Embed Size (px)
Citation preview

Distribution and Morphology ofLuteinising Hormone-Releasing HormoneNeurones in a Species of Wild Antelope,the Springbok (Antidorcas marsupialis)
JANE E. ROBINSON,1* DONAL C. SKINNER,1 JOHN D. SKINNER,2
AND MARTIN A. HAUPT2
1Laboratory of Neuroendocrinology, Department of Neurobiology, The Babraham Institute,Cambridge CB2 4AT, United Kingdom
2Mammal Research Institute, The University of Pretoria, 0002, Pretoria, South Africa
ABSTRACTThe distribution and morphology of luteinising hormone-releasing hormone (LHRH)
neurones varies between species. The primary purpose of this study was to characterize thedistribution and morphology of the LHRH system in a species of antelope, the springbok. Thiswild antelope has a well-defined social structure in which reproductive activity is confined to afew dominant, territorial rams. We also sought to determine whether social or reproductivestatus could be accounted for by differences in the distribution or morphology of hypothalamicLHRH neurones. Eleven anoestrous female, nine breeding territorial male (TM) and eight‘‘bachelor’’ male (BM) springbok were obtained, and their reproductive and body conditionswere assessed. By using standard immunocytochemical techniques, the LHRH system wasvisualised in the brains of four animals from each group. Immunoreactive neurones werelocated in a continuum from the septum to the arcuate nucleus, with the majority at the levelof the organum vasculosum of the lamina terminalis. Neither the distribution nor the numberof cells differed among the three groups. Furthermore, the area of LHRH perikarya wassimilar in both groups of males, suggesting that reproductive differences between TMs andBMs lie at another level of the neuroendocrine axis. The anoestrous females had significantlylarger neurones than males (TM plus BM). This may reflect a sex difference in the LHRHsystem of this wild antelope. However, an alternative explanation is that the male/femaledifference is related to the comparatively inactive reproductive neuroendocrine state of thefemales. J. Comp. Neurol. 389:444–452, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: immunocytochemistry; preoptic area; social status; cell number; cell area
The activity of the reproductive axis of mammals iscontrolled by changes in the release of the decapeptide,luteinising hormone-releasing hormone (LHRH), which issynthesised in neurones located primarily in the hypothala-mus and the preoptic area (poa). The origin of theseneurones is extremely unusual, because they arise in theolfactory placode and migrate into the brain early ingestational life (Schwanzel-Fukuda and Pfaff, 1989; Wrayet al., 1989). In the adult, these neurones are not located indiscrete nuclei; rather, they are scattered in a continuumin the ventral and medial aspects of this region. There aresubstantial species differences in the rostrocaudal locationof LHRH cell bodies. Whereas the majority of cells of therat, golden hamster, Djungarian hamster, and sheep re-side in the relatively rostral regions of the septum, thediagonal band of Broca, and the poa (Jennes and Stumpf,
1980; Witkin et al., 1982; Merchenthaler et al., 1984;Lehman et al., 1986; Caldani et al., 1988; Yellon et al.,1990), those of the ferret, bat, monkey, and human (Silver-man et al., 1982; King and Anthony, 1984; Sisk et al., 1988;Anthony et al., 1989) are located in more caudal regions,including the mediobasal hypothalamus (mbh). There arealso species differences in morphology. For example, inrodents, LHRH cells are normally uni- or bipolar, whereasthey frequently exhibit more complex morphology in thesheep (Lehman et al., 1986) and the rhesus monkey
*Correspondence to: Jane E. Robinson, Laboratory of Neuroendocrinol-ogy, The Babraham Institute, Cambridge CB2 4AT, United Kingdom.E-mail: [email protected]
Received 31 March 1997; Revised 22 July 1997; Accepted 23 July 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 389:444–452 (1997)
r 1997 WILEY-LISS, INC.

(Silverman et al., 1982). Whether these species differencessubserve any function is unclear. Our current knowledge isderived mainly from studies in a few species of laboratoryrodents or from farm animals. The primary purpose of thisstudy was to determine the location and morphologicalcharacteristics of LHRH neurons in a genus of animals inwhich we have no current data; Antidorcas. This genusincludes the springbok (Antidorcas marsupialis), an ante-lope that can be found in southwest Africa. The springbokis of particular interest because of its well-documentedsocial and dominance structure. Studies in a few otherspecies have shown that the number, distribution, ormorphology of LHRH neurones can be altered by ananimal’s social or endocrine status (King et al., 1987; Tangand Sisk, 1992; Francis et al., 1993; Dellovade and Riss-mann, 1994; Rubin and King, 1994). In the springbok, onlya small proportion of males breed, and these breedingmales actively protect their territories with displays ofaggressive behaviour. They are fertile throughout the year(Van Zyl and Skinner, 1970) and are easily identified bytheir solitary station. In contrast, nonbreeding ‘‘bachelor’’males (BMs) form herds that contain either males only ormales, females, and their young. Female springbok do nothave a well-defined breeding season, although individualfemales appear to have a period of anoestrus that is notsynchronised among animals. There is no knowledge of thereproductive neuroendocrine axis in this species. There-fore, a second purpose of this study was to compare thedistribution and morphological characteristics of LHRHneuronees in groups of ‘‘territorial’’ male (TM), BM, andfemale springbok. A preliminary report of this work waspresented at the annual meeting of the Society for theStudy of Fertility in 1995.
MATERIALS AND METHODS
Animals
This study was carried out at a time before the summerrains, when vegetation was relatively sparse. At this time,springbok herds comprised BMs and females with theiryoung, whereas TMs were solitary. Twenty-eight adultspringbok were obtained from two sites in South Africabetween 15 and 29 May as part of a routine cul. Half camefrom the Kalahari Gemsbok National Park, and the remain-der came from the S.A. Lombard Nature Reserve (latitudeof both is between 25° and 27°S). Animals were dividedinto three groups, depending on their sex and social status;females (F; n 5 11); breeding TMs (n 5 9), and nonbreed-ing, BMs (n 5 8). All were shot through the neck by anexpert marksman; death was instantaneous. The car-casses of the animals were weighed, and the gonads,epididymides, and kidney fat (an index of body condition;Vanrooyen, 1993) were dissected free from surroundingtissue and weighed. Epididymal sperm reserves wereassessed for all males (Dott and Skinner, 1967), and thereproductive state of the ovaries was noted. Females werealso examined for evidence of lactation or pregnancy.
LHRH immunocytochemistry
Four animals from each group (all obtained in theKalahari) were used for the immunocytochemical studies.These were perfused through the carotid arteries with 500ml heparinised saline (10,000 units/liter) followed by 3liters of 4% paraformaldehyde in 0.1 M phosphate-
buffered saline, pH 7.6. Solutions were introduced into thevessels by using a small, portable, variable-speed, peristal-tic pump (Easy-load, Masterflex, model 7518-10; ColeParmer Instrument Co., Bishop’s Stortford, United King-dom) driven by a petrol generator. Perfusion began within1–3 minutes of shooting and took approximately 15 min-utes (i.e., flow rate 200 µl/minute). Solutions had beenmade up earlier; these were stored in a polythene con-tainer for up to 1 week and were filtered just before use.Brains were removed from the cranium, immersion fixedfor another 12 hours, and a hypothalamic block wasdissected from each and stored in 30% sucrose solutionuntil it was sectioned.
A series of sections was cut at 30 µm in a coronal planeon a freezing microtome from the level of the septumthrough the hypothalamus to its caudal boundaries, andeach fifth section was processed for LHRH immunoreactiv-ity with the following method. Sections were washed in0.05 M Tris-buffered saline (TBS; 15 minutes), placed in0.3% hydrogen peroxide and 40% methanol for 20 minutes(to deactivate the endogenous peroxidases), and trans-ferred to 2% normal goat serum (2 hours; Vector, Peterbor-ough, United Kingdom) in TBS with 0.3% bovine serumalbumin (BSA) and 0.25% Triton X-100. Free-floatingsections were then reacted with a polyclonal LHRH antise-rum that recognises amino acids 3, 4, 7, 8, 9, and 10 of thedecapeptide (rabbit LR1; 1:40,000) for 48 hours at 4°C.Sections were incubated at room temperature with affinity-purified biotinylated goat anti-rabbit immunoglobulins (2hours; 1:200; Vector) and streptavidin-biotin-peroxidase (2hours; 1:200; Amersham International, Little Chalfont,United Kingdom). Finally, a black reaction product wasformed by a short incubation (5–10 minutes) with 3,38-diaminobenidine tetrahydrochloride and nickel-ammo-nium sulphate (Shu et al., 1988). Control sections wereincubated either without the primary antiserum or withthe antiserum preabsorbed with 200 µg LHRH. In thesesections, no staining was observed (data not shown).Sections were mounted on gelatin-subbed slides and wereviewed under a Leitz Biomed microscope (Thornwood,NY).
Analysis
A Student’s t test was used to compare the sperm countsand gonadal and epididymal weights of the males and ananalysis of variance (ANOVA; with Tukey’s post-hoc test)was used to compare body weights and kidney fat in thethree groups of springbok. All statistical tests were per-formed with InStat for Macintosh (GraphPad SoftwareInc., San Diego, CA).
The number of LHRH neurones was determined for eachanimal by counting the total number of immunoreactivecell bodies in every section processed from the septalregion to the caudal hypothalamus. The area of 20 ran-domly selected (irrespective of morphological characteris-tics) immunoreactive LHRH perikarya per animal wasdetermined at the level of the organum vasculosum of thelamina terminalis (ovlt) by using an image analyzer (JoyceLoebl Magiscan) coupled to a Leica Orthoplan microscope.(Gateshead, UK). Where possible, measurements weremade on cells in which the nucleus was visible. The area ofthe immunoreactive cell body (excluding processes) wasoutlined with a light pencil. For cells in which it was notimmediately apparent where the cell body ended and aprocess began, the boundary was designated as the point
LHRH NEURONS IN THE SPRINGBOK 445

where an imaginary oval would curve round to meet theopposite side of the cell. Calculations were made by asingle observer on tissue from all 12 springboks in arandom order, without knowledge of the sex or socialstatus of the animal. Statistical comparisons were made byusing an ANOVA.
RESULTS
Reproductive status
Comparison of the body weights of the TMs (n 5 9), BMs(n 5 8), and Fs (n 5 11) indicated that Fs were signifi-cantly lighter than the TMs (P , 0.05) but not differentfrom the BMs, which had intermediate body weights(Table 1). All of the animals in the study were in goodphysical condition, as assessed by general external appear-ance and the fact that they all had reserves of fat aroundtheir kidneys. The kidney fat index did not differ betweenthe two groups of rams; however, the TMs had significantlymore kidney fat than the females (P , 0.05; Table 1). TheTM group showed signs of greater reproductive compe-tence than the BM group. Specifically, testicular weightwas significantly greater (P , 0.01) in the TMs than theBMs, as was the weight of the epididymides (P , 0.05).Although the number of sperm per epididymis was higherin the TMs compared with the BMs, this did not quiteachieve statistical significance (P 5 0.07).
None of the 11 Fs was pregnant, although two werelactating. The size and activity of the ovaries varied amonganimals, such that the ovaries of some animals appearedcompletely quiescent, whereas others had many follicleswith diameters up to 5 mm. The mean weight of theovaries was 1.2 6 0.1 g. Interestingly, the right ovary ofthis species was significantly heavier than the left ovary(right 0.71 6 0.08 g vs. left 0.5 6 0.5 g; Student’s t test;P , 0.05), which could reflect a difference in activity. Thiswas not investigated at the time. By chance, all fourfemales used in the immunocytochemical studies hadinactive ovaries characteristic of the anoestrous state,with no corpora lutea or corpora albicantia and only smallfollicles (diameter , 0.1 mm). None was lactating.
Distribution and morphology of LHRHneurones and fibres
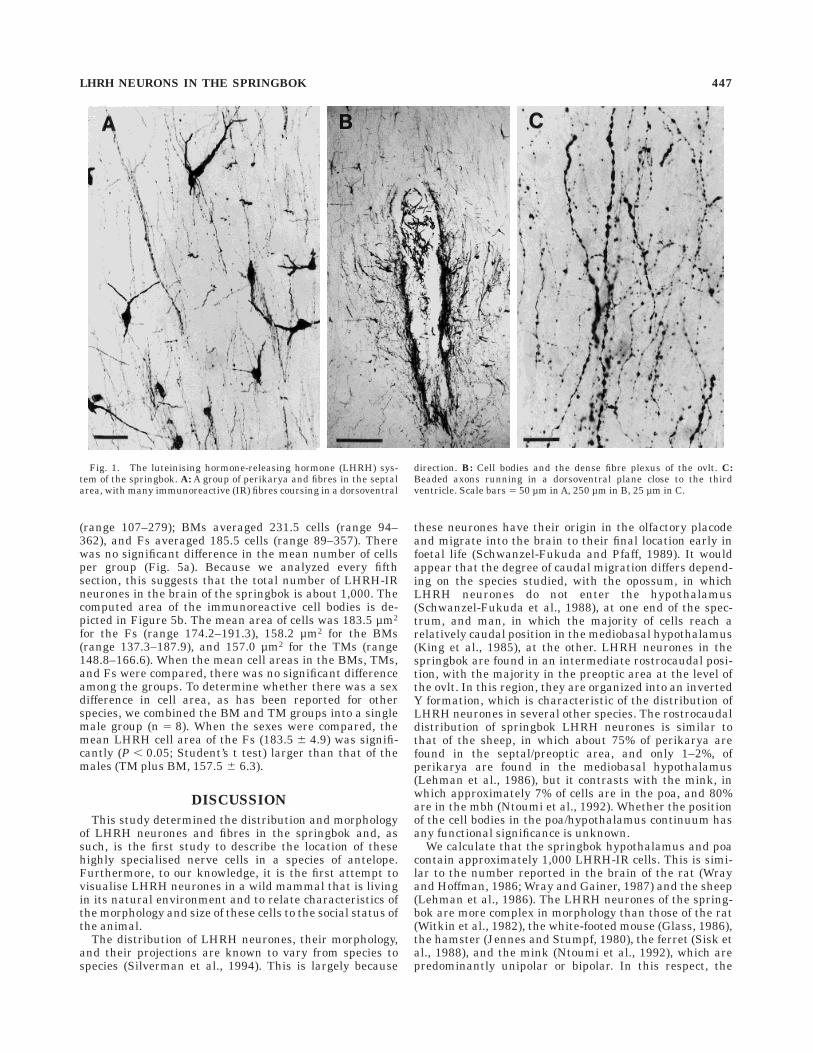
Despite the field conditions under which brain tissuewas fixed, LHRH-immunoreactive (LHRH-IR) cell bodies,terminal fields, and axons were readily visualised in all 12animals (see Figs. 1A–C, 2). LHRH-IR perikarya werelocated in a loose continuum through the septal and poa(Fig. 1A), with the majority at the level of the ovlt (Fig. 1B).In the more dorsal aspects of the hypothalamus, cells werefound close to the midline. Ventral regions showed agreater lateral distribution. In more caudal hypothalamicregions, isolated cell bodies were located in very lateralaspects of the hypothalamus, immediately dorsal to theoptic chiasm (oc) and up to 10 mm from the midline. Asmall and fairly discrete cluster of cells was found in themedian preoptic nucleus (mepon) dorsal to the anteriorcommissure, and another was found just above (but not in)the supraoptic nucleus (son). A few scattered cells werealso located in the arcuate nucleus. Immunoreactive axonscoursing away from the LHRH perikarya were readilyvisible in all animals. At the level of the septum/poa, fibrebundles coursed in a dorsoventral direction in the midline,
intermingled with cell bodies, and could sometimes befollowed for a distance of several millimetres (Fig. 1A). Inthis region, axons frequently were beaded (Fig. 1C), andsome of the beads were large (up to 5 µm diameter). Inother regions, fibres were fine and had either no or verysmall varicosities. Bordering the fornix, bundles of theseneurites could be followed for short distances and had theappearance of short strands of wavy hair. Similar thinfibres wound their way toward the median eminence (me)by a route that passed in the base of the brain over theoptic chiasm.
Extremely dense concentrations of fibres, varicosities,and terminals were found in two distinct regions; the ovltand the me. At the level of the ovlt, fibres were denselystained, heavily beaded, and intermingled with cell bodies(Fig. 1B). In the me, fibres terminated in the external layerand were particularly noticeable in the lateral portions(Fig. 2). This area is bounded by the pars tuberalis (pt) ofthe pituitary gland and, like the pt, is highly vascularized,with portal vessels passing through en route to the rostralface of the anterior pituitary gland. The location ofLHRH-IR perikarya and fibres in a representative Spring-bok is depicted in Figure 3. This distribution was similarin the males and females and, thus, was not dependant oneither the sex or the social status of the animal.
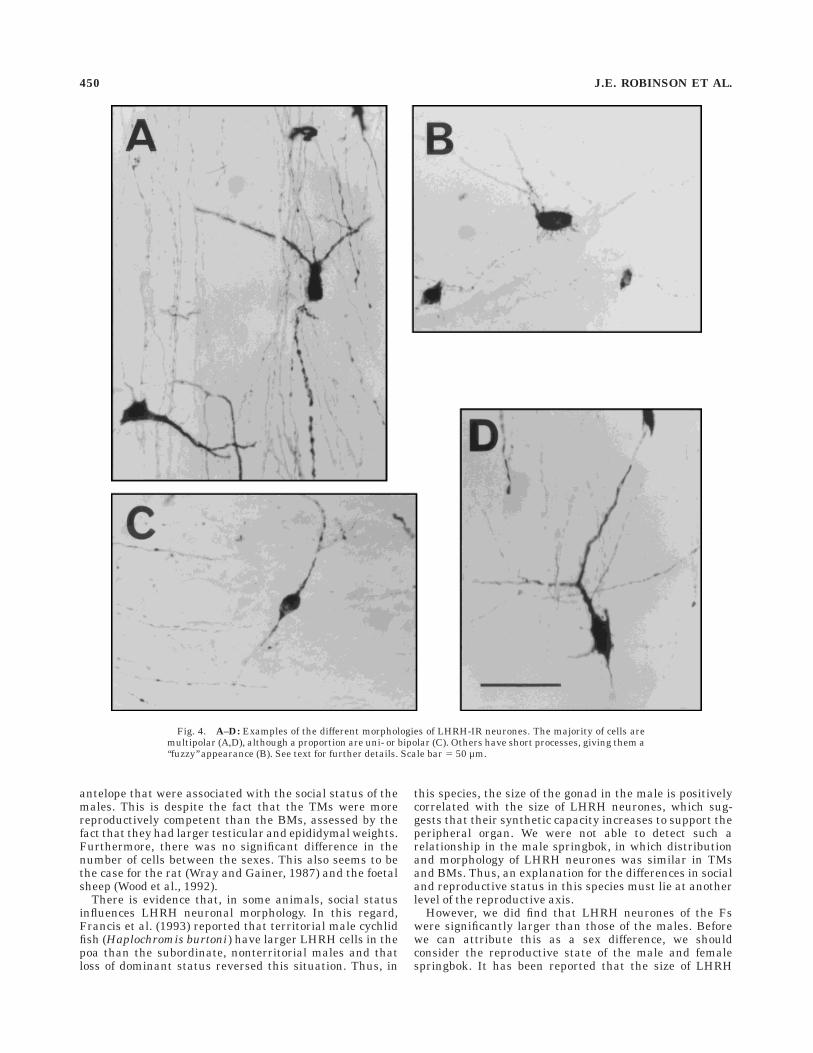
The LHRH neurones in the Springbok exhibit a varietyof different morphologies, examples of which are shown inFigure 4. In general, cells are 10–20 µm in diameter, withlarge, centrally placed nuclei surrounded by cytoplasmthat extends into a variable number of processes. Themajority (approximately 80% of total) of LHRH cells are ofcomplex morphology, with several primary dendrites ex-tending from an oval or triangular-shaped cell body (Fig.4A,D). These multipolar cells are not confined to anyparticular hypothalamic area. Bipolar cells (approxi-mately 15% of total) are also present that are fusiform inshape (Fig. 4C), and these tend to orientate themselveswith their long axes in the direction of fibre bundles. Theyare found lying in a dorsoventral plane in the septum (atthe level of Fig. 3A) but parallel to the base of the brain justabove the oc (at the level of Fig. 3D). A few unipolar cells(,5% of total) are also present. Although the vast majorityof cells in all of these categories have a smooth contour(see, e.g., Fig. 4C), some have a ‘‘fuzzy’’ appearance, withmany short protuberances projecting from the cell body(Fig. 4B). Most cells are found singly, although, in someinstances, two or three cells were found in close associationwith one another. These relationships were notable in theseptum/poa and the mepon, where neurones were ob-served with apparently overlapping cell bodies.
Number and area of LHRH-IR perikarya
There was substantial variation among animals in thenumbers of LHRH cells visualized. Specifically, the num-ber of LHRH-IR neuronees in TMs averaged 175.0 cells
TABLE 1. Body and Reproductive Measurements in the Three Groupsof Springbok (Mean 6 S.E.M.)
Territorial male Bachelor male Female
Weight (Kg) 41.1 6 2.2* 34.9 6 2.8 31.6 6 1.9*Gonadal weight (g) 77.8 6 5.7** 45.5 6 9.7** 1.2 6 0.1Sperm count 57,972 6 17,098 22,109 6 8,350 —Epididymal wt (g) 12.0 6 0.8* 8.8 6 1.5* —Kidney fat (g) 3.7 6 0.7* 2.7 6 0.7 1.5 6 0.3*
*P , 0.05. **, P , 0.01; See text for further details. Wt, weight.
446 J.E. ROBINSON ET AL.
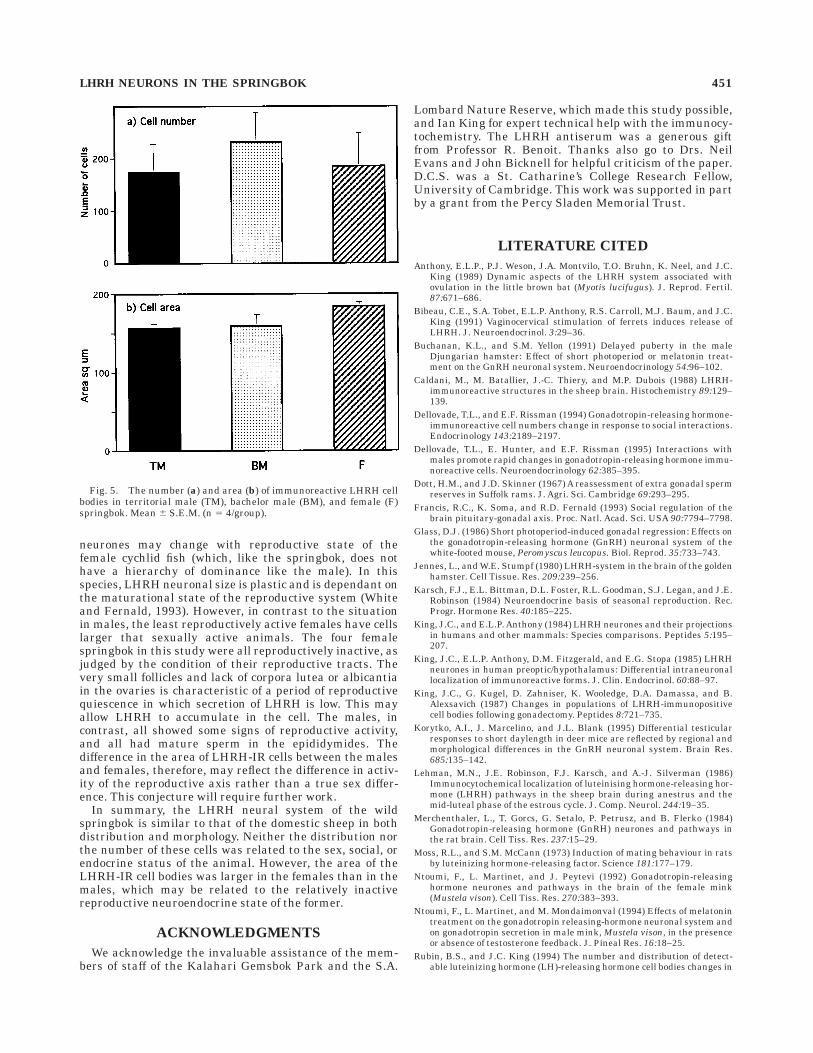
(range 107–279); BMs averaged 231.5 cells (range 94–362), and Fs averaged 185.5 cells (range 89–357). Therewas no significant difference in the mean number of cellsper group (Fig. 5a). Because we analyzed every fifthsection, this suggests that the total number of LHRH-IRneurones in the brain of the springbok is about 1,000. Thecomputed area of the immunoreactive cell bodies is de-picted in Figure 5b. The mean area of cells was 183.5 µm2
for the Fs (range 174.2–191.3), 158.2 µm2 for the BMs(range 137.3–187.9), and 157.0 µm2 for the TMs (range148.8–166.6). When the mean cell areas in the BMs, TMs,and Fs were compared, there was no significant differenceamong the groups. To determine whether there was a sexdifference in cell area, as has been reported for otherspecies, we combined the BM and TM groups into a singlemale group (n 5 8). When the sexes were compared, themean LHRH cell area of the Fs (183.5 6 4.9) was signifi-cantly (P , 0.05; Student’s t test) larger than that of themales (TM plus BM, 157.5 6 6.3).
DISCUSSION
This study determined the distribution and morphologyof LHRH neurones and fibres in the springbok and, assuch, is the first study to describe the location of thesehighly specialised nerve cells in a species of antelope.Furthermore, to our knowledge, it is the first attempt tovisualise LHRH neurones in a wild mammal that is livingin its natural environment and to relate characteristics ofthe morphology and size of these cells to the social status ofthe animal.
The distribution of LHRH neurones, their morphology,and their projections are known to vary from species tospecies (Silverman et al., 1994). This is largely because
these neurones have their origin in the olfactory placodeand migrate into the brain to their final location early infoetal life (Schwanzel-Fukuda and Pfaff, 1989). It wouldappear that the degree of caudal migration differs depend-ing on the species studied, with the opossum, in whichLHRH neurones do not enter the hypothalamus(Schwanzel-Fukuda et al., 1988), at one end of the spec-trum, and man, in which the majority of cells reach arelatively caudal position in the mediobasal hypothalamus(King et al., 1985), at the other. LHRH neurones in thespringbok are found in an intermediate rostrocaudal posi-tion, with the majority in the preoptic area at the level ofthe ovlt. In this region, they are organized into an invertedY formation, which is characteristic of the distribution ofLHRH neurones in several other species. The rostrocaudaldistribution of springbok LHRH neurones is similar tothat of the sheep, in which about 75% of perikarya arefound in the septal/preoptic area, and only 1–2%, ofperikarya are found in the mediobasal hypothalamus(Lehman et al., 1986), but it contrasts with the mink, inwhich approximately 7% of cells are in the poa, and 80%are in the mbh (Ntoumi et al., 1992). Whether the positionof the cell bodies in the poa/hypothalamus continuum hasany functional significance is unknown.
We calculate that the springbok hypothalamus and poacontain approximately 1,000 LHRH-IR cells. This is simi-lar to the number reported in the brain of the rat (Wrayand Hoffman, 1986; Wray and Gainer, 1987) and the sheep(Lehman et al., 1986). The LHRH neurones of the spring-bok are more complex in morphology than those of the rat(Witkin et al., 1982), the white-footed mouse (Glass, 1986),the hamster (Jennes and Stumpf, 1980), the ferret (Sisk etal., 1988), and the mink (Ntoumi et al., 1992), which arepredominantly unipolar or bipolar. In this respect, the
Fig. 1. The luteinising hormone-releasing hormone (LHRH) sys-tem of the springbok. A: A group of perikarya and fibres in the septalarea, with many immunoreactive (IR) fibres coursing in a dorsoventral
direction. B: Cell bodies and the dense fibre plexus of the ovlt. C:Beaded axons running in a dorsoventral plane close to the thirdventricle. Scale bars 5 50 µm in A, 250 µm in B, 25 µm in C.
LHRH NEURONS IN THE SPRINGBOK 447

springbok is more like the sheep, in which the majority ofneurones are multipolar, with fewer bi- and unipolar cells.Although this has not been quantified in adult sheep, inthe foetus, more than half of the immunoreactive neuronesare multipolar, whereas only about 15% are unipolar(Wood et al., 1992). Why such visually striking speciesdifferences should exist in the morphology of LHRH neu-rones is not clear. It could be argued that these relativelycomplex cells have greater potential to form associationswith other hypothalamic cells and thereby to alter or havetheir own activity altered by a greater number of neuralinputs. However, such morphological differences are notobviously associated with any species related variations inthe function of the LHRH neural network or the lifestyle ofthe animal. For example, seasonally breeding species inwhich LHRH neuronal activity is substantially altered byday length, like sheep and hamsters, do not have similarLHRH cell morphologies. Conversely, despite the fact thatspringbok and sheep have very similar LHRH neuralsystems, the former is aseasonal (Skinner and Smithers,1990; Skinner and Louw, 1996), whereas the latter ishighly photoperiodic (Karsch et al., 1984).
In the springbok, certain LHRH cells were found in closeapposition to one another, usually as pairs of neurones butoccasionally as small groups. Recently, Witkin and col-leagues (Witkin et al., 1995) investigated the nature of
similar associations among LHRH cells in the rat andmonkey hypothalamus. These workers describe fusion ofthe cytoplasm of such clusters of perikarya, such that theyform a syncytium, and they have speculated that such anarrangement might have important consequences in coor-dinating cells to release LHRH as a discrete bolus or pulse.Whether a pulse of LHRH is coordinated mainly at thelevel of the cell body or at the terminals in any species isstill a matter of conjecture. Certainly, there is amplepotential for LHRH neurones in the springbok to interactin the me, where a dense plexus of axons was found toterminate in the lateral borders of the external layer.Axons also appeared to terminate in the ovlt. The densityof staining in this circumventricular organ was as great asthat in the me, and this is in contrast to the situation in thesheep, in which the concentration of immunoreactivestructures was reported to be ‘‘much lower’’ in the ovlt thanin the me (Caldani et al., 1988). The function of LHRHneurones that may terminate in the ovlt is presentlyunknown, although they may contribute to the measur-able concentrations of this decapeptide in the cerebrospi-nal fluid of several species (Van Vugt et al., 1985; Skinneret al., 1995) and may play a role in reproductive function,including behaviour (Moss and McCann, 1973).
Thus, in terms of distribution and morphology, LHRHneurones in the springbok are more similar to those de-
Fig. 2. Immunoreactive LHRH axons terminating in the lateral borders of the external layer of themedian eminence (me) close to the pars tuberalis (pt). IIIv, third ventricle. Note how the terminal field ispeppered with small holes where the portal blood vessels link the brain and pituitary gland. Scale bar 5250 µm.
448 J.E. ROBINSON ET AL.

scribed in the sheep (Lehman et al., 1986; Caldani et al.,1988) than in other species that have been studied. Froman evolutionary perspective, this fits well with the relation-ships between the two animals. Both sheep and antelopebelong to the mammalian order Artiodactyla and to thefamily Bovidae. This family includes species like cattle,buffalo, and goats but not species like deer, which mightappear more like antelope but that belong to a differentfamily: the Cervidae. Studies carried out recently haveshown that the number of LHRH-IR neurones may altersubstantially depending on the social status, endocrinesituation, or sex of the species under investigation. In thisregard, the number of detectable LHRH neurones changesin specific hypothalamic regions after gonadectomy (Kinget al., 1987), at the onset of puberty in the Djungarianhamster and ferret (Yellon and Newman, 1991; Tang andSisk, 1992), and during the oestrous cycle of the littlebrown bat, ferret, and rat (Anthony et al., 1989; Bibeau etal., 1991; Rubin and King; 1994). In species in whichbreeding is restricted to a certain time of year, like deermice, white-footed mice, ferret, and Djungarian hamster,changes in the day length or melatonin administrationmay result in altered numbers of LHRH-IR cells (Glass,1986; Buchanan and Yellon, 1991; Ntoumi et al., 1994;
Korytko et al., 1995). However, not all studies have foundchanges in cell number that correlate with changes inreproductive physiology. Thus, cell number has not beenfound to change during puberty or with photoperiod in theSyrian hamster, although other aspects of cell morphologyhave (Urbanski et al., 1991, 1992). Where changes havebeen found, they may be very rapid. Thus, a recent reportindicates that sexually inexperienced female musk shrewshave increased numbers of observable LHRH neurones 30minutes after contact with male conspecifics (Dellovadeand Rissman, 1994; Dellovade et al., 1995). Changes in thenumber of LHRH-IR neurones with reproductive statemay reflect different aspects of the LHRH neural system.Although it is possible that the number of cells actuallydoes alter, it is perhaps more likely that fluctuations in therate of peptide synthesis and/or release result in differentquantities of LHRH in a cell body that alter the chance ofimmunocytochemical detection. In the current study, weobserved substantial interanimal variation in the numberof detectable LHRH neurones in the springbok. Numbersof cells per animal ranged from 89 to 362. Such variationhas also been noted in other species (Lehman et al., 1986;Wood et al., 1992). We did not observe any significantchanges in the number of LHRH-IR neurones in this wild
Fig. 3. Distribution of immunoreactive LHRH cell bodies andfibres in the springbok. A–E represent coronal sections through thepreoptic area and hypothalamus in a rostrocaudal direction; cellbodies are shown as dots, and fibres are shown as short lines. Majorlandmarks are ac, anterior commisure; fx, fornix; me, median emi-
nence; oc, optic chiasm; ot, optic tract; ovlt, organum vasculosum of thelamina terminalis; poa, preoptic area; pvn, paraventricular nucleus;son, supra optic nucleus; IIIv, third ventricle. The majority of cellbodies are in the poa at the level of the ovlt.
LHRH NEURONS IN THE SPRINGBOK 449
antelope that were associated with the social status of themales. This is despite the fact that the TMs were morereproductively competent than the BMs, assessed by thefact that they had larger testicular and epididymal weights.Furthermore, there was no significant difference in thenumber of cells between the sexes. This also seems to bethe case for the rat (Wray and Gainer, 1987) and the foetalsheep (Wood et al., 1992).
There is evidence that, in some animals, social statusinfluences LHRH neuronal morphology. In this regard,Francis et al. (1993) reported that territorial male cychlidfish (Haplochromis burtoni) have larger LHRH cells in thepoa than the subordinate, nonterritorial males and thatloss of dominant status reversed this situation. Thus, in
this species, the size of the gonad in the male is positivelycorrelated with the size of LHRH neurones, which sug-gests that their synthetic capacity increases to support theperipheral organ. We were not able to detect such arelationship in the male springbok, in which distributionand morphology of LHRH neurones was similar in TMsand BMs. Thus, an explanation for the differences in socialand reproductive status in this species must lie at anotherlevel of the reproductive axis.
However, we did find that LHRH neurones of the Fswere significantly larger than those of the males. Beforewe can attribute this as a sex difference, we shouldconsider the reproductive state of the male and femalespringbok. It has been reported that the size of LHRH
Fig. 4. A–D: Examples of the different morphologies of LHRH-IR neurones. The majority of cells aremultipolar (A,D), although a proportion are uni- or bipolar (C). Others have short processes, giving them a‘‘fuzzy’’ appearance (B). See text for further details. Scale bar 5 50 µm.
450 J.E. ROBINSON ET AL.
neurones may change with reproductive state of thefemale cychlid fish (which, like the springbok, does nothave a hierarchy of dominance like the male). In thisspecies, LHRH neuronal size is plastic and is dependant onthe maturational state of the reproductive system (Whiteand Fernald, 1993). However, in contrast to the situationin males, the least reproductively active females have cellslarger that sexually active animals. The four femalespringbok in this study were all reproductively inactive, asjudged by the condition of their reproductive tracts. Thevery small follicles and lack of corpora lutea or albicantiain the ovaries is characteristic of a period of reproductivequiescence in which secretion of LHRH is low. This mayallow LHRH to accumulate in the cell. The males, incontrast, all showed some signs of reproductive activity,and all had mature sperm in the epididymides. Thedifference in the area of LHRH-IR cells between the malesand females, therefore, may reflect the difference in activ-ity of the reproductive axis rather than a true sex differ-ence. This conjecture will require further work.
In summary, the LHRH neural system of the wildspringbok is similar to that of the domestic sheep in bothdistribution and morphology. Neither the distribution northe number of these cells was related to the sex, social, orendocrine status of the animal. However, the area of theLHRH-IR cell bodies was larger in the females than in themales, which may be related to the relatively inactivereproductive neuroendocrine state of the former.
ACKNOWLEDGMENTS
We acknowledge the invaluable assistance of the mem-bers of staff of the Kalahari Gemsbok Park and the S.A.
Lombard Nature Reserve, which made this study possible,and Ian King for expert technical help with the immunocy-tochemistry. The LHRH antiserum was a generous giftfrom Professor R. Benoit. Thanks also go to Drs. NeilEvans and John Bicknell for helpful criticism of the paper.D.C.S. was a St. Catharine’s College Research Fellow,University of Cambridge. This work was supported in partby a grant from the Percy Sladen Memorial Trust.
LITERATURE CITED
Anthony, E.L.P., P.J. Weson, J.A. Montvilo, T.O. Bruhn, K. Neel, and J.C.King (1989) Dynamic aspects of the LHRH system associated withovulation in the little brown bat (Myotis lucifugus). J. Reprod. Fertil.87:671–686.
Bibeau, C.E., S.A. Tobet, E.L.P. Anthony, R.S. Carroll, M.J. Baum, and J.C.King (1991) Vaginocervical stimulation of ferrets induces release ofLHRH. J. Neuroendocrinol. 3:29–36.
Buchanan, K.L., and S.M. Yellon (1991) Delayed puberty in the maleDjungarian hamster: Effect of short photoperiod or melatonin treat-ment on the GnRH neuronal system. Neuroendocrinology 54:96–102.
Caldani, M., M. Batallier, J.-C. Thiery, and M.P. Dubois (1988) LHRH-immunoreactive structures in the sheep brain. Histochemistry 89:129–139.
Dellovade, T.L., and E.F. Rissman (1994) Gonadotropin-releasing hormone-immunoreactive cell numbers change in response to social interactions.Endocrinology 143:2189–2197.
Dellovade, T.L., E. Hunter, and E.F. Rissman (1995) Interactions withmales promote rapid changes in gonadotropin-releasing hormone immu-noreactive cells. Neuroendocrinology 62:385–395.
Dott, H.M., and J.D. Skinner (1967) A reassessment of extra gonadal spermreserves in Suffolk rams. J. Agri. Sci. Cambridge 69:293–295.
Francis, R.C., K. Soma, and R.D. Fernald (1993) Social regulation of thebrain pituitary-gonadal axis. Proc. Natl. Acad. Sci. USA 90:7794–7798.
Glass, D.J. (1986) Short photoperiod-induced gonadal regression: Effects onthe gonadotropin-releasing hormone (GnRH) neuronal system of thewhite-footed mouse, Peromyscus leucopus. Biol. Reprod. 35:733–743.
Jennes, L., and W.E. Stumpf (1980) LHRH-system in the brain of the goldenhamster. Cell Tissue. Res. 209:239–256.
Karsch, F.J., E.L. Bittman, D.L. Foster, R.L. Goodman, S.J. Legan, and J.E.Robinson (1984) Neuroendocrine basis of seasonal reproduction. Rec.Progr. Hormone Res. 40:185–225.
King, J.C., and E.L.P. Anthony (1984) LHRH neurones and their projectionsin humans and other mammals: Species comparisons. Peptides 5:195–207.
King, J.C., E.L.P. Anthony, D.M. Fitzgerald, and E.G. Stopa (1985) LHRHneurones in human preoptic/hypothalamus: Differential intraneuronallocalization of immunoreactive forms. J. Clin. Endocrinol. 60:88–97.
King, J.C., G. Kugel, D. Zahniser, K. Wooledge, D.A. Damassa, and B.Alexsavich (1987) Changes in populations of LHRH-immunopositivecell bodies following gonadectomy. Peptides 8:721–735.
Korytko, A.I., J. Marcelino, and J.L. Blank (1995) Differential testicularresponses to short daylength in deer mice are reflected by regional andmorphological differences in the GnRH neuronal system. Brain Res.685:135–142.
Lehman, M.N., J.E. Robinson, F.J. Karsch, and A.-J. Silverman (1986)Immunocytochemical localization of luteinising hormone-releasing hor-mone (LHRH) pathways in the sheep brain during anestrus and themid-luteal phase of the estrous cycle. J. Comp. Neurol. 244:19–35.
Merchenthaler, L., T. Gorcs, G. Setalo, P. Petrusz, and B. Flerko (1984)Gonadotropin-releasing hormone (GnRH) neurones and pathways inthe rat brain. Cell Tiss. Res. 237:15–29.
Moss, R.L., and S.M. McCann (1973) Induction of mating behaviour in ratsby luteinizing hormone-releasing factor. Science 181:177–179.
Ntoumi, F., L. Martinet, and J. Peytevi (1992) Gonadotropin-releasinghormone neurones and pathways in the brain of the female mink(Mustela vison). Cell Tiss. Res. 270:383–393.
Ntoumi, F., L. Martinet, and M. Mondaimonval (1994) Effects of melatonintreatment on the gonadotropin releasing-hormone neuronal system andon gonadotropin secretion in male mink, Mustela vison, in the presenceor absence of testosterone feedback. J. Pineal Res. 16:18–25.
Rubin, B.S., and J.C. King (1994) The number and distribution of detect-able luteinizing hormone (LH)-releasing hormone cell bodies changes in
Fig. 5. The number (a) and area (b) of immunoreactive LHRH cellbodies in territorial male (TM), bachelor male (BM), and female (F)springbok. Mean 6 S.E.M. (n 5 4/group).
LHRH NEURONS IN THE SPRINGBOK 451

association with the preovulatory LH surge in the brains of young butnot middle-aged female rats. Endocrinology 134:467–474.
Schwanzel-Fukuda, M., and D.W. Pfaff (1989) Origin of luteinizing hormone-releasing hormone neurones. Nature 338:161–163.
Schwanzel-Fukuda, M., B.H. Fadem, M.S. Garcia, and D.W. Pfaff (1988)Immunocytochemical localization of luteinizing hormone-releasing hor-mone (LHRH) in the brain and nervus terminalis of the adult and earlyneonatal gray short-tailed opossum (Monodelphis domestica). J. Comp.Neurol. 276:44–60.
Shu, S., G. Ju, and L. Fan (1988) The glucose oxidase-DAB-nickel method inperoxidase histochemistry of the nervous system. Neurosci. Lett.85:169–171.
Silverman, A.-J., J.L. Antunes, G.M. Abrams, et al. (1982) The luteinizinghormone releasing hormone pathways in rhesus (Macaca mulatta) andpigtailed (Macaca nemistrina) monkeys: New observations on thick,unembedded sections. J. Comp. Neurol. 21: 309–317.
Silverman, A.-J., I. Livne, and J. Witkin (1994) The gonadotropin-releasinghormone (GnRH) neuronal systems: Immunocytochemistry and in situhybridization. In E. Knobil and J.D. Neill (eds): The Physiology ofReproduction, Second Edition. New York: Raven Press Ltd., pp. 1683–1709.
Sisk, C.L., R.L. Moss, and C.A. Dudley (1988) Immunocytochemical localiza-tion of hypothalamic luteinising hormone-releasing hormone in maleferrets. Brain Res. Bull. 20:157–161.
Skinner, D.C., and G.N. Louw (1996) The Springbok. Transvaal MuseumMonograph, 10. Johannesburg: Hortors Print.
Skinner, D.C., B. Malpaux, B. Delaleu, and A. Caraty (1995) Luteinizinghormone (LH)-releasing hormone in third ventricular cerebrospinalfluid of the ewe: Correlation with LH pulses and the LH surge.Endocrinology 136:3230–3237.
Skinner, J., and R.H.N. Smithers (1990) The Mammals of the South AfricanSubregion. Capetown, South Africa: CTP Book Printers.
Tang, Y.P., and C.L. Sisk (1992) LHRH in the ferret—Pubertal decrease inthe number of immunopositive arcuate neurones. Peptides 13:241–247.
Urbanski, H.F., A. Doan, and M. Pierce (1991) Immunocytochemicalinvestigation of luteinizing hormone-releasing hormone neurones inSyrian hamsters maintained under long or short days. Biol. Reprod.44:687–692.
Urbanski, H.F., A. Doan, M. Pierce, W.H. Fahrenbach, and P.M. Collins
(1992) Maturation of the hypothalamo-pituitary-gonadal axis of maleSyrian hamsters. Biol. Reprod. 46:991–996.
Vanrooyen, A.F. (1993) Variation in body condition of impala and nyala inrelation to social-status and reproduction. South African J. Wildlife Res.23:36–38.
Van Vugt, D.A., W.P. Diefenbach, E. Alston, and M. Ferin (1985) Gonadotro-pin-releasing hormone pulses in third ventricular cerebrospinal fluid ofovariectomized rhesus monkeys: Correlation with luteinizing hormonepulses. Endocrinology 117:1550–1558.
Van Zyl, J.H.M., and J.D. Skinner (1970) Growth and development of thespringbok foetus. African Wild Life 24:308–316.
White, S.A., and R.D. Fernald (1993) Gonadotropin-releasing hormonecontaining neurones change size with reproductive state in femalehaplochromis burtoni. J. Neurosci. 13:434–441.
Witkin, J.W., C.M. Paden, and A.-J. Silverman (1982) The luteinizinghormone-releasing hormone (LHRH) systems in the rat brain. Neuroen-docrinology 35:429–438.
Witkin, J.W., H. O’Sullivan, and A.-J. Silverman (1995) Novel associationsamong gonadotropin-releasing hormone neurones. Endocrinology 136:4323–4330.
Wood, R., S. Newman, M.N. Lehman, and D.L. Foster (1992) GnRHneurones in the fetal lamb hypothalamus are similar in males andfemales. Neuroendocrinology 55:427–433.
Wray, S., and H. Gainer (1987) Effect of neonatal gonadectomy on thepostnatal development of LHRH cell subtypes in male and female rats.Neuroendocrinology 45:413–419.
Wray, S., and G. Hoffmann (1986) A developmental study of the quantativedistribution of LHRH neurones within the central nervous system ofpostnatal male and female rats. J. Comp. Neurol. 252:522–531.
Wray, S., P. Grant, and H. Gainer (1989) Evidence that cells expressionluteinizing hormone-releasing hormone mRNAare derived from progeni-tor cells in the olfactory placode. Proc. Natl. Acad. Sci. USA 86:8132–8136.
Yellon, S.M., and S.W. Newman (1991) A developmental study of thegonadotropin-releasing hormone neuronal system during sexual matu-ration in the male Djungarian hamster. Biol. Reprod. 45:440–446.
Yellon, S.M., S.W. Newman, and M.N. Lehman (1990) The gonadotropin-releasing hormone neuronal system of the male Djungarian hamster:Distribution from the olfactory tubercle to the mediobasal hypothala-mus. Neuroendocrinology 51:219–225.
452 J.E. ROBINSON ET AL.





























