Embed Size (px)
Citation preview

Drought enhances folivory by shifting foliar metabolomes inQuercus ilex trees
Albert Rivas-Ubach1,2, Albert Gargallo-Garriga1,2,3, Jordi Sardans1,2, Michal Oravec4, Laia Mateu-Castell1,2,
M�ıriam P�erez-Trujillo3, Teodor Parella3, Rom�a Ogaya1,2, Otmar Urban4 and Josep Pe~nuelas1,2
1CSIC, Global Ecology Unit CREAF-CEAB-CSIC-UAB, Cerdanyola del Vall�es, 08913 Catalonia, Spain; 2CREAF, Cerdanyola del Vall�es, 08913 Catalonia, Spain; 3Servei de Ressonancia
Magnetica Nuclear, Faculty of Sciences and Biosciences, Universitat Aut�onoma de Barcelona, Bellaterra, 08193 Barcelona, Catalonia, Spain; 4Global Change Research Centre, Academy of
Sciences of the Czech Republic, Bĕlidla 4a, CZ-603 00 Brno, Czech Republic
Author for correspondence:Albert Rivas-Ubach
Tel: +34 935 813 355Email: [email protected]
Received: 8 October 2013
Accepted: 17 December 2013
New Phytologist (2014) 202: 874–885doi: 10.1111/nph.12687
Key words: drought, ecology,ecometabolomics, folivory, metabolomics,stoichiometry.
Summary
� At the molecular level, folivory activity on plants has mainly been related to the foliar con-
centrations of nitrogen (N) and/or particular metabolites.� We studied the responses of different nutrients and the whole metabolome of Quercus
ilex to seasonal changes and to moderate field experimental conditions of drought, and how
this drought may affect folivory activity, using stoichiometric and metabolomic techniques.� Foliar potassium (K) concentrations increased in summer and consequently led to higher
foliar K : phosphorus (P) and lower carbon (C) : K and N : K ratios. Foliar N : P ratios were not
lowest in spring as expected by the growth rate hypothesis. Trees exposed to moderate
drought presented higher concentrations of total sugars and phenolics and these trees also
experienced more severe folivory attack.� The foliar increases in K, sugars and antioxidant concentrations in summer, the driest Medi-
terranean season, indicated enhanced osmoprotection under natural drought conditions.
Trees under moderate drought also presented higher concentrations of sugars and phenolics;
a plant response to avoid water loss. These shifts in metabolism produced an indirect relation-
ship between increased drought and folivory activity.
Introduction
Climate change is expected to lead to longer dry spells, higherevaporative demand and more intense droughts in the comingdecades in several regions of the world, including the Mediterra-nean basin (IPCC, 2007). Increased drought has been demon-strated to enhance tree mortality in forests (Martinez-Vilalta &Pinol, 2002; Allen et al., 2010; Galiano et al., 2012) and mayhave multiple effects on ecosystem biodiversity and ecosystemstructure and function (Anderegg et al., 2013; Pe~nuelas et al.,2013a). Furthermore, both xeric and mesic forests may behydraulically vulnerable to drought (Choat et al., 2012).
Several studies showed that herbivores commonly increasetheir activity in response to increased concentrations of solublenitrogen (N) in foliage (Larsson, 1989; Larsson & Bjorkman,1993; Rouault et al., 2006). Plant water status also plays animportant role in the resistance of trees against herbivorous attack(Rouault et al., 2006). Drought may affect foliar nutritional qual-ity and indirectly stimulate insect folivory (White, 1984; Rouaultet al., 2006). Also, plants are able to respond to folivory attack byproducing chemical defenses such as alkaloids, terpenes andphenolics (Bennett & Wallsgrove, 1994; Kessler & Baldwin,2001; Ali & Agrawal, 2012), and these chemical defenses seem tobe more concentrated in those populations of plants historically
more frequently in contact with the insects (Raffa et al., 2013).Several other studies have also demonstrated that plant primarymetabolism is shifted in response to tissue predation or infection(Ehness et al., 1997; Widarto et al., 2006; Sardans et al., 2013a).
Drought is also a potential driver of changes in the elementalcarbon (C) : N : phosphorus (P) : potassium (K) stoichiometriesof different plant organs and ecosystems (Rivas-Ubach et al.,2012; Sardans et al., 2012a,b) with consequent effects on ecologi-cal processes and ultimately the structure and function of ecosys-tems (Elser et al., 1996; Sterner & Elser, 2002; Sardans et al.,2011a, 2012b; Pe~nuelas et al., 2013b). K has been demonstratedto play an important role under low water availability conditionsto avoid water losses, especially in summer drought (Ingram &Bartels, 1996; Sardans et al., 2012a; Wang et al., 2013), throughincreases in its foliar concentrations (Sardans et al., 2013b). Thestudy of foliar K concentrations under natural or experimentaldrought conditions thus requires more attention in the context ofecological stoichiometry (Sardans et al., 2012a). The large varia-tion in C : N : P : K biomass stoichiometries in plants, both intime and in space (latitude and altitude), may be a significant fac-tor, among others, in the selection of foliage with high nutritionalcontent by herbivores (Gusewell & Koerselman, 2002; Lindrothet al., 2002; Oleksyn et al., 2002; Raubenheimer & Simpson,2003; Sardans et al., 2012b). Additionally, the rates of folivory
874 New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research

have been observed to be directly related to the relativeconcentrations of foliar structural compounds such as ligninrather than to foliar N concentrations, with high relative concen-trations of these structural compounds usually being avoided(Choong et al., 1992; Williams et al., 1998). Thus, C : N ratioshave been shown to play a key role in folivory (Ji et al., 2011)through the effects of structural compounds or N concentrations.Furthermore, changes in plant stoichiometry may influence thecoevolution of insects by shifting the composition andconcentration of defensive chemical compounds of plants(Raubenheimer & Simpson, 2003), as most elements, such as C,N and P, generally do not act in isolation but as components ofmolecular compounds (Pe~nuelas & Sardans, 2009a) such as lig-nin and cellulose in lignified structures or various compoundsthat defend against herbivorous attack (Bennett & Wallsgrove,1994; Kessler & Baldwin, 2001).
Most studies examining the relationships between plant metab-olites and herbivory predation at the molecular level have gener-ally focused on the identification of single compounds or familiesof metabolites (Sardans et al., 2013a). The application of the newmetabolomic techniques in ecology and plant physiology (eco-metabolomics) allows the study of the complete metabolome ofan organism, the total set of metabolites in an organism at a spe-cific moment (Fiehn, 2002), and its response to abiotic and bioticenvironmental shifts (Pe~nuelas & Sardans, 2009b; Sardans et al.,2011b; Sardans & Pe~nuelas, 2012; Pe~nuelas et al., 2013a;Rivas-Ubach et al., 2013). Recent ecometabolomic studies havereported metabolomic changes in plants exposed to a biotic stresssuch as herbivorous predation (Jansen et al., 2009; Leiss et al.,2009) or an abiotic stress such as drought (Charlton et al., 2008;Lugan et al., 2009; Sardans et al., 2011b; Rivas-Ubach et al., 2012).The link between shifts in foliar stoichiometries and shifts in foliarmetabolomes throughout the seasons and under experimentaldrought in the field has been recently demonstrated (Rivas-Ubachet al., 2012), so the nutritional quality of food for herbivores maythus also change under stress conditions (Rouault et al., 2006).
The relationship between plant stoichiometry and metabolismand herbivorous attack under drought conditions, however, isstill unclear and warrants further study (Raubenheimer & Simp-son, 2003). The rates of folivory can thus be influenced directlyby food quality and indirectly by plant water status in what couldbe a cascade effect of drought. As a broader and better under-standing of the metabolomic shifts in plants grown under singleand interacting stress factors such as drought and herbivory isgreatly needed, we performed this study where we hypothesizedthat shifts in both foliar elemental stoichiometry and metabolo-mics produced by drought would influence folivory activity. Byaffecting the quality of the food available to herbivores, drought-induced shifts in foliar chemical composition may lead tolong-term cascade effects that could alter trophic webs.
Once per season, we sampled leaves of Quercus ilex trees from amature forest in Catalonia (northeastern Iberian Peninsula)exposed to conditions of moderate experimental drought similarto those projected for the coming decades. The metabolomes andelemental compositions of all samples were analyzed. The rates offolivory were also determined for each sampled tree. Our goal
was to understand: (1) the stoichiometric responses of Q. ilex, themost dominant tree in the forests of the Mediterranean basin, toseasonal changes and to experimental conditions of moderatedrought; (2) the foliar metabolomic responses of Q. ilex to differ-ent seasons and moderate drought; (3) how the foliar stoichiome-tric and metabolic shifts of Q. ilex may affect the selection of foodby herbivores, and (4) how Q. ilex is able to shift its metabolometo respond to herbivore attack. For this purpose, the Q. ilex forestwas used to test our hypothesis that drought, simulated in experi-mental plots, could alter folivorous activity by altering the foliarmetabolome. Our results could thus help to predict the impact ofclimate change on trophic webs (Pe~nuelas & Sardans, 2009a).
Materials and Methods
Study site
This study was performed in a natural Quercus ilex L. forest inthe Prades Mountains in southern Catalonia (see Ogaya &Pe~nuelas, 2007 for details; 41°21′N, 1°2′E). All sampled plotsfaced south-southeast on a 25% slope at 930 m above sea level.The climate of the region is mesic-Mediterranean with a markedsummer drought for 3 months (Supporting Information Fig.S1a). The vegetation consists of a forest dominated by Q. ilex,followed by Phillyrea latyfolia and Arbutus unedo, among others.
Experimental design
Four plots (159 10 m) of mature Q. ilex forest were established15 m apart in March 1999 (Ogaya et al., 2003). Two randomlyassigned plots received a drought treatment, and the other twoserved as control plots. The drought treatment consisted of cover-ing c. 30% of the soil surface with 19 14 m PVC strips orienteddown the slope and 0.5–0.8 m above the soil surface to preventirrigation by rainwater. PVC strips do not produce an increase insoil temperature (Fig. S1d). A ditch 0.8–1 m in depth was dugalong the entire top edge of the treatment plots to intercept ups-lope runoff. All water intercepted by the strips was channeled tothe bottom edge of the drought plots. Soil moisture, air humid-ity, precipitation and air and soil temperatures were monitoredevery 30 min. The drought treatment resulted in an averageannual reduction of 18% in relative soil moisture (Fig. S1b,c; seeBarbeta et al., 2013 for details). This reduction in soil moisturehas increased tree mortality and reduced growth during the last10 yr (Fig. S2).
Sampling of leaves
Five adult Q. ilex individuals 25–50 cm in diameter were ran-domly selected from each plot as study cases (total n = 20). Leaveswere sampled once each season: in February (winter), May(spring), August (summer) and November (autumn). A smallbranch exposed to the sun was removed from each tree with apole, and a sample of the youngest well-developed leaves was fro-zen in liquid nitrogen for the stoichiometric and metabolomicanalyses. The remaining leaves were stored in bags at 6–8°C for
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 875

determination of water content and for photographic analysis ofthe extent of consumption by herbivores.
Calculation of the proportion of leaves consumed byherbivores
Fifteen randomly selected leaves stored at 6–8°C from each treewere placed on a flat white surface and photographed with aNikon D80 camera with a Nikkor AF-S 18-135/3.5-5.6 G DXlens to calculate the percentage of folivory. The area of the leavesconsumed was calculated using Adobe Photoshop CS2 (AdobeSystems Incorporated, San Jose, CA, USA) by following themethod of Pe~nuelas et al. (2013c). The assigned value of con-sumed area for each tree was the mean of the 15 leaves analyzed(see Fig. S3 for details). The values of folivory for each tree werethen standardized by the total foliar biomass of Q. ilex in its plot.All values were thereafter transformed for normality (arcsin(square root(percentage))). Additionally, the fresh leaves werealso used for calculation of fresh weight, dry weight, foliar watercontent, foliar size and leaf mass per unit area (LMA).
Foliar processing for elemental and metabolomic analyses
The processing of the leaves is described in detail in Rivas-Ubachet al. (2013). Briefly, leaves frozen in liquid nitrogen were lyophi-lized and stored in plastic cans at �20°C. Samples were groundwith a ball mill at 1600 rpm for 6 min (Mikrodismembrator-U;B. Braun Biotech International, Melsungen, Germany), produc-ing a fine powder that was then stored at �80°C until the extrac-tion of the metabolites.
Elemental analysis
For the C and N analyses, 1.4 mg of the powder from each samplewas analyzed. The C andN concentrations were determined by ele-mental analysis using combustion coupled to gas chromatographywith a CHNS-O Elemental Analyser (EuroVector, Milan, Italy).
Macroelements (P and K) were extracted by acid digestion in amicrowave reaction system under high pressure and temperature(Sardans et al., 2010). Briefly, 250 mg of leaf powder was placedin a Teflon tube with 5 ml of nitric acid and 2 ml of H2O2.A MARSXpress microwave reaction system (CEM, Mattheus,NC, USA) was used for the acid digestions (see the ‘Chemicalanalyses’ section of Methods S1 for details). The digested mate-rial was transferred to 50-ml flasks and resuspended in Milli-Qwater (EMD Millipore, Darmstadt, Germany) to a final volumeof 50 ml. After digestion, the P and K concentrations were deter-mined by optic emission spectrometry with inductively coupledplasma (ICP-OES) (Optima 4300; Perkin-Elmer Corporation,Norwalk, CT, USA; see Methods S1 for details).
Extraction of metabolites for analysis by nuclear magneticresonance (NMR)
The extraction of foliar metabolites for NMR analysis isdescribed in detail in Rivas-Ubach et al. (2013). First, two sets of
50-ml centrifuge tubes were labeled: set A for the extraction ofmetabolites and set B for lyophilization. A set of crystal jars forthe organic fractions were also labeled. The tubes of set Areceived 200 mg of powdered leaf material of each sample. Sixmilliliters of water/methanol (1 : 1) and 6 ml of chloroform wereadded to each tube. The samples were vortexed for 15 s and thensonicated for 2 min at room temperature. All tubes were centri-fuged at 1100 g for 15 min. Four milliliters of each fraction(aqueous and organic) was collected independently; aqueous frac-tions were transferred to the centrifuge tubes of set B, and organicfractions were transferred to the crystal jars. This procedure wasrepeated twice for two extractions of the same sample. The aque-ous samples, previously resuspended in water to reduce the pro-portion of methanol (< 15% methanol), were lyophilized, andthen 4 ml of water was added to each tube, which was vortexedand centrifuged at 23 000 g for 3 min. The samples were frozenat �80°C and lyophilized again. The organic fractions weretransferred to round-bottomed evaporation flasks and dried in arotary vacuum evaporator. Finally, 1 ml of KH2PO4-NaOD-buffered D2O + 0.01% trimethylsilyl propionic acid sodium salt(TSP; pH 6.0) was added to the dried aqueous fractions,and 1 ml of chloroform-D + 0.01% tetramethylsilyl (TMS) wasadded to the dried organic fractions. TSP and TMS were used asinternal standards. All solutions were resuspended with a micro-pipette, transferred to 2-ml centrifuge tubes and centrifuged at23 000 g for 3 min. The supernatants were transferred to NMRsample tubes.
Extraction of metabolites for liquid chromatography–massspectrometry (LC-MS) analysis
The extraction of metabolites followed the protocol of t’Kindtet al. (2008) with minor modifications. Two sets of Eppendorftubes were labeled: set A for extractions and set B for the extractsfrom set A.
The tubes of set A received 100 mg of sample powder fromeach sample, and then 1 ml of MeOH/H2O (80 : 20) was addedto each tube. The tubes were vortexed for 15 min, sonicated for5 min at room temperature and then centrifuged at 23 000 g for5 min. After centrifugation, 0.6 ml of the supernatant from eachtube was transferred to the corresponding Eppendorf tubes ofset B. This procedure was repeated for two extractions of thesame sample. The tubes of set B were centrifuged at 23 000 gfor 5 min. The supernatants were collected using crystal syrin-ges, filtered through 0.22-lm pore microfilters and transferredto a labeled set of high-performance liquid chromatography(HPLC) vials. The vials were stored at �80°C until the LC-MSanalysis.
LC-MS analysis
LC-MS chromatograms were obtained using a Dionex Ultimate3000 HPLC system (Thermo Fisher Scientific/Dionex RSLC,Waltham, MA, USA) coupled to an LTQ Orbitrap XL high-resolution mass spectrometer (Thermo Fisher Scientific)equipped with a heated electrospray ionization (HESI II) source.
New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist876

Chromatography was performed on a reversed-phase C18 Hyper-sil gold column (1509 2.1 mm, 3 lm particle size; ThermoFisher Scientific) at 30°C. The mobile phases consisted of aceto-nitrile (A) and water (0.1% acetic acid) (B). Both mobile phaseswere filtered and degassed for 10 min in an ultrasonic bath beforeuse. At a flow rate of 0.3 ml min�1, the elution gradient began at10% A (90% B) and was maintained for 5 min, and then waschanged to 10% B (90% A) for the next 20 min. The initial pro-portions (10% A; 90% B) were gradually recovered over the next5 min, and the column was then washed and stabilized for 5 minbefore injecting the next sample. The injection volume of thesamples was 5 ll. All samples were injected twice, once with theHESI operating in negative ionization mode (�H) and once inpositive ionization mode (+H). The Orbitrap mass spectrometerwas operated in Fourier transform mass spectrometry(FTMS) full-scan mode with a mass range of 50–1000m/z andhigh-resolution (60 000). The resolution and sensitivity of thespectrometer were monitored by injecting a standard of caffeineafter every 10 samples, and the resolution was further monitoredwith lock masses (phthalates). Blank samples were also analyzedduring the sequence.
NMR analysis
NMR experiments were performed on a Bruker AVANCE 600spectrometer working at a magnetic field of 14.1 T (1H and 13CNMR frequencies of 600.13 and 150.13MHz, respectively) andequipped with an automatic sample changer, a multinuclear tri-ple resonance TBI probe and a temperature control unit (BrukerBiospin, Rheinstetten, Germany). The temperature into the pro-behead was previously calibrated and maintained constant for allthe experiments at 298.0 K; for this purpose a temperatureequilibration delay of 2 min is left for each sample before theshimming process. All NMR sample handling, automation,acquisition and processing was controlled using TOPSPIN 3.1 soft-ware (Bruker Biospin). Spectra were referenced to TSP (1H and13C at d 0.00 ppm) in the case of polar samples and to the signalof the residual solvent, CHCl3 (
1H at d 7.26 ppm and 13C at d77.16 ppm), in the case of nonpolar samples.
1H NMR fingerprinting
All extract samples were analyzed by high-resolution one-dimensional (1D) 1H NMR spectroscopy following the condi-tions described in Rivas-Ubach et al. (2013) and using standardpulse-acquisition 1D 1H-NMR experiments. In the case of thewater/methanol extract samples, proton spectra were acquiredwith suppression of the residual water resonance. The water reso-nance signal was presaturated at a power level of 55 dB, corre-sponding to an effective field of 30 Hz during a relaxation delayof 2 s. Each experiment was acquired as a set of 32 000 datapoints, over a spectral width of 16 ppm, as the sum of 128 tran-sients and with an acquisition time of 1.7 s. The resulting freeinduction decays (FIDs) were Fourier-transformed and the spec-tra obtained were phased and baseline-corrected. The FIDs ofpolar samples were multiplied by an exponential apodization
function equivalent to 0.2-Hz line broadening before the Fouriertransform. The experimental time was c. 8 min per sample.
NMR metabolite identification Standard 2D NMR experi-ments (1H–1H correlated spectroscopy (COSY), 1H–1H total cor-relation spectroscopy (TOCSY), 1H–13C heteronuclear singlequantum correlation (HSQC) and 1H–13C heteronuclear multiplebond correlation (HMBC)), and 1D selective 1H TOCSYexperiments were performed on representative polar andnonpolar samples for the identification of metabolites. For thewater–methanol extract samples, 2D experiments were carriedout with standard presaturation of the residual water peak duringthe relaxation delay. Experiments were acquired using standardBruker pulse sequences and routine conditions (see Methods S2for details). The procedure followed was that described byRivas-Ubach et al. (2013). All elucidated metabolites were furtherconfirmed using data reported in the literature (see Tables S1 andS2 for references).
Processing of LC-MS chromatograms
The raw data files from the spectrometer were processed usingMZMINE 2.10 (Pluskal et al., 2010). Chromatograms were base-line-corrected, deconvoluted, aligned and filtered before thenumerical database was exported in CSV format (Table S3 fordetails). Metabolites were assigned by exact mass and retentiontime from the measurements of the standards in the MS andMSn modes of the spectrometer (Table S4 for details). Differentassigned variables corresponding to the same molecular com-pounds were summed. The numerical values of the variablesextracted from the LC-MC chromatograms correspond to theabsolute peak areas of the chromatograms detected by the spec-trometer. The area is directly proportional to the concentrationof the variable, so we use the term ‘concentration’ in this articlewhen referring to changes in the amount of metabolites betweenthe studied factors (control vs drought or between seasons).
NMR bucketing
The processing of 1H NMR spectra is detailed in Rivas-Ubachet al. (2013). Briefly, before the exportation of the 1H NMRnumerical databases, all spectra were phased, baseline-correctedand referenced to the resonance of the internal standard (TSP forpolar and TMS for nonpolar samples) at d 0.00 ppm with TOP-
SPIN 3.1. A variable-size bucketing was thus applied to all 1HNMR spectra using AMIX software (Bruker Biospin), scaling thebuckets relative to the internal standard (TMS or TSP). Theoutput was a data set containing the integral values for eachassigned 1H NMR spectral peak in the described pattern. Thebuckets corresponding to the same molecular compound weresummed.
Statistical analyses
To test for differences between seasons and drought treatments infoliar elemental stoichiometry and metabolome, the LC-MS and
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 877
NMR metabolomic fingerprints of the Q. ilex leaves were sub-jected to PERMANOVA analysis (Anderson et al., 2008) usingthe Euclidean distance, with season (spring, summer, autumnand winter) and treatment (control and drought) as fixed factorsand folivory as a covariable. The number of permutations wasset at 999. The PERMANOVA analysis was conducted withPERMANOVA+ for the PRIMER v.6 software (Anderson et al.,2008). One-way ANOVAs were performed for each individualstoichiometric or metabolite variable (Tables S5, S6). Statisticallysignificant results of those ANOVAs are indicated by asterisks inFigs 1 and 2.
Additionally, to understand how the foliar stoichiometry andmetabolome of Q. ilex shifted with the factors studied (seasonand climatic treatment), the foliar stoichiometric and metabolo-mic fingerprints were also subjected to principal componentanalysis (PCA) and partial least squares discriminant analysis(PLS-DA). The seasonal PCA and PLS-DA included the finger-prints of all seasons combined (Figs 1, S4). Trees in summerseason present the accumulated foliar signals of folivory in theMediterranean basin; for this reason, in order to study the effectsof drought on folivory, the fingerprints from summer leaves wereadditionally submitted to separate PCAs and PLS-DA (Figs 2,S5). The PCAs and PLS-DA were performed using the pca andpls.da functions, respectively, of the mixOmics package of R (RDevelopment Core Team, 2011). The score coordinates of thevariables of the PCAs were subjected to one-way ANOVAs
to detect statistical differences among groups (see supportinginformation of Rivas-Ubach et al., 2013). A Kolmogorov–Smirnov (KS) test was performed on each variable to test fornormality. All assigned and identified metabolites were normallydistributed, and any unidentified metabolomic variable that was
(a)
(b)
(c)
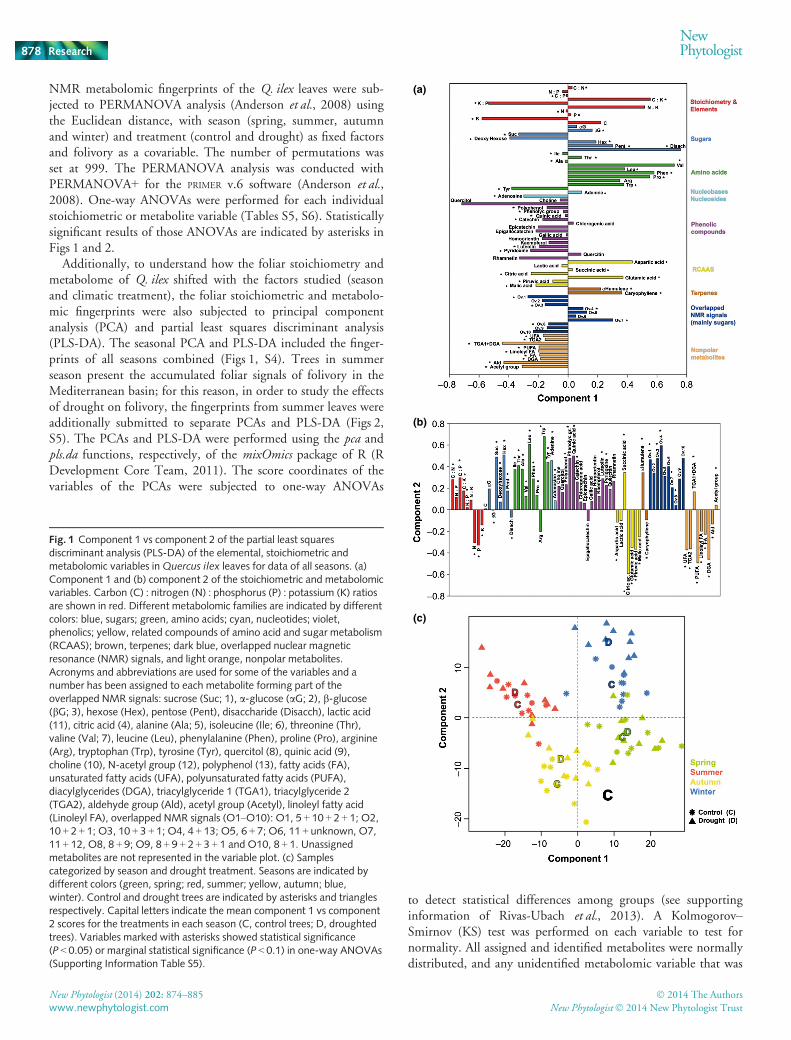
Fig. 1 Component 1 vs component 2 of the partial least squaresdiscriminant analysis (PLS-DA) of the elemental, stoichiometric andmetabolomic variables inQuercus ilex leaves for data of all seasons. (a)Component 1 and (b) component 2 of the stoichiometric and metabolomicvariables. Carbon (C) : nitrogen (N) : phosphorus (P) : potassium (K) ratiosare shown in red. Different metabolomic families are indicated by differentcolors: blue, sugars; green, amino acids; cyan, nucleotides; violet,phenolics; yellow, related compounds of amino acid and sugar metabolism(RCAAS); brown, terpenes; dark blue, overlapped nuclear magneticresonance (NMR) signals, and light orange, nonpolar metabolites.Acronyms and abbreviations are used for some of the variables and anumber has been assigned to each metabolite forming part of theoverlapped NMR signals: sucrose (Suc; 1), a-glucose (aG; 2), b-glucose(bG; 3), hexose (Hex), pentose (Pent), disaccharide (Disacch), lactic acid(11), citric acid (4), alanine (Ala; 5), isoleucine (Ile; 6), threonine (Thr),valine (Val; 7), leucine (Leu), phenylalanine (Phen), proline (Pro), arginine(Arg), tryptophan (Trp), tyrosine (Tyr), quercitol (8), quinic acid (9),choline (10), N-acetyl group (12), polyphenol (13), fatty acids (FA),unsaturated fatty acids (UFA), polyunsaturated fatty acids (PUFA),diacylglycerides (DGA), triacylglyceride 1 (TGA1), triacylglyceride 2(TGA2), aldehyde group (Ald), acetyl group (Acetyl), linoleyl fatty acid(Linoleyl FA), overlapped NMR signals (O1–O10): O1, 5 + 10 + 2 + 1; O2,10 + 2 + 1; O3, 10 + 3 + 1; O4, 4 + 13; O5, 6 + 7; O6, 11 + unknown, O7,11 + 12, O8, 8 + 9; O9, 8 + 9 + 2 + 3 + 1 and O10, 8 + 1. Unassignedmetabolites are not represented in the variable plot. (c) Samplescategorized by season and drought treatment. Seasons are indicated bydifferent colors (green, spring; red, summer; yellow, autumn; blue,winter). Control and drought trees are indicated by asterisks and trianglesrespectively. Capital letters indicate the mean component 1 vs component2 scores for the treatments in each season (C, control trees; D, droughtedtrees). Variables marked with asterisks showed statistical significance(P < 0.05) or marginal statistical significance (P < 0.1) in one-way ANOVAs(Supporting Information Table S5).
New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist878
not normally distributed was removed from the data set. The KStests were performed with the ks.test function of the tuncogofpackage of R (R Development Core Team, 2011).
Folivory begins in spring and early summer, and trees do notproduce new leaves in summer, so the sampled leaves had accu-mulated signs of folivory by the middle of the summer. An addi-tional PCA was thus conducted with summer data excluding theproportions of herbivorous consumption (Fig. S6). The PC1
scores of this PCA, including the entire elemental, stoichiometricand metabolomic variation of the case of trees in summer, wereplotted against the degree of folivory (Fig. 3). Folivory wasanalyzed with a general linear model (GLM) as a function of theclimatic treatment and the metabolomic variation (PC1 scores).Another GLM analyzed the total foliar sugar and phenolic con-centrations as functions of climatic treatment and season. STATIS-TICA v8.0 (StatSoft, Tulsa, OK, USA) was used for the one-wayANOVAs and the post hoc tests of the score coordinates of thePCAs. GLM analyses were performed with the lme function ofthe nlme package of R (R Development Core Team, 2011). Allstatistical analyses were performed at the individual level.
Results
The PERMANOVA analysis performed with all elemental, stoi-chiometric and metabolomic data (assigned and unassignedmetabolites) indicated that leaves in different seasons, underdrought treatment and with different degrees of folivory had dif-ferent foliar chemistries and metabolisms (folivory: pseudo-F = 2.4832; P < 0.001; season: pseudo-F = 2.4749; P < 0.001;and treatment: pseudo-F = 3.1031; P < 0.001).
Elemental, stoichiometric and metabolomic shifts acrossseasons and experimental drought treatments
The PCA conducted with all seasons, including all the elemental,stoichiometric and metabolomic data, indicated that more than50% of the total variance was already gathered by the first fourPCs (PC1 = 15.2%, PC2 = 14.7%, PC3 = 14.1% andPC4 = 12.7%; Fig. S4). Differences between seasons wereexplained by PC1 (P < 0.001), PC3 (P < 0.05) and PC4
(a)
(b)
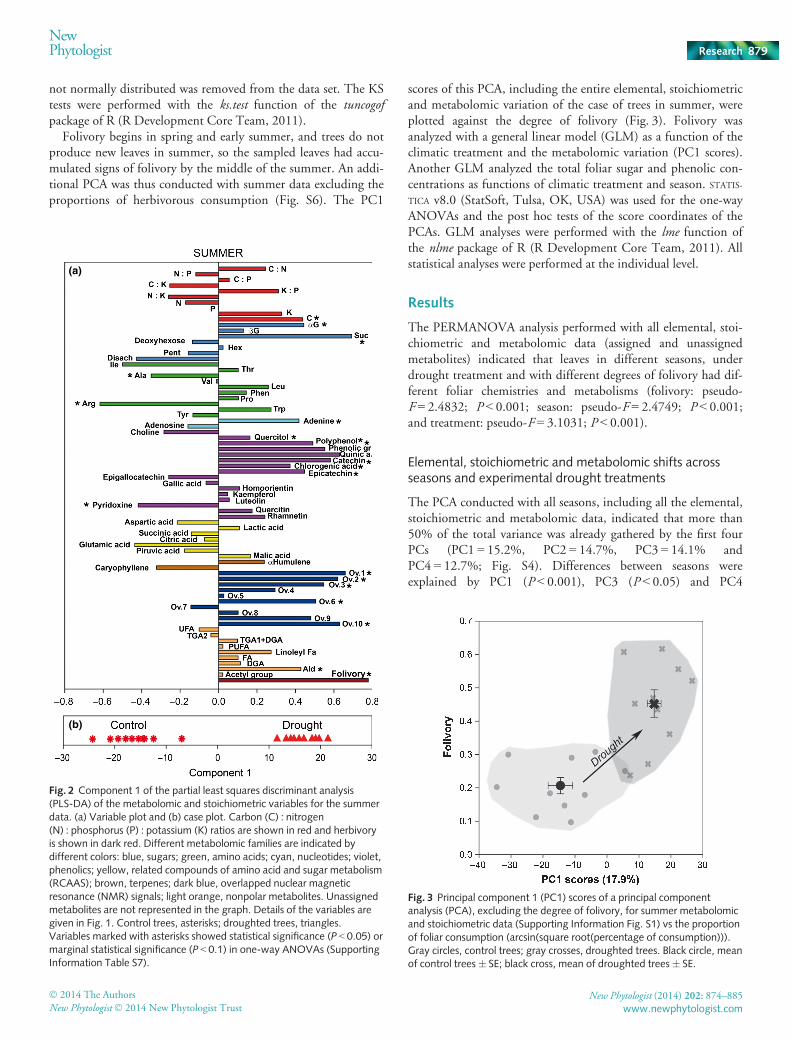
Fig. 2 Component 1 of the partial least squares discriminant analysis(PLS-DA) of the metabolomic and stoichiometric variables for the summerdata. (a) Variable plot and (b) case plot. Carbon (C) : nitrogen(N) : phosphorus (P) : potassium (K) ratios are shown in red and herbivoryis shown in dark red. Different metabolomic families are indicated bydifferent colors: blue, sugars; green, amino acids; cyan, nucleotides; violet,phenolics; yellow, related compounds of amino acid and sugar metabolism(RCAAS); brown, terpenes; dark blue, overlapped nuclear magneticresonance (NMR) signals; light orange, nonpolar metabolites. Unassignedmetabolites are not represented in the graph. Details of the variables aregiven in Fig. 1. Control trees, asterisks; droughted trees, triangles.Variables marked with asterisks showed statistical significance (P < 0.05) ormarginal statistical significance (P < 0.1) in one-way ANOVAs (SupportingInformation Table S7).
Fig. 3 Principal component 1 (PC1) scores of a principal componentanalysis (PCA), excluding the degree of folivory, for summer metabolomicand stoichiometric data (Supporting Information Fig. S1) vs the proportionof foliar consumption (arcsin(square root(percentage of consumption))).Gray circles, control trees; gray crosses, droughted trees. Black circle, meanof control trees � SE; black cross, mean of droughted trees � SE.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 879
(P < 0.001). Post hoc analysis of the score coordinates of the dif-ferent case trees showed that the stoichiometries and metabolo-mes of summer leaves differed significantly from those of leavesin other seasons on the PC1 axis. Droughted trees differed fromcontrol trees in PC1 (P < 0.001), PC2 (P < 0.05) and PC4(P < 0.001). PLS-DA with all seasons including all the elemental,stoichiometric data showed clear separation between seasons(Fig. 1).
The C, N, P and K foliar concentrations and the ratios C : N,C : P, C : K, N : K and K : P, but not N : P, changed with the sea-sons (Fig. 1, Table S5). The highest K : P and K : N concentra-tion ratios occurred in summer, coinciding with the highest foliarK concentrations. Foliar N and P concentrations were highest inautumn, thus resulting in the lowest C : N and C : P ratios (seeTable S5 and Sardans et al., 2013b for more details).
Most assigned and identified metabolites in the leaves ofQ. ilex shifted across the seasons (Fig. 1, Table S5). The mainchanges were between spring and summer, two of the most criti-cal seasons in the Mediterranean basin, because spring is theperiod of growth and summer is the driest season. Summer leavesgenerally had higher concentrations of polyphenolics and sucrose,whereas spring leaves had higher concentrations of amino acids,some related compounds of amino acid and sugar metabolism(RCAAS) and some sugars such as pentoses and disaccharides,products directly related to growth. The effects of experimentaldrought were also detected on the seasonal PLS-DA and PCAplots (Figs 1, S4, Tables S6, S7). In the case plot for both analy-ses, PLS-DA and PCA, droughted trees tended to be distributedin the same direction as the summer trees; summer is the driestseason in Mediterranean climates (Fig. S1a).
Total concentrations of amino acids, sugars andpolyphenolics
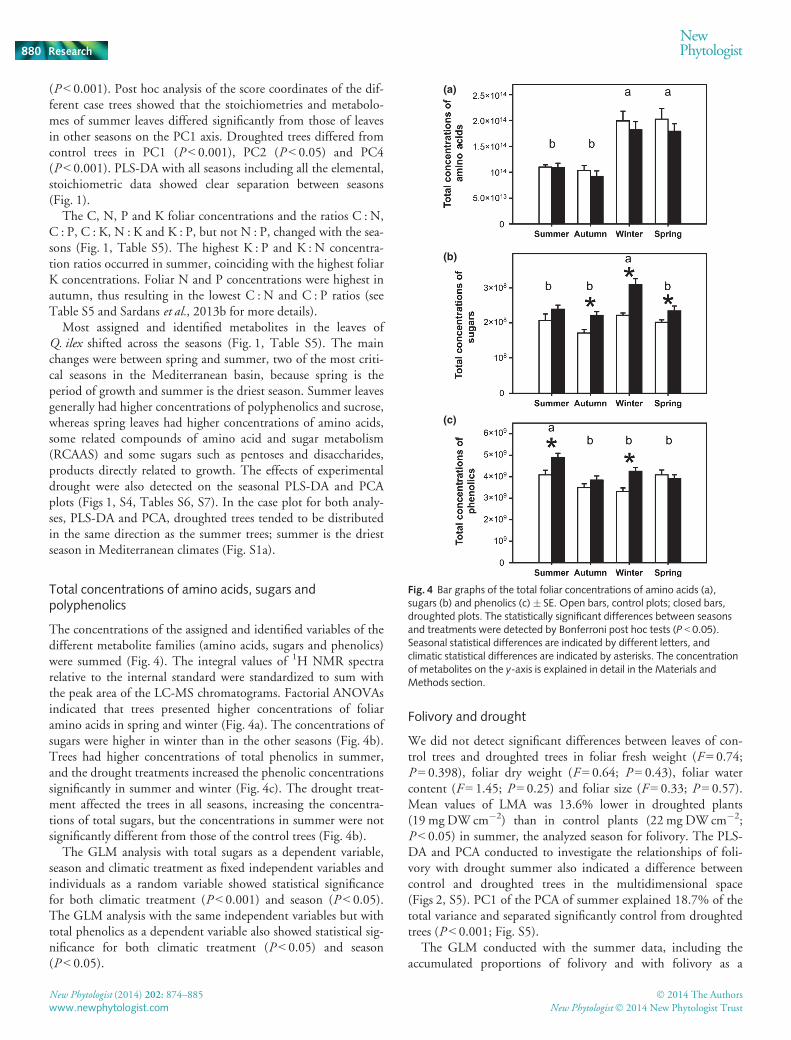
The concentrations of the assigned and identified variables of thedifferent metabolite families (amino acids, sugars and phenolics)were summed (Fig. 4). The integral values of 1H NMR spectrarelative to the internal standard were standardized to sum withthe peak area of the LC-MS chromatograms. Factorial ANOVAsindicated that trees presented higher concentrations of foliaramino acids in spring and winter (Fig. 4a). The concentrations ofsugars were higher in winter than in the other seasons (Fig. 4b).Trees had higher concentrations of total phenolics in summer,and the drought treatments increased the phenolic concentrationssignificantly in summer and winter (Fig. 4c). The drought treat-ment affected the trees in all seasons, increasing the concentra-tions of total sugars, but the concentrations in summer were notsignificantly different from those of the control trees (Fig. 4b).
The GLM analysis with total sugars as a dependent variable,season and climatic treatment as fixed independent variables andindividuals as a random variable showed statistical significancefor both climatic treatment (P < 0.001) and season (P < 0.05).The GLM analysis with the same independent variables but withtotal phenolics as a dependent variable also showed statistical sig-nificance for both climatic treatment (P < 0.05) and season(P < 0.05).
Folivory and drought
We did not detect significant differences between leaves of con-trol trees and droughted trees in foliar fresh weight (F = 0.74;P = 0.398), foliar dry weight (F = 0.64; P = 0.43), foliar watercontent (F = 1.45; P = 0.25) and foliar size (F = 0.33; P = 0.57).Mean values of LMA was 13.6% lower in droughted plants(19 mg DW cm�2) than in control plants (22 mg DW cm�2;P < 0.05) in summer, the analyzed season for folivory. The PLS-DA and PCA conducted to investigate the relationships of foli-vory with drought summer also indicated a difference betweencontrol and droughted trees in the multidimensional space(Figs 2, S5). PC1 of the PCA of summer explained 18.7% of thetotal variance and separated significantly control from droughtedtrees (P < 0.001; Fig. S5).
The GLM conducted with the summer data, including theaccumulated proportions of folivory and with folivory as a
(a)
(b)
(c)
Fig. 4 Bar graphs of the total foliar concentrations of amino acids (a),sugars (b) and phenolics (c) � SE. Open bars, control plots; closed bars,droughted plots. The statistically significant differences between seasonsand treatments were detected by Bonferroni post hoc tests (P < 0.05).Seasonal statistical differences are indicated by different letters, andclimatic statistical differences are indicated by asterisks. The concentrationof metabolites on the y-axis is explained in detail in the Materials andMethods section.
New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist880

dependent variable and climatic treatment and PC1 scores of theadditional PCA (without the folivory variable; Fig. S6) as inde-pendent variables (P < 0.001; R2 = 0.60), indicated significanteffects of the drought treatment (P < 0.05) and no significanteffect of the PC1 scores (P = 0.53). These results imply that theexistence of a significant relationship between folivory andthe PC1 score coordinates of the PCA is mainly attributable tothe fixed effect of drought (Fig. 3).
Discussion
Seasonality and drought
The foliar stoichiometry and metabolome of Q. ilex varied withseason (PERMANOVA P < 0.001; Figs 1, S4). Seasonal PLS-DAand PCA identified significant stoichiometric and metabolomicshifts between summer and the other seasons, although the differ-ences between seasons were not as marked as those of an earlierstudy with the Mediterranean shrub Erica multiflora (Rivas-Ubach et al., 2012). Trees in spring, the Mediterranean growingseason, had high concentrations of metabolites directly related togrowth such as amino acids, some RCAAS and some sugars suchas pentoses and disaccharides, in accordance with the earlier study(Rivas-Ubach et al., 2012; Fig. 1), even though the total concen-tration of amino acids did not differ from the levels in winter(Fig. 4a), the coldest season in the Mediterranean basin. InQ. ilex, foliar N : P ratios did not present the lowest values inspring as expected from the growth rate hypothesis (Elser et al.,1996; Rivas-Ubach et al., 2012; Figs 1, S4, Table S5), which pro-poses that organisms with high rates of growth require high con-centrations of P (low N : P ratios) to meet the demands ofprotein synthesis (Elser et al., 2003). Our results are neverthelessin accordance with those of other studies in plants. The highfoliar concentrations of amino acids in winter trees might beattributable to accumulation under cold conditions. It has beenobserved that some amino acids such as proline and glycine areable to buffer the NADP+ : NADPH ratio in plants (Xu et al.,2013) as a consequence of low photosynthetic activity in winterto provide reducing agents for the generation of ATP in mito-chondria (Hare & Cress, 1997; Xu et al., 2013). However, Mat-zek & Vitousek (2009) found no significant relationshipsbetween the N : P ratio and increased growth in 14 species ofpines, concluding that terrestrial plants must invest in fundamen-tal functions other than growth, such as storage and defense, thatoften require different investments in N and P (Herms & Matt-son, 1992; Pe~nuelas & Estiarte, 1998).
As observed in the multivariate ordination analyses, the con-centrations of the various sugar compounds detected were distrib-uted differentially among the seasons (Figs 1, S4), but droughtedtrees in winter had the highest concentrations of total sugars(Fig. 4b), which may also be related to their accumulation andallocation from other organs (e.g. wood and lignotubers) inresponse to a cold environment to leaves or further preparationfor the growth season (Grimaud et al., 2013; Xu et al., 2013).Quercus ilex is an evergreen tree with slow growth; it presentslarge lignotubers, swollen woody structures at the base of the
stem (James, 1984) that are able to store essential nutrients andmetabolites such as carbohydrates to ensure rapid growth aftersevere stress (Canadell & Zedler, 1995). Our stoichiometric andmetabolomic data support the idea that trees, with large woodystructures such as trunks, lignotubers, and/or extensive rootnetworks, are able to buffer the metabolomic changes in foliarontogeny by storing large amounts of nutrients, allowing the allo-cation of essential resources to photosynthetic organs and thusleaf growth and resistance to severe environmental disturbances(Canadell & Zedler, 1995; Landhausser & Lieffers, 2002;Anderegg & Callaway, 2012; Galiano et al., 2012).
The largest stoichiometric shift between seasons appeared tobe related to the responses to drought conditions (Figs 1, S4,Table S5). Foliar concentrations of K were higher in trees in sum-mer, the driest season. The resulting higher foliar K : P and lowerN : K and C : K concentration ratios (Sardans et al., 2013b) werethus in agreement with the findings of other Mediterranean sea-sonal studies (Sardans et al., 2012a, 2013b). K participates in thewater economy of plants (Babita et al., 2010) through osmoticcontrol (Babita et al., 2010; Laus et al., 2011) and improves thefunctioning of foliar stomata (Khosravifar et al., 2008). Theseresults demonstrate the important role of K in the ecology ofterrestrial plants (Cakmak, 2005; Sardans et al., 2012a, 2013b).Moreover, these shifts in K concentrations in summer leaveswere accompanied by higher concentrations of sucrose, whichcan also act as an osmolyte that, together with K, can help to pre-vent the loss of water through osmotic control (Ingram & Bartels,1996).
As seen in previous studies, a decrease in growth and anincrease in tree mortality are among the main consequences ofthis prolonged moderate drought (Barbeta et al., 2013; Fig. S2).Our seasonal PLS-DA and PCA with metabolomic and stoichi-ometric data also showed interesting differences between the treesin the drought plots and those in the control plots (Figs 1, S4;PERMANOVA P < 0.001). Droughted trees tended to have thesame pattern as summer trees on the Component 2 of case plotof PLS-DA (Fig. 1) and PC1 and PC2 of PCA (Fig. S4), inaccordance with a previous study (Rivas-Ubach et al., 2012), thusindicating a foliar elemental-metabolomic response to droughtindependent of plant ontogeny. Summer trees and droughtedtrees in all seasons tended to have higher foliar concentrations offlavonoids (Figs 1, 4c, S4), the largest group of naturally occur-ring polyphenols (Strack et al., 1994), but the flavonoid composi-tion differed between summer and the drought treatment.Summer trees had higher concentrations of quinic acid, catechinand luteolin, among other polyphenolics, whereas droughtedtrees had higher concentrations of catechin, quercitol, homoori-entin and quercetin (Figs 1, S4, Table S6). These differences cor-related with the differential effects of drought treatment in thedifferent seasons (Fig. 4c). Many biological functions have beenascribed to flavonoids, especially their role as antioxidants (Burda& Oleszek, 2001; Lee et al., 2003). Their antioxidant activity ismainly a result of their role as electron donors (RiceEvans et al.,1997) and their ability to alter the kinetics of peroxidation (Aroraet al., 2000). As expected in a water-limited Mediterraneanecosystem, oxidative stress in plants tends to increase under
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 881

conditions of drought (Price et al., 1989; Dat et al., 2000;Munn�e-Bosch & Pe~nuelas, 2004; Pe~nuelas et al., 2004). Querci-tol has been associated with the avoidance of osmotic stress byreducing the osmotic potential during drought in Quercus species(Passarinho et al., 2006; Spiess et al., 2012). Our study found nosignificant differences (P = 0.12) between choline concentrationsin droughted and control trees but did identify a tendency towardhigher concentrations in droughted plants (Table S6). Choline isalso involved in osmotic protection (McNeil et al., 2001).Drought will probably have a significant impact on the metabol-omes of Mediterranean plants, because dry periods will becomemore frequent and intense in the coming decades, as predicted byprojections of climate change (IPCC, 2007).
Droughted trees also presented higher concentrations of totalsoluble sugars in their leaves (Fig. 4c), supporting the premisethat sugars act as osmolytes to prevent the loss of water by regulat-ing water potential and draw water into leaves during periods ofdrought (Ingram & Bartels, 1996). Control trees in summer didnot have significantly lower foliar concentrations of sugars relativeto droughted trees (Fig. 4b), even though droughted trees had atendency toward higher concentrations, presenting significantlyhigher concentrations of sucrose and a-glucose. The small differ-ences in foliar sugar concentrations between control and drough-ted leaves may be mainly attributable to the naturally drysummers of the Mediterranean basin that increased the foliarsugar concentrations in both control and droughted trees (Fig. 4).
Folivory
The highest herbivorous activity by insects occurs mainly inspring and early summer in the Mediterranean basin (Powell &Logan, 2005; Bonal et al., 2010), and usually the leaves of treeshave accumulated most signs of folivory by the middle ofthe summer. Interestingly, folivorous activity was higher in thedroughted trees in summer (Figs 2, 3). We did not detect any sig-nificant difference between control trees and droughted trees infoliar fresh weight, foliar dry weight, foliar water content or foliarsize that could explain the higher folivory activity in summer.The small LMA differences between controls and droughted treeswere attributable to differences in thickness and not to the foliardensity as found in past studies (Ogaya & Pe~nuelas, 2006), whichshould not influence consumption through folivory.
GLM analyses for the summer season indicated that the rela-tionship between folivory and PC1 score coordinates of the addi-tional PCA for foliar stoichiometry and metabolomics (Fig. S6)was mainly attributable to the fixed effect of drought, becausefolivory was not correlated with PC1 within treatments (control,drought; P > 0.05; Fig. 3). Our results thus suggest that the stoi-chiometric and metabolomic shifts were mainly caused by theexperimental drought, which subsequently indirectly increasedthe degree of folivory.
Our stoichiometric results identified no significant relation-ships between foliar N concentrations and foliar C : N ratios andthe degree of folivory, as reported in other studies (Larsson,1989; Larsson & Bjorkman, 1993; Rouault et al., 2006), indicat-ing that, in our study, folivory was not influenced only by foliar
N concentrations (Choong et al., 1992; Williams et al., 1998). Asdiscussed, droughted trees tended to have higher concentrationsof sugars and polyphenols (flavonoids) than did the controls(Fig. 4). Flavonoids such as quercetin have been shown to act asphagostimulants in herbivorous insects (Diaz Napal et al., 2010;Kosonen et al., 2012). Despite the scarcity of supporting evi-dence, our results also indicated a higher folivorous activity indroughted trees, supporting the premise of the high palatabilityof some flavonoids that are not directly related to defense but actas antioxidants (Diaz Napal et al., 2010; Kosonen et al., 2012).Also, droughted leaves had higher concentrations of sugars(Fig. 4), especially sucrose and a-glucose (Fig. 2), which couldthus provide a source of rapid energy for herbivores. The higherconcentrations of sugars and some flavonoids found in the foliageof droughted Q. ilex trees appeared to be related to the increase inherbivorous activity (Figs 3, S7).
Not all the assigned polyphenols, however, provide only anti-oxidant protection in plants. The role of polyphenolics as achemical defense against predation by herbivores has been widelyreported (Berg, 2003; Lokvam & Kursar, 2005; Treutter, 2006;Kosonen et al., 2012; Rani & Pratyusha, 2013). Moreover, Koso-nen et al. (2012) reported that some polyphenolics could be toxicto specialist herbivores but could increase the palatability ofplants for generalist herbivores and suggested that climate changemay reduce the damage caused by specialist herbivores andincrease that caused by generalists. Phenolic acids could thuspotentially be used for pest management (Rani & Pratyusha,2013). It has been reported that even mammals can be affectedby the polyphenols of plants. For example, Berg (2003) reportedthat elevated concentrations of catechin negatively affected theconsumption of plants by collared lemmings. In our study, nearlyall the assigned polyphenols were associated with drought.
Fig. 5 Summary of the relationship between foliar metabolic shiftsproduced by drought and folivory activity. As drought increases (a) there isa shift in the foliar metabolome ofQuercus ilex, mainly by increasing theconcentrations of antioxidants and sugars (b). This increase in antioxidantsand sugars stimulates folivory activity on droughted trees relative tocontrol trees (c). The increase in folivory inQ. ilex leaves may later causean increase in the synthesis of defense compounds (d).
New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist882

Most of the assigned flavonoids have antioxidant activity, butdroughted trees also had a significantly higher foliar concentra-tion of catechin (Figs 2, S5), thus supporting the idea that plant-induced defenses are not only produced by drought but also by ahigher degree of herbivorous attack as many previous studies havesuggested (Raubenheimer & Simpson, 2003; Fig. 5).
Our results suggest that seasonal drought influences the stoichi-ometry and metabolome of plants and that experimental droughtindirectly affects folivorous activity by shifting the metabolism ofplants, making them more attractive for folivores (Fig. 3). Themore severe and frequent droughts predicted for the Mediterra-nean basin (IPCC, 2007) may thus have an indirect impact ontrophic webs by changing the amount of herbivory through foliarstoichiometric and metabolomic shifts (Fig. 3). This impact maylead to more intensive and extensive outbreaks of pests and to pos-sible changes in the distribution of food between specialist andgeneralist herbivores (Kosonen et al., 2012).
Conclusions and final remarks
Foliar N : P ratios of Q. ilex did not significantly change betweenseasons and were not lowest in spring, the growing season, asexpected from the growth rate hypothesis. Foliar K concentra-tions, however, did change, demonstrating the importance of Kin natural summer droughts. The lack of significant variation inN and P concentrations in Q. ilex may be attributable to the buf-fering effect of lignotubers in this woody species.
Moderate experimental drought increased the concentrationsof sugars and polyphenolic compounds with antioxidant functionin the leaves of Q. ilex. Our results showed a relationship betweenthese metabolomic shifts and foliar consumption by herbivores.Herbivores may also induce plant responses, increasing the con-centrations of defensive polyphenolic compounds.
Our data indicated that summer and droughted trees couldavoid or respond to a reduction in water availability by differentmeans, although both had higher foliar concentrations of flavo-noids. Summer trees tended to use more K as an osmolyte thandid droughted trees.
The present study is an example of how coupling stoichiometricwith metabolomic techniques is useful for identifying the molecu-lar responses of plants to changing environmental conditions suchas drought, for understanding the mechanisms and functions thatunderlie the responses of plants to these new conditions and forinterpreting the implications of drought for trophic webs.
Acknowledgements
The authors thank Gemma Montalban and Sara F�erez for theirfield and laboratory support. A.R.-U. appreciates the financialsupport of a research fellowship (JAE) from the CSIC. Thisresearch was supported by the Spanish Government (projectsCGL2010 17172/BOS, Consolider-Ingenio Montes CSD2008-00040 and CTQ2012-32436), the Catalan Government (projectSGR 2009-458) and the European Research Council (Synergygrant SyG-2013-610028, IMBALANCE-P). M.O. and O.U.were supported by the grant project M200871201 (AS CR).
References
Ali JG, Agrawal AA. 2012. Specialist versus generalist insect herbivores and plant
defense. Trends in Plant Science 17: 293–302.Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N,
Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH et al.2010. A global overview of drought and heat-induced tree mortality reveals
emerging climate change risks for forests. Forest Ecology and Management259: 660–684.
Anderegg WRL, Callaway ES. 2012. Infestation and hydraulic consequences of
induced carbon starvation. Plant Physiology 159: 1866–1874.Anderegg WRL, Kane JM, Anderegg LDL. 2013. Consequences of widespread
tree mortality triggered by drought and temperature stress. Nature ClimateChange 3: 30–36.
Anderson MJ, Gorley RN, Clarke KR. 2008. Permanova+ for PRIMER: guide tosoftware and statistical methods. Plymouth, UK: Primer-E.
Arora A, Byrem TM, Nair MG, Strasburg GM. 2000.Modulation of liposomal
membrane fluidity by flavonoids and isoflavonoids. Archives of Biochemistry andBiophysics 373: 102–109.
Babita M, Maheswari M, Rao LM, Shanker AK, Rao DG. 2010.Osmotic
adjustment, drought tolerance and yield in castor (Ricinus communis L.)hybrids. Environmental and Experimental Botany 69: 243–249.
Barbeta A, Ogaya R, Pe~nuelas J. 2013. Dampening effects of long-term
experimental drought on growth and mortality rates of a Holm oak forest.
Global change biology 19: 3133–3144.Bennett RN, Wallsgrove RM. 1994. Secondary metabolites in plant
defense-mechanisms. New Phytologist 127: 617–633.Berg TB. 2003. Catechin content and consumption ratio of the collared
lemming. Oecologia 135: 242–249.Bonal R, Munoz A, Maria Espelta J. 2010.Mismatch between the timing of
oviposition and the seasonal optimum. The stochastic phenology of
Mediterranean acorn weevils.. Ecological Entomology 35: 270–278.Burda S, Oleszek W. 2001. Antioxidant and antiradical activities of flavonoids.
Journal of Agricultural and Food Chemistry 49: 2774–2779.Cakmak I. 2005. The role of potassium in alleviating detrimental effects of
abiotic stresses in plants. Journal of Plant Nutrition and Soil Science-ZeitschriftFur Pflanzenernahrung Und Bodenkunde 168: 521–530.
Canadell J, Zedler P. 1995. Underground structures of woody plants in
Mediterranean ecosystems of Australia, California and Chile. In: Ecology andbiogeography of Mediterranean ecosystems in Chile, California and Australia. New
York, NY, USA: Ecological Studies, Springer-Verlag, 108: 170–210.Charlton AJ, Donarski JA, Harrison M, Jones SA, Godward J, Oehlschlager S,
Arques JL, Ambrose M, Chinoy C, Mullineaux PM et al. 2008. Responses ofthe pea (Pisum sativum L.) leaf metabolome to drought stress assessed by
nuclear magnetic resonance spectroscopy.Metabolomics 4: 312–327.Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ,
Feild TS, Gleason SM, Hacke UG et al. 2012. Global convergence in the
vulnerability of forests to drought. Nature 491: 752–755.Choong MF, Lucas PW, Ong JSY, Pereira B, Tan HTW, Turner IM. 1992.
Leaf fracture-toughness and sclerophylly – their correlations and ecologicalimplications. New Phytologist 121: 597–610.
Dat J, Vandenabeele S, Vranova E, Van Montagu M, Inze D, Van Breusegem F.
2000. Dual action of the active oxygen species during plant stress responses.
Cellular and Molecular Life Sciences 57: 779–795.Diaz Napal GN, Defago MT, Valladares GR, Palacios SM. 2010. Response of
Epilachna paenulata to two flavonoids, pinocembrin and quercetin, in a
comparative study. Journal of chemical ecology 36: 898–904.Ehness R, Ecker M, Godt DE, Roitsch T. 1997. Glucose and stress
independently regulate source and sink metabolism and defense mechanisms
via signal transduction pathways involving protein phosphorylation. Plant Cell9: 1825–1841.
Elser JJ, Acharya K, Kyle M, Cotner J, Makino W, Markow T, Watts T, Hobbie
S, Fagan W, Schade J et al. 2003. Growth rate-stoichiometry couplings in
diverse biota. Ecology Letters 6: 936–943.Elser JJ, Dobberfuhl DR, MacKay NA, Schampel JH. 1996.Organism size, life
history, and N : P stoichiometry. BioScience 46: 674–684.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 883

Fiehn O. 2002.Metabolomics – the link between genotypes and phenotypes.
Plant Molecular Biology 48: 155–171.Galiano L, Martinez-Vilalta J, Sabate S, Lloret F. 2012. Determinants of
drought effects on crown condition and their relationship with depletion of
carbon reserves in a Mediterranean holm oak forest. Tree Physiology 32:478–489.
Grimaud F, Renaut J, Dumont E, Sergeant K, Lucau-Danila A, Blervacq A-S,
Sellier H, Bahrman N, Lejeune-Henaut I, Delbreil B et al. 2013. Exploringchloroplastic changes related to chilling and freezing tolerance during cold
acclimation of pea (Pisum sativum L.). Journal of Proteomics 80: 145–159.Gusewell S, Koerselman M. 2002. Variation in nitrogen and phosphorus
concentrations of wetland plants. Perspectives in Plant Ecology, Evolution andSystematics 5: 37–61.
Hare PD, Cress WA. 1997.Metabolic implications of stress-induced proline
accumulation in plants. Plant Growth Regulation 21: 79–102.Herms DA, Mattson WJ. 1992. The dilemma of plants – to grow or defend.
Quarterly Review of Biology 67: 283–335.Ingram J, Bartels D. 1996. The molecular basis of dehydration tolerance in
plants. Annual Review of Plant Physiology and Plant Molecular Biology 47:377–403.
IPCC. 2007. Core Writing Team, Pachauri RK, Reisinger A. Fourth AssessmentReport: Climate change 2007 (AR4) Contribution of Working Groups I, II and IIIto the Fourth Assessment Report of the Intergovernmental Panel on ClimateChange. Geneva, Switzerland: IPCC.
James S. 1984. Lignotubers and burls – their structure, function and ecological
significance in Mediterranean ecosystems. Botanical Review 50: 225–266.Jansen JJ, Allwood JW, Marsden-Edwards E, van der Putten WH, Goodacre R,
van Dam NM. 2009.Metabolomic analysis of the interaction between plants
and herbivores.Metabolomics 5: 150–161.Ji L-Z, An L-L, Wang X-W. 2011. Growth responses of gypsy moth larvae to
elevated CO2: the influence of methods of insect rearing. Insect Science 18:409–418.
Kessler A, Baldwin IT. 2001. Defensive function of herbivore-induced plant
volatile emissions in nature. Science 291: 2141–2144.Khosravifar S, Yarnia M, BenamMBK, Moghbeli AHH. 2008. Effect of
potassium on drought tolerance in potato cv. Agria. Journal of Food, Agriculture& Environment 6: 236–241.
t’Kindt R, De Veylder L, Storme M, Deforce D, Van Bocxlaer J. 2008. LC-MS
metabolic profiling of Arabidopsis thaliana plant leaves and cell cultures:optimization of pre-LC-MS procedure parameters. Journal of ChromatographyB-Analytical Technologies in the Biomedical and Life Sciences 871: 37–43.
Kosonen M, Keski-Saari S, Ruuhola T, Constabel CP, Julkunen-Tiitto R. 2012.
Effects of overproduction of condensed ttannins and elevated temperature on
chemical and ecological traits of genetically modified hybrid aspens (Populustremula9 P tremuloides). Journal of chemical ecology 38: 1235–1246.
Landhausser SM, Lieffers VJ. 2002. Leaf area renewal, root retention and
carbohydrate reserves in a clonal tree species following above-ground
disturbance. Journal of Ecology 90: 658–665.Larsson S. 1989. Stressful times for the plant stress – insect performance
hypothesis. Oikos 56: 277–283.Larsson S, Bjorkman C. 1993. Performance of chewing and phloem-feeding
insects on stressed trees. Scandinavian Journal of Forest Research 8: 550–559.Laus MN, Soccio M, Trono D, Liberatore MT, Pastore D. 2011. Activation of
the plant mitochondrial potassium channel by free fatty acids and acyl-CoA
esters: a possible defence mechanism in the response to hyperosmotic stress.
Journal of Experimental Botany 62: 141–154.Lee KG, Shibamoto T, Takeoka GR, Lee SE, Kim JH, Park BS. 2003.
Inhibitory effects of plant-derived flavonoids and phenolic acids on
malonaldehyde formation from ethyl arachidonate. Journal of Agricultural andFood Chemistry 51: 7203–7207.
Leiss KA, Choi YH, Abdel-Farid IB, Verpoorte R, Klinkhamer PGL. 2009.
NMR metabolomics of thrips (Frankliniella occidentalis) resistance in Seneciohybrids. Journal of chemical ecology 35: 219–229.
Lindroth RL, Osier TL, Barnhill HRH, Wood SA. 2002. Effects of genotype
and nutrient availability on phytochemistry of trembling aspen (PopulustremuloidesMichx.) during leaf senescence. Biochemical Systematics and Ecology30: 297–307.
Lokvam J, Kursar TA. 2005. Divergence in structure and activity of phenolic
defenses in young leaves of two co-occurring Inga species. Journal of ChemicalEcology 31: 2563–2580.
Lugan R, Niogret M-F, Kervazo L, Larher FR, Kopka J, Bouchereau A. 2009.
Metabolome and water status phenotyping of Arabidopsis under abiotic stress
cues reveals new insight into ESK1 function. Plant, Cell & Environment 32:95–108.
Martinez-Vilalta J, Pinol J. 2002. Drought-induced mortality and hydraulic
architecture in pine populations of the NE Iberian Peninsula. Forest Ecology andManagement 161: 247–256.
Matzek V, Vitousek PM. 2009. N : P stoichiometry and protein : RNA ratios in
vascular plants: an evaluation of the growth-rate hypothesis. Ecology Letters 12:765–771.
McNeil SD, Nuccio ML, Ziemak MJ, Hanson AD. 2001. Enhanced synthesis of
choline and glycine betaine in transgenic tobacco plants that overexpress
phosphoethanolamine N-methyltransferase. Proceedings of the NationalAcademy of Sciences, USA 98: 10001–10005.
Munn�e-Bosch S, Pe~nuelas J. 2004. Drought-induced oxidative stress in
strawberry tree (Arbutus unedo L.) growing in Mediterranean field conditions.
Plant Science 166: 1105–1110.Ogaya R, Pe~nuelas J. 2006. Contrasting foliar responses to drought inQuercusilex and Phillyrea latifolia. Biologia Plantarum 50: 373–382.
Ogaya R, Pe~nuelas J. 2007. Tree growth, mortality, and above-ground biomass
accumulation in a holm oak forest under a five-year experimental field drought.
Plant Ecology 189: 291–299.Ogaya R, Pe~nuelas J, Martinez-Vilalta J, MangironM. 2003. Effect of drought on
diameter increment ofQuercus ilex, Phillyrea latifolia, and Arbutus unedo in aholm oak forest of NE Spain. Forest Ecology andManagement 180: 175–184.
Oleksyn J, Reich PB, Zytkowiak R, Karolewski P, Tjoelker MG. 2002. Needle
nutrients in geographically diverse Pinus sylvestris L. populations. Annals ofForest Science 59: 1–18.
Passarinho JAP, Lamosa P, Baeta JP, Santos H, Ricardo CPP. 2006. Annual
changes in the concentration of minerals and organic compounds of Quercussuber leaves. Physiologia Plantarum 127: 100–110.
Pe~nuelas J, Estiarte M. 1998. Can elevated CO2 affect secondary metabolism and
ecosystem function? Trends in Ecology & Evolution 13: 20–24.Pe~nuelas J, Munne-Bosch S, Llusia J, Filella I. 2004. Leaf reflectance and photo-
and antioxidant protection in field-grown summer-stressed Phillyreaangustifolia. Optical signals of oxidative stress? New Phytologist 162: 115–124.
Pe~nuelas J, Sardans J. 2009a. Elementary factors. Nature 460: 803–804.Pe~nuelas J, Sardans J. 2009b. Ecological metabolomics. Chemistry and Ecology25: 305–309.
Pe~nuelas J, Sardans J, Estiarte M, Ogaya R, Carnicer J, Barbeta A, Rivas-Ubach
A, Garbulsky M, Filella I. 2013a. Evidence of current impact of climate
change on life: a walk from genes to the biosphere. Global Change Biology 19:2303–2338.
Pe~nuelas J, Sardans J, Llusi�a J, Silva J, Owen S, Bala-Ola B, Linatoc AC,
Dalimin MN, Niinemets €U. 2013c. Foliar chemistry and standing folivory of
early and late-successional species in a Bornean rainforest. Plant Ecology andDiversity 6: 245–256.
Pe~nuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Boucher O,
Godderis Y, Hinsinger P, Llusia J et al. 2013b.Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe.
Nature Communications 4: 2934.Pluskal T,Castillo S, Villar-Briones A,OresicM. 2010.MZmine 2:modular
framework for processing, visualizing, and analyzingmass spectrometry-based
molecular profile data.BMCBioinformatics 11: doi: 10.1186/1471-2105-11-395.Powell JA, Logan JA. 2005. Insect seasonality: circle map analysis of
temperature-driven life cycles. Theoretical Population Biology 67: 161–179.Price AH, Atherton NM, Hendry GAF. 1989. Plants under drought-stress
generate activated oxygen. Free Radical Research Communications 8: 61–66.R Development Core Team. 2011. R: a language and environment for statisticalcomputing. Vienna, Austria: R Foundation for Statistical Computing. ISBN
3-900051-07-0, URL http://www.R-project.org/.
Raffa KF, Powell EN, Townsend PA. 2013. Temperature-driven range
expansion of an irruptive insect heightened by weakly coevolved plant defenses.
Proceedings of the National Academy of Sciences, USA 110: 2193–2198.
New Phytologist (2014) 202: 874–885 � 2014 The Authors
New Phytologist� 2014 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist884

Rani PU, Pratyusha S. 2013. Defensive role of Gossypium hirsutum L.
anti-oxidative enzymes and phenolic acids in response to Spodoptera litura F.feeding. Journal of Asia-Pacific Entomology 16: 131–136.
Raubenheimer D, Simpson SJ. 2003. Nutrient balancing in grasshoppers:
behavioural and physiological correlates of dietary breadth. Journal ofExperimental Biology 206: 1669–1681.
RiceEvans CA, Miller J, Paganga G. 1997. Antioxidant properties of phenolic
compounds. Trends in Plant Science 2: 152–159.Rivas-Ubach A, P�erez-Trujillo M, Sardans J, Gargallo-Garriga A, Parella T,
Pe~nuelas J. 2013. Ecometabolomics: optimized NMR-based method.Methodsin Ecology and Evolution 4: 464–473.
Rivas-Ubach A, Sardans J, P�erez-Trujillo M, Estiarte M, Pe~nuelas J. 2012.
Strong relationship between elemental stoichiometry and metabolome in
plants. Proceedings of the National Academy of Sciences, USA 109: 4181–4186.Rouault G, Candau J-N, Lieutier F, Nageleisen L-M, Martin J-C, Warzee N.
2006. Effects of drought and heat on forest insect populations in relation to the
2003 drought in Western Europe. Annals of Forest Science 63: 613–624.Sardans J, Gargallo-Garriga A, P�erez-Trujillo M, Parella T, Seco R, Filella I,
Pe~nuelas J. 2013a.Metabolic responses of Quercus ilex seedlings to woundinganalysed with nuclear magnetic resonance. Plant Biology. doi:10.1111/plb.12032
Sardans J, Montes F, Pe~nuelas J. 2010. Determination of As, Cd, Cu, Hg and Pb
in biological samples by modern electrothermal atomic absorption
spectrometry. Spectrochimica Acta Part B-Atomic Spectroscopy 65: 97–112.Sardans J, Pe~nuelas J. 2012. The role of plants in the effects of global change on
nutrient availability and stoichiometry in the plant-soil system. Plant Physiology160: 1741–1761.
Sardans J, Pe~nuelas J, Coll M, Vayreda J, Rivas-Ubach A. 2012a. Stoichiometry
of potassium is largely determined by water availability and growth in
Catalonian forests. Functional Ecology 26: 1077–1089.Sardans J, Pe~nuelas J, Rivas-Ubach A. 2011b. Ecological metabolomics: overview
of current developments and future challenges. Chemoecology 21: 191–225.Sardans J, Rivas-Ubach A, Estiarte M, Ogaya R, Pe~nuelas J. 2013b.
Field-simulated droughts affect elemental leaf stoichiometry in Mediterranean
forests and shrublands. Acta Oecologica 50: 20–31.Sardans J, Rivas-Ubach A, Pe~nuelas J. 2011a. Factors affecting nutrientconcentration and stoichiometry of forest trees in Catalonia (NE Spain). ForestEcology and Management 262: 2024–2034.
Sardans J, Rivas-Ubach A, Pe~nuelas J. 2012a. The C : N : P stoichiometry of
organisms and ecosystems in a changing world: a review and perspectives.
Perspectives in Plant Ecology, Evolution and Systematics 14: 33–47.Sardans J, Rivas-Ubach A, Pe~nuelas J. 2012b. The elemental stoichiometry of
aquatic and terrestrial ecosystems and its relationships with organismic lifestyle
and ecosystem structure and function: a review and perspectives.
Biogeochemistry 111: 1–39.Spiess N, Oufir M, Matusikova I, Stierschneider M, Kopecky D, Homolka A,
Burg K, Fluch S, Hausman J-F, Wilhelm E. 2012. Ecophysiological and
transcriptomic responses of oak (Quercus robur) to long-term drought exposure
and rewatering. Environmental and Experimental Botany 77: 117–126.Sterner RW, Elser JJ. 2002. Ecological stoichiometry: the biology of elements frommolecules to the biosphere. Princeton, NJ, USA: Princeton University Press.
Strack D, Wray V, Harborne JB. 1994. The flavonoids, advances in research since1986. Boca Raton, FL, USA: Chapman & Hall.
Treutter D. 2006. Significance of flavonoids in plant resistance: a review.
Environmental Chemistry Letters 4: 147–157.Wang M, Zheng Q, Shen Q, Guo S. 2013. The critical role of potassium in plant
stress response. International Journal of Molecular Sciences 14: 7370–7390.White TCR. 1984. The abundance of invertebrate herbivores in relation to the
availability of nitrogen in stressed food plants. Oecologia 63: 90–105.Widarto HT, Van Der Meijden E, Lefeber AWM, Erkelens C, Kim HK, Choi
YH, Verpoorte R. 2006.Metabolomic differentiation of Brassica rapafollowing herbivory by different insect instars using two-dimensional nuclear
magnetic resonance spectroscopy. Journal of chemical ecology 32: 2417–2428.Williams WP, Davis FM, Buckley PM, Hedin PA, Baker GT, Luthe DS. 1998.
Factors associated with resistance to fall armyworm (Lepidoptera: Noctuidae)
and southwestern corn borer (Lepidoptera: Crambidae) in corn at different
vegetative stages. Journal of Economic Entomology 91: 1471–1480.
Xu J, Li Y, Sun J, Du L, Zhang Y, YuQ, Liu X. 2013.Comparative physiological
and proteomic response to abrupt low temperature stress between two winter
wheat cultivars differing in low temperature tolerance. Plant Biology 15: 292–303.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Soil moisture and temperature of sampled plots.
Fig. S2 Stem mortality and aboveground biomass of sampledplots.
Fig. S3Methodology for the calculation of herbivore attack.
Fig. S4 PC1 vs PC2 of a seasonal PCA of Quercus ilex leaves.
Fig. S5 PC1 vs PC2 of a PCA of Quercus ilex leaves in spring andsummer.
Fig. S6 PC1 vs PC2 of a PCA of Quercus ilex leaves in summerexcluding folivory as a variable.
Fig. S7 Foliar polyphenolic concentrations vs foliar consumptionin summer.
Table S1 NMR assignments of the metabolites identified inpolar samples
Table S2 NMR assignments of the main peaks of nonpolar sam-ples
Table S3 Details of LC-MS chromatogram processing
Table S4 Retention time and exact mass of metabolite assign-ment by LC-MS
Table S5 Seasonal one-way ANOVAs of identified metabolitesin leaves of Quercus ilex
Table S6 One-way ANOVAs of identified metabolites in leavesof Quercus ilex of control vs droughted trees with data of allseasons
Table S7 One-way ANOVAs of identified metabolites in leavesof Quercus ilex of control vs droughted trees with summer data
Methods S1 Details of chemical analyses.
Methods S2 Details of NMR metabolite elucidation.
Please note: Wiley Blackwell are not responsible for the contentor functionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
� 2014 The Authors
New Phytologist� 2014 New Phytologist TrustNew Phytologist (2014) 202: 874–885
www.newphytologist.com
NewPhytologist Research 885





























