset, W.I., Miller, A., Musselman, D.L.,
2003.Paroxetine use in medically ill patients.
Psychopharmacol.Bull. 37 (Suppl 1), 108–122.
Szabo, Z., Kao, P.F., Mathews, W.B., Ravert, H.T., Musachio,
J.L.,Scheffel, U., Dannals, R.F., 1996. Positron emission
tomographyof 5-HT reuptake sites in the human brain with C-11
McN5652extraction of characteristic images by artificial neural
networkanalysis. Behav. Brain Res. 73, 221–224.
Truchot, L., Costes, S.N., Zimmer, L., Laurent, B., Le Bars,
D.,Thomas-Anterion, C., Croisile, B., Mercier, B., Hermier,
M.,Vighetto, A., Krolak-Salmon, P., 2007. Up-regulation
ofhippocampal serotonin metabolism in mild cognitiveimpairment.
Neurology 69, 1012–1017.
Underwood, M.D., Khaibulina, A.A., Ellis, S.P., Moran, A., Rice,
P.M.,Mann, J.J., Arango, V., 1999. Morphometry of the dorsal
raphenucleus serotonergic neurons in suicide victims.Biol.
Psychiatry 46, 473–483.
van Luijtelaar, M.G., Steinbusch, H.W., Tonnaer, J.A.,
1989.Similarities between aberrant serotonergic fibers in the
agedand 5,7-DHT denervated young adult rat brain. Exp. Brain
Res.78, 81–89.
Varnas, K., Halldin, C., Hall, H., 2004. Autoradiographic
distributionof serotonin transporters and receptor subtypes in
humanbrain. Hum. Brain Mapp. 22, 246–260.
Wetherell, J.L., Gatz, M., Johansson, B., Pedersen, N.L., 1999.
Historyof depression and other psychiatric illness as risk factors
forAlzheimer disease in a twin sample. Alzheimer Dis. Assoc.Disord.
13, 47–52.
Wiklund, L., Bjorklund, A., 1980. Mechanisms of regrowth in
thebulbospinal serotonin system following
5,6-dihydroxytryptamineinduced axotomy. II. Fluorescence
histochemical observations.Brain Res. 191, 109–127.
Wilson,M.A., Molliver, M.E., 1994.Microglial response to
degenerationof serotonergic axon terminals. Glia 11, 18–34.
Zhou, F.C., Azmitia, E.C., 1986. Induced homotypic sprouting
ofserotonergic fibers in hippocampus. II. An
immunocytochemistrystudy. Brain Res. 373, 337–348.
Zhou, F.C., Bledsoe, S., Lumeng, L., Li, T.K., 1994.
Reducedserotonergic immunoreactive fibers in the forebrain
ofalcohol-preferring rats. Alcohol Clin. Exp. Res. 18,571–579.
Zhou, F.C., Xu, Y., Bledsoe, S., Lin, R., Kelley, M.R., 1996.
Serotonintransporter antibodies: production, characterization,
andlocalization in the brain. Brain Res. Mol. Brain Res.
43,267–278.
Dystrophic serotonergic axons in neurodegenerative
diseasesIntroductionResults5-HTT patterns in neuropathologically
normal brainsAscending projectionsTerminal distribution
5-HTT patterns in the brain in neurodegenerative diseasesDiffuse
Lewy-body dementiaParkinson's diseaseFrontal lobe
dementiaMorphometric analysis of pathologic changes
DiscussionSerotonin system and neurodegenerationComparison of
axonal pathologyPossible relevance of 5-HTT axon degeneration to
symptoms of neurodegenerative diseasesStudy limitations
Experimental proceduresBrainsSelection of regions for
studyOrientationImmunocytochemistryMorphometric methodsStatistical
analysis
AcknowledgmentsReferences