Embed Size (px)
Citation preview

volunteer group, our data include challenge with 2 differ-ent sources of endotoxin. Although the response to eachendotoxin source was not statistically different, there weredecreased PMNs present in the CCRE versus Sigmagroups, which, when both groups are combined, modestlyincreases the variability of this measure. Because of thisincreased variability and smaller sample size, we recog-nize that a larger sample size may have revealed astatistically significant correlation that we did not observefor the normal cohort. Second, the group with asthma hadincreased PMN responses relative to normal volunteers,but these differences were not statistically significant.Third, our study population was predominantly female,and it is possible that female sex may be important inasthma and response to pollutants. Finally, it is importantto note that all of the patients with asthma studied inthese protocols had mild intermittent disease and requiredno chronic controller medication. Nonetheless, withinthe asthma group (all of whom underwent challenge withthe same source of endotoxin), we observed a significantcorrelation between endotoxin induced increases in air-way inflammation and BMI.
To our knowledge, this is the first report examining arelationship between BMI and inflammatory response toinhaled endotoxin in asthma. Endotoxin has been linked toasthma severity in ambient air and domestic and occupa-tional environments and is a good model for acute eventsassociated with asthma exacerbation. There are a numberof potential mechanisms by which BMI may be linked tothe inflammatory response to inhaled endotoxin in asthma.We have reported that allergic airway inflammation maymodify mCD14 expression in airway macrophages andmonocytes, which may enhance their response to endo-toxin. It is important to note, however, that without aller-gic and nonatopic controls with asthma, it is presentlyunclear whether atopy, asthma, or the combination ofthe 2 is responsible for the differences we observedbetween our study cohorts. Our group has also observedthat even with modest changes in BMI, there is increasedparticle deposition in the airways. Thus, in patients withasthma, increased deposition of endotoxin related tobody mass may allow an increased effective airway doseof endotoxin to interact with primed monocytes and mac-rophages. It is also possible that increased BMI reflectschanges in inflammatory biology reported to occur inobesity, including increased leptin, which has recentlybeen reported to promote eosinophil survival. Thus, obesepatients with asthma may have modified response toendotoxin at a cellular level.
Regardless of the precise mechanism, it is important toconsider the potential interaction between body mass andasthma exacerbation. Obesity likely represents an impor-tant target for intervention in many patients with moderateand severe asthma. Our data demonstrate that in asthma,further examination of both the effect of BMI on airwayphysiology relative to deposition of inhaled bioactiveparticles, as well as the effect of obesity on innate andacquired immune responses, is needed to understand therole of obesity in asthma pathogenesis and exacerbation.
Neil E. Alexis, PhD
David B. Peden, MD
From the Center for Environmental Medicine, Asthma and Lung Biology,
Division of Pediatric Immunology and Infectious Diseases, Department of
Pediatrics, 104 Mason Farm Road, CB#7310, University of North Carolina
School of Medicine, Chapel Hill, NC 27516-7310.
Supported by National Institutes of Health grants RO1-HL62624, RO1-
HL66559, GCRC grant RR-000046, and EPA CR829522.
Disclosure of potential conflict of interest: D. Peden has consultant arrange-
ments with GlaxoSmithKline, Merck, and AstraZeneca, and has received
grants from and is on the speakers bureau for GlaxoSmithKline. N. Alexis
has declared that he has no conflict of interest.
REFERENCES
1. Gilliland FD, Berhane K, Islam T, McConnell R, Gauderman WJ, Gilliland
SS, et al. Obesity and the risk of newly diagnosed asthma in school-age
children. Am J Epidemiol 2003;158:406-15.
2. Shore SA, Fredberg JJ. Obesity, smooth muscle, and airway hyper-
responsiveness. J Allergy Clin Immunol 2005;115:925-7.
3. Weiss ST. Obesity: insight into the origins of asthma. Nat Immunol
2005;6:537-9.
4. Rivera-Sanchez YM, Johnston RA, Schwartzman IN, Valone J, Silverman
ES, Fredberg JJ, et al. Differential effects of ozone on airway and tissue
mechanics in obese mice. J Appl Physiol 2004;96:2200-6.
5. Bernstein JA, Alexis N, Barnes C, Bernstein IL, Bernstein JA, Nel A,
et al. Health effects of air pollution. J Allergy Clin Immunol 2004;
114:1116-23.
6. Bennett WD, Zeman KL. Effect of body size on breathing pattern and
fine particle deposition in children. J Appl Physiol 2004;97:821-6.
7. Alexis N, Eldridge M, Reed W, Bromberg P, Peden DB. CD14-dependent
airway neutrophil response to inhaled LPS: role of atopy. J Allergy Clin
Immunol 2001;107:31-5.
8. Boehlecke B, Hazucha M, Alexis N, Bromberg P, Peden DB. Low dose
endotoxin enhances bronchial responsiveness to inhaled allergen in
atopic asthmatics. J Allergy Clin Immunol 2003;112:1241-3.
9. Alexis NE, Peden DB. Blunting airway eosinophilic inflammation results
in a decreased airway neutrophil response to inhaled LPS in patients
with atopic asthma: a role for CD14. J Allergy Clin Immunol 2001;108:
577-80.
10. Alexis NE, Lay JC, Almond M, Bromberg PA, Patel DD, Peden DB.
Acute LPS inhalation in healthy volunteers induces dendritic cell matu-
ration in vivo. J Allergy Clin Immunol 2005;115:345-50.
Available online February 7, 2006.doi:10.1016/j.jaci.2005.12.1305
J ALLERGY CLIN IMMUNOL
MAY 2006
1186 Letters to the Editor
Lette
rsto
the
Edito
r
Dominance of human innate immuneresponses in primary Francisella tularensislive vaccine strain vaccination
To the Editor:Francisella tularensis is the etiologic agent of the
zoonotic disease, tularemia. An inoculum as small as 10bacteria can cause a flulike disease with substantial mor-bidity and mortality among infected humans.1,2 Humantularemia presents in ulceroglandular, oculoglandular, oro-pharyngeal, pneumonic, and septic forms.1,3 Rapid admin-istration of antibiotics prevents mortality in the majority ofhuman cases if exposure doses are low and nonaerosol.2-4
Without early diagnosis and administration of antibiotics,high-dose aerosol exposure progresses rapidly to life-threatening pleuropneumonitis and systemic infection.3
The relative abundance of F tularensis in nature and the rel-ative ease with which it may be administered raise concernsover its exploitation as a biothreat agent.1,3,4
J ALLERGY CLIN IMMUNOL
VOLUME 117, NUMBER 5
Letters to the Editor 1187
Lett
ers
toth
eEd
itor
In the 1950s, an attenuated strain of F tularensis wasdeveloped into an Investigational New Drug status livevaccine (live vaccine strain [LVS]) administered by intra-dermal scarification.1,5,6 LVS vaccination has signifi-cantly lowered reports of laboratory-acquired tularemia,although the mechanism of protection (humoral or cell-mediated) is still unclear.1,3,7 Microagglutination assaysperformed at 28 days postvaccination (indicating anti–F tularensis IgG and IgM) are the clinical standard forgauging successful vaccination yet show poor correlationwith specific, vigorous lymphocyte responses in LVS-vaccinated and naturally infected humans.8 Human anti–F tularensis immune serum resulting from vaccinationwith LVS is protective only against strains of reducedvirulence, yet immunospecific and long-lasting cell-mediated immunity is the key to protection.5,7,9 Substantialdata suggest that cell-mediated immunity may be moreimportant than humoral immunity for providing long-lasting immunity against virulent strains of F tularensis.5,7,9
Therefore, we examined human immune responses toLVS vaccination to establish early cellular correlates ofLVS-mediated protection predictive of successful vaccineoutcomes.
Volunteers were recruited from US Army MedicalResearch Institute of Infectious Diseases (USAMRIID)personnel at risk of laboratory exposure to F tularensis.A minimal risk protocol to collect peripheral blood sam-ples was approved by institutional review boards at theUSAMRIID (Human Use Committee FY04-16). Donorsprovided informed consent and met eligibility criteria.Six healthy adults (4 males and 2 females, 22-54 yearsold) received a primary LVS vaccination and donatedperipheral blood prevaccination and postvaccination.Mononuclear cells were purified by Ficoll gradients andassessed for changes in immune cell populations usingflow cytometry and quadruple stained using directlyconjugated mAbs (BD-Immunocytometry Systems andBD-Pharmingen, La Jolla, Calif). Cytometric bead arrayanalysis was performed on serum samples (BD-Pharmin-gen). Analysis was performed using FlowJo (TreeStar,Inc, Ashland, Ore) and GraphPad Prism (GraphPad PrismSoftware, San Diego, Calif). All 6 subjects were immuno-logically naive before vaccination. All vaccinations hadpositive responses as indicated by initial formation ofa small pustule/papule and subsequent ulceration.
Cell surface analysis of mononuclear cells revealed biastoward activation of innate versus acquired immunesystem components. The greatest changes in cellularityoccurred on day 11 (P < .001 for day 11 versus all otherdays), a time frame consistent with innate immunity, buttoo short for unprimed acquired responses (Fig 1, A andB). When acquired immune system cells (CD31, CD41,CD81, T-cell receptor [TCR] ab1, CD45RO1) wereanalyzed over all time points, only CD41 and CD81
cells were significantly changed at day 11 (P < .05 andP < .01, respectively).
Although CD41 and CD81 cells increased significantlyon day 11 (Fig 1, A), these cells were not conventionalTCRab1 T cells, but rather TCRgd1 T cells (CD81/
gdTCR1), natural killer (NK) T cells (CD561/CD81),and monocytes (CD41/CD141) whose kinetics mirroredinnate immune responses (Fig 1, B). When innate immunesystem components (CD561, CD1a1, TCRgd1, humanleukocyte antigen [HLA]-DR1, CD161, CD141) weretracked over the course of the vaccination, NK cells,gdT cells, monocytes, granulocytes, and dendritic cellsshowed considerable changes in cellularity on day 11.Dramatic increases in gdT cells (Fig 2, A) and NK cells(Fig 2, B) were noted on day 11 (P < .05 and P < .01,respectively), with NK cells having the most prominentchanges. LVS vaccination induced a strong proliferativesignal for innate lymphocytes as measured by the upregu-lation of the IL-2 receptor high-affinity a chain, CD25(Fig 2, C), whose induction is linked with progressioninto cell cycle.10 CD25 upregulation was highest on day11 for all donors (P < .01) and was noted for NKT,gdT, and NK cells. No similar correlations were seen be-tween CD25 and TCRab1-CD41 or TCRab1-CD81 Tcells, even at time points associated with primary acquiredimmune responses, days 18 to 114.
Our data strongly suggest that cellularity changes afterhuman LVS vaccination strongly parallel innate immunesystem kinetics. These data are in accord with the murineLVS model in which mice deficient in T cells are still ableto resist lethal LVS challenge for 3 to 4 weeks.5 This pro-tection can be traced to proinflammatory cytokine produc-tion by innate immune components, chiefly NK and NKTcells.5 Although serum cytokine levels in our study werebelow the limit of detection (<20 pg/mL), the upregula-tion of the high-affinity IL-2 receptor chain (CD25) mayindicate that a proinflammatory TH1-type response was
FIG 1. Cellularity changes in acquired and innate immune response
to LVS vaccination. Peripheral blood was monitored for changes in
acquired (A) and innate (B) immune cell surface markers over the
course of several days prevaccination and postvaccination (as indi-
cated). Symbols represent the mean percentage positive cells with
n 5 6.
J ALLERGY CLIN IMMUNOL
MAY 2006
1188 Letters to the Editor
Lette
rsto
the
Edito
r
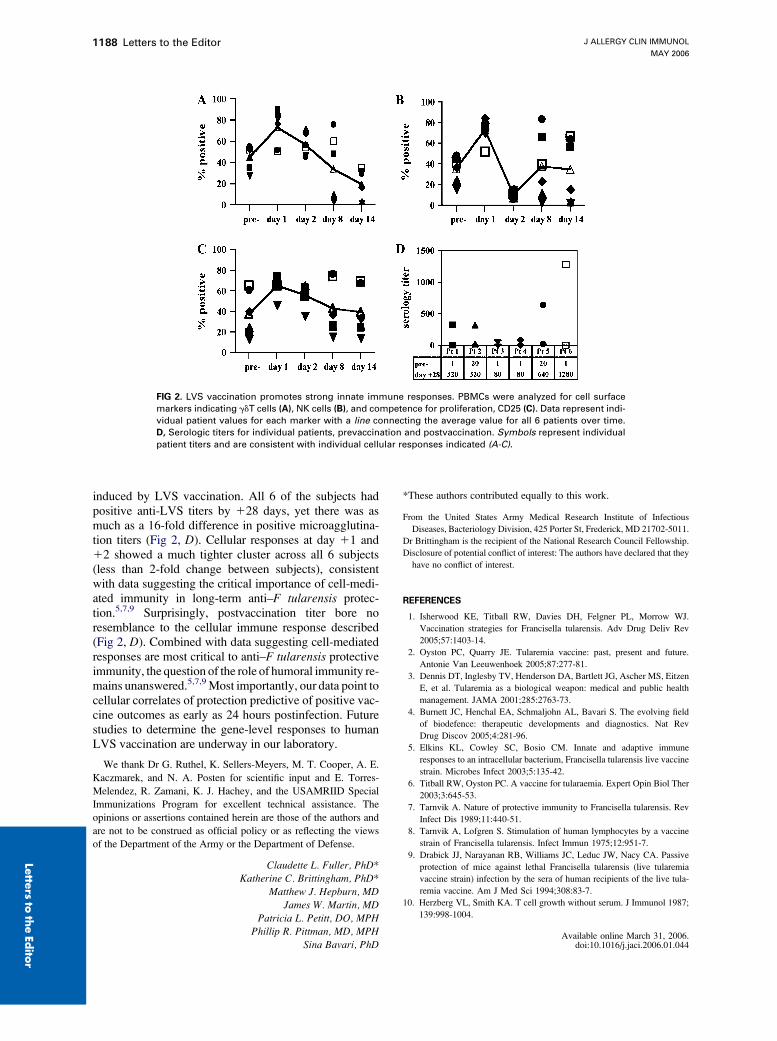
FIG 2. LVS vaccination promotes strong innate immune responses. PBMCs were analyzed for cell surface
markers indicating gdT cells (A), NK cells (B), and competence for proliferation, CD25 (C). Data represent indi-
vidual patient values for each marker with a line connecting the average value for all 6 patients over time.
D, Serologic titers for individual patients, prevaccination and postvaccination. Symbols represent individual
patient titers and are consistent with individual cellular responses indicated (A-C).
induced by LVS vaccination. All 6 of the subjects hadpositive anti-LVS titers by 128 days, yet there was asmuch as a 16-fold difference in positive microagglutina-tion titers (Fig 2, D). Cellular responses at day 11 and12 showed a much tighter cluster across all 6 subjects(less than 2-fold change between subjects), consistentwith data suggesting the critical importance of cell-medi-ated immunity in long-term anti–F tularensis protec-tion.5,7,9 Surprisingly, postvaccination titer bore noresemblance to the cellular immune response described(Fig 2, D). Combined with data suggesting cell-mediatedresponses are most critical to anti–F tularensis protectiveimmunity, the question of the role of humoral immunity re-mains unanswered.5,7,9 Most importantly, our data point tocellular correlates of protection predictive of positive vac-cine outcomes as early as 24 hours postinfection. Futurestudies to determine the gene-level responses to humanLVS vaccination are underway in our laboratory.
We thank Dr G. Ruthel, K. Sellers-Meyers, M. T. Cooper, A. E.
Kaczmarek, and N. A. Posten for scientific input and E. Torres-
Melendez, R. Zamani, K. J. Hachey, and the USAMRIID Special
Immunizations Program for excellent technical assistance. The
opinions or assertions contained herein are those of the authors and
are not to be construed as official policy or as reflecting the views
of the Department of the Army or the Department of Defense.
Claudette L. Fuller, PhD*Katherine C. Brittingham, PhD*
Matthew J. Hepburn, MD
James W. Martin, MD
Patricia L. Petitt, DO, MPHPhillip R. Pittman, MD, MPH
Sina Bavari, PhD
*These authors contributed equally to this work.
From the United States Army Medical Research Institute of Infectious
Diseases, Bacteriology Division, 425 Porter St, Frederick, MD 21702-5011.
Dr Brittingham is the recipient of the National Research Council Fellowship.
Disclosure of potential conflict of interest: The authors have declared that they
have no conflict of interest.
REFERENCES
1. Isherwood KE, Titball RW, Davies DH, Felgner PL, Morrow WJ.
Vaccination strategies for Francisella tularensis. Adv Drug Deliv Rev
2005;57:1403-14.
2. Oyston PC, Quarry JE. Tularemia vaccine: past, present and future.
Antonie Van Leeuwenhoek 2005;87:277-81.
3. Dennis DT, Inglesby TV, Henderson DA, Bartlett JG, Ascher MS, Eitzen
E, et al. Tularemia as a biological weapon: medical and public health
management. JAMA 2001;285:2763-73.
4. Burnett JC, Henchal EA, Schmaljohn AL, Bavari S. The evolving field
of biodefence: therapeutic developments and diagnostics. Nat Rev
Drug Discov 2005;4:281-96.
5. Elkins KL, Cowley SC, Bosio CM. Innate and adaptive immune
responses to an intracellular bacterium, Francisella tularensis live vaccine
strain. Microbes Infect 2003;5:135-42.
6. Titball RW, Oyston PC. A vaccine for tularaemia. Expert Opin Biol Ther
2003;3:645-53.
7. Tarnvik A. Nature of protective immunity to Francisella tularensis. Rev
Infect Dis 1989;11:440-51.
8. Tarnvik A, Lofgren S. Stimulation of human lymphocytes by a vaccine
strain of Francisella tularensis. Infect Immun 1975;12:951-7.
9. Drabick JJ, Narayanan RB, Williams JC, Leduc JW, Nacy CA. Passive
protection of mice against lethal Francisella tularensis (live tularemia
vaccine strain) infection by the sera of human recipients of the live tula-
remia vaccine. Am J Med Sci 1994;308:83-7.
10. Herzberg VL, Smith KA. T cell growth without serum. J Immunol 1987;
139:998-1004.
Available online March 31, 2006.doi:10.1016/j.jaci.2006.01.044









![Clinical Characterization of Aerosolized Francisella ... · Clinical Characterization of Aerosolized . Francisella tularensis. ... and spleen [2]. ... If collection from the CVC was](https://img.pdfslide.us/doc/110x75/5b1c35bd7f8b9a46258f8250/clinical-characterization-of-aerosolized-francisella-clinical-characterization.jpg)



















