Embed Size (px)
Citation preview

Acc
epte
d A
rtic
le
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an 'Accepted Article', doi: 10.1111/tpj.12630 This article is protected by copyright. All rights reserved.
Received Date : 28-May-2014 Revised Date : 21-Jul-2014 Accepted Date : 24-Jul-2014 Article type : Original Article DNA methylation maintenance consolidates RNA-directed DNA methylation and
transcriptional gene silencing over generations in Arabidopsis thaliana
Markus Kuhlmann§1,3, Andreas Finke§1,4, Martin Mascher2 and Michael Florian Mette*1,5
§ These authors contributed equally to the work.
1 Research Group Epigenetics, Leibniz Institute of Plant Genetics and Crop Plant Research
(IPK), D-06466 Gatersleben, Germany;
2 Research Group Bioinformatics and Information Technology, Leibniz Institute of Plant
Genetics and Crop Plant Research (IPK), D-06466 Gatersleben, Germany;
3 Current affiliation: Research Group Abiotic Stress Genomics, Interdisciplinary Center for
Crop Plant Research (IZN), D-06120 Halle (Saale), Germany;
4 Current affiliation: Research Group Genome and Epigenome Maintenance, Max Planck
Institute for Plant Breeding Research, D-50829 Köln, Germany;
5 Current affiliation: Research Group Quantitative Genetics, Leibniz Institute of Plant
Genetics and Crop Plant Research (IPK), D-06466 Gatersleben, Germany;
*Author for correspondence: email [email protected], phone ++49 39482 5181, fax
++49 39482 5137;

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Email contacts of other authors: [email protected], [email protected],
Running title: Consolidation of RdDM and TGS
Keywords (up to 10): RNA-directed DNA methylation, transcriptional gene silencing, DNA
methylation maintenance, transgenerational effect, Arabidopsis thaliana, DRM2, At5g14620;
Accession numbers: PRJEB6549 (whole genome shotgun reads of Col-0 and nrd3-2)
Summary
In plants, 24-nucleotide short interfering RNAs serve as a signal to direct cytosine
methylation at homologous DNA regions in the nucleus. If the targeted DNA has promoter
function, this RNA-directed DNA methylation can result in transcriptional gene silencing. In
a genetic screen for factors involved in RNA-directed transcriptional silencing of a ProNOS-
NPTII reporter transgene in Arabidopsis thaliana, we captured alleles of DOMAINS
REARRANGED METHYLTRANSFERASE 2, the gene encoding the DNA methyltransferase
mainly responsible for de novo DNA methylation in the context of RNA-directed DNA
methylation. Interestingly, reporter gene ProNOS methylation was not erased completely in
these mutants, but persisted in symmetric CG context, indicating that RNA-directed DNA
methylation had been consolidated by DNA methylation maintenance. Taking advantage of
the segregation of the transgenes giving rise to ProNOS short interfering RNAs and carrying
the ProNOS-NPTII reporter in our experimental system, we found that ProNOS DNA
methylation maintenance became first evident after two generations of ongoing RNA-
directed DNA methylation, and then increased in its extent with further generations. As

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
ProNOS DNA methylation already had reached its final level in the first generation of RNA-
directed DNA methylation, our findings suggest that setting up DNA methylation at a
particular region can be divided into distinct stages. An initial phase of efficient, but still fully
reversible de novo DNA methylation and transcriptional gene silencing is followed by the
transition to efficient maintenance of cytosine methylation in symmetric sequence context
accompanied by persistence of gene silencing.
Introduction
Proper temporal and spatial regulation of gene expression is essential for all complex
organisms. In eukaryotes, confinement of gene transcription is supported by chromatin
marks. One of the most prominent marks in plants is methylation at position 5 of cytosines in
genomic DNA, which is mainly associated with transcriptionally silent repetitive sequences
and transposable elements and is typical for heterochromatin. DNA methylation in plants can
not only be found in CG but also in CHG and CHH (with H standing for C, A, T) sequence
context (Cokus et al,. 2008; Lister et al., 2008). In A. thaliana DNA preparations,
approximately 7% of all cytosines are methylated (Rozhon et al., 2008), with methylation
levels amounting to 24% in CG, 6.7% in CHG and 1.7% in CHH context (Cokus et al.,
2008).
Cytosine methylation is introduced after DNA replication by DNA methyltransferases
(DMTases). The A. thaliana genome encodes four classes of genes displaying sequence
homology to conserved DMTase domains, the METHYLTRANSFERASE 1 (MET1) group
belonging to the Dnmt1 family, the DOMAIN REARRANGED METYHLTRANSFERASE
(DRM) group sharing similarity to Dnmt3, the CHROMOMETHYLASE (CMT) group specific
to plants and one putative member of the Dnmt2 family (Finnegan and Kovac, 2000).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Different DMTase families are thought to be responsible for cytosine methylation in different
sequence contexts.
Members of the DRM group are required for de novo DNA methylation in all sequence
contexts and for the propagation of CHH context methylation. Only DRM1 and DRM2 genes
encode proteins that contain all invariant residues necessary for catalytic activity (Cao et al.,
2000; Cao and Jacobsen, 2002; Naumann et al., 2011; Jullien et al., 2012), while DRM3 is
not expected to display catalytic activity and most likely has auxiliary functions (Henderson
et al., 2010). Sequence specificity is conferred to DRM1 and DRM2-mediated methylation by
short interfering (si) RNAs (Zhong et al., 2014) in a mechanism termed RNA-directed DNA
methylation (RdDM) that was first observed in tobacco plants infected with the RNA
pathogen potato spindle tuber viroid (Wassenegger et al., 1994).
A prerequisite for the generation of a siRNA signal guiding RdDM in plants is the formation
of double-stranded (ds) RNA, as was demonstrated by means of Pol II-mediated transcription
of inverted repeat structures (Mette et al., 2000). Resulting transcripts with partial self-
complementarity, able to fold intra-molecularly to form dsRNA stems, could efficiently
trigger DNA hypermethylation of homologous sequences in trans, which, if targeted to
promoter sequences, could lead to transcriptional silencing of affected gene(s) (Mette et al.,
2000; Aufsatz et al., 2002b; Mette et al,. 2005). However, the extent of promoter RdDM and
related RNA-directed transcriptional gene silencing (RdTGS) at different target loci was
reported to vary depending on transgene structure (Khaitova et al., 2011) and insertion
context (Fischer et al., 2008). Once established, DNA methylation can persist to variable
degrees after removal of the inducing trigger (Jones et al., 2001; Lunerova-Bedrichova et al.,
2008; Khaitova et al., 2011). A number of RdDM-reporter systems making use of naturally
occurring as well as engineered transcribed inverted repeats were designed and employed in

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
forward and reverse genetic screens to dissect the molecular pathways underlying RdDM and
RdTGS (Aufsatz et al., 2002a; Bologna and Voinnet, 2014).
Semi-conservative DNA replication in the course of the cell cycle diminishes DNA
methylation in all contexts as the newly synthesised DNA strand is free of methylation.
However, there are mechanisms that allow the re-establishment of pre-replication
methylation patterns at cytosines in symmetric context. In A. thaliana, like in mammals, CG
context sites show a stark correlation of the methylation on one strand with the methylation
on the opposite (Bird, 2002; Cokus et al., 2008). The re-establishment of full CG context
methylation on hemi-methylated DNA is thought to involve the activity of DMTase MET1,
chromatin remodelling factor DECREASE IN DNA METHYLATION1 (DDM1) and
VARIANT IN METHYLATION (VIM) proteins 1 to 3 (Vongs et al., 1993; Woo et al,.
2007; Woo et al,. 2008). Furthermore, the interaction of MET1 with HISTONE
DEACETYLASE 6 (HDA6) is necessary to maintain CG context methylation at various loci
(Aufsatz et al., 2002b; To et al., 2011; Liu et al., 2012; Blevins et al., 2014). However, the
details of the molecular mechanisms underlying CG context methylation maintenance in
plants are still only partly understood.
The maintenance of DNA methylation in symmetric CHG context is unique to plants and
differs significantly from CG methylation maintenance (Cokus et al., 2008). In A. thaliana,
the majority of CHG context methylation depends on the plant-specific DMTase
CHROMOMETHYLASE3 (CMT3) and on the H3-K9-specific SUPPRESSOR OF
VARIAGATION3-9 HOMOLOG (SUVH) histone methyltransferases SUVH4, SUVH5,
SUVH6 (Jackson et al., 2002; Malagnac et al., 2002; Ebbs and Bender, 2006; Pontvianne et
al., 2012). Experiments that showed an in vitro affinity of the chromodomain of CMT3 to
K9-methylated H3-derived peptides and a decrease of CHG methylation in H3K9me2-
deficient mutants suggest that H3-K9-methylation serves as a label for CMT3 target sites

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
(Lindroth et al., 2004), which is also consistent with results from the genome-wide
superimposition of CMT3 binding sites and H3K9me2-marked nucleosomes (Du et al.,
2012). Moreover, the involvement of histone deacetylase HDA6 in CHG methylation
maintenance at specific loci is also well documented (To et al., 2011; Liu et al., 2012;
Blevins et al., 2014).
Here we employ a transgene system undergoing efficient RdDM and RdTGS in order to
analyse the interdependence of de novo DNA methylation and DNA methylation
maintenance in A. thaliana. By means of newly identified drm2 alleles defective in RNA-
directed DNA methylation and by taking advantage of the segregation of transgene loci to
remove the promoter-siRNA source from the target promoter, we demonstrate that DNA
methylation induced by RdDM can be consolidated by CG context DNA methylation
maintenance if RdDM is acting for more than one generation. The initiation of the silenced
state per se by RdDM and the maintenance of CG context DNA methylation are not
associated with histone-H3-lysine-9-dimethylation (H3K9me2) that is otherwise typical for
repressed chromatin in plants.
Results
RNA-directed transcriptional gene silencing is released in nrd3 mutants
We have performed a screen for ethyl methanesulfonate (EMS)-induced mutants (no rna-
directed transcriptional silencing, nrd) that reactivate expression of a silenced reporter
transgene in A. thaliana (Finke et al., 2012a) . The line K/K;H/H submitted to mutagenesis
was doubly homozygous for a SILENCER (H) transgene on chromosome 4 containing an
inverted repeat (IR) of the NOPALIN SYNTHASE promoter (ProNOS) sequence under control

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
of the cauliflower mosaic virus 35S promoter (Pro35) (Aufsatz et al., 2002a) and a TARGET
(K; Kchr1-10 in Fischer et al., 2008) transgene on chromosome 1 containing a NEOMYCIN
PHOSPHOTRANSFERASE II (NPTII) reporter gene controlled by a ProNOS conferring
resistance to kanamycin (Figure 1a) that was previously found to show efficient DNA
methylation and silencing in the presence of the SILENCER (Fischer et al., 2008). Thus,
K/K;H/H plants homozygous for TARGET and SILENCER are sensitive to kanamycin (Figure
1b). F4 K/K;H/H seeds were incubated with EMS, sown on soil and grown to M1 plants,
which were allowed to self-pollinate (Figure S1). The resulting M2 seeds, the first generation
in which an EMS-induced mutation can be homozygous and thus, if recessive, show its
impact on the phenotype, were germinated on medium containing 200 mg/l kanamycin to
screen for individuals that became kanamycin resistant (KanR). Presence and integrity of
TARGET and SILENCER transgenes in resulting KanR M2 K/K;H/H nrd candidate plants
were verified via PCR using primer combinations specific for different parts of the
transgenes. For sake of readability, in the following “nrd” will be used in the sense of “KanR
K/K;H/H nrd” unless specified otherwise. M2 nrd plants were allowed to self-pollinate and
kanamycin resistance was verified for the resulting M3 generation. M3 seedlings of
independent mutant lines nrd3-1 and nrd3-2 showed consistent resistance when grown on
medium containing kanamycin (Figure 1b).
As kanamycin resistance can also arise in A. thaliana mutants by loss of chloroplast-localised
transporter proteins required for kanamycin uptake (Aufsatz et al., 2009; Conte et al., 2009)
rather than by reactivated NPTII expression, NPTII protein levels in mutants were tested by
ELISA (Figure 1c). M3 nrd3-1 and nrd3-2 plants showed clearly more NPTII than K/K;H/H
plants, indicating that their kanamycin resistance was due to a reactivation of NPTII
expression. However, NPTII levels measured in M3 nrd3-1 and nrd3-2 did not reach that of
K/K control plants. To address whether nrd3-1 and nrd3-2 mutations release RdDM, we

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
analysed ProNOS DNA methylation at the ProNOS-NPTII reporter gene in the TARGET
transgene by methylation-sensitive restriction enzyme cleavage-quantitative PCR (msRE-
qPCR) in M3 mutant plants (Figure 1d). The result showed an almost complete loss of
cytosine methylation in non-CG context (NheI, Alw26I), but only partial reduction in CG
context (Psp1406I), which is a hallmark of mutations affecting RdDM (Kanno et al., 2004;
Kanno et al., 2005; Kanno et al., 2008; Daxinger et al., 2009; Finke et al., 2012a; Finke et
al., 2012b). Cleavage with NcoI, whose recognition site lies outside the region targeted by
RdDM and thus is not methylated, served as control for the accessibility of genomic DNA
(Fischer et al., 2008). To check for possible effects on ProNOS-IR-derived siRNAs, RNA
preparations enriched for small RNAs from M3 nrd3-1 and nrd3-2 were analysed in Northern
blots in comparison to RNA from wild-type K/K;H/H plants and a nrpd2a-55 mutant
defective in the second-largest subunit of Pol IV as a representative for a mutation affecting
RdDM (Finke et al., 2012a). No gross differences in SILENCER transgene-derived 24nt, 22nt
and 21nt siRNAs were detected (Figure 1e). This indicates a defect downstream of siRNA
formation in the analysed nrd3-1 and nrd3-2 mutants.
Map-based cloning and next generation sequencing identifies nrd3-1 and nrd3-2 as new drm2
alleles
M3 nrd3-1 plants were crossed with A. thaliana accession Ler and F2 generation seedlings
obtained from these crosses were screened for individuals resistant to hygromycin (HygR,
SILENCER present) and kanamycin (KanR, TARGET present, homozygous for mutation
releasing RdTGS) (Methods S1). Genotyping of these HygR KanR F2 plants using an Illumina
GoldenGate® genotyping assay (Figure S2) identified a section on chromosome 5 (Figure 2a)
including genes DRM2 (At5g14620) and DRM1 (At5g15380) known to have roles in de novo

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
DNA methylation in RdDM (Cao and Jacobsen, 2002; Cao et al., 2003; Jullien et al.; 2012;
Naumann et al., 2011). PCR amplification and Sanger sequencing of DRM2 in five M3
individuals of nrd3-1 identified consistently a C to T transition at position 2908 in exon 9
according to the TAIR 10 gene model, causing a premature stop codon at position 547 of the
protein (Figure 2b). The resulting protein lacks motives III, IV and V of the DMTase domain,
which are highly conserved among DMTases (Cheng, 1995; Cao et al., 2000) and therefore is
most likely not functional. That nrd3-1 is allelic to drm2 was verified via complementation,
for which the intact coding region of DRM2 under the control of its native promoter
(ProDRM2:DRM2) was amplified from wild-type plants via PCR and introduced into M3
plants of nrd3-1 via Agrobacterium tumefaciens mediated transformation (Figure 2c). The
mutation in nrd3-2 was determined by whole genome sequencing of a M3 nrd3-2 plant in
comparison to a non-mutagenized control K/K;H/H plant using Illumina HiSeq2000®
technology. Sequence comparison identified 1071 unique SNPs between the M3 nrd3-2 plant
and the non-mutagenized control. 80 of these SNPs were predicted to cause non-synonymous,
homozygous mutations in open reading frames of protein coding genes (Table S1). One SNP
was found at position 2467 (relative to A of the START codon) in exon 9 of DRM2 according
to the TAIR 10 gene model (Figure 2b). This mutation causes a premature stop codon at
position 435 of the protein, thus resulting in a protein lacking all conserved DNMTase motifs.
The presence and homozygous nature of the mutation was confirmed by PCR amplification
and Sanger sequencing of DRM2 of five M3 nrd3-2 plants. Thus, in nrd3-2 an additional
drm2 allele was identified.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
CG context methylation is maintained to different degrees at the TARGET ProNOS and
endogenous RdDM tragets in nrd3 mutants
As methylation sensitive restriction cleavage-qPCR (msRE-qPCR) (Figure 1d) can test
methylation only at the suitable restriction sites available, the overall methylation of the
ProNOS in the TARGET ProNOS-NPTII reporter gene in wild-type K/K;H/H as well as in M3
nrd3-1 and nrd3-2 plants carrying drm2 alleles was determined by bisulfite sequencing
(Figure 3a). The results showed that cumulative cytosine methylation in the ProNOS region
undergoing RdDM that is 68% in wild-type K/K;H/H plants is reduced to 27% and 19% in
nrd3-1 and nrd3-2, respectively. This reduction is primarily due to extensive loss of CHH
context and less pronounced loss of CHG context methylation. ProNOS methylation in CG
context, which was 83% in wild-type K/K;H/H plants, is with 74% almost unchanged in M3
nrd3-1 and with 52% moderately reduced in M3 nrd3-2. The persistence of CG context
methylation is in particular illustrated by the Psp1406I site in the TARGET ProNOS, where in
M3 nrd3-1 and nrd3-2 methylation is present in more than a half of DNA molecules
according to msRE-qPCR (Figure 1d) and bisulfite sequencing (Figure S3). In K/K;H/H
plants, methylation is distributed evenly over the whole length of the TARGET ProNOS
despite of a prevalence of cytosines in CG context in its 5’ and in CHH context in its 3’ half
(Figure S3). As persistence of CG context methylation was rather equal over all CG context
sites within the ProNOS, more remaining methylation was found in the 5’ than the 3’
ProNOS half in nrd3-1 and nrd3-2 mutants.
To test whether this effect was specific for the analysed transgenic ProNOS of TARGET Kchr1-
10 (Fischer et al., 2008) or a general feature, DNA methylation at the endogenous RdDM
targets AtSN1 (Zilberman et al., 2003; Kuhlmann and Mette, 2012) (Figure 3b), MEA-ISR
(Cao and Jacobsen, 2002) (Figure 3c) and AtMU1 (Bäurle et al., 2007) (Figure 3d) was

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
determined in M3 nrd3-1 and nrd3-2 by bisulfite sequencing. In addition, DNA methylation
at AtCOPIA4 was analysed as a RdDM-independent control (Johnson et al., 2007) (Figure
3e). In M3 nrd3-1 and nrd3-2 plants, DNA methylation is reduced at AtSN1 in CG, CHG and
CHH context, which differs from the pattern observed for the TARGET ProNOS. In contrast,
CG context methylation is well maintained in nrd3-1 and nrd3-2 at MEA-ISR and in nrd3-1 at
AtMU1, while CHG and CHH context methylation are reduced. Methylation at AtCOPIA4 is
unaltered in all contexts in M3 nrd3-1 and nrd3-2 plants. Thus, methylation patterns at
transgenic and endogenous RdDM targets are affected by the loss of de novo methylation in
nrd3 mutants in a selective way, with AtSN1 acting distinct from the TARGET ProNOS,
MEA-ISR and AtMU1.
Persistence of CG context methylation at the TARGET ProNOS increases with extended
exposure to RNA-directed DNA methylation
The high persistence of CG context methylation at the TARGET ProNOS in M3 nrd3-1 and
nrd3-2 mutants was striking, as this was already the second generation in which the analysed
plants had been homozygous for the nrd3 mutant alleles and thus defective in RNA-directed
DNA methylation (Figure S1). This prompted us to approach the persistence of TARGET
ProNOS CG context methylation in a systematic way by taking advantage of the independent
segregation of the genetically unlinked transgene loci TARGET Kchr1-10 on chromosome 1 and
SILENCER loci H on chromosome 4, respectively, in our system (Fischer et al., 2008). If
K/K;H/- plants homozygous for the TARGET and hemizygous for the SILENCER are allowed
to self-pollinate, 25% of the progeny will be of genotype K/K;H/H, 50% of K/K;H/- and 25%
of K/K, having lost the SILENCER by segregation. In these K/K* individuals (*indicating
that the TARGET transgene was exposed to the SILENCER in the previous generation),

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
maintenance of methylation in absence of the inducing signal can be analysed in wild-type
plants (Figure 4a). The K/K;H/- plants in such a segregating population can serve as a source
for the next generation of K/K* individuals, in which the TARGET now has been exposed for
one generation more to the SILENCER. Parental (P) plants homozygous for either TARGET K
or SILENCER H where crossed and the obtained F1 plants hemizygous for both transgenes
(K/-;H/-) were allowed to self-pollinate. Among the resulting F2 and their follow up progeny,
individuals with the desired K/K;H/- and K/K* genotypes were identified by PCR and
reporter gene activity tests.
TARGET ProNOS methylation was analysed in DNA preparations from rosette leaves of
individual mature plants by msRE-qPCR for members of the F1, F3, F4, and F5 generation
(Figure 4b) and bisulfite sequencing for members of the F1, F2, F3, and F4 generation (Figure
4c). ProNOS DNA methylation in the TARGET transgene reached its final level already when
encountering the SILENCER in the F1 generation and then stayed essentially the same in the
following F2 to the F5 in presence of the SILENCER (Figure 4b top, Figure 4c top, Figure S4).
This applied in particular to CG context methylation (F1 78%, F2 82%, F3 77%, F4 80%)
determined by bisufite sequencing. If at all, there might have been a limited decrease in non-
CG context methylation over generations, as seen at the sites recognised by NheI and Alw26I
in the assay using methylation-sensitive restriction enzyme cleavage and for CHH context
methylation (F1 67%, F2 59%, F3 47%, F4 51%) in bisulfite sequencing data. Further, no
change in the distribution pattern of DNA methylation over the cytosine sites of the TARGET
ProNOS was seen in bisulfite sequencing data over generations (Figure S4). However, we
found indication for slight spreading of DNA methylation into the area 5´ of the region
covered by ProNOS siRNAs. In particular, the first CG site adjacent to the 5´end of the
region covered by ProNOS siRNAs showed methylation after 3 generations in the presence
of the SILENCER (Figure S4, F3 K/K;H/-, F4 K/K;H/-) that was not observed earlier (Figure

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
S4, F1 K/-;H/-). No DNA methylation spread into the area 3´ of the region covered by
ProNOS siRNAs (Figure S4). This is in line with the results from msRE-qPCR for enzyme
NcoI, for which no consistent shift to cleavage inhibition was seen (Figure 4b).
In K/K* individuals that lost the SILENCER by segregation, persisting ProNOS methylation
was first detected at a low level in F2 plants and then at stepwise increasing levels in F3, F4
and F5 plants with increasing numbers of generations of the TARGET having been exposed to
the SILENCER (Figure 4b bottom, Figure 4c bottom). Thus, DNA methylation persistence on
the TARGET ProNOS accumulates over generations. Mainly methylation in symmetric CG
context was inherited, which was consistently detected by msRE-qPCR for Psp1406I (F3, F4,
F5) and by bisulfite sequencing (F2 12%, F3 39%, F4 43%) (Figure S5). Again, analysis of the
Psp1406I site was particularly informative, with methylation detected in more than a half of
DNA molecules in F4 and F5 K/K* plants according to msRE-qPCR (Figure 4b) and F3 and F4
K/K* according to bisulfite sequencing (Figure S5). Persistence of CG context cytosine
methylation was rather equal over all sites present in the TARGET ProNOS, thus showing a
slight preference in the 5´ part of the region that had been covered by SILENCER-derived
ProNOS siRNAs due to the higher prevalence of CG context cytosines (Figure S4; Figure
S5). Further, in F3 and F4 K/K* plants, persistence of some CHG methylation could be
observed at the 5´end of the ProNOS siRNA-target region. Similar as in the analysed F4
K/K;H/- plants, in F4 K/K* plants some DNA methylation was found at the first CG site
adjacent to the 5´end of the ProNOS siRNA-target region. In contrast, the methylation at
asymmetric context cytosines in flanking regions did not persist. In comparison, in control
K/K and K/- plants containing the naive TARGET, only minor apparent ProNOS methylation
was indicated by both methods, most likely resulting from incomplete restriction cleavage or
incomplete chemical conversion of actually unmethylated DNA.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Inherited TARGET ProNOS CG context methylation compromises NPTII expression.
The 43% of CG context methylation together with the lower levels of CHG and CHH
methylation at the TARGET ProNOS detected by bisulfite sequencing in F4 K/K* plants
accounted to in total 14% of methylated cytosines. To see whether this is sufficient to affect
ProNOS-NPTII reporter gene expression, NPTII protein levels in extracts from mature rosette
leaves were measured by ELISA (Figure 5). NPTII levels in F4 K/K* plants were reduced in
comparison to K/K control plants that contained the naive TARGET, showing that even
inheritance of moderate levels of promoter methylation can affect gene expression.
The silenced TARGET ProNOS does not accumulate histone modification H3K9me2
As proposed above, an explanation for the increasing heritability of CG context methylation
at the TARGET ProNOS would be that the ProNOS undergoing RdDM attracts some mark
that accumulates over generations. This is most likely not cytosine methylation itself, as
DNA methylation already reaches its final level in F1 K/-;H/- plants, while little methylation
inheritance is seen in F2 K/K* derived from them. Histone-H3-lysine-9-dimethylation
(H3K9me2) can be associated with transcriptionally silenced transgenes in plants (Foerster et
al., 2011) and thus represents an obvious candidate for such a mark. Chromatin
immunoprecipitation combined with quantitative PCR (ChIP qPCR) was performed to
quantify H3K9me2 relative to total H3 associated with different regions of the TARGET
ProNOS in one-week-old F4 K/K and F4 K/K;H/H seedlings (Figure 6). The tightly silenced
transposable element Ta3 known to be associated with H3K9me2 was used as a reference,
while the active housekeeping gene PHOSPHOFRUCTOKINASE (PFK) served as a negative
control (Johnson et al., 2002; Mathieu et al., 2003; Mathieu et al., 2005). However, no

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
increase of TARGET ProNOS-associated H3K9me2 was visible for the transcriptionally
silenced state, indicating that RdTGS and RdDM of the transgene promoter are independent
of histone modification H3K9me2. Consistently, introgression of mutant suvh4R302 allele
known to affect DNA methylation via formation of H3K9me2 at endogenous silencing
targets (Jackson et al., 2002; Ebbs and Bender, 2006) did not affect TARGET ProNOS
silencing and DNA methylation persistence (Figure S6).
Discussion
Utilising a two-component transgene system with a ProNOS-NPTII target gene highly
susceptible for RdTGS (Fischer et al., 2008; Finke et al., 2012a), two no rna-directed
transcriptional silencing 3 (nrd3) mutants with suppressed RdTGS and RdDM were isolated.
These showed restored resistance to kanamycin and presence of NPTII protein combined
with a strong reduction of asymmetric DNA methylation in the targeted ProNOS while
ProNOS-derived siRNAs were still present in unchanged levels. All these features are
characteristic for mutants defective in RdDM (Law and Jacobsen, 2010; Eun et al., 2012;
Matzke and Mosher, 2014). This was confirmed by the reduced DNA methylation (mainly in
non-CG context) found at the endogenous RdDM targets AtSN1 (Zilberman et al., 2003;
Kuhlmann and Mette, 2012), MEA-ISR (Cao and Jacobsen, 2002) and AtMU1 (Bäurle et al.,
2007). Map-based cloning and next generation sequencing identified in nrd3-1 and nrd3-2
the two new drm2 alleles nrd3-1/drm2-8 and nrd3-2/drm2-9 carrying premature translational
stop codons upstream of the essential DMTase domain of DOMAIN REARRANGED
METYHLTRANSFERASE 2. The identification of DRM2 in the screen for mutants defective
in RdDM is consistent with the predicted predominant role of DRM2 in comparison to DRM1
in RdDM (Cao and Jacobsen, 2002; Cao et al., 2003) and its repeated identification in

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
independent mutagenesis studies (Naumann et al., 2011). The maintenance of up to 70% of
symmetric context ProNOS DNA methylation in our nrd3 lines to the second generation (M3)
is remarkable and was associated with a reduction of NPTII accumulation, but it is unclear
whether this was due to persistent transcriptional silencing or to pleiotropic effects of
mutagenesis. Similar maintenance of RdDM-induced CG context methylation at transgenic
reporter genes in RdDM-releasing mutants has been observed previously (Aufsatz et al.,
2002b; Daxinger et al., 2009; Finke et al., 2012a; Sasaki et al., 2014), but was not followed
up in detail.
At the endogenous RdDM targets AtSN1, AtMU1, and MEA-ISR, CG context methylation
was maintained to different degrees in nrd3 mutants, while CHG and CHH context
methylation was clearly reduced. This is consistent with previous observations for mutants
affected in RdDM, for example nrd1/idn2 or nrd2/nrpd2a (Finke et al., 2012a) or a suvh2
suvh9 double mutant (Kuhlmann et al., 2012). For ATSN1, increased accumulation of
transcripts in the presence of remaining DNA methylation has been seen in several mutants
affecting RdDM (He et al., 2009; Gao et al., 2010), including the suvh2 suvh9 double mutant
showing remaining AtSN1 CG and CNG context methylation levels similar to the ones in
nrd3-1 and nrd3-2 (Kuhlmann et al., 2012). However, as, in contrast to transgenic reporter
genes, completely unmethylated endogenous RdDM targets that would allow the
determination of reference transcript levels are usually not available, the effect of persistent
DNA methylation on transcript accumulation cannot be determined. The maintenance of
symmetric TARGET ProNOS methylation in our material must have been established within
the four generations between the first encounter of TARGET and SILENCER in the F1 and the
M2 generation equivalent to F5 in which the RdDM pathway became first defective.
However, as the level of siRNAs with ProNOS-homology was unaltered in drm2 plants, it
could not be distinguished whether ProNOS CG context methylation was maintained by a

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
mechanism solely based on DNA methylation per se (e.g. via MET1) independent from the
siRNA signal or by a so far unknown mechanism dependent on siRNAs but independent of
DRM2.
In order to address this point and to get insights into a possible cumulative effect of multiple
generations of exposure to the siRNA signal also reported by others (Lunerova-Bedrichova et
al., 2008; Khaitova et al., 2011), persistence of DNA methylation at the TARGET ProNOS
was analysed over successive generations with the SILENCER transgene presence or after
segregating out the SILENCER transgene as the source of ProNOS siRNAs in sexual
reproduction. Consistent with previous observations (Fischer et al., 2008), the particular
TARGET transgene used in our study showed already in the first generation (F1) of encounter
with the SILENCER the final level of approx. 80% cytosine methylation in all contexts. In the
following F3 to F5 generations with the SILENCER present, TARGET ProNOS methylation
stayed essentially the same, with a slight tendency to reduced asymmetric DNA methylation
in later generations. Such a possible partial mutual exclusion of DNA methylation
maintenance (via MET1 at CG context sites only) and RdDM-mediated de novo methylation
(in all contexts) has been discussed previously based on observations made in met1 mutants
(Mathieu et al., 2007). After losing the SILENCER in the F2 by segregation after one
generation of encounter, only little DNA methylation in mainly CG context persisted at the
TARGET ProNOS. However, persisting DNA methylation increased with each following
generation of joint presence of TARGET and SILENCER transgenes. At latest in the F4
generation, more than half of the DNA molecules in the TARGET ProNOS maintained CG
context methylation after segregation of the SILENCER. Thus, from this stage on, it can be
excluded that only one of the two copies present in plants homozygous for the TARGET
remained methylated, for example due to differential inheritance of CG context methylation
via male versus the female gametophytes (Saze et al., 2003). In the F4 generation, the

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
persisting approx. 40% DNA methylation were sufficient to reduce the TARGET ProNOS-
NPTII gene expression by approx. 30%, proving that maintained symmetric context promoter
methylation can substantially reduce gene expression levels. Despite the presence of some
CHG methylation in F3 and F4 K/K* plants, we did not find evidence that the persisting DNA
methylation and its effect on gene expression was related to histone modification H3K9me2,
as the silenced ProNOS was not associated with increased H3K9me2 and a suvh4 mutant
background did not affect reporter gene silencing. This is consistent with the rather limited
involvement of H3K9me2 in RdDM reported by others, for example in the case of trans-
chromosomal methylation induction (Greaves et al., 2014). Loss of histone acetylation via
histone deacetylase HDA6 might be an alternative candidate for involved histone
modifications (Blevins et al., 2014).
In summary, our observations made using a transgene-based experimental system suggest
that setting up DNA methylation at a particular region in the A. thaliana genome can be
divided into an initial phase of immediate, but still fully reversible de novo methylation and
transcriptional gene silencing, that is followed by the setting of so far unknown additional
chromatin mark(s) maintenance over several generations, resulting in efficient maintenance
of cytosine methylation in symmetric sequence context connected with consolidation of gene
silencing (Figure 7). This is important for the role of RNA-directed DNA methylation in
genome-wide epigenetic regulation (Bond and Baulcombe, 2014). In terms of using RNA-
directed DNA methylation of endogenous promoter regions in plant biotechnology (Okano et
al., 2008; Kasai and Kanazawa, 2013), persistence of DNA methylation and transcriptional
silencing after a number of generations would allow “non-transgenic” lines with modulated
gene expression after removal of the SILENCER transgene. Differences in the time scale
needed for the initial de novo methylation to be consolidated by methylation maintenance
might affect the way in which RNA-directed DNA methylation contributes to heterosis

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
(Birchler et al., 2010) via trans-chromosomal changes of DNA methylation (Greaves et al.,
2012; Greaves et al., 2014). Further, it is tempting to speculate that separating short-term
responses from establishing long-term epigenetic memory would allow the plant to integrate
external stress exposure over extended time periods (Luna and Ton, 2012; Thellier and
Lüttge, 2013).
Experimental procedures
Additional detailed information is provided online in supporting experimental procedures
(Methods S1).
Plant material and EMS mutant screen
Transgenic A. thaliana double homozygous for TARGET and SILENCER transgenes (Kchr1-
10/Kchr1-10;H/H) in accession Col-0 have been described in Fischer et al. (2008). Plant
cultivation, testing for antibiotics resistances and PCR-based detection of transgenes was
performed as indicated there. Seed EMS (ethyl-methanesulfonate) mutagenesis was
performed by Lehle Seeds. No rna-directed transcriptional silencing (nrd) mutations were
screened for as described in Finke et al. (2012a) and (2012b).
NPTII protein quantification
Amounts of NPTII protein were determined by ELISA using an Agdia PathoScreen Kit for
NPTII (Agdia), with total protein in the same extracts determined using a Pierce BCA Protein
Assay kit (Pierce).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
DNA methylation analysis and small RNA analysis
ProNOS DNA methylation analysis by methylation sensitive restriction cleavage-qPCR was
done according to Finke et al. (2012a) and (2012b) with data analysis using the ΔΔCt method
according to Pfaffl (2001). Primers are listed in Table S2. Analysis by bisulfite genomic
sequencing was done as described in Kuhlmann and Mette (2012) and Finke et al. (2012a)
using primers listed in Table S3. DNA methylation patterns were analysed using CyMATE
software (Hetzl et al., 2007). For analysis of ProNOS-derived siRNAs and Mir167 as control
(for probe, see Table S2), a RNA preparation enriched in small RNAs was submitted to
Northern blot analysis as described previously (Mette et al., 2005; Finke et al., 2012a).
Mapping of mutations and complementation by Agrobacterium-mediated transformation
Mapping of nrd3-1 was performed using Illumina VeraCode GoldenGate® genotyping (Table
S4) and InDel markers (Salathia et al., 2007; Table S5) as described (Finke et al., 2012a).
The wild-type DRM2 ORF including the 3’UTR and around 450 bp upstream of the
transcriptional start site was amplified from A. thaliana accession Col-0 and cloned in binary
vector pCMBL2 (Finke et al., 2012a) to yield pCMBL2+DRM2. pCMBL2+DRM2 was
introduced into A. tumefaciens strain pGV2260 and transferred to M3 nrd3-1 via the floral dip
method (Clough and Bent, 1998).
Next generation sequencing and bioinformatics analysis
Next generation sequencing (NGS) of plant genomic DNA was performed by IPK’s in-house
sequencing facility using an Illumina HiSeq2000 device according to manufacturer’s
protocols. Sequencing was performed in paired-end read mode (2 times 100 bp). In total, 47
194 081 (M3 nrd3-2) and 62 130 438 (control K/K;H/H) paired-end reads (2 times 100 bp)
were obtained. Sequencing reads were mapped to the A. thaliana reference sequence (version

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
TAIR10; http://www.arabidopsis.org/) using BWA 0.6.1 (Li and Durbin, 2009). The
parameter "-q 15" was used for "bwa aln" to trim off low quality bases. 30 158 845 (63.9%;
M3 nrd3-2) and 56 782 470 (91.4%; control K/K;H/H) reads were mapped as proper pairs to
the A. thaliana nuclear genome to reach on average 22-fold and 41-fold coverage,
respectively. SNP calling was performed with the SAMtools mpileup/bcftools pipeline
version 0.1.18 (Li, 2011) using default parameters. The program snpEff 2.1b (Cingolani et
al., 2012) was used for variant annotation and effect prediction based on the TAIR10 genome
annotation. For the identification of mutations in M3 nrd3-2 in comparison to the non-
mutagenized K/K;H/H control, the respective SNP lists were imported into MS Excel and
compared manually. SNPs occurring in both, mutant and control plants were considered as
pre-existing in the transgenic line submitted to mutagenesis and excluded from further
analysis. Further, SNPs affecting promoters, introns, 5’ and 3’ UTRs as well as “silent”
nucleotide changes in exons not expected to compromise gene function were excluded from
the initial search. The resulting list of gene loci potentially containing the mutation causative
for the release of transcriptional silencing in nrd3-2 is provided in Table S1. Original data
were deposited in the NCBI Short Read Archive under accession number PRJEB6549.
Chromatin immunoprecipitation-qPCR analysis
Chromatin immunoprecipitation (ChIP) was performed as outlined in Kuhlmann and Mette
(2012). Primers used for amplification of Ta3, ProNOS 5´part, ProNOS 3´part and PFK ChIP
are described in Table S6.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acknowledgements
We thank Christa Fricke, Inge Glaser, Beate Kamm, Doreen Stengel and Ines Walde for
excellent technical assistance, Lothar Altschmied, Axel Himmelbach and Uwe Scholz for
helpful discussions and Ingo Schubert and Renate Schmidt for critical comments on the
manuscript. This work received support from the German Research Foundation (DFG)
collaborative research centre (SFB) 648 “Molecular Mechanisms of Information Processing
in Plants” (M.K.) and IPK Gatersleben (A.F.).
Short legends for Supporting Information
Figure S1: Outline of the mutagenesis approach
Figure S2: Rough mapping of mutation nrd3-1
Figure S3: TARGET ProNOS methylation patterns in K/K;H/H wild-type control, M3 nrd3-1
and nrd3-2 plants
Figure S4: TARGET ProNOS methylation patterns in F1, F2, F3 and F4 plants in presence of
the SILENCER
Figure S5: TARGET ProNOS methylation patterns in F2, F3 and F4 plants after segregation of
the SILENCER
Figure S6: Mutation suvh4 does not affect TARGET ProNOS RNA-directed transcriptional
gene silencing and DNA methylation maintenance.
Table S1: Candidate genes for nrd3-2
Table S2: Primers for methylation-sensitive restriction cleavage-quantitative PCR and
miRNA hybridisation

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Table S3: Primers for bisulfite sequencing
Table S4: Illumina GoldenGate® markers for Col-0 / Ler polymorphisms
Table S5: Primers for PCR-based detection of Col-0 / Ler polymorphisms
Table S6: Primers for chromatin immunoprecipitation-quantitative PCR
Methods S1: Supporting experimental procedures
References
Aufsatz, W., Mette, M.F., van der Winden, J., Matzke, A.J.M. and Matzke, M. (2002a)
RNA-directed DNA methylation in Arabidopsis. Proc. Natl. Acad. Sci. USA, 99S4,
16499-16506.
Aufsatz, W., Mette, M.F., van der Winden, J., Matzke, M. and Matzke, A.J. (2002b)
HDA6, a putative histone deacetylase needed to enhance DNA methylation induced
by double-stranded RNA. EMBO J., 21, 6832-6841.
Aufsatz, W., Nehlin, L., Voronin, V., Schmidt, A., Matzke, A.J. and Matzke, M. (2009)
A novel strategy for obtaining kanamycin resistance in Arabidopsis thaliana by
silencing an endogenous gene encoding a putative chloroplast transporter. Biotechnol.
J., 4, 224-229.
Bäurle, I., Smith, L., Baulcombe, D.C. and Dean, C. (2007) Widespread role for the
flowering-time regulators FCA and FPA in RNA-mediated chromatin silencing.
Science, 318, 109-112.
Birchler, J.A., Yao, H., Chudalayandi, S., Vaiman, D. and Veitia, R.A. (2010) Heterosis.
Plant Cell, 22, 2105-2112.
Bird, A. (2002) DNA methylation patterns and epigenetic memory. Genes & Dev., 16, 6-21.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Blevins, T., Pontvianne, F., Cocklin, R., Podicheti, R., Chandrasekhara, C., Yerneni, S.,
Braun, C., Lee, B., Rusch, D., Mockaitis, K., Tang, H. and Pikaard, C.S. (2014) A
two-step process for epigenetic inheritance in Arabidopsis. Mol. Cell, 54, 30-42.
Bologna, N.G. and Voinnet, O. (2014) The diversity, biogenesis, and activities of
endogenous silencing small RNAs in Arabidopsis. Annu. Rev. Plant Biol., 65, 473-
503.
Bond, D.M., and Baulcombe, D.C. (2014) Small RNAs and heritable epigenetic variation in
plants. Trends Cell Biol., 24, 100-107.
Cao, X., Aufsatz, W., Zilberman, D., Mette, M.F., Huang, M.S., Matzke, M. and
Jacobsen, S.E. (2003) Role of the DRM and CMT3 methyltransferases in RNA-
directed DNA methylation. Curr. Biol., 13, 2212-2217.
Cao, X. and Jacobsen, S.E. (2002) Role of the arabidopsis DRM methyltransferases in de
novo DNA methylation and gene silencing. Curr. Biol., 12, 1138-1144.
Cao, X., Springer, N.M., Muszynski, M.G., Phillips, R.L., Kaeppler, S. and Jacobsen,
S.E. (2000) Conserved plant genes with similarity to mammalian de novo DNA
methyltransferases. Proc. Natl. Acad. Sci. USA, 97, 4979-4984.
Cheng, X.D. (1995) Structure and function of DNA methyltransferases. Annu. Rev. of
Biophys. .Biomol. Struc., 24, 293-318.
Cingolani, P., Platts, A., Wang, l.L., Coon, M., Nguyen, T., Wang, L., Land, S.J., Lu, X.
and Ruden, D.M. (2012) A program for annotating and predicting the effects of
single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila
melanogaster strain w1118; iso-2; iso-3. Fly (Austin), 6, 80-92.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for Agrobacterium-
mediated transformation of Arabidopsis thaliana. Plant J., 16, 735-743.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Cokus, S.J., Feng, S., Zhang, X., Chen, Z., Merriman, B., Haudenschild, C.D., Pradhan,
S., Nelson, S.F., Pellegrini, M. and Jacobsen, S.E. (2008) Shotgun bisulphite
sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature,
452, 215-219.
Conte, S., Stevenson, D., Furner, I. and Lloyd, A. (2009) Multiple antibiotic resistance in
Arabidopsis is conferred by mutations in a chloroplast-localized transport protein.
Plant Physiol., 151, 559-573.
Daxinger, L., Kanno, T., Bucher, E., van der, W.J., Naumann, U., Matzke, A.J. and
Matzke, M. (2009) A stepwise pathway for biogenesis of 24-nt secondary siRNAs
and spreading of DNA methylation. EMBO J., 28, 48-57.
Du, J., Zhong, X., Bernatavichute, Y.V., Stroud, H., Feng, S., Caro, E., Vashisht, A.A.,
Terragni, J., Chin, H.G., Tu, A., Hetzel, J., Wohlschlegel, J.A., Pradhan, S.,
Patel, D.J. and Jacobsen, S.E. (2012) Dual binding of chromomethylase domains to
H3K9me2-containing nucleosomes directs DNA methylation in plants. Cell, 151,
167-180.
Ebbs, M.L. and Bender, J. (2006) Locus-specific control of DNA methylation by the
Arabidopsis SUVH5 histone methyltransferase. Plant Cell, 18, 1166-1176.
Eun, C., Lorkovic, Z.J., Sasaki, T., Naumann, U., Matzke, A.J. and Matzke, M. (2012)
Use of forward genetic screens to identify genes required for RNA-directed DNA
methylation in Arabidopsis thaliana. Cold Spring Harbor Symp. Quant. Biol., 77, 195-
204.
Finke, A., Kuhlmann, M. and Mette, M.F. (2012a) IDN2 has a role downstream of siRNA
formation in RNA-directed DNA methylation. Epigenetics, 7, 950-960.
Finke, A., Mette, M.F. and Kuhlmann, M. (2012b) Genetic analysis of RNA-mediated
gene silencing in Arabidopsis thaliana. J. Verbrauch. Lebensm., 7, 27-33.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Finnegan, E.J. and Kovac, K.A. (2000) Plant DNA methyltransferases. Plant Mol. Biol., 43,
189-201.
Fischer, U., Kuhlmann, M., Pecinka, A., Schmidt, R. and Mette, M.F. (2008) Local DNA
features affect RNA-directed transcriptional gene silencing and DNA methylation.
Plant J., 53, 1-10.
Foerster, A.M., Dinh, H.Q., Sedman, L., Wohlrab, B. and Mittelsten, S.O. (2011)
Genetic rearrangements can modify chromatin features at epialleles. PLoS Genet., 7,
e1002331.
Gao, Z., Liu, H.-L., Daxinger, L., Pontes, O., He, X., Qian, W., Lin, H., Xie, M.,
Lorcovic, Z.J., Zhang, S., Miki, D., Zhan, X., Pontier, D., Lagrange, T., Jin, H.,
Matzke, A.J.M., Matzke, M., Pikaard, C.S., and Zhu, J.-K. (2010) An RNA
polymerase II- and AGO4-associated protein acts in RNA-directed DNA methylation.
Nature, 465, 106-109
Greaves, I.K., Groszmann, M., Wang, A., Peacock, W.J. and Dennis, E.S. (2014)
Inheritance of Trans Chromosomal Methylation patterns from Arabidopsis F1
hybrids. Proc. Natl. Acad. Sci. USA, 111, 2017-2022.
Greaves, I.K., Groszmann, M., Ying, H., Taylor, J.M., Peacock, W.J. and Dennis, E.S.
(2012) Trans chromosomal methylation in Arabidopsis hybrids. Proc. Natl. Acad. Sci.
USA, 109, 3570-3575.
He, X.-J., Hsu, Y.-F., Zhu, S., Wierzbicki, A.T., Pontes, O., Pikaard, C.S., Liu, H.-L.,
Wang, C.-S., Jin, H., and Zhu, J.-K. (2009) An effector of RNA-directed DNA
methylation in Arabidopsis is an ARGONAUTE4- and RNA-binding protein. Cell,
137, 498-508.
Henderson, I.R., Deleris, A., Wong, W., Zhong, X., Chin, H.G., Horwitz, G.A., Kelly,
K.A., Pradhan, S. and Jacobsen, S.E. (2010) The de novo cytosine

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
methyltransferase DRM2 requires intact UBA domains and a catalytically mutated
paralog DRM3 during RNA-directed DNA methylation in Arabidopsis thaliana. PLoS
Genet., 6, e1001182.
Hetzl, J., Foerster, A.M., Raidl, G. and Mittelsten, S.O. (2007) CyMATE: a new tool for
methylation analysis of plant genomic DNA after bisulphite sequencing. Plant J., 51,
526-536.
Jackson, J.P., Lindroth, A.M., Cao, X. and Jacobsen, S.E. (2002) Control of CpNpG
DNA methylation by the KRYPTONITE histone H3 methyltransferase. Nature, 416,
556-560.
Johnson, L., Cao, X. and Jacobsen, S. (2002) Interplay between two epigenetic marks.
DNA methylation and histone H3 lysine 9 methylation. Curr. Biol., 12, 1360-1367.
Johnson, L.M., Bostick, M., Zhang, X., Kraft, E., Henderson, I., Callis, J. and Jacobsen,
S.E. (2007) The SRA methyl-cytosine-binding domain links DNA and histone
methylation. Curr. Biol., 17, 379-384.
Jones, L., Ratcliff, F., and Baulcombe, D.F. (2001) RNA-directed transcriptional gene
silencing in plants can be inherited independently of the RNA trigger and requires
MET1 for maintenance. Curr. Biol., 11, 747-757.
Jullien, P.E., Susaki, D., Yelagandula, R., Higashiyama, T. and Berger, F. (2012) DNA
methylation dynamics during sexual reproduction in Arabidopsis thaliana. Curr.
Biol., 22, 1825-1830.
Kanno, T., Bucher, E., Daxinger, L., Huettel, B., Bohmdorfer, G., Gregor, W., Kreil,
D.P., Matzke, M. and Matzke, A.J. (2008) A structural-maintenance-of-
chromosomes hinge domain-containing protein is required for RNA-directed DNA
methylation. Nat. Genet., 40, 670-675.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Kanno, T., Huettel, B., Mette, M.F., Aufsatz, W., Jaligot, E., Daxinger, L., Kreil, D.P.,
Matzke, M. and Matzke, A.J. (2005) Atypical RNA polymerase subunits required
for RNA-directed DNA methylation. Nat. Genet., 37, 761-765.
Kanno, T., Mette, M.F., Kreil, D.P., Aufsatz, W., Matzke, M. and Matzke, A.J. (2004)
Involvement of putative SNF2 chromatin remodeling protein DRD1 in RNA-directed
DNA methylation. Curr. Biol., 14, 801-805.
Kasai, M., and Kanazawa, A. (2013) Induction of RNA-directed DNA methylation and
heritable transcriptional gene silencing as a tool to engineer novel traits in plants.
Plant Biotech., 30, 233-241.
Khaitova, L.C., Fojtova, M., Krizova, K., Lunerova, J., Fulnecek, J., Depicker, A., and
Kovarik, A. (2011) Paramutation of tobacco transgenes by small RNA-mediated
transcriptional gene silencing. Epigenetics, 6, 650-660.
Kuhlmann, M. and Mette, M.F. (2012) Developmentally non-redundant SET domain
proteins SUVH2 and SUVH9 are required for transcriptional gene silencing in
Arabidopsis thaliana. Plant Mol. Biol., 79, 623-633.
Law, J.A. and Jacobsen, S.E. (2010) Establishing, maintaining and modifying DNA
methylation patterns in plants and animals. Nat. Rev. Genet., 11, 204-220.
Li, H. (2011) A statistical framework for SNP calling, mutation discovery, association
mapping and population genetical parameter estimation from sequencing data.
Bioinformatics, 27, 2987-2993.
Li, H. and Durbin, R. (2009) Fast and accurate short read alignment with Burrows-Wheeler
transform. Bioinformatics, 25, 1754-1760.
Lindroth, A.M., Shultis, D., Jasencakova, Z., Fuchs, J., Johnson, L., Schubert, D.,
Patnaik, D., Pradhan, S., Goodrich, J., Schubert, I., Jenuwein, T.,
Khorasanizadeh, S. and Jacobsen, S.E. (2004) Dual histone H3 methylation marks

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
at lysines 9 and 27 required for interaction with CHROMOMETHYLASE 3. EMBO
J., 23, 4286-4296.
Lister, R., O'Malley, R.C., Tonti-Filippini, J., Gregory, B.D., Berry, C.C., Millar, A.H.
and Ecker, J.R. (2008) Highly integrated single-base resolution maps of the
epigenome in Arabidopsis. Cell, 133, 523-536.
Liu, X., Yu, C.W., Duan, J., Luo, M., Wang, K., Tian, G., Cui, Y. and Wu, K. (2012)
HDA6 directly interacts with DNA methyltransferase MET1 and maintains
transposable element silencing in Arabidopsis. Plant Physiol., 158, 119-129.
Luna, E. and Ton, J. (2012) The epigenetic machinery controlling transgenerational
systemic acquired resistance. Plant Signal. Behav., 7, 615-618.
Lunerova-Bedrichova, J., Bleys, A., Fojtova, M., Khaitova, L., Depicker, A., and
Kovarik, A. (2008) Trans-generation inheritance of methylation patterns in a tobacco
transgene following a post-transcriptional silencing event. Plant J., 54, 1049-1062.
Malagnac, F., Bartee, L. and Bender, J. (2002) An Arabidopsis SET domain protein
required for maintenance but not establishment of DNA methylation. EMBO J., 21,
6842-6852.
Mathieu, O., Jasencakova, Z., Vaillant, I., Gendrel, A.V., Colot, V., Schubert, I. and
Tourmente, S. (2003) Changes in 5S rDNA chromatin organization and transcription
during heterochromatin establishment in Arabidopsis. Plant Cell, 15, 2929-2939.
Mathieu, O., Probst, A.V. and Paszkowski, J. (2005) Distinct regulation of histone H3
methylation at lysines 27 and 9 by CpG methylation in Arabidopsis. EMBO J., 24,
2783-2791.
Mathieu, O., Reinders, J., Caikovski, M., Smathajitt, C. and Paszkowski, J. (2007)
Transgenerational stability of the Arabidopsis epigenome is coordinated by CG
methylation. Cell, 130, 851-862.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Matzke, M.A., and Mosher, R.A. (2014) RNA-directed DNA methylation: an epigenetic
pathway of increasing complexity. Nat. Rev. Genet., 15, 394-408.
Mette, M.F., Aufsatz, W., Kanno, T., Daxinger, L., Rovina, P., Matzke, M. and Matzke,
A.J. (2005) Analysis of double-stranded RNA and small RNAs involved in RNA-
mediated transcriptional gene silencing. Methods Mol. Biol., 309, 61-82.
Mette, M.F., Aufsatz, W., van der Winden, J., Matzke, M.A. and Matzke, A.J. (2000)
Transcriptional silencing and promoter methylation triggered by double-stranded
RNA. EMBO J., 19, 5194-5201.
Naumann, U., Daxinger, L., Kanno, T., Eun, C., Long, Q., Lorkovic, Z.J., Matzke, M.
and Matzke, A.J. (2011) Genetic evidence that DNA methyltransferase DRM2 has a
direct catalytic role in RNA-directed DNA methylation in Arabidopsis thaliana.
Genetics, 187, 977-979.
Okano, Y., Miki, D., and Shimamoto, K., (2008) Small interfering RNA (siRNA) targeting
of endogenous promoters induces DNA methylation, but not necessarily gene
silencing, in rice. Plant J., 53, 65-77.
Pfaffl, M.W. (2001) A new mathematical model for relative quantification in real-time RT-
PCR. Nucleic Acids Res., 29, e45.
Pontvianne, F., Blevins, T., Chandrasekhara, C., Feng, W., Stroud, H., Jacobsen, S.E.,
Michaels, S.D. and Pikaard, C.S. (2012) Histone methyltransferases regulating
rRNA gene dose and dosage control in Arabidopsis. Genes Dev., 26, 945-957.
Rozhon, W., Baubec, T., Mayerhofer, J., Mittelsten, S.O. and Jonak, C. (2008) Rapid
quantification of global DNA methylation by isocratic cation exchange high-
performance liquid chromatography. Anal. Biochem., 375, 354-360.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Sasaki, T., Lorkovic, Z.J., Liang, S.C., Matzke, A.J. and Matzke, M. (2014) The ability to
form homodimers is essential for RDM1 to function in RNA-directed DNA
Methylation. PloS One, 9, e88190.
Saze, H., Mittelsten Scheid, O., and Paszkowski, J. (2003) Maintenance of CpG
methylation is essential for epigenetic inheritance during plant gametogenesis. Nat.
Genet., 34, 65-69.
Thellier, M. and Lüttge, U. (2013) Plant memory: a tentative model. Plant Biol., 15, 1-12.
To, T.K., Kim, J.M., Matsui, A., Kurihara, Y., Morosawa, T., Ishida, J., Tanaka, M.,
Endo, T., Kakutani, T., Toyoda, T., Kimura, H., Yokoyama, S., Shinozaki, K.
and Seki, M. (2011) Arabidopsis HDA6 regulates locus-directed heterochromatin
silencing in cooperation with MET1. PLoS Genet., 7, e1002055.
Vongs, A., Kakutani, T., Martienssen, R.A. and Richards, E.J. (1993) Arabidopsis
thaliana DNA methylation mutants. Science, 260, 1926-1928.
Wassenegger, M., Heimes, S., Riedel, L. and Sanger, H.L. (1994) RNA-directed de novo
methylation of genomic sequences in plants. Cell, 76, 567-576.
Woo, H.R., Dittmer, T.A. and Richards, E.J. (2008) Three SRA-domain methylcytosine-
binding proteins cooperate to maintain global CpG methylation and epigenetic
silencing in Arabidopsis. PLoS Genet., 4, e1000156.
Woo, H.R., Pontes, O., Pikaard, C.S. and Richards, E.J. (2007) VIM1, a methylcytosine-
binding protein required for centromeric heterochromatinization. Genes Dev., 21, 267-
277.
Zhong, X., Du, J., Hale, C.J., Gallego-Bartolome, J., Feng, S., Vashisht, A.A., Chory, J.,
Wohlschlegel, J.A., Patel, D.J., and Jacobsen, S.E. (2014) Molecular mechanism of
action of plant DRM de novo DNA methyltransferases. Cell, 157, 1050-1060.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Zilberman, D., Cao, X. and Jacobsen, S.E. (2003) ARGONAUTE 4 control of locus-
specific siRNA accumulation and DNA and histone methylation. Science, 299, 716-
719.
Figure legends
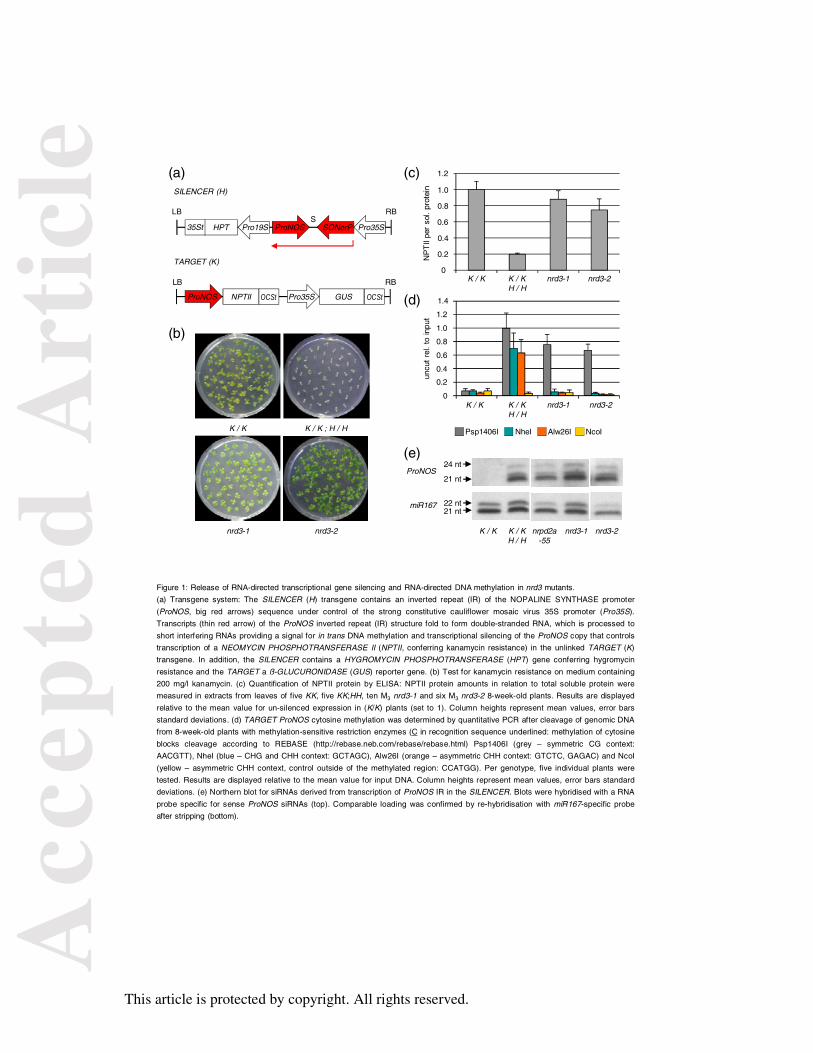
Figure 1: Release of RNA-directed transcriptional gene silencing and RNA-directed DNA
methylation in nrd3 mutants.
(a) Transgene system: The SILENCER (H) transgene contains an inverted repeat (IR) of the
NOPALINE SYNTHASE promoter (ProNOS, big red arrows) sequence under control of the
strong constitutive cauliflower mosaic virus 35S promoter (Pro35S). Transcripts (thin red
arrow) of the ProNOS inverted repeat (IR) structure fold to form double-stranded RNA,
which is processed to short interfering RNAs providing a signal for in trans DNA
methylation and transcriptional silencing of the ProNOS copy that controls transcription of a
NEOMYCIN PHOSPHOTRANSFERASE II (NPTII, conferring kanamycin resistance) in the
unlinked TARGET (K) transgene. In addition, the SILENCER contains a HYGROMYCIN
PHOSPHOTRANSFERASE (HPT) gene conferring hygromycin resistance and the TARGET a
ß-GLUCURONIDASE (GUS) reporter gene. (b) Test for kanamycin resistance on medium
containing 200 mg/l kanamycin. (c) Quantification of NPTII protein by ELISA: NPTII
protein amounts in relation to total soluble protein were measured in extracts from leaves of
five KK, five KK;HH, ten M3 nrd3-1 and six M3 nrd3-2 8-week-old plants. Results are
displayed relative to the mean value for un-silenced expression in (K/K) plants (set to 1).
Column heights represent mean values, error bars standard deviations. (d) TARGET ProNOS
cytosine methylation was determined by quantitative PCR after cleavage of genomic DNA
from 8-week-old plants with methylation-sensitive restriction enzymes (C in recognition

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
sequence underlined: methylation of cytosine blocks cleavage according to REBASE
(http://rebase.neb.com/rebase/rebase.html) Psp1406I (grey – symmetric CG context:
AACGTT), NheI (blue – CHG and CHH context: GCTAGC), Alw26I (orange – asymmetric
CHH context: GTCTC, GAGAC) and NcoI (yellow – asymmetric CHH context, control
outside of the methylated region: CCATGG). Per genotype, five individual plants were
tested. Results are displayed relative to the mean value for input DNA. Column heights
represent mean values, error bars standard deviations. (e) Northern blot for siRNAs derived
from transcription of ProNOS IR in the SILENCER. Blots were hybridised with a RNA probe
specific for sense ProNOS siRNAs (top). Comparable loading was confirmed by re-
hybridisation with miR167-specific probe after stripping (bottom).
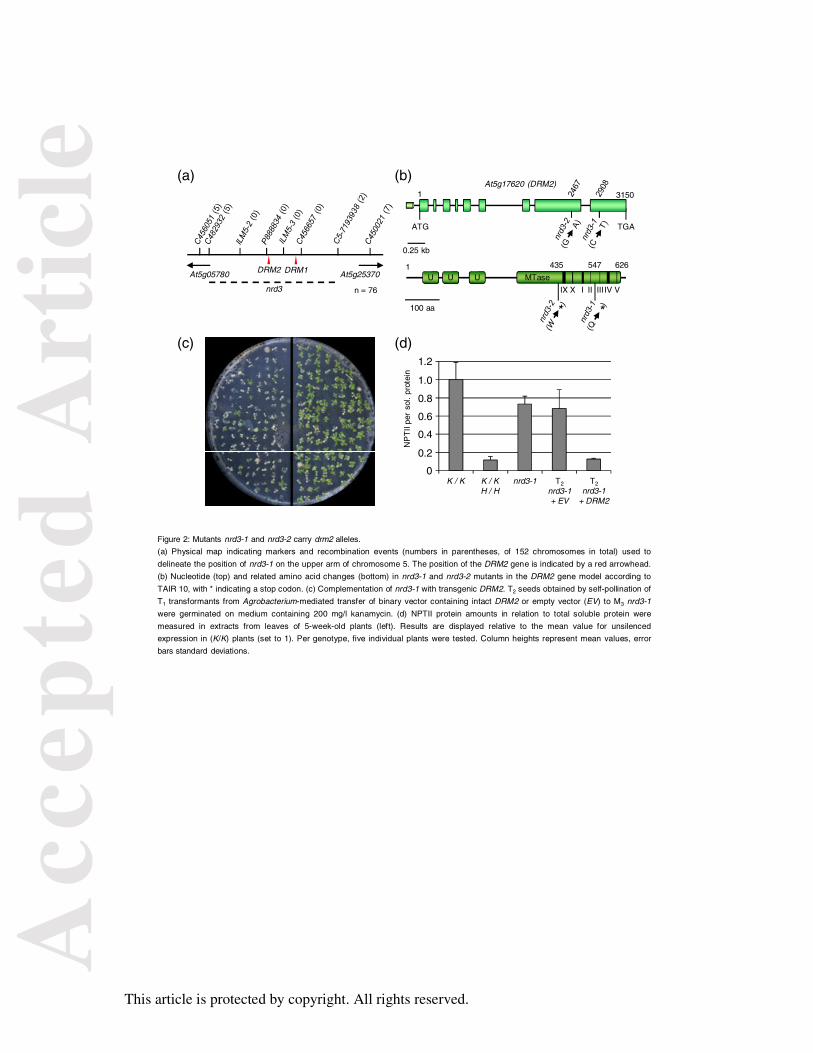
Figure 2: Mutants nrd3-1 and nrd3-2 carry drm2 alleles.
(a) Physical map indicating markers and recombination events (numbers in parentheses, of
152 chromosomes in total) used to delineate the position of nrd3-1 on the upper arm of
chromosome 5. The position of the DRM2 gene is indicated by a red arrowhead. (b)
Nucleotide (top) and related amino acid changes (bottom) in nrd3-1 and nrd3-2 mutants in
the DRM2 gene model according to TAIR 10, with * indicating a stop codon. (c)
Complementation of nrd3-1 with transgenic DRM2. T2 seeds obtained by self-pollination of
T1 transformants from Agrobacterium-mediated transfer of binary vector containing intact
DRM2 or empty vector (EV) to M3 nrd3-1 were germinated on medium containing 200 mg/l
kanamycin. (d) NPTII protein amounts in relation to total soluble protein were measured in
extracts from leaves of 5-week-old plants (left). Results are displayed relative to the mean
value for unsilenced expression in (K/K) plants (set to 1). Per genotype, five individual plants
were tested. Column heights represent mean values, error bars standard deviations.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
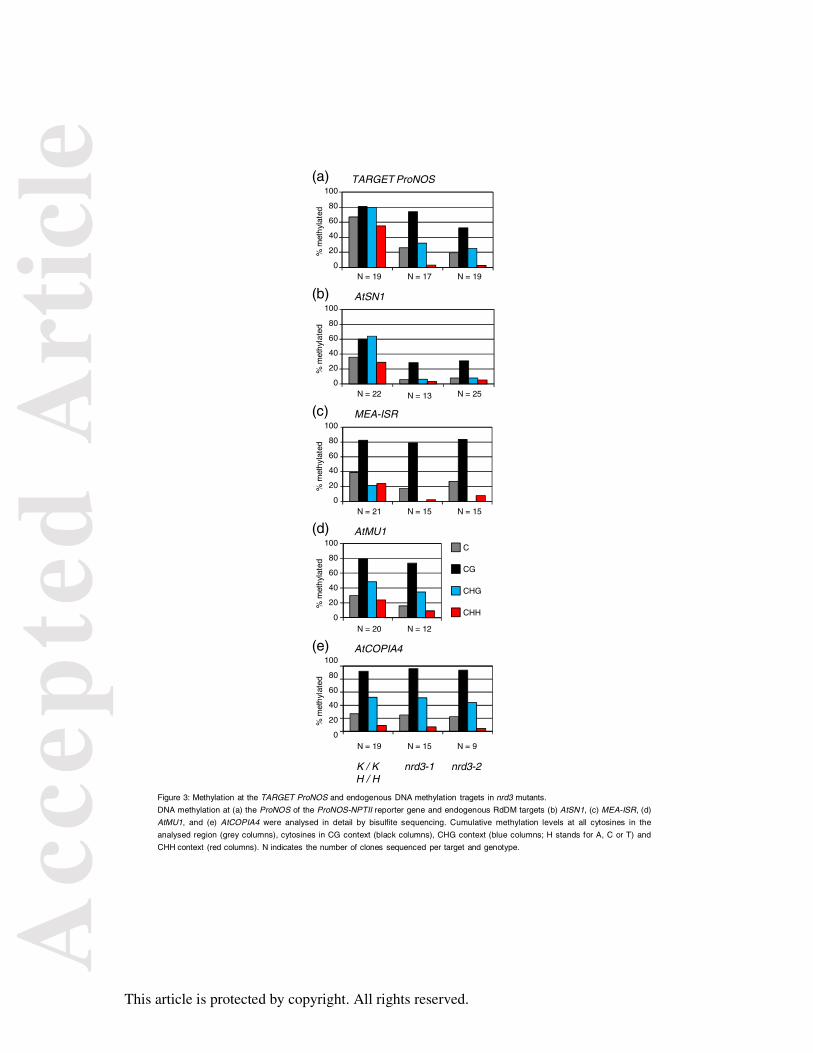
Figure 3: Methylation at the TARGET ProNOS and endogenous DNA methylation tragets in
nrd3 mutants.
DNA methylation at (a) the ProNOS of the ProNOS-NPTII reporter gene and endogenous
RdDM targets (b) AtSN1, (c) MEA-ISR, (d) AtMU1, and (e) AtCOPIA4 were analysed in
detail by bisulfite sequencing. Cumulative methylation levels at all cytosines in the analysed
region (grey columns), cytosines in CG context (black columns), CHG context (blue
columns; H stands for A, C or T) and CHH context (red columns). N indicates the number of
clones sequenced per target and genotype.
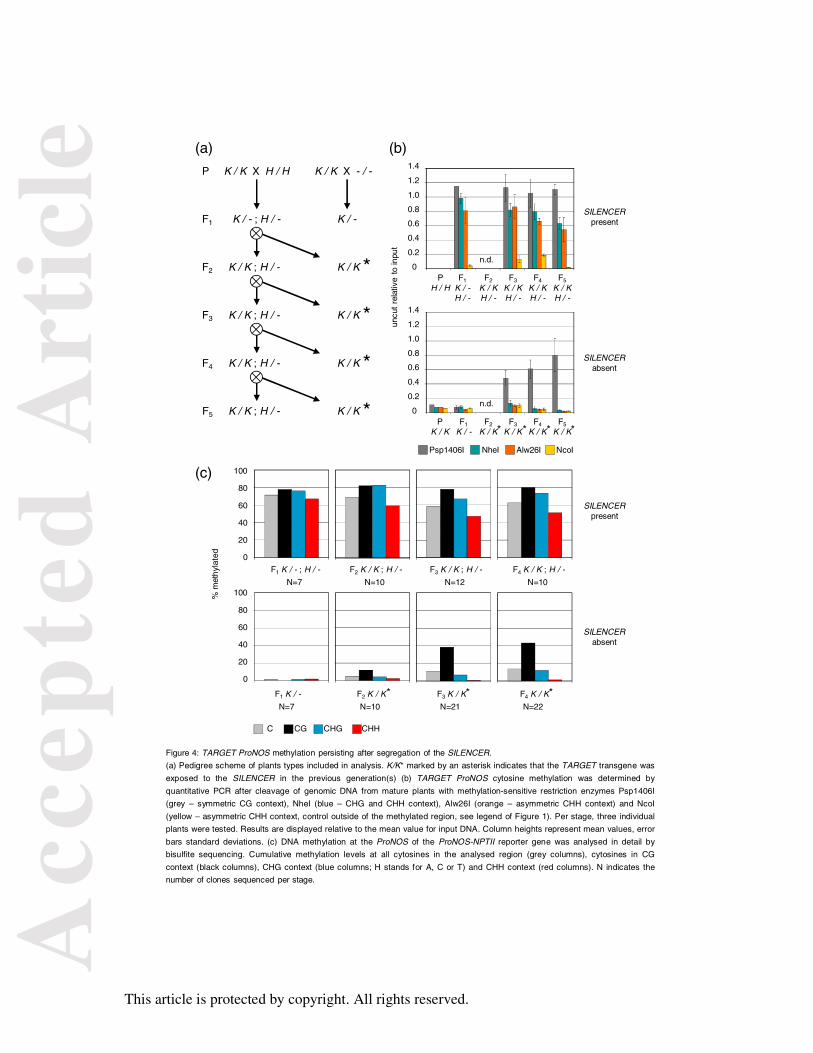
Figure 4: TARGET ProNOS methylation persisting after segregation of the SILENCER.
(a) Pedigree scheme of plants types included in analysis. K/K* marked by an asterisk
indicates that the TARGET transgene was exposed to the SILENCER in the previous
generation(s) (b) TARGET ProNOS cytosine methylation was determined by quantitative
PCR after cleavage of genomic DNA from mature plants with methylation-sensitive
restriction enzymes Psp1406I (grey – symmetric CG context), NheI (blue – CHG and CHH
context), Alw26I (orange – asymmetric CHH context) and NcoI (yellow – asymmetric CHH
context, control outside of the methylated region, see legend of Figure 1). Per stage, three
individual plants were tested. Results are displayed relative to the mean value for input DNA.
Column heights represent mean values, error bars standard deviations. (c) DNA methylation
at the ProNOS of the ProNOS-NPTII reporter gene was analysed in detail by bisulfite
sequencing. Cumulative methylation levels at all cytosines in the analysed region (grey
columns), cytosines in CG context (black columns), CHG context (blue columns; H stands
for A, C or T) and CHH context (red columns). N indicates the number of clones sequenced
per stage.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 5: NPTII expression in presence of persisting TARGET ProNOS CG context
methylation
Quantification of NPTII protein by ELISA: NPTII protein amounts in relation to total soluble
protein were measured in extracts from leaves of 8-week-old plants. Results are displayed
relative to the mean value for un-silenced expression in K/K plants (set to 1). Per genotype,
four (F1 K/-;H/-, F4 K/K;H/-, F4 K/K*) or two (F4 K/K, F4 K/K;H/H) individual plants were
tested. Column heights represent mean values, error bars standard deviations.
Figure 6: H3K9me2 is not increased at the TARGET ProNOS undergoing RNA-directed
DNA methylation.
Chromatin immunoprecipitation combined with quantitative PCR was performed on 1 week
old seedling with antibodies specific to H3 and H3K9me2 and primers specific for transposon
Ta3, TARGET ProNOS 5´side and 3´side regions and housekeeping gene
PHOSPHOFRUCTOKINASE (PFK). Results were calculated separately as the ratio of the
H3K9me2-specific signal to the H3-specific signal in relation to the ratio obtained for Ta3
(set to 1.0) for each chromatin preparation. Column heights indicate the median, error bars
the absolute deviation from five (Ta3, ProNOS 5´side and 3´side) or four (PFK) biological
replicates, respectively.
Figure 7: Transition from immediate response to epigenetic memory over generations
In the first generation G1 of de novo DNA methylation of a promoter via RNA-directed DNA
methylation, transcriptional gene silencing sets in as an immediate response. However, this
silencing is still reversible. Only with the following generations G2, G3 and G4 of ongoing
RNA-directed DNA methylation, DNA methylation maintenance and thus an epigenetic
memory is established.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
K / K K / K ; H / H
nrd3-1 nrd3-2
(b)
K / KH / H
0
0.2
0.4
0.6
0.8
1.0
1.2
NP
TII
per
sol.
prot
ein
nrd3-1 nrd3-2K / K
(c)
(e)ProNOS
miR167
K / K K / KH / H
nrd3-1nrpd2a-55
nrd3-2
24 nt
21 nt
21 nt22 nt
SPro35SProNOS SONorPPro19SHPT35St
LB RB
SILENCER (H)
ProNOS NPTII Pro35SOCSt OCStGUS
LB RB
TARGET (K)
(a)
(d)
K / K K / KH / H
nrd3-1 nrd3-2
Psp1406I NheI Alw26I NcoI
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
uncu
t re
l. to
inpu
t
Figure 1: Release of RNA-directed transcriptional gene silencing and RNA-directed DNA methylation in nrd3 mutants.
(a) Transgene system: The SILENCER (H) transgene contains an inverted repeat (IR) of the NOPALINE SYNTHASE promoter
(ProNOS, big red arrows) sequence under control of the strong constitutive cauliflower mosaic virus 35S promoter (Pro35S).
Transcripts (thin red arrow) of the ProNOS inverted repeat (IR) structure fold to form double-stranded RNA, which is processed to
short interfering RNAs providing a signal for in trans DNA methylation and transcriptional silencing of the ProNOS copy that controls
transcription of a NEOMYCIN PHOSPHOTRANSFERASE II (NPTII, conferring kanamycin resistance) in the unlinked TARGET (K)
transgene. In addition, the SILENCER contains a HYGROMYCIN PHOSPHOTRANSFERASE (HPT) gene conferring hygromycin
resistance and the TARGET a ß-GLUCURONIDASE (GUS) reporter gene. (b) Test for kanamycin resistance on medium containing
200 mg/l kanamycin. (c) Quantification of NPTII protein by ELISA: NPTII protein amounts in relation to total soluble protein were
measured in extracts from leaves of five KK, five KK;HH, ten M3 nrd3-1 and six M3 nrd3-2 8-week-old plants. Results are displayed
relative to the mean value for un-silenced expression in (K/K) plants (set to 1). Column heights represent mean values, error bars
standard deviations. (d) TARGET ProNOS cytosine methylation was determined by quantitative PCR after cleavage of genomic DNA
from 8-week-old plants with methylation-sensitive restriction enzymes (C in recognition sequence underlined: methylation of cytosine
blocks cleavage according to REBASE (http://rebase.neb.com/rebase/rebase.html) Psp1406I (grey – symmetric CG context:
AACGTT), NheI (blue – CHG and CHH context: GCTAGC), Alw26I (orange – asymmetric CHH context: GTCTC, GAGAC) and NcoI
(yellow – asymmetric CHH context, control outside of the methylated region: CCATGG). Per genotype, five individual plants were
tested. Results are displayed relative to the mean value for input DNA. Column heights represent mean values, error bars standard
deviations. (e) Northern blot for siRNAs derived from transcription of ProNOS IR in the SILENCER. Blots were hybridised with a RNA
probe specific for sense ProNOS siRNAs (top). Comparable loading was confirmed by re-hybridisation with miR167-specific probe
after stripping (bottom).
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
DRM2
nrd3 n = 76
At5g05780 At5g25370
(a) (b)
0.25 kb
At5g17620 (DRM2)
ATG TGA
31501
MTase
IX X I II III IV V
U U U1 626
100 aa
435
*
547
*
(c)
T2 nrd3-1 + DRM2 T2 nrd3-1 + EV
T2
nrd3-1+ EV
K / K K / KH / H
T2
nrd3-1 + DRM2
nrd3-10
0.2
0.4
0.6
0.8
1.0
1.2
NP
TII
per
sol.
prot
ein
(d)
DRM1
Figure 2: Mutants nrd3-1 and nrd3-2 carry drm2 alleles.
(a) Physical map indicating markers and recombination events (numbers in parentheses, of 152 chromosomes in total) used to
delineate the position of nrd3-1 on the upper arm of chromosome 5. The position of the DRM2 gene is indicated by a red arrowhead.
(b) Nucleotide (top) and related amino acid changes (bottom) in nrd3-1 and nrd3-2 mutants in the DRM2 gene model according to
TAIR 10, with * indicating a stop codon. (c) Complementation of nrd3-1 with transgenic DRM2. T2 seeds obtained by self-pollination of
T1 transformants from Agrobacterium-mediated transfer of binary vector containing intact DRM2 or empty vector (EV) to M3 nrd3-1
were germinated on medium containing 200 mg/l kanamycin. (d) NPTII protein amounts in relation to total soluble protein were
measured in extracts from leaves of 5-week-old plants (left). Results are displayed relative to the mean value for unsilenced
expression in (K/K) plants (set to 1). Per genotype, five individual plants were tested. Column heights represent mean values, error
bars standard deviations.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
020406080
100
020406080
100
TARGET ProNOS
AtSN1
MEA-ISR
AtCOPIA4
N = 19 N = 17 N = 19
N = 22 N = 13 N = 25
N = 21 N = 15 N = 15
K / KH / H
nrd3-1 nrd3-2
(b)
(c)
(d) AtMU1
N = 20 N = 12
02040
6080
100100
80
60
40
20
0
% m
eth
ylat
ed
(a)
020406080
100%
met
hyla
ted
100
80
60
40
20
0
100
80
60
40
20
0
% m
eth
ylat
ed
(e)
% m
eth
ylat
ed
100
80
60
40
20
0
% m
ethy
late
d
100
80
60
40
20
0N = 19 N = 15 N = 9
C
CG
CHG
CHH
Figure 3: Methylation at the TARGET ProNOS and endogenous DNA methylation tragets in nrd3 mutants.
DNA methylation at (a) the ProNOS of the ProNOS-NPTII reporter gene and endogenous RdDM targets (b) AtSN1, (c) MEA-ISR, (d)
AtMU1, and (e) AtCOPIA4 were analysed in detail by bisulfite sequencing. Cumulative methylation levels at all cytosines in the
analysed region (grey columns), cytosines in CG context (black columns), CHG context (blue columns; H stands for A, C or T) and
CHH context (red columns). N indicates the number of clones sequenced per target and genotype.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 4: TARGET ProNOS methylation persisting after segregation of the SILENCER.
(a) Pedigree scheme of plants types included in analysis. K/K* marked by an asterisk indicates that the TARGET transgene was
exposed to the SILENCER in the previous generation(s) (b) TARGET ProNOS cytosine methylation was determined by
quantitative PCR after cleavage of genomic DNA from mature plants with methylation-sensitive restriction enzymes Psp1406I
(grey – symmetric CG context), NheI (blue – CHG and CHH context), Alw26I (orange – asymmetric CHH context) and NcoI
(yellow – asymmetric CHH context, control outside of the methylated region, see legend of Figure 1). Per stage, three individual
plants were tested. Results are displayed relative to the mean value for input DNA. Column heights represent mean values, error
bars standard deviations. (c) DNA methylation at the ProNOS of the ProNOS-NPTII reporter gene was analysed in detail by
bisulfite sequencing. Cumulative methylation levels at all cytosines in the analysed region (grey columns), cytosines in CG
context (black columns), CHG context (blue columns; H stands for A, C or T) and CHH context (red columns). N indicates the
number of clones sequenced per stage.
(b)(a)
uncu
t re
lativ
e to
inpu
t
(c)
K / K
K / K
K / K
K / K
F5
K / K X - / -
K / -
K / K X H / H
K / K ; H / -
K / - ; H / -
K / K ; H / -
K / K ; H / -
K / K ; H / -
F4
P
F1
F2
F3 *
*
*
*
Psp1406I NheI Alw26I NcoI
% m
ethy
late
d
100
80
60
40
20
0
C CG CHG CHH
n.d.
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
PH / H
n.d.
F1
K / -H / -
F2
K / KH / -
F3
K / KH / -
F4
K / KH / -
F5
K / KH / -
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
PK / K
F1
K / -F2
K / KF3
K / KF4
K / KF5
K / K
n.d.
* * * *
100
80
60
40
20
0
F1 K / -
N=7
F3 K / K
N=21
F4 K / K
N=22
F2 K / K
N=10
* * *
F1 K / - ; H / -
N=7
F3 K / K ; H / -
N=12
F4 K / K ; H / -
N=10
F2 K / K ; H / -
N=10
SILENCERpresent
SILENCERabsent
SILENCERpresent
SILENCERabsent
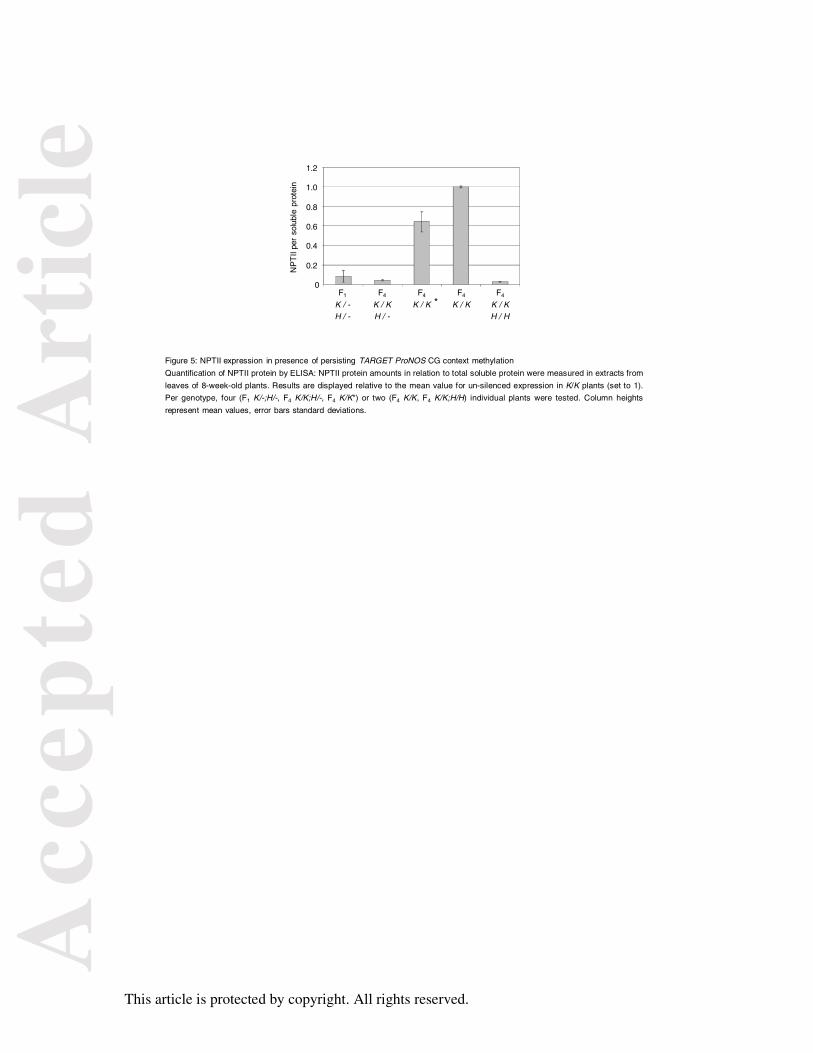
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 5: NPTII expression in presence of persisting TARGET ProNOS CG context methylation
Quantification of NPTII protein by ELISA: NPTII protein amounts in relation to total soluble protein were measured in extracts from
leaves of 8-week-old plants. Results are displayed relative to the mean value for un-silenced expression in K/K plants (set to 1).
Per genotype, four (F1 K/-;H/-, F4 K/K;H/-, F4 K/K*) or two (F4 K/K, F4 K/K;H/H) individual plants were tested. Column heights
represent mean values, error bars standard deviations.
0
0.2
0.4
0.6
0.8
1.0
1.2
F1
K / -H / -
F4
K / KH / -
F4
K / K
F4
K / K
F4
K / KH / H
NP
TII
per
solu
ble
prot
ein
*
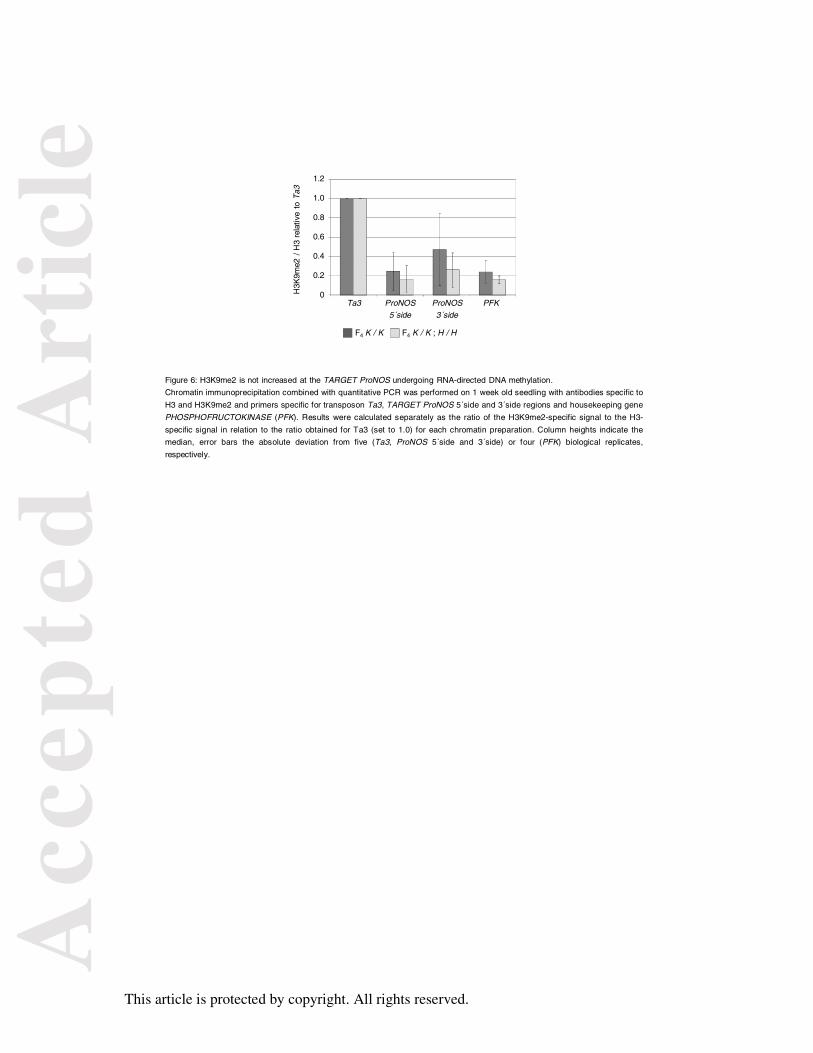
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
F4 K / K F4 K / K ; H / H
Ta3 ProNOS
5´side
ProNOS
3´side
PFK
1.2
1.0
0.8
0.6
0.4
0.2
0H3K
9me2
/ H
3 re
lativ
e to
Ta3
Figure 6: H3K9me2 is not increased at the TARGET ProNOS undergoing RNA-directed DNA methylation.
Chromatin immunoprecipitation combined with quantitative PCR was performed on 1 week old seedling with antibodies specific to
H3 and H3K9me2 and primers specific for transposon Ta3, TARGET ProNOS 5´side and 3´side regions and housekeeping gene
PHOSPHOFRUCTOKINASE (PFK). Results were calculated separately as the ratio of the H3K9me2-specific signal to the H3-
specific signal in relation to the ratio obtained for Ta3 (set to 1.0) for each chromatin preparation. Column heights indicate the
median, error bars the absolute deviation from five (Ta3, ProNOS 5´side and 3´side) or four (PFK) biological replicates,
respectively.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 7: Transition from immediate response to epigenetic memory over generations
In the first generation G1 of de novo DNA methylation of a promoter via RNA-directed DNA methylation, transcriptional gene
silencing sets in as an immediate response. However, this silencing is still reversible. Only with the following generations G2, G3
and G4 of ongoing RNA-directed DNA methylation, DNA methylation maintenance and thus an epigenetic memory is established.
de novomethylation
methylationmaintenance
generation G1 G2 G3 G4
memoryresponse





























