Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Nov. 2008, p. 4959–4967 Vol. 76, No. 110019-9567/08/$08.00�0 doi:10.1128/IAI.00664-08Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Diminished ICAM-1 Expression and Impaired Pulmonary Clearance ofNontypeable Haemophilus influenzae in a Mouse Model of Chronic
Obstructive Pulmonary Disease/Emphysema�
Bing Pang,1 Wenzhou Hong,1† Shayla L. West-Barnette,1‡ Nancy D. Kock,2 and W. Edward Swords1*Departments of Microbiology and Immunology,1 and Pathology and Comparative Medicine,2
Wake Forest University Health Sciences, Winston-Salem, North Carolina
Received 28 May 2008/Returned for modification 2 July 2008/Accepted 3 September 2008
The airways of patients with chronic obstructive pulmonary disease (COPD) are continually colonized withbacterial opportunists like nontypeable Haemophilus influenzae (NTHi), and a wealth of evidence indicates thatchanges in bacterial populations within the lung can influence the severity of COPD. In this study, we used amurine model for COPD/emphysema to test the hypothesis that COPD affects pulmonary clearance. Mice weretreated with a pulmonary bolus of elastase, and as reported previously, the lungs of these mice were patho-logically similar to those with COPD/emphysema at �1 month posttreatment. Pulmonary clearance of NTHiwas significantly impaired in elastase-treated versus mock-treated mice. While histopathologic analysis re-vealed minimal differences in localized lung inflammation between the two groups, lower levels of intercellularadhesion molecule 1 (ICAM-1) were observed for the airway epithelial surface of elastase-treated mice than forthose of control mice. Following infection, elastase-treated mice had lung pathology consistent with pneumoniafor as long as 72 h postinfection, whereas at the same time point, mock-treated mice had cleared NTHi andshowed little apparent pathology. Large aggregates of bacteria were observed within damaged lung tissue of theelastase-treated mice, whereas sparse individual bacteria were observed in lungs of mock-treated mice at thesame time point postinfection. Additional infection studies showed that NTHi mutants with biofilm defectswere less persistent in the elastase-treated mice than the parent strain. These findings establish a model forCOPD-related infections and support the hypotheses that ICAM-1 promotes clearance of NTHi. Furthermore,the data indicate that NTHi may form biofilms within the context of COPD-related infections.
Chronic obstructive pulmonary disease (COPD) is a pro-gressive lung disease that includes emphysema, chronic bron-chitis, and bronchiectasis and is among the leading publichealth problems worldwide (8, 34). COPD affects over 10 mil-lion adults in the United States alone (22) and is the fourthleading cause of death in the United States (13). The estimatedtotal economic impact of COPD in the United States is over$20 billion/year (44). While the primary cause of COPD issmoking or exposure to other inhaled pollutants, the progres-sion and severity of COPD may be promoted by opportunisticairway infections (37). While this has been an area of somecontroversy (9), it is undeniable that the management ofCOPD is, at best, seriously complicated by bacterial and viralinfections (31). The agents that cause COPD-related airwayinfections are found predominantly within the normal flora ofthe nasopharynx and include nontypeable Haemophilus influ-enzae (NTHi), Streptococcus pneumoniae, Moraxella catarrhalis,and Pseudomonas aeruginosa (25, 26, 37, 38). Patients may alsobe chronically infected with H. haemolyticus, which does notseem to be a significant pathogen but is associated with asymp-tomatic carriage (27). In patients with COPD, carriage of theseorganisms is not limited to the nasopharynx, as is the case in
healthy patients, but extends into the upper and lower airways(23, 24, 29).
The composition of the bacterial population within theCOPD lung is extremely dynamic, with individual strains/clonesexhibiting variable persistence and with incoming strains sup-planting other strains (35–39). Patients with COPD can be col-onized by several different strains simultaneously, and thelength of persistence varies dramatically between the differentstrains (30). Patient studies have demonstrated that somestrains of NTHi can persist within individual patients formonths or even years and that exacerbations of COPD aresignificantly correlated with the acquisition of a new bacterialstrain (35, 36, 39), apparently independently of the bacterialload (39) but in accordance with host-pathogen interactionsthat may be specific to the individual bacterial strain (3). No-tably, most animal models for pulmonary infection fail tomimic the degree of bacterial persistence observed for humanpatients. For example, it is well established that mice thatreceive a pulmonary infection of NTHi reproducibly clear theinfection within 4 days postinoculation (33, 49, 50). There is aneed for an animal model that better reflects the persistentinfections that occur in the context of COPD.
One of the hallmarks of COPD/emphysema is tissue destruc-tion by elastase released by neutrophils within the lung, result-ing in pulmonary fibrin deposition and decreased lung volume(41, 42). In prior studies, COPD-like conditions have beenestablished in mice by chronic exposure to cigarette smoke(40–42) or by the introduction of a bolus of elastase into thelung (15, 20, 32). In the latter model, mice treated with elastase
* Corresponding author. Mailing address: 5101A Gray Building,Medical Center Boulevard, Winston-Salem, NC 27157. Phone: (336)713-5049. Fax: (336) 716-9928. E-mail: [email protected].
† Present address: Department of Otolaryngology, Medical Collegeof Wisconsin, Milwaukee, WI.
‡ Present address: NIDCD/NIH, Bethesda, MD.� Published ahead of print on 15 September 2008.
4959
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

exhibited lung damage consistent with COPD, including tissuedestruction within the lung, enlargement of airspaces, and fi-brotic deposits within the lung alveolar spaces. We reasonedthat this model system could provide a better way to examinebacterial clearance from the COPD lung. Therefore, we com-pared bacterial clearance following pulmonary infection withNTHi in mice treated with elastase with that of controlstreated with vehicle (phosphate-buffered saline [PBS]) alone.The results show that clearance of NTHi from the lung wassignificantly impaired following elastase treatment, in accor-dance with the formation of large bacterial communities thatwere not observed for the control mice. Furthermore, immu-nohistochemical analysis revealed diminished expression of in-tercellular adhesion molecule 1 (ICAM-1) on the airway epi-thelial surfaces of the elastase-treated mice following infectioncompared to that of the control groups. We thus conclude thatpulmonary infections in mice with normal clearance may notfully represent the host-pathogen interactions that determinethe outcome of infections in COPD and that the outcome of apulmonary bacterial infection in this setting may be deter-mined by the interplay of host clearance events initiated byICAM-1 expression on the airway epithelium and by bacterialpersistence mechanisms that may include biofilm formation.
MATERIALS AND METHODS
Bacterial strains and growth conditions. All NTHi strains were cultivated onsupplemented brain-heart infusion (Difco) medium supplemented with NAD(Sigma) and hemin (ICN), as described previously (45–48). NTHi 2019 is awell-characterized strain that was originally isolated from the sputum of a patientwith chronic bronchitis (2), and all of the mutant strains were derived from thisstrain background. A list of bacterial strains, along with primary references andphenotypes, is provided in Table 1.
Elastase treatment. Healthy C57BL/6 mice were purchased from CharlesRiver Laboratories (Wilmington, MA). Mice were anesthetized with Avertin(2,2,2-tribromoethanol), and a 50-�l bolus of elastase (Sigma) suspended insterile PBS was instilled intratracheally into the lung. The amount of elastaseused was determined by dose-response experiments as the minimal amount ofenzyme necessary to generate lung damage consistent with COPD. Various dosesof elastase (1 to 9 units) were intratracheally instilled into mice (five/group), andthe mice were then euthanized 21 days posttreatment. Histopathologic analysisrevealed tissue fibrosis and reduction in airway space, consistent with COPD, at3, 6, and 9 U of elastase (data not shown), with no apparent pathology in micethat received vehicle (PBS) alone (Fig. 1 and 2). As 3 U of elastase was theminimal dose needed to elicit COPD-like pathology, this was the amount chosenfor infection studies. Animals were allowed to recover for 21 days after elastasetreatment, before histopathology and/or infection studies were performed.
Infections. NTHi bacteria were harvested from overnight plate cultures andsuspended in PBS. Bacterial counts were estimated by optical density and sus-pended in PBS solution as described previously (45). The estimated bacterialdensity was confirmed by plate count. Approximately 107 CFU was used to infectmice (five animals/group). The mice were anesthetized as described before andinfected intratracheally, and the bacterial load in the inocula was confirmed byplate count. At the times indicated, mice were euthanized, and their lungs were
excised. For each animal, the left lung was homogenized and used for platecount. Plate count data were analyzed by unpaired t test analysis with Welch’scorrection for unequal variance; groups with P values of less than 0.05 weredeemed significantly different from the control. The right lungs were fixed in 4%paraformaldehyde-PBS for histopathology and cryosection. The elastase treat-ment and infection protocols were approved by the Wake Forest UniversityHealth Sciences animal care and use committee.
Histopathology. Portions of fixed lung tissue were dehydrated and embeddedin paraffin according to standard methods. Sections (5 �m) were cut fromparaffin-embedded blocks with a microtome and mounted from warm water(40°C) onto adhesive microscope slides. After serial deparaffinization and rehy-dration, tissue sections were stained with hematoxylin and eosin for histopatho-logic assessment. Stained slides were provided as a blinded set to a veterinarypathologist (N.K.) and were scored for markers of inflammation (neutrophilicinflux, edema, etc.).
Immunostaining. To determine ICAM-1 expression, paraffin sections werestained using monoclonal antibody 3E2, recognizing mouse ICAM-1 (BD Phar-mingen) according to the manufacturer’s instructions, essentially as reported by
TABLE 1. NTHi strains used in this study
NTHi Strain Description Referenceor source
2019 Bronchial isolate 22019 siaB strain Sialylated mutant 162019 htrB strain Lipid A acylation defect 212019 pgmB strain Phosphoglucomutase mutant 452019 licD strain Phosphorylcholine-negative mutant 452019 licON strain Constitutive phosphorylcholine-
positive mutant11
FIG. 1. Elastase-treated mice have fibrotic lung damage consistentwith COPD. Mice were treated by nonsurgical intratracheal instillationof elastase (see Materials and Methods) and allowed to recover for 21days posttreatment. Lung tissue samples from euthanized mice receiv-ing vehicle (PBS) (A), elastase (B), or heat-inactivated (HI) elastase(C) were compared by hematoxylin-eosin staining of paraffin sections.COPD-like pulmonary damage was observed for the elastase-treatedgroup that was not observed for either of the control groups. Magni-fication, �4 for left panels and �40 for right panels.
FIG. 2. Impaired pulmonary clearance of NTHi from elastase-treatedmice. Mice (five/group) were infected via the intratracheal route with�107 CFU of NTHi 2019 and euthanized at the indicated times postin-fection (see Materials and Methods). Symbols are CFU counts obtainedfrom homogenized lung tissue from mock-treated (filled circles) or elas-tase-treated mice (open circles). The dotted line indicates the lower limitof detection, and symbols below that line indicate a mouse from which nobacteria were recovered. Asterisks indicate groups in which bacterialnumbers were significantly higher than those in controls, as assessed bynonparametric statistical analysis.
4960 PANG ET AL. INFECT. IMMUN.
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

others (6). For visualization of NTHi bacteria, portions of fixed lung tissuesamples were rinsed with 1� PBS at room temperature and placed into aCryomold (Sakura Finetek USA, Torrance, CA). Tissue-Tek OCT compound(Sakura Finetek USA, Torrance, CA) was added, and the blocks were frozen at�70°C for 1 h. Serial 5-�m sections were cut using an Accu-Edge low-profileblade (Feather Safety Razor Co., Japan) at �20°C and stored at �70°C. Immu-nofluorescence staining was performed using rabbit antisera recognizing NTHi,essentially as described previously (33). For both sets of sections, pixel quanti-zation was performed for all tissue sections and is presented as the mean num-bers of pixels for ICAM-1 or bacterial staining.
RESULTS
Pulmonary elastase treatment results in damage consistentwith COPD/emphysema and impaired bacterial clearance. Toelicit pulmonary fibrotic damage consistent with COPD, micewere treated with a pulmonary bolus of elastase, vehicle (PBS),or heat-inactivated elastase delivered via nonsurgical intratra-cheal instillation. After 21 days, mice in each group were eu-thanized, and their lungs were sectioned and stained with he-matoxylin and eosin for histopathologic analysis. The resultsclearly showed significant pulmonary damage in the elastase-treated mice, whereas there was minimal damage observed forthe control groups (Fig. 1).
The effect of this treatment on the clearance of NTHi strain2019 from the lung was determined with pulmonary infectionstudies using elastase-treated mice and mock-treated mice.While bacterial counts obtained from lung homogenates werecomparable at an early time point (6 h postinfection), signifi-cantly higher numbers of bacteria were recovered from theelastase-treated mice at 24 h, 48 h, and 72 h postinfection (Fig.2). In the control group, the numbers of bacteria declinedsignificantly over time, and the majority of mice had pulmonarybacterial loads below the threshold of detection by 72 h postin-
fection. Thus, we concluded that mice treated with elastase hadimpaired pulmonary clearance of NTHi.
Histopathologic analysis of lung tissue from infected mice.Lung tissue from each group of mice was embedded in paraffin,sectioned, and stained for histopathologic assessment (as de-scribed in Materials and Methods). Stained slides were exam-ined as a blinded set by a veterinary pathologist (N.K.) andscored for parameters of airway inflammation, which werecompiled into a total score for inflammation. Figure 3 showsrepresentative images from each group at the different timepoints postinfection. Notably, there was a dramatic loss ofalveolar lung tissue, along with fibrosis, observed for the elas-tase-treated mice even prior to infection (Fig. 3C to E). Lo-calized tissue responses, including edema and cellular infiltratethat included neutrophils, were observed for both groups ofanimals. Total histopathology scores were compiled for eachgroup of sections as blinded sets. The only differences in scor-ing occurred with samples from the latest time point (72 hpostinfection), where inflammation remained notable in theelastase-treated group. In contrast, all indicators of inflamma-tion were decreased in the mock-treated group at this time-point (Fig. 3F to G). As shown in Fig. 2, elastase-treated micefailed to clear NTHi bacteria from the lung within 72 h postin-fection. For the mock-treated mice, epithelial cells were ob-served sloughed into a bronchiolar lumen, and the interstitiumwas mildly infiltrated by neutrophils. In contrast, the sectionsfrom the elastase-treated mice showed severe pneumonia withmarked infiltration of neutrophils, causing consolidation ofthe lung.
Histopathology scores. The stained sections were examinedas a blinded set and assigned scores (1–4) for markers ofinflammation. A composite score (1–10) was compiled based
FIG. 3. Histopathologic assessment of lung tissue from infected animals. Lung tissue was sectioned and stained for histopathology analysis asdescribed in Materials and Methods. Each panel is a portion of a representative hematoxylin-eosin-stained section from the indicated group(Control or Elastase group, left margin) at a given time point (Mock, 6 h, 24 h, 48 h, or 72 h).
VOL. 76, 2008 IMPAIRED BACTERIAL CLEARANCE IN MOUSE MODEL FOR COPD 4961
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

on the total scores. The results are depicted in Fig. 4. A sig-nificant increase in vascular degeneration was observed in theelastase treatment group early after infection (Fig. 4A). Bron-chial epithelial responses peaked later and to a greater degreein the elastase treatment group (Fig. 4B), as was observed forpneumonia (Fig. 4C), alveolar macrophages (Fig. 4D), and
airway inflammation (Fig. 4E). Similarly, the composite in-flammation scores were higher for the elastase treatmentgroup at the later time points postinfection (Fig. 4F). No sig-nificant differences were noted in the number of total lympho-cytes (data not shown). Taken together, these results indicatea slower pulmonary inflammatory response that reached
FIG. 4. Histopathology scores. Hematoxylin-eosin-stained sections from mock-treated (open bars) and elastase-treated (filled bars) mice wereexamined as a blinded set and scored for markers of pulmonary inflammation (see Materials and Methods).
4962 PANG ET AL. INFECT. IMMUN.
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

higher levels later after infection for the elastase treatmentgroup.
Diminished surface expression of ICAM-1 in elastase-treatedmice. ICAM-1 is expressed on the airway epithelial surfaceunder many different pulmonary inflammatory conditions andserves to facilitate neutrophil recruitment into the lung (4, 7,51, 52). Surface expression of ICAM-1 has been demon-strated to promote NTHi clearance from mouse lung (6),although there are also indications that NTHi may utilizeICAM-1 as a receptor for attachment to epithelial surfaces(1). Therefore, we determined whether ICAM-1 expressionwas altered in the elastase-treated mice. Paraffin sections oflung tissue from infected mice were examined by immuno-staining for ICAM-1. Figure 5 shows representative imagesfrom light microscopic examination of tissue from eachgroup of animals, along with quantization of ICAM-1 stain-ing as a percentage of total pixels in all sections. In the
control group, ICAM-1 expression increased significantly asearly as 6 h postinfection and remained elevated above thebaseline throughout the infection study. However, the levelof expression in the elastase-treated animals was markedlylower at all time points and did not show an increase until 24to 48 h postinfection. Therefore, based on these data, weconclude that ICAM-1 expression is diminished and tempo-rally changed in the elastase-treated mice.
Presence of multicellular NTHi bacterial communitieswithin the lungs of elastase-treated mice. To visualize NTHibacteria within the lungs of infected mice, cryosections wereprepared from mock-treated and elastase-treated mice at thevarious time points after infection and stained with polyclonalrabbit antisera directed against NTHi. For mock-treated ani-mals, limited reactivity was observed that correlated in sizewith individual bacteria dispersed throughout the lung tissuetaken 48 h postinfection (Fig. 6A). However, in the elastase-
FIG. 5. Experimental COPD reduces the level and kinetics of immunohistochemical staining for ICAM-1 expression within lung tissue duringinfection. ICAM-1 levels in paraffin sections of lung tissue were assessed by immunohistochemical staining with monoclonal antibody 3E2(BD/Pharmingen) as described in Materials and Methods. Sections were counterstained with hematoxylin. Panels show representative sectionsfrom the groups indicated at left. Graphs depict total ICAM-1 staining in all sections examined in the group as a percentage of total pixels.Magnification, �20 for left panels and �40 for right panels.
VOL. 76, 2008 IMPAIRED BACTERIAL CLEARANCE IN MOUSE MODEL FOR COPD 4963
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

treated animals, larger regions of reactivity were visible indiscrete locations within the lung tissue at this time point (Fig.6B). Quantization of the fluorescence from microscopy imagesfrom all infection groups showed an �40-fold increase in bac-terial density within the lung tissue of elastase-treated mice atthis time point (Fig. 6). Examination of sections at a highermagnification revealed that these regions were more than 10�m in diameter, which correlates in size with a multicellularcommunity of NTHi bacteria (Fig. 6C). Moreover, as clearlyvisible by differential interference contrast/Nomarski imaging,the NTHi communities were present in damaged regions of thelung with fibrotic deposits. Paired histopathologic staining ofserial sections immediately adjacent to those stained for im-munofluorescence revealed the presence of neutrophils sur-rounding many of these communities (Fig. 6C).
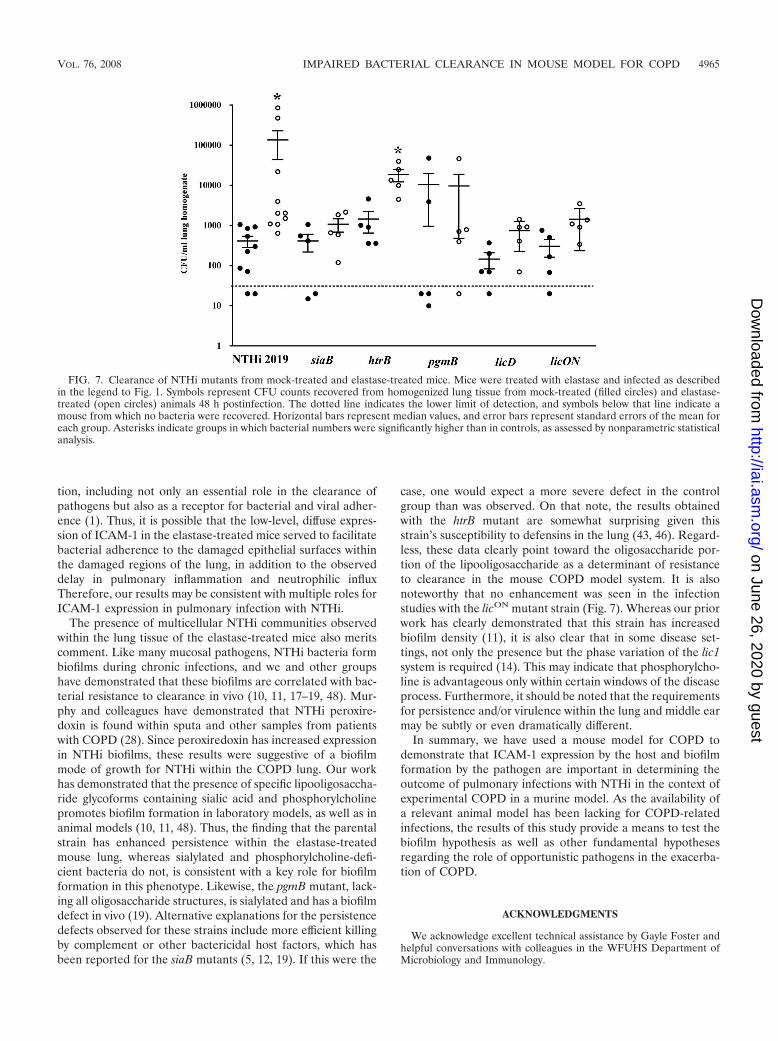
Infection studies using mutant NTHi strains. To furtherclarify the role(s) of specific persistence-related surface moi-eties in NTHi persistence within the elastase-treated mice, weperformed infection studies using a panel of mutant strains(Fig. 7). For the purpose of comparison, we chose the 48-htime point postinfection, as bacteria were consistently recov-ered from both the control and elastase-treated mice withmaximal differences at this time point. As in the initial studiesshown in Fig. 1, significantly higher numbers of NTHi 2019bacteria were recovered from elastase-treated mice. However,the counts from control and elastase-treated mice infected witha sialylated (siaB) mutant strain were indistinguishable. Simi-larly, counts from mice infected with a “rough” mutant lackingthe oligosaccharide portion of the LOS moiety (pgmB mutant)or with mutants with altered expression of phosphorylcholine-positive lipooligosaccharides (licD and licON mutants, the lat-ter harboring an in-frame deletion of the CAAT repeat region
in licA), were indistinguishable between the control and elas-tase-treatment groups. However, mice infected with the NTHi2019 htrB strain, which has an underacylated lipid A, hadsignificantly higher resistance to clearance in the elastase-treated group than in the control group, similar to the parentalstrain. It is notable that all of the mutations that affected thepersistence of the elastase treatment group affected the oligo-saccharide portion of the lipooligosaccharides on the NTHisurface. Therefore, based on these data, we conclude thatmoieties contained within the carbohydrate portion of the en-dotoxins on the NTHi surface promote persistence within ela-stase-treated mice. The implications of these data for therole(s) of biofilms in the increased persistence phenotype willbe further outlined in the discussion.
DISCUSSION
While it is clear that patients with COPD/emphysema haveincreased susceptibility to many respiratory pathogens, thereremains a need for a better understanding of the mechanismsfor this susceptibility. In this study, we adapted an existingmodel of COPD for infection studies. The results clearly showthat following pulmonary treatment with elastase to elicit aCOPD-like condition, mice had a significant clearance defectfor NTHi bacteria compared to mock-treated mice. This clear-ance defect was correlated with a delayed expression ofICAM-1 on the airway epithelial surfaces, a host response thatpromotes the influx of neutrophils and the resolution of NTHiinfection (6). It is notable that the basal level of ICAM-1expression observed for the elastase-treated mice was signifi-cantly higher than that observed for mock-treated mice.ICAM-1 appears to play several roles in NTHi and viral infec-
FIG. 6. Immunofluorescent staining reveals multicellular bacterial communities within lung tissue of elastase-treated animals. Lung tissue wascryosectioned (see Materials and Methods) and stained with rabbit anti-H. influenzae sera (53) and fluorescent antibody conjugate (JacksonLaboratories). Panels A and B show merged differential interference-contrast/fluorescent images from a mock-treated animal (A) and elastase-treated animal (B) 48 h postinfection. The graph depicts quantified bacterial staining as the percentage of total pixels and was obtained fromsections of tissue from all animals. Panels in C show sequential sections from the same tissue block stained to show the distribution of bacteria(fluorescent image marked “NTHi”) and cellular infiltrate within the lung (light microscopy image marked “H & E”). Magnification, �20 for leftset of panels in C and �100 for right set of panels.
4964 PANG ET AL. INFECT. IMMUN.
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
tion, including not only an essential role in the clearance ofpathogens but also as a receptor for bacterial and viral adher-ence (1). Thus, it is possible that the low-level, diffuse expres-sion of ICAM-1 in the elastase-treated mice served to facilitatebacterial adherence to the damaged epithelial surfaces withinthe damaged regions of the lung, in addition to the observeddelay in pulmonary inflammation and neutrophilic influxTherefore, our results may be consistent with multiple roles forICAM-1 expression in pulmonary infection with NTHi.
The presence of multicellular NTHi communities observedwithin the lung tissue of the elastase-treated mice also meritscomment. Like many mucosal pathogens, NTHi bacteria formbiofilms during chronic infections, and we and other groupshave demonstrated that these biofilms are correlated with bac-terial resistance to clearance in vivo (10, 11, 17–19, 48). Mur-phy and colleagues have demonstrated that NTHi peroxire-doxin is found within sputa and other samples from patientswith COPD (28). Since peroxiredoxin has increased expressionin NTHi biofilms, these results were suggestive of a biofilmmode of growth for NTHi within the COPD lung. Our workhas demonstrated that the presence of specific lipooligosaccha-ride glycoforms containing sialic acid and phosphorylcholinepromotes biofilm formation in laboratory models, as well as inanimal models (10, 11, 48). Thus, the finding that the parentalstrain has enhanced persistence within the elastase-treatedmouse lung, whereas sialylated and phosphorylcholine-defi-cient bacteria do not, is consistent with a key role for biofilmformation in this phenotype. Likewise, the pgmB mutant, lack-ing all oligosaccharide structures, is sialylated and has a biofilmdefect in vivo (19). Alternative explanations for the persistencedefects observed for these strains include more efficient killingby complement or other bactericidal host factors, which hasbeen reported for the siaB mutants (5, 12, 19). If this were the
case, one would expect a more severe defect in the controlgroup than was observed. On that note, the results obtainedwith the htrB mutant are somewhat surprising given thisstrain’s susceptibility to defensins in the lung (43, 46). Regard-less, these data clearly point toward the oligosaccharide por-tion of the lipooligosaccharide as a determinant of resistanceto clearance in the mouse COPD model system. It is alsonoteworthy that no enhancement was seen in the infectionstudies with the licON mutant strain (Fig. 7). Whereas our priorwork has clearly demonstrated that this strain has increasedbiofilm density (11), it is also clear that in some disease set-tings, not only the presence but the phase variation of the lic1system is required (14). This may indicate that phosphorylcho-line is advantageous only within certain windows of the diseaseprocess. Furthermore, it should be noted that the requirementsfor persistence and/or virulence within the lung and middle earmay be subtly or even dramatically different.
In summary, we have used a mouse model for COPD todemonstrate that ICAM-1 expression by the host and biofilmformation by the pathogen are important in determining theoutcome of pulmonary infections with NTHi in the context ofexperimental COPD in a murine model. As the availability ofa relevant animal model has been lacking for COPD-relatedinfections, the results of this study provide a means to test thebiofilm hypothesis as well as other fundamental hypothesesregarding the role of opportunistic pathogens in the exacerba-tion of COPD.
ACKNOWLEDGMENTS
We acknowledge excellent technical assistance by Gayle Foster andhelpful conversations with colleagues in the WFUHS Department ofMicrobiology and Immunology.
FIG. 7. Clearance of NTHi mutants from mock-treated and elastase-treated mice. Mice were treated with elastase and infected as describedin the legend to Fig. 1. Symbols represent CFU counts recovered from homogenized lung tissue from mock-treated (filled circles) and elastase-treated (open circles) animals 48 h postinfection. The dotted line indicates the lower limit of detection, and symbols below that line indicate amouse from which no bacteria were recovered. Horizontal bars represent median values, and error bars represent standard errors of the mean foreach group. Asterisks indicate groups in which bacterial numbers were significantly higher than in controls, as assessed by nonparametric statisticalanalysis.
VOL. 76, 2008 IMPAIRED BACTERIAL CLEARANCE IN MOUSE MODEL FOR COPD 4965
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

This work was supported by a grant from NIH/NIAID (AI054425; toW.E.S.).
Shayla West-Barnette was supported by an individual NIH fellow-ship (AI061830).
REFERENCES
1. Avadhanula, V., C. A. Rodriguez, G. C. Ulett, L. O. Bakaletz, and E. E.Adderson. 2006. Nontypeable Haemophilus influenzae adheres to intercellu-lar adhesion molecule 1 (ICAM-1) on respiratory epithelial cells and up-regulates ICAM-1 expression. Infect. Immun. 74:830–838.
2. Campagnari, A. A., M. R. Gupta, K. C. Dudas, T. F. Murphy, and M. A.Apicella. 1987. Antigenic diversity of lipooligosaccharides of nontypeableHaemophilus influenzae. Infect. Immun. 55:882–887.
3. Chin, C. L., L. J. Manzel, E. E. Lehman, A. L. Humlicek, L. Shi, T. D.Starner, G. M. Denning, T. F. Murphy, S. Sethi, and D. C. Look. 2005.Haemophilus influenzae from patients with chronic obstructive pulmonarydisease exacerbation induce more inflammation than colonizers. Am. J.Respir. Crit. Care Med. 172:85–91.
4. Churchill, L., R. H. Gundel, L. G. Letts, and C. D. Wegner. 1993. Contri-bution of specific cell-adhesive glycoproteins to airway and alveolar inflam-mation and dysfunction. Am. Rev. Respir. Dis. 148:S83–S87.
5. Figueira, M. A., S. Ram, R. Goldstein, D. W. Hood, E. R. Moxon, and S. I.Pelton. 2007. Role of complement in defense of the middle ear revealed byrestoring the virulence of nontypeable Haemophilus influenzae siaB mutants.Infect. Immun. 75:325–333.
6. Frick, A. G., T. D. Joseph, L. Pang, A. M. Rabe, J. W. St. Geme III, and D. C.Look. 2000. Haemophilus influenzae stimulates ICAM-1 expression on respi-ratory epithelial cells. J. Immunol. 164:4185–4196.
7. Gundel, R. H., C. D. Wegner, C. A. Torcellini, and L. G. Letts. 1992. The roleof intercellular adhesion molecule-1 in chronic airway inflammation. Clin.Exp. Allergy 22:569–575.
8. Halbert, R. J., J. L. Natoli, A. Gano, E. Badamgarav, A. S. Buist, and D. M.Mannino. 2006. Global burden of COPD: systematic review and meta-anal-ysis. Eur. Respir. J. 28:523–532.
9. Hirschmann, J. V. 2000. Do bacteria cause exacerbations of COPD? Chest118:193–203.
10. Hong, W., K. Mason, J. A. Jurcisek, L. A. Novotny, L. O. Bakaletz, and W. E.Swords. 2007. Phosphorylcholine decreases early inflammation and pro-motes the establishment of stable biofilm communities of nontypeable Hae-mophilus influenzae strain 86-028NP in a chinchilla model of otitis media.Infect. Immun. 75:958–965.
11. Hong, W., B. Pang, S. West-Barnette, and W. E. Swords. 2007. Phosphoryl-choline expression by nontypeable Haemophilus influenzae correlates withmaturation of biofilm communities in vitro and in vivo. J. Bacteriol. 189:8300–8307.
12. Hood, D. W., K. Makepeace, M. E. Deadman, R. F. Rest, P. Thibault, A.Martin, J. C. Richards, and E. R. Moxon. 1999. Sialic acid in the lipopoly-saccharide of Haemophilus influenzae: strain distribution, influence on serumresistance and structural characterization. Mol. Microbiol. 33:679–692.
13. Hoyert, D. L., E. Arias, B. L. Smith, S. L. Murphy, and K. D. Kochanek.2001. Deaths: final data for 1999. Natl. Vital Stat. Rep. 49:1–113.
14. Humphries, H. E., and N. J. High. 2002. The role of licA phase variation inthe pathogenesis of invasive disease by Haemophilus influenzae type b. FEMSImmunol. Med. Microbiol. 34:221–230.
15. Inoue, S., H. Nakamura, K. Otake, H. Saito, K. Terashita, J. Sato, H.Takeda, and H. Tomoike. 2003. Impaired pulmonary inflammatory responsesare a prominent feature of streptococcal pneumonia in mice with experi-mental emphysema. Am. J. Respir. Crit. Care Med. 167:764–770.
16. Jones, P. A., N. A. Samuels, N. J. Phillips, R. S. Munson, J. A. Bozue, J. A.Arseneau, W. A. Nichols, A. Zaleski, B. W. Gibson, and M. A. Apicella. 2002.Haemophilus influenzae type B strain A2 has multiple sialyltransferases in-volved in lipooligosaccharide sialylation. J. Biol. Chem. 277:14598–14611.
17. Jurcisek, J. A., and L. O. Bakaletz. 2007. Biofilms formed by nontypeableHaemophilus influenzae in vivo contain both double-stranded DNA and typeIV pilin protein. J. Bacteriol. 189:3868–3875.
18. Jurcisek, J. A., J. Bookwalter, B. Baker, S. Fernandez, L. A. Novotny, R. S.Munson, Jr., and L. O. Bakaletz. 2007. The PilA protein of nontypeableHaemophilus influenzae plays a role in biofilm formation, adherence to ep-ithelial cells and colonization of the mammalian upper respiratory tract. Mol.Microbiol. 65:1288–1299.
19. Jurcisek, J. A., L. Greiner, H. Watanabe, A. Zaleski, M. A. Apicella, andL. O. Bakaletz. 2005. Role of sialic acid and complex carbohydrate biosyn-thesis in biofilm formation by nontypeable Haemophilus influenzae in thechinchilla middle ear. Infect. Immun. 73:3210–3218.
20. Karlinsky, J. B., and G. L. Snider. 1978. Animal models of emphysema. Am.Rev. Respir. Dis. 117:1109–1133.
21. Lee, N.-G., M. G. Sunshine, J. J. Engstrom, B. W. Gibson, and M. A.Apicella. 1995. Mutation of the htrB locus of Haemophilus influenzae non-typeable strain 2019 is associated with modifications of lipid A and phos-phorylation of the lipo-oligosaccharide. J. Biol. Chem. 270:27151–27159.
22. Mannino, D. M., D. M. Homa, L. J. Akinbami, E. S. Ford, and S. C. Redd.
2002. Chronic obstructive pulmonary disease surveillance–United States,1971-2000. MMWR Surveill Summ. 51:1–16.
23. Monso, E., J. Ruiz, A. Rosell, J. Manterola, J. Fiz, J. Morera, and V. Ausina.1995. Bacterial infection in chronic obstructive pulmonary disease. A studyof stable and exacerbated outpatients using the protected specimen brush.Am. J. Respir. Crit. Care Med. 152:1316–1320.
24. Murphy, T. F. 2000. Haemophilus influenzae in chronic bronchitis. Semin.Respir. Infect. 15:41–51.
25. Murphy, T. F., A. L. Brauer, K. Eschberger, P. Lobbins, L. Grove, X. Cai,and S. Sethi. 2008. Pseudomonas aeruginosa in chronic obstructive pulmo-nary disease. Am. J. Respir. Crit. Care Med. 177:491–497.
26. Murphy, T. F., A. L. Brauer, A. T. Schiffmacher, and S. Sethi. 2004. Persis-tent colonization by Haemophilus influenzae in chronic obstructive pulmo-nary disease. Am. J. Respir. Crit. Care Med. 170:266–272.
27. Murphy, T. F., A. L. Brauer, S. Sethi, M. Kilian, X. Cai, and A. J. Lesse.2007. Haemophilus haemolyticus: a human respiratory tract commensal to bedistinguished from Haemophilus influenzae. J. Infect. Dis. 195:81–89.
28. Murphy, T. F., C. Kirkham, S. Sethi, and A. Lesse. 2005. Expression of aperoxiredoxin-glutaredoxin by Haemophilus influenzae in bioflims andduring human respiratory tract infection. FEMS Immunol. Med. Micro-biol. 44:81–89.
29. Murphy, T. F., and S. Sethi. 1992. Bacterial infection in chronic obstructivepulmonary disease. Am. Rev. Respir. Dis. 146:1067–1083.
30. Murphy, T. F., S. Sethi, K. L. Klingman, A. B. Brueggemann, and G. V.Doern. 1999. Simultaneous respiratory tract colonization by multiple strainsof nontypeable Haemophilus influenzae in chronic obstructive pulmonarydisease: implications for antibiotic therapy. J. Infect. Dis. 180:404–409.
31. Murphy, T. F., S. Sethi, and M. S. Niederman. 2000. The role of bacteria inexacerbations of COPD. A constructive view. Chest 118:204–209.
32. Otto-Verberne, C. J., A. A. Ten Have-Opbroek, C. Franken, J. Hermans, andJ. H. Dijkman. 1992. Protective effect of pulmonary surfactant on elastase-induced emphysema in mice. Eur. Respir. J. 5:1223–1230.
33. Pang, B., D. Winn, R. Johnson, W. Hong, S. West-Barnette, N. Kock, andW. E. Swords. 2008. Lipooligosaccharides containing phosphorylcholine de-lay pulmonary clearance of nontypeable Haemophilus influenzae. Infect. Im-mun. 76:2037–2043.
34. Rennard, S. I. 1998. COPD: overview of definitions, epidemiology, andfactors influencing its development. Chest 113(Suppl.):235S–241S.
35. Sethi, S., N. Evans, B. J. Grant, and T. F. Murphy. 2002. New strains ofbacteria and exacerbations of chronic obstructive pulmonary disease.N. Engl. J. Med. 347:465–471.
36. Sethi, S., J. Maloney, L. Grove, C. Wrona, and C. S. Berenson. 2006. Airwayinflammation and bronchial bacterial colonization in chronic obstructivepulmonary disease. Am. J. Respir. Crit. Care Med. 173:991–998.
37. Sethi, S., and T. F. Murphy. 2001. Bacterial infection in chronic obstructivepulmonary disease in 2000: a state-of-the-art review. Clin. Microbiol. Rev.14:336–363.
38. Sethi, S., R. Sethi, K. Eschberger, P. Lobbins, X. Cai, B. J. Grant, and T. F.Murphy. 2007. Airway bacterial concentrations and exacerbations of chronicobstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 176:356–361.
39. Sethi, S., C. Wrona, K. Eschberger, P. Lobbins, X. Cai, and T. F. Murphy.2007. Inflammatory profile of new bacterial strain exacerbations of chronicobstructive pulmonary disease. Am. J. Respir. Crit. Care Med.
40. Shapiro, S. D. 2002. Neutrophil elastase: path clearer, pathogen killer, or justpathologic? Am. J. Respir. Cell Mol. Biol. 26:266–268.
41. Shapiro, S. D. 1995. The pathogenesis of emphysema: the elastase:antiela-stase hypothesis 30 years later. Proc. Assoc. Am. Physicians 107:346–352.
42. Shapiro, S. D., N. M. Goldstein, A. M. Houghton, D. K. Kobayashi, D.Kelley, and A. Belaaouaj. 2003. Neutrophil elastase contributes to cigarettesmoke-induced emphysema in mice. Am. J. Pathol. 163:2329–2335.
43. Starner, T. D., W. E. Swords, M. A. Apicella, and P. B. McCray, Jr. 2002.Susceptibility of nontypeable Haemophilus influenzae to human �-defensinsis influenced by lipooligosaccharide acylation. Infect. Immun. 70:5287–5289.
44. Sullivan, S. D., S. D. Ramsey, and T. A. Lee. 2000. Economic burden ofCOPD. Chest 117:5S–9S.
45. Swords, W. E., B. Buscher, K. Ver Steeg, W. Nichols, A. Preston, J. N.Weiser, B. Gibson, and M. A. Apicella. 2000. Nontypeable Haemophilusinfluenzae adhere to and invade human bronchial epithelial cells by aninteraction of lipooligosaccharide with the PAF receptor. Mol. Microbiol.37:13–27.
46. Swords, W. E., D. L. Chance, L. A. Cohn, J. Shao, M. A. Apicella, and A. L.Smith. 2002. Acylation of the lipooligosaccharide of Haemophilus influenzaeand colonization: an htrB mutation diminishes the colonization of humanairway epithelial cells. Infect. Immun. 70:4661–4668.
47. Swords, W. E., M. R. Ketterer, J. Shao, C. A. Campbell, J. N. Weiser, andM. A. Apicella. 2001. Binding of the nontypeable Haemophilus influenzaelipooligosaccharide to the PAF receptor initiates host cell signaling. CellMicrobiol. 8:525–536.
48. Swords, W. E., M. L. Moore, L. Godzicki, G. Bukofzer, M. J. Mitten, andJ. VonCannon. 2004. Sialylation of lipooligosaccharides promotes biofilmformation by nontypeable Haemophilus influenzae. Infect. Immun. 72:106–113.
4966 PANG ET AL. INFECT. IMMUN.
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

49. Toews, G. B., D. A. Hart, and E. J. Hansen. 1985. Effect of systemic immu-nization on pulmonary clearance of Haemophilus influenzae type b. Infect.Immun. 48:343–349.
50. Toews, G. B., S. Viroslav, D. A. Hart, and E. J. Hansen. 1984. Pulmonaryclearance of encapsulated and unencapsulated Haemophilus influenzaestrains. Infect. Immun. 45:437–442.
51. Tosi, M. F., J. M. Stark, C. W. Smith, A. Hamedani, D. C. Gruenert, andM. D. Infeld. 1992. Induction of ICAM-1 expression on human airway
epithelial cells by inflammatory cytokines: effects on neutrophil-epithelialcell adhesion. Am. J. Respir. Cell Mol. Biol. 7:214–221.
52. Wegner, C. D., R. H. Gundel, P. Reilly, N. Haynes, L. G. Letts, and R.Rothlein. 1990. Intercellular adhesion molecule-1 (ICAM-1) in the patho-genesis of asthma. Science 247:456–459.
53. West-Barnette, S., A. Rockel, and W. E. Swords. 2006. Biofilm growth in-creases phosphorylcholine content and decreases potency of nontypeableHaemophilus influenzae endotoxins. Infect. Immun. 74:1828–1836.
Editor: J. N. Weiser
VOL. 76, 2008 IMPAIRED BACTERIAL CLEARANCE IN MOUSE MODEL FOR COPD 4967
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from











![Journal of Biomedical Science BioMed Centrallar adhesion molecule-1 (ICAM-1) is a crucial step in thrombosis/atherogenesis [12]. Suppressed expression of ICAM-1 was associated with](https://img.pdfslide.us/doc/110x75/60a4dc9698be37434147c54d/journal-of-biomedical-science-biomed-central-lar-adhesion-molecule-1-icam-1-is.jpg)

















