Embed Size (px)
Citation preview

BRAINA JOURNAL OF NEUROLOGY
Dopaminergic neuroprotection of hormonalreplacement therapy in young and agedmenopausal rats: role of the brainangiotensin systemAna I. Rodriguez-Perez,1,2 Rita Valenzuela,1,2 Begona Villar-Cheda,1,2 Maria J. Guerra1,2 andJose L. Labandeira-Garcia1,2
1 Laboratory of Neuroanatomy and Experimental Neurology, Department of Morphological Sciences, Faculty of Medicine, University of Santiago de
Compostela, 15782 Santiago de Compostela, Spain
2 Networking Research Centre on Neurodegenerative Diseases (CIBERNED), 28031, Spain
Correspondence to: Jose L. Labandeira-Garcia MD, PhD,
Department of Morphological Sciences,
Faculty of Medicine,
University of Santiago de Compostela,
15782 Santiago de Compostela, Spain
E-mail: [email protected]
There is a lack of consensus about the effects of the type of menopause (surgical or natural) and of oestrogen replacement
therapy on Parkinson’s disease. The effects of the timing of replacement therapy and the female’s age may explain the observed
differences in such effects. However, the mechanisms involved are poorly understood. The renin-angiotensin system mediates
the beneficial effects of oestrogen in several tissues, and we have previously shown that dopaminergic cell loss is enhanced by
angiotensin via type 1 receptors, which is activated by ageing. In rats, we compared the effects of oestrogen replacement
therapy on 6-hydroxydopamine-induced dopaminergic degeneration, nigral renin-angiotensin system activity, activation of the
nicotinamide adenine dinucleotide phosphate oxidase complex and levels of the proinflammatory cytokine interleukin-1b in
young (surgical) menopausal rats and aged menopausal rats. In young surgically menopausal rats, the renin-angiotensin system
activity was higher (i.e. higher angiotensin converting enzyme activity, higher angiotensin type-1 receptor expression and lower
angiotensin type-2 receptor expression) than in surgically menopausal rats treated with oestrogen; the nicotinamide adenine
dinucleotide phosphate oxidase activity and interleukin-1b expression were also higher in the first group than in the second
group. In aged menopausal rats, the levels of nigral renin-angiotensin and nicotinamide adenine dinucleotide phosphate oxidase
activity were similar to those observed in surgically menopausal rats. However, oestrogen replacement therapy significantly
reduced 6-hydroxydopamine-induced dopaminergic cell loss in young menopausal rats but not in aged rats. Treatment with
oestrogen also led to a more marked reduction in nigral renin-angiotensin and nicotinamide adenine dinucleotide phosphate
oxidase activity in young surgically menopausal rats (treated either immediately or after a period of hypo-oestrogenicity) than in
aged menopausal rats. Interestingly, treatment with the angiotensin type-1 receptor antagonist candesartan led to remarkable
reduction in renin-angiotensin system activity and dopaminergic neuron loss in both groups of menopausal rats. This suggests
that manipulation of the brain renin-angiotensin system may be an efficient approach for the prevention or treatment of
Parkinson’s disease in oestrogen-deficient females, together with or instead of oestrogen replacement therapy.
Keywords: ageing; angiotensin; oestrogen; menopause; Parkinson’s disease
doi:10.1093/brain/awr320 Brain 2012: 135; 124–138 | 124
Received March 25, 2011. Revised September 7, 2011. Accepted October 3, 2011. Advance Access publication December 20, 2011
� The Author (2011). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved.
For Permissions, please email: [email protected]
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

IntroductionSex steroids have been shown to have a neuroprotective role in
several models of neurological diseases, although the effects of the
loss of ovarian function and hormonal replacement therapy in
humans are controversial. Numerous observational studies have
supported the concept that oestrogen therapy in postmenopausal
females protects against ageing-related diseases, including cardio-
vascular diseases, stroke and neurodegeneration. This was not
confirmed in several randomized controlled trials that reported
no or even detrimental effects (Rossouw et al., 2002;
Chlebowski et al., 2010). Several possible explanations have
been proposed for the discrepant results (Clarkson and
Mehaffey, 2009). However, the age of the females receiving the
treatment appears to be a major factor. The vast majority of fe-
males participating in these trials were on average 565 years, and
had started oestrogen therapy 12 years after undergoing meno-
pause (Turgeon et al., 2004, 2006); on the contrary, in observa-
tional studies that reported beneficial effects, most females had
initiated oestrogen therapy in their perimenopausal period
(Harman et al., 2005; Miller et al., 2005, 2009). The mechanisms
underlying the differences in the effects of oestrogen therapy be-
tween young perimenopausal females (including early surgical
menopause) and senescent females are still poorly understood
(Sohrabji and Bake, 2006; Suzuki et al., 2007).
Experiments with animal models (Dluzen, 1997; Murray et al.,
2003), epidemiological and clinical evidence (Shulman, 2002;
Ragonese et al., 2006a, b; Liu and Dluzen, 2007) suggest that
sex hormones have a beneficial influence on the risk, onset and
severity of Parkinson’s disease, and that oestrogen modulates
dopaminergic function. The anti-inflammatory effects of oestrogen
appear to play a major role in the neuroprotective effects on
Parkinson’s disease (Suzuki et al., 2007; Vegeto et al., 2008).
However, conflicting findings have been reported from some clin-
ical trials and there is still a lack of consensus about the effects of
the type of menopause (i.e. surgical versus natural) and of the
oestrogen therapy on Parkinson’s disease (Ragonese et al.,
2006a, b; Miller et al., 2009). As already mentioned for other
diseases, the effects of the timing of postmenopausal oestrogen
therapy and the age of the females receiving the treatment may
be a major factor.
The renin-angiotensin system mediates the beneficial effects of
oestrogen in several tissues (Nickenig et al., 1998; Dean et al.,
2004; Chen et al., 2008), and interactions between oestrogen and
angiotensin receptors have also been observed (Liu et al., 2002;
Tasuda et al., 2005; Xue et al., 2007; Hoshi-Fukushima et al.,
2008). In addition, the local renin-angiotensin system has been
reported to be involved in age-related degenerative changes in
several tissues (Heymes et al., 1998; Basso et al., 2005; Benigni
et al., 2009). The peptide angiotensin, via type 1 (AT1) receptors,
is one of the most important known inducers of inflammation and
oxidative stress, and produces reactive oxygen species by activa-
tion of the NADPH oxidase complex (Seshiah et al., 2002; Touyz
et al., 2002; Cai et al., 2003), which is the most important intra-
cellular source of reactive oxygen species apart from mitochondria
(Babior, 1999, 2004). The brain possesses a local renin-angiotensin
system (Mckinley et al., 2003; Savedra et al., 2005), and we have
previously shown that the dopaminergic cell loss is enhanced by
angiotensin via AT1 receptors (Rey et al., 2007; Rodriguez-Pallares
et al., 2008; Joglar et al., 2009). In addition, we have observed
that oestrogen decreases and ageing increases the effect of the
nigral renin-angiotensin system on enhancing dopaminergic
neuron degeneration in young female rats and aged male rats,
respectively (Rodriguez-Perez et al., 2010; Villar-Cheda et al.,
2010b). It is therefore possible that ageing modifies the beneficial
effects of oestrogen therapy in postmenopausal females, and that
the nigral renin-angiotensin system plays a major role. In the pre-
sent study, we compared the effects of oestrogen therapy on
dopaminergic degeneration, nigral renin-angiotensin system activ-
ity, activation of the NADPH oxidase complex and levels of the
pro-inflammatory cytokine interleukin-1b (IL-1b) in rat models of
untreated and treated (either immediate or delayed oestrogen
therapy) surgical (young ovariectomized rats) and natural (aged
menopausal rats) menopause, and investigated if renin-angiotensin
system manipulation may improve or replace the effects of oes-
trogen therapy.
Materials and methods
Experimental designYoung adult and aged acyclic female Sprague-Dawley rats
(10 weeks and 24 months old at the beginning of the experiments,
respectively) were used. All experiments were carried out in ac-
cordance with the ‘Principles of laboratory animal care’ (NIH
publication No. 86-23, revised 1985) and approved by the corres-
ponding committee at the University of Santiago de Compostela.
All surgery was performed under ketamine/xylazine anaesthesia,
and rats were fed with 2019S Teklad Rodent Maintenance Diet
(Harlan Laboratories) to minimize the occurrence of natural phyto-
estrogens. Five rat models corresponding to five major possibilities
in clinical menopause were studied: untreated surgical menopause
(young ovariectomized rats), surgical menopause immediately
treated with oestrogen therapy, surgical menopause treated with
oestrogen therapy after a period of hypo-oestrogenicity, untreated
natural menopause (untreated aged rats), and natural menopause
treated with oestrogen therapy. In addition, a group of aged rats
were ovariectomized and treated with oestrogen to study the po-
tential effects of the presence of ovaries on renin-angiotensin
system activity in aged rats.
The rats were divided into six groups. Rats in Group A were
young rats (10-weeks-old at the beginning of the experiments),
which were ovariectomized (i.e. menopause was induced surgi-
cally) and given empty implants (n = 38). Rats in Group B were
young rats that were ovariectomized and simultaneously received
implants containing 17b-oestradiol (n = 31). Rats in Group C were
treated as Group B rats but the oestrogen implants were given
3 weeks after ovariectomy. Rats in Group D were aged rats
(24 months old; aged; n = 31) given empty implants (i.e. natural
senescent menopause). Rats in Group E were aged rats given im-
plants containing 17b-oestradiol (n = 25). Rats in Group F were
aged rats that were ovariectomized and given implants containing
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 125
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

17b-oestradiol (n = 5). Two series of experiments were carried out
with the different groups of rats (Groups A–F). In the first series of
experiments, young ovariectomized or aged menopausal rats
(n = 55) were used to determine the effect of oestrogen and can-
desartan (i.e. inhibition of renin-angiotensin system activity by
this AT1 receptor antagonist) or treatment with oestro-
gen + candesartan on the dopaminergic degeneration induced by
the neurotoxin 6-hydroxydopamine, relative to rats in the different
groups treated with 6-hydroxydopamine alone and compared with
the corresponding controls injected with vehicle. A second series
of experiments was carried out to investigate the effect of treat-
ment with oestrogen or candesartan on markers of NADPH oxi-
dase activity/oxidative stress, inflammation and renin-angiotensin
system activity in young ovariectomized and aged menopausal rats
(n = 87). Rats in the first series of experiments were injected
intrastriatally with 6-hydroxydopamine or vehicle (controls) and
treated with oestrogen and/or candesartan or vehicle (controls),
then killed for immunohistochemical studies (i.e. quantification of
dopaminergic cell death), as described below. Rats in the second
series of experiments were killed by decapitation 3 weeks after
ovariectomy and/or treatment with implants. The brains were rap-
idly removed and coronal slices of the mesencephalon were cut
with a tissue chopper set to 1 mm. To obtain substantia nigra
compacta, the individual 1-mm tissue slides were dissected on a
pre-cooled glass plate under a stereoscopic microscope (Leica
M220). The substantia nigra compacta was dissected according
to Paxinos and Watson (1985), frozen on dry ice, and stored at
�80�C until processed for investigation of expression of AT1 and
AT2 receptors, expression of the NADPH oxidase cytosolic subunit
p47phox and expression pro-inflammatory cytokine IL-1b by west-
ern blot and reverse transcriptase–polymerase chain reaction
studies.
Expression of the NADPH oxidase cytosolic subunit p47phox is
an indicator of the level of activation of the NADPH oxidase com-
plex. The NADPH oxidase complex is composed of membrane-
bound and cytosolic subunits such as p47phox, which is considered
key for NADPH oxidase activation (Li and Shah, 2003).
Translocation of cytosolic subunits to the membrane, which
leads to reactive oxygen species generation, is a necessary step
for NADPH oxidase activation. The level of the NADPH oxidase
subunit p47phox is correlated with NADPH oxidase activity and
NADPH-derived superoxide formation (Rueckschloss et al., 2002;
Touyz et al., 2002). Finally, angiotensin converting enzyme activ-
ity was determined by lucigenin-enhanced chemiluminescence (see
below).
Oestrogen and candesartanadministrationRats were bilaterally ovariectomized through a dorsal incision and
simultaneously (Groups A, B and F) or 3 weeks later (Group C)
received Silastic implants, placed subcutaneously in the midscapu-
lar region (Dziuk and Cook, 1966; Febo et al., 2005). Aged acyclic
rats (Groups D and E) received similar implants without prior ovari-
ectomy. Silastic implants were prepared with Silastic� tubing
(1.47 mm inner diameter � 1.95 mm outer diameter, Dow
Corning 508-006; VWR Scientific), as described by Febo et al.
(2005). Briefly, 5-mm long sections of tubing were sealed at one
end with Silastic silicone sealant (Dow Corning 732; VWR) and left
to dry for 30 min. The implants were then either filled with crys-
talline 17-b-oestradiol (17-b-oestradiol benzoate; Sigma-Aldrich;
Groups B, C, E and F) or were left empty (Groups A and D);
the open end was then sealed in the same way as the other
end. Implants were air-dried and incubated in sterile saline for at
least 12–16 h to allow the initial surge of high oestradiol levels to
be released before use. It has been observed that such implants
achieve stable levels of plasma oestradiol over 30 days, with a
release rate of 75–100 pg/ml per 24 h (Febo et al., 2005), as con-
firmed in our previous studies (Rodriguez-Perez et al., 2010).
However, stable levels of oestrogen have also been found to per-
sist for only 7–24 days (Mannino et al., 2005). Therefore, rats
were killed 3 weeks after oestrogen implantation (i.e. 2 weeks
after 6-hydroxydopamine injection).
Some rats in the different groups received candesartan in
their drinking water (candesartan cilexetil, AstraZeneca; 3 mg/kg/
day) from 7 days before the empty or oestrogen implants
were fitted until they were killed. The oral bioavailability of
candesartan cilexetil is 5–10% oral (Kondo et al., 1996; Zorad
et al., 2006); the drug was diluted (0.025 mg/ml) following
the protocol suggested by AstraZeneca and administered to the
rats in their drinking water (25–30 ml/day/rat). It has been re-
ported that candesartan is the most effective AT1 antagonist in
crossing the blood–brain barrier, and that low doses have little
effect on blood pressure and block brain angiotensin effects
(Unger, 2003).
Intrastriatal injection of6-hydroxydopamineOne week after receiving empty or oestrogen implants, some rats
in the different groups were injected intrastriatally with
6-hydroxydopamine or vehicle. Thirty minutes prior to intrastriatal
injection with 6-hydroxydopamine or vehicle, rats were treated
with the selective inhibitor for the norepinephrine transporter
desipramine (Sigma, 25 mg/kg i.p.) to prevent uptake of
6-hydroxydopamine by noradrenergic terminals. The rats were in-
jected in the right striatum with 7 mg of 6-hydroxydopamine (in
3 ml of saline containing 0.2% ascorbic acid; Sigma). Stereotaxic
coordinates were 0.8 mm anterior to bregma, 3.0 mm right of
midline and 5.0 mm ventral to the dura; tooth bar at �3.3.
Control animals were injected with 3ml of vehicle alone. Rats
were killed 2-week post-lesion (i.e. 3 weeks post-implant).
Previous studies on the time course of 6-hydroxydopamine lesions
have shown that the loss of tyrosine hydroxylase immunoreactive
neurons is complete (Przedborski et al., 1995) or practically com-
plete (Sauer and Oertel, 1994) 2 weeks after administration of
intrastriatal injections. Although a few dopaminergic neurons
may degenerate after the 2-week period, we considered it more
important to kill the rats before any possible loss of oestrogen
levels (i.e. 3 weeks after implantation and 2 weeks after
6-hydroxydopamine injection).
126 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

RadioimmunoassayIn order to confirm the efficiency of ovariectomies and implants,
blood samples were taken from the rats by cardiac puncture just
before the animals were killed. Blood was collected on ice and
serum samples immediately frozen at �70� until analysis for
17b-oestradiol by radioimmunoassay, with the Diagnostic
Products Corporation Coat-a-Count kit and protocol. The samples
were counted for 1 min in a gamma counter, and assayed in
triplicate.
RNA extraction and real-timequantitative reversetranscriptase–polymerase chain reactionTotal RNA from the nigral region was extracted with TRIzol�
(Invitrogen), according to the manufacturer’s instructions. Total
RNA (2.5 mg) was reverse-transcribed to complementary DNA
with deoxynucleotide triphosphates, random primers and
Moloney murine leukaemia virus reverse transcriptase (M-MLV;
200 U; Invitrogen). Real-time polymerase chain reaction was
used to examine relative levels of angiotensin receptors type 1
(AT1a) and type 2 (AT2) messenger RNA, p47 and IL-1b.
Experiments were performed with a real-time iCyclerTM polymer-
ase chain reaction platform (Bio-Rad). b-Actin was used as house-
keeping gene and was amplified in parallel with the genes of
interest. The comparative Ct method was used to examine the
relative messenger RNA expression. The expression of each gene
was obtained as relative to the housekeeping transcripts. The data
were then normalized to the values of the control group of the
same batch (i.e. expressed as a percentage of the control; 100%)
to counteract any possible variability among batches. Two house-
keeping genes [b-actin and GAPDH (glyceraldehyde 3-phosphate
dehydrogenase)] were simultaneously included in some experi-
ments to exclude any potential effect of oestrogen on the house-
keeping gene: no significant differences were detected. Finally, the
results were expressed as mean � SEM. Primers sequences were as
follows: for AT1a, forward 50-TTCAACCTCTACGCCAGTGTG-30,
reverse 50-GCCAAGCCAGCCATCAGC-30; for AT2, forward
50-AACATCTGCTGAAGACCAATAG-30, reverse 50-AGAAGGTC
AGAACATGGAAGG-30; for p47phox, forward 50-CCACAC
CTCTTGAACTTCTTC-30, reverse 50-CTCGTAGTCAGCGATGGC-30;
for b-actin, forward 50-TCGTGCGTGACATTAAAGAG-30, reverse
50-TGCCACAGGATTCCATACC-30; for and IL-1b forward 50-
ATCTCACAGCAGCATCTC, reverse 50-TAGCAGGTCGTCATCATC.
Western blot analysis and angiotensinconverting enzyme activityFor western blot analysis, tissue was homogenized in radioimmu-
noprecipitation buffer containing protease inhibitor cocktail
(P8340, Sigma) and phenylmethylsulphonyl fluoride (P7626,
Sigma). The homogenates were centrifuged and protein concen-
trations were determined with the Bradford protein assay. Equal
amounts of protein were separated by 5–10% bis–Tris polyacryl-
amide gel, and transferred to nitrocellulose membrane. The
membranes were incubated overnight with primary antibodies
(1:200) against AT1 receptor (sc-31181), AT2 receptor
(sc-9040), p47-phox (sc-7660) and IL-1b (sc-1252), all from
Santa Cruz Biotechnology. The horseradish peroxidase conjugated
secondary antibodies used were Protein A (NA9120V, GE
Healthcare) and Protein G (18-161, Upstate-Millipore).
Immunoreactivity was detected with an Immun-StarTM HRP
Chemiluminescent Kit (170-5044, Bio-Rad) and imaged with a
chemiluminescence detection system (Molecular Imager
ChemiDoc XRS System, Bio-Rad). Blots were stripped and
re-probed for anti-GAPDH (G9545, Sigma; 1:25 000) as loading
control. In each animal, protein expression was measured by
densitometry of the corresponding band and expressed as relative
to the GAPDH band value. The data were then normalized to the
values of the control group of the same batch (i.e. expressed as a
percentage of the control values; 100%) to counteract any pos-
sible variability among batches. GAPDH and b-actin were simul-
taneously included as loading control in some experiments to
exclude any potential effects of oestrogen on the internal control:
no significant differences were detected. Finally, the results were
expressed as means � SEM.
Angiotensin converting enzyme activity in ventral mesencephalic
tissue was assayed with hippuryl-L-histidyl-L-leucine (Hip-His-Leu;
Sigma) as substrate, as described by Hemming et al. (2007).
Fluorescence was assayed in 96-well plates in an Infinite M200
multi-well plate reader (TECAN; excitation, 355; emission, 535)
and determined as nmol his-leu/mg protein/min. The data
were then normalized to the values of the control group of
the same batch (i.e. expressed as a percentage of the control
values; 100%)
Immunohistochemistry anddopaminergic neuron quantificationThe animals used for immunohistochemistry (i.e. those injected
intrastriatally with 6-hydroxydopamine or vehicle) were first per-
fused with 0.9% saline and then with cold 4% paraformaldehyde
in 0.1 M phosphate buffer, pH 7.4. The brains were removed and
subsequently washed and cryoprotected in the same buffer con-
taining 20% sucrose, and finally cut into 40-mm sections on a
freezing microtome. The sections were incubated for 1 h in 10%
normal swine serum with 0.25% Triton X-100 in 20 mM potas-
sium phosphate-buffered saline containing 1% bovine serum al-
bumin and then incubated overnight at 4�C with a mouse
monoclonal anti-tyrosine hydroxylase (Sigma; 1:10 000), as dopa-
minergic marker. The sections were subsequently incubated, first
for 60 min with the corresponding biotinylated secondary anti-
body, and then for 90 min with avidin–biotin–peroxidase complex
(ABC, 1:100, Vector). Finally the labelling was revealed by treat-
ment with 0.04% hydrogen peroxide and 0.05% 3-
30diaminobenzidine (DAB, Sigma). In all experiments the control
sections, in which the primary antibody was omitted, were immu-
nonegative for these markers.
The total number of tyrosine hydroxylase immunoreactive
neurons in the substantia nigra compacta was estimated by
an unbiased stereological method (the optical fractionator).
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 127
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

The stereological analysis was carried out with the Olympus
CAST-Grid system (Computer Assisted Stereological Toolbox;
Olympus). Uniform randomly chosen sections through the sub-
stantia nigra (every fourth section) were analysed for the total
number of tyrosine hydroxylase immunoreactive cells by means
of a stereological grid (fractionator), and the nigral volume was
estimated according to Cavalieri’s method (Gundersen et al.,
1988). To confirm that 6-hydroxydopamine induces cell death,
series of sections through the entire substantia nigra of control
rats and rats treated with 6-hydroxydopamine were counter-
stained with Cresyl violet, and the total number of neurons in
the substantia nigra was estimated by the unbiased stereology
method described above for tyrosine hydroxylase immunoreactive
cells. Neurons were distinguished from glial cells on a morpho-
logical basis, and neurons with visible nuclei were counted as
above for tyrosine hydroxylase immunoreactive neurons (see Rey
et al., 2007 for details).
Statistical analysisAll data were obtained in triplicate (i.e. from each rat at least three
samples were independently processed and the mean value for
each rat was used for statistical analysis). Data were expressed
as means � SEM. Two group comparisons were analysed by a
Student’s t-test and multiple comparisons were analysed by
one-way ANOVA followed by a post hoc Bonferroni test. The
normality of populations and homogeneity of variances were
tested before each ANOVA. Differences were considered signifi-
cant at P50.05. Statistical analyses were carried out with
SigmaStat 3.0 from Jandel Scientific.
Results
Effect of the dopaminergic neurotoxin6-hydroxydopamine on different groupsof menopausal ratsIntrastriatal injection of 6-hydroxydopamine induced a marked loss
of dopaminergic neurons in the substantia nigra of aged rats and
young ovariectomized rats, which was significantly higher than
that induced by 6-hydroxydopamine in young ovariectomized
rats immediately treated with oestrogen, or young ovariectomized
rats treated with oestrogen 3 weeks after ovariectomy. The loss of
dopaminergic neurons induced by the neurotoxin in aged rats
treated with oestrogen tended to be lower than in untreated
aged rats, but the difference was not significant. Interestingly,
however, the loss of dopaminergic neurons was significantly
lower in both groups of menopausal rats treated with the
AT1 antagonist candesartan, and treatment with oestro-
gen + candesartan did not increase the neuroprotective effect
induced by treatment with candesartan alone (Fig. 1A–E).
Therefore, the present results indicate that either oestrogen or
candesartan induced significant neuroprotection against
6-hydroxydopamine in young menopausal rats (surgically induced
menopause). However, only candesartan induced significant neu-
roprotection in aged menopausal rats (natural menopause).
Renin-angiotensin system activity inyoung surgically menopausal and agedmenopausal ratsBoth groups of menopausal rats showed very low blood levels of
oestrogen. Blood levels of oestrogen in young ovariectomized rats
were 18.6 � 2.4 pg/ml, and similar to those observed in aged
menopausal rats (26.2 � 4.5 pg/ml). Comparison between both
groups of menopausal rats did not reveal any significant differ-
ences in the expression of AT2 and the NADPH subunit p47phox
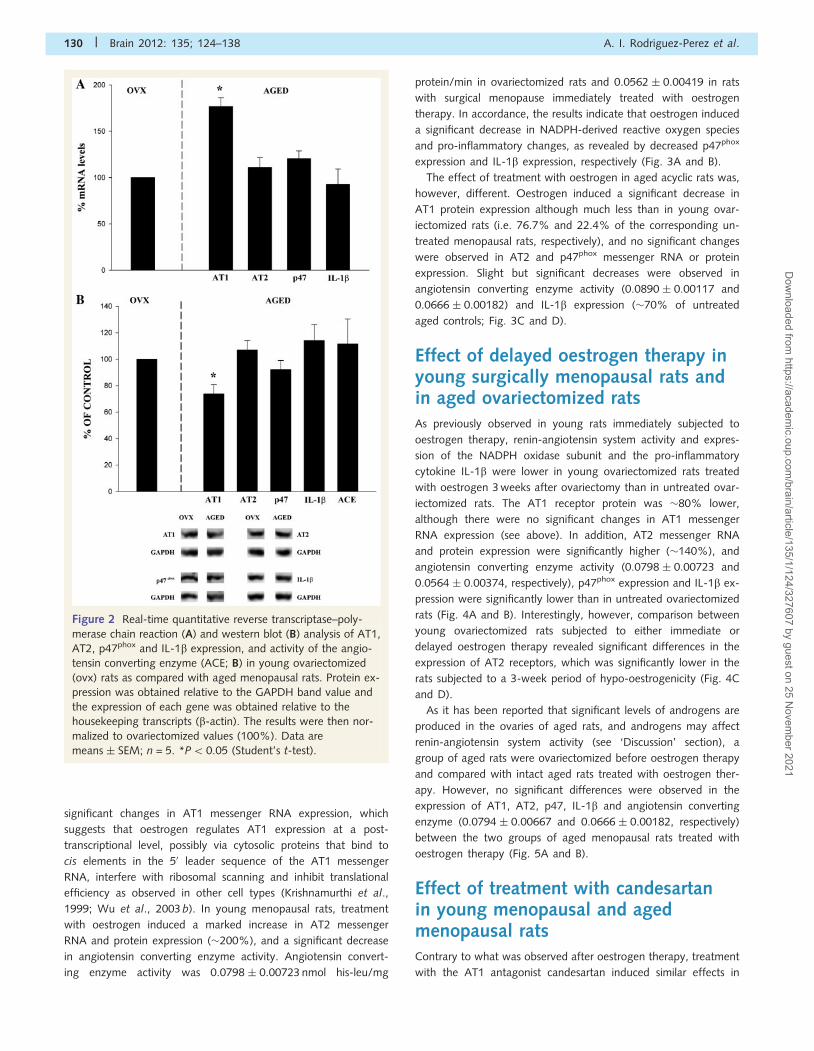
and IL-b (Fig. 2A and B). However, differences were observed
between both groups of menopausal rats in AT1 receptor expres-
sion; AT1 messenger RNA expression was higher in aged rats.
Since the present and previous studies have shown that oestrogen
affects AT1 expression at post-transcriptional levels (see below),
the increased messenger RNA expression in aged rats must be
related to ageing-related factors other than low levels of oestro-
gen. Interestingly, AT1 protein expression was higher in young
ovariectomized rats, which suggests that the lack of oestrogen
affects the expression of AT1 receptors to a greater extent in
young ovariectomized rats than in aged rats. Similar levels of
AT2 messenger RNA and protein expression in young ovariecto-
mized and aged rats also suggest that ageing-related factors other
than low levels of oestrogen reduce the expression of AT2 recep-
tors in aged rats, because the lack of oestrogen reduced AT2 re-
ceptor levels in young rats but did not significantly change AT2
receptor levels in aged rats (see below). Angiotensin converting
enzyme activity was 0.0798 � 0.00723 nmol his-leu/mg protein/
min in ovariectomized rats and 0.0890 � 0.0117 in aged rats.
Effect of oestrogen therapy in youngsurgically menopausal and agedmenopausal rats compared with thecorresponding untreated menopausalratsIn another set of experiments, we tested the effects of oestrogen
therapy with respect to the corresponding untreated menopausal
rats. Blood levels of oestrogen in young ovariectomized rats that
received implants with oestrogen were 144.8 � 16.7 pg/ml, which
were similar to values observed in normal rats in proestrous (refer
to ‘Discussion’ section). Plasma oestrogen concentrations in aged
menopausal rats that received implants with oestrogen were not
significantly different from those of young ovariectomized rats
treated with oestrogen (151.1 � 22.4 pg/ml immediately before
sacrifice). However, the effects of oestrogen treatment on
renin-angiotensin system activity were different. In young surgi-
cally ovariectomized rats, treatment with oestrogen at the time of
ovariectomy induced a marked decrease in renin-angiotensin
system activity as well as in the expression of the NADPH oxidase
subunit and the pro-inflammatory cytokine IL-1b. The AT1 recep-
tor protein decreased by �80%. However, there were no
128 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
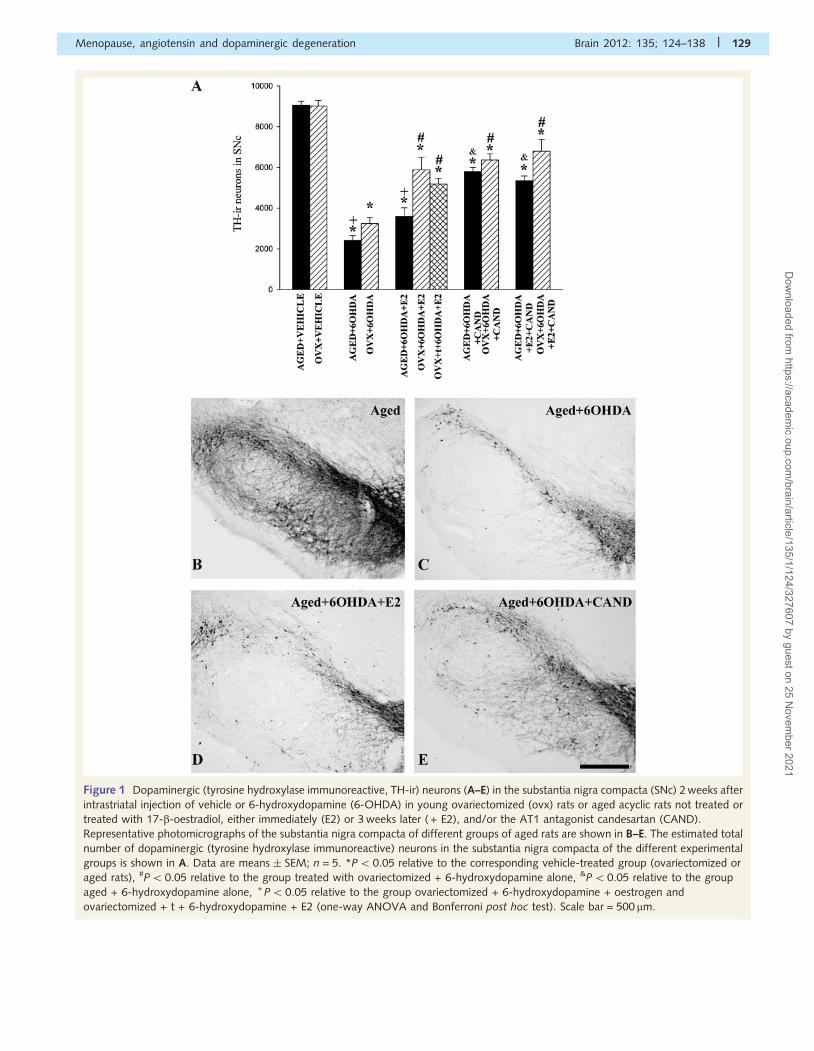
Figure 1 Dopaminergic (tyrosine hydroxylase immunoreactive, TH-ir) neurons (A–E) in the substantia nigra compacta (SNc) 2 weeks after
intrastriatal injection of vehicle or 6-hydroxydopamine (6-OHDA) in young ovariectomized (ovx) rats or aged acyclic rats not treated or
treated with 17-b-oestradiol, either immediately (E2) or 3 weeks later ( + E2), and/or the AT1 antagonist candesartan (CAND).
Representative photomicrographs of the substantia nigra compacta of different groups of aged rats are shown in B–E. The estimated total
number of dopaminergic (tyrosine hydroxylase immunoreactive) neurons in the substantia nigra compacta of the different experimental
groups is shown in A. Data are means � SEM; n = 5. *P5 0.05 relative to the corresponding vehicle-treated group (ovariectomized or
aged rats), #P50.05 relative to the group treated with ovariectomized + 6-hydroxydopamine alone, &P50.05 relative to the group
aged + 6-hydroxydopamine alone, + P50.05 relative to the group ovariectomized + 6-hydroxydopamine + oestrogen and
ovariectomized + t + 6-hydroxydopamine + E2 (one-way ANOVA and Bonferroni post hoc test). Scale bar = 500 mm.
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 129
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
significant changes in AT1 messenger RNA expression, which
suggests that oestrogen regulates AT1 expression at a post-
transcriptional level, possibly via cytosolic proteins that bind to
cis elements in the 50 leader sequence of the AT1 messenger
RNA, interfere with ribosomal scanning and inhibit translational
efficiency as observed in other cell types (Krishnamurthi et al.,
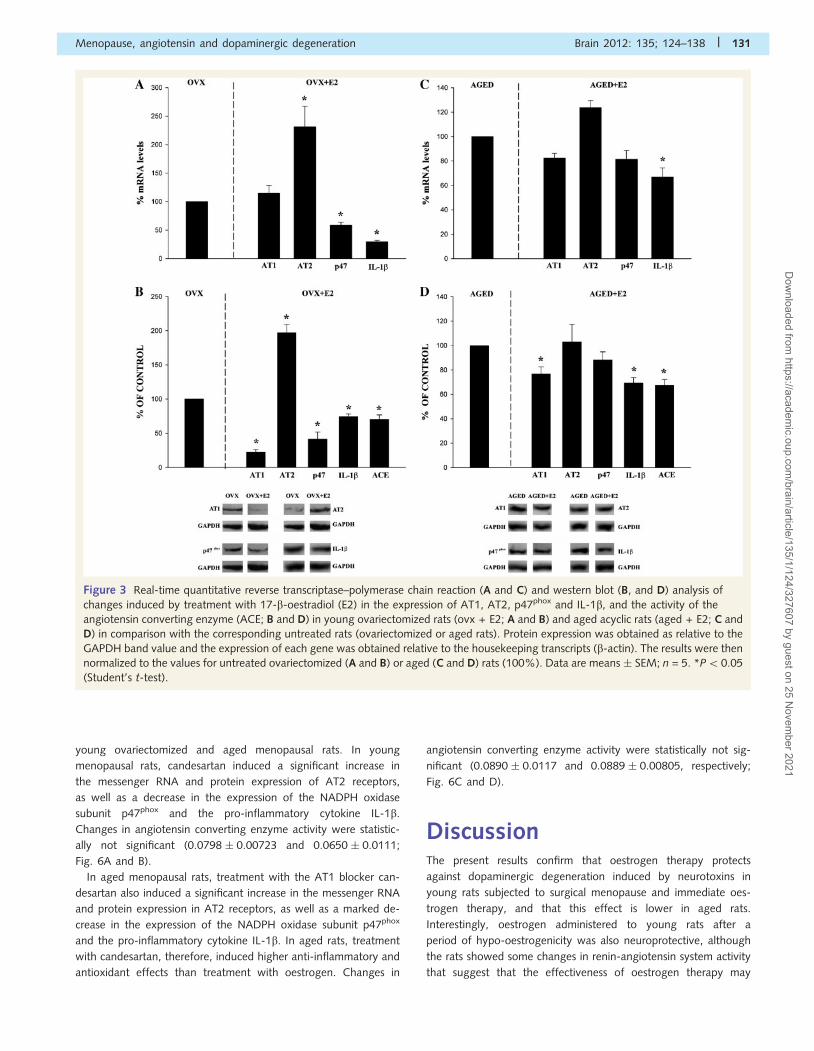
1999; Wu et al., 2003 b). In young menopausal rats, treatment
with oestrogen induced a marked increase in AT2 messenger
RNA and protein expression (�200%), and a significant decrease
in angiotensin converting enzyme activity. Angiotensin convert-
ing enzyme activity was 0.0798 � 0.00723 nmol his-leu/mg
protein/min in ovariectomized rats and 0.0562 � 0.00419 in rats
with surgical menopause immediately treated with oestrogen
therapy. In accordance, the results indicate that oestrogen induced
a significant decrease in NADPH-derived reactive oxygen species
and pro-inflammatory changes, as revealed by decreased p47phox
expression and IL-1b expression, respectively (Fig. 3A and B).
The effect of treatment with oestrogen in aged acyclic rats was,
however, different. Oestrogen induced a significant decrease in
AT1 protein expression although much less than in young ovar-
iectomized rats (i.e. 76.7% and 22.4% of the corresponding un-
treated menopausal rats, respectively), and no significant changes
were observed in AT2 and p47phox messenger RNA or protein
expression. Slight but significant decreases were observed in
angiotensin converting enzyme activity (0.0890 � 0.00117 and
0.0666 � 0.00182) and IL-1b expression (�70% of untreated
aged controls; Fig. 3C and D).
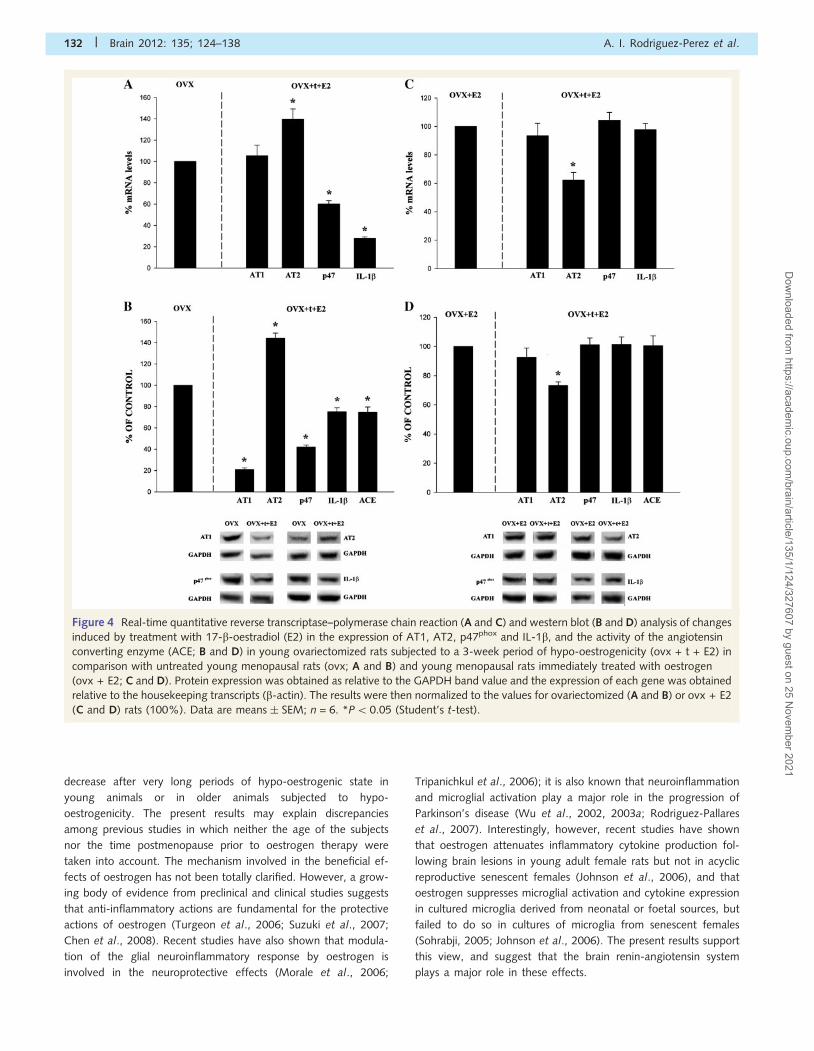
Effect of delayed oestrogen therapy inyoung surgically menopausal rats andin aged ovariectomized ratsAs previously observed in young rats immediately subjected to
oestrogen therapy, renin-angiotensin system activity and expres-
sion of the NADPH oxidase subunit and the pro-inflammatory
cytokine IL-1b were lower in young ovariectomized rats treated
with oestrogen 3 weeks after ovariectomy than in untreated ovar-
iectomized rats. The AT1 receptor protein was �80% lower,
although there were no significant changes in AT1 messenger
RNA expression (see above). In addition, AT2 messenger RNA
and protein expression were significantly higher (�140%), and
angiotensin converting enzyme activity (0.0798 � 0.00723 and
0.0564 � 0.00374, respectively), p47phox expression and IL-1b ex-
pression were significantly lower than in untreated ovariectomized
rats (Fig. 4A and B). Interestingly, however, comparison between
young ovariectomized rats subjected to either immediate or
delayed oestrogen therapy revealed significant differences in the
expression of AT2 receptors, which was significantly lower in the
rats subjected to a 3-week period of hypo-oestrogenicity (Fig. 4C
and D).
As it has been reported that significant levels of androgens are
produced in the ovaries of aged rats, and androgens may affect
renin-angiotensin system activity (see ‘Discussion’ section), a
group of aged rats were ovariectomized before oestrogen therapy
and compared with intact aged rats treated with oestrogen ther-
apy. However, no significant differences were observed in the
expression of AT1, AT2, p47, IL-1b and angiotensin converting
enzyme (0.0794 � 0.00667 and 0.0666 � 0.00182, respectively)
between the two groups of aged menopausal rats treated with
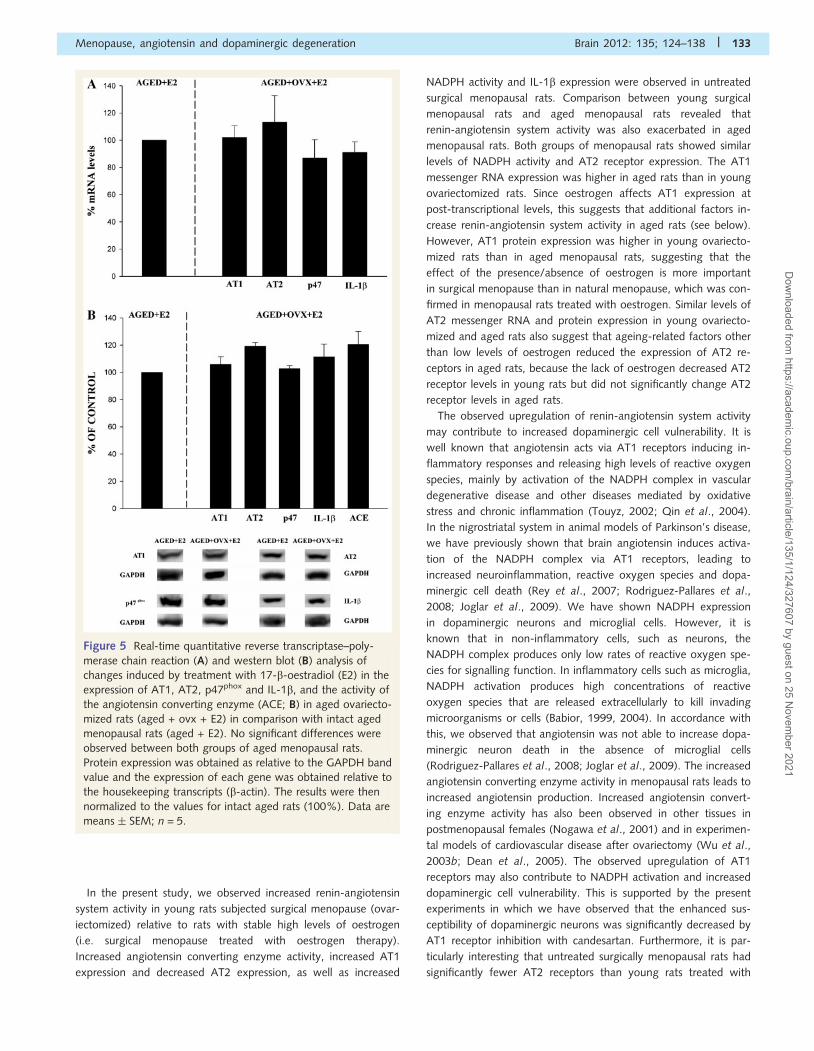
oestrogen therapy (Fig. 5A and B).
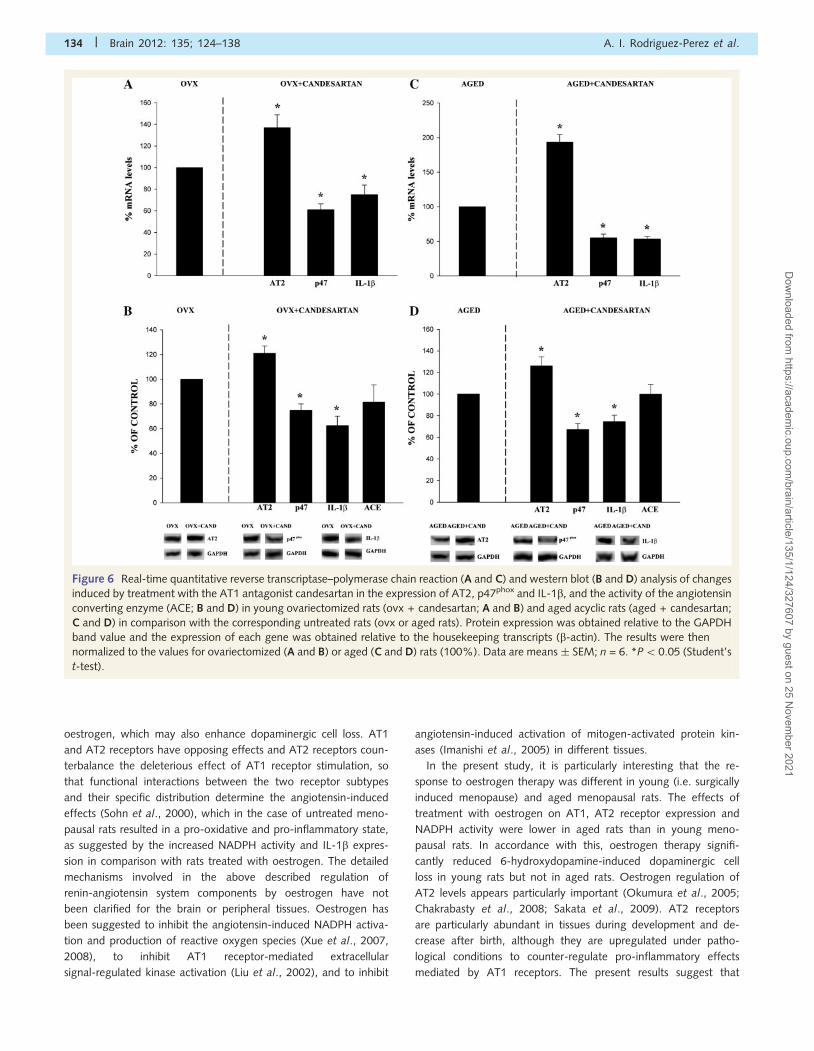
Effect of treatment with candesartanin young menopausal and agedmenopausal ratsContrary to what was observed after oestrogen therapy, treatment
with the AT1 antagonist candesartan induced similar effects in
Figure 2 Real-time quantitative reverse transcriptase–poly-
merase chain reaction (A) and western blot (B) analysis of AT1,
AT2, p47phox and IL-1b expression, and activity of the angio-
tensin converting enzyme (ACE; B) in young ovariectomized
(ovx) rats as compared with aged menopausal rats. Protein ex-
pression was obtained relative to the GAPDH band value and
the expression of each gene was obtained relative to the
housekeeping transcripts (b-actin). The results were then nor-
malized to ovariectomized values (100%). Data are
means � SEM; n = 5. *P5 0.05 (Student’s t-test).
130 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
young ovariectomized and aged menopausal rats. In young
menopausal rats, candesartan induced a significant increase in
the messenger RNA and protein expression of AT2 receptors,
as well as a decrease in the expression of the NADPH oxidase
subunit p47phox and the pro-inflammatory cytokine IL-1b.
Changes in angiotensin converting enzyme activity were statistic-
ally not significant (0.0798 � 0.00723 and 0.0650 � 0.0111;
Fig. 6A and B).
In aged menopausal rats, treatment with the AT1 blocker can-
desartan also induced a significant increase in the messenger RNA
and protein expression in AT2 receptors, as well as a marked de-
crease in the expression of the NADPH oxidase subunit p47phox
and the pro-inflammatory cytokine IL-1b. In aged rats, treatment
with candesartan, therefore, induced higher anti-inflammatory and
antioxidant effects than treatment with oestrogen. Changes in
angiotensin converting enzyme activity were statistically not sig-
nificant (0.0890 � 0.0117 and 0.0889 � 0.00805, respectively;
Fig. 6C and D).
DiscussionThe present results confirm that oestrogen therapy protects
against dopaminergic degeneration induced by neurotoxins in
young rats subjected to surgical menopause and immediate oes-
trogen therapy, and that this effect is lower in aged rats.
Interestingly, oestrogen administered to young rats after a
period of hypo-oestrogenicity was also neuroprotective, although
the rats showed some changes in renin-angiotensin system activity
that suggest that the effectiveness of oestrogen therapy may
Figure 3 Real-time quantitative reverse transcriptase–polymerase chain reaction (A and C) and western blot (B, and D) analysis of
changes induced by treatment with 17-b-oestradiol (E2) in the expression of AT1, AT2, p47phox and IL-1b, and the activity of the
angiotensin converting enzyme (ACE; B and D) in young ovariectomized rats (ovx + E2; A and B) and aged acyclic rats (aged + E2; C and
D) in comparison with the corresponding untreated rats (ovariectomized or aged rats). Protein expression was obtained as relative to the
GAPDH band value and the expression of each gene was obtained relative to the housekeeping transcripts (b-actin). The results were then
normalized to the values for untreated ovariectomized (A and B) or aged (C and D) rats (100%). Data are means � SEM; n = 5. *P50.05
(Student’s t-test).
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 131
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
decrease after very long periods of hypo-oestrogenic state in
young animals or in older animals subjected to hypo-
oestrogenicity. The present results may explain discrepancies
among previous studies in which neither the age of the subjects
nor the time postmenopause prior to oestrogen therapy were
taken into account. The mechanism involved in the beneficial ef-
fects of oestrogen has not been totally clarified. However, a grow-
ing body of evidence from preclinical and clinical studies suggests
that anti-inflammatory actions are fundamental for the protective
actions of oestrogen (Turgeon et al., 2006; Suzuki et al., 2007;
Chen et al., 2008). Recent studies have also shown that modula-
tion of the glial neuroinflammatory response by oestrogen is
involved in the neuroprotective effects (Morale et al., 2006;
Tripanichkul et al., 2006); it is also known that neuroinflammation
and microglial activation play a major role in the progression of
Parkinson’s disease (Wu et al., 2002, 2003a; Rodriguez-Pallares
et al., 2007). Interestingly, however, recent studies have shown
that oestrogen attenuates inflammatory cytokine production fol-
lowing brain lesions in young adult female rats but not in acyclic
reproductive senescent females (Johnson et al., 2006), and that
oestrogen suppresses microglial activation and cytokine expression
in cultured microglia derived from neonatal or foetal sources, but
failed to do so in cultures of microglia from senescent females
(Sohrabji, 2005; Johnson et al., 2006). The present results support
this view, and suggest that the brain renin-angiotensin system
plays a major role in these effects.
Figure 4 Real-time quantitative reverse transcriptase–polymerase chain reaction (A and C) and western blot (B and D) analysis of changes
induced by treatment with 17-b-oestradiol (E2) in the expression of AT1, AT2, p47phox and IL-1b, and the activity of the angiotensin
converting enzyme (ACE; B and D) in young ovariectomized rats subjected to a 3-week period of hypo-oestrogenicity (ovx + t + E2) in
comparison with untreated young menopausal rats (ovx; A and B) and young menopausal rats immediately treated with oestrogen
(ovx + E2; C and D). Protein expression was obtained as relative to the GAPDH band value and the expression of each gene was obtained
relative to the housekeeping transcripts (b-actin). The results were then normalized to the values for ovariectomized (A and B) or ovx + E2
(C and D) rats (100%). Data are means � SEM; n = 6. *P50.05 (Student’s t-test).
132 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
In the present study, we observed increased renin-angiotensin
system activity in young rats subjected surgical menopause (ovar-
iectomized) relative to rats with stable high levels of oestrogen
(i.e. surgical menopause treated with oestrogen therapy).
Increased angiotensin converting enzyme activity, increased AT1
expression and decreased AT2 expression, as well as increased
NADPH activity and IL-1b expression were observed in untreated
surgical menopausal rats. Comparison between young surgical
menopausal rats and aged menopausal rats revealed that
renin-angiotensin system activity was also exacerbated in aged
menopausal rats. Both groups of menopausal rats showed similar
levels of NADPH activity and AT2 receptor expression. The AT1
messenger RNA expression was higher in aged rats than in young
ovariectomized rats. Since oestrogen affects AT1 expression at
post-transcriptional levels, this suggests that additional factors in-
crease renin-angiotensin system activity in aged rats (see below).
However, AT1 protein expression was higher in young ovariecto-
mized rats than in aged menopausal rats, suggesting that the
effect of the presence/absence of oestrogen is more important
in surgical menopause than in natural menopause, which was con-
firmed in menopausal rats treated with oestrogen. Similar levels of
AT2 messenger RNA and protein expression in young ovariecto-
mized and aged rats also suggest that ageing-related factors other
than low levels of oestrogen reduced the expression of AT2 re-
ceptors in aged rats, because the lack of oestrogen decreased AT2
receptor levels in young rats but did not significantly change AT2
receptor levels in aged rats.
The observed upregulation of renin-angiotensin system activity
may contribute to increased dopaminergic cell vulnerability. It is
well known that angiotensin acts via AT1 receptors inducing in-
flammatory responses and releasing high levels of reactive oxygen
species, mainly by activation of the NADPH complex in vascular
degenerative disease and other diseases mediated by oxidative
stress and chronic inflammation (Touyz, 2002; Qin et al., 2004).
In the nigrostriatal system in animal models of Parkinson’s disease,
we have previously shown that brain angiotensin induces activa-
tion of the NADPH complex via AT1 receptors, leading to
increased neuroinflammation, reactive oxygen species and dopa-
minergic cell death (Rey et al., 2007; Rodriguez-Pallares et al.,
2008; Joglar et al., 2009). We have shown NADPH expression
in dopaminergic neurons and microglial cells. However, it is
known that in non-inflammatory cells, such as neurons, the
NADPH complex produces only low rates of reactive oxygen spe-
cies for signalling function. In inflammatory cells such as microglia,
NADPH activation produces high concentrations of reactive
oxygen species that are released extracellularly to kill invading
microorganisms or cells (Babior, 1999, 2004). In accordance with
this, we observed that angiotensin was not able to increase dopa-
minergic neuron death in the absence of microglial cells
(Rodriguez-Pallares et al., 2008; Joglar et al., 2009). The increased
angiotensin converting enzyme activity in menopausal rats leads to
increased angiotensin production. Increased angiotensin convert-
ing enzyme activity has also been observed in other tissues in
postmenopausal females (Nogawa et al., 2001) and in experimen-
tal models of cardiovascular disease after ovariectomy (Wu et al.,
2003b; Dean et al., 2005). The observed upregulation of AT1
receptors may also contribute to NADPH activation and increased
dopaminergic cell vulnerability. This is supported by the present
experiments in which we have observed that the enhanced sus-
ceptibility of dopaminergic neurons was significantly decreased by
AT1 receptor inhibition with candesartan. Furthermore, it is par-
ticularly interesting that untreated surgically menopausal rats had
significantly fewer AT2 receptors than young rats treated with
Figure 5 Real-time quantitative reverse transcriptase–poly-
merase chain reaction (A) and western blot (B) analysis of
changes induced by treatment with 17-b-oestradiol (E2) in the
expression of AT1, AT2, p47phox and IL-1b, and the activity of
the angiotensin converting enzyme (ACE; B) in aged ovariecto-
mized rats (aged + ovx + E2) in comparison with intact aged
menopausal rats (aged + E2). No significant differences were
observed between both groups of aged menopausal rats.
Protein expression was obtained as relative to the GAPDH band
value and the expression of each gene was obtained relative to
the housekeeping transcripts (b-actin). The results were then
normalized to the values for intact aged rats (100%). Data are
means � SEM; n = 5.
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 133
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021
oestrogen, which may also enhance dopaminergic cell loss. AT1
and AT2 receptors have opposing effects and AT2 receptors coun-
terbalance the deleterious effect of AT1 receptor stimulation, so
that functional interactions between the two receptor subtypes
and their specific distribution determine the angiotensin-induced
effects (Sohn et al., 2000), which in the case of untreated meno-
pausal rats resulted in a pro-oxidative and pro-inflammatory state,
as suggested by the increased NADPH activity and IL-1b expres-
sion in comparison with rats treated with oestrogen. The detailed
mechanisms involved in the above described regulation of
renin-angiotensin system components by oestrogen have not
been clarified for the brain or peripheral tissues. Oestrogen has
been suggested to inhibit the angiotensin-induced NADPH activa-
tion and production of reactive oxygen species (Xue et al., 2007,
2008), to inhibit AT1 receptor-mediated extracellular
signal-regulated kinase activation (Liu et al., 2002), and to inhibit
angiotensin-induced activation of mitogen-activated protein kin-
ases (Imanishi et al., 2005) in different tissues.
In the present study, it is particularly interesting that the re-
sponse to oestrogen therapy was different in young (i.e. surgically
induced menopause) and aged menopausal rats. The effects of
treatment with oestrogen on AT1, AT2 receptor expression and
NADPH activity were lower in aged rats than in young meno-
pausal rats. In accordance with this, oestrogen therapy signifi-
cantly reduced 6-hydroxydopamine-induced dopaminergic cell
loss in young rats but not in aged rats. Oestrogen regulation of
AT2 levels appears particularly important (Okumura et al., 2005;
Chakrabasty et al., 2008; Sakata et al., 2009). AT2 receptors
are particularly abundant in tissues during development and de-
crease after birth, although they are upregulated under patho-
logical conditions to counter-regulate pro-inflammatory effects
mediated by AT1 receptors. The present results suggest that
Figure 6 Real-time quantitative reverse transcriptase–polymerase chain reaction (A and C) and western blot (B and D) analysis of changes
induced by treatment with the AT1 antagonist candesartan in the expression of AT2, p47phox and IL-1b, and the activity of the angiotensin
converting enzyme (ACE; B and D) in young ovariectomized rats (ovx + candesartan; A and B) and aged acyclic rats (aged + candesartan;
C and D) in comparison with the corresponding untreated rats (ovx or aged rats). Protein expression was obtained relative to the GAPDH
band value and the expression of each gene was obtained relative to the housekeeping transcripts (b-actin). The results were then
normalized to the values for ovariectomized (A and B) or aged (C and D) rats (100%). Data are means � SEM; n = 6. *P50.05 (Student’s
t-test).
134 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

ageing disrupts the ability of the cell to respond to oestrogen by
upregulation of AT2 receptors (Armando et al., 2002; Suarez
et al., 2004). Interestingly, the results observed in rats subjected
to delayed oestrogen therapy indicate that the treatment is able to
reverse changes induced by a period of hypo-oestrogenicity
in young animals (i.e. the increase in renin-angiotensin system
activity, and in markers of oxidative stress, neuroinflam-
mation and dopaminergic neuron vulnerability). However, the
oestrogen-induced upregulation of AT2 receptor expression was
significantly lower than in young ovariectomized rats treated im-
mediately with oestrogen therapy. It is therefore possible that very
long periods of hypo-oestrogenicity in young animals and/or
delayed oestrogen therapy in older rats decreases the neuropro-
tective effects of oestrogen therapy.
Different responses to oestrogen therapy in young menopausal
rats and aged rats have also been observed in other experimental
models. Thus, oestrogen therapy has been shown to protect
against other diseases such as atherosclerosis, although only
when the vessels are healthy and not when atherosclerosis is al-
ready established (Clarkson, 2007, 2009; Antonicelli et al., 2008).
However, the mechanism responsible for the different response to
oestrogen therapy in young and aged animals (i.e. surgical and
natural menopause) has not been clarified. In the present study,
young ovariectomized rats and aged rats treated with oestrogen
did not show significant differences in blood levels of oestrogen,
which were similar to those previously observed with similar im-
plants (Febo et al., 2002; Sandoval and Witt, 2011), and similar to
those observed in intact rats in proestrous (Nequin et al., 1979).
The above reported differences are therefore due to differences in
the response to oestrogen between young and aged rats. Several
recent studies have reported that significant levels of androgens
are produced in the ovaries of aged rats (Fogle et al., 2008; Korse
et al., 2009), but not in young ovariectomized rats (surgically
induced menopause), and it has been suggested that administra-
tion of androgens induce activation of renin-angiotensin system in
several tissues (Fisher et al., 2002; Henriques et al., 2008; Ojeda
et al., 2010). Androgens derived from aged ovaries may therefore
counteract the inhibitory effect of oestrogen therapy on the nigral
renin-angiotensin system. However, we have observed that
oestrogen therapy did not have significantly different effects in
aged ovariectomized rats and intact aged rats. Furthermore,
renin-angiotensin system activity has also been found to be
higher in aged males than in young males (Villar-Cheda et al.,
2012). Interestingly in the case of the dopaminergic system, it
has been shown that loss of oestrogen decreases striatal dopamin-
ergic levels and oestrogen administration increases dopaminergic
release (Becker, 1999; Ohtani et al., 2001; Shulman, 2002; Liu
and Dluzen, 2007), and that there is a counter-regulatory mech-
anism between the dopaminergic and renin-angiotensin system
(Gildea, 2009; Villar-Cheda et al., 2010), which may also explain
the increased renin-angiotensin system activity in menopausal rats
and the decrease in renin-angiotensin system activity by oestrogen
therapy. Decreased dopaminergic levels have also been observed
in aged male animals (i.e. independently of oestrogen depletion;
McCormack et al, 2004; Cruz-Muros et al., 2009), which may
also induce increased renin-angiotensin system activity in males.
However, different responses to oestrogen in young and aged
animals have also been observed in other tissues (independently
of dopaminergic levels; see above) and decreased levels of dopa-
mine with age is not the only factor involved.
It has been suggested that prolonged hypo-oestrogenicity dis-
rupts the ability of injured tissue to upregulate oestrogen receptors
that mediate the anti-inflammatory and neuroprotective actions of
oestrogen, thus eliminating the beneficial effects of oestrogen
treatment (Suzuki et al., 2007). However, it has also been sug-
gested that a prolonged period of oestrogen deprivation may
upregulate oestrogen receptors in senescent females, and exogen-
ous oestrogen therapy may be ineffective or neurotoxic when
acting on supraphysiological receptor substrates (Bake et al.,
2008; Selvamani and Sohrabji, 2010). The present results in
young ovariectomized rats subjected to delayed oestrogen therapy
reveal that a prolonged hypo-oestrogenic state may modify the
effects of oestrogen on renin-angiotensin system activity, particu-
larly the oestrogen-induced upregulation of AT2 receptors.
However, several studies in different tissues suggest that pro-
longed hypo-oestrogenicity is not the only factor responsible for
the different responses to oestrogen therapy in young and aged
rats. Normal ageing is associated with a proinflammatory,
pro-oxidant state that may favour an exaggerated response to
injury and degenerative diseases, in males as well as in females
(Csiszar et al., 2003; Ungvari et al., 2004; Choi et al., 2008);
particularly, we have observed increased renin-angiotensin
system activity in the substantia nigra of aged males in comparison
with young males (i.e. independently of oestrogen levels or
hypo-oestrogenic periods; Villar-Cheda et al., 2012). Factors
other than oestrogen are therefore involved in the upregulation
of renin-angiotensin system in aged rats, an effect that cannot be
counteracted by oestrogen therapy alone.
Parkinson’s disease is usually considered a multifactorial process
in which low and apparently non-toxic doses of several pathogenic
factors can act synergistically to cross the threshold of dopamin-
ergic cell degeneration (Gao et al., 2003), and oxidative stress and
inflammation play major roles in the synergistic process (Wu et al.,
2003a; Andersen, 2004). A pro-oxidative and proinflammatory
state in both groups of menopausal rats may therefore be related
to the increased risk of Parkinson’s disease reported in menopausal
females in several epidemiological studies (Currie et al., 2004;
Ragonese et al., 2006a, b). Furthermore, untreated surgical or
natural menopause may increase the progression of the dopamin-
ergic degeneration in patients with initial or moderate stages of
Parkinson’s disease.
In conclusion, the results suggest that oestrogen constitutes a
major factor for inhibition of renin-angiotensin system activity in
young females. The loss of oestrogen after ovariectomy (surgical
menopause) increases renin-angiotensin system activity and the
susceptibility to neuroinflammation oxidative damage and dopa-
minergic neuron neurodegeneration, and oestrogen therapy in-
duces pronounced renin-angiotensin system inhibition and
neuroprotection. In aged animals, other factors in addition to
the lack of oestrogen contribute to the increase in
renin-angiotensin system activity, and oestrogen therapy can
only partially counteract the renin-angiotensin system hyperactiv-
ity, leading to no significant neuroprotection in the present experi-
mental conditions. In addition, changes in oestrogen receptors or
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 135
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

oestrogen-induced intracellular signalling with ageing may also
lead to partial or ineffective renin-angiotensin system inhibition
by oestrogen therapy in aged animals. In any case, it is
particularly interesting that direct manipulation of the
renin-angiotensin system with the AT1 antagonist candesartan
led to remarkable reduction in renin-angiotensin system activity
(increased either by the lack of oestrogen in young surgically
induced menopausal rats or by the lack of oestrogen and addition-
al factors in aged rats) and significantly protected against dopa-
minergic neuron loss in both types of menopausal rats. This
suggests that manipulation of the brain renin-angiotensin system
may be an efficient approach for the prevention or treatment of
Parkinson’s disease in oestrogen-deficient females together with or
instead of oestrogen replacement therapy.
AcknowledgementsThe authors thank Pilar Aldrey, Iria Novoa and Jose A. Trillo for
their excellent technical assistance. The authors are thankful to
AstraZeneca for providing candesartan for the experiments.
FundingSpanish Ministry of Science and Innovation; Spanish Ministry of
Health (RD06/0010/0013, TERCEL and CIBERNED); Galician
Government (XUGA) and European Regional Development Fund.
ReferencesAndersen JK. Oxidative stress in neurodegeneration: cause or conse-
quence. Nat Med 2004; 10: S18–25.
Antonicelli R, Olivieri F, Morichi V, Urbani E, Mais V. Prevention of
cardiovascular events in early menopause: a possible role for hormone
replacement therapy. Int J Cardiol 2008; 130: 140–6.Armando I, Jezova M, Juorio AV, Terron JA, Falcon-Neri A, et al.
Estrogen upregulates renal angiotensin II AT(2) receptors. Am J
Physiol Renal Physiol 2002; 283: F934–43.
Babior B. NADPH oxidase: an update. Blood 1999; 93: 1464–76.
Babior BM. NADPH oxidase. Curr Opin Immunol 2004; 16: 42–7.Bake S, Ma L, Sohrabji F. Estrogen receptor-alpha overexpression sup-
presses 17beta-estradiol-mediated vascular endothelial growth factor
expression and activation of survival kinases. Endocrinology 2008; 149:
3881–9.
Basso N, Paglia N, Stella I, de Cavanagh EM, Ferder L, del Rosario Lores
Arnaiz M, et al. Protective effect of the inhibition of the
renin-angiotensin system on aging. Regul Pept 2005; 128: 247–52.
Becker JB. Gender differences in dopaminergic function in striatum and
nucleus accumbens. Pharmacol Biochem Behav 1999; 64: 803–12.
Benigni A, Corna D, Zoja C, Sonzogni A, Latini R, Salio M, et al.
Disruption of the Ang II type 1 receptor promotes longevity in mice.
J Clin Invest 2009; 119: 524–30.
Cai H, Griendling KK, Harrison DG. The vascular NAD(P)H oxidases as
therapeutic targets in cardiovascular diseases. Trends Pharmacol Sci
2003; 24: 471–8.Chakrabarty A, Blacklock A, Svojanovsky S, Smith PG. Estrogen elicits
dorsal root ganglion axon sprouting via a renin-angiotensin system.
Endocrinology 2008; 149: 3452–60.
Chen J, Yang S, Hu S, Choudhry MA, Bland KI, Chaudry IH. Estrogen
prevents intestinal inflammation after trauma-hemorrhage via
downregulation of angiotensin II and angiotensin II subtype I receptor.
Am J Physiol Gastrointest Liver Physiol 2008; 295: G1131–7.
Chlebowski RT, Anderson GL, Gass M, Lane DS, Aragaki AK, Kuller LH,
et al. Estrogen plus progestin and breast cancer incidence and mortal-
ity in postmenopausal women. JAMA 2010; 304: 1684–92.
Choi DY, Zhang J, Bing G. Aging enhances the neuroinflammatory re-
sponse and alpha-synuclein nitration in rats. Neurobiol Aging 2010;
31: 1649–53.
Clarkson TB. Estrogen effects on arteries vary with stage of reproductive
life and extent of subclinical atherosclerosis progression. Menopause
2007; 14: 373–84.Clarkson TB, Appt SE. Controversies about HRT–lessons from monkey
models. Maturitas 2005; 51: 64–74.Clarkson TB, Mehaffey MH. Coronary heart disease of females: les-
sons learned from nonhuman primates. Am J Primatol 2009; 71:
785–93.
Cruz-Muros I, Afonso-Oramas D, Abreu P, Perez-Delgado MM,
Rodrıguez M, Gonzalez-Hernandez T. Aging effects on the dopamine
transporter expression and compensatory mechanisms. Neurobiol
Aging 2009; 30: 973–86.
Csiszar A, Ungvari Z, Koller A, Edwards JG, Kaley G. Aging-induced
proinflammatory shift in cytokine expression profile in coronary
arteries. FASEB J 2003; 17: 1183–5.Currie LJ, Harrison MB, Trugman JM, Bennett JP, Wooten GF.
Postmenopausal estrogen use affects risk for Parkinson disease. Arch
Neurol 2004; 61: 886–8.
Dean SA, Tan J, O’Brien ER, Leenen FH. 17beta-estradiol downregulates
tissue angiotensin-converting enzyme and ANG II type 1 receptor in
female rats. Am J Physiol Regul Integr Comp Physiol 2004; 288:
R759–66.
Dluzen D. Estrogen decreases corpus striatal neurotoxicity in response to
6-hydroxydopamine. Brain Res 1997; 767: 340–4.
Dziuk PJ, Cook B. Passage of steroids through silicone rubber.
Endocrinology 1966; 78: 208–11.
Febo M, Ferris CF, Segarra AC. Estrogen influences cocaine-induced
blood oxygen level-dependent signal changes in female rats.
J Neurosci 2005; 25: 1132–6.Febo M, Jimenez-Rivera CA, Segarra AC. Estrogen and opioids interact
to modulate the locomotor response to cocaine in the female rat. Brain
Res 2002; 943: 151–61.
Fischer M, Baessler A, Schunkert H. Renin angiotensin system and
gender differences in the cardiovascular system. Cardiovasc Res
2002; 53: 672–7.
Fogle RH, Stanczyk FZ, Zhang X, Paulson RJ. Ovarian androgen produc-
tion in postmenopausal women. J Clin Endocrinol Metab 2007; 92:
3040–3.
Gao HM, Liu B, Zhang W, Hong JS. Synergistic dopaminergic neurotox-
icity of MPTP and inflammogen lipopolysaccharide: relevance to the
etiology of Parkinson’s disease. FASEB J 2003; 17: 1957–9.Gildea JJ. Dopamine and angiotensin as renal counterregulatory systems
controlling sodium balance. Curr Opin Nephrol Hypertens 2009; 18:
28–32.
Gundersen HJG. Bendsen, T.F., Korbo, L., Marcussen, N., Moller, A.,
Nielsen, K., Some new, simple and efficient stereological methods
and their use in pathological research and diagnosis. APIMS 1988;
96: 379–94.
Harman SM, Naftolin F, Brinton EA, Judelson DR. Is the estrogen con-
troversy over? Deconstructing the Women’s Health Initiative study: a
critical evaluation of the evidence. Ann NY Acad Sci 2005; 1052:
43–56.
Hemming ML, Selkoe DJ, Farris W. Effects of prolonged
angiotensin-converting enzyme inhibitor treatment on amyloid
b-protein metabolism in mouse models of Alzheimer disease.
Neurobiol Dis 2007; 26: 273–81.
Henriques T, Zhang X, Yiannikouris FB, Daugherty A, Cassis LA.
Androgen increases AT1a receptor expression in abdominal aortas to
promote angiotensin II-induced AAAs in apolipoprotein E-deficient
mice. Arterioscler Thromb Vasc Biol 2008; 28: 1251–6.
136 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

Heymes C, Silvestre JS, Llorens-Cortes C, Chevalier B, Marotte F,
Levy BI, et al. Cardiac senescence is associated with enhanced expres-
sion of angiotensin II receptor subtypes. Endocrinology 1998; 139:
2579–87.
Hoshi-Fukushima R, Nakamoto H, Imai H, Kanno Y, Ishida Y,
Yamanouchi Y, et al. Estrogen and angiotensin II interactions deter-
mine cardio-renal damage in Dahl salt-sensitive rats with heart failure.
Am J Nephrol 2008; 28: 413–23.Imanishi T, Hano T, Nishio I. Estrogen reduces angiotensin II-induced
acceleration of senescence in endothelial progenitor cells. Hypertens
Res 2005; 28: 263–71.Joglar B, Rodriguez-Pallares J, Rodrıguez-Perez AI, Rey P, Guerra MJ,
Labandeira-Garcia JL. The inflammatory response in the MPTP
model of Parkinson’s disease is mediated by brain angiotensin: rele-
vance to progression of the disease. J Neurochem 2009; 109: 656–69.
Johnson AB, Bake S, Lewis DK, Sohrabji F. Temporal expression of
IL-1beta protein and mRNA in the brain after systemic LPS injection
is affected by age and estrogen. J Neuroimmunol 2006; 174: 82–91.
Kondo T, Yoshida K, Yoshimura Y, Motohashi M, Tanayama S.
Disposition of the new angiotensin II receptor antagonist candesartan
cilexetil in rats and dogs. Arzneimittelforschung 1996; 46: 594–600.
Korse CM, Bonfrer JM, van Beurden M, Verheijen RH, Rookus MA.
Estradiol and testosterone levels are lower after oophorectomy than
after natural menopause. Tumour Biol 2009; 30: 37–42.
Krishnamurthi K, Verbalis JG, Zheng W, Wu Z, Clerch LB, Sandberg K.
Estrogen regulates angiotensin AT1 receptor expression via cytosolic
proteins that bind to the 50 leader sequence of the receptor mRNA.
Endocrinology 1999; 140: 5435–8.Li JM, Shah AM. Mechanism of endothelial cell NADPH oxidase activa-
tion by angiotensin II. Role of the p47phox subunit. J Biol Chem 2003;
278: 12094–100.Liu B, Dluzen DE. Oestrogen and nigrostriatal dopaminergic neurodegen-
eration: animal models and clinical reports of Parkinson’s disease. Clin
Exp Pharmacol Physiol 2007; 34: 555–65.
Liu HW, Iwai M, Takeda-Matsubara Y, Wu L, Li JM, Okumura M, et al.
Effect of estrogen and AT1 receptor blocker on neointima formation.
Hypertension 2002; 40: 451–7.
Mannino CA, South SM, Inturrisi CE, Quinones-Jenab V.
Pharmacokinetics and effects of 17beta-estradiol and progesterone
implants in ovariectomized rats. J Pain 2005; 6: 809–16.
McCormack AL, Di Monte DA, Delfani K, Irwin I, DeLanney LE,
Langston WJ, et al. Aging of the nigrostriatal system in the squirrel
monkey. J Comp Neurol 2004; 471: 387–95.
McKinley MJ, Albiston AL, Allen AM, Mathai ML, May CN,
McAllen RM, et al. The brain reenin-angiotensin system: location
and physiological roles. Int J Biochem Cell Biol 2003; 35: 901–18.
Miller VM, Black DM, Brinton EA, Budoff MJ, Cedars MI, Hodis HN,
et al. Using basic science to design a clinical trial: baseline character-
istics of women enrolled in the Kronos Early Estrogen Prevention Study
(KEEPS). J Cardiovasc Transl Res 2009; 2: 228–39.Miller VM, Clarkson TB, Harman SM, Brinton EA, Cedars M, Lobo R,
et al. Women, hormones, and clinical trials: a beginning, not an end.
J Appl Physiol 2005; 99: 381–3.Morale MC, Serra PA, L’episcopo F, Tirolo C, Caniglia S, Testa N, et al.
Estrogen, neuroinflammation and neuroprotection in Parkinson’s dis-
ease: glia dictates resistance versus vulnerability to neurodegeneration.
Neuroscience 2006; 138: 869–78.
Murray HE, Pillai AV, McArthur SR, Razvi N, Datla KP, Dexter DT,
Gillies GE. Dose- and sex-dependent effects of the neurotoxin
6-hydroxydopamine on the nigrostriatal dopaminergic pathway of
adult rats: differential actions of estrogen in males and females.
Neuroscience 2003; 116: 213–22.Nequin LG, Alvarez J, Schwartz NB. Measurement of serum steroid and
gonadotropin levels and uterin and ovarian variables throughout 4-day
and 5-day estrous cycles in the rat. Biol Reprod 1979; 20: 659–70.Nickenig G, Baumer AT, Grohe C, Kahlert S, Strehlow K, Rosenkranz S,
et al. Estrogen modulates AT1 receptor gene expression in vitro and
in vivo. Circulation 1998; 97: 2197–201.
Nogawa N, Sumino H, Ichikawa S, Kumakura H, Takayama Y,
Nakamura T, et al. Effect of long-term hormone replacement therapy
on angiotensin-converting enzyme activity and bradykinin in postme-
nopausal women with essential hypertension and normotensive post-
menopausal women. Menopause 2001; 8: 210–5.
Ohtani H, Nomoto M, Douchi T. Chronic estrogen treatment replaces
striatal dopaminergic function in ovariectomized rats. Brain Res 2001;
900: 163–8.
Ojeda NB, Royals TP, Black JT, Dasinger JH, Johnson JM, Alexander BT.
Enhanced sensitivity to acute angiotensin II is testosterone dependent
in adult male growth-restricted offspring. Am J Physiol Regul Integr
Comp Physiol 2010; 298: R1421–7.
Okumura M, Iwai M, Ide A, Mogi M, Ito M, Horiuchi M. Sex difference
in vascular injury and the vasoprotective effect of valsartan are related
to differential AT2 receptor expression. Hypertension 2005; 46:
577–83.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. New
York: Academic Press; 1985.
Przedborski S, Levivier M, Jiang H, Ferreira M, Jackson-Lewis V,
Donaldson D, et al. Dose-dependent lesions of the dopaminergic
nigrostriatal pathway induced by intrastriatal injection of
6-hydroxydopamine. Neuroscience 1995; 67: 631–47.Qin L, Liu Y, Wang T, Wei SJ, Block ML, Wilson B, et al. NADPH oxidase
mediates lipopolysaccharide-induced neurotoxicity and proinflamma-
tory gene expression in activated microglia. J Biol Chem 2004; 279:
1415–21.Ragonese P, D’Amelio M, Callari G, Salemi G, Morgante L, Savettieri G.
Age at menopause predicts age at onset of Parkinson’s disease. Mov
Disord 2006a; 21: 2211–4.Ragonese P, D’Amelio M, Savettieri G. Implications for estrogens in
Parkinson’s disease: an epidemiological approach. Ann NY Acad Sci
2006b; 1089: 373–82.Rey P, Lopez-Real A, Sanchez-Iglesias, Munoz A, Soto-Otero R,
Labandeira-Garcia JL. Angiotensin type-1-receptor antagonists reduce
6-hydroxydopamine toxicity for dopaminergic neurons. Neurobiol
Aging 2007; 28: 555–67.
Rodriguez-Pallares J, Parga JA, Munoz A, Rey P, Guerra MJ, Labandeira-
Garcia JL. Mechanism of 6-hydroxydopamine neurotoxicity: the role of
NADPH oxidase and microglial activation in 6-hydroxydopamine-
induced degeneration of dopaminergic neurons. J Neurochem 2007;
103: 145–56.
Rodriguez-Pallares J, Rey P, Parga JA, Munoz A, Guerra MJ, Labandeira-
Garcia JL. Brain angiotensin enhances dopaminergic cell death via
microglial activation and NADPH-derived ROS. Neurobiol Dis 2008;
31: 58–73.
Rodriguez-Perez AI, Valenzuela R, Villar-Cheda B, Guerra MJ,
Lanciego JL, Labandeira-Garcia JL. Estrogen and angiotensin inter-
action in the substantia nigra. Relevance to postmenopausal
Parkinson’s diseas. Exp Neurol 2010; 224: 517–26.
Rossouw JE, Anderson GL, Prentice RL, LaCroix AZ, Kooperberg C,
Stefanick ML, et al. Risks and benefits of estrogen plus progestin in
healthy postmenopausal women: principal results From the Women’s
Health Initiative randomized controlled trial. JAMA 2002; 288:
321–33.Rueckschloss U, Quinn MT, Holtz J, Morawietz H. Dose-dependent
regulation of NAD(P)H oxidase expression by angiotensin II in
human endothelial cells: protective effect of angiotensin II type 1 re-
ceptor blockade in patients with coronary artery disease. Arterioscler
Thromb Vasc Biol 2002; 22: 1845–51.
Saavedra JM. Brain angiotensin II: new developments, unanswered ques-
tions and therapeutic opportunities. Cell Mol Neurobiol 2005; 25:
485–12.
Sakata A, Mogi M, Iwanami J, Tsukuda K, Min LJ, Fujita T, et al.
Sex-different effect of angiotensin II type 2 receptor on ischemic
brain injury and cognitive function. Brain Res 2009; 1300: 14–23.
Sandoval KE, Witt KA. Age and 17b-estradiol effects on blood–brain
barrier tight junction and estrogen receptor proteins in ovariectomized
rats. Microvascular Research 2011; 81: 198–205.
Menopause, angiotensin and dopaminergic degeneration Brain 2012: 135; 124–138 | 137
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021

Sauer H, Oertel WH. Progressive degeneration of nigrostriatal dopamineneurons following intrastriatal terminal lesions with
6-hydroxydopamine: a combined retrograde tracing and immunohis-
tochemical study in the rat. Neuroscience 1994; 59: 401–15.
Selvamani A, Sohrabji F. Reproductive age modulates the impact of focalischemia on the forebrain as well as the effects of estrogen treatment
in female rats. Neurobiol Aging 2010; 31: 1618–28.
Seshiah PN, Weber DS, Rocic P, Valppu L, Taniyama Y, Griendling KK.
Angiotensin II stimulation of NAD(P)H oxidase activity: upstream me-diators. Circ Res 2002; 91: 406–13.
Shulman LM. Is there a connection between estrogen and Parkinson’s
disease? Parkinsonism Relat Disord 2002; 8: 289–95.Sohn HY, Raff U, Hoffmann A, Gloe T, Heermeier K, Galle J, et al.
Differential role of angiotensin II receptor subtypes on endothelial
superoxide formation. Br J Pharmacol 2000; 131: 667–72.
Sohrabji F, Bake S. Age-related changes in neuroprotection: is estrogenpro-inflammatory for the reproductive senescent brain? Endocrine
2006; 29: 191–7.
Sohrabji F. Estrogen: a neuroprotective or proinflammatory hormone?
Emerging evidence from reproductive aging models. Ann NY AcadSci 2005; 1052: 75–9.
Suarez C, Dıaz-Torga G, Gonzalez-Iglesias A, Cristina C, Becu-
Villalobos D. Upregulation of angiotensin II type 2 receptor expression
in estrogen-induced pituitary hyperplasia. Am J Physiol EndocrinolMetab 2004; 286: E786–94.
Suzuki S, Brown CM, Dela Cruz CD, Yang E, Bridwell DA, Wise PM.
Timing of estrogen therapy after ovariectomy dictates the efficacy ofits neuroprotective and antiinflammatory actions. Proc Natl Acad Sci
USA 2007; 104: 6013–8.
Touyz RM, Chen X, Tabet F, Yao G, He G, Quinn MT, et al. Expression
of a functionally active gp91phox-containing neutrophil-type NAD(P)Hoxidase in smooth muscle cells from human resistance arteries, regu-
lation by angiotensin II. Circ Res 2002; 14: 1205–13.
Tripanichkul W, Sripanichkulchai K, Finkelstein DI. Estrogen
down-regulates glial activation in male mice following1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine intoxication. Brain Res
2006; 1084: 28–37.
Tsuda M, Iwai M, Li JM, Li HS, Min LJ, Ide A, et al. Inhibitory effects ofAT1 receptor blocker, olmesartan, and estrogen on atherosclerosis via
anti-oxidative stress. Hypertension 2005; 45: 545–51.
Turgeon JL, Carr MC, Maki PM, Mendelsohn ME, Wise PM. Complex
actions of sex steroids in adipose tissue, the cardiovascular system, andbrain: insights from basic science and clinical studies. Endocr Rev 2006;
27: 575–605.
Turgeon JL, McDonnell DP, Martin KA, Wise PM. Hormone therapy:physiological complexity belies therapeutic simplicity. Science 2004;
304: 1269–73.
Unger T. Inhibiting angiotensin receptors in the brain: possible thera-
peutic implications. Curr Med Res Opin 2003; 19: 449–51.Ungvari Z, Csiszar A, Kaley G. Vascular inflammation in aging. Herz
2004; 29: 733–40.
Vegeto E, Benedusi V, Maggi A. Estrogen anti-inflammatory activity in
brain: a therapeutic opportunity for menopause and neurodegenera-tive diseases. Front Neuroendocrinol 2008; 29: 507–19.
Villar-Cheda B, Rodrıguez-Pallares J, Muno A, Valenzuela R, Guerra MJ,
Baltatu OC, Labandeira-Garcia JL. Nigral and striatal regulation ofangiotensin receptor expression by dopamine and angiotensin in
rodents: implications for progression of Parkinson’s disease. Eur J
Neurosci 2010; 32: 1695–706.
Villar-Cheda B, Valenzuela R, Rodriguez-Perez AI, Guerra MJ,Labandeira-Garcia JL. Aging-related changes in the nigral angiotensin
system enhances proinflammatory and pro-oxidative markers and
6-hydroxydopamine-induced dopaminergic degeneration. Neurobiol
Aging 2012; 33: 204.e1–204.e11.Wu D, Teisman P, Tieu K, Vila M, Jackson-Lewis V, Ischiropoulos H,
et al. NADPH oxidase mediates oxidative stress in the
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson’s
disease. Proc Natl Acad Sci USA 2003a; 100: 6145–50.Wu DC, Jackson-Lewis V, Vila M, Tieu K, Teismann P, Vadseth C, et al.
Blockade of microglial activation is neuroprotective in the
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model ofParkinson disease. J Neurosci 2002; 22: 1763–71.
Wu Z, Zheng W, Sandberg K. Estrogen regulates adrenal angiotensin
type 1 receptors by modulating adrenal angiotensin levels.
Endocrinology 2003b; 144: 1350–6.Xue B, Johnson AK, Hay M. Sex differences in angiotensin II- induced
hypertension. Braz J Med Biol Res 2007a; 40: 727–34.
Xue B, Pamidimukkala J, Lubahn DB, Hay M. Estrogen receptor-alpha
mediates estrogen protection from angiotensin II-induced hypertensionin conscious female mice. Am J Physiol Heart Circ Physiol 2007b; 292:
H1770–6.
Xue B, Zhao Y, Johnson AK, Hay M. Central estrogen inhibition of angio-tensin II-induced hypertension in male mice and the role of reactive
oxygen species. Am J Physiol Heart Circ Physiol 2008; 295: H1025–32.
Zorad S, Dou J, Benicky J, Hutanu D, Tybitanclova K, Zhou J,
Saavedra JM. Long-term angiotensin II AT1 receptor inhibition pro-duces adipose tissue hypotrophy accompanied by increased expression
of adiponectin and PPAR�. Eur J Pharmacol 2006; 552: 112–22.
138 | Brain 2012: 135; 124–138 A. I. Rodriguez-Perez et al.
Dow
nloaded from https://academ
ic.oup.com/brain/article/135/1/124/327607 by guest on 25 N
ovember 2021





























