Embed Size (px)
Citation preview

1
Report of Project Hoona – 2003
Diversity of Nilgala Fire Savannah, Sri Lanka: with special reference to its
herpetofauna
Suraj Goonewardene, Zoe Hawke, Vannessa Vanneck
University of Edinburgh
Antoine Drion University of Napier, UK
Anslem de Silva
Herpetologist Sri Lanka
Ravi Jayarathne , Janaka Perera Rajarata University, Sri Lanka
2003

2
List of Contents Page Project Summary 3 Introduction to the project 5 Acknowledgements 7 Study area 9 Introduction Geology and climate Landscape and Physical Anthropological Methodology 11 Results 14 Tree inventory 14 Grassland plants inventory 15 Fauna in Nilgala Fire Savannah Insects, crustaceans, arachnids and Gastropods 16 Amphibians and Reptiles 17 Calodactylodes illingworthorum 18 Hemidactylus maculatus hunae 25 Cnemespis 28 Bird inventory 31 Mammal inventory 32 Folklore and Cultural traits of Veddas 33 Threats to ecosystem 35 Management Recommendations 37 References 38 Photographs 44 Annexure 1 - Accounts Annexure 2 - Notes from Seligman on Veddas Annexure 3 - Journal articles Annexure 4 - 1 KAP Annexure 5 - 2 Reptile survey Annexure 6 - Maps

3
Project summary
a) Funders Davis Fund (University of Edinburgh), James Rennie Bequest (UoE), Student Travel Fund (UoE), Weir Fund (UoE), Peoples Trust for endangered species
b) Title Project: Diversity of Nilgala Fire Savannah,
Sri Lanka: with special reference to its herpetofauna
c) Where research was carried: Main study was conducted at the Nilgala
Fire Savannah, and general observations conducted at Lahugala, Knuckles, Horton Plains, Gampola, and Ampara
d) Team leader: Suraj Goonewardene e) Local consultant: Ans lem de Silva, Herpetologist f) Team: Zoe Hawke
Venessa Vanneck Antoine Droin
Alex Wasserman ( for 1 week)
g) Date of Commencement: 15th June, 2003 h) Date Completed: 7th August, 2003 i) Citation of periodicals reporting work done under this project:
1. PRELIMINARY OBSERVATIONS ON SOME BEHAVIORIAL ASPECTS OF OPHISOPS (REPTILIA: LACERTIDAE) AT NILGALA FIRE-SAVANNAH, SRI LANKA. Anslem de Silva, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck, Antoine Drion, Aaron Bauer and Chris Austin. IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus , special issue) (in press)
2. OTOCRYPTIS WIEGMANNI (REPTILIA: AGAMIDAE): A GECKO
PREDATOR AT NILGALA FIRE-SAVANNAH, SRI LANKA. Antoine Drion, Alexandra Fabry, Anslem de Silva, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck and Aaron Bauer. IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)

4
3. PARASITES OF SOME REPTILIS IN HABITING THE NILGALA FIRE SAVANNAH, SRI LANKA. R.P.V.J.Rajapaksa, Anslem de Silva, Suraj Goonawardena, and Aaron Bauer. IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)
4. SOME CLUTURAL TRAITS OF THE INHABITATNTS OF THE NILGALA
FIRE SAVANNAH, SRI LANKA TOWARDS ANIMALS. J. Perera, Ravi Jayarathne, Anslem de Silva, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck and Aaron Bauer.IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)
5. SOME CLUTURAL TRAITS TOWARDS GECKOS BY THE INHABITATNTS
OF THE NILGALA FIRE SAVANNAH, SRI LANKA. J. Perera, Ravi Jayarathne , Anslem de Silva, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck and Aaron Bauer.IN: Gecko (Journal of the Global gecko Association) (in press)
6. THE HERPETOFAUNA OF THE NILGALA FIRE SAVANNAH, SRI LANKA.
Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck, Antoine Drion, Aaron Bauer and Chris Austin. J. Perera, Ravi Jayarathne , Anslem de Silva, and Aaron Bauer.IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)
7. ECOLOGY OF CALODACTYLODES ILLNGWORTHORUM (REPTILIEA:
GECKONIDAE) IN SRI LANKA: PRELIMINARY FINDINGS. Anslem de Silva, Aaron Bauer, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck and Antoine Drion. IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)
8. CALODACTYLODES ILLNGWORTHORUM (REPTILIEA: GECKONIDAE)
EGGS: SOME THREATS OBSERVED IN NILGALA FIRE SAVANNAH, SRI LANKA. Anslem de Silva, Aaron Bauer, Suraj Goonawardena, Zoe Hawke, Vanessa Vanneck and Antoine Drion. IN: Anslem de Silva (Eds), Proceedings of the Fourth World Congress of Herpetology (part): and recent research on reptiles of Sri Lanka. (Lyriocephalus, special issue) (in press)

5
Introduction to the project
Additions: Mayers “hotspot”, highest number of amphibian species per unit area in the
world (WHT). Reptile 4th highest, Asian region highest number of amphibian, reptile,
mammal, tree species per unit area
Sri Lanka has many unique ecosystems, each with its own charm and specific flora and
fauna. Of these some of the best known are Sinharaja (World Heritage Site), Ritigala,
Knuckles, Horton Plains, and Nilgala Fire Savannah. The present work deals with the
Nilgala Fire Savannah ecosystem.
Preliminary studies were conducted during June 17th 2003 to August 19th, 2003. This
work investigated the nature of the fauna, with special reference to the herpetofauna and
characteristics of their habitats, flora, and archeology of Nilgala Fire Savannah and
cultural traits of its inhabitants. The objectives were to record and identify the
herpetofaunal species inhabiting the Canopy, assess their relative abundance, population
status, interspecies associations, habitat features, distribution within the park and to
identify the threats to these animals and their habitats. On the basis of this data,
recommendations regarding future research directions and management of the Nilgala
Fire Savannah could be put forward. The field technique employed was to make an
inventory of the amphibians and reptiles found in Rathugala and Nilgala mountains
(popularly ending with the suffix “gala” = rock), forest and anthropogenic vegetation in
the Nilgala Fire Savannah. Estimation of species diversity, relative abundance,
population densities and threats faced by amphibians and reptiles were also conducted.
Additional nocturnal investigations were conducted on the amphibian and reptile species
inhabiting the forests.
During the investigations 35 species of reptiles and 15 species of amphibians were
located and identified. Of the species of amphibians and reptiles observed during the
survey, 15 more specimens could not be identified using available literature; thus these
species may be new to science and currently their DNA is being checked to prove these
differences in a genetic level in the US by Aaron Bauer Ph.D. Villanova University and
Chris Austin Ph.D., University of North Dakota.

6
The relative abundance, population densities, food and feeding habits and reproductive
patterns, and some of the threats faced by these animals and to their habitats were
studied.
The results of this survey have highlighted the urgent need to conduct detailed studies
on the following major aspects:
1. To quantify and take steps to minimize threats to amphibians and reptiles, and to their
habitats in this unique ecosystem.
2. To mount in situ and ex situ breeding programs for the un-described reptiles and
amphibian species.
3. To conduct an awareness program and also to educate the inhabitants of the savannah
on the sustainable utilization of wild species.
4. This study and earlier studies have indicated that a significant number of animals are
killed by vehicles on roads within Nilgala Fire Savannah area. Further studies will
build a better picture of the extent of this problem which can then be dealt with in
appropriate ways with the help of local authorities (Dept. of Forests and Wildlife
staff). Measures such as road humps and road signs warning of animals crossing and
to reduce speed are some of the options.
5. To conduct more extensive investigations on fossorial reptiles such as Nessia and
Uropeltids which inhabit the fire savannah. Rare species of Acontinae (Sauria:
Scincidae) have been ‘accidentally’ discovered by excavating the upper soil layers.

7
Acknowledgements:
We thank the Edinburgh University Courts and the University Expeditions Panel for
accepting this proposal and making this expedition possible.
?? Davis Fund (University of Edinburgh)
?? James Rennie Bequest (University of Edinburgh)
?? Student Travel Fund (University of Edinburgh)
?? Weir Fund (University of Edinburgh)
?? Peoples Trust for endangered species
for providing grants for this expedition.
Anslem de Silva, for attending to all local arrangements, accompanying us in the field
and teaching us field techniques, identification etc.
Director General, Dept. of Wildlife Conservation and Assistant Director, Dept. of
Wildlife Conservation, Eastern for permission to conduct the project.
Director General, Dept of Forests and Divisional Forest Officer, Monaragala for
permission to conduct the project. , Mr. Sisira Kumara, Forest Officer, Bibile for his
support throughout the project.
Dr. Manel Gunawardena, Head Dept. of Zoology, Rajarata University, Polgolla, for her
support and all herpetology students of that institution for their support
Dr. R. P. V. J. Rajapaksa, Faculty of Veterinary Science, University of Peradeniya for
parasitological investigations. Kelum Nalinda Manamendra-Arachchi for help in
identification of some amphibians.

8
Dr. Neil Stuart from the Edinburgh University Geography department on how GPS
would be used (i.e. using in combination with GIS we can model population distribution,
habitats, diversity indicators which can be used in management plans, education etc.)
Dr. Scot Ramsay regarding methodology (Edinburgh University Biology department.)
Erik McClements and Alex Wasserman who were brilliant in helping us to construct this
web page.
Margaret Jackson (expedition’s committee secretary) for all the support and patience
from start to finish.
Exped (Edinburgh University Expeditions Society) particularly to Nina Rzechorzek and
Alex Phythian-Adams. And Bjorn Schulte Herbruggen (Project leader of Save the
monkeys, Peru) for all the helpful tips.
Derek Scott of the BTO labs for providing the equipment which was vital to carry out the
work in the field.
Edinburgh Zoo for providing us with handling experience with reptiles and the use of
their library.
Police Officers in charge of Bibile, Bulupitiya for their support.
Villagers at Serawa especially Mr.Gune , TB Lokubanda, “Chutta” (Nihal Luxman) and
his wife and host of other guides and helpers at Nilgala. Our driver Mr. Dissanayake .
Our parents for their support and encouragement.
Finally Anslem de Silva helped with the report by providing valuable suggestions and
comments.
9
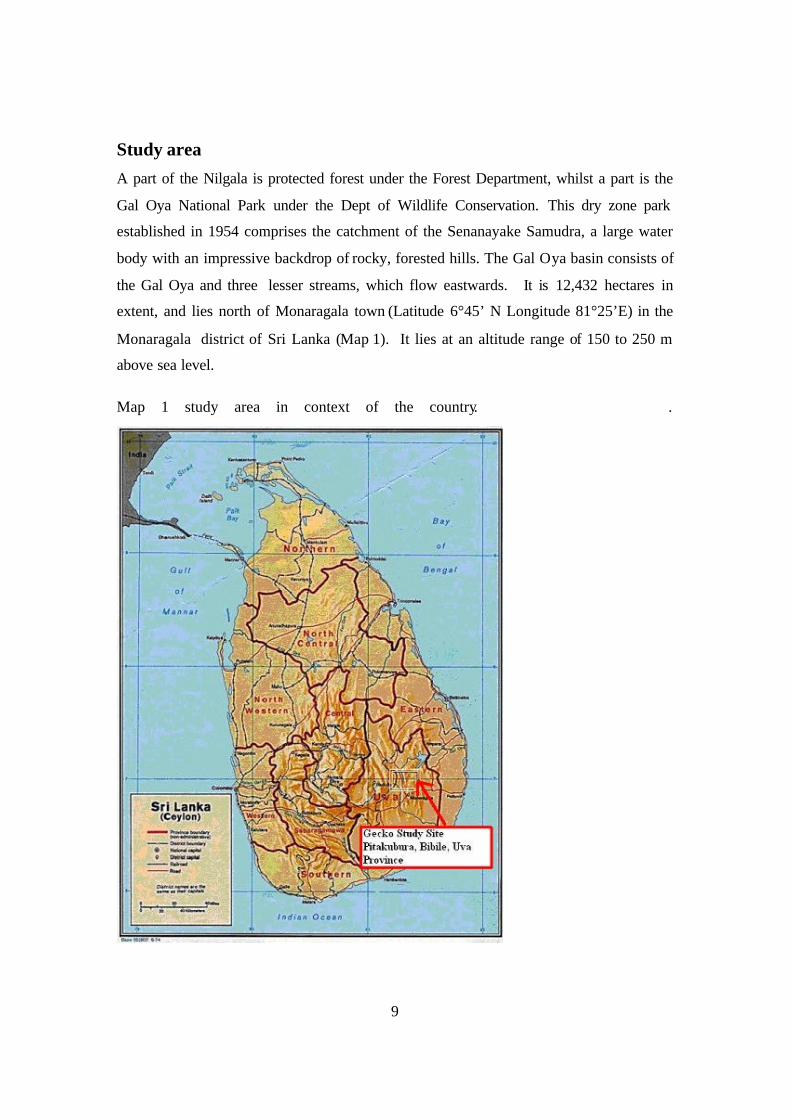
Study area
A part of the Nilgala is protected forest under the Forest Department, whilst a part is the
Gal Oya National Park under the Dept of Wildlife Conservation. This dry zone park
established in 1954 comprises the catchment of the Senanayake Samudra, a large water
body with an impressive backdrop of rocky, forested hills. The Gal Oya basin consists of
the Gal Oya and three lesser streams, which flow eastwards. It is 12,432 hectares in
extent, and lies north of Monaragala town (Latitude 6°45’ N Longitude 81°25’E) in the
Monaragala district of Sri Lanka (Map 1). It lies at an altitude range of 150 to 250 m
above sea level.
Map 1 study area in context of the country. .

10
The terrain is flat with veins of boulders and occasional mountains (figures 56). The
dominant vegetation covers of extensive grasslands (figures 47, 48) and the dominant
tree species in the savannah are the Aralu (Terminalia chebula), Bulu (Terminalia
bellerica) and Nelli (Phyllanthus emblica). Kahata (Careya arborea). There are many
valuable timer trees such as Kruwala (Diospyros crassiflora-Ebony) confined to the
mountains. Swietenia Macrophylla (mahogany), Tectona grandis (teak) was a rapidly
depleting resource due to the exploitation of this timber in natural forests.
The lentic habitats consist of few “Tanks” – figures 55. These are man commencing pre
Christian times for agricultural purposes. The Nilgala savannah has many pre and post
Christian settlements and caves in habited by man, figures 79 (showing dip ledge sign of
prehistoric habitation). Including a road constructed during pre Christian times, which
runs from one part of the savannah. The following vegetation zones can be distinguished:
1. Savannah figures 47, 48
2. Monsoon forests figures 49
3. Anthropogenic vegetations (paddy fields and chena cultivations) figures 50,51,58,60
4. Tertiary vegetation (weed species eg: Eraminiya Zizyphus napeca and Katuhinguru
Lantana camera) figures 83
Description of Nilgala Fire Savannah
Nilgala is situated in the Uva Province and Moanaragala District. It approximate area is
12,432 hectares. The Geographic Coordinates of Nilgala savannah are: 81º 20’, 7º 12’
The Altitude varies from 150 – 250 m in the savannah and the scattered mountains may
raise up to 350 – 750 m. Part of Nilgala savannah is owned by the Forest department and
one part by the Dept. of Wildlife Conservation.
Geology and Landscape
Low Country – major soil groups Reddish Brown Earth, immature brown loams and low
humic gley soils. Terrain is scattered with “Rock knob plains and eroded lands” and
“erosional remnants”, alluvial soils of variable drainage and texture; flat terrain. Terrain
is rolling, hilly and undulating (Figure 56).

11
The type of rock according to PG Cooray (1967) is pre Cambrian Vijayan Complex.
Climate
Dry zone - average annual rain fall (75 % expectancy value) > 1500 mm. – temperature –
north east monsoon (rain during October to January)
Methodology
The study was conducted during the period of 20th June 2003 and 04th August 2003. A
total of approximately 248 hours were spent in the field.
Field methods
1. The field map was compiled using the Survey Department maps of Bibile (sheet No.
63 (1: 50,000) published 2002) and Padiyatalawa (sheet No. 65 (1: 50,000)
published in 1989) (Map 2). An enlarged photocopy (40 x 40 cm) in which the
vegetation zones, rock caves, mountains were highlighted for clarity in the field. This
map was laminated so that it could be used in the field even when raining.
2. Canopy quadrates (10m x 10m) for sampling forest canopy were adopted by de Silva
(2001) was adopted in the present study. A white nylon chord encircling 10 x 10
meter plot was demarcated. All trees over 250 cm girth at breast height (gbh) were
selected for the survey.
3. The following tools were taken by the persons who climb trees: penlight torch,
measuring tape, flexible probe to check tree holes, pH strips, thermometer, hand axe
to widen the tree holes if required and a cloth bag tied to a 15 m long nylon rope.
4. Environmental data within the quadrate were recorded in the structured survey form
(Annex 1) while the field survey proceeded. Field notes were recorded in a field
notebook. The altitude in meters (measured using Compens Altimeter (Sundo-Alpina,
Germany), and relative humidity 1.5 m above the ground inside the quadrate were
recorded, the latter using a Hair-Hygrometer (Gischer - Germany). Air temperature

12
(one and half meters above the ground), and soil temperature (one cm inside soil) in
centigrade, soil type, and litter depth.
5. The GPS location reading inc luding the altitude was recorded of all study locations
including places where geckos, skinks and other reptiles were observed
6. If an amphibian was present in a tree hole, the measurements of this hole was
recorded, if water is present the pH and the temperature inside the hole too were
recorded. Time of detection, height of the particular tree and the height of the hole
from ground, the tree species too were recorded. In most instances the particular tree
was photographed (Figure 24, 25).
7. Identity of the species (if possible) and its sex, length from tip of the snout to the vent
(SV length), measurements taken in millimeters using a micromatic 1m steel tape,
status (gravid or not), presence of ecto-parasites and other defects if observed too
were recorded in the survey form (Annex 2)
8. Stools samples were collected into special vials with 10% formaldehyde solution.
These were labeled and numbered to be later taken to University of Peradeniya for
investigations.
9. Amphibians and reptiles were identified using manuals by Deraniyagala (1953), de
Silva (1990), Dutta and Manamendra-Arachchi (1996), and Greer (1991).
10. Botanical specimens obtained were identified at the Herbarium, Royal Botanical
Gardens, Peradeniya. Manuals Ashton et al (1997) and Gunatilleke (1996) too were
used.
11. Invertebrate fauna inhabiting the particular tree too were recorded.
12. The mountain-wise inventory data of amphibians and reptiles are given in Table 04.

13
13. Visual encounter searches (VES) were the main method used as well as Patch
Encounter Studies (PES). The specimens encountered were collected, measured,
sexed, and checked for defects, gravid females recorded. All animals were released
back in the spots where they were collected.
14. Random stomach contents were obtained for analysis from a few reptiles and
amphibians by using an intragastric cannula and distilled water. The distilled water
was gently squirted into the stomach, while the mouth of the animal was held
downwards, thereby flushing the stomach contents, which were collected into sterile
bottles. The animals were subsequently released in the same locality. The stomach
contents were preserved in 10% formaldehyde and examined in the laboratory under a
dissecting microscope.
15. Voucher specimens of geckos and skinks (male and female, usually a single
specimen) were collected for taxonomical investigations (with the approval from the
Dept. of Wildlife Conservation (DWLC), and Forest for Anslem de Silva).
16. Observations were made of road kills of amphibians and reptiles along study are road.
17. Postmortem examinations were conducted on road kill animals.
18. All the species of amphibians, reptiles, invertebrates, parasites collected and habitats
(terrestrial and lentic) were photographed using Minolta Dynax 7, Nikon CoolPix 775
and Pentax M250 cameras using Minolta 35-70 with macro, Minolta 70-300
telemacro, 2x and 4x close-up adapter lenses. Fuji Chrome and Fuji Colors, 800, 400
and 200 ASA, 36 exposure films were used).
19. Statistical analysis: data were analyzed using SAS. Area of the vegetation habitats
from Arc-info package, map by the GIS Unit of the Natural Resource Center,
Peradeniya, were used for calculating the populations in respective vegetation areas.

14
20. The temperature and relative humidity was recorded using Max-Min thermo
hygrometer.
Results
The results of the surveys carried out for amphibians, reptiles and their habitats during
the project are given in Tables 4 and 5.
The vegetation in Nilgala Fire Savannah
Table 02 Trees / Timber in Bible Region Observed during the expedition
The following tree species were observed by the team during field work. 1) Euphorbia thymofolia 2) Scutia myrtina 3) Adina cordifolia 4) Diospyros ebenum 5) Diospyros quaestia 6) Diospyros oocarpa 7) Albizia odoratissima 8) Berrya cordifolia 9) Terminalia arjuna 12) Alseodaphne semecarpifolia 13) Madhuca longifolia / neriifolia 14) Terminalia chebula 15) Terminalia belerica 16) Terminalia catappa 17) Phyllanthus emblica 18) Schleichera oleosa 20) Woodfordia fructicosa 23) Strychnos potatorum 24) Myristica dactyloides 25) Mangifera zeylanica 26) Sterculia foetida 27) Salmalia insignis / malabarica 28) Alstonia scholaris 29) Tetrameles nudiflora
30) Muntingia calabura 32) Dialium ovoideum 33) Vitex pinnata 34) Mesua ferrea 35) Garallia brachiata 36) Garcinia morella 37) Urandra apicalis 38) Chukrasia velutina 40) Phyllanthus indicus 41) Syzygium operculatum 42) Syzygium assimile / cordifolium 43) Cassia fistula 44) Cassia siamea 45) Pongamia pinnata 46) Soymida febrifuga 47) Neolitsea cassia 48) Litsea glutinosa 50) Gmelina arborea 53) Bridelia retusa 54) Careya arborea 58) Hydnocarpus veneata 59) Sterospermum personatum
The following trees were not listed in ‘The glossary of Sinhala and Tamil names of the plants of Sri Lanka’. Hence, the local vernacular names collected are listed. 10) Wilan 11) Panakka

15
19) Patollkanda 21) Bakini 22) Damania 31) Mora 39) Bathik 49) Helemba 51) Dadakirala
52) Galkulu 55) Nabada 56) Riti 57) Kalupennie 60) Hagilla 61) Bala 62) Lawennie
Table 03 Medicinal plant species observed during the expedition
List of medicinal plants 1. Wattakaka volubilis (L.f.) Stapf 2. Paederia foetida L. 3. Tephrosia purpurea (L.) Pers. 4. Ipomoea mauritiana Jacq. 5. Eclipta prostrate (L.) L. 6. Amorphophallus paeoniifolius (D.) 7. Terminalia arjuna (Roxb.) Wight 8. Caesalpinia bonduc (L.) Roxb. 9. Salicia reticulate Wight. 10. Bauhinia variegate L. 11. Aloe Barbadensis Mill 12. Pterocapus marsupium Roxb. 13. Leea indica (Burm.f.) Merr. 14. Centella asiatica (L.) Urban 15. Garcinia quaesita Pierre. 16. Croton tiglium L. 17. Piper longum L. 18. Operculina turpethum (L.) S. Manso 19. Costus speciosus 20. Oroxylum indicum (L.) Vent 21. Euphorbia antiquorum L. 22. Berberis aristata DC. 23. Trichosanthes cucumerina L. 24. Celastrus paniculatus Willd. 25. Cissampelos pareira L. var. hirsute
26. Pagiantha dichotoma (Roxb.) 27. Citrus aurantifolia 28. Calophyllum inophyllum L. 29. Vitex negundo L. 30. Phyllanthus embilica L. 31. Stereospermum suaveolens DC. 32. Phyllanthus debilis Klein ex Willd. 33. Boerhavia diffusa L. 34. Entada pusaetha DC. 35. Aervalanata (L.) Juss. ex Schult. 36. Semecarpus coriaceae Thw. 37. Erythroxylum moonii Hochr. 38. Aegle marmelos (L.) Correa. 39. Woodfordia fruticosa (L.) Kruz. 40. Premna obtusifolia R. Br. 41. Crotalaria laburnifolia L. 42. Atlantia rotundifolia (Thw.) Tanaka. 43. Cassia auriculata L. 44. Crataeva adansonii DC. ssp. odora 45. Coscinium fenestratum (Gaertn.) 46. Aristolochia indica L. 47. Jasminum grandiflorum L. 48. Vetiveria zizanioides (L) Nash 49. Santalum album L. 50. Osbeckia octandra (L.) DC.

16
Fauna in Nilgala Fire Savannah Insects
1. stick insects 2. praying mentis 3. butterflies 4. dragon flies 5. moths 6. bees and beetles
Crustaceans and arachnids Crabs Cardina Spiders Scorpions Centipedes Millipedes Gastropods Eight species of gastropods and one aquatic species were observed during the survey, which currently being identified in the University of Peradeniya. Amphibians and Reptiles The amphibian and reptile species observed during the expedition are listed in table below.
Table 04 Amphibian and reptile species observed during the expedition
Species list of Bibile: Species name Common name Endemic Number
observed
Testudines
1 Melanochelys trijuga Black turtle No 3 2 Geochlone elegans Indian star tortoise No 1 Agamidae 3 Calotes calotes Green garden lizard No 3 4 Calotes ceylonensis Painted lip lizard Yes 1 5 Calotes versicolor
versicolor Common garden lizard No Many
6 Otocryptis wiegmanii Sri Lankan Kangaroo lizard Yes Many 7 Sitana ponticeriana Fanthroat lizard No 2
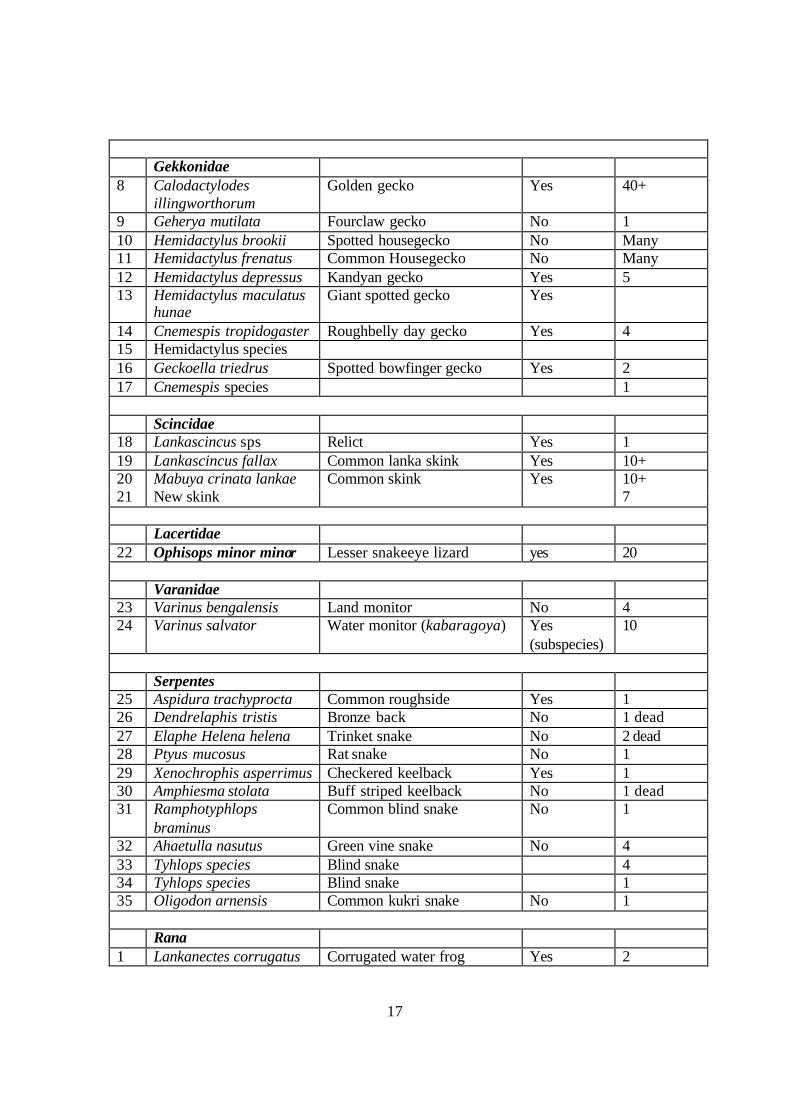
17
Gekkonidae 8 Calodactylodes
illingworthorum Golden gecko Yes 40+
9 Geherya mutilata Fourclaw gecko No 1 10 Hemidactylus brookii Spotted housegecko No Many 11 Hemidactylus frenatus Common Housegecko No Many 12 Hemidactylus depressus Kandyan gecko Yes 5 13 Hemidactylus maculatus
hunae Giant spotted gecko Yes
14 Cnemespis tropidogaster Roughbelly day gecko Yes 4 15 Hemidactylus species 16 Geckoella triedrus Spotted bowfinger gecko Yes 2 17 Cnemespis species 1 Scincidae 18 Lankascincus sps Relict Yes 1 19 Lankascincus fallax Common lanka skink Yes 10+ 20 Mabuya crinata lankae Common skink Yes 10+ 21 New skink 7 Lacertidae 22 Ophisops minor minor Lesser snakeeye lizard yes 20 Varanidae 23 Varinus bengalensis Land monitor No 4 24 Varinus salvator Water monitor (kabaragoya) Yes
(subspecies) 10
Serpentes 25 Aspidura trachyprocta Common roughside Yes 1 26 Dendrelaphis tristis Bronze back No 1 dead 27 Elaphe Helena helena Trinket snake No 2 dead 28 Ptyus mucosus Rat snake No 1 29 Xenochrophis asperrimus Checkered keelback Yes 1 30 Amphiesma stolata Buff striped keelback No 1 dead 31 Ramphotyphlops
braminus Common blind snake No 1
32 Ahaetulla nasutus Green vine snake No 4 33 Tyhlops species Blind snake 4 34 Tyhlops species Blind snake 1 35 Oligodon arnensis Common kukri snake No 1 Rana 1 Lankanectes corrugatus Corrugated water frog Yes 2
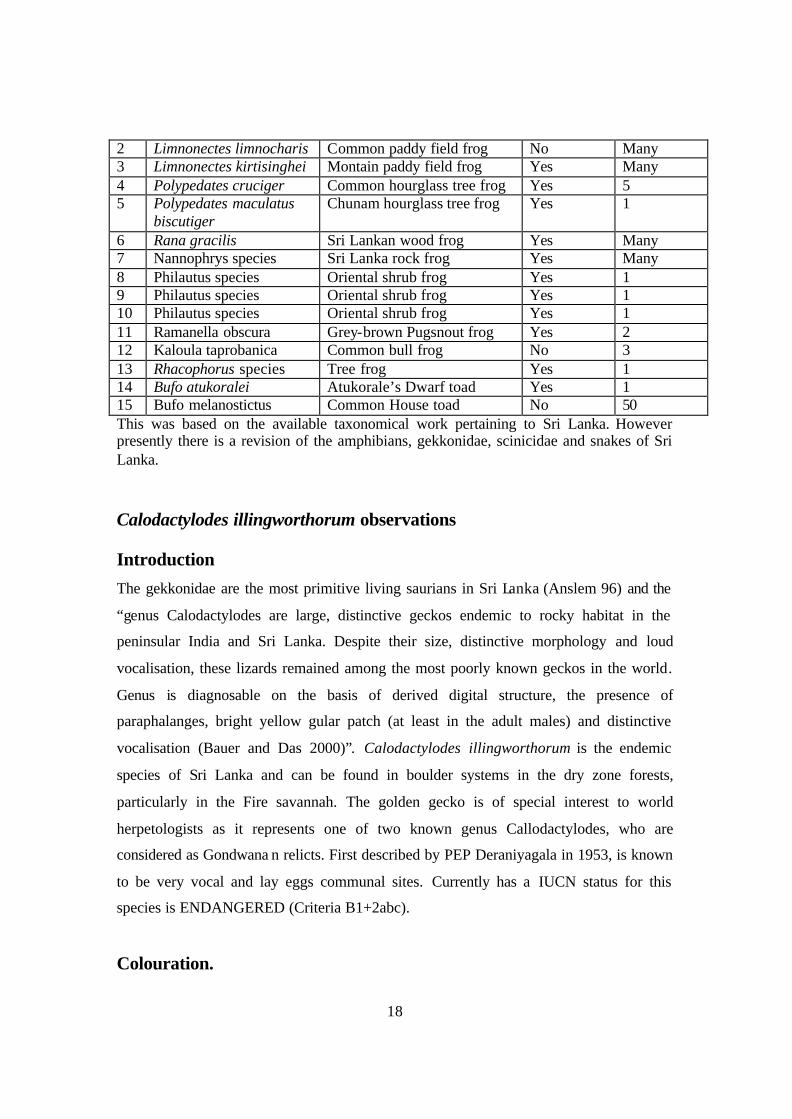
18
2 Limnonectes limnocharis Common paddy field frog No Many 3 Limnonectes kirtisinghei Montain paddy field frog Yes Many 4 Polypedates cruciger Common hourglass tree frog Yes 5 5 Polypedates maculatus
biscutiger Chunam hourglass tree frog Yes 1
6 Rana gracilis Sri Lankan wood frog Yes Many 7 Nannophrys species Sri Lanka rock frog Yes Many 8 Philautus species Oriental shrub frog Yes 1 9 Philautus species Oriental shrub frog Yes 1 10 Philautus species Oriental shrub frog Yes 1 11 Ramanella obscura Grey-brown Pugsnout frog Yes 2 12 Kaloula taprobanica Common bull frog No 3 13 Rhacophorus species Tree frog Yes 1 14 Bufo atukoralei Atukorale’s Dwarf toad Yes 1 15 Bufo melanostictus Common House toad No 50 This was based on the available taxonomical work pertaining to Sri Lanka. However presently there is a revision of the amphibians, gekkonidae, scinicidae and snakes of Sri Lanka. Calodactylodes illingworthorum observations Introduction
The gekkonidae are the most primitive living saurians in Sri Lanka (Anslem 96) and the
“genus Calodactylodes are large, distinctive geckos endemic to rocky habitat in the
peninsular India and Sri Lanka. Despite their size, distinctive morphology and loud
vocalisation, these lizards remained among the most poorly known geckos in the world.
Genus is diagnosable on the basis of derived digital structure, the presence of
paraphalanges, bright yellow gular patch (at least in the adult males) and distinctive
vocalisation (Bauer and Das 2000)”. Calodactylodes illingworthorum is the endemic
species of Sri Lanka and can be found in boulder systems in the dry zone forests,
particularly in the Fire savannah. The golden gecko is of special interest to world
herpetologists as it represents one of two known genus Callodactylodes, who are
considered as Gondwana n relicts. First described by PEP Deraniyagala in 1953, is known
to be very vocal and lay eggs communal sites. Currently has a IUCN status for this
species is ENDANGERED (Criteria B1+2abc).
Colouration.

19
Colour is variable and changing from site to site (camouflage) and across age group, sex
and mood.
Dorsal pattern has mottled ashy brown body and bands of ash and cream on limbs and tail
separated by alternating thicker bands of black and cream. The vertebral midline bears
series of dark patches from nape to base of tail, each separated by a cream dash. Throat
coloration of adult males is yellow, the brightest belonging to the dominant male.
Distribution
It was found to be widespread in appropriate rocky habitats and may not be as uncommon
as previously reported. Table 1 shows distribution and numbers of individuals sighted in
each of the study sites. Ideal habitat appeared to be boulder systems, especially those with
caves, surrounded by either fire savannah or monsoon forest. They often share this
habitat with bats, many species of invertebrate and sometimes Hemidactylus maculatus
hunae.. Almost all suitable sites have gecko sightings as well as current egg sites of
Calodactylodes illingwortherum indicating that the species is successful despite some
cases of human intervention.
Table 5 Calodactylodes illingworthorum distribution in the Monaragala district , Sri
Lanka.
Some interesting points to note about these results are that despite the human
threats in cluster 3, the Buddhist monk’s cave, the population of Golden geckoes is still
thriving here. 9 adults were sighted in total and 2 current egg sites were also observed,
although these were nowhere near as extensive as the abandoned ones.
The lack of golden geckos, gecko calls or eggs in the areas of Hangala, Tottileketiya, and
Walasgalge (Fig. 81) is also significant. It looks as though excessive burning in the
Hangala region could be the cause of diminished gecko populations, as it has been noted
that no Giant Spotted inhabit this area either (Fig. 78). In Tottileketiya and Walasgalge
however, thriving populations of Giants can be found, suggesting that environmental

20
factors are not to blame for lack of Calodactylodes illingworthorum. A population
dynamics theory could be used to explain this phenomenon although further studies must
be done to see if this is the case.
Maligathenna, a protected forest, showed very good golden population in all boulders
examined. Total 16 adults, 14 subs and 6 juveniles. Egg counts were also good; 6 unused
sites and 4 current sites, one in cave totaling 89 eggs at various stages of development
and some recently hatched. Some of the eggs were dry and empty inside and it has been
suggested that this could be caused by attack by ants, which were also noted at the site.
Typical data for ? adult, ? adult and juvenile
Weight (g) Snout-Vent
(mm)
Tail length
(mm)
Juvenile C2 0.7 36 41
Gravid ? C3 9.2 82 102
Adult ? C3 ? 85 98
Habitat and activity
Daytime behavior
It has been suggested from previous studies that golden geckos are largely
nocturnal as this is when they go into the trees to feed. We observed that during daylight
hours they were mostly quiet and mainly remained in caves and crevices in the boulder
system. This made them quite difficult to find without searching crevices with a torch. A
few calls were heard but these were infrequent and had large pauses. Our studies may
have altered their call behavior also as they would have been warning others of our
presence in their habitat and been unable to rest as normal.
‘Active’ periods
There are certain ‘active’ times at dawn and dusk when behavior changes and the
geckos become very vocal and begin to move. Calls were heard at 17:00 in Mahahela and
18:00 in Gangodagama Dambagahawela temple, A169 and also during the 24 hour study

21
in Cluster 1 unit 1. Geckos were heard calling to each other between 16:00 and 17:30
from within the cave. Calls were loud and frequent with only very short pauses.
Individuals start moving out of caves by 6pm, seem to climb tree with haste. Most of
them were out of cave system by 19:30. A solitary Hemidactylus maculatus hunae was
also observed leaving at the same time.
The geckos were then extremely hard to observe in the dense trees overnight but were
quiet until 4:30 the following morning when the loud bursts of calls could again be heard
and they made their way back down into the cave. All had returned by 6:30.
Social behavior
Territorial behavior was also observed in the course of the 24 hour study. Adults
were very vocal and aggressive, displaying defensive arch posture (also adopted in
response to torchlight) and biting each other. The winner of disputes was observed
retaining the arch position, while the loser ran off. Other geckos appeared to gather in a
circle around winner. A separate adult was observed primarily defending territory against
a juvenile for over an hour.
It was also observed that during the resting and moving in and out of cave, many of the
geckos remained in pairs of approximately the same age. Behavior was relaxed within
these close quarters and during daytime hours a soft chuckling vocal sound was heard
between members of a pair, very different to calls heard during fights and active time.
Previous research has suggested that a system of dominant males may exist in golden
gecko communities and that these males have yellow throats. We did not discover
dominant males in most of our sites, although there appeared to be 2 living in the cave in
cluster 3.
Other behavior
Calodactylodes illingworthorum are adapted for cave and boulder systems of a
certain rock type. They are extremely agile and fast and have been observed jumping
over 1 foot in order to span a vertical crevice. It was found that Golden gecko feet are

22
highly adapted to their habitat, only adhering to certa in surfaces. Three individuals were
observed when placed on different species of tree. It was found that when placed on
Parnaka or Kalupenni all three ran quickly up the trunk. However when placed on
Kiricone, they could not grip and fell to the ground. This is due to the courser texture of
the bark and indicates that the feet are highly specialised and the areas they inhabit must
have certain properties in order for them to traverse them effectively.
Unusual behavior
Our research indicates that Calodactylodes illingworthorum remain in the caves
and rock systems during the day and only come out at night. However, an adult was
spotted on a tree near to the main cave at cluster 3 during daylight hours around 15:00.
This was not the only time that Callodactylus ilingworthorum was seen in trees before
dusk as a juvenile was observed resting at base of a young tree at 11:00 for 2 hours,
leaving, then returning after 30minutes and remaining there for a further 3 hours. This
was during the 24 hour study in cluster 1 unit 1.
Another unusual phenomenon was observed in Rathugala, where a golden adult was
found in a crevice infested with ants and remained motionless while the ants crawled all
over its body.
Diet
It has been reported by Deraniyagala that Calodactylodes illingworthorum diet
include beetle larvae and glow worms (Bauer and Das 2000).
Dietary composition was determined by stomach flushing of all adult and sub adult
specimens (n=…) Most specimens contained very little stomach contents of prey items.
This may have something to do with time of day they were caught i.e. all caught at
daytime after 10am so all prey items eaten during previous night would be too digested to
identify. But all prey items retrieved were arthropod (large muscle blocks in the upper-leg
of unidentified insects were observed in stomach contents).
Reproduction

23
Location of egg sites
See Table 1 for details of egg clutches at each study site
Most of the egg sites found adhered to a strict set of properties in order to be
suitable. The most extensive current egg sites were found in caves and archways where
temperatures remain stable at around 28? C and darkness is constant. Smaller clutches of
around 6-15 eggs were frequently sighted underneath overhangs and ledges and in swifts’
nests, and were generally at least a metre from the ground. In cluster 2 sites were only
found in areas that were far from recently burnt savannah. Occasionally 2 solitary eggs
were found under small ledges away from the communal sites. This may be because the
female did not have time to reach the site and had to settle for the most appropriate
alternative. In cluster 1 only a small area is used for breeding, around the main cave and
archway leading to it. The main egg site in the cave was observed one various dates to
monitor its progress.
20th June 99 eggs of various colours
1st July 70 eggs, many juveniles observed in area
1st August 88 eggs after rains,18 more than 1st July.
Abandoned sites
Numerous abandoned sites were discovered at almost every site we visited. This
suggests that although these sites were once used for egg laying, changes have meant that
they are no longer suitable. An example of this is an abandoned egg site observed in an
overhang in cluster 2 which had water damage and fungus. Savannah burning and tree
felling have also exposed some sites to sunlight making them unsuitable. However with
most sites it is unclear why they are currently unused. They may just have gone out of
favour due to more appropriate sites being available, or may be not needed due to
diminishing population caused by various threats, for example human intervention. This
is most likely the case in C3, where although there are good numbers of individuals, the
huge abandoned sites suggest that there was once a far more extensive population.

24
However in Hamanawa, a recently abandoned human construction has provided an
artificial cave which has actually benefited the gecko population here. 3 clutches and 7
individuals were observed in the building, which had also become inhabited by bats.
Unusual egg sites
Several egg sites were discovered in areas which did not appear to fit the usual
trends that had been followed by most of the clutches. In cluster 2 a small abandoned site
was found inside onion weathering on the side of a rock. This is strange as it provides no
overhead cover. In Rathugala, a medium sized egg cluster with 4 current eggs was also
found in crevice which was shared with ant nest. Ants were crawling all over and in the
eggs.
Empty/diseased eggs
The phenomenon of empty eggs was first observed at Bulapitya Hela where two
apparently large and healthy sites in a low cave were discovered to be less than 50%
viable (see table 1) . New eggs had also been laid over complete unhatched eggs, which
first highlighted the situation as unusual. Some eggs were completely empty and easy to
break, whereas others still had dry embryos inside. Dried foetal remains were removed
form several eggs for DNA. It was noticed that affected eggs had a tiny hole and
appeared flakier in texture than healthy eggs.
This new finding was also recorded at several other sites, including Yakunahela,
Hamapola rock, Maligathenna and Gal kotte. The cause of these empty eggs is not known
for certain, but there is a strong possibility that it could be caused by invasion by
Thelkumbi ants, which were observed crawling over the eggs in Gal kotte, Maligathenna
and Yakunahela. The site at Gal kotte was especially unusual as its structure had become
globular (not flat to rock) due to eggs being laid over intact eggs several times. It was
estimated that the site contained at least 50% empty eggs and many eggs were noted to
have orange colouration with white fungal spots. A dry and dead foetus was removed
from an affected egg for analysis. Despite its appearance as an obviously unhealthy site, a
female was later observed attempting to lay eggs here but was disturbed by our presence.

25
Another type of non-viable eggs was discovered at Hamanawa monk house where a
clutch of 13 new eggs was discovered. One egg was broken by Anslem and was not dry
but full of spoilt yellow gunk. This was also removed for testing.
Egg laying
On two occasions, one in Gal Kotte and one in Palamugala, females were also
seen arching in a similar stance to the threat position over current egg sites. However in
both instances the female was disturbed by our presence and left without laying any eggs.
Hemidactylus maculatus hunae observations
Introduction
The Giant Spotted gecko is one of the largest species of Hemidactylus in the world
measuring at 250mm+. First described by PEP Derianiyagala in 1937 has a IUCN status
of ENDANGERED (Criteria B1+2bc).
Distribution
As shown in table 1, five Hemidactylus maculatus hunae were found in Palamugala unit
one Figure 17, coinciding with Calodactylodes illingwortherum, but distributed over a
wider area than the Calodactylodes illingwortherum which were more concentrated. Two
Hemidactylus maculatus hunae were also found in unit two, where no Calodactylodes
illingwortherum were found. One or two Hemidactylus maculatus hunae were seen in
other boulder systems; Rathugala, Gal kotte, Avalahela and Maligathenna. But the
greatest populations were found in regions where there were few or no Calodactylodes
illingwortherum.
In Totillaketiya where there were no Calodactylodes illingwortherum, eleven
Hemidactylus maculatus hunae were found, and similarly in Walasgalge where there
were no sightings of Calodactylodes illingwortherum, ten Hemidactylus maculatus hunae
were seen. Regions such as Cluster 1 unit 2, and Maligathenne, shaded cave systems
surrounded by fire savannah, which would be ideally suited as Hemidactylus maculatus

26
hunae habitats, have thriving Calodactylodes illingwortherum populations, but have
remarkably few Hemidactylus maculatus hunae.
This would suggest that the two species compete for the same niche and cannot readily
co-exist, so in many boulder systems one species dominates.
Habitat and activity
Hemidactylus maculatus hunae were found mainly in the same habitat as
Calodactylodes illingwortherum, within cave and rock systems. However we made an
unusual discovery in Gal Kotte; an adult Hemidactylus maculatus hunae was found under
the bark removed from a dead Atabo tree. It was quite dark in colour, very similar to the
tree colour, and was possibly another species of arborial Hemidactylus. Unfortunately it
escaped us so we were not able to take it back for identification.
From the distribution observed, it would appear that Totillaketiya is the ideal habitat for
Hemidactylus maculatus hunae. Over our relatively brief investigation of the site (seven
hours) we sighted a total of eleven individuals.
Totillaketiya had extensive cave systems with very few other geckos to compete with
the Hemidactylus maculatus hunae , and no bats which could predate on them.
One early gravid female was caught, and she displayed interesting behaviour, moving
towards the torch light Figure 15. A usual response would be to freeze or move in the
opposite direction to try and escape. A very large Hemidactylus maculatus hunae was
sighted approximately 280mm, which would put this specimen as the contender for the
‘biggest Hemidactylus maculatus hunae’ record, but it was inaccessible and could not be
caught.
Reproduction
Hemidactylus maculatus hunae lay their eggs in solitary pairs in leaf litter or rock
crevices. As they are not gathered in communal egg laying sites, or attached to rocks,
they are much more difficult to find than the egg clusters of Calodactylodes

27
illingwortherum. However we did come across a few Calodactylodes illingwortherum
eggs in our investigations.
In Palamugala unit two a Hemidactylus maculatus hunae egg was discovered semi-
submerged in leaf litter in the rotting stump of a tree. It was on the verge of hatching, and
when touched, the baby Hemidactylus maculatus hunae leaped out. We managed to catch
it, and believed it to be a new species for severa l days until its colouration matured into
that of a Hemidactylus maculatus hunae. (Figure 18)
In Rathugala, two solitary eggs, presumed to be Hemidactylus maculatus hunae, were
found in a small rock very close to an old Calodactylodes illingwortherum egg site. This
is interesting as we think that the two are rival species.
In Totillaketiya we discovered a suspected Hemidactylus maculatus hunae egg in a
crevice where two giant spotted geckos were also found (in boulder on periphery of
system) The temperature in the 16cm wide crevice was 26.6ºC, the outside temperature
was 30.6ºC, and the relative humidity was 90%.
We found two more Hemidactylus eggs of undetermined species, submerged under leaf
litter during the canopy study near the lacerid rock. One of these was kept.
Other species of Hemidactylus
On the 11th July at 4.30, Hemidactylus depressus Figure 21 individuals were found under
the peeling bark of a dead tree in Panaka tree. The tree was in direct sunlight growing on
a rock. A darker coloured Hemidactylus depressus was then observed on a shady rock
under an overhang near a porcupine den. A Hemidactylus depressus was also found in
boulder systems near the Lacertid rock.
In Walasgalge an unidentified, possibly new Hemidactylus species was caught from an
exposed rock. It’s total length was 90mm Snout-tail, and an egg was also found on the

28
same rock Figure 19. On the same day another Hemidactylus individual, possibly of a
new species, was found in a fallen decomposing Dambe tree.
In Lahugala two species of Hemidactylus, H. frenatus (common housegecko) Figure
20,22 and Hemidactylus leschenaultia (bark or sycamore gecko), were found in trees
around the dried marshland. We then found three H. depressus geckoes in the trees
around a cave system nearby.
Cnemespis observations Distribution
Cnemespis species were found to be very widespread throughout our study area. We
found them living in almost all sites, but most notably Maligathenna, where a huge
population exists in the rock systems alongside an equally successful Calodactylodes
illingworthorum community. Cnemespis species are characteristically small, which
makes them very difficult to find and this may account for the small numbers sighted
earlier on in the research. It is also very difficult to determine exact species without
capturing and there is a lot of classification work still to be done involving Cnemespis.
For this reason, many of the geckos we caught are currently unknown.
Table 1 shows the areas where Cnemespis species were sighted.
Habitat and activity
Boulder system habitat
Initially we were searching the same rock systems as the golden habitat in order to look
for Cnemespis. 3 Cnemespis tropidogaster were found among the rocks high up the hill at
Rathugala. 2 of these were caught and found to be gravid females. An immense
population of C. tropidogaster was also discovered living in rocks at Maligathenna where
a total of 13 individuals were recorded. These Cnemespis were sharing are able to coexist
successfully. Another example of this was at the Hamanewa monk cave where a

29
Cnemepsis pair was sighted above the doorway to the manmade cave which was largely
populated by Calodactylodes illingworthorum.
However, we later discovered that the Cnemespis habitat range is far more diverse,
including not only rocks and caves but also tall, wide-girth trees, especially those with
peeling bark, such as Hama, or with deep grooves in the bark, such as Galiyabala. In
Hamapola, two Cnemespis of unknown species were discovered living underneath sheets
of peeling bark on a huge dying Hama tree. Another was sighted high up in a Galiyabala
tree in Maligathenna.
Canopy studies
This discovery prompted us to begin actively searching the forest canopy for Cnemepsis
species. Canopy studies were carried out mainly around the lacertid rock area, 13km post
Ampara road. On the 31st July a total of five Cnemespis were sighted in trees assessed in
this area. Trees were of wide girth (GBH>162cm) and individuals were usually between
4 and 6m above ground level. A Golden band was observed on the dorsal side of four of
these individuals, although catching attempts were unsuccessful. This type of Cnemepsis
was found two per tree which suggests they may live in pairs. A different Cnemespis
species was also caught from at tree with paler bark. This gecko was unusual in that a
greenish skin tone could be observed in certain light.
Several other Cnemepsis were spotted in similar trees, such as large Mille, although were
not caught. The pair trend was however supported by these sightings.
Additional habitats
Also, whilst traveling in Sri Lanka outside our study area we discovered Cnemespis
species in other types of habitat. In the Monatane forests surrounding the Pissa waterfall
two individuals were caught from rock near to the river. These are expected to be
different to those caught in fire savannah sites as the vegetation here is mostly Diocarps
and the climate much cooler and damper. Specimens have been sent to America for

30
classification. Two other individuals were also caught from rocks at the roadside amongst
pine forest between Knuckles and Bibile.
Diet
As some species of Cnemespis live in the same habitat as Calodactylodes
illingworthorum, i.e. boulder systems surrounded by fire savannah, it would be
reasonable to assume that they predate the same types of invertebrates. However the diet
is difficult to research as we were unable to do stomach flushes on Cnemespis caught due
to their size. Cnemespis found in the Hama tree were however surrounded by termites
(white ants) which could act as a potential food source for this type.
Reproduction
Cnemespis eggs are very small in size (approx 4mm in diameter) and have been
discovered in several locations throughout our study. It was noted that when laid under
rock ledges, the sites appeared to be used over and over and were communal as in
Calodactylodes illingworthorum. However when eggs were found in trees they were
often solitary or in pairs, although this does not mean that sites are not communal,
because old egg fragments may not remain in bark as they do on rock. Intact eggs were
discovered in bark of Athumbe tree in Rathugala (temp 22.1?C), a Patirande tree in
Hamanewa (22.1?C) and in the Hama tree at Hamapola (26.8?C). A sample egg was also
removed from a tree in Gal Kotte.
Cnemespis eggs were also discovered on rock near the ground of a cave in Rathugala,
adhered to the surface. One of the pair was removed for tests. No other intact Cnemespis
eggs were discovered on rock surfaces, although several small currently unused sites
were found under ledges in Palamugala and Maliga thenna. Gravid females were also
caught in Rathugala.

31
New species
It is not possible to classify many of the specimens sighted and caught as there is
currently a lot of re-working involving Cnemespis species. However at least 3 of the
animals found are thought to be previously unseen:
?? Grey tone species found in rocks at Gal Kotte
?? Arboreal species found in Athumbe tree at Yakunahela
?? Green tinge species found during canopy work near lacertid rock
Birds Bird species observed in Nilgala area
Common Name Scientific Name
1. Brown-headed Barbet Thereiceryx zeylanicus zeylanicus 2. Black-headed Oriole Oriolus Xanthornus ceylonensis 3. Spoonbill Platalea leucorodia 4. Spotted Dove Streptopelia chinensis ceylonensis 5. Black Indian Robin Saxicoloides fulicata leucoptera 6. Ceylon Black Drongo Dicrurus macrocercus minor 7. Ceylon Grey Hornbill Tockus gingalensis 8. Common Babbler Turdoides striatus striatus 9. Red-vented Bulbul Molpastes cafer cafer 10. Ceylon Bronze-wing Pigeon Chalcophaps indica robinsoni 11. White-breasted Kingfisher Halcyon smyrnensis fusca 12. Ceylon Common Kingfisher Alcedo atthis taprobana 13. Blue-eared Kingfisher,A2b, A3d1 Alcedo meninting 14. Black Crow Corvus macrorhynchos culminates 15. Rose-ringed Parakeet Psittacula krameri manillensis 16. Emerald-collared Parakeet Psittacula calthorpae 17. Large Ceylon Parakeet Psittacula eupatria eupatria 18. Pond Heron Ardeola grayii 19. Eastern Large Egret Egretta alba modesta 20. Median Egret Egretta intermedia intermedia 21. Little Egret Egretta garzetta garzetta 22. Spotted Munia Uroloncha punctulata lineoventer 23. Orange Minivet Pericrocotus flammeus 24. Green Imperial Pigeon Ducula aenea pusilla 25. Ceylon Crested Goshawk Accipter trivirgatus layardi 26. Black Eagle Ictinaetus malayensis perniger 27. Painted Stork Ibis leucocephalus 28. Southern Magpie Robin Copsychus saularis ceylonensis 29. Ceylon Swallow Hirundo daurica hyperythra

32
30. Yellow-fronted Barbet Cyanops flavifrons 31. Indian Roller Coracias benghalensis indica 32. Koel Eudynamys scolopaceus scolopaceus 33. Indian Peafowl Pavo cristatus 34. Ceylon Paradise Flycatcher Tchitrea paradise paradisi 35. Red-wattled Lapwing Lobivanellus indicus indicus 36. Common Grackle Eulobes religiosa indica 37. Common Coucal Centropus sinensis parroti 38. Ceylon Jungle Bush Quail Perdicula asiatica ceylonensis 39. Little Cormorant Phalacrocorax niger 40. Indian Shag Phalacrocorax fuscicollis 41. Indian White-necked Stork Dissoura episcopus episcopus 42. Sri Lanka Junglefowl, A2a Gallus lafayeti
Field Ornithology Group of Sri Lanka. 2003. Preliminary IBA Site Directory. Notes on List of Important Birds A2a Endemic Species A2b Nationally Threatened Species A3d1 Uva Avifaunal Zone Mammals The following mammals were observed during the field work.
Mammal species observed in Nilgala area
Common Name Scientific Name Sri Lanka Elephant Elephas maximus maximus Giant Squirrel Ratufa macroura Mongoose x2 Herpestes edwardsi lanka,
Herpestes smithi zeylanicus Spotted Deer Axis (axis) axis ceylonensis Sambha Deer Cervus unicolor unicolor Wild Boar Sus scrofa cristatus Bats Microchiroptera sps Jungle Cat Felis chaus kelaarti Mouse Deer Tragulus meminna Palm Squirrel Funambulus palmarum Bandicoot Bandicota indica Porcupine Hystrix indica Pangolin Manis crassicaidata Macaques Macaca sinica sinica Ceylon Grey Langur Semnopithecus priam thersites Sloth Bear Melursus ursinus inornatus

33
Sri Lanka Leopard Panthera pardus lankae Flying Squirrel Petaurista petaurista lanka Common Flying fox Pteropus giganteus giganteus Loris Loris tardigradus Black napped hare Lepus nigricollis singhala Palm Cat Paradoxurus hermaphroditus Otter Lutra lutra (ceylonica) nair Shrews Crocidurinae sps
Folklore
In Sri Lanka people don’t harm geckos due to many cultural beliefs. One such belief in
the area is that gecko bites are deadly and there is no cure for it. Another such belief
across Sri Lanka is that if a gecko cries when the occupant leaves on a journey, it
signifies a warning and should not leave the house immediately (Anslem 96)
Traditional knowledge of the Vedda
Summary of our findings from the KAP study (knowledge and attitudes of people) on the
vedda aborigines of Nilgala and Rathugala :
As Seligman predicted a century ago when he first made his study on the declining vedda
population of Sri Lanka, the vedda aborigines have now almost totally disappeared due to
the effects of modernization. The Rathugala region where we were studying was once at
the center of their lands and is one of the last regions in Sri Lanka to be inhabited by this
ancient people. However even here there are no longer any true veddas, and only
descendants of the true veddas and remnants of their culture remain.
Vedda knowledge is passed on solely by word of mouth, and by speaking with the
vedda descendents who lived in the Nilgala and Rathugala districts we were able to learn
about their culture. Although this is only a fragment of what Seligman described. The
traditional vedda way of life is sustained by the forest, and hence a knowledge of the
natural resources in the forest around them is central to their survival. This knowledge is
now being lost, or used by young vedda descendents to exploit the forest ie for illegal
hunting, collecting honey and medicinal plants.

34
Respect of nature is ingrained in vedda culture; the chief we met said he would not kill a
deadly viper even after he was bitten by it. They kill only for necessity, and when hunting
observe their prey to see if it is female with the potential to breed, in which case they will
not kill it. They also do not kill sacred ‘key stone’ species ie leopard, bear, elephant and
snakes.
However the vedda descendents we came across have come a long way from the veddas
described by Seligman, who lived secluded in the forests in caves and hunted for
sustenance. A whole vedda tribe was moved by the government in the Mahaweli project
and settled in Galgamuwa (Rathugala), far from Danigala, their ancestral land. The
remaining families we met in Nilgala and Rathugala live along the roadside in houses
given to them by the government, and cultivate plots of land by “chena cultivation” and
receive government funded medical care. Their primitive way of life has been lost and
some have been reduced to collecting money by the road side.
The families we met were impoverished and very large, with evidently no family
planning in use, and the numerous small children did not go to school. It was apparent
that women have little place in society amongst these people, as they were always
encumbered with small children and allowed the men to speak for them.
The characteristic vedda traits are greatly reduced in the descendents we saw due to
intermarriage and integration with Sinhalese local people. However they appeared
noticeably smaller and shorter in stature than pure Sinhalese people, a characteristic
Seligman noted when he studied their ancestors, suggesting that their genetic identity is
still primarily vedda. They no longer retain any of their religious traditions or belief in the
Nae Yaku, and have adopted the Buddhist or Hindu beliefs, worshiping Lord Buddah or
Lord Ganesh. They have also lost their own langua ge which has been replaced by
Sinhalese. The old chief was the only one who could speak their ancestral language.
However they still remember traditional vedda dances which they perform for
entertainment in special events.

35
During our exploration of the Nilgala forest we came across many caves which were
once inhabited by veddas long ago. They were naturally formed caves under boulders
with the characteristic drip ledge carved into the rock around the top rim using sharp
implements to divert the flow of rain water away from the mouth of the cave. However,
besides providing temporary shelter to locals whilst hunting, these have not been lived in
for many generations.
See Annexure 4 for further details- Notes on Veddas from Seligman and Punchihewas
books.
Threats and Management
Threats to the Gecko populations with a special reference to
Calodactylus illingworthorum.
Humans have shared the gecko habitat for 1000s of years (Fig. 84). Drip ledges and
treasure stones constructed by veddas were found at several of our sites (Figs, 79 and
80). However. it became apparent during our studies that there are many recent human
threats faced by golden gecko populations due to expansion of farmland and changes of
traditional ways. It will be interesting to see if these threats have any effect on the
population numbers we have counted in years to come.
Threats caused by human behaviour include:
?? Clearing of trees and lyanas cluster1 by forestry department surveyors, exposing the
rock face to sunlight which may result in changes in temperature, humidity and light
intensity of the rock.
?? Encroachment by paddy fields and other changes in land use i.e. Segregation of rock
outcrops due to local paddy field farmers creating thrashing floor in cluster1.

36
?? Clearing for thrashing floor leads to tertiary vegetation, as thorny weed species
(Eraminiya Zizyphus napeca and Katuhinguru Lantana camera) engulf natural
vegetation of Kirikone, Mille, Thelabo, Dhikande, Kalupanni (Fig. 83)
Note – these thorny species do not allow geckoes to move around easily
?? Grazing by cattle cluster 2 (Fig.85)
?? Honey collecting by destructive methods
?? Burning savannah for Arulu and bedi leave collection. Note: time of year when the
burning begins as its not too dry. Once started the fires go one to for about 4 months
until it hits a fire break. (Fig. 77)
?? chopping of woody saplings for fence making cluster2
?? illegal extraction of timber (Fig. 86)
?? human disturbance, i.e. in Cluster 3
*Building construction and building materials
*Dynamite and debris
*Graffiti
*Children playing
Other threats to the populations are caused by parasites. Red ectoparasitic mites were
discovered on several individuals including a juvenile in cluster2 and adult male in
cluster 3. These may hinder adhesion and could pass infections or result in high blood
loss if an individual is badly affected. Thelkumbi ants predating unhatched embryos is
also a serious problem as seemingly healthy looking sites of many eggs could result in no
new offspring if infestation is heavy. Work on identification of reptile parasites is
ongoing in the Vetinary Faculty, University of Peradeniya, Sri Lanka. This work will
give a clearer picture of the effects of these parasites on the population.
Threats to other reptiles and amphibians
There are three distinct predators of the house gecko: domestic cats, snakes (Boiga sps,
Oligodon sps, Lycodon aulicus, Elephe Helena, Oligodon sps feed on eggs as well) and
poultry and birds especially the common Coucal (Centropus sinensis parroti) a big
predator of all herpetofauna (Anslem 96).

37
Many road kills were observed en route, for example Dendrelaphis tristis- bronze back and further examples see Table 04.
Other Threats to the area.
?? Overexploitation of medicinal plants in the fire savannah which include most
species listed in Table 03 and other plants like wild tabbaco, bidi and
binkohomba.
?? Threats to archeological monuments etc from destruction by treasure hunters.
?? Hunting of protected mammals i.e. finding Pangolin scales ( Manis
crassicaidata ) on Lacertid rock Figure 53.
?? Illegal gem mining especially on the banks of the Gal Oya river and its tributaries
seen in Thotilleketiya.
Management recommendations

38
References
?? Henry, G.M. 1971. A Guide to the Birds of Ceylon. Oxford University Press. Pg. 457.
?? Field Ornithology Group of Sri Lanka. 2003. Preliminary IBA Site Directory.
?? Bandaranayake, W. M., Sultanbawa, M. U. S., Weerasekara, S. C.,
Balasubramaniam, S. 1974. A Glossary of Sinhala and Tamil Names of the plants of Sri Lanka. Forester, 6 (3 & 4): pp.
?? Allmon WD (1991) A plot study of forest floor frogs, Central Amazon, Brazil.
Journal of Tropical Ecology, 7: 503-522.
?? Ashton P S and CVS Gunatilleke (1987) New light on the plant geography of Ceylon 1 -
?? Historical Plant Geography. Journal of Biogeography 14: 249-285
?? Ashton M, S Gunatilleke, N de Zoysa, MD Dassanayake, N Gunatilleke & Siril Wijesinghe (1997) A field guide to the common trees and shrubs of Sri Lanka. WHT Publishers (Pvt.) Ltd. 432 pp.
?? Balasubramaniam S, S Ratnayake and R White (1993) The montane forests of the
Horton Plains Nature Reserve. IN: Ecology and Landscape Management in Sri Lanka. W Erdelen, C Preu, N Ishwaran, C M Madduma Bandara (Eds.). Margraf Scientific Books, PP 95 -108.
?? Bambaradeniya C N B & K B Ranawana (1998) Some amphibians observed in
three montane zone forests of Sri Lanka and their habitats In: Biology & Conservation of the amphibians, reptiles and their habitats in South Asia. pp. 108-113. Anslem de Silva et al (Eds.). ARROS.
?? Bauer A. and Das I., A review of the gekkonid genus Calodactylodes (Reptilia:
Squamata) from India and Sri Lanka. Journal of South Asian Natural History. 2000. Vol.5, No. 1, pp.25-35.
?? Bauer A M & A P Russell (1994) Regional integumentary loss as an antipredator
strategy in lizards. Abstracts, Second World Congress of Herpetology, University of Adelaid, Australia pp.16-17.
?? Bickford D P (1994) Diversity and abundance of a montane leaf-litter
herpetofauna in Costa Rica: implications of elevational patterns. Abstracts, Second World Congress of Herpetology , University of Adelaide, Australia. p.11

39
?? Central Environmental Authority (1997) Wetland site report and conservation management plan, Horton Plains National Park. CEA/Euroconsult. 70 pp.
?? Chandraratne R M M (1990) Archaeological explorations at Horton Plains: a
review. Paper presented at Conservation and management of Ritigala Kanda and Horton Plains. 9-10 August 1990. Natural Resources, Energy and Science Authority of Sri Lanka. 6 pp.
?? Costa H H (1974) Limnology and fishery biology of the streams at Horton
Plains, Sri Lanka (Ceylon). Bull. Fisheries Research Station, Sri Lanka (Ceylon). 25(1&2): 15-26.
?? Crusz H & V Sanmugasunderam (1974a) Parasites of the relict fauna of Ceylon.
III. Nematodes from a Rhacophoid frog and reptiles of the hill country. Annals de Parasitologie (Paris), 48(6): 767-795.
?? Crusz H & Sanmugasunderam (1974b) Parasites of the relict fauna of Ceylon.
IV.. Zeylanurotrema lyriocephali gen. et. sp. nov and other trematodes from mountain lizards and a rodent. Annals de Parasitologie (Paris), 48(6): 797-810.
?? Crusz H & CC Ching (1975) Parasites of the relict fauna of Ceylon. VI.. More
new helminths from amphibians and reptiles, with a new host-record and redescription of Acanthocephalus serendibensis Crusz & Mills, 1970. Annals de Parasitologie (Paris), 50(5): 531- 558.
?? Das I (1995) Amphibians and reptiles recorded from Batu Apoi, a lowland
dipterocarp forest in Brunei Darussalam Raffles Bulletin of Zoology 43: 157-180
?? Das I (1996) Spatio-temporal resource utilization by a Bornean rainforest herpetofauna: preliminary results In: Tropical rainforest research: current issues. pp. 315-323. D Edward’s, W Booth & S Choy (Eds.) Kluwer Academic Pub. Dordrecht.
?? Das I & de Silva Anslem (1998) Species diversity and species richness of the leaf
litter herpetofauna in Sri Lanka: preliminary results. In: Biology and conservation of the amphibians, reptiles and their habitats in South Asia. pp. 285-293. Anslem de Silva et al (Eds.). ARROS.
?? Davis A M M (1953) Python at Horton Plains. Loris 6(3): 122
?? Deraniyagala P E P (1953) A colour atlas of some vertebrates from Ceylon. Vol.2
Tetrapod Reptiles. Ceylon National Museum Publications. 101 pp.
?? Deraniyagala S (1971) Archaeological explorations in Ceylon. Part 1: Horton Plains. Spolia Zeylanica. 33(1): 13-22

40
?? de Silva, Anslem (1990) Colour guide to the snakes of Sri Lanka R & A Publications ltd. Portishead, UK, 130 pp.
?? de Silva, Anslem (1996) The herpetofauna of Sri Lanka: a brief review. Author,
Graphic Land, Kandy. 99 pp + 15 plates.
?? de Silva, Anslem (1996) The amphibia of Sri Lanka: a checklist and an annotated bibliography. Dept. of Wildlife Conservation/GEF/UNDP/FAO. 48 pp
?? de Silva, Anslem (1997a)(publ. 1998) Lankascincus taprobanensis (Kelaart,
1854) a skink with a prehensile tail Lyriocephalus 3(2): 26-27
?? de Silva, Anslem (1999a) New amphibian parasites from Sri Lanka. Froglog.. No.31. p 2.
?? de Silva, Anslem (2001) Preliminary observations of the amphibian fauna at
Horton Plains National Park. IN: The Amphibia of Sri Lanka: current research. Anslem de Silva et al (Eds.) pp. 64 – 88.
?? de Silva, Anslem (2001) Species-specific factors preventing detection of
Microhyla zeylanica at Horton Plains National Park. . IN: The Amphibia of Sri Lanka: current research. Anslem de Silva et al (Eds.). pp. 141 - 144.
?? de Silva, Anslem (2001) Some aquatic insects: predators of anuran tadpoles at
Horton Plains National Park. . IN: The Amphibia of Sri Lanka: current research. Anslem de Silva et al (Eds.) pp. 145 - 146.
?? de Silva, Anslem, RPVJ Rajapaksa and S Karunaratne (2001) Preliminary
observations of the Proteocephalan cestodes from Polypedates eques inhabiting Horton Plains National Park. IN: The Amphibia of Sri Lanka: current research. Anslem de Silva et al (Eds.). pp. 153 – 165.
?? RPVJ Rajapaksa and de Silva, Anslem (2001) Preliminary observations of the
paasites of amphibian fauna at Horton Plains National Park. IN: The Amphibia of Sri Lanka: current research. Anslem de Silva et al (Eds.). pp. 166 – 167.
?? Diamond, JM (1975) The island dilemma: lessons of modern biographic studies
for the design of natural reserves. Biological Conservation. 7: 129-146.
?? Dutta S K & K Manamendra-Arachchi (1996) The amphibian fauna of Sri Lanka Wildlife Heritage Trust of Sri Lanka. 230 pp.
?? Erdelen W (1984) The genus Calotes (Sauria, Agamidae) in Sri Lanka:
distribution patterns. Journal of Biogeography. 11: 515-525

41
?? Erdelen W (1988) Population dynamics and dispersal in three species of agamid lizards in Sri Lanka: Calotes calotes, C versicolor and C. nigrilabris. Journal of Herpetology 22(1): 42-52.
?? Erdelen W (1998) The Genera Otocryptis and Sitana (Sauria, Agamidae);
Geographic distribution, microhabitat use, and morphometric relations. IN: Biology and Conservation of amphibians, reptiles and their habitats in South Asia, Eds. Anslem de Silva et al. pp. 232-246.
?? Fernando C H (1990) Freshwater fauna and fisheries of Sri Lanka. Natural
Resources Energy & Science Authority of Sri Lanka. p.p. 444.
?? Fernando T K & Chandrapala L (1991) Global warming and rainfall variability - Sri Lanka situation. Proceedings 47 Ann. Sess. Sri Lanka Asso. Adv. of Science. Part 1: 138
?? Gans C and Fetcho J R (1982) The Sri Lankan Genus Aspidura (Serpentes.
Reptilia, Colubridae) Annals of Carnage Museum 51(14): 271-316.
?? Gosner K L (1960) A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16: 183-190.
?? Greer A E (1991) Lankascincus, a new genus of scincid lizard from Sri Lanka,
with description of three new species. Journal of Herpetology. 25(1): 59-64
?? Gunawardena ERN, P Rosier, H Davies (1997) Climate of Horton Plains. A publication of University of Peradeniya-Oxford Institute (UK), Forest Research Link Project. 4 pp.
?? Gunatilleke C V S (1996) A nature guide to the World’s end trail, Horton Plains.
Science Education Unit, University of Peradeniya, 48 pp.
?? Haly A (1890) Report of Director of the Colombo museum for 1888. Ceylon Administration Reports 1888 . 4: 152D.
?? Heinen J L (1992) Comparison of the leaf-litter herpetofauna in abandoned cacao
plantations and primary rain forests in Costa Rica: some implications for faunal restoration. Biotropica 24(3): 431-439
?? Inger R F (1980a) Densities of floor -dwelling frogs and lizards in lowland forests
of Southeast Asia and Central America. American Naturalist 115(6): 761-770.
?? Inger R F (1980b) Reletive abundance of frogs and lizards in forests of Southeast Asia. Biotropica 12(1): 14-22

42
?? Jaeger RG & RF Inger (1994) Quadrat sampling. In: Measuring and monitoring biological diversity: standard methods for amphibians. pp. 97-102. W A Heyer et al (Eds.) Smithsonian Institute, USA.
?? Karunaratne P B (1986) Random notes on the amphibians of the Kanneliya
jungles. Loris 17(4): 140-142.
?? Kirtisinghe P (1957) The amphibia of Ceylon. Author,William Clowes & Sons, London, 112 pp.
?? Lloyd M, R F Inger & F W King (1968) On the diversity of reptiles and
amphibian species in a Bornean rain forest. American Nat. 102: 497-515.
?? Manamendra-Arachchi K (1997) Gecko. Sri Lanka Nature 1(1): 45-55.
?? Naggs F (1996) A coloured guide to the land and freshwater mollusca of Sri Lanka. The Natural History Museum. 36 pp.
?? Olson D H, W P Leonard, R B Bury (1997) Sampling amphibians in lentic
habitats. Society for Northwestern Vertebrate Biology. Washington, 134 pp.
?? Pethiyagoda R & K Manamendra-Arachchi (1998) Evaluating Sri Lanka’s amphibian diversity. Occasional Papers No. of the Wildlife Heritage Trust. pp 1-12.
?? Rajapaksa R P V J & Anslem de Silva (1999a) Some parasites observed in frogs
inhabiting Horton Plains National park. Lyriocephalus, 4(1): accepted
?? Rajapaksa R P V J & Anslem de Silva (1999b) Preliminary observations of a subcutaneous cestode from a Rhacophorid frog inhabiting Horton Plains Lyriocephalus, 4(1): accepted
?? Silva E I L, PALC Manuweera, NP Athukorale, I Tumpela & KTDPR Jayapala
(1994) Buffer intensities: risk of acidification of aquatic ecosystems in Sri Lanka. Annual Review 1994. Institute of Fundamental Studies, Kandy. p. 77.
?? de Silva, Anslem (1997 “1998”) Crab feeding on a tadpole. Lyriocephalus 3(2):
28
?? de Silva, Anslem (1999a) The habitat preferences and the status of amphibians and reptiles at Horton Plains National Park. National Science Foundation of Sri Lanka, Zoological Survey of Sri Lanka, 67 pp (unpublished report).
?? de Silva, Anslem (1999b) Some aquatic hemiptran insects: predators of anuran
larvae at the Horton Plains National Park. Lyriocephalus 4 (1):

43
?? Heatwole H (1961) Habitat selection and activities of the wood frog, Rana sylvatica. American Midl. Nat. 66: 301-313.
?? Parker, H W & W C Osman Hill (1948, publ 1949) Frogs of the Genus Microhyla
from Ceylon. Annals & Magazine of Natural History. Ser. 12 vol. I: 759-764.
?? Punzo F (1976) The effects of early experience on habitat selection in tadpoles of the Malayan painted frog, Kaloula pulchra (Anura: Microhylidae). Journal of Bombay Nat. Hist. Soc. 73: 270-272.
?? Wiens J A (1970) Effects of early experience on substrate pattern selection in
Rana aurora tadpoles. Copeia (1970): 543-548.
?? Arnold EN (1988) Caudal autotomy as a defens IN: Biology of the Reptilia. Vol.16 Ecology B Defence and Life History. (Eds.) C Gans and RB Huey. Alan R Liss., Inc. New York. pp. 235-373.
?? Bauer, A M & A P Russell (1994) Regional integumentary loss as an antipredator
strategy in lizards. Abstracts. Second World Congress of Herpetology, University of Adelaide, Australia. pp. 16-17.
?? de Silva, Anslem (1997 “1998”) Crab feeding on a tadpole. Lyriocephalus 3(2):
28
?? de Silva, Anslem (1999) The habitat preferences and the status of amphibians and reptiles at Horton Plains National Park. National Science Foundation of Sri Lanka, Zoological Survey of Sri Lanka, 67 pp (unpublished).
?? Fernando C H (1990) Freshwater fauna and fisheries of Sri Lanka. Natural
Resources, Energy and Science Authority of Sri Lanka. Colombo., 444 pp.

44
Photographs Pictures were taken by Minolta Dynax 7,Nikon CoolPix 775, Minolta 35-70 with macro, Minolta 70-300 telemacro, 2x and 4x closeup adapter lenses, FujiFilm 800 ASA unless otherwise stated.
Figure 1 Adult golden gecko profile Calodactylodes illingworthorum, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
Figure 2 Calodactylodes illingworthorum body colouration, taken by Sura j Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 3 Calodactylodes illingworthorum, showing its ability to camouflage, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
? AntoineDrion

45
Figure 4 Calodactylodes illingworthorum coloration from the front, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 5 Gravid female Calodactylodes illingworthorum, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 6 Juvenile Calodactylodes illingworthorum, taken by Antoine Drion

46
Figure 7 Calodactylodes illingworthorum egg clutch, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 8 Calodactylodes illingworthorum eggs, taken by Antoine Drion.
Figure 9 Calodactylodes illingworthorum ectoparisties, taken by Antoine Drion.
47
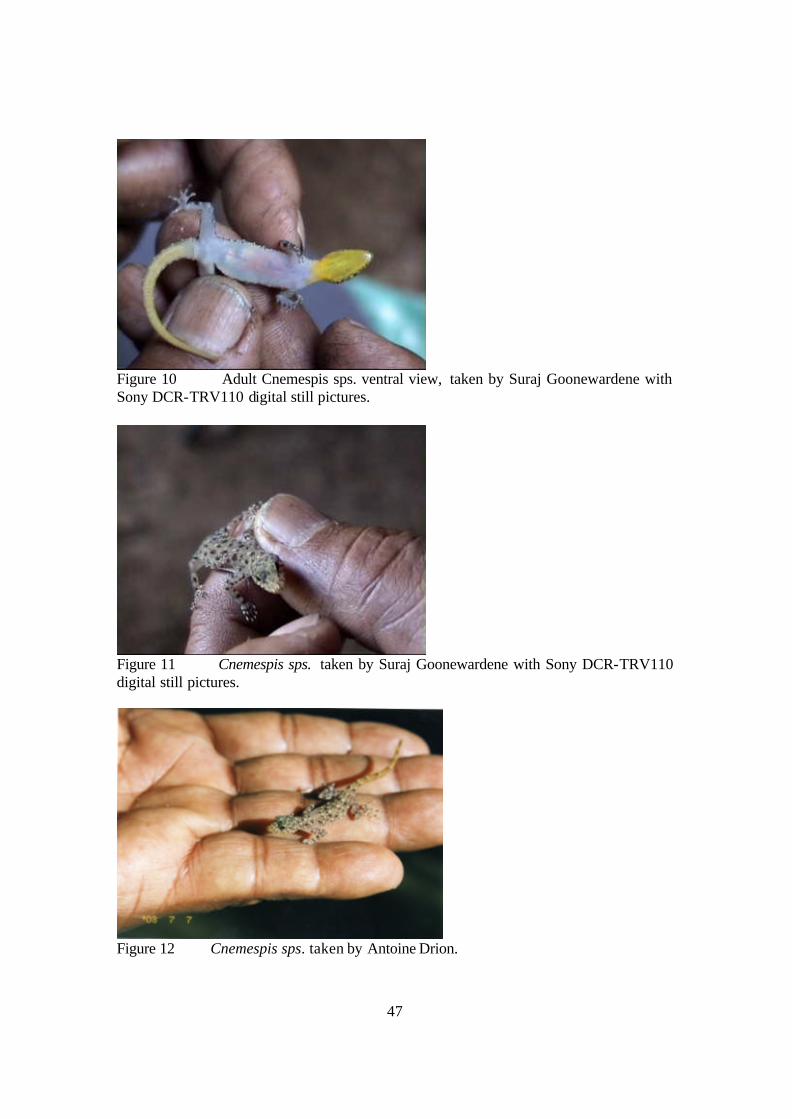
Figure 10 Adult Cnemespis sps. ventral view, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 11 Cnemespis sps. taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 12 Cnemespis sps. taken by Antoine Drion.

48
Figure 13 Adult Cnemespis sps. dorsal view, taken by Antoine Drion
Figure 14 Cnemespis sps. eggs, taken by Antoine Drion.
Figure 15 Adult Female Hemidactylus maculates hunae, dorsal view, taken by Antoine Drion

49
Figure 16 Hemidactylus maculatus hunae foot, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
Figure 17 Adult male Hemidactylus maculates hunae head, taken by Antoine Drion.
? AntoineDrione

50
Figure 18 Juvenile Hemidactylus maculatus hunae dorsal view, taken by Antoine Drion.
Figure 19 New Hemidactylus sps. dorsal view, taken by Antoine Drion
Figure 20 Hemidactylus frenatus profile, taken by Antoine Drion

51
Figure 21 Hemidactylus depressus , taken by Antoine Drion
Figure 22 Hemidactylus frenatus eggs, taken by Antoine Drion Other reptiles and amphibians
Figure 23 New Philautus sps , taken by Antoine Drion

52
Figure 24 Polypedatus crucigur. Picture, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
Figure 25 Kaloula taprobanica, taken by Antoine Drion
? AntoineDrione

53
Figure 26 New Nannophrys sps. taken by Antoine Drion
Figure 27 Ophisops minor minor, taken by Antoine Drione with Pentax M250 camera and Fuji chrome ASA 100 film.
Figure 28 Ophisops minor minor, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
? AntoineDrione
? AntoineDrione

54
Figure 29 Lankascincus fallax, taken by Antoine Drion
Figure 30 Mabuya Crinata , taken by Antoine Drion
Figure 31 Lankascincus fallax, taken by Antoine Drion

55
Figure 32 Sitana ponticeriana , taken by Antoine Drion
Figure 33 Calotes versicolor, taken by Antoine Drion
Figure 34 Adult Otocryptis wiegmanii eating golden hatchling, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
? AntoineDrione

56
Figure 35 Male Otocryptus wiegmanii, taken by Antoine Drion
Figure 36 Otocryptis wiegmanii, taken by Antoine Drion
Figure 37 Boiga Ceylonensis, taken by Antoine Drion

57
Figure 38 Gerdon Polygodon, taken by Antoine Drion
Figure 39 Dendrelaphis sps, taken by Antoine Drion
Figure 40 Dendrelaphis tristis, taken by Antoine Drion

58
Figure 41 Ahaetulla nasutus, taken by Antoine Drion
Figure 42 Ahaettulla nasutus, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
? AntoineDrione

59
Figure 43 Geochlone elegans, taken by Antoine
Figure 44 Melanochelys trijuga , taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 45 Varinus bengalensis, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.

60
Figure 46 Varinus salvator , taken by Antoine Drion Locations and habitat
Figure 47 fire savannah, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures
Figure 48 Team in Manung grass, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.

61
Figure 49 Monsoon forest, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 50 Palamugala 1, taken by Antoine Drion
Figure 51 Palamugala 2, taken by Antoine Drion

62
Figure 52 Rathugala, taken by Antoine Drion
Figure 53 Lacertid rock, taken by Antoine Drion
Figure 54 Yakunehela, taken by Antoine Drion

63
Figure 55 Reservoir near study area, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 56 Mountain range near area, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures. Other fauna and flora
Figure 57 Giant Centipede, taken by Antoine Drion

64
Figure 58 Beetle, taken by Antoine Drion
Figure 59 Scorpion. , taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
? AntoineDrione

65
Figure 70 Elephant, taken by Antoine Drion with Pentax M250 camera and Fuji chrome ASA 100 film.
Figure 71 White lily, taken by Antoine Drion
? AntoineDrione

66
Figure 72 Bat, taken by Antoine Drion
Figure 73 Dragonfly, taken by Antoine Drion
Figure 74 Caterpillar, taken by Antoine Drion

67
Figure 75 bat, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 76 Annelid, taken by Antoine Drion Threats
Figure 77 Burnt savannah, taken by Antoine Drion

68
Figure 78 Hangala burnt savannah, taken by Antoine Drion
Figure 79 Drip ledge, taken by Antoine Drion
Figure 80 Treasure stone, taken by Antoine Drion
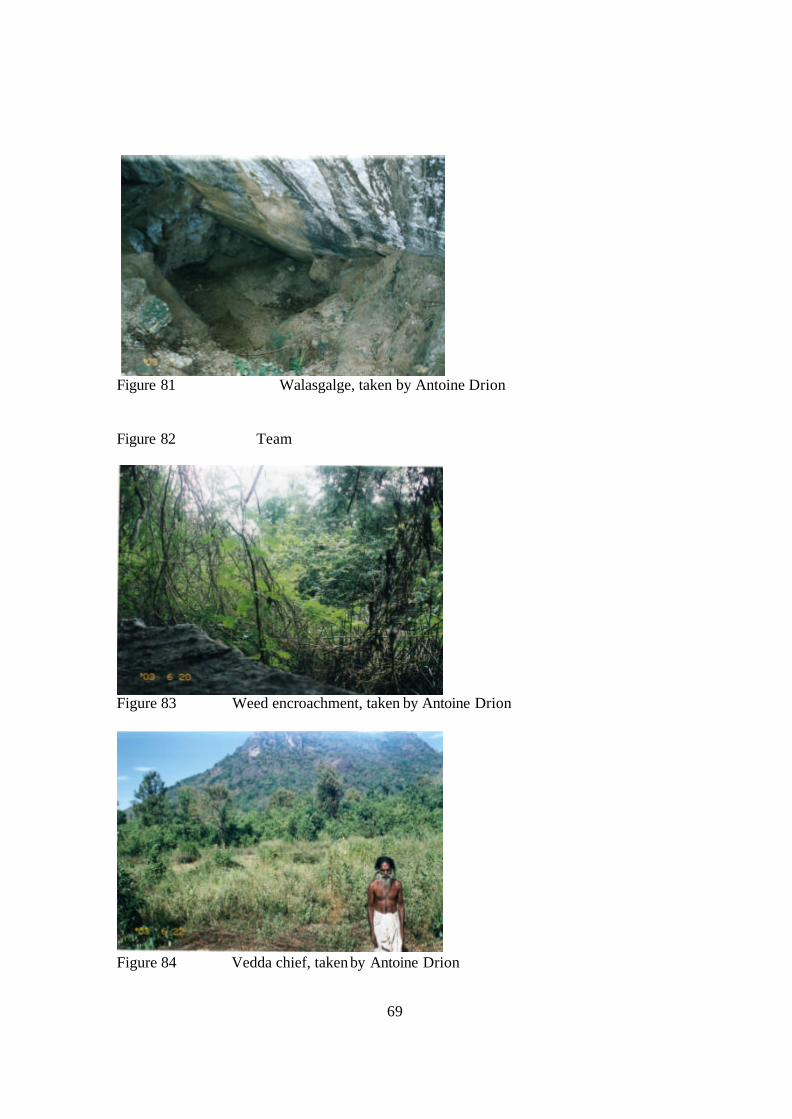
69
Figure 81 Walasgalge, taken by Antoine Drion Figure 82 Team
Figure 83 Weed encroachment, taken by Antoine Drion
Figure 84 Vedda chief, taken by Antoine Drion

70
Figure 85 Cattle grazing, taken by Antoine Drion
Figure 86 Illegal logging , taken by Antoine Drion

71
Management
Figure 87 Educating the Veddah (dance) , taken by Antoine Drion
Figure 88 KAP students, taken by Antoine Drion
Figure 89 KAP teachers, taken by Antoine Drion

72
Figure 90 Searching boulder systems, taken by Antoine Drion
Figure 91 De-weeding, taken by Antoine Drion

73
Figure 92 Janaka Perera, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
Figure 93 Ravi Jayarathne, taken by Suraj Goonewardene with Sony DCR-TRV110 digital still pictures.
74
Annexure 1- Treasury report of Project Hoona Pre Reconnaissance visit £750
Administration: i.e. postage, telephone, printing, photocopying £300
Flights £3,240
Insurance £350
Vaccinations £150
Medical kit £75
Photographic £147
Laptop computer £950
General field equipment( head torches etc) £500
Training,1st aid, workshops etc £120
In field Fuel and van hire £400
Visa extension £75
Calls and internet £25
Tips £325
Food and supplies £350
Accommodation £1100
Experts costs £400
Other(entry to National Parks) £330
Post- Printing reports & Postage to sponsors Budgeted £800
Colour separation £100
Photography: developing Budgeted £300
Camera repairs and lost equipment £72.48
Transportation of equipment and travel for research £152.25
Total £11,012

75
Annexure 2 - Notes from Seligman on Veddas Anthropological
Rathugala vedda Seligman and Punchihewa.
The Vedda Aborigines.
They constitute an insignificant fraction of the Tamil and Sinhalese in habitants before
whom they are rapidly disappearing, partly by intermarriage and absorption, partly
owing to high death rate brought about by sheer inability to change with world around
them. There is no reason to doubt that the veddas are identical with the 'Yakkas' of the
Mahavansa and other native chronicles. These wild people were hunter gathers and
were notorious for the hunting game (their use of bow and arrow) and gathering honey.
But of late most have given up these old traditions and taken up sinhalese style chena
cultivation.
In stature the veddas are short (men about 5 feet in height) and never cut their hair but
tie it up. The hair is wavy to the extent it is curly, and in old age rarely turns white.
There is little hair on the body and facial hair is limited to a thin moustache and sparse
goatee beard. Skin colour is deep brown-black. The brow ridges are well marked so
that the eyes appear deeply set, moderately prominent cheekbones and the nostrils are
moderately broad, the root of the nose is depressed.
Vedda religion is essentially a cult of the dead, it is not surprising that the propitiation
of the spirits of dead relatives, called the Nae Yaku. The most important of these is
Kande Yaka, a celebrated hunter who lived many generations ago and whose
assistance is invoked for good hunting. But over the years veddas have taken up
worshipping Buddhist and Hindu gods.
They were well known to be jealous of their women and did not let early visitors come
close to their caves when they were near. Every vedda belongs to a waruge or clan, for
example uru waruge (to which the Dabani gamm Veddas belong), tala waruge,
unapane waruge etc. The Nilgala hills (the east of the Badulla Batticaloa road) lie in
what is considered the best of the vedda domain by Seligmann.

76
The Veddas are an ancient race of people, separate to the sinhalese and Tamil
Sri Lankans whose ancestry is shaped by the frequent invasions from Northern
and Southern India. They have survived over the centuries by withdrawing deep into
the forests, retaining their primitive lifestyle despite the waves of invasions and
modernisation that swept through the rest of the country.
However there are now very few true veddas left due to intermarriage and
absorption. Also there is a high death rate associated with this kind of
lifestyle. Veddas used to belong to clans, named by female lineage and territorially
grouped, but now there are so few left that they just exist in a few family
assemblies scattered about.
General appearance: Short stature (average height is 5 feet), dark skinned
(lighter than Tamils and darker than Sinhalese), wild, wavy hair. Generally
they are scantily clad wearing just a loin cloth for modesty and do not
decorate themselves with jewelery or other garments.
True Veddas are hunter gatherers and do not cultivate the land, but move from
place to place living off animals they hunt in the forest. Mainly this is deer
which they hunt with bows and arrows, as well as the monitor lizard and anything
else they come across. They also eat a lot of honey which they gather from
hives or hollow trees which have to be split open using an axe. Traditionally
they would preserve any excess of meat they had caught in hollow trees filled
with honey. They eat their meat merely browned over the embers of their fire,
and no part of a kill is wasted, from lungs to intestines. When food is scarce,
they chew on bark to appease hunger.
As their settlements are temporary and they have few building resources, they
do not build houses, rather shelter under trees near water, or in caves which
some decorate with simple pictures of people and animals, drawn using simply
their fingers and a mixture of saliva and charcoal. They make eating implements

77
out of clay which they dry in the sun and then bake, and make their bows and
arrows from wood in the forest.
Religion- the veddas have no legends or theories of the origin of man and the
universe, but they have a vague belief in a host of nameless spirits that
inhabit the natural features around them- air, rocks, trees, glades and
springs. They also have faith in the spirits of the dead or Nae Yaku,
especially of those recently dead or more important. They appeal to these
spirits for protection and success in hunting, and fear the spirits will harm
them if ignored. Closely associated with the Nae Yaku are the spirits of a few
long dead heroes, mighty hunters, handed down by tradition. Eg Kande Yaka and
his brother Bilindi Yake. These spirits and others are invoked at ceremonies.
Rituals are carried out and complex ceremonial songs and dances, symbolically
acting out the lives of the particular spirit being invoked, are undertaken to
awaken the spirit of the deceased in the body of the dancer. Different dances
are used in different situations, and different spirits called upon. The
simplest is the arrow dance, which is performed when a spell of ill luck has
attended hunting. The Veddas will accompany the sound of their voices with
rhythmic smacking of their chest and abdomen.
Marriage: A Vedda man may marry his father’s sister’s daughter, but the children
of two sisters or two brothers may not marry as this is considered incestuous.
Individuals make life -long bonds of marriage at an early age, although the
marriage is accompanied with little ceremony. Likewise death is accompanied by
no kind of ritual. The relatives will simply cover the body with leaves and
vacate that area for several years, after which they may move back again,
sweeping the dried bones aside.

78
Annexure 3 - Journal article For GGA journal – Gecko – Article 1 OTOCRYPTIS WIEGMANNI (REPTILIA: AGAMIDAE): A GECKO PREDATOR
AT NILGALA FIRE-SAVANNAH, SRI LANKA.
Anslem de Silva [email protected]
Aaron M. Bauer
Department of Biology, Villanova University, 800 Lancaster Avenue Villanova, Pennsylvania 19085, USA
Antoine Drion
Napier University, 10, Colinton Road, Edinburgh, EH10 5DT.
Alexandra Fabry
Viator Publications (Pvt) Ltd, Sri Lanka
S. Goonawardena, Zoe Hawke and Vanessa Vanneck University of Edinburgh, UK
Otocryptis wiegmanni Wagler, 1830 (English: Sri Lankan kangaroo lizard, Sinhala: Gomu talikatussa = Forest Pendant lizard; Pinum katussa = Jumping lizard, Tali katussa = pendant lizard, Kala katussa = artistic agamid) is a small slender agamid, with adults reaching a maximum total length of 275 mm (80 mm SVL). It is a common species widely distributed in Sri Lanka from lowlands up to at least 1000 m. Although Otocryptis is mainly a ground dwelling form it may occasionally be seen climbing shrubs, trees and boulders. However, at night they climb into small shrubs where they sleep at the periphery of branches. It is an endemic reptile which is of Lower Risk – near threatened according to IUCN Red List criteria (de Silva et al., 2000). Although Otocryptis wiegmanni has never been the focus of a detailed dietary study, all previous sources have regarded them as exclusive insectivores (Buhle 1983; Manthey 1985) or as insectivores that occasionally ingest plant material, such as tender leaves (Deraniyagala 1953; Matuschka 1978). One of us (AdS) has observed Otocryptis feeding on grasshoppers and ants. However, on 22nd June 2003 around 1200 noon we observed in the Nilgala forest a male Otocryptis wiegmanni (SV = 57 mm and Tail = 162 mm long) devouring a hatchling Geckoella triedra (SVL = 00 mm and Tail = 00 mm long) (Figure 1 & 2) head first. It took approximately 5 minutes for it to ingest the gecko completely. This particular Otocryptis was captured immediately and the swallowed gecko was extracted manually with the help of forceps (Figure 3), after which the agamid was released.

79
Agamids in general are typically insectivorous, but some, especially larger species, are occasional predators on other vertebrates (Daniel 2002) and some may even feed on other lizards with some regularity (Meek et al. 2001). Agamid predation on geckos, however, has rarely been noted, perhaps in part due to a lack of overlap in activity pattern between putative predator and prey in the case of nocturnal geckos. The common garden lizard Calotes versicolor has been reported to feed on Hemidactylus brookii brookii in western India (Sharma 1992), but our observation constitutes the first published record of vertebrate predation by a member of the genus Otocryptis and the first record of gecko predation by a Sri Lankan agamid. We have observed several (usually about 10) specimens of Otocryptis wiegmanni around the boulders where Callodactylodes illingworthorum and Hemidactylus maculatus hunae abound. There are several such boulder units in the Nilgala forest. Otocryptis wiegmanni were often observed on boulders occupied by these geckos. However, the above was the first instance in which we observed an Otocryptis wiegmanni feeding on a gecko hatchling. Whether Otocryptis wiegmanni is a habitual predator on geckos needs to be investigated. A survey of the herpetofaunal knowledge of Nilgala inhabitants (de Silva et al., in prep.), however, revealed that two respondents had observed agamids feeding on geckos (interestingly, residents in the area have reported more frequent predation on geckos by frogs, possibly Rana gracilis). The relatively small size of Otocryptis suggests that only hatchling geckos or perhaps adults of the smallest species of Cnemaspis are at risk and that Otocryptis does not pose a threat to gecko populations. While it is possible that Otocryptis may locate nocturnal geckos in their diurnal retreats, this seems unlikely, as it, like virtually all agamids, is chiefly a sit-and-wait predator, ambushing mobile prey items as they come within striking range. Although Geckoella triedra is generally considered nocturnal (Deraniyagala 1953), it appears likely that the hatchling was diurnally active or was accidentally exposed by movements of its predator. Cnemaspis species are perhaps the most likely victims of O. wiegmanni predation, as they share a diurnal activity pattern. The young of the golden gecko, Calodactylodes illingworthorum, are also potential prey, as these lizards may be active by day and we have observed individuals in vegetation adjacent to the same boulder cluster where predat ion on Geckoella triedra occurred. Acknowledgments We thank Philippe Fabry for the photograph. We thank the Department of Wildlife Conservation (WL/3/2/1/14/12) and Forest Department (FRC/5 and FRC/6) for the permission granted to Anslem de Silva. Mr. H. D. Ratnayake, Director Research and Training, Department of Wildlife Conservation for his kind support in the research project. References

80
BUHLE, M. 1983. Nachzucht und Verhaltensweise von Otocryptis wiegmanni Wagler, 1830. Sauria 5(1):5-10. DANIEL, J.C. 2002. The Book of Indian Reptiles and Amphibians. Bombay Natural History Society, Mumbai. viii + 238 pp. DERANIYAGALA, P.E.P. 1931. Some Ceylon lizards. Spolia Zeylanica. 16(2):139-180. DERANIYAGALA, P.E.P. 1953. A colored atlas of some vertebrates from Ceylon. Volume 2. National Museums, Colombo, 101 pp.
DE SILVA, ANSLEM., A.BAUER, B.J.K.. PERERA, R.L.JAYARATNE, M. GOONASEKERA, S. GOONAWARDENA, Z. HAWKE, V. VANNECK & A. DRION. ( )Some aspects of the knowledge and attitude towards geckos by the students living in and around the Nilgala forest, Sri Lanka MANTHEY, U. 1985. Otocryptis wiegmanni Wagler. Sauria 7(2) (Supplement):11-12. MATUSCHKA, F.R. 1978. Beobachtungen bei der Haltung von Otocryptis wiegmanni (Reptilia: Sauria: Agamidae). Salamandra 14(4):207-211. MEEK, R., R. AVERY & E. WEIR. 2001. Physignathus lesuerii (Australian water dragon): predation on a skink (Lampropholis delicate). Herpetological Bulletin 76:31-32. PAHATHKUMBURA, M., A. JAYAWICKRAMA & ANSLEM DE SILVA. 1966. Otocryptis wiegmanni: preliminary observations. Abstracts, International Conference on the Biology & Conservation of the amphibians & reptiles of South Asia. p.23. SHARMA, S.K. 1992. Common garden lizard Calotes versicolor preying on Brook’s gecko Hemidactylus brooki. Journal of the Bombay Natural History Society 88(3):459. TAYLOR. E.H. 1953. A review of the lizards of Ceylon. Univ. of Kansas Science Bulletin. 35(12): 1525-1585.

81
SOME ASPECTS OF THE KNOWLEDGE AND ATTITUDE
TOWARDS GECKOS BY THE STUDENTS LIVING IN AND AROUND THE
NILGALA FOREST, SRI LANKA
Anslem de Silva 15/1 Dolosbage road, Gampola, Sri Lanka. Email: [email protected]
Aaron M. Bauer Department of Biology, Villanova University, 800 Lancaster Avenue
Villanova, Pennsylvania 19085, USA B.J.K.. Perera, R.L.Jayaratne & M. Goonasekera
Dept of Biological Sciences, Rajarata University of Sri Lanka S. Goonawardena, Z. Hawke, & V. Vanneck
University of Edinburgh, UK A. Drion
Napier University, Edinburgh, UK
Introduction Geckos play a significant role in the culture of Sri Lankan society. It is widely believed that various activities of geckos, such as vocalization, defecation, or falling from a roof or wall on to one’s body presage either good luck or misfortune. This belief is so deep-rooted that even today many people, when leaving their houses, may wait for some time if they hear a gecko vocalizing as it is believed that the gecko foresees some impending misfortune to that particular person. The most popular almanac in Sri Lanka, “Epa panchanga litha,” which has been published since 1855, continues to have a section on “hunu handa palapala” (= predictions based on gecko cries), which forecasts predictions based on gecko calls, and another on the “hunu enga vetimae palapala” (= predictions based on the part of the body on which a gecko falls). The predictive value of the direction of gecko calls and of the part of the body on which a gecko falls is also maintained in much of India, but these predictions are especially detailed in south India, where they are recorded in Tamil almanacs (Mahendra, 1936). The similarities between the Indian and Sri Lankan almanacs suggest a common origin for these beliefs. The Nilgala fire savannah is a main habitat of the Sri Lankan Golden gecko (Callodactylodes illingworthorum), and the Spotted Giant-gecko (Hemidactylus maculatus hunae). The former is locally known by the villagers as Gal húná (= rock gecko), however, to the Veddha (hunter-gatherer descendents of the ancient aboriginal race of Sri Lanka) Callodactylodes illingworthorum is known as Galpahuro (= rock climbers or rock rafters). The Spotted Giant-gecko is known locally as Davanta tit húná, and the Veddhas call it Kotakka. These geckos face many threats, natural and anthropogenic. In fact, we observed three former Callodactylodes illingworthorum sites that have been completely deserted by geckos, possibly over five years ago. These were almost certainly abandoned due to human interference, or the gecko population may have been deliberately killed. In order to prevent such actions and to conserve and manage gecko populations it is vital that we understand the socio-economic circumstances of the local human inhabitants. In particular, an understanding of their knowledge, attitudes and practices towards the fauna and flora is essential. Thus, as part of a recent 36 day field program designed to survey

82
the herpetofauna of the Nilgala fire savannah, East Sri Lanka, we investigated the knowledge, beliefs, and actions of school students in the region with respect to geckos. Information was gathered by means of a structured questionnaire administered on 3 July 2003 to eighty-one school children between ages 11 to 18 from two schools (Galgamuwa and Bulupitiya). Results The responses of the students are given in tables 1 to 4 Discussion and recommendations Of the 81 students who answered the questionnaire 74 (91%) students have observed Callodactylodes illingworthorum. Of these, six students have seen seven or more spec imens of this species. In India, despite its large size, conspicuous color, semi-diurnal habits and distinctive vocalization, the related species C. aureus has escaped notice by most people where it is common (Bauer & Das, 2000). However, in Sri Lanka many villagers, when asked about the golden gecko, replied that they had seen it and were able to guide us to localities supporting this species. Our data suggest that local children have a close association with nature and knowledge of the fauna and flora (T able 2). However, 14 (17 %) of the students have killed geckos (Table 4), possibly because most (52%) believe that C. illingworthorum is poisonous and 79% considered other geckos as poisonous. This is consistent with widely held beliefs. Indeed, according to the traditional medical system, both geckos and skinks are considered poisonous. Many books on traditional medicine for snakebite treatment record this (Roberts, 1919; de Silva, 1993). The view of geckos as poisonous is widespread around the world (Rösler, 1995). Elsewhere in Asia they have been thought to cause leprosy and skin eruptions and to cause poisoning either through their bites or through the fouling of milk by their saliva (Mahendra, 1936), and in Africa they are believed to cause hysteria, ultimately resulting in death (Auerbach 1995). School children near Nilgala report having observed a variety of predation events involving geckos (Table 3). Although our questions did not distinguish between the types of setting (forest versus homes) in which predation events were observed, the listing of dogs and cats as predators suggest that many events were observed in houses or compounds. However, as most of these people are encroachers into the Nilgala forest, thus the so called “compound” is in matter of fact part of the forest. Cats in particular, were significant predators on geckos (56% of predation events) as has been documented in earlier surveys (de Silva, 1996). Although there are no previously published records of agamid predation on geckos in Sri Lanka, two students claim to have observed such activity. During our field work in the Nilgala forest we also observed an Otocryptis wiegmanni Wagler, 1830 (English: Sri Lankan kangaroo lizard) feeding on a gecko (Geckoella triedra; de Silva, et al 2003a) and so were able to confirm the student’s report of agamid predation on geckos. Two students also reported ant predation on geckos. We have not observed attacks on adult or juvenile geckos, but we have documented ant predation on the eggs of Calodactylodes illingworthorum (de Silva, et al 2003b). Among native predators reported by students, frogs are perhaps the most surprising; however, we observed several large ranids (Rana gracilis and Polypedates cruciger) in Golden gecko and Spotted Giant-gecko habitats in the forest, and it is possible that these large species could swallow a juvenile or sub adult gecko. We also observed several species of snakes, some of which are voracious feeders on geckos (e.g., Boiga ceylonensis).
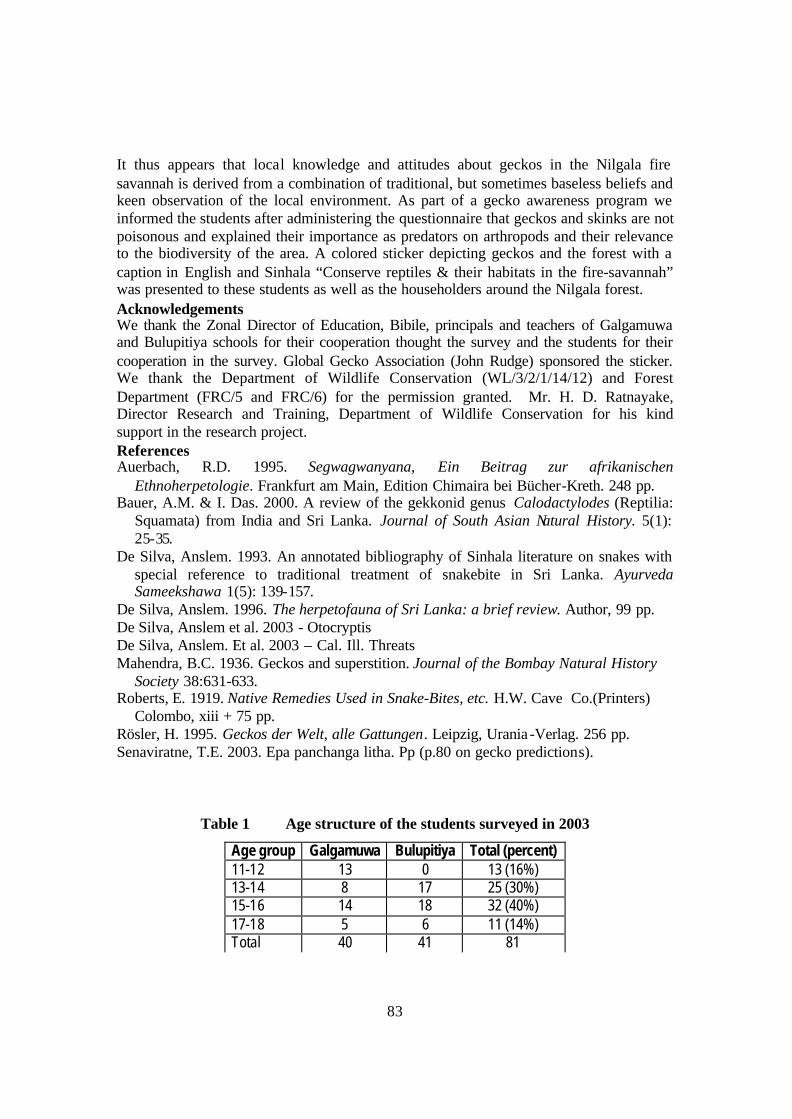
83
It thus appears that local knowledge and attitudes about geckos in the Nilgala fire savannah is derived from a combination of traditional, but sometimes baseless beliefs and keen observation of the local environment. As part of a gecko awareness program we informed the students after administering the questionnaire that geckos and skinks are not poisonous and explained their importance as predators on arthropods and their relevance to the biodiversity of the area. A colored sticker depicting geckos and the forest with a caption in English and Sinhala “Conserve reptiles & their habitats in the fire-savannah” was presented to these students as well as the householders around the Nilgala forest. Acknowledgements We thank the Zonal Director of Education, Bibile, principals and teachers of Galgamuwa and Bulupitiya schools for their cooperation thought the survey and the students for their cooperation in the survey. Global Gecko Association (John Rudge) sponsored the sticker. We thank the Department of Wildlife Conservation (WL/3/2/1/14/12) and Forest Department (FRC/5 and FRC/6) for the permission granted. Mr. H. D. Ratnayake, Director Research and Training, Department of Wildlife Conservation for his kind support in the research project. References Auerbach, R.D. 1995. Segwagwanyana, Ein Beitrag zur afrikanischen
Ethnoherpetologie. Frankfurt am Main, Edition Chimaira bei Bücher-Kreth. 248 pp. Bauer, A.M. & I. Das. 2000. A review of the gekkonid genus Calodactylodes (Reptilia:
Squamata) from India and Sri Lanka. Journal of South Asian Natural History. 5(1): 25-35.
De Silva, Anslem. 1993. An annotated bibliography of Sinhala literature on snakes with special reference to traditional treatment of snakebite in Sri Lanka. Ayurveda Sameekshawa 1(5): 139-157.
De Silva, Anslem. 1996. The herpetofauna of Sri Lanka: a brief review. Author, 99 pp. De Silva, Anslem et al. 2003 - Otocryptis De Silva, Anslem. Et al. 2003 – Cal. Ill. Threats Mahendra, B.C. 1936. Geckos and superstition. Journal of the Bombay Natural History
Society 38:631-633. Roberts, E. 1919. Native Remedies Used in Snake-Bites, etc. H.W. Cave Co.(Printers)
Colombo, xiii + 75 pp. Rösler, H. 1995. Geckos der Welt, alle Gattungen. Leipzig, Urania -Verlag. 256 pp. Senaviratne, T.E. 2003. Epa panchanga litha. Pp (p.80 on gecko predictions).
Table 1 Age structure of the students surveyed in 2003
Age group Galgamuwa Bulupitiya Total (percent) 11-12 13 0 13 (16%) 13-14 8 17 25 (30%) 15-16 14 18 32 (40%) 17-18 5 6 11 (14%) Total 40 41 81
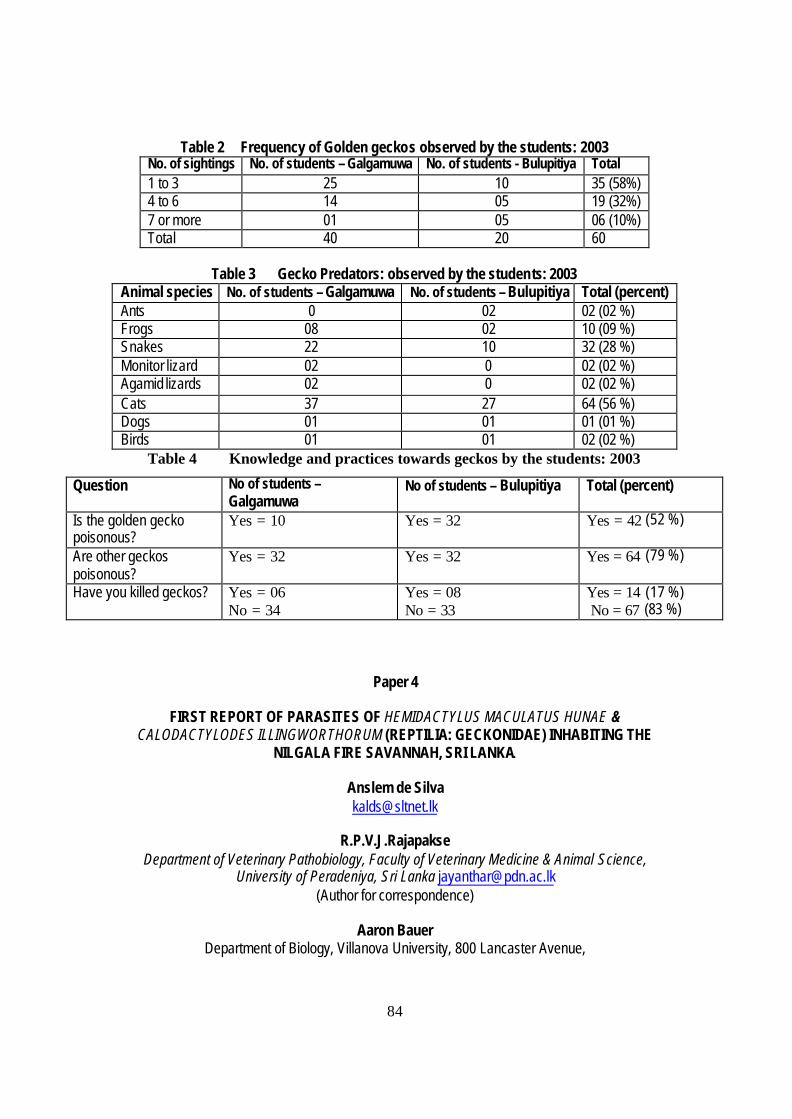
84
Table 2 Frequency of Golden geckos observed by the students: 2003 No. of sightings No. of students – Galgamuwa No. of students - Bulupitiya Total 1 to 3 25 10 35 (58%) 4 to 6 14 05 19 (32%) 7 or more 01 05 06 (10%) Total 40 20 60
Table 3 Gecko Predators: observed by the students: 2003
Animal species No. of students – Galgamuwa No. of students – Bulupitiya Total (percent) Ants 0 02 02 (02 %) Frogs 08 02 10 (09 %) Snakes 22 10 32 (28 %) Monitor lizard 02 0 02 (02 %) Agamid lizards 02 0 02 (02 %) Cats 37 27 64 (56 %) Dogs 01 01 01 (01 %) Birds 01 01 02 (02 %)
Table 4 Knowledge and practices towards geckos by the students: 2003
Question No of students – Galgamuwa
No of students – Bulupitiya Total (percent)
Is the golden gecko poisonous?
Yes = 10
Yes = 32
Yes = 42 (52 %)
Are other geckos poisonous?
Yes = 32
Yes = 32
Yes = 64 (79 %)
Have you killed geckos?
Yes = 06 No = 34
Yes = 08 No = 33
Yes = 14 (17 %) No = 67 (83 %)
Paper 4
FIRST REPORT OF PARASITES OF HEMIDACTYLUS MACULATUS HUNAE & CALODACTYLODES ILLINGWORTHORUM (REPTILIA: GECKONIDAE) INHABITING THE
NILGALA FIRE SAVANNAH, SRI LANKA.
Anslem de Silva [email protected]
R.P.V.J.Rajapakse
Department of Veterinary Pathobiology, Faculty of Veterinary Medicine & Animal Science, University of Peradeniya, Sri Lanka [email protected]
(Author for correspondence)
Aaron Bauer Department of Biology, Villanova University, 800 Lancaster Avenue,

85
Villanova, Pennsylvania 19085, USA
W. M. J. de Silva Department of Veterinary Pathobiology, Faculty of Veterinary Medicine & Animal Science,
University of Peradeniya, Sri Lanka
Christopher C. Austin Museum of Natural Science, Louisiana State University,
119 Foster Hall, Baton Rouge, LA 70803 USA
S. Goonawardena University of Edinburgh, UK
M. Goonasekera
Dept of Biological Sciences, Rajarata University of Sri Lanka
Introduction The two largest geckos in Sri Lanka are the golden gecko, Calodactylodes illingworthorum, (Figure 000) and the spotted giant gecko, Hemidactylus maculatus hunae (Figure 00). The former species reaches a snout vent length (SVL) of 100 mm, whereas the latter is one of the largest species of the diverse genus Hemidactylus in the world. Specimens measuring 280 to 300 mm in total long have been observed (de Silva, et al, 2004). The Golden gecko has been postulated to be a Gondwanan relict (Russell and Bauer 1990; Bauer and Das 2000). It is found in appreciable numbers on rock outcrops (large boulders and cave systems) in the savannah and monsoon forests of the Monaragala and Ampara districts. Until very recently almost nothing was known about the golden gecko and observations on its biology remain very limited (Wickramasinghe and Somaweera 2003). The same may be said of H. maculatus hunae. Indeed, despite the pioneering efforts of Deraniyagala (1931, 1953), this remains true for most Sri Lankan geckos today. While much information is available regarding the parasites of Indian species, as well as some Sri Lankan species that are also widespread in India (von Linstrow, 1906; Baylis, 1936; Soond, 1999), parasitism remains a particularly neglected aspect of Sri Lankan gecko biology. The first significant contributions, however, were studies on the helminth parasites of Hemidactylus brookii parvimaculatus, H. triedrus lankae and Cnemaspis kandianus from Sri Lanka conducted by Crusz and coworkers (Crusz & Ching, 1975, Crusz & Nugaliyadde, 1977, Crusz, 1984) as part of a broader survey of the parasites of the endemic elements of the Sri Lankan vertebrates. Crusz (1984) reported Paradistomum sp. from Hemidactylus brooki parvimaculatus and Haemogrgarina treidri, Trypanosoma pertenue from Hemidactylus triedrus lankae. The helminth Acanthocephalus serndibensis was reported from the small intestine of the dwarf day-gecko Cnemaspis kandianus, collected from Haputale (1,418 m) (Crusz & Ching, 1975) (Table 4). However, no parasitological investigations have been conducted on Calodactylodes illingworthorum and Hemidactylus maculatus hunae. We here report for the first time preliminary investigations on the parasites of these large endemic Sri Lankan geckos. We also present data on the distribution of the parasites according to habitat type.
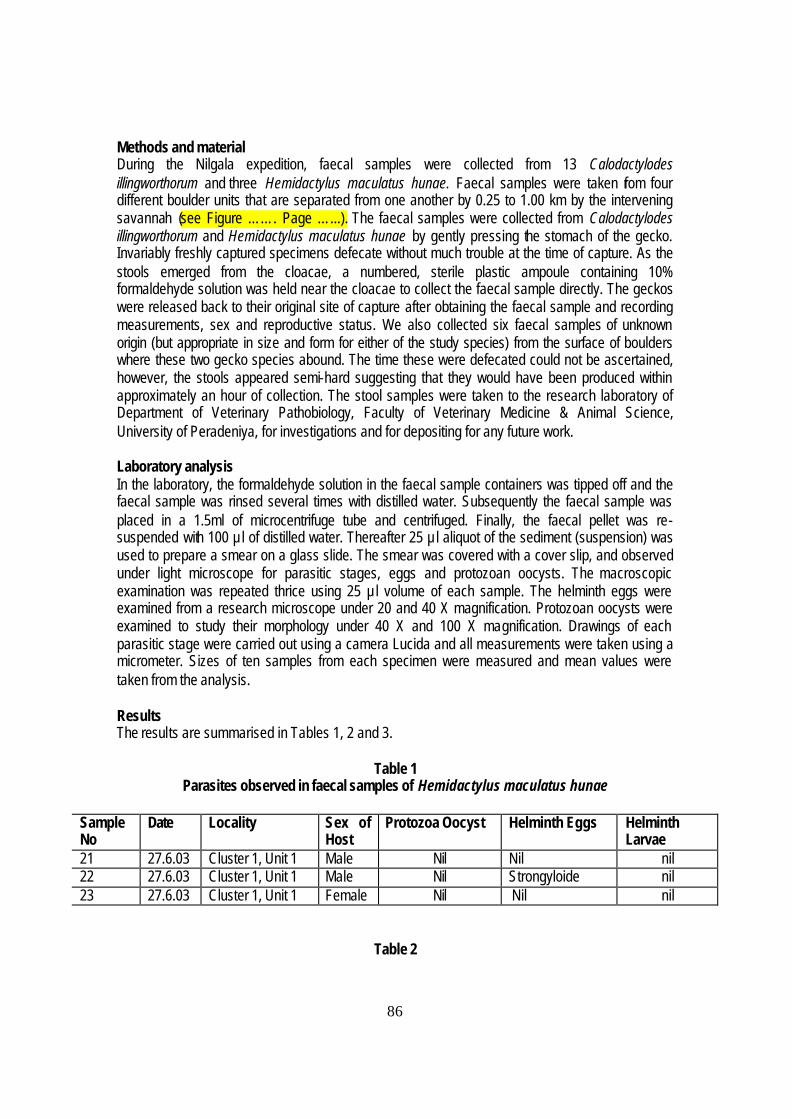
86
Methods and material During the Nilgala expedition, faecal samples were collected from 13 Calodactylodes illingworthorum and three Hemidactylus maculatus hunae. Faecal samples were taken from four different boulder units that are separated from one another by 0.25 to 1.00 km by the intervening savannah (see Figure ……. Page …...). The faecal samples were collected from Calodactylodes illingworthorum and Hemidactylus maculatus hunae by gently pressing the stomach of the gecko. Invariably freshly captured specimens defecate without much trouble at the time of capture. As the stools emerged from the cloacae, a numbered, sterile plastic ampoule containing 10% formaldehyde solution was held near the cloacae to collect the faecal sample directly. The geckos were released back to their original site of capture after obtaining the faecal sample and recording measurements, sex and reproductive status. We also collected six faecal samples of unknown origin (but appropriate in size and form for either of the study species) from the surface of boulders where these two gecko species abound. The time these were defecated could not be ascertained, however, the stools appeared semi-hard suggesting that they would have been produced within approximately an hour of collection. The stool samples were taken to the research laboratory of Department of Veterinary Pathobiology, Faculty of Veterinary Medicine & Animal Science, University of Peradeniya, for investigations and for depositing for any future work. Laboratory analysis In the laboratory, the formaldehyde solution in the faecal sample containers was tipped off and the faecal sample was rinsed several times with distilled water. Subsequently the faecal sample was placed in a 1.5ml of microcentrifuge tube and centrifuged. Finally, the faecal pellet was re-suspended with 100 µl of distilled water. Thereafter 25 µl aliquot of the sediment (suspension) was used to prepare a smear on a glass slide. The smear was covered with a cover slip, and observed under light microscope for parasitic stages, eggs and protozoan oocysts. The macroscopic examination was repeated thrice using 25 µl volume of each sample. The helminth eggs were examined from a research microscope under 20 and 40 X magnification. Protozoan oocysts were examined to study their morphology under 40 X and 100 X magnification. Drawings of each parasitic stage were carried out using a camera Lucida and all measurements were taken using a micrometer. Sizes of ten samples from each specimen were measured and mean values were taken from the analysis. Results The results are summarised in Tables 1, 2 and 3.
Table 1
Parasites observed in faecal samples of Hemidactylus maculatus hunae
Sample No
Date Locality Sex of Host
Protozoa Oocyst Helminth Eggs Helminth Larvae
21 27.6.03 Cluster 1, Unit 1 Male Nil Nil nil 22 27.6.03 Cluster 1, Unit 1 Male Nil Strongyloide nil 23 27.6.03 Cluster 1, Unit 1 Female Nil Nil nil
Table 2
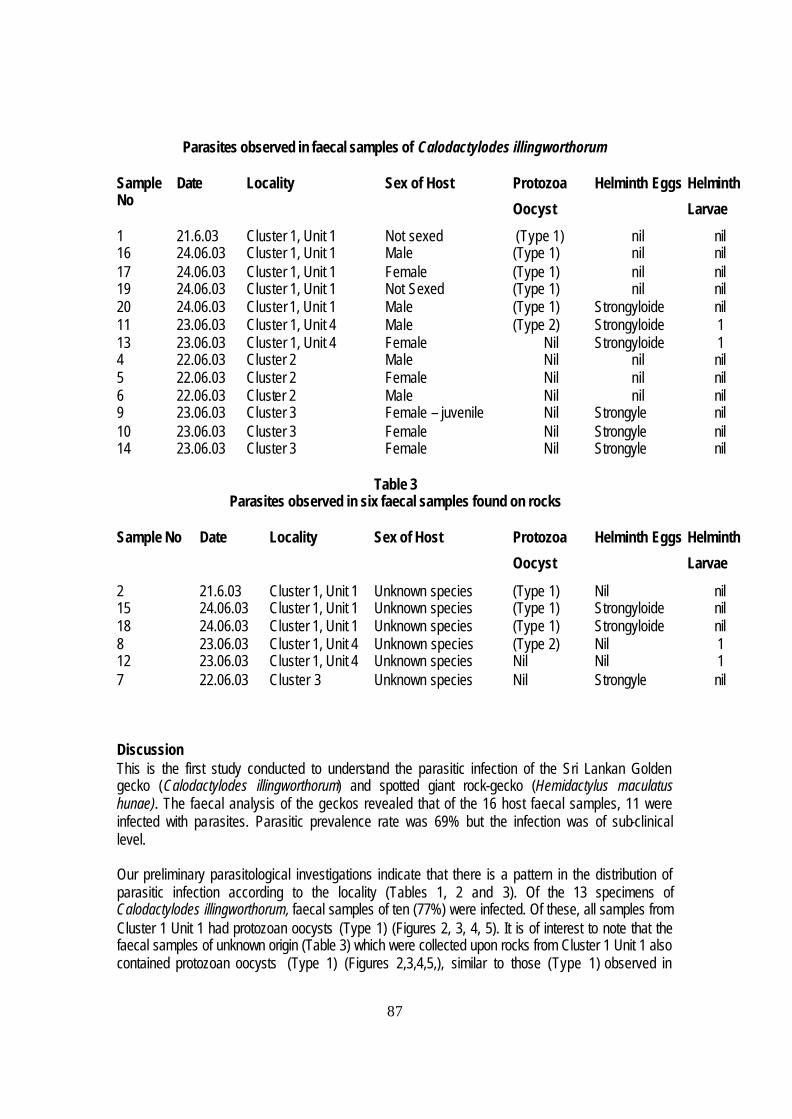
87
Parasites observed in faecal samples of Calodactylodes illingworthorum Sample No
Date Locality Sex of Host Protozoa
Oocyst
Helminth Eggs Helminth
Larvae
1 21.6.03 Cluster 1, Unit 1 Not sexed (Type 1) nil nil 16 24.06.03 Cluster 1, Unit 1 Male (Type 1) nil nil 17 24.06.03 Cluster 1, Unit 1 Female (Type 1) nil nil 19 24.06.03 Cluster 1, Unit 1 Not Sexed (Type 1) nil nil 20 24.06.03 Cluster 1, Unit 1 Male (Type 1) Strongyloide nil 11 23.06.03 Cluster 1, Unit 4 Male (Type 2) Strongyloide 1 13 23.06.03 Cluster 1, Unit 4 Female Nil Strongyloide 1 4 22.06.03 Cluster 2 Male Nil nil nil 5 22.06.03 Cluster 2 Female Nil nil nil 6 22.06.03 Cluster 2 Male Nil nil nil 9 23.06.03 Cluster 3 Female – juvenile Nil Strongyle nil 10 23.06.03 Cluster 3 Female Nil Strongyle nil 14 23.06.03 Cluster 3 Female Nil Strongyle nil
Table 3
Parasites observed in six faecal samples found on rocks Sample No Date Locality Sex of Host Protozoa
Oocyst
Helminth Eggs Helminth
Larvae
2 21.6.03 Cluster 1, Unit 1 Unknown species (Type 1) Nil nil 15 24.06.03 Cluster 1, Unit 1 Unknown species (Type 1) Strongyloide nil 18 24.06.03 Cluster 1, Unit 1 Unknown species (Type 1) Strongyloide nil 8 23.06.03 Cluster 1, Unit 4 Unknown species (Type 2) Nil 1 12 23.06.03 Cluster 1, Unit 4 Unknown species Nil Nil 1 7 22.06.03 Cluster 3 Unknown species Nil Strongyle nil Discussion This is the first study conducted to understand the parasitic infection of the Sri Lankan Golden gecko (Calodactylodes illingworthorum) and spotted giant rock-gecko (Hemidactylus maculatus hunae). The faecal analysis of the geckos revealed that of the 16 host faecal samples, 11 were infected with parasites. Parasitic prevalence rate was 69% but the infection was of sub-clinical level. Our preliminary parasitological investigations indicate that there is a pattern in the distribution of parasitic infection according to the locality (Tables 1, 2 and 3). Of the 13 specimens of Calodactylodes illingworthorum, faecal samples of ten (77%) were infected. Of these, all samples from Cluster 1 Unit 1 had protozoan oocysts (Type 1) (Figures 2, 3, 4, 5). It is of interest to note that the faecal samples of unknown origin (Table 3) which were collected upon rocks from Cluster 1 Unit 1 also contained protozoan oocysts (Type 1) (Figures 2,3,4,5,), similar to those (Type 1) observed in

88
Calodactylodes illingworthorum. Thus, we could assume that these could be the faecal samples of Calodactylodes illingworthorum. As regards the faecal samples from the three Hemidactylus maculatus hunae, only one was infected with Strongyloide type parasite [Anslem – it would be useful if the parasitologist can provided a brief – one sentence or so, definition of strongyle vs strongyloide types as this will be unfamiliar to most herpetologists]. This was similar to the Strongyloide type parasite observed in Calodactylodes illingworthorum of the same boulder unit, Cluster 1 Unit 1. If indeed these strongyloides are identical, this would indicate that this nematode parasite is not specific to a single host, but possibly specific to the locality of host. The two faecal samples of Calodactylodes illingworthorum from Cluster 1 Unit 4 had a Strongyloide type of parasite similar to the ones observed in specimens from Cluster 1 Unit 1. However, one (sample 11) had different protozoan oocysts (Type 2). Similarly, the unknown host faecal sample from Cluster 1 Unit 4 also contained oocysts of this type (Table 3). All three samples of C. illingworthorum from Cluster 2 were negative, whilst all the samples from Cluster 3 had Strongyle type helminth eggs. It is of interest to note that the host unknown faecal sample from Cluster 3 too had Strongyle type helminth eggs (Table 3). Four animals had protozoan infections and their oocysts were similar to coccidian parasites, however, further investigations are needed to identify the species. However, according to morphological appearances of the nematode eggs, we grouped them in to five categories according to the size of the eggs [should there be a table or figure citation here? It is confusing how these categories relate to the two categories below]. Two types of helminth eggs were observed in the present study. From the morphological appearances of the eggs, they can be divided into two strongyle eggs where as eggs with morala stages and eggs of other group is very similar to strogyloides where fully developed larvae too were observed. Therefore according there morphological feature the eggs seen in this study, two distinct categories: strongyle and strongyloides eggs (Figures 1, 2, 00, 00, 00 & 00) were identified. Only faecal samples from geckos (including the unknown host faecal sample from this locality) showed helminth larval stages in the faeces. Morphologically this larva was similar to stronglye type (Figure 3, 00). Similarly, it has been noticed that 10 (48%) of 21 animals are infected with protozoan oocysts which are very similar to coccidian oocysts. However according to morphology of the oocysts, two type of oocysts were observed: Type 1 in cluster 1 unit 1 and Type 2 cluster 1 unit 4 (Figure 4 and 5). Following are some parasites that have been identified from three species of endemic geckos by Crusz and his team (Table 4).
Table 4 Parasites observed in Sri Lankan geckos in previous studies
Gecko species Parasite Authority

89
Cnemaspis kandianus Acanthocephalus serndibensis Crusz & Ching, 1975 Hemidactylus brooki parvimaculatus Paradistomum sp. Crusz, 1984 Hemidactylus triedrus lankae. Haemogregarina treidri,
Trypanosoma pertenue (both protozoan parasites)
Crusz, 1984
Chabaud & Brygoo (1964) have also shown that isolation has had its effects in the evolution of particular parasite species peculiar to the host(s) on which they are found. From Sri Lanka, Crusz and his co-workers too demonstrated this fact from a series of studies on parasites of the relict fauna of Sri Lanka (Crusz & Ching, 1975; Crusz, 1984). Our preliminary investigations show some uniformity in the spread of parasitic infestation at particular localities. This isolation could be a recent phenomenon due to heavy logging and clearing of the monsoon forests which have resulted in the formation of the savannah, which in turn restricts the geckos from moving freely from one boulder unit to the others. Thus, could it lead to some sub-populations of these animals. This study also indicates the importance of identification of these parasitic species, whether these parasites cause any clinical effects to the host, and if they are of any zoonotic significance. [Anslem – please check the paragraph above – the wording is not quite clear, I have tried to modify and clarify it in some parts] Undoubtedly the patterns of parasite infestation of Sri Lankan geckos are complex and reflect a combination of host-specificity and site-specificity with broader host utilization. The latter pattern is suggested by our data, but our samples are too restricted to generalize, even for the two host species studied. Further, any interpretation of the evolution of current parasite distributions in Sri Lanka will require a more complete knowledge of the systematics and life histories of the parasites involved. Acknowledgments We thank to N. A. N. D. Perera, Department of Veterinary Pathobiology for the photographs. Z. Hawke, V. Vanneck and A. Drion, members of the University of Edinburgh, UK expedition team for their assistance during the survey. The Department of Wildlife Conservation (WL/3/2/1/14/12) and Forest Department (FRC/5 and FRC/6) for the permission granted to AdS, AB and CA. Mr. H. D. Ratnayake, Director Research and Training, Department of Wildlife Conservation for his kind support in the research project. Reference BAUER, A.M. & I. DAS. 2000. A review of the gekkonid genus Calodactylodes (Reptilia: Squamata) from India and Sri Lanka. Journal of South Asian Natural History. 5(1): 25-35. BAYLIS, H.A. 1936. The fauna of British India, including Ceylon and Burma CRUSZ, H. AND C. C. CHING. 1975. Parasites of the relict fauna of Ceylon, vi more new helminths form amphibians and reptiles, with a new host-record and redescription of Acanthocephalus serendibensis Crusz and Mills, 1970. Annals de Parasitologie (Paris) 50(5): 531-558 CHABAUD, A. G. & E. R. BRYGOO. 1964. L’endémisme chez les helminthes de Madascar. C. R. Society of Biogéography. 356: 3-13.

90
CRUSZ, H. AND L. NUGALIYADDE. 1977. Parasites of the relict fauna of Ceylon; VII. General consideration and first host-parasite checklist; C.R. Soc. Biogeor. 477: 85-106. Nematoda Vol. I. (Ascaroidea and Strongyloidea). Taylor and Francis, London. 408 pp. CRUSZ, H. 1984. Parasites of endemic and relict vertebrates: a biogeographical review. In: Ecology and Biogeography in Sri Lanka (Ed.) C. H. Fernando. Dr. W. Junk Publishers, The Hague. pp.321-351 CRUSZ, H. 1986. The vertebrates of Sri Lanka: endemism and other aspects. Rep. Soc. Res. Native Livestock 11: 65-85. DERANIYAGALA, P.E.P. 1931. Some Ceylon lizards. Spolia Zeylanica. 16(2):139 -180, pls. 33-38. DERANIYAGALA, P.E.P. 1953. A coloured atlas of some vertebrates from Ceylon . Volume 2. National Museums, Colombo. xviii + 101 pp., 11 black and white pls., 35 color plates. LINSTROW, O. VON. 1906. Helminths from the collection of the Colombo Museum. Spolia Zeylanica 3(11):163-168, pls. i-iii. RUSSELL, A. P. and A. M. BAUER. 1989. The morphology of the digits of the golden gecko, Calodactylodes aureus (Reptilia: Gekkonidae) and its implications for the occupation of rupicolous habitats. Amphibia-Reptilia 10:125-140. SOOD, M.L. 1999. Reptilian nematodes from South Asia. International Book Distributors, Dehra Dun. xi + 299 pp. WICKRAMASINGHE, M. and R. SOMAWEERA. 2003. Distribution and current status of the endemic geckos of Sri Lanka. Gekko 3(1):2-13. Not completed
FIRST REPORT OF MITES from CALODACTYLODES ILLINGWORTHORUM & MABUYA …………..INHABITING THE NILGALA FIRE SAVANNAH, SRI LANKA.
R.P.V.J.Rajapakse Department of Veterinary Pathobiology, Faculty of Veterinary Medicine & Animal Science,
University of Peradeniya, Sri Lanka [email protected]
Anslem de Silva [email protected]
Aaron Bauer.

91
Department of Biology, Villanova University, 800 Lancaster Avenue, Villanova, Pennsylvania 19085, USA
Christopher C. Austin
Museum of Natural Science, Louisiana State University, 119 Foster Hall, Baton Rouge, LA 70803 USA
D. Ekanayake Department of Veterinary Pathobiology, Faculty of Veterinary Medicine & Animal Science,
University of Peradeniya, Sri Lanka
S. Goonawardena University of Edinburgh, UK
Ecto parasites are common among wild reptiles in Sri Lanka. Some of these are the …………. …………… Snakes place attached Tortoises place attached Flapshell turtle place attached Geckos place attached Skinks place attached Biology of the mite





























