Embed Size (px)
Citation preview

Distribution and Systematicsof the Rabbits (Sylvilagus)of West-Central Mexico
VICTOR E. DIERSING
and
DON E. WILSON
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 297

SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION
Emphasis upon publication as a means of "diffusing knowledge" was expressedby the first Secretary of the Smithsonian. In his formal plan for the Institution, JosephHenry outlined a program that included the following statement: "It is proposed topublish a series of reports, giving an account of the new discoveries in science, andof the changes made from year to year in all branches of knowledge." This themeof basic research has been adhered to through the years by thousands of titles issuedin series publications under the Smithsonian imprint, commencing with SmithsonianContributions to Knowledge in 1848 and continuing with the following active series:
Smithsonian Contributions to AnthropologySmithsonian Contributions to Astrophysics
Smithsonian Contributions to BotanySmithsonian Contributions to the Earth Sciences
Smithsonian Contributions to PaleobiologySmithsonian Contributions to Zoo/ogySmithsonian Studies in Air and Space
Smithsonian Studies in History and Technology
In these series, the Institution publishes small papers and full-scale monographsthat report the research and collections of its various museums and bureaux or ofprofessional colleagues in the world cf science and scholarship. The publications aredistributed by mailing lists to libraries, universities, and similar institutions throughoutthe world.
Papers or monographs submitted for series publication are received by theSmithsonian Institution Press, subject to its own review for format and style, onlythrough departments of the various Smithsonian museums or bureaux, where themanuscripts are given substantive review. Press requirements for manuscript and artpreparation are outlined on the inside back cover.
S. Dillon RipleySecretarySmithsonian Institution

S M I T H S O N I A N C O N T R I B U T I O N S T O Z O O L O G Y • N U M B E R 2 9 7
Distribution and Systematicsof the Rabbits {Sylvilagus)of West-Central Mexico
Victor E. Diersingand Don E. Wilson
SMITHSONIAN INSTITUTION PRESS
City of Washington
1980

A B S T R A C T
Diersing, Victor E., and Don E. Wilson. Distribution and Systematics of theRabbits (Sylvilagus) of West-Central Mexico. Smithsonian Contributions toZoology, number 297, 34 pages, 12 figures, 3 tables, 1980.—Distribution patternsof Sylvilagus cunicularius, S. floridanus, S. graysoni, and S. audubonii are docu-mented for west-central Mexico including the states of Sinaloa, Durango, Nayarit,Zacatecas, Aguascalientes, Jalisco, Guanajuato, Colima, and Michoacan. A taxo-nomic revision of the four species in this area is presented recognizing two sub-species of S. cunicularius, three of S. floridanus, two of S. graysoni, and two of5. audubonii. Two subspecies of S. floridanus (S. /. restrictus and S. /. subcinctus)are relegated to the synonomy of S. /. orizabae. Two subspecies are described asnew, one each of 5. graysoni and S. floridanus. The karyotype of S. graysoni ispresented and figured. External and cranial measurements for all species andsubspecies are presented.
OFFICIAL PUBLICATION DATE is handstamped in a limited number of initial copies and is recordedin the Institution's annual report, Smithsonian Year. SERIES COVER DESIGN: The coral Montastreacavernosa (Linnaeus).
Library of Congress cataloging in Publication DataDiersing, Victor EDistribution and systematics of the rabbits (Sylvilagus) of west-central Mexico.(Smithsonian contributions to zoology ; no. 297Bibliography: p.1. Cottontails—Classification. 2. Cottontails—Geographical distribution. 3. Mammals—Classi-
fication. 4. Mammals—Geographical distribution. 5. Mammals—Mexico. I. Wilson, Don E.,joint author. II. Title. III. Series: Smithsonian Institution. Smithsonian contributions tozoology ; no. 297.
QL737.L32D53 599'.322 79-607046

Contents
PageIntroduction 1
Acknowledgments 1Methods 2Recognition of Species 3Evolution of Sylvilagus graysoni 6Sylvilagus cunicularius insolitus (J. A. Allen) 7Sylvilagus cunicularius cunicularius (Waterhouse) 8Sylvilagus floridanus macrocorpus, new subspecies 8Sylvilagus floridanus holzneri (Mearns) 12Sylvilagus floridanus orizabae (Merriam) 13Sylvilagus graysoni graysoni (J. A. Allen) 14Sylvilagus graysoni badistes, new subspecies 15Sylvilagus audubonii parvulus (J. A. Allen) 15Sylvilagus audubonii goldmani (Nelson) 17Summary 17Appendix: Measurements and Discriminant Multipliers 19Literature Cited 34
in


Distribution and Systematic'sof the Rabbits {Sylvilagus)of West-Central Mexico
Victor E. Diersing
and Don E. Wilson
Introduction
There has been no comprehensive account of therabbits of west-central Mexico since Nelson's (1909)revision of North American rabbits. More recentregional studies within west-central Mexico includea survey of the rabbits of Sinaloa by Armstrong andJones (1971) and notes on the rabbits of Jalisco byGenoways and Jones (1973).
Presently, four species of rabbits are known fromwesteentral Mexico: Mexican cottontail, Sylvilaguscunicularius (Waterhouse, 1848); eastern cottontail,S. floridanus (J. A. Allen, 1890); Tres Marias cotton-tail, 5. graysoni (J. A. Allen, 1877); and desert cot-tontail, S. audubonii (Baird, 1857). West-centralMexico is that area including Sinaloa and Durangosouthward through Nayarit, Zacatecas, Aguascalien-tes, Jalisco, Guanajuato, Colima, and Michoacan.
Recent collecting in Nayarit by personnel of theNational Fish and Wildlife Laboratory, and on theTres Marias Islands (approximately 86 km west ofmainland Nayarit), has yielded specimens repre-senting three of the four species known from west-central Mexico. This material, combined with thatavailable in other collections, makes it possible tomore clearly define the distribution and relation-ships of members of the genus Sylvilagus in west-central Mexico.
Victor E. Diersing, Museum of Natural History, Universityof Illinois, Urbana, Illinois, 61801. Don E. Wilson, U. S. Fishand Wildlife Service, National Fish and Wildlife Laboratory,National Museum of Natural History, Washington, D. C.20560.
Many individuals of S. floridanus and S. cuni-cularius from mainland Nayarit and adjacent areas,could not be assigned to either species using pub-lished methods. Also, the rabbits from San JuanitoIsland differ markedly from those on adjacentMaria Madre Island. This report presents distribu-tional data and methods for recognizing all speciesof Sylvilagus throughout west-central Mexico. Theinsular 5. graysoni is compared to the mainlandspecies.
ACKNOWLEDGMENTS.—The research on which thisreport is based was assisted by a Smithsonian Insti-tution summer fellowship awarded to Diersing in1975. In addition, funds for Diersing's travel toexamine rabbits housed in collections throughoutthe United States and Canada were made availableby the Theodore Roosevelt Fund of the AmericanMuseum of Natural History. Our appreciation isextended to those people who gave their permissionto examine specimens under their care. Thesepeople and their institutions (abbreviations usedin listing specimens examined follow the name ofeach collection) are: Sidney Anderson, AmericanMuseum of Natural History (AMNH), New York;Richard D. Estes, Philadelphia Academy of NaturalSciences (ANSP); Robert T. Orr, California Aca-demy of Sciences (CAS), Golden Gate Park, SanFrancisco; David G. Huckaby, Biology Department,California State University (CSLB), Long Beach;Luis de la Torre, Field Museum of Natural History(FMNH), Chicago; Robert S. Hoffmann, Museum
1

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
of Natural History (KU), University of Kansas,Lawrence; Donald R. Patten, Los Angeles CountyMuseum (LACM), Los Angeles; James S. Findley,Museum of Southwestern Biology (MSB), Universityof New Mexico, Albuquerque; Rollin H. Baker,The Museum, Michigan State University (MSU),East Lansing; William Z. Lidicker, Jr., Museumof Vertebrate Zoology (MVZ), University of Cali-fornia, Berkeley; J. Mary Taylor, Vertebrate Muse-um, University of British Columbia (UBC), Van-couver; William H. Burt, University of ColoradoMuseum (UCM), Boulder; Donald F. Hoffmeister,Museum of Natural History, University of Illinois(UIMNH), Urbana; Emmet T. Hooper, Museumof Zoology, University of Michigan (UMMZ), AnnArbor; National Museum of Natural History,Smithsonian Institution (USNM, collection of theformer United States National Museum).
Our studies in Mexico were facilitated by permitsand other assistance provided by the DireccionGeneral de la Fauna Silvestre, and especially by theformer Director, Mario Luis Gabucio. Field andmuseum assistance was provided by B. A. Bacon,C. A. Blount, R. D. Fisher, K. N. Geluso, P. Huerta,C. B. Robbins, and N. J. Scott, Jr.; A. L. Gardnerprovided laboratory assistance with the karyotypicanalysis; M. A. Bogan, A. L. Gardner, C. Jones, andR. W. Thorington, Jr. helpfully criticized earlierversions of this report.
Methods
When available, total length, tail length, hindfoot length, and ear length from the notch weretaken from the original skin tag. Body length wascalculated by subtracting tail length from the totallength. Dry ear length was measured from the notchto the tip of the fleshy ear. All 26 quantitative cra-nial measurements were taken to the nearest 0.5 mmusing dial calipers. The cranial measurements andmethods for taking them are: upper incisor length,I1 (first upper incisor) length measured along itsgroove; palatal length, least length; greatest skulllength, anterior face of I1 to the posterior point ofthe supraoccipital shield; basal length, ventro-median notch of the foramen magnum to theanterior face of I1; zygomatic breadth, greatestbreadth across both zygomatic arches; braincasebreadth, greatest breadth usually taken underneaththe free projection of each zygomatic arch; nasal
length, greatest length of a nasal bone; greatest nasalbreadth, greatest breadth across both nasals; maxil-lary toothrow, alveolar length of the toothrow,PM2-M3; maxillary toothrow breadth, taken acrossboth toothrows at the labial alveolus of PM3;postdental breadth, least breadth across the ptery-goid processes posterior to the maxillary toothrows;incisive foramen, greatest length of an incisive fora-men; basioccipital length, midventral length of thebasioccipital; basioccipital breadth, greatest breadthbetween the auditory bullae; diastema length, pos-terior alveolar border of I2 to the anterior alveolarborder of PM2; rostrum depth, taken from the mostventral point of the rostrum (the point where thesuture is formed by the premaxillary and maxillary)dorsally in a line perpendicular to the length of theventral surface of the rostrum to the most dorsalsurfaces of the nasals; bullar length, greatest antero-posterior length of the bulla; bullae breadth, dis-tance between the most lateral margins of bothbullae; shield-bullae depth, depth of the skull fromthe postero-dorsal surface of the supraoccipitalshield to a plain across the antero-ventral surfacesof both auditory bullae; skull depth, greatest heightof the skull above a glass slide positioned suchthat the upper incisors and mastoid processes restupon the slide; carotid foramina breadth, distancebetween the two foramina; infraorbital canalsbreadth, distance between the canals taken acrossthe ridge of bone forming the lateral border ofeach canal; mandible height, most anteroventralextension of the angular process to the most dorsalpart of the coronoid process; mandible length, an-terior point of the alveolus of PM3 to the mostposterior extension of the angular process; ramusheight, taken according to Findley, et al. (1975:86),from the bottom of the most anterior labial notchof the alveolus of PM4 ventrally to the most ventralextent of the ramus; mandibular toothrow, an-terior alveolar border of PM3 to the posterioralveolar of M3. Methods for taking these measure-ments are further explained in Diersing (1978).The color of the dorsal and tail pelages were re-corded as grayish or reddish.
Statistical procedures include discriminant func-tion analysis, principal components analysis, Stu-dent's t-test of means, scattergrams, means, stand-ard deviations, and coefficients of variation. Allcomputer techniques were employed on the Uni-versity of Illinois IBM 360 computer.

NUMBER 297
All specimens were aged by the method of Hoff-meister and Zimmerman (1967). Only adults wereused in statistical analyses. Because females aver-aged only slightly larger than males, both sexeswere combined in statistical treatments.
Recognition of Species
Four species of rabbits are known from west-central Mexico, S. floridanus, S. cunicularius, S.audubonii, and S. graysoni. The Omilteme rabbit,S. insonus, and forest rabbit, S. brasiliensis, areknown from areas to the south in Guerrero andOaxaca, respectively. In the interior Mexican Pla-teau of west-central Mexico, the geographic rangeof S. floridanus partially overlaps that of S. audu-bonii. Along the lower western foothills of theSierra Madre Occidental and the Transverse Vol-canic Axis, the ranges of S. floridanus and S. cuni-cularius are parapatric. In most of these areas thesespecies are distinguished from one another withdifficulty, and some specimens have been misidenti-fied. Sylvilagus graysoni is known only from theTres Marias Islands of Nayarit. Sylvilagus audu-bonii also occurs in the arid sections of north-western Sinaloa, where it is parapatric with S. flori-danus of the adjacent Sierra Madre. The bestmethods for distinguishing these species are pre-sented below. Refer to Table A for additionalquantitative data. Geographic variation withineach species is discussed under "Taxonomy."
Sylvilagus floridanus from 5. cunicularius: Insouthern Sinaloa southward to western Michoacan,S. floridanus occurs on the western forested slopesof the Sierra Madre Occidental and western marginsof the Transverse Volcanic Axis. Immediately westof the mountains, S. cunicularius occurs on thetropical coastal plain from the sea to mountainslopes. These two species are parapatric and aredifficult to distinguish where the tropical coastalplain meets the western mountain slopes. Fromsouthern Sinaloa southward to western Michoacan,each species increases in overall size, which com-plicates recognition of the two species.
In Sinoloa, Nayarit, western Jalisco, Colima, andwestern Michoacan, 5. cunicularius has long earsand a short tail with short reddish-colored hairsdorsally. Sylvilagus floridanus has medium-sizedears and medium-sized tail with long reddish toreddish black hairs dorsally. In general, the breadth
of the tail of S. cunicularius (including the hairs)is less than 35 mm and the ears (as measured dryfrom the notch) are usually longer than 61 mm,whereas in 5. floridanus the breadth of the tail isusually greater than 40 mm and the ears are usu-ally shorter than 61 mm. The two species canusually be separated on the basis of these twoexternal differences.
Cranially, S. cunicularius differs from S. flori-danus primarily in having a larger skull (especiallyin greatest length) with a much deeper mandible,larger auditory bullae, longer maxillary and man-dibular toothrows, and greater breadth across thecarotid foramina. A scattergram analysis (Figure 1)plotting mandibular depth and bullar lengthagainst alveolar lengths of the mandibular andmaxillary toothrows, separates the two species. In-dividuals of grouped samples 2 (Sinaloa specimensonly), 3, and 4 of S. floridanus, and samples I, 2,and 3 of S. cunicularius were used. Grouped sam-ples within the figure correspond to those of Fig-
fHR
OW
* TO
O1
' (m
m)
_j °
»OTH
»i
OF
MAF
LAR
Y TC
AR
LE
NG
THS
PLU
S M
AX
ILLV
EO
L
35.0
34.0
33.0
32.0
31.0
300
29.0
28.0
27.0
-
-
4 > " -
M A
\ >
2 f \
j
"I7]j
/ \ \A
• i i i
20.0 21.0 22.0 23.0 24.0 25.0 260 Z7.0
MANDIBULAR RAMUS DEPTH PLUS BULLA LENGTH (mm)
FIGURE 1.—Scattergram analysis comparing individuals ofS. cunicularius with S. floridanus from extreme western Mex-ico (encircled area = morphological dispersion of pooled sam-ple; number associated with each sample corresponds tonumber used in Figures 3 and 4; subscripts "c" or "t" byeach number refer to cunicularius or floridanus; sample 2fis composed only of specimens from Sinaloa; within eachspecies, southern samples are morphologically larger thannorthern samples).
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
9.0
8.0
I 7.0
S 6.03
5.0-1
S. cunicularius
-S. floridanus
5. graysoni
10.0 1.0 12.0 13.0 14.0
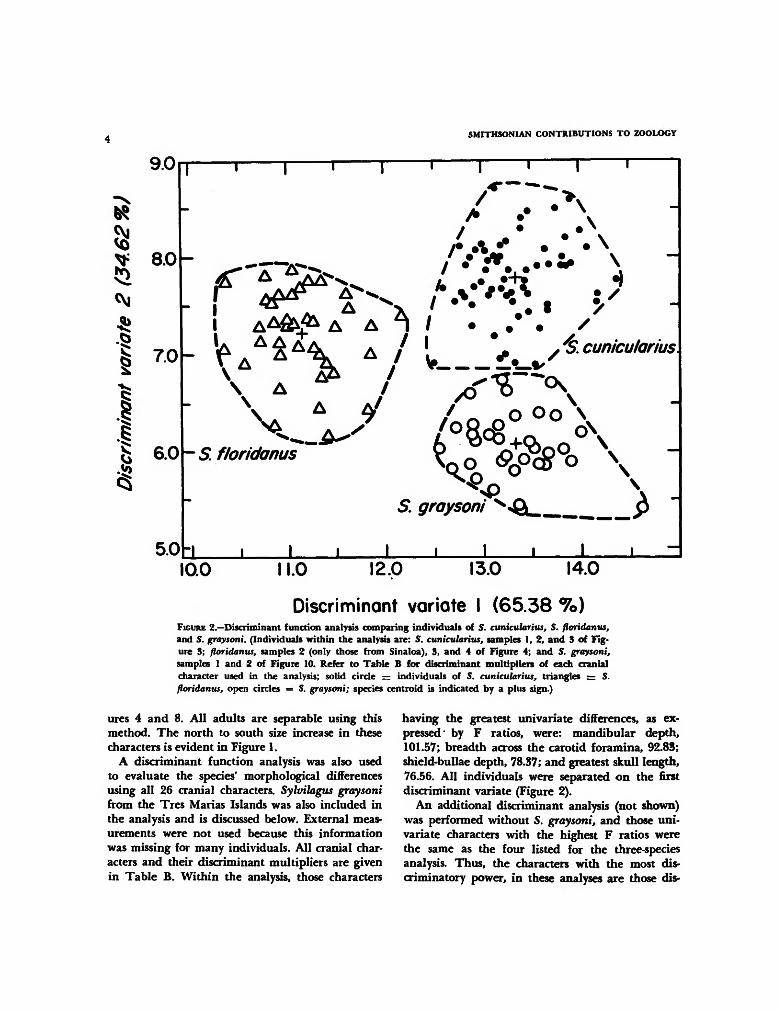
Discriminant variate I (65.38 7o)FIGURE 2.—Discriminant function analysis comparing individuals of 5. cunicularius, S. floridanus,and S. graysoni. (Individuals within the analysis are: S. cunicularius, samples 1, 2, and 3 of Fig-ure 3; floridanus, samples 2 (only those from Sinaloa), 3, and 4 of Figure 4; and 5. graysoni,samples 1 and 2 of Figure 10. Refer to Table B for discriminant multipliers of each cranialcharacter used in the analysis; solid circle = individuals of S. cunicularius, triangles — S.floridanus, open circles = S. graysoni; species centroid is indicated by a plus sign.)
ures 4 and 8. All adults are separable using thismethod. The north to south size increase in thesecharacters is evident in Figure 1.
A discriminant function analysis was also usedto evaluate the species' morphological differencesusing all 26 cranial characters. Sylvilagus graysonifrom the Tres Marias Islands was also included inthe analysis and is discussed below. External meas-urements were not used because this informationwas missing for many individuals. All cranial char-acters and their discriminant multipliers are givenin Table B. Within the analysis, those characters
having the greatest univariate differences, as ex-pressed by F ratios, were: mandibular depth,101.57; breadth across the carotid foramina, 92.83;shield-bullae depth, 78.37; and greatest skull length,76.56. All individuals were separated on the firstdiscriminant variate (Figure 2).
An additional discriminant analysis (not shown)was performed without 5. graysoni, and those uni-variate characters with the highest F ratios werethe same as the four listed for the three-speciesanalysis. Thus, the characters with the most dis-criminatory power, in these analyses are those dis-

NUMBER 297
tinguishing S. floridanus from the other two species.Armstrong and Jones (1971) considered 5. flori-
danus and S. cunicularius to be allopatric in Sina-loa except at San Ignacio, 215 m, where two speci-mens of S. cunicularius and one of S. floridanuswere recorded. The two specimens referred to S.cunicularius (KU 90781 and 90782) were includedin our discriminant function analysis as S. flori-danus} and have individual discriminant scores onthe first axis of 11.65 and 11.40, typical for 5. flori-danus rather than S. cunicularius. Examination ofthese specimens confirmed their identity.
Allen (1906:212) referred a specimen (AMNH24694) from Sinaloa, Arroyo de Taquaco, 1220 m,to S. cunicularius. This is an unusually high ele-vation for the species in Sinaloa and Nayarit. Allother records from these two states are from eleva-tions under 90 m. The recorded elevation of thisspecimen (individual discriminant score of 13.52on the first discriminant variate) is probably in-correct. The town "Escuinapa", written on theskull box, has an elevation of approximately 30 m.
In interior Mexico, S. floridanus is smaller than
it is in the west and S. cunicularius is larger.Therefore, the difference in overall size betweenthe two species is much greater. For example, inthe northern half of Michoacan and southern Ja-lisco (samples 8 and 10 of Figure 4), the largestindividual of S. floridanus measured 75.85 mm and66.7 mm in greatest skull and dry ear length, re-spectively. The smallest specimen of S. cunicularius(sample 4 of Figure 3) from the same general areameasured 82.30 mm and 70.0 mm. There is nooverlap in these characters and the two species areeasily distinguished from one another.
FIGURE 3.—Geographic distribution of 5. cunicularius in West-Central Mexico (circles «= individual localities of 5. cuni-cularius insolitus; triangles = S. cunicularius cunicularius;number enclosed by dashes — pooled sample used in allstatistical comparisons; see Table A) .
FIGURE 4.—Geographic distribution of S. floridanus in West-Central Mexico (triangles = individual localities of 5.floridanus holzneri; solid circles — S. f. macrocopus; opencircles -» S. F. orizabae; number enclosed by dashes — pooledsample used in statistical comparisons in Table A; distribu-tion of S. floridanus holzneri in extreme northeastern Sinaloais not shown).
Sylvilagus graysoni from S. floridanus: Exter-nally, these species differ little. Cranially, S. gray-soni differs in being slightly larger with a markedlydeeper mandible, longer palate, greater breadthacross the maxillary toothrows, and greater breadthacross the carotid foramina.
A scattergram analysis (not shown), plottingdepth of the mandible against palatal length, sepa-

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
rated 61 of 63 individuals of the two species. Syl-vilagus floridanus from samples 2 (Sinaloa speci-mens only), 3, and 4 were used in the scattergram.In the discriminant function analysis, all individ-uals of the two species are separable on the firstdiscriminant function (Figure 2). There is addi-tional separation on the second discriminant func-tion.
Sylvilagus graysoni from S. cunicularius: Exter-nally, S. graysoni differs in having markedly shorterears, which range in length (dry) from 50.8 to 60.4mm; whereas ear lengths of S. cunicularius (sam-ples 1, 2, and 3) range from 59.8 to 74.4 mm. Asingle specimen of S. cunicularius is responsiblefor the overlap. Cranially, S. graysoni differs pri-marily in having much shorter and narrower na-sals. The discriminant function analysis, (Figure2; Table B) correctly allocated all individuals ofthe two species.
Sylvilagus floridanus and S. audubonii: On theMexican Plateau, S. floridanus always has a well-pigmented rufous tail, whereas in S. audubonii thetail has a narrow middorsal gray streak borderedby white. Externally and cranially, S. floridanusis larger, especially in greatest skull length, nasallength, palate length, maxillary toothrow (no over-lap), and mandibular toothrow. Sylvilagus audu-bonii has relatively longer ears and larger auditorybullae. Detailed measurements are given inTable A.
Sylvilagus floridanus of the Sierra Madre of Sina-loa and adjacent Durango differs from S. audu-bonii from northwestern Sinaloa in the same waysthat the two species differ in interior Mexico,except that S. audubonii is slightly larger, withsmaller auditory bullae. However, the grayish tailof S. audubonii and the rufous tail of S. floridanusreadily distinguishes the two species.
Evolution of Sylvilagus graysoni
The Tres Marias Islands are continental islandsseparated from mainland Nayarit by water depthsof up to approximately 365 m. During maximumPleistocene glaciation when ocean levels may havebeen lowered by as much as 120 m (Zweifel, 1960),these islands may either have been connected tothe mainland or separated by a narrow strait ofwater. The presence of fresh water fish (Nelson,1899) on the islands suggests a land connection.
Rabbits most likely colonized the islands duringthis period.
Morphologically, S. graysoni is more like S. cuni-cularius than S. floridanus. Sylvilagus graysoni andS. cunicularius both differ from S. floridanus inhaving a markedly deeper mandible, greater breadthacross the carotid foramina, and larger skulls.These differences from S. floridanus were evidentin the discriminant function analysis (Figure 2)where 5. floridanus was separated from S. graysoniand S. cunicularius on the first discriminant func-tion.
Biogeographical evidence supports the origin ofS. graysoni from S. cunicularius-like stock. Sylvi-lagus cunicularius occurs along the coastal plainof mainland Mexico whereas S. floridanus occupiesthe higher foothills of the Sierra Madre. The dis-persal of S. cunicularius-like rabbits from coastalMexico to the Tres Marias Islands is, therefore,much more likely than is the dispersal of S. flori-danus-like stock to the islands. Nelson (1909:238)maintained that S. graysoni "was probably derivedfrom S. c. insolitus of the adjacent mainland" butpresented no definite supportive evidence.
When and how the progenitors of S. graysoniarrived on the islands is unknown. However, basedon the comparisons of the morphology of S. gray-soni and mainland S. cunicularius it is evidentthat individuals of S. graysoni from all four islandsshare several morphological properties (chieflyshort and narrow nasals and short ears) indicatinggene exchange between the island populationsafter isolation from the mainland stock. Gene ex-change may have been accomplished by any of thefollowing methods: (1) passive dispersal betweenislands by man, or rafting; (2) active dispersal be-tween islands by swimming; or (3) dispersal whenthe four islands comprised a larger single island.Subsequently, the rabbits on San Juanito Island,the northernmost of the four islands, have second-arily differentiated (see species account, page 15.)
The karyotype (Figure 5) of S. graysoni may bedescribed as follows: 2n=42, FN=78, 15 pairs ofmedium-sized to small meta- and submetacentrics,and 5 pairs of small acrocentric or subtelocentricautosomes. Some of these latter 5 pairs may bearsatellites on the short arms as noted for S. flori-danus by Hsu and Benirschke (1971). The X chro-mosome is a small acrocentric or subtelocentricand the Y is a very small acrocentric. The sex

NUMBER 297
XX
44* A
XIXX
IAA4
MX X I
A * * •
X Y
MM
• A * A
FIGURE 5.—Karyotype of Sylwlagus graysoni.
chromosomes are morphologically distinct fromthose figured for S. floridanus by Hsu andBenirschke (1971).
Unfortunately, 5. cunicularius has not beenkaryotyped, precluding comparison with that spe-cies. We would predict that S. cunicularius will befound to share the diploid number of 42 withS. graysoni and S. floridanus, and the morphologyof the sex chromosomes or banding patterns of theautosomes may be useful in further elucidating rela-tionships among S. graysoni, S. cunicularius, andS. floridanus.
In addition to the morphological distinctness ofS. graysoni, it is relatively unafraid of man. Thislack of escape behavior was particularly apparentin S. graysoni from San Juanito Island, althoughrabbits from the other islands showed far less fearof man than do their mainland counterparts. Be-cause of the above reasons, S. graysoni seems bestconsidered a distinct species.
Sylvilagus cunicularius insolitus (J. A. Allen)
L-pus insolitus J. A. Allen, 1890:189.Sylvilagus cunicularius insolitus.—Nelson, 1909:243.
TYPE.—AMNH 2655/2133, adult, male, skin andskull, from the plains of Colima, Mexico. Holo-type examined.
RANGE.—Coastal plains of the southern third ofSinaloa, as well as Nayarit, Jalisco, and Colima.Marginal records are: SINALOA: El Limon; Ro-sario; 9.7 km W La Concha. NAYARIT: Aca-poneta; Platanares, 16.1 km E. Ruiz; 29 km E SanBias; 3.2 km SW San Juan de Abajo. JALISCO:Ixtepa; Chamala Bay. COLIMA: Colima; Armeria.
DIAGNOSIS.—Externally: size small for the spe-cies; small tail and short ears. Skull: small in mostmeasurements, especially in length of upper in-cisor, greatest skull length, breadth of the brain-case, and mandibular height. Color: body reddishdorsally with black hairs intermixed, tail brightreddish dorsally.
COMPARISONS.—Sylvilagus c. insolitus differs fromS. c. cunicularius from south-central Jalisco andMichoacan as follows: externally smaller, especiallyin ear and tail length; cranially smaller, particu-arly in greatest skull length and braincase breadth;and color of body and tail reddish dorsally ratherthan grayish. All individuals of the two subspeciescan be easily distinguished using these characters(see Table A).

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
REMARKS.—Genoways and Jones (1973:4) referredspecimens from Jilotlan de los Dolores and 12.8km E Jilotlan de los Dolores in south-centralJalisco to S. c. insolitus. These specimens insteadrepresent S. c. cunicularius as shown by large size,long grayish tail (KU 111660), and broad brain-cases.
There is some geographic variation within S. c.insolitus. Specimens from the northern part of therange are slightly smaller in overall dimensionsthan are individuals from the southern parts oithe range in Colima.
SPECIMENS EXAMINED (74).—COLIMA: Armeria, sea levelto 60 m, 6 (USNM); Colima, 1 (USNM); plains of Colima, 1(AMNH); Hacienda Magdalena, 455 m, 2 (USNM); Manza-nillo, 15 m, 1 (USNM).
JALISCO: Chamala Bay, 4 (UMMZ); Ixtepa, 90 in, »(USNM).
NAYARIT: Acaponeta, 60 m, 5 (USNM); Chacala, 1(USNM); Paso de Soquilpa, 14.2 km E San Bias, 1 (USNM);Platanares, 16.1 km E Ruiz, 2 (KU); San Bias, 30 m, 1(USNM); 8.1 km E, 6.4 km S San Bias, 1 (MSB); 20.8 km(by road) E San Bias, 1 (KU); 29 km E San Bias, 1 (CSLB);3.2 km SW San Juan de Abajo, 2 (USNM); Santiago, 61 m,4 (USNM); 2.3 km (by road) Tacote, 3 (USNM).
SINALOA: Arroyo de Taquaco, 1220 m (probably incor-rect; see page 5), 1 (AMNH); 32.2 km SE Autlan, Isla Palmitode la Virgin, 1 (UMMZ); 9.7 km W La Concha, 3 m, 3 (KU);Escuinapa, 13 (12 AMNH, 1 UCM); El Limon, 1 (LACM);Mazatlan, 7 (6 FMNH, 1 CAS); near Mazatlan, 30 to 90 m,2 (USNM); Palmito, 6 m, 2 (KU); Rosario, 30 m, 2 (USNM);9.7 km NNW Teacapan. 2 (KU).
Sylvilagus cunicularius cunicularius (Waterhouse)
Lepus cunicularius cunicularius Waterhouse, 1848:132.Sylvilagus cunicularius.—Nelson, 1909:239.Lepus verae-crucis Thomas, 1890:74. [Type from Las Vigas,
Veracruz, Mexico. Holotype not examined.]
RANGE.—Mountains of the Transverse VolcanicAxis from southern Jalisco eastward to Veracruz,Sierra Madre del Sur of Guerrero, and SistemaMontanoso of northern Oaxaca. Marginal recordsin western Mexico are: JALISCO: Jilotlan de losDolores, 730 m. MICHOACAN: Cerro Tancitaro;Patzcuaro.
DIAGNOSIS.—Externally: large in all features.Skull: large in all characters, particularly greatestskull length, braincase breadth, and mandibularheight. Color: body grayish dorsally, tail with anarrow middorsal gray streak bordered by muchwhite.
COMPARISONS.—For comparisons of S. c. cunicu-larius with S. c. insolitus, see the latter subspecies.
REMARKS.—Although not presented here, com-parisons of S. c. cunicularius from Jalisco andMichoacan with samples to the east across theMexican Plateau reveal little differentiation. Inaddition, specimens from coastal Guerrero differfrom both S. c. insolitus and 5. c. cunicularius andare referable to S. c. pacificus.
SPECIMENS EXAMINED (15) .—JALISCO: Jilotlan de losDolores, 730 m, 1 (KU); 12.9 km E Jilotlan de los Dolores.610 m, 2 (KU).
MICHOACAN: Cerro Tancitaro, vicinity Apo, 1 (UMMZ);Patzcuaro, 2130 m, 10 (USNM); Tancitaro, 1 (FMNH).
Sylvilagus floridanus macrocorpus, new subspecies
TYPE.—USNM 509011, adult, female, skin andskull, from Estanzuela, 1372 m, Nayarit, Mexico,collected 20 March 1975 by Clyde Jones, originalno. 5251.
RANGE.—Mountains of extreme southwesternMichoacan, mountains of southwestern Jalisco, andNayarit. Marginal records are: NAYARIT: Mesadel Nayar, 1372 m; Estanzuela; Estancia (possiblyacross the border in Jalisco). JALISCO: La Cie-nega; Volcan de Fuego, 2985 m; Las Canoas.MICHOACAN: 22.6 km W Aquililla, Dos Aguas,2130 m. JALISCO: 32.2 km SE Autlan, 2740 m;24.2 km S, 14.5 km E Talpa de Allende, 2105 m;La Laguna, Sierra de Juanacatlan, 1980 m. NAYA-RIT: 3.2 km E San Pedro Lagunillas, E sidelake; Tepic.
DIAGOSIS.—Externally: large body with medium-sized ears. Skull: large in all measured features ex-cept with medium-sized auditory bullae, but espe-cially large in length of the maxillary toothrow,length of the mandibular toothrow, postdentalbreadth, rostral depth, and breadth across the max-illary toothrows. Color: varying geographically, inMichoacan and extreme southwestern Jalisco allspecimens are reddish dorsally with numerousblackish hairs intermixed, grayish and ochraceouson the body sides, and whitish ventrally with adusky neck patch. In Nayarit and bordering Jaliscosome specimens are grayer dorsally than reddishand all have grayish body sides.
COMPARISONS.—Sylvilagus f. macrocorpus differsfrom S. /. orizabae primarily in larger size, particu-larly in greatest skull length, length of the maxil-lary toothrow, postdental breadth, and breadthacross the maxillary toothrows. In addition, S. f.

NUMBER 297
macrocorpus has relatively shorter ears. Both sub-species are polychromic and color is not a reliablecharacter for distinguishing between them.
The larger size of S. f. macrocorpus relative toS. f. orizabae is shown in Figure 6. Individuals ofS. /. orizabae from centra] and eastern Jalisco andnorth-central Michoacan (samples 6, 7, and 8 ofFigure 3) are almost completely separable fromindividuals of S. /. macrocorpus from westernMichoacan, western Jalisco, and Nayarit (samples3 and 4 of Figure 3). The open triangles (sample5 of Figure 3) represent the holotype, topotypes,and near topotypes of S. f. restrictus (now referredto S. /. orizabae). These specimens cluster moreclosely with S. f. orizabae.
Discriminant function analysis also affirms thedesignation of S. f. restrictus as a synonym of S. /.orizabae. For the analysis, the reference sample ofS. f. macrocorpus consisted of samples 3 and 4 fromNayarit, western Jalisco, and northwestern Mich-
oacan. The reference sample of S. f. orizabae con-sisted of samples 7 and 8 from northern Michoa-can, east-central Jalisco, and Aguascalientes. Thetest sample (sample 5) consisted of the holotype,topotypes, and near topotypes of 5. /. restrictusfrom central Jalisco. The 26 cranial measurementsused and their discriminant multipliers are givenin Table C. The individual discriminant scores forthe reference samples and the test sample are givenin Figure 7. All but one of the individuals in thetest sample are referable to S. /. orizabae.
Sylvilagus f. macrocorpus differs from S. f. holz-neri in larger size except in bullar and ear length.Sylvilagus f. macrocorpus is considerably larger ingreatest skull length, depth of the rostrum, andin the lengths of the maxillary and mandibulartoothrows (Table C). Typically, S. f. macrocorpusis reddish dorsally and S. f. holzneri is grayish dor-sally. A scattergram analysis (Figure 8) illustratesthe larger size of S. /. macrocorpus.
15.8
15.4
15.0
14.6
14.2
13.8
13.4
13.0
_
-
_ ///
hi ,
orizabae
A
A A
1 1 1 1 1 1 1 1
//
* //
//
f
1
/ ° ° o"°~~ - ^o o -Q
O O 00 O /
'̂Ctf00 oo /
macrocorpus
i • i . i . i . i . i . i
184 18.8 19.2 19.6 20.0 20.4 208 21.2 21.6 22.0 22.4 22.8 23.2
Breadth across nhaxillary toothrows (mm)FIGURE 6.—Scattergram used to compare S. f. orizabae and S. /. macrocorpus (circles z= Sylvilagusf. macrocorpus (Figure 4, samples 3, 4); solid triangles — S. /. orizabae (Figure 4, samples 7, 8);open triangles = the type, topotypes, and near topotypes of S. f. restrictus (Figure 4, sample 5)herein considered a synonym of S. f. orizabae; see also "Comparisons," page 8).

10 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
-0. _E_0_
orizabae - reference\
macrocorpus - reference
P, np9.6 10.0 10.4 10.8 1.2 12.0 12.4
Individual discriminant scoresFIGURE 7.—Discriminant function analysis of S. /. macrocorpus (Figure 4, samples 3, 4) with5. /. orizabae (samples 7, 8). (Sample 5 of S. /. orizabae, top graph, was used as a test sample;square containing the symbol "R" — the type of S. f. restrictus.)
II1
|
I
16.1
15.7
153
14.9
14.5
14.1
13.7
13.3
_
-
-
-
-
-
-l
//
z
/o
y
/
/
/
o
)
1
oo
o °o°o
i
/
//
/ • •
v i ^T^
O ^
holzneri
1 L 1 1
macrocorpus
•
//
1 « ' i i
19.0 19.4 19.8 20.2 20.6 21.0 21.4 21.8 22.2 22.6 23.0 23.4
Breadth across maxillary toothrows (mm)
FIGURE 8.—Scattergram analysis of S. f. macrocorpus (Figure 4, samples 3, 4) and 5. /. holzneri(sample 2). (Solid circles = S. /. macrocorpus; open circles — 5. /. holzneri.)
NUMBER 297 11
j
5.0
3.0
1.0
-1.0
-3.0
holzneri
.'' V
- orizabae ^~-
-~ (9mocrocorpus
-6.0 -4.0 -2.0 0 2.0 4.0 6.0 8.0 10.0Principal component I ( 7 8 . 5 % )
FIGURE 9.—Principal components analysis of S. /. macrocorpus, S. f. holzneri, and S. f. orizabaebased upon 26 cranial variables, dry ear length, total length, and hind foot length (samplenumbers correspond to those of Figure 4; sample "A" of S. f. holzneri, not pictured in Figure 4.is composed of specimens from central Chihuahua, southern Sonora, and northern Sinaloa;see also "Comparisons," page 8).
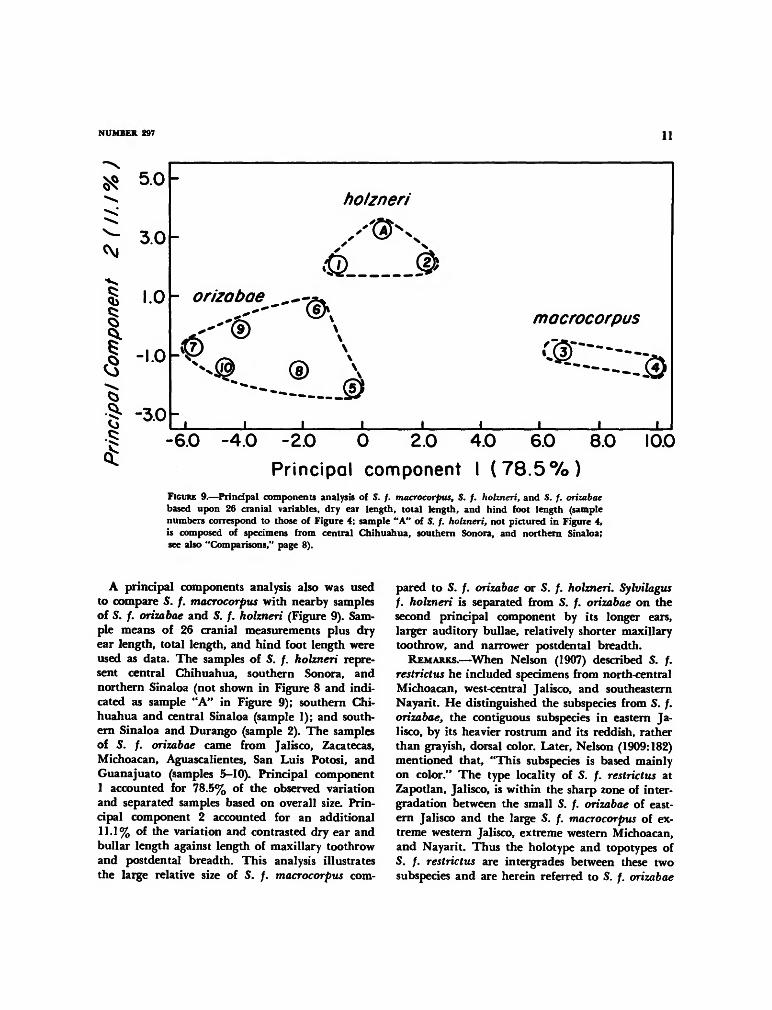
A principal components analysis also was usedto compare S. f. macrocorpus with nearby samplesof S. /. orizabae and S. /. holzneri (Figure 9). Sam-ple means of 26 cranial measurements plus dryear length, total length, and hind foot length wereused as data. The samples of S. /. holzneri repre-sent central Chihuahua, southern Sonora, andnorthern Sinaloa (not shown in Figure 8 and indi-cated as sample "A" in Figure 9); southern Chi-huahua and central Sinaloa (sample 1); and south-ern Sinaloa and Durango (sample 2). The samplesof S. f. orizabae came from Jalisco, Zacatecas,Michoacan, Aguascalientes, San Luis Potosi, andGuanajuato (samples 5-10). Principal component1 accounted for 78.5% of the observed variationand separated samples based on overall size. Prin-cipal component 2 accounted for an additional11.1% of the variation and contrasted dry ear andbullar length against length of maxillary toothrowand postdental breadth. This analysis illustratesthe large relative size of S. /. macrocorpus com-
pared to S. f. orizabae or S. /. holzneri. Sylvilagusf. holzneri is separated from S. f. orizabae on thesecond principal component by its longer ears,larger auditory bullae, relatively shorter maxillarytoothrow, and narrower postdental breadth.
REMARKS.—When Nelson (1907) described S. /.restrictus he included specimens from north-centralMichoacan, west-central Jalisco, and southeasternNayarit. He distinguished the subspecies from S. /.orizabae, the contiguous subspecies in eastern Ja-lisco, by its heavier rostrum and its reddish, ratherthan grayish, dorsal color. Later, Nelson (1909:182)mentioned that, "This subspecies is based mainlyon color." The type locality of S. /. restrictus atZapotlan, Jalisco, is within the sharp zone of inter-gradation between the small S. /. orizabae of east-ern Jalisco and the large S. /. macrocorpus of ex-treme western Jalisco, extreme western Michoacan,and Nayarit. Thus the holotype and topotypes ofS. f. restrictus are intergrades between these twosubspecies and are herein referred to S. /. orizabae

12 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
because of their small size, especially in alveolarlength of the maxillary toothrow, postdentalbreadth, breadth across the maxillary toothrows,greatest skull length, and alveolar length of themandibular toothrow. Reddish and grayish speci-mens are represented among the topotypes of S. /.restrictus from Zapotlan, Jalisco.
Nelson had no specimens from extreme westernMichoacan, and only a few individuals from north-central Michoacan, extreme western Jalisco, andNayarit. Specimens are now available from thisarea where S. f. macrocorpus is known to occur.Nelson (1909) had one adult specimen from LaLaguna, Sierra de Juanacatlan, 1980 m, Jaliscowhich he referred to S. f. restrictus. He prescientlynoted (1909:183) that it "differs so much in skullcharacters and in general appearance from bothrestrictus and orizabae that I hesitate to place itwith either." Nelson also considered a skull onlyfrom Tepic, Nayarit, as representing typical S. f.restrictus. This subadult specimen is much largerthan other specimens Nelson referred to 5. /. re-strictus. After Nelson had written most of his 1909account of 5. /. restrictus, he received specimensof rabbits from several localities in western Jaliscofrom J. A. Allen. Nelson also identified these mor-phologically much larger animals as typical S. f.restrictus.
The type of 5. /. restrictus was taken from Zapot-lan, Jalisco. Two additional specimens, labelled"Sierra Nevada de Colima," have "Zapotlan" writ-ten in pencil on the skull boxes. In addition, oneis recorded from an elevation of "about 4500 ft"(1372 m) and the other from "about 5000 ft" (1525m). Zapotlan (today = Ciudad Guzman) is at thebase of the Sierra Nevada de Colima. These twoapparent topotypes are slightly smaller than thetype, and all three rabbits probably came from afew miles southwest of Zapotlan towards the slopeof the Sierra Nevada de Colima (Field notes, E. W.Nelson, 1892). All three are intergrades between5. /. macrocorpus and S. f. orizabae, but best re-ferred to 5. /. orizabae.
Three rabbits from Las Canoas, Jalisco (AMNH26148-50) average larger in all dimensions thando the three specimens from near Zapotlan. Al-though intermediate in many respects, they arebest referred to S. f. macrocorpus. The exact loca-tion of Las Canoas is unknown, however, Geno-ways (1973) located it 10 km NW Tuxpan and
10 km SSW Ciudad Guzman (=Zapotlan). Thus,the specimens taken near Zapotlan (type and topo-types of S. f. restrictus) may have come from alower elevation and from only a few miles farthereast than the specimens herein referred to S. f.macrocorpus from Las Canoas.
In Michoacan and Jalisco S. f. macrocorpus isknown only from the higher pine-oak forests,where specimens have been recorded from 1980 to2985 m. However, Nayarit S. f. macrocorpus oftenoccur at lower elevations, the lowest at Estanzuela,1372 m. The specimen from Mesa del Nayar showsmorphological intergradation with S. /. holzneri,but is here referred to S. f. macrocorpus. Two fromsouthern Sinaloa, one taken near San Ignacio, 215m, and the other near Palmito, overlap with speci-mens of S. /. macrocorpus in Figure 8. These tworabbits are best referred to S. f. holzneri.
The eastern limit of S. /. macrocorpus in Nay-arit, Jalisco, and Michoacan, corresponds closelyto the eastern limit of the Pacific Region, as de-fined by Rzedowski and McVaugh (1966).
ETYMOLOGY.—The Latin macrocorpus ("largebody") refers to the large size of this subspeciesrelative to the smaller contiguous subspecies.
SPECIMENS EXAMINED (32).—JALISCO: Atenguillo (possiblylocated across the border in Nayarit), 1 (USNM); 32.2 kmSE Autlan, 2740 m, 3 (KU); La Cienega, 2 (AMNH); LaLaguna, Sierra de Juanacatlan, 1980 m, 1 (USNM); LasCanoas, 3 (AMNH); 245 km S, 14.5 km E Talpa de Allende,2100 m, 4 (KU); 19.3 km S Toliman, 2345 m, 8 (KU); Volcande Fuego, 2985 m, 1 (KU).
MICHOACAN: 22.4 km W Aquililla, Dos Aguas, 215 m, 1(UMMZ).
NAYARIT: Estancia (possibly located across the border inJalisco), 2 (AMNH); Estanzuela, 2 (USNM); Llano y Casco(possibly located across the border in Jalisco), 1 (AMNH);Mesa Del Nayar, 1370 m, 1 (USNM); 3.2 km E San PedroLagunillas, E side lake. 1 (USNM); Tepic, 1 (USNM).
ADDITIONAL RECORDS.—Nelson (1909:183) listedspecimens probably referable to this subspeciesfrom the border of western Jalisco and southeast-ern Nayarit. These localities are Garabatos, RioAmeca, and Ojo de Agua (near Amatlan). We haveexamined these specimens and found them to besubadults that we are reluctant to allocate to sub-species, except on geographic grounds.
Sylvilagus floridanus holzneri (Mearns)
Lepus sylvaticus holzneri Mearns, 1896:554.

NUMBER 297
Sylvilagus (SyMlagus) floridanus holzneri.—Lyon, 1904:356.[Lepus sylvaticus] subspecies rigidus Mearns, 1896:555. [Type
from Carrizalillo, Luna Co., New Mexico. Holotype ex-amined.]
Lepus (Sylvilagus) durangae J. A. Allen, 1903:609. [Type fromRancho Bailon, northwestern Durango, Mexico. Holotypeexamined.]
TYPE.—USNM 58937, adult, female, skin andskull, from the Douglas spruce (=Douglas fir) zonenear the summit of the Huachuca Mountains,Cochise County, Arizona. Holotype examined.
RANGE.—Mountains of the southwestern UnitedStates southward along the Sierra Madre Occiden-tal of Sonora, Chihuahua, Sinaloa, and Durango.Marginal records in Durango and Sinaloa are:DURANGO: Inde, 1830 m, 6.4 km NW La Pila;45.1 km S, 27.4 km W Vicente Guerrero, 2545 m.SINALOA: Plomosas, 22 km E Matatan, 760 m;San Ignacio, 215 m; 2.4 km N Badiraguato, 230m; 16 km NNE Choix, 520 m.
DIAGNOSIS.—Externally: medium-sized to largewith long ears. Skull: medium-sized to large in allfeatures except small in rostrum depth, short andnarrow maxillary and mandibular toothrows, andshallow mandible, but with large auditory bullae.Color: grayish dorsally, pale ochraceous buff later-ally, pale gray rump patch, and creamy whiteventrally except for a dusky neck patch.
COMPARISONS.—For comparisons with S. /. macro-corpus see that subspecies. Sylvilagus f. holzneridiffers from S. f. orizabae: externally, larger sizeand longer ear; cranially, larger skull particularlyevident in greatest skull length and bullar length;and color, pale grayish dorsally rather than darkgrayish.
REMARKS.—Nelson (1909) and Baker and Greer(1962) assigned specimens from Zacatecas to S. f.holzneri. Admittedly, intergradation between S. f.holzneri and S. /. orizabae is gradual in Durangoand Zacatecas, and the assignment of intergradesto either subspecies is subjective. However, speci-mens from Zacatecas (sample 6) are significantlysmaller in greatest skull length (P<.01) and havesmaller auditory bullae than have specimens fromDurango and Sinaloa (sample 2). The small skulland small auditory bullae of Zacatecan specimensis characteristic of S. /. orizabae, the subspecies towhich they are here referred.
SPECIMENS EXAMINED (68).—CHIHUAHUA: 9.7 km N Cero-cahui, 1890 m, 1 (KU); 17.7 km S, 12.9 km W Creel, 245 m.
1 (KU); 25.8 km S, 9.7 km F. Creel, 2225 m, 1 (KU): 48.3 kmSW Gallego, Arroyo del Nido, 2135 m, 1 (MVZ); 64.4 km OEGallego, 1524 m, 1 (ANSP); 5.6 km ESE Los Lamentos, 1(KU); 3.2 Km W Minaca, 2100 m, 1 (KU); Mojarachic, 2(USNM); 6.4 km NW San Francisco de Baja, 1735 m, 1 (KU);25 km NE Santa Clara, W side Sierra del Nido, 2485 m,1 (MVZ); Sierra del Nido, Arroyo Mesteno, 2315 m. 1 (MVZ);Sierra del Nido, Arroyo Mesteno, 2375 m, 1 (MVZ); Sierradel Nido, Arroyo Mesteno, 2440 m, 1 (MVZ); Sierra del Nido,Canon del Alamo, 2225 m, 3 (MVZ); Sierra Madre, nearCuadalupe de Calvo, 2135 m, 1 (USNM); Sierra Tarahumare,"Samachique", 2 (FMNH); 4.8 km NE Temoris. 1705 m.1 (KU).
DURANGO: 12 km E Cosala, Santa Ana, 395 m, 2 (KU);Coyotes, 1 (FMNH); 38.6 km SSE Durango, 2120 m, 1 (MSU);El Salto, 2315-2560 m, 3 (USNM); 8.1 km NE El Salto, Ha-cienda Coyotes, 1 (CAS); 9.7 km SW El Salto, 2530-2545 m,2 (MSU); Guanacevi, 2 (AMNH); mtns, near Guanacevi,2375 m, 2 (USNM); Inde, 1830 m, 1 (USNM); 6.4 km NWLa Pila, 2 (MSU); 2.4 km S Laguna del Progresso, 1(AMNH); 8.1 km W Las Adjuntas, 3050 m, 1 (KU); 11.3 kmSW Las Adjuntas, 2715 m, 1 (KU); Rancho Bailon, 2(AMNH); 9-7 km NW Rodeo, Rio Nazaz, 1280 m, 1 (KU);Sierra Madre, 2740 m (Cerro Prieto), 1 (USNM); 45.1 km S,27.4 km W Vicente Guerrero, 2545 m, 2 (MSU).
SINALOA: 2.4 km N Badiraguato, 230 m, 2 (KU); 20.9 kmESE Badiraguato, 245 m, 1 (KU); 20 Km N, 5 km E Badira-guato, 550 m, 2 (KU); 1.6 km S El Cajon, 550 m, 2 (KU);16 km NNE Choix, 520 m, 2 (KU); 5 km SW Palmito. 1860m, 5 (KU); Plomosas, 22 km E Matatan, 760 m, 1 (KU);San Ignacio, 215 m, 2 (Klft; 10 km S, 38 km E Sinaloa, 245 m,2 (KU); 15 km N, 65 km E Sinaloa, 1435 m, 1 (KU).
SONORA: 11.3 km WNW Alamos, 1 (UIMNH); 12.9 kmWNW Alamos, 1 (UIMNH).
Sylvilagus floridanus orizabae (Merriam)
Lepus orizabae Merriam, 1893:143.Sylvilagus floridanus orizabae.—Nelson, 1909:183.Lepus floridanus subcinctus Miller, 1899:386. [Type from
Hacienda El Molino, near Negrete, Michoacan, Mexico.Holotype examined.]
Lepus floridanus persultator Elliot, 1903:147. [Type fromPuebla. Puebla, Mexico. Holotype examined.]
Sylvilagus floridanus restrictus Nelson, 1907:82. [Type fromZapotlan, Jalisco, Mexico. Holotype examined.]
TYPE.—USNM 53318, adult, female, skin andskull, from Mount Orizaba, Puebla, Mexico. Holo-type examined.
RANGE.—Sierra Madre Occidental of the north-ern panhandle of Jalisco and adjacent southwest-ern Zacatecas; higher mountains of the CentralMexican Plateau from Aguascalientes, eastern Ja-lisco, adjacent portions of southwestern San LuisPotosi, and northwestern Guanajuato; TransverseVolcanic Axis from central Jalisco eastwardthroughout the northern half of Michoacan, ex-

14 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
treme southern Guanajuato, Mexico, Distrito Fe-deral, Morelos, Tlaxcala, central third of Puebla,and extreme west-central Veracruz; Sierra MadreOriental of extreme southeastern Coahuila, south-ern third of Nuevo Leon, extreme southwesternTamaulipas, central third of San Luis Potosi,Queretaro, and Hidalgo. Marginal records in west-central Mexico: MICHOACAN: Quiroga, Tacam-baro Hwy km 54, 2835 m; San Juan, Cerro Tanci-taro, 2300 m. JALISCO: Sierra Nevada de Colima(Zapotlan), 1370-1525 m; 1.6 km SSE Ameca, 1220m; Etzatlan, 1065 m; 3.2 km E Bolanos, 1080 m.ZACATECAS: Hacienda San Juan Capistrano;Valparaiso Mountains; 6.4 km N Villanueva, 2010m. AGUASCALIENTES: 4.8 km N Cerro delJaguez, 2500 m, Sierra Fria. SAN LUIS POTOSI:Cerro Penon Blanco, 2360 m.
DIAGNOSIS.—Externally: small to medium-sizedwith long ears. Skull: small to medium-sized ingreatest length, zygomatic breadth, length of themaxillary and mandibular toothrows, mandibularramus depth, and rostrum depth; however, withlarge auditory bullae. Color: variable, usually darkgrayish dorsally but reddish gray dorsally in someareas of its range, ochraceous laterally, blackishgray rump patch, and whitish ventrally with adusky neck patch.
COMPARISONS.—For comparisons of S. f. orizabaewith S. /. macrocorpus and 5. /. holzneri, see ac-counts of those subspecies.
REMARKS.—Sylvilagus f. orizabae varies in colorgeographically. Typically the subspecies is grayishdorsally. However, in northwestern Michoacan,many of the specimens are reddish gray, and insome cases (Patzcuaro and Los Reyes in Michoa-can), are more reddish than grayish. Likewise, incentral Jalisco many individuals intergrading withthe reddish S. f. macrocorpus are also more reddishthan grayish. All specimens within samples 7 and 9are grayish dorsally.
Topotypical and near topotypical S. /. orizabaefrom Puebla, Tlaxcala, Mexico, and the DistritoFederal were compared to more western samplespreviously referred to S. /. orizabae, S. f. restrictus,and S. f. subcinctus. Excluding color, the only sig-nificant feature noted among these samples was aclinal east to west increase in overall size across theMexican Plateau and Transverse Volcanic Axis.The largest individuals are within sample 5 and
these intergrade with the much larger S. /. macro-corpus.
Since geographic variation within all of thesesamples is largely clinal and color has already beenshown to be variable, even within a population,S. /. subcinctus and S. /. restrictus are both regardedas synonyms of S. /. orizabae.
SPECIMENS EXAMINED (81).—AGUASCALIENTES: 4.8 km NCerro del Jaguez, 2500 m, Sierra Fria, 1 (MVZ); 12.9 kmWNW Colonia Presidente Calles, 2285 m, 1 (MVZ); Vena-verros, 21 km W Aquascalientes, 2 (MVZ).
JALISCO: 4.8 km N Amatitan, 1235 m, 1 (KU); 1.6 kmSSE Ameca, 1220 m, 1 (KU); 3.2 km E Bolanos, 1080 m,1 (KU); 9.7 km ENE Bolanos, 1630 m, 2 (KU); Top of CerroViejo de Curjuflam, 2955 m, 30.6 km S, 14.5 km W Guadalajara, 1 (KU); 72 km NE Comanja dc Corona, 2440 m, 2 (KU);Etzatlan, 1065 m, 9 (USNM); 6.4 km W, 3.2 km S Guadala-jara, 1555 m, 1 (KU); 25.8 km S Guadalajara, 1 (KU); 8.1 kmNE Huejuquilla, 1890 m, 1 (KU); 1.6 km S Jalostoiitlan,1735 m, 1 (KU); La Barca, 1 (USNM); La Mesa Maria deLeon, 2255 m, 2 (KU); Lagos, 1875 m, 2 (USNM); 22.5 km SELagos de Moreno, 2040 m, 1 (KU); 16.1 km NW Matanzas,2440 m, 2 (KU); Ocotlan, 5 (2 FMNH. 1 USNM); SierraNevada de Colima, (Zapotlan) 1370-1525 m, 3 (USNM); 4 kmE Tuxcueca, S side Lago de Chapala, 1525 m, 1 (KU); 6.4 kmW Villa Guerrero, 1675 m, 2 (KU); 3.2 km S, 14.5 km WYahualica, 2135 m, 1 (KU); 1.6 km SW Zapotlanejo, 1 (KU);3.2 km E Zapotlanejo, 1 (KU).
MICHOACAN: Barranca Seca, 1 (UMMZ); Hacienda ElMolino, near Negrete, 2 (USNM); 17.7 km W Jiquilpan, 2040m, 1 (KU); 3.2 km E Los Reyes, 2 (UMMZ); Mt. Tancitaro,2745-P660 m, 1 (USNM); Nuevo San Juan (Los Conejos), 8.1km SW Uruapan, 1 (UMMZ); Patzcuaro, 2135 m, 9 (USNM);9 km S Patzcuaro, on Hwy to Tacambaro, 2440 m, 1 (UMMZ);Puerto Murillos, 59.6 km E Morelia, 1 (MVZ); Quiroga,Tacambaro Hwy, km 54, 2835 m, 1 (UMMZ); 6 km NWQuiroga, Lake Patzcuaro, 2745 m, 1 (MVZ); San Juan, CerroTancitaro, 2300 m, 1 (UMMZ); 7.2 km NE Tarecuato, 1(KU); 4.8 km N Volcan Paricutin, San Juan, 7200 ft, 1(UMMZ).
ZACATECAS: Hacienda San Juan Capistrano, 2 (USNM);Monte Escobedo, 2285 m, 1 (KU); Plateado, 1 (USNM);Valparaiso Mtns, 1 (USNM); 6.4 km N Villanueva, 2010 m,1 (CAS).
Sylvilagus graysoni graysoni (J. A. Allen)
Lepus graysoni J. A. Allen, 1877:347.Sylvilagus (Sylvilagus) graysoni.—Lyon, 1904:336.
TYPE.—USNM 8318, adult, female, skin withskull inside, "undoubtedly from Maria Madre,"according to Nelson (1899:16), Tres Marias Islands,Nayarit, Mexico. Holotype examined.
RANGE.—The southern three islands of the Tres

NUMBER 297 15
22"
21*JO'
If
107*
1
-
107*
/c\ 1
0
0
\\
5
S 10
106*30'
1
n,,o
N
Xf l ?
IS 20 K
1IO6"JO'
Modrt
\
wMill!
lomtltr
PACIFICOCEAN
106'
1
o
Cltotoi
1106*
22"
lo'
21*
FIGURE 10.—Geographic distribution of 5. graysoni on theTres Marias Islands and San Juanito Island (individuals weregrouped into two pooled samples and compared in Figure 11).
Marias (Maria Madre, Maria Magdalena, andMaria Cleofas), Nayarit, Mexico (Figure 10).
DIAGNOSIS.—Externally: Medium-sized to largeexcept with short ears. Skull: medium-sized tolarge with long rostrum (reflected in long diastemaand incisive foramina), short maxillary toothrow,and narrow basioccipital. Color: reddish dorsallywith the nape and rump brightest, body sides apaler reddish, venter whitish except for the brown-ish throat patch.
COMPARISONS.—For comparisons with S. graysonifrom San Juanito Island, see below.
REMARKS.—Only two adults are available fromMaria Magdalena and one from Maria Cleofas. Allthree compare with topotypical specimens fromMaria Madre and all are referable to 5. g. graysoni.Specimens from Maria Magdalena are the darkestin color.
Short-term morphological evolution of insularfaunas is often assumed. To test the possibilityof rapid change in the Tres Marias rabbits, 15specimens collected from Maria Madre in 1897were compared to five collected in 1960 and 1976(2 from Maria Madre, 2 from Maria Magdalena,and 1 from Maria Cleofas). There were no signifi-
cant differences between the two samples (univari-ate Student's Mests, P>0.05).
SPECIMENS EXAMINED (28).—NAYARIT: Tres Marias Islands:Maria Madre, 22 (20 USNM, 2 CAS); Maria Magdalena, 4(S USNM, 1 UBC); Maria Cleoras, 1 (USNM); and un-identified island, 1 (FMNH).
Sylvilagus graysoni badistes, new subspecies
TYPE.—USNM 512550, adult, male, skin andskull, from San Juanito Island of the Tres MariasIslands, Nayarit, Mexico, collected 24 March 1976by Don E. Wilson, original no. 3730.
RANGE.—Occurs only on San Juanito Island,Nayarit, Mexico (Figure 10).
DIAGNOSIS.—Externally: medium-sized exceptwith short ears. Skull: medium-sized with shortrostrum (reflected in short diastema and short in-cisive foramina), long maxillary toothrows, andwide basioccipital. Color: variable, from reddishto brownish dorsally, pale reddish to brownish onthe sides; venter whitish except for a brown throatpatch.
COMPARISONS.—Sylvilagus g. badistes differs sig-nificantly from S. g. graysoni in having a smallerskull as measured by greatest skull length (P<0.05),wider basioccipital (P<0.01), longer maxillarytoothrow (P<0.001), shorter incisive foramina(P<0.001), and shorter diastema (P<0.001).
A scattergram analysis (Figure 11) of maxillarytoothrow length against length of the incisive fora-men, separates all individuals of the two subspe-cies. Table 3 details the morphological differencesbetween S. g. badistes and S. g. graysoni.
ETYMOLOGY.—The name badistes, from theGreek stem badio ("to advance slowly, step bystep") refers to the unusual lack of escape behaviorin these rabbits.
SPECIMENS EXAMINED (11).—NAYARIT: TresMarias Islands, San Juanito Island, 11 (USNM).
Sylvilagus audubonii parvulus (J. A. Allen)
Lepus (Sylvilagus) parvulus J. A. Allen 1904:34.Sylvilagus audubonii parvulus.—Nelson, 1909:236.
TYPE.—AMNH 12578/10888, adult, sex un-known, skin and skull, from Apam, Hidalgo, Mex-ico, approximately 2440 m. Holotype examined.
RANGE.—Arid deserts of the Mexican Plateau.Marginal records in west-central Mexico are:

16 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
55
1
16.8
16.4
16.0
15.6
15.2
14.8
badistes-
o /" p ° ° ,''
\ s
\ •\ /
\ /
i
A
o••
•
•1
%
= San Juanito= Maria Madre= Maria Magdalena= Maria Cleofas= unknown island
\
•
\^*~* graysoni
i i i
17.7 18.1 18.5 18.9 19.3 19.7 20.1 20.5 20.9 21.3
Incisive foramen length (mm)FIGURE 11.—Scattergram analysis comparing 5. g. badistes from San Juanito Island (Figure 10,
sample 1) with S. g. graysoni from the Tres Marias Islands (sample 2).
DURANGO: 6.4 km NNE Boquilla, 1920 m;Durango City. ZACATECAS: 4.8 km SW Som-brerete, 2345 m. JALISCO: 4.8 km S Huejucar,1800 m; 4.8 km S Yahualica, 1800 m; 4.8 km SWTeatitlan. GUANAJUATO: 4.8 km E Tepezala,2345 m. QUERETARO: Caderata [= Cadereyta],2100 m.
DIAGNOSIS.—Externally: size small except withlarge ears. Skull: small size especially evident ingreatest skull length, nasal length, and palatelength, however, with large auditory bullae andmedium depth mandibular ramus. Color: dorsumgray and tail gray middorsally broadly marginedby white.
COMPARISONS.—Sylvilagus a. parvulus differsfrom Sinaloan S. a. goldmani (sample 3 of Figure12) in smaller size overall, particularly in palatelength, nasal length, and breadth across the carotidforamina. Sylvilagus a. parvulus does have mark-
edly larger auditory bullae.These characters distinguish all individuals of
the two subspecies. Refer to Table 1 for quantita-tive data.
REMARKS.—The two samples (1 and 2 of Figure12) of S. a. parvulus differ significantly only inlength of the auditory bullae (P<0.05). Topotypi-cal and near topotypical specimens of S. a. minorfrom near El Paso, Texas were compared withS. a. parvulus and found to average smaller in sizeand paler gray in color. Nevertheless, differencesbetween samples from Texas and Durango areslight.
SPECIMENS EXAMINED (44).—AGUASCALIENTES: Chicha-lote, 3 (USNM); 4.8 km E Tepezala, 2345 m, 1 (MVZ).
DURANGO: 2 km E Atotonilco, 2035 m, 2 (MSU); 3.1 kmSE Atotonilco, 2035 m, 1 (MSU); 4.8 km SE Atotonilco, 2035m, 1 (MSU); 15 km NW Atotonilco, 1980 m, 1 (MSU); 6.4km NNE Boquilla, 1920 m, 1 (MSU); 9.7 km N Chocolate,

NUMBER 297 17
FIGURE 12—Geographic distribution of S. audubonii (cir-cles - individual localities of 5. a. parvulus; triangles = S. a.goldmani; each pooled sample was treated statistically inTable A).
1250 m, 1 (MSU); Durango City, 2 (USNM); 12 km NEDurango, 1860 m, 1 (MVZ); 6.4 km NW La Pila, 1 (MSU);11.3 km NNW La Zarca, 185 m, 2 (MSU); 56 km W Mapimi,1675 m, 1 (MSU); San Juan. 19.3 km W Lerdo, 1160 m,
2 (UMMZ); San Juan, 20 km W Ciudad Lerdo, 1160 m,1 (MVZ).
GUANAJUATO: Silao, 1 (USNM).JALISCO: 12.9 km W Encarnacion de Diaz, 1 (KU); 4.8 km
S Huejucar, 1800 m, 3 (KU); Lagos de Moreno, 1 (USNM);4.8 km SW Tepatitlan, 4.8 km S Yahualica, 1800 m, 1 (KU);8.1 km S, 1.6 km E Yahualica, 1 (KU); 16.1 km NE Yahualica,1 (KU).
QUERETARO: Caderata [=Cadereyta], 2100 m, 1 (UMMZ).SAN LUIS POTOSI: 1.6 km W Hacienda la Parada, 32.2
km NW San Luis Potosi, 1 (MVZ); 2 km E Illescas, 2(LSUMZ); Leoncito, 1 (LSUMZ); 7 km NW Palma, 2380 m,1 (LSUMZ); Salinas, 1 (LSUMZ); 3 km S Santo Domingo,1 (LSUMZ).
ZACATECAS: Berriozabal, 2 (USNM); Canitas, 1 (USNM);32.2 KM S Concepcion del Oro, 1830 m, 1 (CAS); 35.4 kmSW Concepcion del Oro, 1 (CAS); 4.8 km SW Sombrerete,2345 m, 1 (CAS).
Sylvilagus audubonii goldmani (Nelson)
Lepus arizonae goldmani Nelson, 1904:107.Sylvilagus audubonii goldmani.—Nelson, 1909:225.
TYPE.—USNM 96812, adult, female, skin andskull, from Culiacan, Sinaloa, Mexico. Holotypeexamined.
RANGE.—Arid coastal plains of the northern halfof Sinaloa. Marginal records: SINALOA: 19.3 kmSW El Fuerte; Sinaloa; 1.6 km S Pericos; 3.2 kmE Aguacaliente, 245 m; 9.7 km N El Dorado; Islade Tachichilte, 6 km E Isla de Altamura; 17.7 kmN Ahome, 45 m.
DIAGNOSIS.—Externally: medium-sized. Skull: me-dium-sized in greatest skull length, nasal length,maxillary toothrow length, and zygomatic breadth,but with small auditory bullae and narrow mandi-ble. Color: dorsum gray and tail gray middorsallyand narrowly margined by white.
COMPARISONS.—For comparisons with S. a. parvu-lus see that subspecies.
REMARKS.—In Sinaloa the ranges of S. auduboniiand S. floridanus are complementary. The range ofS. audubonii may also be complementary with 5.cunicularius, although specimens of either are notavailable from the coastal region of central Sinaloa.
SPECIMENS EXAMINED (30).—SINALOA: 3.2 km E Agua-caliente, 245 m, 2 (KU); 17.7 km N Ahome, 45 m, 1 (MVZ);Bacubirito. 1 (USNM>; Culiacan, 2 (USNM); 9.7 km NWCuliacan, 1 (MVZ); 19.3 km N Culiacan, 2 (KU); 12.9 kmWNW El Carrizo, 3 m, 1 (KU); 9.7 km N El Dorado, 1 (KU);9.7 km N, 2.4 km E El Dorado, 3 (KU); 11.3 km NE ElFuerte, 2 (KU); 19.3 km SW El Fuerte, 1 (UIMNH); Isla deTachichilte, 6 km E Isla de Altamura, 7 (KU); 4 km SWNavolato, 6 m, 2 (KU); S side Rio Sinaloa, 1.6 km SW Sinaloa,35 m, 2 (MVZ); Sinaloa, 1 (USNM); 16 km SE Topolobampo,6 m. 1 (KU).
Summary
The geographic distribution of populations ofSylvilagus in west-central Mexico may be summar-ized as follows: The northern part of the coastalplain in Sinaloa is inhabited by S. audubonii gold-mani. This area is characterized by arid climate andthorn scrub vegetation. The coastal plain fromsouthern Sinaloa through Nayarit, Jalisco, andColima is more humid and contains subtropicaldeciduous forest. This region is inhabited by S.cunicularius insolitus. Sylvilagus cunicularius cuni-cularius extends eastward across the TransverseVolcanic Axis from montane Jalisco into easternMexico. The southern extension of the SierraMadre Occidental in eastern Nayarit, Jalisco, andwestern Michoacan is inhabited by S. floridanus

18 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
macrocorpus. These animals are primarly restrictedto the pine-oak woodlands of intermediate to higherelevations on the western flanks of the Sierra MadreOccidental. They intergrade in the north with S. /.holzneri, the form inhabiting the Sierra MadreOccidental northwards into the United States. Tothe east, they intergrade with s. f. orizabae, whichextends westward from the Mexican Plateau alongthe eastern edges of the Sierra Madre Occidental.The more arid northern parts of the MexicanPlateau are inhabited by S. audubonii parvulus.The insular species, Sylvilagus graysoni, has twosubspecies, the nominate form of the three majorislands of the Tres Marias, and S. g. badistes onSan Juanito Island.
Reasons for the subspeciation of Sylvilagus gray-soni on the Tres Marias Islands are not readilyapparent. However, the taxonomic conclusions(endemic species with insular subspecies) supportthe hypothesis that these rabbits are not adept atcrossing water barriers. Such an hypothesis is inline with our view that the species S. graysonishared a common mainland ancestor with S. cuni-cularius, and was separated from it at a time whenthe Islands were closer to, or attached to the main-
land. More recently, the Islands were probablycloser together or possibly united. It is possible thatthe rabbits on San Juanito were separated from thenearest populations on Maria Madre at a timewhen rising sea levels created the situation we seetoday. Although the water gap between the twoislands is narrow (3 kms), there is a strong currentbetween the islands which makes passage by boathazardous. Perhaps this current adds to the effectof distance to preclude interchange between thepopulations.
Finally, it should be noted that most of thegeographic variation found in these rabbits is dueto size. Since many, if not all, of the mensuralcharacters are correlated with size, this is not sur-prizing. However, some populations do show allo-metric relationships that result in the separationseen on the second discriminant function of thoseanalyses. The most notable general pattern is thenorth-to-south size increase demonstrated by almostall taxa. In some forms, especially S. floridanus, asimilar trend occurs from east-to-west. The ecologi-cal reasons for such size differences are beyond thescope of this study, but should provide a fruitfulfield of inquiry for future research.
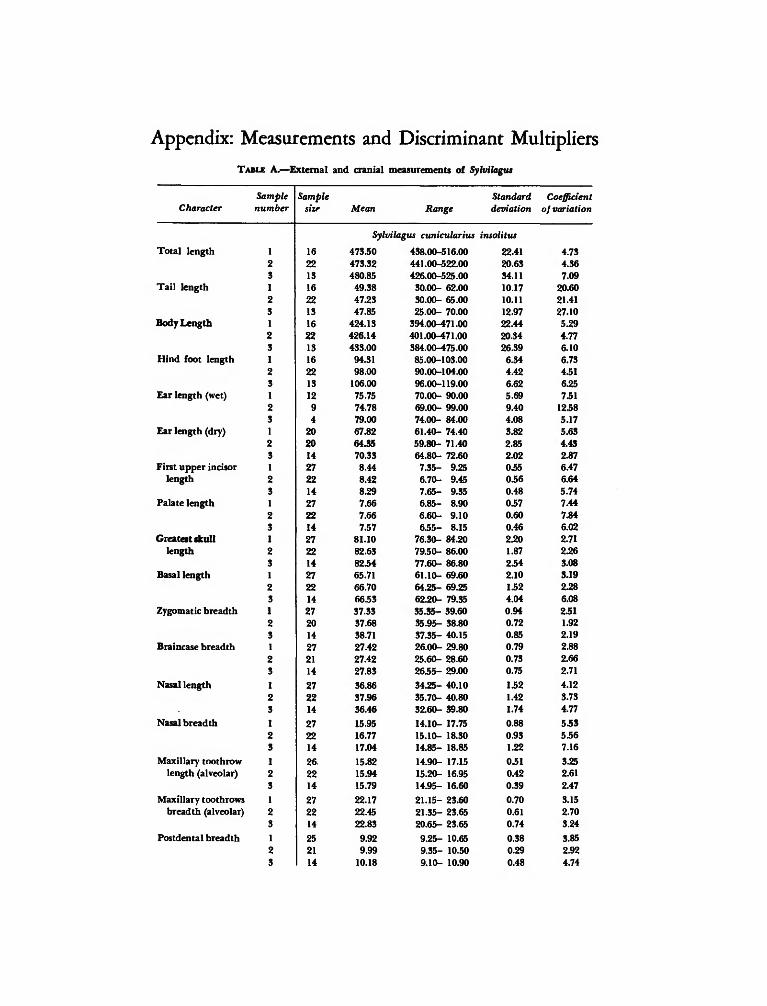
Appendix: Measurements and Discriminant Multipliers
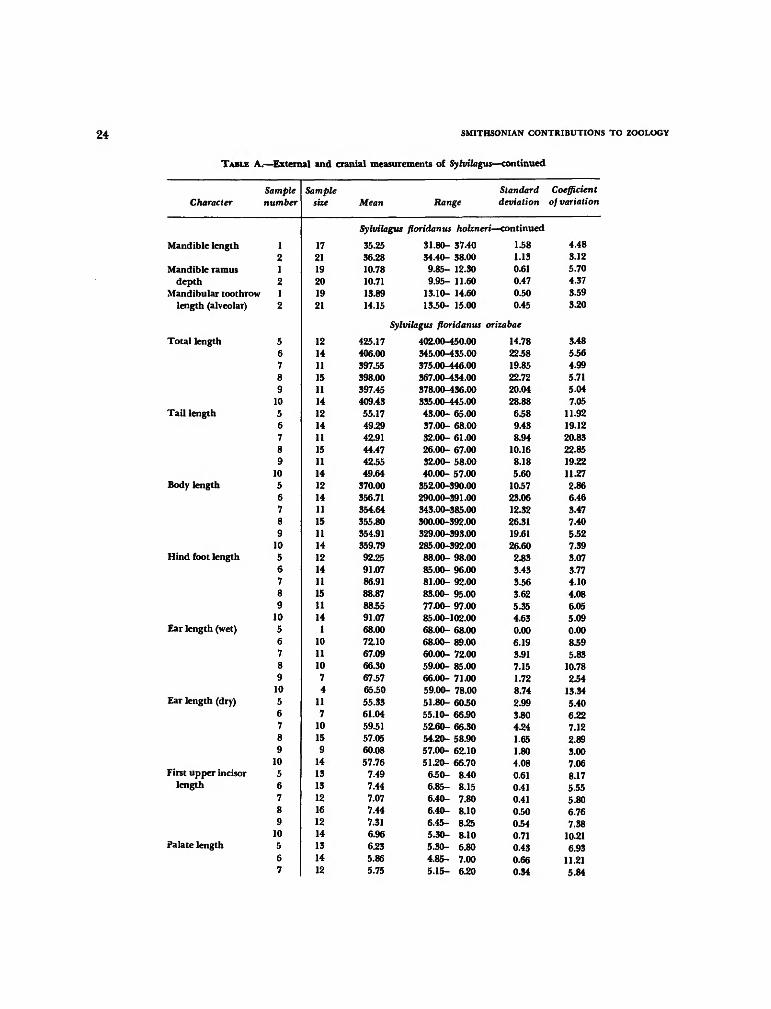
TABLE A.—External and cranial measurements of Sylvilagus
Character
Total length
Tail length
Body Length
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Greatest skulllength
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
Samplenumber
12312312312312312312312S123123123123
123
123
123
123
123
Samplesize
1622131622131622131622131294
202014272214272214272214272214272014272114
272214
272214262214
272214
252114
Mean Range
Sylvilagus cunicularius
47350473.32480.85
49.3847.2347.85
424.13426.14433.00
94.3198.00
106.0075.7574.7879.0067.8264.3570.338.448.428.297.667.667.57
81.1082.63825465.7166.7066.5337.3337.6838.7127.4227.4227.83
36.8637.9636.46
15.9516.7717.04
15.8215.9415.79
22.1722.4522.83
9.929.99
10.18
438.00-516.00441.00-522.00426.00-525.00
30.00- 62.0030.00- 65.0025.00- 70.00
394.00-471.00401.00-471.00384.00-475.0085.00-103.0090.00-104.0096.00-119.0070.00- 90.0069.00- 99.0074.00- 84.0061.40- 74.4059.80- 71.4064.80- 72.60
7.35- 9.256.70- 9.457.65- 9.356.85- 8.906.60- 9.106.55- 8.15
76.30- 84.2079.50- 86.0077.60- 86.8061.10- 69.6064.25- 69.2562.20- 79.3535.35- 39.6035.95- 38.8037.35- 40.1526.00- 29.8025.60- 28.602655- 29.00
34.25- 40.1035.70- 40.8032.60- 39.80
14.10- 17.7515.10- 18.3014.85- 18.85
14.90- 17.1515.20- 16.9514.95- 16.60
21.15- 23.6021.35- 23.6520.65- 23.65
9.25- 10.659.35- 10.509.10- 10.90
Standarddeviation
insolitus
22.4120.6334.1110.1710.1112.9722.4420.3426.39
6.344.426.625.699.404.083.822.852.020550560.480570.600.462.201.872542.101.524.040.940.720.850.790.730.75
1521.421.74
0.880.931.22
0510.420.39
0.700.610.74
0.380.290.48
Coefficientof variation
4.734.367.09
20.6021.4127.105.294.776.106.734.516.25751
12585.175.634.432.876.476.645.747.447.846.022.712.263.083.192.286.082511.922.192.882.662.71
4.123.734.77
5535567.16
3.252.612.47
3.152.703.24
3.852.924.74

20 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character i
Incisive foramenlength
Basioccipital length
Basioccipital breadth
Diastema length
Rostrum depth
Bulla length
Bullae breadth
Shield-bullae depth
Skull depth
Carotid foraminabreadth
Infraorbital canalsbreadth
Mandible height
Mandible length
Samplenumber
12312312312312312312312312S123123123123
Mandible ramus depth 1
Mandibular toothrowlength (alveolar)
Total lengthTail lengthBody lengthHind foot lengthEar length (wet)Ear length (dry)
23123
444444
Samplesize
272214272214272214272214272214272214272114272214272214272214262214261914242113272214272214
131313134
12
Mean
Sylvilagus
19.9320.2320.7010.5910.569.85
10.0610.0310.0522.0322.5022.4716.9917.0616.6411.8512.14115328.0528.0027.6123.8924.3223.6734.0334.4234.2512.3912.2820.7819.9819.6920.7839.4839.9339.9640.9741.55405112.4912.6412.6816.0916.2916.38
RangeStandarddeviation
Coefficientof variation
cunicularius insolitus—continued
17.60- 21.8018.80- 21.6520.10- 22.208.95- 11.609.50- 11.359.10- 10.559.25- 10.859.30- 10.659.25- 11.35
19.75- 23.9520.75- 24.0521.15- 24.3015.75- 18.2015.85- 18.3515.50- 17.8011.25- 12.6011.35- 12.7510.90- 12.1026.90- 29.7025.90- 28.9026.00- 28.7021.90- 25.0022.85- 25.6021.60- 24.6531.90- 35.5031.95- 36.0033.00- 36.1511.00- 14.3011.45- 13.0018.75- 22.2018.20- 21.4018.20- 21.0518.75- 22.2037.15- 42.9038.40- 42.8537.35- 42.1038.20- 43.4539.80- 42.8038.30- 41.9011.60- 13.7011.40- 13.6511.15- 13.8015.00- 17.6515.70- 17.0015.30- 17.00
0.920.880.550.560.530.390.420.390.541.100,900.830.740.700.630.340.420.360.820.840.860.620.800.941.011.130.930.770.470.830.890.780.831.331.201.251.200.941.000.480.550.620560.330 5 0
Sylvilagus cunicularius cunicularius
509.6962.69
447.00109.0081.2575.29
490.00-525.0028.00- 78.00
422.00-467.0010200-118.0078.00- 83.0070.00- 78.80
12.4312.6813.984.182.362.79
4.614.332.685.324.993.944.153.855.345.014.003.714.344.113.782.913.473.092.933.003.122.603593.952.963.282.716.223.824.014.473.954.013.383.003.122.922.272.473.834.354.913502.033.03
2.4420.14
3.133.842.913.71

NUMBER 297 21
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
First upper incisorlength
Palate lengthGreatest skull lengthBasal lengthZygomatic breadthBraincase breadthNasal lengthNasal breadthMaxillary toothrow
length (alveolar)Maxillary toothrows
breadth (alveolar)Postdental breadthIncisive foramen
lengthBasioccipital lengthBasioccipital breadthDiastema lengthRostrum depthBulla lengthBullae breadthShield-bullae depthSkull depthCarotid foramina
breadthInfraorbital canals
breadthMandible heightMandible length
Samplenumber
4
44444444
4
44
444444444
4
44
Mandible ramus depth 4Mandibular toothrow
length (alveolar)
Total length
Tail length
Body length
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Greatest skull length
4
343434343434343434
Samplesize
14
1414141414141414
13
1414
141414141414141414
14
14141414
9129
129
129
128
12109
151615161516
Mean RangeStandarddeviation
Coefficientof variation
Sylvilagus cunicularius cunicularius—continued
9.28
8.2586.1869.2639.7229.8737.3516.8216.41
23.44
9.9620.18
10.1110.6823.4317.2212.0628.9225.2936.2212.08
21.46
42.1643.9813.4316.90
8.10- 9.99
7.40- 9.3582.30- 88.9066.40- 70.9538.05- 41.4528.80- 30.5034.75- 39.2015.50- 18.0515.70- 17.25
21.95- 25.10
9.40- 10.3519.25- 20.95
9.25- 10.7010.00- 11.7022.00- 24.1016.20- 18.1511.20- 12.7027.90- 30.8024.35- 26.5034.65- 37.7511.35- 13.15
20.45- 23.45
40.65- 43.6042.45- 45.1012.75- 14.5016.20- 17.70
Sylvilagus floridanus
437.78443.0845.2237.00
392.5640555
96.2294.1765.3863.0857.3357.44
7.597.686.586.90
76.0476.82
369.00-464.00400.00-485.00
38.00- 55.0021.00- 50.00
331.00-424.00370.00-140.00
83.00-105.0091.00- 99.0061.00- 71.0058.00- 69.0050.10- 64.8053.50- 60.60
6.60- 8.456.55- 8.805.80- 7.305.10- 7.85
73.70- 78.3071.05- 80.35
0.57
0.551.691.370.850.531.410.690.40
0.93
0.320.50
0.380.560.710.610.420.830.570.810.55
0.85
0.880.850.470.52
macrocorpus
28.2325.27
6.598.02
27.4719.966.343.133.422.944.942.490.590.700.450.761.562.77
6.09
6.711.961.982.141.783.784.132.41
3.99
3.202.47
3.745.293.053.533.522.862.252.234.58
3.96
2.091.943.533.07
6.455.70
14.5821.687.004.926.593.325.234.668.614.337.759.106.86
11.002.063.60

22 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and cranial measurements of Sylvilagus—continued
SampleCharacter number
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
Incisive foramenlength
Basioccipital length
Basioccipital breadth
Diastema length
Rostrum depth
Bulla length
Bullae breadth
Shield-bullae depth
Skull depth
Carotid foraminabreadth
Infraorbilal canalsbreadth
Mandible height
Mandible length
Mandible ramus depth
Mandibular toothrowlength (alveolar)
Total length
Tail length
Body length
3434343434343434343434343434343434343434343434
121212
Samplesize
15161516151615161516151615161516151615161516151615161516151615161516151615161516151614161416
142114201420
Mean
SyliHlagus
61.9962.9936.1736.6226.5127.2134.8134.6715.7815.9214.7215.1621.2021.63
9.4010.0218.9119.209.339.639.61
10.1321.0221.2015.3815.7210.9711.0825.7426.2922.1922.3132.0232.4510.5510.9518.7219.4836.1737.2238.3239.3611.1711.3415.0515.35
RangeStandarddeviation
Coefficientof variation
floridanus macrocorpus—continued
59.80- 63.9057.20- 65.8534.65- 37.8034.25- 39.2524.65- 27.6025.60- 28.8531.00- 37.7030.30- 37.4013.55- 17.8513.75- 17.8014.15- 15.1014.40- 15.6520.65- 22.2520.50- 23.20
8.15- 10.609.40- 11.00
17.95- 20.0016.55- 21.207.85- 10.458.50- 10.758.60- 10.409.00- 11.70
19.45- 22.3518.25- 22.6514.60- 16.5513.75- 17.8010.50- 11.6010.20- 12.1024.50- 27.8023.95- 28.0521.05- 23.1520.70- 23.8030.45- 33.2030.50- 33.60
9.75- 11.659.75- 12.40
17.15- 20.2018.00- 20.7534.20- 38.2533.50- 40.1036.95- 39.9534.95- 42.5010.20- 11.9510.10- 11.9514.30- 15.9013.75- 16.25
Sylvilagus floridanus
416.43423.5250.2149.50
366.50375.20
371.00-448.00398.00-461.0035.00- 62.0033.00- 73.00
336.00-397.00345.00-409.00
1.392.610.951.310.901.041.962.231.301.090.280.360.420.690.630.480.711.110.670.630.540.640.861.270.671.230.330.461.091.130.600.740.830.840.580.800.870.751.131.870.872.060.560.560.450.61
holzneri
21.5918.658.00
11.6118.9018.43
2.234.142.623.573.403.835.626.438.216.851.882.391.983.186.674.823.765.797.166.575.656.334.086.014.337.862.984.174.234.312.713.312.592.595.517.304.653.843.125.012.265.244.984.933.004.00
5.194.40
15.9423.46
5.164.91
NUMBER 297 23
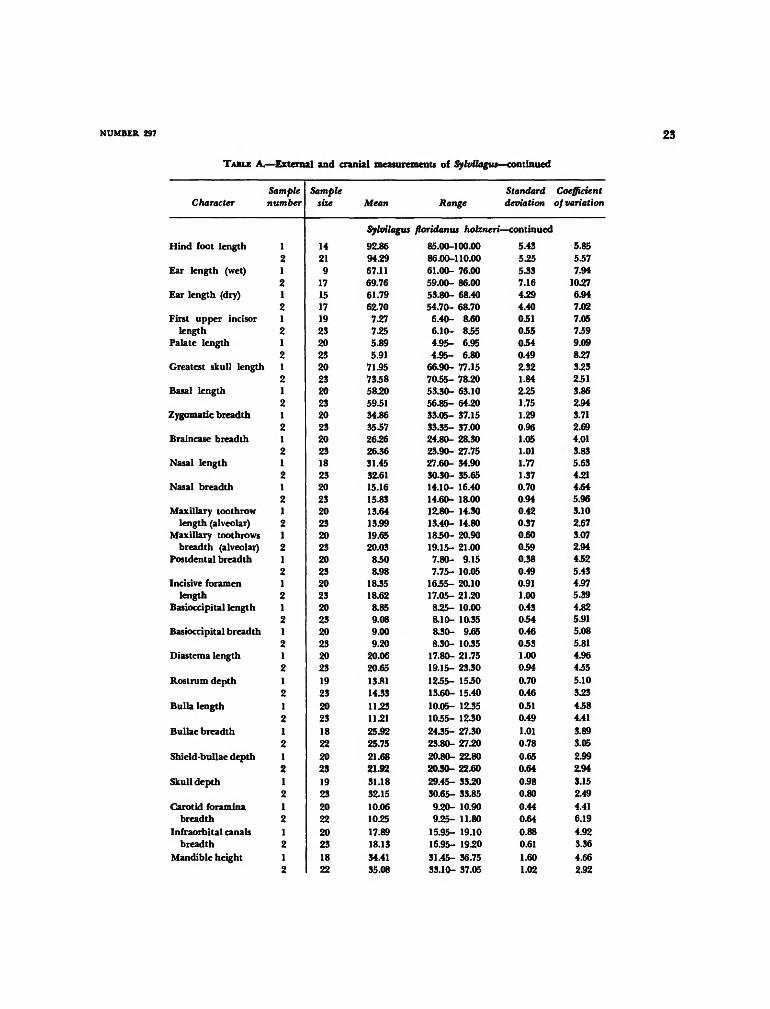
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Greatest skull length
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
Incisive foramenlength
Basioccipital length
Basioccipital breadth
Diastema length
Rostrum depth
Bulla length
Bullae breadth
Shield-bullae depth
Skull depth
Carotid foraminabreadth
Infraorbital canalsbreadth
Mandible height
Samplenumber
12121212121212121212121212121212121
2121212121212121
2
Samplesize
14219
17151719232023202320232023202318232023202320232023202320232023202319232023182220231923202220231822
Mean
Sylvilagus
92.86945967.1169.7661.7962.70
7.277.255.895.91
71.9573.5858.2059.5134.86355726.2626.3631.4532.6115.1615.8313.6413.9919.6520.03
8.508.98
18.3518.628.859.089.009.20
20.0620.6513.8114.3311.2311.2125.9225.7521.6821.9231.1832.1510.0610.2517.8918.1334.4135.08
RangeStandarddeviation
floridanus holzneri—continued
85.00-100.0086.00-110.0061.00- 76.0059.00- 86.0053.80- 68.4054.70- 68.70
6.40- 8.606.10- 8554.95- 6.954.95- 6.80
66.90- 77.1570.55- 78.2053.30- 63.1056.85- 64.2033.05- 37.1533.35- 37.0024.80- 28.3023.90- 27.7527.60- 34.9030.30- 35.6514.10- 16.4014.60- 18.0012.80- 14.3013.40- 14.8018.50- 20.9019.15- 21.007.80- 9.157.75- 10.05
1655- 20.1017.05- 21.208.25- 10.008.10- 10.358.30- 9.658.30- 10.35
17.80- 21.7519.15- 23.3012.55- 155013.60- 15.4010.05- 12.3510.55- 12.3024.35- 27.3023.80- 27.2020.80- 22.8020.80- 22.6029.45- 335030.65- 33.859.20- 10.909.25- 11.80
15.95- 19.1016.95- 19.2031.45- 36.7533.10- 37.05
5.435555.337.164.294.400.510550540.492.321.842.251.751.290.961.051.011.771.370.700.940.420.370.600590.380.490.911.000.430 5 40.460 5 31.000.940.700.460.510.491.010.780.650.640.980.800.440.640.880.611.601.02
Coefficientof variation
5.855577.94
10.276.947.027.057599.098.273532513.862.943.712.694.013.835.634.214.645.963.102.673.072.944525.434.975.394.825.915.085.814.964555.103.234584.413.893.052.992.943.152.494.416.194.923.364.662.92
24 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Mandible length
Mandible ramusdepth
Mandibular toothrowlength (alveolar)
Total length
Tail length
Body length
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Samplenumber
1
21212
56789
1056789
1056789
1056789
1056789
10
56789
1056789
10567
Samplesize
172119201921
121411
151114121411151114121411151114121411151114
110111074
11
710159
14131312161214131412
Mean
Sylvilagus
35.2536.2810.7810.7113.8914.15
RangeStandarddeviation
floridanus holzneri—continued
31.80- 37.4034.40- 38.009.85- 12.309.95- 11.60
13.10- 14.6013.50- 15.00
Sylvilagus floridanus
425.17406.00397.55398.00397.45409.4355.1749.2942.9144.4742.5549.64
370.00356.71354.64355.80354.91359.7992.2591.0786.9188.8788.5591.0768.0072.1067.0966.3067.5765.5055.3361.0459.5157.0560.0857.76
7.497.447.077.447.316.966.235.865.75
402.00-450.00345.00-435.00375.00-446.00367.00-434.00378.00-436.00335.00-445.0043.00- 65.0037.00- 68.0032.00- 61.0026.00- 67.0032.00- 58.0040.00- 57.00
352.00-390.00290.00-391.00343.00-385.00300.00-392.00329.00-393.00285.00-392.0088.00- 98.0085.00- 96.0081.00- 92.0083.00- 95.0077.00- 97.0085.00-102.0068.00- 68.0068.00- 89.0060.00- 72.0059.00- 85.0066.00- 71.0059.00- 78.0051.80- 60.5055.10- 66.9052.60- 66.3054.20- 58.9057.00- 62.1051.20- 66.70
6.50- 8.406.85- 8.156.40- 7.806.40- 8.106.45- 8.255.30- 8.105.30- 6.804.85- 7.005.15- 6.20
1.581.130.610.470.500.45
rrizabae
14.7822.5819.8522.7220.0428.886.589.438.94
10.168.185.60
10.5723.0612.3226.3119.6126.602.833.433.563.625.354.630.006.193.917.151.728.742.993.804.241.651.804.080.610.410.410.500.540.710.430.660.34
Coefficientof variation
4.483.125.704.373.593.20
3.485.564.995.715.047.05
11.9219.1220.8322.8519.2211.272.866.463.477.405.527.393.073.774.104.086.055.090.008.595.83
10.782.54
13.345.406.227.122.893.007.068.175.555.806.767.38
10.216.93
11.215.84

NUMBER 297 25
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Greatest skull
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
Samplenumber
89
1056789
1056789
1056789
1056789
1056789
1056789
1056789
1056789
1056
Samplesize
1612131314121612131314121612111314121612111313121612131314121612131314121612131314121612131314121612131314
Mean
Sylvilagus
6.035.945.76
725472.0670.1671.3971.1970.40585457.9056.5157.2357.3556.4535.2134.7433.7934.6934.2133.53265826.0925.4226.4026.0425.6931.9831.33305832.2431.0731.1914.9315.0314.7315.2114.6814.9114.2313.8113.6913.8713.6713.5819.9719.6819.29195619.2019.248.91855
RangeStandarddeviation
floridanus orizabae—continued
4.40- 7.105.40- 6.405.05- 6.70
70.30- 75.8069.40- 74.0567.30- 73.3067.70- 75.4568.80- 76.8068.15- 75.8556.10- 62.0555.05- 60.955255- 60.0053.45- 60.7055.10- 61.705350- 57.7533.95- 37.0033.20- 37.0032.40- 35.9032.85- 36.3533.35- 35.8031.25- 35.0525.35- 28.2025.25- 27.3023.80- 26.952450- 27.7024.90- 27.1023.70- 26.7530.15- 35.302955- 32.9527.05- 33.7029.60- 36.1528.90- 335028.40- 35.0013.80- 16.7513.80- 16.9513.45- 17.0013.30- 17.451350- 16.1512.65- 17.001355- 14.7513.05- 14.3513.05- 14.3013.05- 14.8513.00- 14.6013.00- 14.4518.35- 20.9518.80- 215518.40- 20.6518.40- 20.8518.35- 20.9518.20- 2050855- 9.707.85- 8.95
0.740.380521.771.472.152.292.171.921.591.772.082.911.751.140.840.981.071.080.800.990.860580.900.980.680.831.601.062.011.731.481550.820.971.241.090.931.180.330.460.380.540.390.430.660.700.760.670.670.770.410.32
Coefficientof variation
12.256.349.052.442.043.063.213.052.732.723.063.683523.052.012.382.823.173.102.352.973232.223533.722.633544.993.406585.354.764.955526.478.427.186.327.912.343.312.793.902.843.133.28
3583.943.443.474.014.633.73

26 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Incisive foramenlength
Basioccipital length
Basioccipital breadth
Diastema length
Rostrum depth
Bulla length
Bullea breadth
Shield-bullae depth
Samplenumber
789
1056789
10
56789
10
56789
10
56789
10
56789
1056789
1056789
1056789
10
Samplesize
12151211131412161212
131412161213
131412161213
131412161212
131412161213131412161214131412151210131412161212
Mean
Sylvilagus
8.418.688.368.62
17.9918.4017.7217.9518.2318.19
8.968.778.678.668.698.80
9.138.968.659.128.759.27
19.6519.9119.2219.6019.6619.65
14.0813.9313.7614.0113.31135710.6010.9110.72105610.68105024.8125.2424.3524.9824.7324.4321.2421.6121.09215421.3821.18
RangeStandarddeviation
floridanus orizabae—continued
7.50- 8.907.70- 9.607.70- 9.007.90- 9.25
17.10- 18.9517.20- 19.0016.15- 19.301650- 19.6516.90- 20.6517.60- 1955
8.60- 9.608.25- 9507.90- 9.157.75- 9.608.30- 9.307.65- 9.60
8.60- 9.758.45- 10.60755- 9558.25- 9.808.05- 9.808.50- 10.10
18.70- 21.3018.75- 20.7017.55- 21.6018.20- 21.4518.45- 21.9018.80- 21.25
12.70- 15.0012.85- 14.7013.00- 14.9013.40- 14.8512.10- 14.1513.15- 14.9010.00- 11.4010.40- 11.7510.05- 11.8010.15- 11.309.95- 11.659.85- 11.45
23.65- 26.5023.80- 275522.80- 26.0023.15- 26.8022.80- 25.7022.85- 255020.45- 22.4021.00- 22.6519.85- 22.4020.70- 22.8020.20- 22.4520.80- 2155
0.470500.350.380.490510.940.850.980.54
0.290.320.320.610.31052
0.330.600.690.490580.60
0.640.481.220.780.980.80
0590.600550.430590.41
0.420.400.490.360.480.410.800.951.071.090.810.84
0.590540.700560.560.27
Coefficientof variation
5.575.764.214.432.732.775.314.715.372.97
3.193.673.747.083.575.88
3.606.717.945.406.686.47
3.262.436.364.984.994.09
4.194.343.983.104.463.274.003.694.613.414.483.91
3.233.764.394.363.263.442.762513.322582.601.25

NUMBER 297 27
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Skull depth
Carotid foraminabreadth
Infraorbital canalsbreadth
Mandible height
Mandible length
Mandible Ramusdepth
Mandibular toothrowlength (alveolar)
Total length
Tail length
Body length
Samplenumber
56789
1056789
1056
789
1056789
1056789
1056789
1056789
10
2a2b2c2a2b2c2a2b2c
Samplesize
1314
1216121113141216121113141216121313131215121313
121214
12131314121594
1314121594
1921
1921
192I
Mean
Sylvilagus
31.1130.9430.4831.2431.0230.4810.2110.079.70
10.069.87
10.4117.8917.6617.3317.4517.1616.9234.0633.9832.9233.2133.1732.6635.7735.3334.2235.1034.3334.071026103010.1710.3510.159.46
14.3214.1413.7514.0713.7013.25
RangeStandarddeviation
floridanus orizabae—continued
29.70- 32.5030.20- 31.8028.80- 33.1029.20- 33.7530.05- 32.0029.40- 31.50
9.35- 11.159.05- 11.358.50- 10.758.90- 11.409.30- 10.959.95- 10.90
17.25- 19.6016.35- 18.801535- 18.9516.30- 18.9516.30- 18.5015.35- 18.6032.30- 35.8532.65- 36.1031.25- 35.7030.75- 36.5030.85- 36.0530.40- 34.7533.40- 37.7033.00- 36.8532.75- 35.7031.80- 38.3532.50- 37.4032.40- 35.70930- 11.159.60- 11.409.60- 11.359.30- 11.109.20- 11.459.25- 9.80
13.00- 15.1513.40- 14.9013.10- 14.8013.10- 15.4512.75- 14.4513.00- 1335
0.710.461.111.050.650.610300.590.630.630.510.440.620.600.990.800.630.951.100.911.311.641.691.521.451.060.841.911.380.980.49030030
0320.670.240.620.420.470.610.48053
Sylvilagus graysoni graysoni466.11502.00480.0050.2143.0036.00
415.89459.00444.00
400.00-490.00470.00-534.00
28.00- 65.0030.00- 56.00
371.00-438.00440.00-478.00
23.24
9.94
17.27
Coefficientof variation
2.281.483.653.372.101.994.915.886.496.255.144.193.463.385.704.563.665.623.242-693.994.945.114.654.063.002.445.434.012.874.764.764.885.016392 3 34.352-943.444.353.481.77
4.99
19.79
4.15

28 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and cranial measurements of Sylvilagus—continued
Character
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Greatest skull length
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
Incisive foramenlength
Basiocdpital length
Basiocdpital breadth
Diastema length
Samplenumber
2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c
Samplesize
1921411
1921
192I
id21
1921
1921
1921
1921
19
21
1921
1921
1921
1921
1921
1921
19
21
1921
Mean
Sylvilagus
95.4297.5095.0063.7566.0063.0056.8358.2557.50
8.938.007308.048.7S8.15
79.9283.38813564.83675066.0037.0037.9036.9027 3327.85273534.3135.0336.7514.9016.3314.85155314.9814.9522.6023.1023.0010.1510.00103519.9320.0519.7510.0910.909.759.739.80930
22.0522.7022.80
RangeStandarddeviation
graysoni graysoni—continued
90.00-102.0095.00-100.00
63.00- 65.00
53.00- 60.4054.90- 60.00
7.75- 9.957.60- 8.40
7.00- 9308.40- 9.05
76.00- 84.2083.05- 83.70
6155- 68.306630- 67.90
3530- 38.9537.60- 38.20
2630- 28.052735- 28.15
32.00- 36.8534.85- 3550
13.65- 16.7016.05- 16.60
14.80- 16.3014.80- 15.15
21.70- 23.7022.85- 23.35
9.65- 10.959.90- 10.10
18.85- 21.4019.80- 2030
930- 10.8010.80- 11.00
9.25- 10.70950- 10.40
20.55- 23.6022.60- 22.80
3.17
0.96
1.94
0.52
0.61
2.21
1.88
0.95
0.49
1.50
0.86
0.43
0 3 3
0.37
0.79
0.45
0.32
0.88
Coefficientof variation
3.32
1.50
3.41
6.18
7.60
2.76
2 9 0
236
1.78
4.38
5.77
2.80
2 3 3
3.64
3.99
4 3 0
3.32
3.99

NUMBER 297 29
TABLE A.—External and
Character
Rostrum depth
Bulla length
Bullae breadth
Shield-bullaedepth
Skull depth
Carotid foraminabreadth
Infraorbital canalsbreadth
Mandible height
Mandible length
Samplenumber
2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c2a2b2c
Mandible ramus depth 2a
Mandibular toothrowlength (alveolar)
Total lengthTail lengthBody lengthHind foot lengthEar length (wet)Ear length (dry)First upper incisor
lengthPalate lengthGreatest skull lengthBasal lengthZygomatic breadthBraincase breadthNasal lengthNasal breadthMaxillary toothrow
length (alveolar)Maxillary toothrows
breadth (alveolar)
2b2c
r 2a2b2c
1
]
I
1
I
Samplesize
1921
1921
1921
1921
1921
1921
1921
1921
1921
1921
1921
111111111111
1111111111111111
11
11
cranial measurements of Sylvilagus—continued
Mean
Sylvilagus
16.4717.0517.3011.31115310.6527.4827.5827.0023.0323.9522.7033.3133.8333.5511.9912.1511.5519.7120.3319.1538.8440.6540.7041.15423341.8012.7312.9312.6015.7115.7815.00
RangeStandarddeviation
graysoni graysoni—continued1530- 17.8016.10- 18.00
10.60- 12.3011.05- 11.40
26.50- 28.8027.40- 27.75
21.70- 24.4023.60- 24.30
31.80- 34.8033.80- 33.85
11.10- 12.9511.95- 12.35
17.95- 20.7019.95- 20.70
36.30- 42.604035- 40.75
39.30- 45.1041.90- 43.15
11.80- 13.9512-75- 13.10
15.10- 16.3015.65- 15.90
059
034
0.73
0.73
0.85
052
0.72
150
1.44
0.62
0.37
Sylvilagus graysoni badistes436.913255
404.3690.7362.1857.11
9.147.84
785263.95375727.5734.4214.97
16.09
22.49
412.00-472.0026.00- 45.00
373.00-427.0086.00- 95.0055.00- 65.0050.80- 59.70
850- 9.957.15- 9.10
75.90- 79.956155- 65.6036.15- 38.7526.65- 28.7032.45- 36.0014.15- 15.80
1555- 16.60
21.85- 23.35
17.005.94
17.132.692.602.43
0.600521.361.470.940.700.990.49
0.40
0.44
Coefficientof variation
3.61
4.78
2.64
3.18
255
4.32
3.67
3.87
330
4.85
2.35
3.8918554542.964.18455
6526.661.742592512522.88358
250
1.96

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A.—External and
Character
Postdental breadthIncisive foramen
lengthBasioccipital lengthBasioccipital breadthDiastema lengthRostrum depthBulla lengthBullae breadthShield-bullae depthSkull depthCarotid foramina
breadthInfraorbital canals
breadthMandible heightMandible length
Samplenumber
1
]
1
]
Mandible ramus depthMandibular toothrow
length (alveolar)
Total length
Tail length
Body length
Hind foot length
Ear length (wet)
Ear length (dry)
First upper incisorlength
Palate length
Greatest skull length
Basal length
Zygomatic breadth
Braincase breadth
Nasal length
Nasal breadth
Maxillary toothrowlength (alveolar)
Maxillary toothrowsbreadth (alveolar)
Postdental breadth
]
IILIIIIII
II
1212121212121212121212121212121212
Samplesize
11
111111111111111111
1111
11111111
117
107
107
117
11797
17101710171117101610161017101710171017101510
cranial measurements of Sylvilagus—continued
Mean RangeStandarddeviation
Sylvilagus graysoni badistes—continued10.23
18.579.99
10.0720.1916.1511.4827.0423.1933.20
11.9720.11
38.4040.6412.6615.89
9.60- 10.60
17.75- 19.359.10- 10.609.65- 10.65
18.90- 21.2015.35- 16.8510.75- 11.8026.35- 28.2022.70- 23.7532.25- 33.90
11.30- 12.8019.20- 21.15
37.40- 40.2539.75- 41.7012.00- 13.3515.40- 16.80
Sylvilagus audubonii351.64339.7143.4038.29
321.20301.43773575.2975.0067.0062.0860.946.946.864.954.97
64.5764.27513151.4132.2732.2224.6924.%26.6427.0012.8512.9412.0712.0417.2716.957.277.09
302.00-410.00320.00-360.0021.00- 64.0028.00- 52.00
281.00-386.00280.00-315.0067.00- 95.0071.00- 78.0065.00- 91.0064.00- 70.0057.90- 65.8058.10- 63.506.15- 7.705.25- 8.254.10- 5.654.35- 5.40
59.20- 68.3062.00- 67.3546.55- 54.5048.95- 53.4529.75- 33.9030.70- 33.0023.10- 26.4023.95- 25.7524.45- 28.8524.90- 29.0511.50- 13.7511.90- 13.7011.35- 12.6011.25- 12.6016.10- 18.6015.80- 17.806.20- 8.306.75- 7.60
0.30
0.520.450.320.660.470.300.520.300.58
0.430.61
0.960.640.410.47
barvulus31.4214.9710.998.75
35.8112357.482.368.962.312361.840.490.870.370.312.341.692.001.421.110.860.800371.351.350.760.630.360.470.740.650.460J26
Coefficientof variation
2.90
2.774.533.153.272.942.591.921.301.74
3373.06
2301.583.232.98
8.944.41
25.3422.8611.154.179.643.14
11.943.454.123.027.11
12.757356.153.622.633.862.753.432.653.232595.084.995.904.892.973.944.273.836.373.64

NUMBER 297 31
TABLE A.—External and cranial measurements of Sylvilagus—continued
SampleCharacter number
Incisive foramenlength
Basioccipital length
Basioccipital breadth
Diastema length
Rostrum depth
Bulla length
Bullae breadth
Shield-bullae depth
Skull-depth
Carotid foraminabreadth
Infraorbital canalsbreadth
Mandible height
Mandible length
Mandible ramus depth
Mandibular toothrowlength (alveolar)
Total lengthTail lengthBody lengthHind foot lengthEar length (wet)Ear length (dry)First upper incisor
lengthPalate lengthGreatest skull lengthBasal lengthZygomatic breadthBraincase breadthNasal lengthNasal breadthMaxillary toothrow
length (alveolar)Maxillary toothrows
breadth (alveolar)Postdental breadthIncisive foramen
length
12121212121212121212121212
. 1212
333333
33333333
33
3
3
Samplesize
171017101710171017101711149
179
149
161017101510139
1510159
66666
10
1010101010101010
1010
10
10
Mean
Sylvilagus15.5615.908.278.317.69739
16.8717.0612.0712.1112.8512.3926.6325.7821.7621.6629.3029.678.388.16
15.8015.7831.6131.3631.9531.3910.2410.3312.2712.41
RangeStandarddeviation
Coefficientof variation
audubonii parvulus—continued14.15- 16.4014.85- 17.157.65- 9.007.60- 9.007.05- 9.006.95- 8.75
15.25- 18.3015.40- 18.4011.05- 12.6011.45- 13.7012.00- 13.9511.75- 13.1524.50- 29.252430- 26.9020.55- 23.1520.85- 22.9028.30- 30.8528.00- 30.457.40- 9.157.10- 9.25
14.75- 17.1015.00- 163527.25- 34.1029.95- 33.0027.80- 33.5529.60- 34.90
9.35- 10.858.95- 11.30
11.70- 12.651130- 12.95
0.660.750.390.370.400340.870.870.410.660350.411.320.870.640.640.640.770320.580.680301.901.011321350370.650.280.48
Sylvilagus audubonii goldmani369.2544.00
325.257530685060.08
7.345.82
66.7853.6133.3424.96283413.96
12.9618.21
8.05
16.73
363.00-380.0043.00- 46.00
320.00-334.0072.00- 78.0060.00- 80.005630- 62.20
6.70- 7.70530- 6.30
64.15- 69.6052.00- 56.3531.35- 34.4023.75- 25.952755- 29.9013.00- 14.65
12.45- 13.3517.65- 18.95
7.30- 8.80
15.65- 17.15
7.461.416.082.658.392.60
0.420.332531.931.190.911.240.65
0.390.48
035
0.63
4.254.724.774.495.277.075.135.103.445.454563.304.973.372.962.942.192.606.177.064.283.156.003534.764.955326562.323.90
2.023.211.87330
12544.34
5.695393.343393.563.664.344.68
2.992.65
6.89
3.76

32 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE A
Character
Basioccipital lengthBasioccipital breadthDiastema lengthRostrum depthBulla lengthBullae breadthShield-bullae depthSkull depthCarotid foramina
breadthInfraorbital canals
breadthMandible heightMandible lengthMandible ramus
depthMandibular toothrow
length (alveolar)
—External and cranial measurements of Sylvilagus—continuec
Samplenumber
33333333
3
333
3
3
Samplesize
1010101099
1010
10
101010
10
10
Mean
Sylvilagus
8.968.39
18.0913.0911.3824.5121.3129.29
9.87
16.9432.1633.01
10.32
13.34
RangeStandarddeviation
Coefficientof variation
audubonii goldmani—continued
8.75- 9.157.95- 8.75
17.15- 19.1512.60- 13.608.95- 12.40
23.45- 25.5520.65- 22.3028.05- 30.15
9.35- 10.35
16.25- 17.7529.90- 33.7031.95- 34.85
9.50- 10.85
12.60- 14.00
0.190.350.820.361.000.840.630.81
0.36
0.541.350.88
0.48
0.45
2.154.124.552.738.813.422.972.75
3.63
3.174.202.67
4.70
3.35
TABLE B.—List of 26 cranial characters and their discriminant multipliers used in a discriminantfunction analysis comparing individuals of S. floridanus, S. cunicularius, and 5. graysoni fromwestern Mexico
Character
Upper incisor lengthLeast palatal lengthGreatest skull lengthBasal lengthGreatest zygomatic breadthBraincase breadthGreatest nasal lengthGreatest nasal breadthMaxillary toothrow lengthMaxillary toothrows breadthPostdental breadthIncisive foramen lengthBasioccipital lengthBasioccipital breadthDiastema lengthRostrum depthBulla lengthBullae breadthShield-bullae depthSkull depthCarotid foramina breadthInfraorbital canals breadthMandible heightMandible lengthMandible ramus heightMandibular toothrow length
First discriminantmultiplier
.128990598270.126300
-.028489-.012512-552760-.114320
.052802
.050421
.049597
.167790509620.158450
-.425280-.351690-.061170
.175600
.070920-.033701
.102300
.455340-.084252
.049897-.056153
.334970
.086564
Second discriminantmultiplier
-.203200-.311270
.020535-.018176
.063484-.010618
.152860
.086462
.050469-558320-.095209-.022496-.077523-514120
.044509
.063670
.333940-.005489
.117760
.018456
.154000
.040451
.161480-.224970-.461430
.498630

NUMBER 297 33
TABLE C.—List of 26 characters and their discriminant multipliers used in a discriminantfunction analysis comparing individuals of S. f. macrocorpus and S. f. orizabae from westernMexico (see "Comparisons" under 5. /. macrocorpus for discussion)
Character
Upper incisor lengthLeast palatal lengthGreatest skull lengthBasal lengthGreatest zygomatic breadthBraincase breadthGreatest nasal lengthGreatest nasal breadthMaxillary toothrow lengthMaxillary toothrows breadthPostdental breadthIncisive foramen lengthBasioccipital length
Discriminantmultiplier
.013404
.126090-.150300
.343930-.086063-.159110
.128430-.143300-.025437
.398370
.241650-.034595-.280460
Character
Basioccipital breadthDiastema lengthRostrum depthBulla lengthBullae breadthShield-bullae depthSkull depthCarotid foramina breadthInfraorbital canals breadthMandible heightMandible lengthMandible ramus heightMandibular toothrow length
Discriminantmultiplier
.071662-.349320
.338130
.288650
.066839
.127060
.022495-.198880
.086752-.012280-.083788-.269720-.052596

Literature Cited
Allen, J. A.1877. Leporidae. In E. Coues and J. A. Allen, Monographs
of North American Rodentia, pages 265-378. Wash-ington, D.C.: United States Geological Survey ofthe Territories.
1890. Notes on Collections of Mammals made in Centraland Southern Mexico, by Dr. Audley C. Buller,with Descriptions of New Species of the GeneraVespertilio, Sciurus, and Lepus. Bulletin of theAmerican Museum of Natural History, 3:175-194.
1903. List of Mammals Collected by Mr. J. H. Batty inNew Mexico and Durango, with Descriptions of NewSpecies and Subspecies. Bulletin of the AmericanMuseum of Natural History, 19:587-612.
1904. Mammals from Southern Mexico and Central andSouth America. Bulletin of the American Museumof Natural History, 20:29-80.
1906. Mammals from the States of Sinaloa and Jalisco,Mexico, Collected by J. H. Batty during 1904 and1905. Bulletin of the American Museum of NaturalHistory, 22:191-262.
Armstrong, D. M., and J. K. Jones, Jr.1971. Mammals from the Mexican State of Sinaloa,
I: Marsupialia, Insectivora, Edentata, Lagomorpha.Journal of Mammalogy, 52:747-757.
Baird, S. F.1857. North American Mammals, xxxiv + 757 pages, 60
plates. Philadelphia: J. B. Lippincott & Co.Baker, R. H., and J. K. Greer
1962. Mammals of the Mexican State of Durango. Publi-cations of the Museum, Michigan State University,Biological Series, 2:25-154.
Coues, E., and J. A. Allen1877. Monographs of North American Rodentia. x + 1091
pages. Washington, D.C.: United States GeologicalSurvey of the Territories.
Diersing, V. E.1978. A Systematic Rex>ision of Several Specis of Cotton-
tails (Genus Sylvilagus) from North and SouthAmerica. 872 pages. Doctoral dissertation, Universityof Illinois, Urbana.
Elliot, D. G.1903. A List of a Collection of Mexican Mammals with
Descriptions of Some Apparently New Forms. FieldColumbian Museum Publication, 71, ZoologicalSeries, 3:141-149.
Findley, J. S., A. H. Harris, D. E. Wilson, and C. Jones1975. Mammals of New Mexico. 360 pages. Albuquerque:
University of New Mexico Press.Genoways, H. H.
1973. Systematics and Evolutionary Relationships of theSpiny Pocket Mice of the Genus Liomys. SpecialPublications of the Museum, Texas Tech University,5:1-368.
Genoways, H. H., and J. K. Jones, Jr.1973. Notes on Some Mammals from Jalisco, Mexico.
Occasional Papers of the Museum, Texas TechUniversity, 9:1-22.
Hoffmeister, D. F., and E. G. Zimmerman1967. Growth of the Skull in the Cottontail (Sylvilagus
floridanus) and Its Application to Age-Determina-tion. The American Midland Naturalist, 78:198-206.
Hsu, T. C, and K. Benirschke1971. An Atlas of Mammalian Chromosomes. Volume 5,
folio 212. New York: Springer-Verlag.Lyon, M. W., Jr.
1904. Classification of the Hares and Their Allies. Smith-sonian Miscellaneous Collections, 45:321—447.
Mcarns, E. A.1896. Preliminary Description of a New Subgenus and
Six New Species and Subspecies of Hares, from theMexican Border of the United States. Proceedingsof the United States National Museum, 18:551-565.
Merriam, C. H.1893. Preliminary Descriptions of Four New Mammals
from Southern Mexico, Collected by E. W. Nelson.Proceedings of the Biological Society of Washington,8:143-146.
Miller, G. S., Jr.1899. Descriptions of Six New American Rabbits. Pro-
ceedings of the Academy of Natural Sciences ofPhiladelphia, 51:380-390.
Nelson, E. W.1899. General Account of the [Tres Marias] Islands, with
Reports on Mammals and Birds. In E. W. Nelson,L. Stejneger, M. J. Rathbun, and J. M. Rose,Natural History of the Tres Marias Islands,Mexico. North American Fauna, 14:1-97.
1904. Descriptions of Seven New Rabbits from Mexico.Proceedings of the Biological Society of Washington,17:103-110.
1907. Descriptions of New American Rabbits. Proceed-ings of the Biological Society of Washington, 20:81-84.
1909. The Rabbits of North America. North AmericanFauna, 29:1-314.
Rzedowski, J., and R. McVaugh1966. La vegetacion de Nueva Galicia. Contributions of
the Herbarium, University of Michigan, 9:1-123.Thomas, O.
1890. On a Collection of Mammals from Central VeraCruz, Mexico. Proceedings of the Zoological Societyof London, 4 Feb:71-76.
Waterhouse. G. R.1848. A Natural History of the Mammalia. Volume II,
500 pages. London: Hippolyte Bailliere.Zweifel, R. G.
1960. Results of the Puritan-American Museum of NaturalHistory Expedition to Western Mexico, 9: Herpe-tology of the Tres Marias Islands. Bulletin of theAmerican Museum of Natural History, 119:81-128.
34



REQUIREMENTS FOR SMITHSONIAN SERIES PUBLICATION
Manuscripts intended for series publication receive substantive review within theiroriginating Smithsonian museums or offices and are submitted to the Smithsonian InstitutionPress with approval of the appropriate museum authority on Form SI-36. Requests forspecial treatment—use of color, foldouts, casebound covers, etc.—require, on the sameform, the added approval of designated committees or museum directors.
Review of manuscripts and art by the Press for requirements of series format and style,completeness and clarity of copy, and arrangement of all material, as outlined below, willgovern, within the judgment of the Press, acceptance or rejection of the manuscripts and art.
Copy must be typewritten, double-spaced, on one side of standard white bond paper,with iyA" margins, submitted as ribbon copy (not carbon or xerox), in loose sheets (notstapled or bound), and accompanied by original art. Minimum acceptable length is 30 pages.
Front matter (preceding the text) should include: title page with only title and authorand no other information, abstract page with author/title/series/etc., following the establish-ed format, table of contents with indents reflecting the heads and structure of the paper.
First page of text should carry the title and author at the top of the page and an unnum-bered footnote at the bottom consisting of author's name and professional mailing address.
Center heads of whatever level should be typed with initial caps of major words, withextra space above and below the head, but with no other preparation (such as all caps orunderline). Run-in paragraph heads should use period/dashes or colons as necessary.
Tabulations within text (lists of data, often in parallel columns) can be typed on the textpage where they occur, but they should not contain rules or formal, numbered table heads.
Formal tables (numbered, with table heads, boxheads, stubs, rules) should be sub-mitted as camera copy, but the author must contact the series section of the Press for edito-rial attention and preparation assistance before final typing of this matter.
Taxonomic keys in natural history papers should use the alined-couplet form in thezoology and paleobiology series and the multi-level indent form in the botany series. Ifcross-referencing is required between key and text, do not include page references within thekey, but number the keyed-out taxa with their corresponding heads in the text.
Synonymy in the zoology and paleobiology series must use the short form (taxon,author, year:page), with a full reference at the end of the paper under "Literature Cited."For the botany series, the long form (taxon, author, abbreviated journal or book title, volume,page, year, with no reference in the "Literature Cited") is optional.
Footnotes, when few in number, whether annotative or bibliographic, should be typedat the bottom of the text page on which the reference occurs. Extensive notes must appear atthe end of the text in a notes section. If bibliographic footnotes are required, use the shortform (author/brief title/page) with the full reference in the bibliography.
Text-reference system (author/year/page within the text, with the full reference in a"Literature Cited" at the end of the text) must be used in place of bibliographic footnotes inall scientific series and is strongly recommended in the history and technology series:"(Jones, 1910:122)" or ".. . . Jones (1910:122)."
Bibliography, depending upon use, is termed "References," "Selected References," or"Literature Cited." Spell out book, journal, and article titles, using initial caps in all majorwords. For capitalization of titles in foreign languages, follow the national practice of eachlanguage. Underline (for italics) book and journal titles. Use the colon-parentheses systemfor volume/number/page citations: "10(2):5-9." For alinement and arrangement ofelements, follow the format of the series for which the manuscript is intended.
Legends for illustrations must not be attached to the art nor included within the text butmust be submitted at the end of the manuscript—with as many legends typed, double-spaced, to a page as convenient.
Illustrations must not be included within the manuscript but must be submitted sepa-rately as original art (not copies). All illustrations (photographs, line drawings, maps, etc.)can be intermixed throughout the printed text. They should be termed Figures and shouldbe numbered consecutively. If several "figures" are treated as components of a single largerfigure, they should be designated by lowercase italic letters (underlined in copy) on the illus-tration, in the legend, and in text references: "Figure 9b.." If illustrations are intended to beprinted separately on coated stock following the text, they should be termed Plates and anycomponents should be lettered as in figures: "Plate 9.b_." Keys to any symbols within anillustration should appear on the art and not in the legend.
A few points of style: (1) Do not use periods after such abbreviations as "mm, ft,yds, USNM, NNE, AM, BC." (2) Use hyphens in spelled-out fractions: "two-thirds." (3)Spell out numbers "one" through "nine" in expository text, but use numerals in all othercases if possible. (4) Use the metric system of measurement, where possible, instead ofthe English system. (5) Use the decimal system, where possible, in place of fractions.(6) Use day/month/year sequence for dates: "9 April 1976." (7) For months in tabular list-ings or data sections, use three-letter abbreviations with no periods: "Jan, Mar, Jun," etc.
Arrange and paginate sequentially EVERY sheet of manuscript—including ALL frontmatter and ALL legends, etc., at the back of the text—in the following order: (1) title page,(2) abstract, (3) table of contents, (4) foreword and/or preface, (5) text, (6) appendixes,(7) notes, (8) glossary, (9) bibliography, (10) index, (11) legends.

«•
,111/.,





























