Embed Size (px)
Citation preview

D
Sa
b
a
ARRAA
KCCDDET
1
1c
tommt(M(B
Aebdm
s
Hf
1h
DNA Repair 12 (2013) 1152– 1158
Contents lists available at ScienceDirect
DNA Repair
j ourna l ho me p age : www.elsev ier .com/ locate /dnarepai r
NA modifications repaired by base excision repair are epigenetic
tephen P.G. Moorea, Kimberly J. Toomirea, Phyllis R. Straussa,b,∗
Department of Biology, Northeastern University, Boston, MA, 02115, United StatesDepartment of Biology, 134 Mugar Hall, Northeastern University, 360 Huntington Avenue, Boston, MA, 02115, United States
r t i c l e i n f o
rticle history:eceived 23 July 2013eceived in revised form 4 October 2013ccepted 7 October 2013vailable online 9 November 2013
eywords:hromatinREB
a b s t r a c t
CREB controls ∼25% of the mammalian transcriptome. Small changes in binding to its consensus (CRE)sequence are likely to be amplified many fold in initiating transcription. Here we show that DNA lesionsrepaired by the base excision repair (BER) pathway modulate CREB binding to CRE. We generated Kd
values by electrophoretic mobility shift assays using purified human CREB and a 39-mer double-strandedoligonucleotide containing modified or wild-type CRE. CRE contains two guanine residues per strand,one in a CpG islet. Alterations in CRE resulted in positive or negative changes in Kd over two orders ofmagnitude depending on location and modification. Cytosine methylation or oxidation of both guaninesgreatly diminished binding; a G/U mispair in the CpG context enhanced binding. Intermediates in the BER
NA base excision repairNA–protein interactionpigeneticsranscription factors
pathway at one G residue or the other resulted in reduced binding, depending on the specific location,while there was no change in binding when the single G residue outside of the CpG islet was oxidized.CREB recruits other partners after dimers form on DNA. Only UpG increased DNA.CREB dimer formation.Since oxidation is ongoing and conversion of cytosine to uracil occurs spontaneously or at specific timesduring differentiation and development, we propose that BER substrates are epigenetic and modulatetranscription factor recognition/binding.
. Introduction
.1. Oxidative damage to DNA is ongoing in all cells and notonsidered to be epigenetic
Epigenetics encompasses DNA methylation, histone modifica-ion, nucleosome localization and synthesis of non-coding RNA, allf which modulate access to the transcription machinery. Theseodifications are central to differentiation, embryonic develop-ent and carcinogenesis [1]. Other DNA modifications including
automeric shifts, deamination, conversion of cytosine to uracilU), oxidation of bases and spontaneous base loss are ongoing.
ost of these lesions are repaired by the base excision DNA repairBER) pathway and have never been considered epigenetic, becauseER efficiently removes them. However, oxidative DNA damage
Abbreviations: AID, activation induced deaminase; AP, abasic; APEX1, humanP endonuclease 1; ATF-1, ATF-2, activating transcription factor-1,-2; BER, basexcision repair; CRE, cAMP response element; CREB, cyclic AMP response elementinding protein 1; CREM, cAMP response element modulator; ◦G, 8-oxo-7,8-ihydroguanine; hmC, hydroxymethylcytosine; hmU, 5-hydroxylmethyluracil;C, 5-methylcytosine; OGG1, oxoguanine glycosylase; RNS, reactive nitrogen
pecies; ROS, reactive oxygen species; THF, tetrahydrofuran.∗ Corresponding author at: Northeastern University, Department of Biology, 360untington Avenue, Boston, MA 02115, United States. Tel.: +1 617 3733492;
ax: +1 617 3733724.E-mail address: [email protected] (P.R. Strauss).
568-7864/$ – see front matter © 2013 Published by Elsevier B.V.ttp://dx.doi.org/10.1016/j.dnarep.2013.10.002
© 2013 Published by Elsevier B.V.
due to reactive oxygen species (ROS) or reactive nitrogen species(RNS) from either intra- or extracellular sources is a continuousprocess in all cells [2], with oxidation of guanine (G) to 8-oxo-7,8-dihydroguanine (◦G) being especially common. Repair is efficientbut not perfect and a low level of ◦G and U can be found in thegenome.
1.2. The BER pathway, which repairs most oxidative damage andremoves uracil in DNA, is a cascade of critical enzymes
The BER pathway is initiated by specific DNA glycosylases thatlargely remove the aberrant base without cleaving the DNA back-bone and leave an abasic (AP) site, the accumulation of which istoxic [3]. The endonuclease function of the enzyme AP endonu-clease (APEX1) then recognizes and cleaves this AP site on the 5′
side of the phosphodeoxyribose, after which a single nucleotide isinserted by DNA polymerase � [4]. However, in some cases, such aswith ◦G, the lesion is recognized by oxoguanine glycosylase (OGG1)that not only removes the aberrant base but frequently cleaves theAP site on the 3′ side requiring the removal of the sugar by APEX1[3]. The loss of APEX1 is embryonic lethal in mice [5], while partial
knockdown of Apex1 in zebrafish embryos leads to improper devel-opment of heart and neural tissue, so that development continuesuntil heart function is required several days later and the embryosdie [6]. Furthermore, knockdown of Apex1 in primary murine cells
S.P.G. Moore et al. / DNA Repai
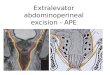
Fig. 1. Co-crystal of the transcription factor CREB1 bound as a dimer to the CRE-siteoow
aA
aldwcyg5r
1i
taoticfC([tt(orfth
fiogowirWo
three independent times, except for two experiments that were
f a ds-oligonucleotide. The G1, G2, and C positions of the CpG islet on a single strandf DNA are shown in space fill to indicate proximity to the protein. This structureas obtained from PDB 1DH3.
nd zebrafish embryos results in a p53-independent loss of cyclicMP response element binding protein (CREB) [7].
Cytosine can be modified by a number of different mechanismsssociated with chromatin modification and gene activation. Inymphocytes, C is converted directly to U by activation inducedeaminase (AID) during immunoglobulin processing [8,9]. Mean-hile, for chromatin to be activated, mC residues need to be
onverted to C through the removal of the methyl group. Demeth-lation of mC occurs through several enzymatic pathways whichenerate various intermediates such as T, hmC, 5-carboxylcytosine,-formylcytosine or 5-hydroxylmethyluracil (hmU). BER is thenequired to convert the resulting intermediates to C [10].
.3. The oxidative state of DNA could be epigenetic and affectmportant transcription binding factors like CREB
Cellular redox status, reflected in the levels of glutathione andhioredoxin/thioredoxin reductase, is usually construed as the rel-tive ability to make or break disulfide bonds [11]. Despite thengoing production of ROS and resultant oxidative damage to DNA,he oxidative state of DNA is overlooked. We have chosen to exam-ne the ability of CREB to bind to its CRE consensus sequenceontaining U or selected oxidative damage, as CREB is responsibleor the transcriptional activation of ∼25% of the genome [12,13].REB binds to the eight-base palindromic cAMP-response elementCRE) TGACGTCA, or the non-palindromic half-site motif CGTCA14]. The full length CRE-site contains two G residues per strandhat could be susceptible to oxidation, one located in a CpG islethat is subject to methylation/demethylation or conversion to UpGFig. 1). CREB commonly functions as a homodimer but is capablef forming heterodimers with other bZIP members including cAMPesponse element modulator (CREM) and activating transcriptionactor 1 or 2 (ATF-1 or ATF-2) [15]. Despite the name “activatingranscription factor,” several of the ATF’s including ATF-2 in theeterodimer serve as repressors [16].
Since CREB is the foundation on which many other transcriptionactors build the complex that will recruit RNA polymerase andnitiate transcription, subtle changes in CREB binding could amplifyr depress transcription many fold. Since even small changes inene expression can have an enormous impact on cellular devel-pment, changes in oxidative state or the presence of a U residueithin promoter sites may be epigenetic [17,18]. In this study we
nvestigated the possibility that lesions and modifications in DNAepaired by BER might alter CREB binding to its CRE target site.
e present quantitative data indicating that the oxidative statusf DNA itself and, in particular, lesions repaired by BER, are an
r 12 (2013) 1152– 1158 1153
important component in regulating recognition of transcriptionfactors to their cognate sequences and, therefore, are likely to playan epigenetic role in cell physiology. It is even possible that futureexperiments will show that other covalent modifications to DNAmight also have epigenetic effects.
2. Methods
2.1. Oligonucleotides
39-mer oligonucleotides containing a CRE-site with or with-out various DNA modifications (Table 1) were sourced either fromMidland Certified Reagent Co., Inc. (Midland, Texas) or from Inte-grated DNA Technologies (Coralville, Iowa). G1 and G2 AP sitesand G1 5′ and 3′ dRP modifications were formed with tetrahydro-furan (THF). Probes that had nicks between nucleotides or weremissing nucleotides were constructed by synthesizing two top orbottom strands that when annealed together with a full lengthcomplementary strand resulted in a gap or nick being present atthe desired position. Either upper or lower strands were labeled atthe 5′ end with [�32P]ATP (PerkinElmer, Waltham, Massachusetts)by polynucleotide kinase (PNK) (New England Biolabs, Ipswich,Massachusetts) as previously described [19,20]. After the label-ing reaction was complete, PNK was heat-inactivated at 95 ◦C for5 min and the desired complement was added at a 1:1 molar ratio.The mixture was slow-cooled, separated from unincorporated 32Pusing IllustraTM MicroSpinTM G-25 columns (GE Healthcare, LittleChalfont, United Kingdom), and stored at −20 ◦C.
2.2. Protein
Purified recombinant human CREB protein was the kind gift ofDr. Jennifer Nyborg (Colorado State University, Fort Collins, Col-orado). Before use CREB was diluted into a buffer containing 50 mMTris–HCl (pH7.9), 100 mM KCl, 12.5 mM MgCl2, 1 mM EDTA (pH8.0), 20% glycerol, and 2 mM DTT.
2.3. Electrophoretic Mobility Shift Assay (EMSA)
EMSAs were performed as described by Lopez et al. [21]. Briefly,varying concentrations (0 to 50 nM) of purified CREB proteinwere incubated in a reaction buffer (25 mM Tris [pH 7.9], 50 mMKCl, 6.25 mM MgCl2, 0.5 mM EDTA, 10% glycerol, 0.1% Tween-20,and 2 mM DTT) with 5 ng poly(dA-dT)·poly(dA–dT), 0.25 �g/�LBSA, and 32P-end labeled double stranded oligonucleotide probe(0.5 nM) on ice for 1 h in a final volume of 20 �L. Samples werethen loaded into the gel without tracking dye and resolved atroom temperature on pre-electrophoresed native 5% (49:1 acry-lamide:bisacrylamide) gels in a buffer composed of 40 mM Tris–HCl(pH 8.5), 306 mM glycine and 0.1% Nonidet P-40. Electrophoresiswas performed at 90 V for 2.5 h until a tracking lane containingbromphenol blue dye had reached ∼2/3 down the gel.
2.4. Binding kinetics
Vacuum dried gels were exposed to a PhosphoImager screen andvisualized with a GE StormTM PhosphorimagerTM; data were quan-tified with Molecular Dynamics ImageQuant (GE Healthcare, LittleChalfont, United Kingdom). Experiments were performed at least
conducted twice. Kd values were calculated using the equation:
Kd = [unbound DNA][unbound CREB][bound CREB]
(1)

1154 S.P.G. Moore et al. / DNA Repair 12 (2013) 1152– 1158
Table 1Modifications to CRE used in this study. The residue/s and positions within the CRE-site that were modified are indicated in the Location column, whereas the Modificationcolumn indicates the changes at that site. The unmodified control 39-mer oligonucleotide is shown with the CRE site in bold (5′- TAC CAT GCC TTG1 ACG2 TCA GAG AGCATT CGT AAT CAT GGT-3′). Numerals 1 and 2 indicate the guanine or cytosine residue/s that were modified. Some probes had modifications in both top and bottom strands;top and bottom strands are differentiated by T or B in the Sequence column. Probes that had nicks between nucleotides or were missing nucleotides were constructed bysynthesizing two top or bottom strands that when annealed together with a full length complementary strand resulted in a gap or nick being present at the desired position.These are indicated by | for a nick and � for a gap. The 5′ oligonucleotide ends at the nick or gap sites did not have phosphate groups present.
Dt
oc
2
ImtIuw
Bound CREB was calculated by quantifying the amount of boundNA; unbound CREB was calculated as the difference between ini-
ial CREB and bound CREB.For substrates where saturation binding was not achieved under
ur assay conditions, we used the initial slope to estimate a con-entration of CREB where half maximal binding would be expected.
.5. Statistical analyses
Data were analyzed using SPSS 19.0 (IBM, Armonk, New York).ndependent t-tests were performed between data for each DNA
odification and the unmodified control sequence. Before each t-
est was applied, a Levene’s test for equal variance was conducted.f the assumption of equal variances was violated, a t-test withnequal variances was utilized. Significance for all statistical testsas set a priori at 0.05.3. Results
3.1. Purified CREB recognizes and binds the CRE-site cleanly
DNA.protein complexes (monomer and dimer) formed betweenpurified recombinant CREB and the 39-mer sequence 5′- TAC CATGCC TTG1 ACG2 TCA GAG AGC ATT CGT AAT CAT GGT-3′ (CREsequence shown in bold) resolved cleanly using the experimen-tal protocols employed in this study (Figs. 2 and 3). The 5′-most Gin the sequence is indicated as G1, while the G residue in the CpGislet is indicated as G2. As the concentration of CREB increased, themonomer form (DNA.CREB) appeared first followed at higher con-
centrations by the dimer form. The Kd for total CREB binding tocontrol 39-mer was 4.8 ± 0.2 nM (SE,3) (Figs. 3 and 4). After CREBinitially bound to the CRE-site as a monomer (DNA.CREB), a secondmolecule of CREB bound to DNA.CREB to form a dimer (DNA.CREB2),
S.P.G. Moore et al. / DNA Repair 12 (2013) 1152– 1158 1155
Fig. 2. Purified recombinant CREB binds to a 39-mer oligonucleotide containing the8-mer CRE site first as a monomer and then as a dimer. Purified CREB at increas-ing concentrations was incubated on ice with a 39-mer 32P-labeled CRE-containingprobe (0.5 nM) and resolved on a 5% nondenaturing PAGE. The experiment dis-played contains a G/U mispair at the G2 position (CpG). Note the clean separationof monomer and dimer.
Fig. 3. Selected DNA modifications alter the overall affinity of CREB for the CRE-site both positively and negatively. Graph showing quantitative binding of CREBto 39-mer oligonucleotide containing select modifications as a function of CREBc ◦
up
woK(
3bl
aSt0airUcUCtprerrvt
Fig. 4. The type of DNA modification and its position within the CRE-site has a dra-matic effect upon recognition and binding of purified CREB. (A) Modifications at G1.Modifications at the G1 position are generally detrimental to total CREB bindingexcept when ◦G, a nick on the 3′ side of G1, or a G/U mispair is present. (B) Modi-fications at G2. While a single ◦G did not affect total binding, a G/U mispair in theG2 position enhanced total CREB binding. All other modifications in the G2 positionwere deleterious to total CREB binding. Each experiment was performed indepen-dently three times, except for when a nick on the 3′ side of G1, or a nick on the 5′ side
oncentration. �, control; �, G1 G; �, G2 U; �, CpG islet with C residues on bothpper and lower strands methylated and one G residue as ◦G. Each experiment waserformed three times with error bars representing ± SE.
hich is in keeping with published reports that CREB forms dimersn CRE only after the DNA.monomer complex has formed [22].inetics of dimer formation are dissected from overall data below
Fig. 5).
.2. CREB’s ability to bind to its consensus sequence is affectedoth positively and negatively by specific modifications and theirocations
A G/U mispair positioned at G1 did not interfere with CREB’sbility to recognize and bind to the CRE-site (Kd G1 G/U 4.2 ± 0.7 nM,E,3). However, when a G/U mispair was positioned at G2 withinhe CpG islet, CREB binding was enhanced 6-fold (Kd G2 G/U.8 ± 0.1 nM, SE,3) relative to the unmodified control (Fig. 4And B). Unlike the case with a single G/U mispair, CREB bind-ng was enhanced only 3-fold (Kd 1.9 ± 0.12 nM, SE,3) when Cesidues in the CpG islet in both strands were replaced with. The differences in binding between control and a single U,ontrol and two U residues, and between a single U and two
residues were significant at p ≤ 0.001. The ability of purifiedREB to recognize and bind to the CRE-site was not affected byhe presence of a single ◦G modification at either the G1 or G2ositions (Kd G1 ◦G 3.7 ± 1.0 nM, SE,3; Kd G2 ◦G 5.0 ± 0.5 nM, SE,3)elative to the unmodified control sequence (Fig. 4A and B). How-ver, when both CRE-site G residues on the same strand or G
◦
esidues on both strands in the CpG islet were replaced with Gesidues, saturable binding could not be achieved (Fig. 6A). Kdalues, calculated from the initial binding slope at low concentra-ions, were 36 ± 3.3 nM (SE, 3) and 36.7 ± 2.5 nM (SE, 3) respectivelyof the C in the CpG islet were present, which were performed twice. In the former,error bars represent ± S.E.; in the latter error bars represent ± range and significancewas not calculated. P-values are represented as ***≤0.001, **≤0.01, *≤0.05.
(Fig. 6A). An AP site at either the G1 or G2 positions (Fig. 4A andB) was detrimental for CREB binding (Kd G1 AP 8.1 ± 0.5 nM, SE,3;Kd G2 AP 13.9 ± 1.5 nM, SE,3). Other modifications to the G1 or G2residue, or the C residue within the CpG islet were deleterious tototal CREB binding, except for the presence of a nick in the back-bone on the 5′ side of the G1 guanine (Kd G1 5′Nick 5.5 ± 1.2 nM,SE,3) (Fig. 4A and B). This lesion would have been generated bythe 5′ incision repair activity of APEX1 [23]. Methylation of both Cresidues on both strands in the CpG islet, e.g. methylation on bothupper and lower strands, resulted in extremely poor CREB binding(Kd hypermethylated 99.1 ± 15.7 nM, Fig. 6A), while hemimethy-lation resulted in reduced binding affinity (Kd hemimethylated17.4 ± 3.2 nM, SE,3, Fig. 4B). Failure of CREB to bind to its CREsequence when both C residues in the CpG context are methylatedhas been reported previously [24].
3.3. DNA modification and spatial position within the CRE-siteaffects CREB’s ability to form a dimer
Since CREB is only transcriptionally active when two proteinmolecules (either CREB or CREB and one of its binding part-ners) have bound to each CRE-site [15], we investigated howmodifications to selected residues in the CRE-site might affectdimer formation, which was easily detected under our conditions(Figs. 2 and 3). When 50 nM CREB was added to the reaction mix,
62% of the total amount of bound CREB was bound as a dimeron the control consensus sequence (0.28 ± 0.02 nM, SE,3) (Fig. 5Aand B). Several G1 modifications including a nick in the DNA back-bone 5′ to the G1 residue (0.26 ± 0.02 nM, SE,3) and a U opposite
1156 S.P.G. Moore et al. / DNA Repair 12 (2013) 1152– 1158
Fig. 5. Only a U in the CpG context enhances dimer formation on DNA. (A) Dimer for-mation after modification at G1. CREB ability to form a dimer remained unchangedwhen a nick in the DNA backbone 5′ to the G1 residue, or a G/U mispair was presentat the G1 position. Other modifications to the G1 position including the presence of◦G resulted in impaired dimer formation by CREB. (B) Dimer formation after mod-ification of G2. The presence of a G/U mispair at the G2 position enhanced CREB’sability to form a dimer on the CRE-site, while all other modifications to the CpGislet inhibited CREB dimerization relative to the unmodified control. The amount ofDNA.CREB2 as dimer was calculated as part of the kinetic binding studies, using thedata from the 50 nM CREB concentration. Each experiment was performed indepen-dently three times, except for when a nick on the 3′ side of G1, or a nick on the 5′ sideoew
tfer(rpGC
4
4
ircm(Cwcp
Fig. 6. Some DNA modifications within the CRE-site were especially deleteriousto CREB binding so that saturation was not achieved under our conditions. (A) Kd
values in these cases were estimated from the initial linear binding slope and calcu-lating the concentration of CREB at which half concentration would occur. Bindingwas reduced up to 20-fold with these modifications. (B) Dimerization at 50 nM wasseverely reduced up to 9-fold for substrates where saturation did not occur. Allexperiments were performed independently three times except for fully methyl-
f the C in the CpG islet was present, which were performed twice. In the former,rror bars represent ± S.E.; in the latter error bars represent ± range and significanceas not calculated. P-values are represented as ***≤0.001, **≤0.01, *≤0.05.
he G1 residue (0.26 ± 0.01 nM, SE,3) did not interfere with dimerormation despite changes in overall Kd values (Fig. 5A). How-ver, all other modifications to the G1 residue that we testedesulted in reduced dimer formation, including a single ◦G at G10.23 ± 0.01 nM, SE,3) (Fig. 5A). When modifications were made toesidues in the CpG context, dimer formation was generally sup-ressed (Fig. 5B and Fig. 6B) except for a G/U mispair (Fig. 5B). A/U mispair enhanced dimer formation relative to the unmodifiedRE-site (0.33 ± 0.01 nM, SE,3).
. Discussion
.1. Major conclusions
Here we have shown that DNA modifications of guanine residuesn the CREB consensus sequence, the presence of U, or lesionsepaired through the BER pathway alter CREB binding to its CREonsensus sequence positively or negatively depending on theodification and its location. Selected G or C residues were oxidized
◦G), converted to U, T, methylated C (mC), 5-hydroxymethylated
(hmC), or a range of intermediates arising during BER. The effectsere specific: conversion of a single C residue within the CpGontext to U enhanced total binding 6-fold, while a G/T mis-air, oxidation of both G residues or a methylated C was highly
ated CpG, which was performed twice. In the former, error bars represent ± S.E.,while in the latter error bars represent ± range and significance was not calculated.P-values are represented as ***≤0.001, **≤0.01, *≤0.05.
deleterious, decreasing binding up to 25-fold of the control level,thus expanding the range of CREB binding over two orders of mag-nitude. Furthermore, substitution with all intermediates of the BERpathway was also deleterious. In agreement with others [24], CREBbinds poorly to its target site when the cytosine residues in theCpG islet of the CRE-site were hyper- or fully methylated, presum-ably preventing indiscriminant and wasteful transcription. We alsoexamined the ability of highly purified recombinant CREB to formdimers on the CRE sequence. While others have noted effects of aG/U mispair on CREB or activator protein-1 (AP-1) binding from anuclear extract [25], the ability to discriminate monomer and dimerby purified CREB allowed us to quantify useful binding [26], becauseCREB recruits its binding partners only when it has formed a dimeron the CRE.
4.2. Response to oxidative DNA damage is pleomorphic andprobably extends to other transcription factors
It is widely accepted that oxidative DNA damage is harmful.Oxidative base lesions have been linked to carcinogenesis andneurodegenerative diseases such as Alzheimer’s and Parkinsonsdisease [27,28]. However, the response to oxidation is, in fact,graded and depends on the gene in question: in our study CREBbinding tolerated conversion of either G residue individually to ◦Gwithout change in recognition, while conversion of both G residuesto ◦G and all BER intermediates were highly deleterious. Sensitivityto guanine oxidation depends on the transcription factor as well,since single ◦G lesions in the binding sites of the transcription fac-tors AP-1 and Sp1 are sufficient to inhibit binding affinity [18,29,30].
In contrast, ◦G lesions in select residues of the NF-kB consensus siteenhance binding affinity [29], while an 8-oxo-A lesion positioned atthe first adenine residue of the CREB consensus sequence enhancesbinding affinity [31]. Since oxidized adenine occurs at much lower
Repai
fs
fitmwtCdwcosatCb
∼tNftsSccCtiis1hFsmteisiBBg
4
spaataAmuterwoc
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S.P.G. Moore et al. / DNA
requency than ◦G, it may or may not play a role in modulating tran-cription. No data on dimerization are available in these studies.
Conformational changes to DNA imposed by the various modi-cations examined in this study account for many of the changeso CREB binding kinetics. For example, an AP site and its compli-
entary residue often adopt an extrahelical conformation [32,33],hile DNA containing a T:G mispair displays a distorted conforma-
ion with the T often flipped out [34]. Both would interfere withREB binding and dimer formation. The fact that single ◦G lesionso not appear to overly disturb DNA structure [32] is consistentith the observation that the overall binding of CREB when CRE
ontains a single ◦G does not differ with that of control. On thether hand, NMR or electron resonance structural studies of othertructures indicated negligible perturbation to DNA when a nick or
gap is present [32]. However, these studies were performed inhe absence of protein, suggesting that nicks, gaps, or an AP site inRE may increase flexibility that is incompatible with strong CREBinding.
CREB acts as an anchor protein that enables transcription of25% of the eukaryotic genome [12,13]. Thus, one would expect
hat genes containing CRE sites in their promoters, including FOS,FkB, and genes whose promoters include consensus binding sites
or those transcription factors would be more sensitive to oxida-ive damage than others. Transcription factors whose consensusequences are enriched for G residues would also be prime targets.p1 is one such transcription factor whose consensus sequence isomposed solely of G and C residues. Consequently, select genesontaining one or more Sp1 binding sites as well as one or moreRE in their promoter sequences could be especially responsiveo oxidative damage. Genes with both Sp1 and CREB binding sitesnclude nitric oxide synthase 1 and 3, dihydrofolate reductase,nterleukin 10 and the major enzymes involved in base exci-ion repair including 8-oxoguanine glycosylase, AP endonuclease
and DNA polymerase ß. At least one of these genes (APEX1)as 8 CREB binding sites and 11 AP-1 sites. Since AP-1 consists ofOS and JUN hetero or homodimers and, since FOS has 8 CREB1ites in its promoter, transcription of such genes would be evenore CREB dependent and, therefore, would be even more subject
o potential regulation by oxidation of G residues or the pres-nce of U. CREB itself is autoregulated with 13 self binding sitesn its own promoter. The CREB1 promoter also contains 7 AP-1ites, 1 Sp1 site, 4 NFkB sites, 2-p300 sites, and an ATF site. Themportance of oxidation and potential U in these sites to regulateER enzymes is further underscored by the fact that the sites inER promoters are conserved among the human, mouse and ratenomes [35].
.3. DNA damage repaired by BER is likely to be epigenetic
Small changes to gene expression in a single prominent tran-cription factor like CREB1 can have a considerable impact on cellhysiology and development. Given the broad importance of CREBnd the fact that oxidative modification and the presence of Uffect other critical transcription factors as well, we propose thathe lesions repaired by the base excision repair pathway serves modulators for transcription efficiency. It is even possible thatID/APOBEC enzymes might convert C residues to uracil selectively,uch as in the case of mutation and recombination of immunoglob-
lin genes [36]. While higher levels of oxidative damage might alerthe cellular machinery of impending oxygen toxicity, lower lev-
ls or the presence of U could serve as a titration mechanism toegulate the degree of activation of selected genes. Consequently,e propose that selective oxidative alterations and the presencef U in DNA act epigenetically and separately from modification ofhromatin proteins.
[
[
r 12 (2013) 1152– 1158 1157
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgments
The authors thank Dr. Jennifer Nyborg (Colorado State Univer-sity) for the gift of purified human CREB.
This work was supported by the G. Harold and Leila Y. MathersFoundation; Aid for Cancer Research; and funds from NortheasternUniversity.
References
[1] G. Riddihough, L.M. Zahn, What is Epigenetics? Science 330 (2010) 611.[2] S.S. David, V.L. O’Shea, S. Kundu, Base-excision repair of oxidative DNA damage,
Nature 447 (2007) 941–950.[3] D.O. Zharkov, A.P. Grollman, The DNA trackwalkers: principles of lesion search
and recognition by DNA glycosylases, Mutat. Res. 577 (2005) 24–54.[4] D.O. Zharkov, Base excision DNA repair, Cell. Mol. Life Sci. 65 (2008) 1544–1565.[5] S. Xanthoudakis, J.D. Smeyne, J.D. Wallace, T. Curran, The redox/DNA repair
protein, Ref-1, is essential for early embryonic development in mice, Proc. NatlAcad. Sci. USA 93 (1996) 8919–8923.
[6] Y. Wang, C.C. Shupenko, L.F. Melo, P.R. Strauss, DNA repair protein involved inheart and blood development, Mol. Cell. Biol. 26 (2006) 9083–9093.
[7] D.-S. Pei, X.-J. Yang, W. Liu, J.E.J. Guikema, C.E. Schrader, P.R. Strauss, A novelregulatory circuit in base excision repair involving AP endonuclease 1. Creb1and DNA polymerase beta, Nucleic Acids Res. 39 (2011) 3156–3165.
[8] R.W. Maul, H. Saribasak, S.A. Martomo, R.L. McClure, W. Yang, A. Vaisman, H.S.Gramlich, D.G. Schatz, R. Woodgate, D.M. Wilson, P.J. Gearhart, Uracil residuesdependent on the deaminase AID in immunoglobulin gene variable and switchregions, Nat. Immunol. 12 (2011) 70–76.
[9] H.E. Krokan, F. Drabløs, G. Slupphaug, Uracil in DNA – occurrence, consequencesand repair, Oncogene 21 (2002) 8935–8948.
10] S.C. Wu, Y. Zhang, Active DNA demethylation: many roads lead to Rome, Nat.Rev. Mol. Cell Biol. 11 (2010) 607–620.
11] G. Manda, M.T. Nechifor, T.-M. Neagu, Reactive oxygen species, cancer and anti-cancer therapies, Curr. Chem. Biol. 3 (2009) 342–366.
12] X. Zhang, D.T. Odom, S.-H. Koo, M.D. Conkright, G. Canettieri, J. Best, H. Chen,R. Jenner, E. Herbolsheimer, E. Jacobsen, S. Kadam, J.R. Ecker, B. Emerson, J.B.Hogenesch, T. Unterman, R.A. Young, M. Montminy, Genome-wide analysisof cAMP-response element binding protein occupancy, phosphorylation, andtarget gene activation in human tissues, Proc. Natl Acad. Sci. USA 102 (2005)4459–4464.
13] S. Impey, S.R. Mccorkle, H. Cha-Molstad, J.M. Dwyer, G.S. Yochum, J.M. Boss,S. Mcweeney, J.J. Dunn, G. Mandel, R.H. Goodman, N. York, Defining the CREBregulon: a genome-wide analysis of transcription factor regulatory regions, Cell119 (2004) 1041–1054.
14] B. Mayr, M. Montminy, Transcriptional regulation by the phosphorylation-dependent factor CREB, Nat. Rev. Mol. Cell Biol. 2 (2001) 599–609.
15] A.J. Shaywitz, M.E. Greenberg, CREB: a stimulus-induced transcription factoractivated by a diverse array of extracellular signals, Annu. Rev. Biochem. 68(1999) 821–861.
16] T. Hai, M.G. Hartman, The molecular biology and nomenclature of the activatingtranscription factor/cAMP responsive element binding family of transcriptionfactors: activating transcription factor proteins and homeostasis, Gene 273(2001) 1–11.
17] R. Ghosh, D.L. Mitchell, Effect of oxidative DNA damage in promoter elementson transcription factor binding, Nucleic Acids Res. 27 (1999) 3213–3218.
18] D. Mitchell, R. Ghosh, in: M.D. Evans, M.S. Cooke (Eds.), Oxidative Damage toNucleic Acids, Springer, New York, NY, 2007, pp. 91–99.
19] S. Fortier, X. Yang, Y. Wang, R.A.O. Bennett, P.R. Strauss, Base excision repairin early zebrafish development: evidence for DNA polymerase switching andstandby AP endonuclease activity, Biochemistry 48 (2009) 5396–5404.
20] P.R. Strauss, W.A. Beard, T.A. Patterson, S.H. Wilson, Substrate binding by humanapurinic/apyrimidinic endonuclease indicates a Briggs-Haldane mechanism, J.Biol. Chem. 272 (1997) 1302–1307.
21] D.I. Lopez, J.E. Mick, J.K. Nyborg, Purification of CREB to apparent homogeneity:removal of truncation products and contaminating nucleic acid, Protein Expres.Purif. 55 (2007) 406–418.
22] X. Wu, C. Spiro, W.G. Owen, C.T. McMurray, cAMP response element-bindingprotein monomers cooperatively assemble to form dimers on DNA, J. Biol.Chem. 273 (1998) 20820–20827.
23] N.A. Timofeyeva, V.V. Koval, A.A. Ishchenko, M.K. Saparbaev, O.S. Fedorova,Lys98 substitution in human AP endonuclease 1 affects the kinetic mechanismof enzyme action in base excision and nucleotide incision repair pathways, PloSOne 6 (2011) 1–10.
24] S.M. Iguchi-Ariga, W. Schaffner, CpG methylation of the cAMP-responsiveenhancer/promoter sequence TGACGTCA abolishes specific factor binding aswell as transcriptional activation, Genes Dev. 3 (1989) 612–619.
25] A. Verri, P. Mazzarello, G. Biamonti, S. Spadari, F. Focher, The specific bind-ing of nuclear protein(s) to the cAMP responsive element (CRE) sequence

1 Repa
[
[
[
[
[
[
[
[
[
[
158 S.P.G. Moore et al. / DNA
(TGACGTCA) is reduced by the misincorporation of U and increased by thedeamination of C, Nucleic Acids Res. 18 (1990) 5775–5780.
26] D.K. Rogstad, P. Liu, A. Burdzy, S.S. Lin, L.C. Sowers, Endogenous DNA lesionscan inhibit the binding of the AP-1 (c-Jun) transcription factor, Biochemistry41 (2002) 8093–8102.
27] M.S. Cooke, M.D. Evans, M. Dizdaroglu, J. Lunec, Oxidative DNA damage: mech-anisms, mutation, and disease, FASEB J. 17 (2003) 1195–1214.
28] M. Smeyne, R.J. Smeyne, Glutathione metabolism and Parkinson’s disease, FreeRadical Biol. Med. (2013) 1–13.
29] M.K. Hailer-Morrison, J.M. Kotler, B.D. Martin, K.D. Sugden, Oxidized guaninelesions as modulators of gene transcription, Altered p50 binding affinity andrepair shielding by 7,8-dihydro-8-oxo-2’-deoxyguanosine lesions in the NF-kappaB promoter element, Biochemistry 42 (2003) 9761–9770.
30] O. Ramon, S. Sauvaigo, D. Gasparutto, P. Faure, A. Favier, J. Cadet, Effects of 8-
oxo-7,8-dihydro-2’-deoxyguanosine on the binding of the transcription factorSp1 to its cognate target DNA sequence (GC box), Free Radical Res. 31 (1999)217–229.31] J. Abraham, P.J. Brooks, Divergent effects of oxidatively induced modifica-tion to the C8 of 2’-deoxyadenosine on transcription factor binding: 8,5’
[
ir 12 (2013) 1152– 1158
(S)-cyclo-2’-deoxyadenosine inhibits the binding of multiple sequence specifictranscription factors, while 8-oxo-2’-deoxyadenosine, Environ. Mol. Mutagen.52 (2011) 287–295.
32] G. Sicoli, G. Mathis, S. Aci-Sèche, C. Saint-Pierre, Y. Boulard, D. Gasparutto, S.Gambarelli, Lesion-induced DNA weak structural changes detected by pulsedEPR spectroscopy combined with site-directed spin labelling, Nucleic Acids Res.37 (2009) 3165–3176.
33] J. Chen, F.-Y. Dupradeau, D.A. Case, C.J. Turner, J. Stubbe, DNA oligonucleotideswith A, T, G or C opposite an abasic site: structure and dynamics, Nucleic AcidsRes. 36 (2008) 253–262.
34] P. Imhof, M. Zahran, The effect of a G:T mispair on the dynamics of DNA, PloSOne 8 (2013) 1–11.
35] M. Rebhan, V. Chalifa-Caspi, J. Prilusky, D. Lancet, GeneCards: anovel functional genomics compendium with automated data min-
ing and query reformulation support, Bioinformatics 14 (1998)656–664.36] H.C. Smith, R.P. Bennett, A. Kizilyer, W.M. McDougall, K.M. Prohaska, Functionsand regulation of the APOBEC family of proteins, Semin. Cell Dev. Biol. 23 (2012)258–268.