Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Distinctive pattern of cannabinoid receptor type II (CB2)expression in adult and pediatric brain tumors
Aleksandra Ellert-Miklaszewskaa,c, Wieslawa Grajkowskab, Konrad Gabrusiewiczc,Bozena Kaminskac,⁎, Liliana Konarskaa
aDepartment of Biochemistry and Clinical Chemistry, Medical University of Warsaw, PolandbDepartment of Neurosurgery, Children's Memorial Health Institute, Warsaw, PolandcLaboratory of Transcription Regulation, Department of Cell Biology, Nencki Institute of Experimental Biology,3 Pasteur Str., 02-093 Warsaw, Poland
A R T I C L E I N F O
⁎ Corresponding author. Fax: +48 22 822 53 42E-mail address: [email protected]: CB1/CB2, Cannabinoid rec
neuroectodermal tumor; SEGA, Subependym
0006-8993/$ – see front matter © 2006 Elsevidoi:10.1016/j.brainres.2006.12.060
A B S T R A C T
Article history:Accepted 14 December 2006Available online 23 December 2006
The efficacy of cannabinoids against high-grade glioma in animal models, mediated by twospecific receptors, CB1 and CB2, raised promises for targeted treatment of themost frequentand malignant primary brain tumors. Unlike the abundantly expressed CB1, the CB2receptor shows a restricted distribution in normal brain. Although brain tumors constitutethe secondmost commonmalignancy in children and the prevalence of histological types ofbrain tumors vary significantly between the adult and pediatric populations, cannabinoidreceptor expression in pediatric tumors remains unknown. In the present study, wecompared the expression of the CB2 receptor in paraffin-embedded sections from primarybrain tumors of adult and pediatric patients. Most glioblastomas expressed very high levelsof CB2 receptors and the expression correlated with tumor grade. Interestingly, some benignpediatric astrocytic tumors, such as subependymal giant cell astrocytoma (SEGA), whichmay occasionally cause mortality owing to progressive growth, also displayed high CB2immunoreactivity. The high levels of CB2 expression would predestine those tumors to bevulnerable to cannabinoid treatment. In contrast, all examined cases of embryonal tumors(medulloblastoma and S-PNET), the most frequently diagnosed malignant brain tumors inchildhood, showed no or trace CB2 immunoreactivity. Our results suggest that the CB2receptor expression depends primarily on the histopathological origin of the brain tumorcells and differentiation state, reflecting the tumor grade.
© 2006 Elsevier B.V. All rights reserved.
Keywords:Brain tumorCannabinoid receptorGliomaImmunohistochemistryPediatric brain tumor
1. Introduction
The hemp plant-derived, endogenous and synthetic cannabi-noids elicit a wide range of central and peripheral effectsmediated mostly through cannabinoid receptors (Howlett
.(B. Kaminska).eptor type 1/type 2; GBal giant cell astrocytoma
er B.V. All rights reserved
et al., 2002). There are two types of specific Gi/o-protein-coupled receptors cloned so far, called CB1 and CB2 (Matsudaet al., 1990; Munro et al., 1993), although an existence ofadditional cannabinoid-binding receptors has been suggested(Howlett et al., 2002; Stella, 2004). CB1 is abundantly expressed
M, Glioblastoma multiforme; S-PNET, Supratentorial primitive
.

162 B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
in the central nervous system, but it is also present inperipheral nerve terminals and some extra-neural sites,such as testis, uterus, vascular endothelium, eye, spleenand tonsils (Howlett et al., 2002; Matsuda et al., 1990). Bycontrast, the CB2 receptor is primarily expressed in cells andorgans of the immune system (Howlett et al., 2002). How-ever, the CB2 receptors were recently detected in healthyneurons as well as in glial cells under inflammatory con-ditions (Gong et al., 2006; Stella, 2004; Van Sickle et al., 2005;Zhang et al., 2003).
Gliomas, the most common primary brain tumors, accountfor more than 40% of all central nervous system (CNS)neoplasms and are highly resistant to available therapeuticapproaches including radiation and chemotherapy (Galanisand Buckner, 2000; Hosli et al., 1998). Despite current efforts todevelop more effective clinical treatments against gliomas,the median survival time for patients with advanced tumorsremains approximately 1 year (Ohgaki and Kleihues, 2005).Routine therapeutic strategies for the management of allprimary brain tumors are still dependent on surgery andradiotherapy—treatments known to leave survivors withdevastating neurological deficits and frequently with a highrisk of the disease recurrence.
Anti-proliferative effects of cannabinoids have beenreported in various cancer cells in culture, includingmalignantgliomas (De Petrocellis et al., 1998; Duntsch et al., 2005;Guzman et al., 2001; Massi et al., 2004; McAllister et al., 2005;McKallip et al., 2002; Portella et al., 2003; Ruiz et al., 1999;Sanchez et al., 1998; Sarker et al., 2003). Sanchez et al. (1998)showed that (−)-trans-Δ9−tetrahydrocannabinol (Δ9-THC) inhi-bits growth of C6 glioma cells in vitro and induces cell deathwith features typical for apoptosis. More importantly, the anti-tumor actionof cannabinoids,mediated via theCB1and/orCB2cannabinoid receptors, resulted in a significant regression ofrat and human malignant gliomas in drug-treated animals(Duntsch et al., 2005; Galve-Roperh et al., 2000; Massi et al.,2004; Sanchez et al., 2001). These results prompted the clinicaltrials in patients with recurrent glioblastoma multiforme(Guzman et al., 2006). However, the well-known psychotropiceffects of Δ9-THC and related compounds, that act through theCB1 receptors, raise a number of clinical and ethical considera-tions, limiting medicinal usage of cannabinoids. One of thestrategies to avoid the side effects is administration of CB2-selective compounds.
Despite a large number of reports demonstrating anti-tumor action of synthetic cannabinoids, the studies of CBreceptor expression in astrocytic tumors are few and contra-dictory. Expression of the CB1 and CB2 cannabinoid receptorshas been detected in rat C6 glioma cells (Galve-Roperh et al.,2000) and in biopsies from human astrocytomas (Sanchezet al., 2001) using both Western blotting and immunocyto-chemistry. In the single immunohistochemical study onastrocytic tumors, Sanchez and co-workers (2001) havefound that 70% of human astrocytomas express significantlevels of both cannabinoid receptors and the extent of CB2receptor expression is related to tumor malignancy. Incontrast, Held-Feindt et al. (2006) showed that expression ofboth receptor subtypes in astroglial tumors was unrelated tomalignancy, varied between patients and was not signifi-cantly increased in relation to normal brain tissues. It is in
agreement with previous results by Bouaboula et al. (1995),who did not detect CB2 receptor mRNA in 1 GBM and 2astrocytoma tumors. Comprehensive data on the expressionof CB2 receptors in other histopathological types of primarybrain tumors are not available.
In the presentwork, we evaluated the expression of the CB2receptor in benign and malignant brain tumors in adult andpediatric patients. Although primary brain tumors constitutethe second most frequent malignancy in childhood (Kleihuesand Cavenee, 2000), this is the first report describing the CB2receptor expression in pediatric tumors. Our studies demon-strate distinct CB2 expression levels in brain tumors fromadult and pediatric patients.
2. Results
We evaluated the CB2 receptor expression in various humanbrain tumors, representing the most common histopatholo-gical types of primary brain tumors (Kleihues and Cavenee,2000) in adult and pediatric patients. The presence of CB2receptors was analyzed by immunohistochemistry on paraf-fin-embedded surgical specimens from tumors of low (gradesI–II) and high (grades III–IV) malignancy.
The specificity of the anti-human CB2 receptor antibodywas confirmed on the human tonsils sections, shown to bevery rich in this receptor (Carayon et al., 1998; Galiegue et al.,1995). We observed an intensive positive reaction in thefollicular B cell areas, mainly in germinal centers (Fig. 1A).The CB2 immunostaining in the control sections was elimi-natedbypre-absorptionof theprimary antibodywith a specificblocking peptide, thus confirming the reliability of CB2detection.
The specificity of antibody was further confirmed byWestern blot analysis using lysates of Jurkat cells (T-cellleukemia) as a positive control for CB2 expression (McKallip etal., 2002). It revealed the presence of three immunoreactivebands of approximately 59, 45 and 39 kDa (Fig. 1B). The strongband at 39 kDa, which was barely detected, when the anti-CB2antibody was pre-incubated with the blocking peptide, isconsistent with the predicted size of the CB2 protein based onits amino acid sequence. The band at 59 kDa, whose stainingwas totally abolished by pre-absorption of the antibody, aswell as the other less intensive band at 45 kDa, mightcorrespond to glycosylated forms of the CB2 receptor protein.The pattern of CB2 detection in Western analysis was similarto the one observed by Matias et al. (2002) in human dendriticcells.
Representative micrographs of CB2-expressing tumorsfrom adult and pediatric patients are presented in Fig. 2.Human astrocytic tumors, both benign such as pilomyxoidastrocytoma, juvenile pilocytic astrocytoma, SEGA (Figs. 2A,E, G) and malignant-GBM, giant cell glioblastomas (Figs. 2B,F), exhibit significant levels of CB2 expression. Low CB2expression was observed in other types of brain tumors:includsing fibroblastic meningioma, oligodendroglioma andpediatric supratentorial PNET (Figs. 2C, D, H). The CB2receptors were usually located in areas abundant in pro-liferating, invading cells. In contrast to a very strong reactionin areas of dense cellularity and microvascularization

Fig. 1 – Specificity of the anti-human CB2 receptor antibody. (A) Tonsil sections were stained as described in Experimentalprocedures. CB2-positive cells, visualized in brown using DAB, were located to germinal centers of B-cell follicles. Theimmunoreactivity was completely abolished by pre-absorption of the primary antibody on the specific blocking peptide.(B) Western blotting was performed with Jurkat cell lysates. Specific CB2 signal disappeared upon pre-incubation of theanti-CB2 antibody with the blocking peptide.
163B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
observed in GBM, staining in S-PNET section was barelyvisible. No CB2 was detected in necrotic areas (Fig. 2K). CB2immunostaining was abolished, when the primary antibodywas omitted (Fig. 2I) or pre-absorbed (Fig. 2J) with theblocking peptide.
In order to exclude the possibility of microglial infiltrationbeing responsible for the strong positive immunoreactivity inthe analyzed samples, we compared the expression patternsof the CB2 receptor and the HLA-DP,DQ,DR antigen (HLA), amarker of activated microglia, in several tumor tissues. CB2immunoreaction within the tumor was mainly attributed tothe tumor cells and not to activated microglia (Figs. 3A, B);however, the majority of HLA-positive cells showed a co-localization of both antigens in double staining experiments(Fig. 3B).
No CB2-positive reaction was observed in cells of astrocyticappearance in perilesional healthy tissue margin. However, inthe tumor surrounding areas we observed few CB2-positivecells (Figs. 3C, D). As illustrated by staining of adjacentsections for the human HLA antigen and the CB2 receptor,not all of those cells were microglia and instead, few of themresembled neurons (Fig. 3C). The double staining in anothersection showed a co-localization of CB2 and HLA in somemicroglial cells but also a strong CB2-positive reaction incertain cells in the middle layer of cerebellar cortex, mostlikely Purkinje cells (Fig. 3D).
Studying distribution and intensity of CB2 expressionwithin sections of tumors, we found that the intensity ofstaining increased with malignancy progression. Cells withanaplastic features showed high CB2 immunoreaction fadingtowards differentiated parts of the same lesion (Fig. 2D).
As shown in Table 1, among 45 tumors analyzed, 42expressed detectable levels of CB2 and in 38 cases (84%) thelevel of expression was scored asmoderate or high. Due to lowincidence of an early diagnosis in adult brain tumor patients,most of the analyzed cases were lesions of grades III and IV.Nevertheless, our results suggest that among adult braintumors, high CB2 expression characterizes the majority ofglioblastomas, particularly giant cell glioblastomas, and ismore commonly observed in high than in lower grade tumors.Remarkably, benign pediatric brain tumors (WHO grades I andII), including juvenile pilocytic astrocytoma and SEGA (sub-ependymal giant cell astrocytomas—WHO grade I), expressedmoderate or high levels of CB2, comparable to those observedin glioblastomas.
The extent of CB2 receptor expression was related totumor malignancy in astrocytic tumors and other glialneoplasms (Fig. 4). Conversely, all examined cases ofembryonal tumors, the most common malignant braintumors in childhood, composed of undifferentiated or poorlydifferentiated primitive cells, showed no or trace CB2immunoreactivity. Our results suggested that the CB2receptor expression was higher in astrocytomas/glioblasto-mas than in oligodendrogliomas, ependymomas or menin-giomas of the same grade. According to histopathologicevaluation, the case of glioblastoma with a moderate level ofCB2 expression might has developed from oligodendro-glioma. Among all studied astrocytomas (independently ofthe grade), we did not find a case that lacked the CB2receptor expression. The difference between the scored CB2expression levels in low (I+II) and high (III+ IV) gradeastrocytomas was statistically significant (p<0.01). The CB2

Fig. 2 – Expression of cannabinoid receptor type 2 (CB2) in human brain tumors from adult and pediatric patients. Detectionof the CB2 receptor in paraffin sections of brain tumors from adult (A–D) and pediatric (E–H) patientswas performed as describedin Experimental procedures using 3,3′-diaminobenzidine (DAB). Sections were counterstained with hematoxylin to visualizecell nuclei. Specimens incubated without primary antibody (I) or with the antibody pre-absorbed on the blocking peptide(J) were used as negative controls. Human astrocytic tumors, both benign such as pilomyxoid astrocytoma (A), juvenile pilocyticastrocytoma (E), SEGA (G) and malignant GBM (B), giant cell glioblastoma (F), exhibit significant levels of CB2 expression.LowCB2 expressionwas observed in other types of brain tumors: fibroblasticmeningioma (C), anaplastic oligodendroglioma (D)and pediatric supratentorial PNET (H). Note a very strong reaction in areas of dense cellularity andmicrovascularization in GBM(B and F) in contrast to barely visible staining in S-PNET (H). Abundance of CB2 receptor in poorly differentiated cells ofanaplastic oligodendroglioma (D, lower triangle) and sparse expression in cells of a less malignant appearance within distinctparts of the same section (D, upper triangle) show that CB2 expression can be related to tumor grade. Differences inimmunoreactivity between the benign juvenile pilocytic astrocytoma (E) and the malignant giant cell glioblastoma (F) are alsonoticeable. No positive CB2 reaction was detected in necrotic areas (N) surrounded by tumor cells (T) (K).
164 B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
expression in high grade primary brain tumors was judgedas significantly higher than in low grade lesions also whenother glial tumors were included in the statistical analysis(p<0.05) and even when meningiomas were added to thecompared groups (p<0.05). The embryonal tumors wereexcluded from the analysis as non-glial and non-neuronalin origin.
Additional data were obtained from the analysis of mixedglial tumors. In the single case of oligoastrocytomaand the twogangliogliomas examined, the cells of astrocytic appearanceshoweda strongCB2 immunoreactivity,while the reactionwasnegative in the oligodendroglial part of the section or inneuronal (ganglia-type) cells, respectively. Our results suggestthat the CB2 receptor expression depends primarily on the
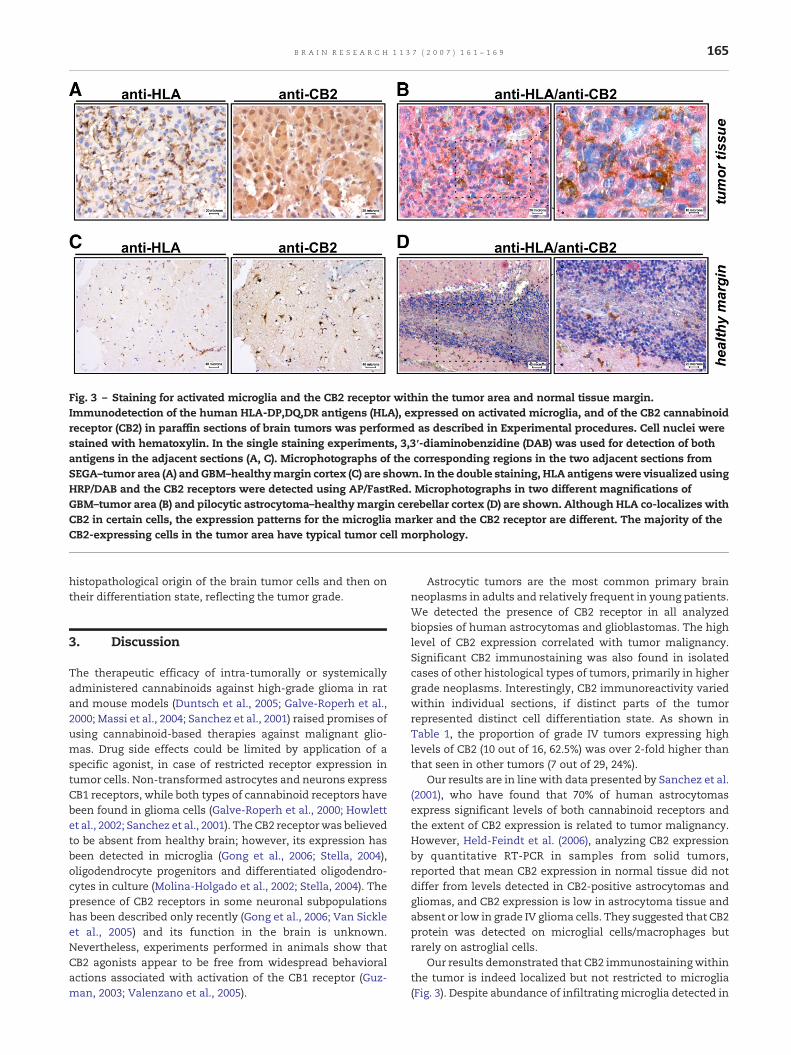
Fig. 3 – Staining for activated microglia and the CB2 receptor within the tumor area and normal tissue margin.Immunodetection of the human HLA-DP,DQ,DR antigens (HLA), expressed on activated microglia, and of the CB2 cannabinoidreceptor (CB2) in paraffin sections of brain tumors was performed as described in Experimental procedures. Cell nuclei werestained with hematoxylin. In the single staining experiments, 3,3′-diaminobenzidine (DAB) was used for detection of bothantigens in the adjacent sections (A, C). Microphotographs of the corresponding regions in the two adjacent sections fromSEGA–tumor area (A) andGBM–healthymargin cortex (C) are shown. In the double staining, HLA antigenswere visualized usingHRP/DAB and the CB2 receptors were detected using AP/FastRed. Microphotographs in two different magnifications ofGBM–tumor area (B) and pilocytic astrocytoma–healthymargin cerebellar cortex (D) are shown. Although HLA co-localizes withCB2 in certain cells, the expression patterns for the microglia marker and the CB2 receptor are different. The majority of theCB2-expressing cells in the tumor area have typical tumor cell morphology.
165B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
histopathological origin of the brain tumor cells and then ontheir differentiation state, reflecting the tumor grade.
3. Discussion
The therapeutic efficacy of intra-tumorally or systemicallyadministered cannabinoids against high-grade glioma in ratand mouse models (Duntsch et al., 2005; Galve-Roperh et al.,2000; Massi et al., 2004; Sanchez et al., 2001) raised promises ofusing cannabinoid-based therapies against malignant glio-mas. Drug side effects could be limited by application of aspecific agonist, in case of restricted receptor expression intumor cells. Non-transformed astrocytes and neurons expressCB1 receptors, while both types of cannabinoid receptors havebeen found in glioma cells (Galve-Roperh et al., 2000; Howlettet al., 2002; Sanchez et al., 2001). The CB2 receptorwas believedto be absent from healthy brain; however, its expression hasbeen detected in microglia (Gong et al., 2006; Stella, 2004),oligodendrocyte progenitors and differentiated oligodendro-cytes in culture (Molina-Holgado et al., 2002; Stella, 2004). Thepresence of CB2 receptors in some neuronal subpopulationshas been described only recently (Gong et al., 2006; Van Sickleet al., 2005) and its function in the brain is unknown.Nevertheless, experiments performed in animals show thatCB2 agonists appear to be free from widespread behavioralactions associated with activation of the CB1 receptor (Guz-man, 2003; Valenzano et al., 2005).
Astrocytic tumors are the most common primary brainneoplasms in adults and relatively frequent in young patients.We detected the presence of CB2 receptor in all analyzedbiopsies of human astrocytomas and glioblastomas. The highlevel of CB2 expression correlated with tumor malignancy.Significant CB2 immunostaining was also found in isolatedcases of other histological types of tumors, primarily in highergrade neoplasms. Interestingly, CB2 immunoreactivity variedwithin individual sections, if distinct parts of the tumorrepresented distinct cell differentiation state. As shown inTable 1, the proportion of grade IV tumors expressing highlevels of CB2 (10 out of 16, 62.5%) was over 2-fold higher thanthat seen in other tumors (7 out of 29, 24%).
Our results are in line with data presented by Sanchez et al.(2001), who have found that 70% of human astrocytomasexpress significant levels of both cannabinoid receptors andthe extent of CB2 expression is related to tumor malignancy.However, Held-Feindt et al. (2006), analyzing CB2 expressionby quantitative RT-PCR in samples from solid tumors,reported that mean CB2 expression in normal tissue did notdiffer from levels detected in CB2-positive astrocytomas andgliomas, and CB2 expression is low in astrocytoma tissue andabsent or low in grade IV glioma cells. They suggested that CB2protein was detected on microglial cells/macrophages butrarely on astroglial cells.
Our results demonstrated that CB2 immunostainingwithinthe tumor is indeed localized but not restricted to microglia(Fig. 3). Despite abundance of infiltrating microglia detected in

Table 1 – Expression level of cannabinoid receptor type 2(CB2) evaluated in adult and pediatric brain tumors
Tumor Immunoreactivityscore (CB2)
Tumor grade
I II III IV
Adult (n=20) Negative – – – –Low – – 2a 1Moderate 2b 2c 3d 2High – – 1 7
Pediatric (n=25) Negative – – – 3e
Low 2f – – –Moderate 6 2g 3h –High 2 – 4i 3
All cases were classified into groups according to diagnosed WHOgrade, regardless of the histopathological type of tumor. Numbersof cases refer to astrocytic tumors, which were the most abundant,and where indicated also include: aanaplastic oligodendroglioma(n=2); bganglioglioma (n=1); coligodendroglioma (n=1) danaplasticependymoma (n=1); emedulloblastoma (n=2) and S-PNET (n=1);ffibroblastic meningioma (n=1); gastrooligodendroglioma (n=1) andependymoma (n=1); hanaplastic ependymoma (n=2) and anaplasticganglioglioma (n=1); ianaplastic meningioma (n=1), anaplasticganglioglioma (n=1) and anaplastic ependymoma (n=1). Immuno-reactivity was scored as negative, low,moderate or high, dependingon the intensity of DAB staining.
Fig. 4 – CB2 immunoreactivity in various brain tumors frompediatric and adult patients in relation to tumor grade.Different histopathological types of tumors are representedby separate symbols. Oligodendroglial, ependymal,ganglioglial and mixed glial tumors are combined as“other glial tumors”. Despite a strong CB2 immunoreactivityamong astrocytic tumors, a correlation between malignancyand CB2 expression level is still clearly visible in thisgroup. The difference in the CB2 immunoreactivity betweenlow- and high-grade tumors is statistically significant(*astrocytic tumors: p<0.01, all glial tumors: p<0.05, all glialand meningeal tumors: p<0.05). No or hardly detectedexpression of the CB2 cannabinoid receptor in embryonaltumors, such as medulloblastoma and S-PNET, is observed.
166 B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
some sections, the majority of CB2-positive cells weretransformed glial cells. Ramified and activated microglia iscommonly detected within astrocytic tumors and the level ofresident tumor microglia seems to correlate with the tumorgrade (Badie and Schartner, 2001; Roggendorf et al., 1996). Inanaplastic astrocytomas and glioblastomas, microglia mayconstitute up to 30% of all tumor cells (Badie and Schartner,2001). Moreover, the CB2 receptor mRNA and protein expres-sion has been also detected in human cerebromicrovascularendothelial cells (HBEC) (Golech et al., 2004), which asconstituents of blood vessels may affect the total level ofCB2 expression in the highly vascularized tumor tissue.
From that perspective, the lack of CB2 mRNA in high gradeglioma in quantitative RT-PCR studies by Held-Feindt et al.(2006) is surprising. One explanation for the observed dis-crepancy is that a specific gene expression in tumor tissuesample may be underestimated due to dilution of the realmRNA level or protein expressed in certain cell population, ina mass of a variety of other cells, thus showing the advantageof in situ detection methods. Higher stability of the CB2receptor protein than mRNA may be another reason. In fact,Bouaboula et al. (1995) did not detect CB2 receptor mRNA in 1GBM and 2 astrocytoma tumors. However, regulation of CB2gene expression and its transcript processing is poorly under-stood (Onaivi et al., 2002) and no data on CB2 mRNA stabilityare currently available.
The specificity of the anti-human CB2 antibody used forimmunohistochemical staining was tested by us (Fig. 1) andothers (Gong et al., 2006; Matias et al., 2002; Sanchez et al.,2001). We showed that the positive reaction on tonsilsections and in Western blot with Jurkat cells extract isspecific to CB2 receptor. Pre-incubation with the specificblocking peptide diminished the immunohistochemicalstaining in control tonsil and tumor sections and the CB2signal in Western blot.
Our results support a notion that CB2 expression isassociated with tumor malignancy. However, we found thatbenign pediatric brain tumors, such as subependymal giantcell astrocytomas SEGA, also express high levels of CB2. SEGAsare distinctive brain tumors that are commonly associatedwith tuberous sclerosis complex (Shepherd et al., 1991).Although histologically benign, they cause both morbidityand occasional mortality owing to progressive growth in somepatients. The high levels of CB2 expression would predestinethose tumors to cannabinoid treatment.
Among the grade IV neoplasms, the three cases with noCB2 immunoreactivity were embryonal tumors, composed ofundifferentiated or poorly differentiated primitive cells withthe capacity to differentiate along neuronal, astrocytic,ependymal, muscular or melanocytic lines (Kleihues andCavenee, 2000). The analyzed cases of embryonal tumorsincluded S-PNET and medulloblastomas; in fact the latter areknown to reveal features of predominantly neuronal differ-entiation. Thus, the CB2 receptor expression seems to dependprimarily on the histopathological origin of the tumor andthen on its grade.
Sanchez et al. (2001) observed no difference in thedistribution of cannabinoid receptors between pure astrocy-tomas and mixed oligoastrocytomas. However, we found thatin the case of mixed glial (oligoastrocytoma) or mixed

167B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
neuronal-glial tumors (ganglioglioma) the CB2 expression washigher in astrocytic compartment than in the other types oftransformed neural cells. Concordantly, we observed that theCB2 staining intensity was much higher in astrocytomas thanin oligodendrogliomas, ependymomas or meningiomas of thesame grade, showing again a strong relationship between CB2expression and tumor histopathology. To our knowledge thisis the first report comparing the CB2 expression in variousprimary brain tumors.
Our results concerning glial tumors, as well as recentlypublished array of data, show that increased CB2 receptorlevels are found in dynamically proliferating tumor cells. Acorrelation between CB2 expression and histologic grade ofthe tumor has been reported in other types of tumors, and thelevel of CB2 expression in transformed cells was higher thanin the respective normal tissue (Caffarel et al., 2006; Carracedoet al., 2006; Sarfaraz et al., 2005). Elevated endocannabinoidlevels were found in glioblastoma tissue as compared to non-tumor brain tissue, suggesting that endocannabinoid signal-ing may play a role in negative control of cell divisions(Petersen et al., 2005).
A strong inhibition of glioma growth after intra-tumoralinjection of natural or synthetic cannabinoids, including a CB2agonist (Galve-Roperh et al., 2000; Sanchez et al., 2001), andthe efficacy of systemically administered cannabinoidsagainst high-grade human glioma in mouse xenograft models(Duntsch et al., 2005; Massi et al., 2004), resulted in trials inhumans (Guzman et al., 2006). McAllister et al. (2005) describedcell death of cultured human glioblastoma cells exposed to Δ9-THC or WIN 55,212-2. Others demonstrated that cannabidiol(Massi et al., 2004), JWH133 (Sanchez et al., 2001) or a novelcannabinoid KM-233 (Duntsch et al., 2005) blocked theproliferation of human astrocytoma cells implanted subcuta-neously to immunodeficient mice. We have reported anapoptotic death triggered by WIN 55,212-2 in rat glioma cells(Ellert-Miklaszewska et al., 2005) and our recent studies showeffectiveness of WIN 55,212-2 and CB2-selective JWH133 toinduce apoptosis of cultured human glioblastoma cells andtumor-derived primary cultures (Ellert-Miklaszewska et al.,submitted for publication).
It is worthmentioning that administration of non-selectivecannabinoids, which are more effective in the induction ofapoptosis (our unpublished observation), could be consideredin pediatric patients. Studies in pre- and postnatal lifedemonstrated the gradual increase of the level of anandamideand its CB1 receptors in childhood, accompanied with devel-opment of susceptibility to the psychoactive action of exogen-ous cannabinoids (Fride, 2004). Thus, possible side effects maybe minor at young age, which has been observed in twoindependent case studies (Fride, 2004). Thus, expression of theCB2 receptor among pediatric glial tumors may be of greatimportance for future clinical applications of cannabinoids.
4. Experimental procedures
4.1. Materials
The anti-human-CB2 receptor antibody and a specific blockingpeptide corresponding to amino acid sequence of the N
terminus of the receptor (20–33 aa) were purchased fromCayman Chemical Company. The anti-human HLA-DP,DQ,DRantigen antibody, hematoxylin, LSAB+ Peroxidase Kit, EnVi-sion Doublestain System and DAB (3,3′-diaminobenzidine)were from DakoCytomation. Streptavidin–peroxidase conju-gate was from Beckman Coulter. Nitrocellulose membraneand enhanced chemiluminescence detection system (ECL)were from Amersham Pharmacia Biotech. The horseradishperoxidase-conjugated anti-rabbit IgG were from Cell Signal-ling. The biotinylated anti-mouse IgG antibody and all otherreagents, unless otherwise stated, were purchased from SigmaAldrich.
4.2. Tumor samples characteristics
Sections of human brain tumors pre-embedded in paraffinwere retrieved from the files of the Neurosurgery Departmentof Children's Memorial Health Institute in Warsaw, MedicalUniversity of Warsaw and The Central Clinical Hospital of theMinistry of Internal Affairs in Warsaw. Diagnosis as well asgrading of the tumors according to the WHO classification(Kleihues and Cavenee, 2000) was confirmed by histologicalstaining of sections with hematoxylin and eosin (H&E).
The tumor types examined included:
- tumors from adult (>18 years old) patients (n=20): glio-blastoma multiforme (GBM) GIV (n=8), giant cell glioblas-toma GIV (n=2), anaplastic astrocytoma GIII (n=3),pilomyxoid astrocytoma GI/II (n=1), pleomorphic xantho-astrocytoma GII (n=1), anaplastic oligodendroglioma GIII(n=2), oligodendroglioma GII (n=1), anaplastic ependy-moma GIII (n=1) and ganglioglioma GI (n=1);
- tumors from pediatric (≤18 years old) patients (n=25):glioblastoma GIV (n=2), giant cell glioblastoma GIV (n=1),anaplastic astrocytoma GIII (n=1), juvenile pilocytic astro-cytoma GI (n=8), subependymal giant cell astrocytoma(SEGA) GI (n=1), oligoastrocytoma GII (n=1), anaplasticganglioglioma GIII (n=2), anaplastic ependymoma GIII(n=3), ependymoma GII (n=1), anaplastic meningiomaGIII (n=1), fibroblastic meningioma GI (n=1), supratentorialprimitive neuroectodermal tumor (S-PNET) GIV (n=1) andmedulloblastoma GIV (n=2).
4.3. Immunohistochemistry
Immunostaining was performed on 5-μm sections of paraffin-embedded tumor tissues. The sections were deparaffinized inxylene, hydrated with a descending ethanol series and rinsedwith deionized water. Endogenous peroxidase activity wasblocked for 20 min with 3% hydrogen peroxide. For eachwashing 0.05 M Tris-buffered saline, pH 7.6, was used. In orderto retrieve the antigen, sections were boiled for 30 min incitrate buffer, pH 6.0. In the single immunostaining for can-nabinoid receptor 2 (CB2) detection, the sections were thenincubated overnight at 4 °C with anti-human-CB2 receptorantibody, diluted 1:300 in 1% bovine serum albumin in Tris-buffered saline. After incubation with the primary antibody,the subsequent steps were performed using LSAB+ PeroxidaseKit. To stain activated microglia, the anti-human HLA-DP,DQ,DR antigen antibody diluted 1:150 was used and the reaction

168 B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
was followed by consecutive incubations with biotinylatedanti-mouse secondary antibody and streptavidin-peroxidaseconjugate. In case of both antigens, peroxidase activity wasrevealed by chromogen 3,3′-diaminobenzidine (DAB) (10 min)and then counterstaining was performed with hematoxylin.Finally, sectionswere dehydrated through ethanols, cleared inxylene and mounted with coverslips using Canada balsam.
Double staining experiments were carried out using EnVi-sion Doublestain System, according to the manufacturer'sprotocol. After antigen retrieval step and blocking of theendogenous peroxidase, the sections were incubated withanti-human HLA-DP,DQ,DR antigen antibody (diluted 1:150).The labeling was revealed with peroxidase (HRP)-conjugatedpolymer (that also carries antibodies to mouse and rabbitimmunoglobulins) and DAB as a chromogen. Following ablocking step using the Doublestain Block reagent, thesections were incubated with the CB2 receptor antibodyusing aforementioned dilutions and next with alkalinephosphatase (AP)-conjugated polymer. The alkaline phospha-tase activity was visualized with Fast-Red and then counter-staining was performed with hematoxylin. Finally, thesections were washed in ammonia water and mounted withcoverslips using Kaiser's Glycerol-Gelatine (Merk).
Primary antibodies were omitted from the staining proce-dure as a routine in a set of control sections. In addition, theCB2 primary antibody pre-absorption experiments werecarried out with the specific blocking peptide (1:20 antibody:peptide ratio). The specificity of CB2 detection was confirmedon human tonsils sections, where high immunoreactivity hasbeen previously described (Carayon et al., 1998; Galiegue et al.,1995).
4.4. Western blot analysis
Western blots were performed according to the previouslydescribed protocol (Ellert-Miklaszewska et al., 2005). Briefly,whole-cell protein extracts from Jurkat cells were resolved onSDS-PAGE before electrophoretic transfer onto a nitrocellulosemembrane. After blocking in 5% low-fat milk in TBS-T (0.1%Tween 20/Tris-buffered saline, pH 7.6), the membranes wereincubated overnight with the CB2 antibody diluted 1:1000 inblocking buffer. For negative controls the CB2 antibody waspre-absorbed for 1 h at 37 °C on the specific blocking peptide(1:20 antibody:peptide ratio) and then used in the aforemen-tioned dilution. The primary antibody reaction was followedby 1-h incubation with horseradish peroxidase-conjugatedanti-rabbit secondary antibody. Immunocomplexes weredetected using an enhanced chemiluminescence detectionsystem and membrane exposure to X-ray film (Kodak). Themolecular weight of proteins was estimated with pre-stainedprotein markers (Sigma).
4.5. Semi-quantitative scoring system and statisticalanalysis
All tumor sections were evaluated and scored by a neuro-pathologist and one or two independent investigators, whowere blinded to the tissue slides. Staining intensity and CB2distribution patterns were analyzed in the whole section butimmunoreactivity scoring was restricted to tumor area (three
to five representative fields under 10×40 magnification).Immunoreactivity was scored as 1=negative, 2=low, 3=mod-erate or 4=high, depending on both the intensity of DABstaining and the percentage of CB2-positive tumor cells. Allcases were classified into groups according to diagnosedWHOgrade. The resultant scores were compared between thegroups of low (grades I and II) and high (grades III and IV)malignancy using the non-parametrical Mann–Whitney Urank sum test. Statistical analysis was performed usingSTATISTICA 7.1 (StatSoft, Inc., USA). Differences were con-sidered statistically significant for p values <0.05.
Acknowledgments
This work was supported by grant from the State Committeefor Scientific Research (Poland) No. 3P05F02224 and grant No.06/2002 from the Polish Pharmacy and Medicine DevelopmentFoundation by Polpharma S.A.We thankMrs. D. Giedronowicz(Department of Pathomorphology, Institute of Tuberculosisand Pulmonary Diseases, Warsaw) for her excellent tech-nical help with immunohistochemistry analysis. A. Ellert-Miklaszewska is a recipient of a scholarship from the Post-graduate School of Molecular Medicine.
R E F E R E N C E S
Badie, B., Schartner, J., 2001. Role of microglia in glioma biology.Microsc. Res. Tech. 54, 106–113.
Bouaboula, M., Bourrie, B., Rinaldi-Carmona, M., Shire, D., Le Fur,G., Casellas, P., 1995. Stimulation of cannabinoid receptor CB1induces krox-24 expression in human astrocytoma cells. J. Biol.Chem. 270, 13973–13980.
Caffarel, M.M., Sarrio, D., Palacios, J., Guzman, M., Sanchez, C.,2006. Delta9-tetrahydrocannabinol inhibits cell cycleprogression in human breast cancer cells through Cdc2regulation. Cancer Res. 66, 6615–6621.
Carayon, P., Marchand, J., Dussossoy, D., Derocq, J.M., Jbilo, O.,Bord, A., Bouaboula, M., Galiegue, S., Mondiere, P., Penarier, G.,Fur, G.L., Defrance, T., Casellas, P., 1998. Modulation andfunctional involvement of CB2 peripheral cannabinoidreceptors during B-cell differentiation. Blood 92, 3605–3615.
Carracedo, A., Gironella, M., Lorente, M., Garcia, S., Guzman, M.,Velasco, G., Iovanna, J.L., 2006. Cannabinoids induce apoptosisof pancreatic tumor cells via endoplasmic reticulumstress-related genes. Cancer Res. 66, 6748–6755.
De Petrocellis, L., Melck, D., Palmisano, A., Bisogno, T., Laezza, C.,Bifulco, M., Di Marzo, V., 1998. The endogenous cannabinoidanandamide inhibits human breast cancer cell proliferation.Proc. Natl. Acad. Sci. U. S. A. 95, 8375–8380.
Duntsch, C., Divi, M.K., Jones, T., Zhou, Q., Krishnamurthy, M.,Boehm, P., Wood, G., Sills, A., Ii, B.M., 2005. Safety and efficacyof a novel cannabinoid chemotherapeutic, KM-233, for thetreatment of high-grade glioma. J. Neuro-oncol. 1–10.
Ellert-Miklaszewska, A., Kaminska, B., Konarska, L., 2005.Cannabinoids down-regulate PI3K/Akt and Erk signallingpathways and activate proapoptotic function of Bad protein.Cell. Signal. 17, 25–37.
Fride, E., 2004. The endocannabinoid-CB(1) receptor system inpre- and postnatal life. Eur. J. Pharmacol. 500, 289–297.
Galanis, E., Buckner, J.C., 2000. Chemotherapy of brain tumors.Curr. Opin. Neurol. 13, 619–625.
Galiegue, S., Mary, S., Marchand, J., Dussossoy, D., Carriere, D.,

169B R A I N R E S E A R C H 1 1 3 7 ( 2 0 0 7 ) 1 6 1 – 1 6 9
Carayon, P., Bouaboula, M., Shire, D., Le Fur, G., Casellas, P.,1995. Expression of central and peripheral cannabinoidreceptors in human immune tissues and leukocyte subpopu-lations. Eur. J. Biochem. 232, 54–61.
Galve-Roperh, I., Sanchez, C., Cortes, M.L., del Pulgar, T.G.,Izquierdo, M., Guzman, M., 2000. Anti-tumoral action ofcannabinoids: involvement of sustained ceramideaccumulation and extracellular signal-regulated kinaseactivation. Nat. Med. 6, 313–319.
Golech, S.A., McCarron, R.M., Chen, Y., Bembry, J., Lenz, F.,Mechoulam, R., Shohami, E., Spatz, M., 2004. Human brainendothelium: coexpression and function of vanilloid andendocannabinoid receptors. Brain Res. Mol. Brain Res. 132,87–92.
Gong, J.P., Onaivi, E.S., Ishiguro, H., Liu, Q.R., Tagliaferro, P.A.,Brusco, A., Uhl, G.R., 2006. Cannabinoid CB2 receptors:immunohistochemical localization in rat brain. Brain Res.1071, 10–23.
Guzman, M., 2003. Cannabinoids: potential anticancer agents. Nat.Rev. Cancer 3, 745–755.
Guzman, M., Sanchez, C., Galve-Roperh, I., 2001. Control of the cellsurvival/death decision by cannabinoids. J. Mol. Med. 78,613–625.
Guzman, M., Duarte, M.J., Blazquez, C., Ravina, J., Rosa, M.C.,Galve-Roperh, I., Sanchez, C., Velasco, G., Gonzalez-Feria, L.,2006. A pilot clinical study of Delta9-tetrahydrocannabinol inpatients with recurrent glioblastoma multiforme. Br. J. Cancer95, 197–203.
Held-Feindt, J., Dorner, L., Sahan, G., Mehdorn, H.M., Mentlein, R.,2006. Cannabinoid receptors in human astroglial tumors.J. Neurochem. 98, 886–893.
Hosli, P., Sappino, A.P., de Tribolet, N., Dietrich, P.Y., 1998.Malignant glioma: should chemotherapy be overthrown byexperimental treatments? Ann. Oncol. 9, 589–600.
Howlett, A.C., Barth, F., Bonner, T.I., Cabral, G., Casellas, P.,Devane, W.A., Felder, C.C., Herkenham, M., Mackie, K., Martin,B.R., Mechoulam, R., Pertwee, R.G., 2002. International union ofpharmacology. XXVII. Classification of cannabinoid receptors.Pharmacol. Rev. 54, 161–202.
Kleihues, P., Cavenee, W.K., 2000. Pathology and Genetics ofTumours of the Nervous System. IARC Press, Lyon.
Massi, P., Vaccani, A., Ceruti, S., Colombo, A., Abbracchio, M.P.,Parolaro, D., 2004. Antitumor effects of cannabidiol, anonpsychoactive cannabinoid, on human glioma cell linesJ. Pharmacol. Exp. Ther. 308, 838–845.
Matias, I., Pochard, P., Orlando, P., Salzet, M., Pestel, J., Di Marzo, V.,2002. Presence and regulation of the endocannabinoid systemin human dendritic cells. Eur. J. Biochem. 269, 3771–3778.
Matsuda, L.A., Lolait, S.J., Brownstein, M.J., Young, A.C., Bonner,T.I., 1990. Structure of a cannabinoid receptor and functionalexpression of the cloned cDNA. Nature 346, 561–564.
McAllister, S.D., Chan, C., Taft, R.J., Luu, T., Abood, M.E., Moore,D.H., Aldape, K., Yount, G., 2005. Cannabinoids selectivelyinhibit proliferation and induce death of cultured humanglioblastoma multiforme cells. J. Neuro-oncol. 74, 31–40.
McKallip, R.J., Lombard, C., Fisher, M., Martin, B.R., Ryu, S., Grant,S., Nagarkatti, P.S., Nagarkatti, M., 2002. Targeting CB2cannabinoid receptors as a novel therapy to treat malignantlymphoblastic disease. Blood 100, 627–634.
Molina-Holgado, E., Vela, J.M., Arevalo-Martin, A., Almazan, G.,Molina-Holgado, F., Borrell, J., Guaza, C., 2002. Cannabinoidspromote oligodendrocyte progenitor survival: involvement ofcannabinoid receptors and phosphatidylinositol-3 kinase/Aktsignaling. J. Neurosci. 22, 9742–9753.
Munro, S., Thomas, K.L., Abu-Shaar, M., 1993. Molecularcharacterization of a peripheral receptor for cannabinoids.Nature 365, 61–65.
Ohgaki, H., Kleihues, P., 2005. Population-based studies onincidence, survival rates, and genetic alterations in astrocyticand oligodendroglial gliomas. J. Neuropathol. Exp. Neurol. 64,479–489.
Onaivi, E.S., Leonard, C.M., Ishiguro, H., Zhang, P.W., Lin, Z.,Akinshola, B.E., Uhl, G.R., 2002. Endocannabinoids andcannabinoid receptor genetics. Prog. Neurobiol. 66, 307–344.
Petersen, G., Moesgaard, B., Schmid, P.C., Schmid, H.H., Broholm,H., Kosteljanetz, M., Hansen, H.S., 2005. Endocannabinoidmetabolism in human glioblastomas and meningiomascompared to human non-tumour brain tissue. J. Neurochem.93, 299–309.
Portella, G., Laezza, C., Laccetti, P., De Petrocellis, L., Di Marzo, V.,Bifulco, M., 2003. Inhibitory effects of cannabinoid CB1 receptorstimulation on tumor growth and metastatic spreading:actions on signals involved in angiogenesis and metastasis.FASEB J. 17, 1771–1773.
Roggendorf, W., Strupp, S., Paulus, W., 1996. Distribution andcharacterization of microglia/macrophages in human braintumors. Acta Neuropathol. (Berl.) 92, 288–293.
Ruiz, L., Miguel, A., Diaz-Laviada, I., 1999. Delta9-tetrahydrocan-nabinol induces apoptosis in human prostate PC-3 cells via areceptor-independent mechanism. FEBS Lett. 458, 400–404.
Sanchez, C., Galve-Roperh, I., Canova, C., Brachet, P., Guzman, M.,1998. Delta9-tetrahydrocannabinol induces apoptosis in C6glioma cells. FEBS Lett. 436, 6–10.
Sanchez, C., de Ceballos, M.L., del Pulgar, T.G., Rueda, D., Corbacho,C., Velasco, G., Galve-Roperh, I., Huffman, J.W., Ramon y Cajal,S., Guzman, M., 2001. Inhibition of glioma growth in vivo byselective activation of the CB(2) cannabinoid receptor. CancerRes. 61, 5784–5789.
Sarfaraz, S., Afaq, F., Adhami, V.M., Mukhtar, H., 2005.Cannabinoid receptor as a novel target for the treatment ofprostate cancer. Cancer Res. 65, 1635–1641.
Sarker, K.P., Biswas, K.K., Yamakuchi, M., Lee, K.Y., Hahiguchi, T.,Kracht, M., Kitajima, I., Maruyama, I., 2003. ASK1-p38 MAPK/JNK signaling cascademediates anandamide-induced PC12 celldeath. J. Neurochem. 85, 50–61.
Shepherd, C.W., Scheithauer, B.W., Gomez, M.R., Altermatt, H.J.,Katzmann, J.A., 1991. Subependymal giant cell astrocytoma: aclinical, pathological, and flow cytometric study. Neurosurgery28, 864–868.
Stella, N., 2004. Cannabinoid signaling in glial cells. Glia 48,267–277.
Valenzano, K.J., Tafesse, L., Lee, G., Harrison, J.E., Boulet, J.M.,Gottshall, S.L., Mark, L., Pearson, M.S., Miller, W., Shan, S.,Rabadi, L., Rotshteyn, Y., Chaffer, S.M., Turchin, P.I., Elsemore,D.A., Toth, M., Koetzner, L., Whiteside, G.T., 2005.Pharmacological and pharmacokinetic characterization of thecannabinoid receptor 2 agonist, GW405833, utilizing rodentmodels of acute and chronic pain, anxiety, ataxia andcatalepsy. Neuropharmacology 48, 658–672.
Van Sickle, M.D., Duncan, M., Kingsley, P.J., Mouihate, A., Urbani,P., Mackie, K., Stella, N., Makriyannis, A., Piomelli, D., Davison,J.S., Marnett, L.J., Di Marzo, V., Pittman, Q.J., Patel, K.D., Sharkey,K.A., 2005. Identification and functional characterization ofbrainstem cannabinoid CB2 receptors. Science 310, 329–332.
Zhang, J., Hoffert, C., Vu, H.K., Groblewski, T., Ahmad, S.,O'Donnell, D., 2003. Induction of CB2 receptor expression in therat spinal cord of neuropathic but not inflammatory chronicpain models. Eur. J. Neurosci. 17, 2750–2754.










![P-ISSN2349–8528 Molecular modeling studies and structure ...Bockaert and Pin., 1999) [5, 4] and There are two distinct cannabinoid receptors, CB1 and CB2, have been identified in](https://img.pdfslide.us/doc/110x75/60efdc26f1ad6e375a074656/p-issn2349a8528-molecular-modeling-studies-and-structure-bockaert-and-pin.jpg)