Embed Size (px)
Citation preview

D I S T R I B U T I O N AND I N H E R I T A N C E OF H E M O G L O B I N V A R I A N T S I N A M E R I C A N C A T T L E 1
D. C. SHREFFLER ~ A~D G. W. SALISBURY Department of Dairy Science, University of Illinois, Urbana
SUMMARY
The relative frequencies and genetic determination of the two electrophoretically distinguishable adult bovine henmglobin types were investigated in a large sample drawn from a number of different breeds and sources. Electrophoretic studies of hemoglobins from 121 mMes and 646 females, representing nine cattle breeds, indicate that the Ayrshire, Holstein, Angus, Hereford, and Milking Shorthorn breeds probably have only Type A hemoglobin; whereas, the Brown Swiss, Guernsey, Jersey, and Sindhi breeds have both Type A and Type B. An analysis of hemoglobin types a~mng progeny from parents of known types substantiates the theory that the occurrenee of these two types of henmglobin is genetically dete~nined by a pair of codominant alleles--HbA, determining Type A, and HbB, determining Type B. Estimates of the frequency of ttbB among the major dairy breeds are: Ayrshire--0.000, Holstein--0.000, Guernsey-- 0.116, Brown Swiss--0.117, Jersey--0.333. No gross physiological differences have been observed to be correlated with differences in hemoglobin type. The trait nmy be of some practical value in parentage determination and in the tracing of breed origins. The most significant feature of these results is that they represent a further demonstra- tion of single-gene determination of a protein specificity.
Electrophoret ical ly distinguishable hemoglobins have been observed ill the blood of cattle of North Afr ica by Cabannes and Serain (3), of Great Br i ta in by Bangham (1), and of the United States by Grimes, Duncan, and Lassiter (6) and by Sal isbury and Shreffler (12). In each instance, two types of hemoglobin have been observed which migra te at different rates in an electrical field. Three studies (1, 6, 12) indicated probable breed differences in the relative frequencies of the two hemoglobin variants, suggesting genetic determinat ion of the trait .
'i2 In the most extensive s tudy repot ted thus far , Bangham (1) presented data indicating that these two types of hemoglobin are genetically controlled by a pair of alleles at a single locus, with neither allele exhibiting any apparen t degree of dominance, so that in a heterozygous individual the blood contains a mixture of the two types of hemoglobin. Sal isbury and Shreffier (1.2) reported pre l iminary observations which seemed to suppor t this theory. The present pape r reports the results of a more extensive survey of cattle of a number of breeds and f rom a number of sources. The objective of this survey was to obtain fu r the r evidence establishing the mode of inheri tance of these hemoglobin types and to obtain estimates of the gene frequencies and genotypie distributions within breeds.
E X P E R I M E N T A L P R O C E D U R E
Animals. Included in this s tudy were 121 males and 646 females, represent- ing nine breeds and drawn f rom four experiment station herds and the two
Received for publication February 25, 1959.
1 Supported in part by funds from the Rockefeller Foundation. 2Present address: Division of Biology, California Institute of Technology, Pasadena,
California. 1147

1148 D. C. S H R E F F L E R AND G. W. S A L I S I ~ U R Y
Illinois artificial breeding centers. The breeds represented were the five major American dai ry breeds plus the Sindhi, Milking Shorthorn, Angus, and Hereford breeds. The latter three breeds were represented only by bulls f rom the arti- ficial breeding centers. The Holsteins, Guernseys, and Ayrshires studied were all in the University of Illinois herd, except 42 Holstein bulls and 21 Guernsey bulls, which were sampled at the two artificial breeding centers. Brown Swiss were sampled from the University of Illinois herd, the artificial breeding centers, the North Louisiana Hill Fa rm Exper iment Station at Homer, Louisiana, and the USDA herd at Beltsville, Maryland. Jersey bloods were obtained from the same sources as the Brown Swiss, plus the Iberia Livestock Dairy Station at Jeanerette, Louisiana. The Sindhis and Sindhi crosses were obtained from the Beltsville, Homer, and Jeaneret te stations.
Because previous observations (1, 6, 12) have indicated that the occurrence of fetal hemoglobin, Type F, in very young individuals may be confused with the adult Type B hemoglobin, no animals were included in this survey which were not at least 3 too. of age. The nature of the similarities and differences between bovine hemoglobin Types B and F has been studied in detail and reported by Grimes, Duncan, and Lassiter (7).
Hemoglobin analysis. A heparinized sample of venous blood was drawn from each animal to be tested. Seven milliliters of 0.9% NaC1 solution were added to a 1-ml. sample of the blood, the sample was centrifuged to sediment the erythro- eytes, and the supernatant was removed. The erythrocytes were washed twice more by addition of 0.9% NaC1 solution, eentrifugation, and removal of the supernatant . The washed cells were then hemo'lyzed with a quant i ty of distilled water sufficient to give a hemoglobin solution of approximately 10% concentra- tion. The prepared solutions were used either fresh or af ter storage at 0 ° F.
For analysis by electrophoresis, 0.01 ml. of the prepared hemoglobin solution was banded upon a strip of Whatman 3 MM filter paper on a Dur rum hanging strip electrophoresis apparatus (2) (manufactured by Spinco). Separations were carried out at room temperature for 3 hr. in veronal buffer, p H 8.6, ionic strength 0.05, at 8 v. per cm. In the initial phases o'f the survey, the strips were dyed in bromphenol blue solution for 6 hr., washed in 5% acetic acid, and fixed in an acetate buffer. However, in the course of the survey, it was established that with the concentrated hemoglobin solution described, the rate of migration could be accurately determined without dyeing, by examination of the visibly colored bands of hemoglobin. Therefore, for the qualitative purposes of this survey, the dyeing was discontinued.
Recent observations indicate also that, for survey work of this type, sepa- ration of the hem(~globins from the plasma proteins is unnecessary. Consistently a,~eurate results were obtained by diluting 1 vol. of whole blood with 3 vol. o~ distilled water, to hemolyze the erythroeytes, and analyzing the resulting solution by eleetrophoresis. The strips can not be dyed, but the position of the hemo- globin can be determined by its characteristic color, as previously discussed. No results were obtained which indicated interference in normal migration of the hemoglobins due to the presence of the plasma proteins.

I N H E R I T A N C E OF ~ E M O G L O B I N V A R I A N T S I N C A T T L E 1149
R E S U L T S
Three specific types of results were obtained in the analysis by paper elee- trophoresis. One type of hemoglobin, designated Type A, migrated a mean dis- tance of 1.5 cm. under the experimental conditions described. Another type, desig- nated Type B, migrated a mean distance of 2.5 cm. under these conditions. A third type of result apparently arose from the presence of both Type A and Type B hemoglobins in the blood of some individuals. This result, called Type AB, was characterized by two bands of hemoglobin, one migrating 1.5 cm., the other migrating 2.5 cm. A nlixture of equal quantities of blood containing only Type A hemoglobin and blood containing only Type B hemoglobin was found to give a similar result. The specific hemeglobin type of any individual is entirely repeatable, both for duplicate analyses of the same sample of blood and for analyses of samples taken at different times. Re-analysis of 35 adult individuals after intervals of a year or more revealed 15o instance of change in hemoglobin type.
Table 1 shows the distributions within breeds of hemoglobin types among the unselected animals analyzed. Those individuals tested as a part of the genetic study because of their presumptive or predicted hemoglobin type are excluded from these data, and only those individuals sampled at random are included. Type A hemoglobin was found among all of the breeds studied, and was found to be the sole type among the Holstein, Ayrshire, Milking Shorthorn, Angus, and Hereford individuals tested. Types B and AB were observed only in the Brown Swiss, Guernsey, Jersey, and Sindhi breeds. The 42 Ayrshires tested provide rather sound evidence that if Type B hemoglobin occurs in this breed, its frequency must be quite low. This is supported by the observations of Bangham (1), who found only Type A among 38 Ayrshire bulls, and of Grimes, Duncan, and Lassiter (6), who found only Type A among eight individuals. The absence of Type B among the large number of Holsteins tested suggests that this trait occurs very. rarely, if at all, within this breed. This is also in agreement with the results (ff Bangham (1) and Grimes, Duncan, and Las- siter (6) from smaller numbers of animals. The numbers of Shorthorn, Angus, and Hereford bulls are much too small to suggest any hemoglobin characteriza- tion of these breeds, but the results, which show the presence of Type A only, are consistent with those of Bangham (1), which included larger numbers of individuals.
The size and the variety of sources of the sample of Brown Swiss should make it possible to characterize this breed reasonably well with respect to this trait. Approximately one-fourth of the Brown Swiss have Type AB hemoglobin, while about 1% are of Type B. The remainder are Type A. Although the sample is smaller, and probably less representative, the results for the Guernseys sampled indicate a frequency of hemoglobin types similar to that found for the Bro~vn Swiss. The sample for the Jersey breed, drawn from five different sources, should also be reasonably representative of the breed. The analysis of this sample indicates a higher frequency of Types B and AB than found for the Guernsey

1150 D. C. S t t R E F F L E R AND G. W. S A L I S B U R Y
TABLE 1 Distribution of hemoglobin types within breeds
Breed
Hemoglobin type
Sex Total A AB B
Ayrshire
Brown Swiss
Guernsey
Holstein
Jersey
Sindhi
Milking Shorthorn
AberdeemAngus
Hereford
Crossbred
Holstein-Guernsey, F~ F 45 Holstein-Guernsey, other F 46
Brown Swiss-Sindhi, F~ F 14 Brown Swiss-Sindhi, other F 46
Jersey-Sindhi, F~ F 9 Jersey-Sindhi, other F 37
Holstein-Sindhi, F~ F 2 Holstein-Sindhi, other F 9
42 42 o 0
F 120 91 28 1 M 17 15 2 0
137 106 30 1
F 51 42 9 0 M 22 15 6 1
73 57 15 1
F 312 112 o 0 1~{ 43 43 0 0
155 155 0 0
F 77 41 25 11 M 13 3 7 3
90 44 32 14
F 1 0 0 1 M 1 0 0 1
,2 o ~
M 12 12 0 0
M 9 9 0 0
M 5 5 0 0
40 5 0 4~6 0 0
0 14 0 23 21 2
2 5 2 14 17 6
0 2 0 4 5 0
and B r o w n Swiss breeds. Over one-half of the i nd iv idua l s tes ted were e i ther of T y p e B or T y p e AB.
A l t h o u g h only two p u r e S indh i s were tested, both of which were Type B, some f u r t h e r ind ica t ion of f r e q u e n c y of Type B hemoglobin in this breed may be ob ta ined f r o m the da t a on crosses o'f the S i n d h i w i th the B r o w n Swiss, Je r sey , and Hols t e in breeds. These resul t s suggest tha t T y p e B hemoglobin is the pre- d o m i n a n t type in the S indh i breed, and pe rhaps the only type, since the f re- quency of T y p e B among the F1 crosses of S indh i w i th E u r o p e a n breeds is h ighe r t han tha t f o u n d in any of the E u r o p e a n breeds invo lved in the crosses. E x a m i n a t i o n of da ta on i n d i v i d u a l crosses in which the hemoglobin t y p e of the E u r o p e a n dams is known reveals t h a t in all bu t one ins tance the S indh i sires used in the crosses were T y p e B, assuming tha t the proposed t h e o r y o'f in- he r i t ance of the charac te r i s t i c is correct . I t mus t be po in ted out, however , t ha t

I N H E R I T A N C E OF H E M O G L O B I N V A R I A N T S IIN CATTLE 1151
most of the Sindhi individuals tested, or involved in the crosses the progeny of which were tested, were ra ther closely related. Thus, these observations may reflect the hemoglobin distribution in only a par t icular family or line of the breed, ra ther than the breed in general.
In connection with the two-allele theory of inheri tance of the t ra i t suggested by Bangham (1), the authors have proposed (12) that the genes controlling these hemoglobin types be designated as HbA, controlling the less rap id ly mi- grat ing Type A, and I tbB, controlling the more rap id ly migra t ing Type B. Thus, according to this nomenclature and the suggested mode of inheritance, an individual H b A / H b A would have only Type A hem(yglobin, an individual H b B / H b B would have only Type B, and a hetero'zygous individual, H b A / H b B , would have both Type A and Type B, the condition which has been designated as Type AB.
Table 2 contains the distributions of hemoglobin types among the offspring result ing f rom matings of sires and dams of known types. Included are the actual distributions observed among the offspring in each class of rantings, and the distributions expected with genetic control by a pair of codominant alleles. ,Since the presumpt ive heterozygous condition occurs both in males and in females, and since the ratio's among offspring of reciprocal crosses show no significant differences, the possibility of sex linkage is effectively eliminated. Therefore, in the data presented in Table 2, reciprocal crosses have been pooled. Chi-square and probabi l i ty values for the A × AB crosses indicate how well the observed distribution fits the expected. The numbers of animals in the AB × AB and B × AB classes are too small for any meaningful test of fit, but it will be noted that the observed and expected distributions are quite comparable in these classes. I t should also be observed that no offspring was produced in any of these crosses which was not possible under the two'-allele theory of inheritance. This is of par t icular importance in the critical tests, A × A, B × B, and A × B, in which a]] of the progeny of a given mat ing class must have identical genotypes. Larger numbers among the crosses involving Types B and AB would, of course, be desirable. The numbers are limited because the low frequency of individuals with these types among the breeds f rom which most of the data were drawn makes such matings infrequent.
In Table 3 may be found similar data for a number of instances in which the hemoglobin type of only one parent was known. While the small number of animals in the AB and B groups, and the division o'f these animals among several breeds, makes a test for the fit of the ratios of questionable reliability, values are included for a Chi-square test. The distr ibution of genotypes among the unknown parents was estimated f rom the breed gene frequencies in Table 4. Expected distr ibutions of progeny were calculated on this basis. The Chi-square and probabi l i ty values for these groups indicate that the observed distributions do not show deviations f rom the expected distributions which can not be attr ib- uted to chance. Also, it should again be observed tha t no offspring oceurre~l among these groups which could not possibly occur under the proposed mode of inheritance.

1152 D. C. SHREFFLER AND G. W. SALISBURY
T A B L E 2
D i s t r i b u t i o n of hemoglobin types among of fspr ing of p a r e n t s of known t y p e
Hemoglob in type
P a r e n t a l types Value Tota l A AB B :~2 P r o b a b i l i t y
A X A Obs. 78 78 0 0 ( H b A / H b A X H b A / H b A ) • Exp . 78 78 0 0 . . . . . . . . . . . . . . . .
A × AB Obs. 45 27 18 0 ( t t b A / H b A × H b A / H b B ) Exp . 46 23 23 0 1.778 0 . 2 0 ~ P ~ 0 . 1 0
AB × AB Obs. 3 1 2 0 ( H b A / H b B × H b A / H b B ) Exp . 4 1 2 1 . . . . . . . . . . . . . . . .
A × B Obs. 4 0 4 0 ( H b A / H b A × H b B / H b B ) Exp . 4 0 4 0 . . . . . . . . . . . . . . . .
B × AB Obs. 5 0 3 2 ( H b B / H b B × H b A / H b B ) Exp . 6 0 3 3 . . . . . . . . . . . . . . . .
B × B Obs. 5 0 0 5 ( H b B / H b B × H b B / H b B ) Exp. 5 0 0 5 . . . . . . . . . . . . . . . .
P r e s u m p t i v e genotype .
I f the theory of inheritance is correct, the phenotypic distributions of hemo- globin types shown in Table I are also genotypic distributions, and the frequen- cies of the genes controlling these types may be calculated from these data. Because there were apparent differences in the frequencies and distributions between males and females within breeds having both hemoglobin types, the distributions of males and females within these breeds were first tested separately by the Chi-square test for homogeneity of a binomial distribution (13). This test showed the separate distributions of Brown Swiss, Guernsey, and Jersey males and females to be homogeneo'us with respect to their own frequencies, although the probabili ty that the deviations in Jersey females were due to chance was very close to the 0.05 level. The differences in frequencies between males and females within breeds were then tested (13). I t was found that none of these differences was significant. Therefore, gene frequencies and the test for equilibrium of the breed populations were calculated for the pooled samples of males and females within breeds. The gene frequencies were calculated in the usual manner for a two-allele t ra i t without dominance (10), and are pre- sented in Table 4. I f random mating is assumed, the expected genotypie ar ray may be calculated from these frequencies and compared with the observed genotypie distribution in the usual test for equilibrium of a population (10). The Chi-square and probabil i ty values given in Table 4 for the three breeds in which both hemoglobin types observed frequencies from those system.
occur indicate no significant departure in the expected with random mating in a two-allele
DISCUSSION
The evidence in Tables 2 and 3 is consistent with that obtained by Bang- ham (1), upon which the theory of genetic control was originally based, and serves to support and strengthen that hypothesis. This mode of inheritance is also co~asistent with findings with respect to hemoglobin variants in humans (9), sheep (4), and mice (5), although codomiuanee in the genetic determination of mouse hemoglobins has not yet been demonstrated.

1NHERIT.kNCE OI~ HEMOGLOBIN VARIANTS IN CATTLE
TABLE 3
Distribution of hemoglobin types among offspring of one parent of known type
1153
Hemoglobin type
Parenta l type Value Total A AB B )d Probabi l i ty
A × ? Obs. 116 97 19 0 ( H b A / H b A ) a Exp. 116 101 15 0 1.225 0 . 3 0 > P ~ 0 . 2 0
AB × ? Obs. 25 6 13 6 ( H b A / t t b B ) Exp. 26 9 13 4 2.000 0 . 5 0 > P ~ 0 . 3 0
B × ? Obs. 9 0 5 4 ( I t b B / H b B ) :Exp. 9 0 5 4 0.~00 P = I . 0 0
a Presumpt ive genotype.
While it can not be considered as direct or definite evidence, the hypothesis is also supported by the fact that the tests of breed populations imply equilibrium conditions under this type of genetic control. I t must, of course, be recognized that certain factors, such as nonrandom mating, selection, and lethal effects, might influence this test for equilibrium. Three explanations may be suggested for the positive test for equilibrium: (1) The theory of inheritance is correct and, with respect to this trait, mating is relatively random with no' important lethality or selection. (2) The theory of inheritance is correct, but the effects of lethality, selection, and nonrandom mating act in different directions, in such a way that by chance the test for equilibrium is positive. (3) The theory of in- heritance is not co'rrect, but chance combinations of lethal effects, selection, and nonrandom mating have resulted in a phenotypic a r ray which fits the two-gene hypothesis.
Certainly, selection and nonrandom mating are more the rule than the excep- tion in dairy cattle breeding. However, since the trai t can be recognized only by the methods previously described, no direct selectiou based upon hemoglobin type can have been employed in the animals sampled, nor can any depar ture from random mating have been based upon this trait. Observations in the Illinois station herd have revealed no gross physiological differences within breeds between animals having different types of hemoglobin. Therefore, it does not seem likely that any extensive selectio'n for hemoglobin type due to genetic correlations with other traits has taken place. Also, if either type of hemoglobin caused lethal effects, this would be expected to result in deviations among the ratios o'f genotypes in the data in Tables 2 and 3. On the basis of these considerations, the first explanation seems the most probable, although the others can not be excluded. I t would appear, however, that the evidence presently available dictates the ac- ceptance of the hypothesis of genetic determination by a pair of eodominant alleles, unti l such time as new evidence might disprove it.
I t is not presently known whether these observations are of practical signifi- cance. These hemoglobin variants could be of some value in parentage deternfi- nation in certain breeds. Their occurrence and frequency might be useful in tracing breed origins and relationships among certain breeds. Present evidence indicates some common ancestry for the Brown Swiss, Guernsey, Jersey, and Sindhi breeds. Informat ion on the hemoglobin characterization of other Euro-
F~reed q H b B a
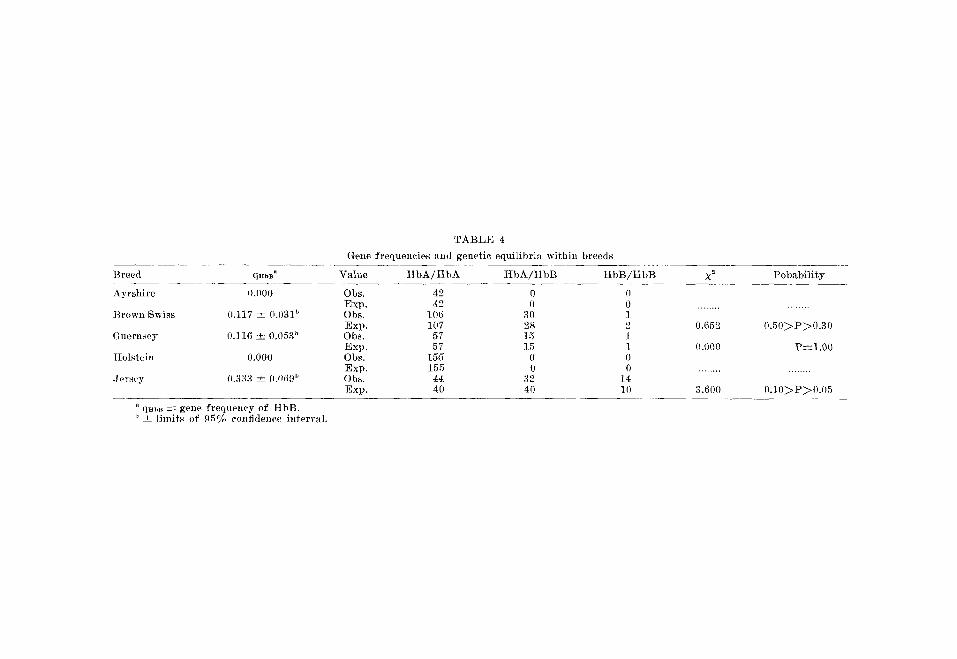
T A B L E 4
Gene f requencies and genet ic equ i l ib r i a wi th in breeds
Va lue t l bA / H b A H b A / H b B HbB/HbB X ~ P o b a b i l i t y
Ayrsh i re 0.000
Brown Swiss 0.117 _+ 0.031 ~)
Guernsey 0.] 16 _ 0.053 ~
Hols t e in 0.000
Je r sey 0.33a ~ 0.0d9 j~
Obs. 42 0 0 Exp . 42 0 0 Obs. 106 30 1 Exp . 107 2~ 2 Obs. 57 15 ] Exp . 57 15 1 Obs. 156 0 0 Exp. 155 0 0 Obs. 44 32 14 Exp . 40 40 10
0.652 0 . 5 0 ~ P ~ 0 . 3 0
0.000 P----1.00
3.600 0 . 1 0 ~ P ~ 0 . 0 5
" qHbB -~- gene f requency of HbB. ~' ± l imi t s of 95% confidence in te rva l .

INHERITANCE OF HEMOGLOBIN VARIANTS IN CATTLE 1155
pean, African, and Asian breeds could be useful in this respect. As previously stated, no gross within-breed differences in health, vigor, livability, conformation, or production, which could be related to differences in hemoglobin type, have been observed in those animals tested in the University of Illinois herd. I t is pos- sible, however, that differences could occur which would be observed only under specific physiological or environmental stresses which " t r i g g e r " some physio- l(~gieal failure, as in the case of sickle-cell and other anemias arising from hemo- globin variants in humans (14).
The quantitative investigations of Grimes, Duncan, and Lassiter (6), show- ing equal concentrations of the two hemoglobin types in what has been shown here to be the heterozygous condition, provide an interesting example o~ genetic dosage effects. Each allele apparent ly has equal efficiency with regard to the quant i ty of hemoglobin produced, but the eleetrophoretic differences indicate that the molecular structures of the two hemoglobins are slightly different. Thus, these two alleles have quantitatively equal, but qualitatively different, effects. That electrophoretic differences reflect structural differences in the hemoglobin molecule has been conclusively demonstrated by the work of Pauling et aI. (11), and the later work of h lgram (8). This difference and the simple genetic deter- mination of it are the most significant features of these results. They consti- tute fur ther evidence support ing the role of genes as determinants of protein specificities.
ACKNOWLEDGMENTS The authors wish to express their sincere appreciation to Dr. R. E. MeDowell, Dairy Cattle
Research Branch, USDA, who supplied blood samples from the USDA herd at Beltsville, Maryland, and arranged for the shipment of samples from other USDA stations. We also wish to thank Dr. F. N. Baker, North Louisiana Hill Farm Experiment Station, Homer, Louisiana, Mr. W. K. Boyd, Southern Illinois Breeding Association, Breese, Illinois, Dr. J. L, Fletcher, Iberia Livestock Dairy Station, Jeanerette, Louisiana, and Mr. W. Goeke, Northern Illinois Breeding Co-Op, Hampshire, Illinois. for their assistance in supplying blood samples from the herds under their supervision.
REFERENCES (l) BA~GHA~ A. D. Distribution of Electrophoretically Different Hemoglobins Among Cattle
Breeds of Great Britain. Nature, 179: 467. 1957. (2) BLOOK, R. J., Dtmaz~L E. L., AND ZWIFPG, G. ~ Man~al of Paper Chromatography a~d
-paper EIectropko~'esis. 2nd ed. Academic Press, New Xork. 1958. (3) CABANNE;S, R., AND SE~AIN, C. Heterogeneite de l 'Hemoglobine des Bovides, Identifica-
tion electrophoretlque de deux Hemoglobines bovines. C. R. soe. biol. -Paris, 149: 7. 1955.
(4) EVANS, J. V., ]~ING, J. W. B., CO~EN, B. L., HAaR]S, H., AN]) WAnREN, F. L. Genetics of Hemoglobin and Blood Potassium Differences in Sheep. Nature, 178: 849. 195.6.
(5) GLUF~OKSOItl~-WAELSCH, S., R.ANNEY, H. M., AND SISKEN, B. 1 ~. The Hereditary Trans- mission of Hemoglobin Differences in :Mice. J. Clin. Invest., 35: 753. 1957.
(6) GRIMEIS, R. M., DUNCAN, C. W., AND LASSITER. C. A. Occurrence of Multiple Hemo- globins in Certain Breeds of Dairy Cattle. J. Dairy Sei., 40: 1338. 1957.
(7) GlCII~IEIS, l~. M., DUNCAlg, C. W., AND LASSITE~ C. A. Bovine Fetal Hemoglobin. I. Post- natal Persistence and Relation to Adult Hemoglobins. J. D(~iry Sci., 41: 1527. 1958.
(8) INGICA]~[, V. M. Gene Mutation in Human Hemoglobin: The Chemical Difference Between Normal and Sickle Cell Haemoglobin. Nat*lre, 180: 326. 1957.

1156 D. C. SHREFFLER AND G. W. SALISBUtCY
(9) ITAN0, H. A. The Human Hemoglobins: Their Properties and Genetic Control. Advances in Protein Che, m., 12: 215. 1957.
(10) LI, C, C. Population Genetics. University of Chicago Press, Chicago. 1955. (11) PAULINe, L., IT,tNO, H. A., SINGER, S. J., AN]) W~LS, I. C. Sickle Oell Anemia, a Molec-
ular Disease. Science, 110: 543. 1949. (12) SXLISBURY, G. W., AND SHR~FFLER, D. C. Hemoglobin Variants in Dairy Cattle. J. Dairy
Sol., 40: 1198. 1957. (13) SNEDECOR, G. W. Statistical Methods. 5th ed. Iowa State College Press, Ames, Iowa.
1956. (14) ZUELZEa, W. W., NEEL, J. V., AND ]:~OBINSON, A. R. Abnormal Hemoglobins. Progress
in Hematology. Grune and Strat ton, New York. 1956.