Embed Size (px)
Citation preview

Dimer-based model for heptaspanningmembrane receptorsRafael Franco1, Vicent Casado1, Josefa Mallol1, Sergi Ferre2, Kjell Fuxe3,
Antonio Cortes1, Francisco Ciruela1, Carmen Lluis1 and Enric I. Canela1
1Department Bioquımica i Biologia Molecular, Universitat de Barcelona, A. Diagonal, 645. 08028 Barcelona, Spain2National Institute on Drug Abuse, NIH, DHHS, Baltimore, MD 21224, USA3Department of Neuroscience; Division of Cellular and Molecular Neurochemistry, Karolinska Institutet (KF), S-171 77 Stockholm,
Sweden
Glossary
Non-linear Scatchard plots: Scatchard plots are used to determine the number
of binding sites and the equilibrium constant (Kd) for agonist, antagonist or any
other ligand. A Scatchard plot is linear when one molecule of ligand binds to
onemolecule of the receptor or when the binding of one molecule of the ligand
has no effect on the affinity of the other molecules for the vacant sites of the
receptor (all the intrinsic affinity constants are identical). Non-linear Scatchard
plots for ligand binding are the general consequence of the binding of more
than one molecule of ligand to one molecule of receptor or when one ligand
binds to two separated or non-interconvertible forms of a receptor having
different affinity constant values.
Hill coefficient: An indicator of cooperativity. When the Hill coefficient is!1 the
cooperativity is negative; if the value is O1, the cooperativity is positive.
Occupational theory of drug action: This theory assumes that themagnitude of
the receptor-mediated effect is proportional to the concentration of the drug-
receptor complex.
F-test: The F-test is used to compare the variances (or standard deviations) of
The existence of intramembrane receptor–receptor
interactions for heptaspanning membrane receptors is
now fully accepted, but a model considering dimers as
the basic unit that binds to two ligand molecules is
lacking. Here, we propose a two-state-dimer model in
which the ligand-induced conformational changes from
one component of the dimer are communicated to the
other. Our model predicts cooperativity in binding,
which is relevant because the other current models fail
to address this phenomenon satisfactorily. Our two-
state-dimer model also predicts the variety of responses
elicited by full or partial agonists, neutral antagonists
and inverse agonists. This model can aid our under-
standing of the operation of heptaspanning receptors
and receptor channels, and, potentially, be important for
improving the treatment of cardiovascular, neurological
and neuropsychyatric diseases.
Background
From the application of the pioneering ‘two-state’ theoryfor receptors [1–3] a variety of models have been devised toexplain the behaviour of heptaspanning, G protein-coupled receptors (GPCRs) [4–14]. Almost all currentmodels are based on the non-cooperative mechanismproposed in 1957 by del Castillo and Katz [15] for nicotinicreceptors, which do not belong to the GPCR superfamily.
Although the binding of agonists to heptaspanning recep-tors leads to non-linear Scatchard plots (see Glossary),current models fail to predict non-linear Scatchard plotsfor agonist binding. Non-linear Scatchard plots displayingcurves with a Hill coefficient of !1 could be explainedby assuming the existence of two separated or non-interconvertible forms of the receptor: a high-affinityform (R* or G protein-coupled) and a low-affinity form(R or G protein-uncoupled). However, existing modelsassume that R and R* are in equilibrium and, therefore,they would account for non-linear Scatchard plots only if(i) the concentration of G protein is lower or similar to thatof the receptor and (ii) the interconversion betweencoupled and uncoupled forms is almost irreversible. Thisis not an accurate assumption for many physiologicalconditions, mainly because G proteins are in excess [16].
Corresponding author: Canela, E.I. ([email protected]).Available online 13 June 2005
www.sciencedirect.com 0968-0004/$ - see front matter Q 2005 Elsevier Ltd. All rights reserved
Moreover, an agonist might induce changes in theproportions of the so-called ‘high-affinity’ and ‘low-affinity’states, which strongly suggests that the two states cannotexist separately but that they are interconnected [17]. Wehave provided compelling evidence that the apparent inter-conversion between states is independent of G proteins [18].If the agonist varies the proportion of high- and low-affinity sites, the existing models cannot accuratelyrepresent the behaviour of the receptors. Moreover,these models cannot explain positive cooperativity,which has been reported for 5-hydroxytryptamine1A
receptor [19,20]. Non-linear Scatchard plots require amodel of cooperativity that can explain both positive andnegative cooperativity.
A general model that is able to predict positive, neutraland negative cooperativity should consider inter- and/orintra-molecular interactions resulting from a multivalentreceptor molecule in terms of the number of ligand-binding sites per molecule of receptor; that is, a moleculecapable of binding more than one agonist molecule or,alternatively, the receptor molecule being oligomeric[21,22]. Cooperativity in the binding of agonists toreceptor oligomers would come as naturally as in thecase of oligomeric enzymes or oligomeric functionalproteins such as haemoglobin.
The hypothesis on the existence of intramembranereceptor–receptor interactions was introduced in the early
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005
two sets of data to determine whether they differ significantly. The classical
t-test is a particular case of the F-test.
. doi:10.1016/j.tibs.2005.05.010
α K
µθ K
L
α L
θα L
K
µK
A + A + (RR) A + A + (RR)*
A + A(RR)*
A(RR)*AA(RR)A
A + A(RR)
Ti BS
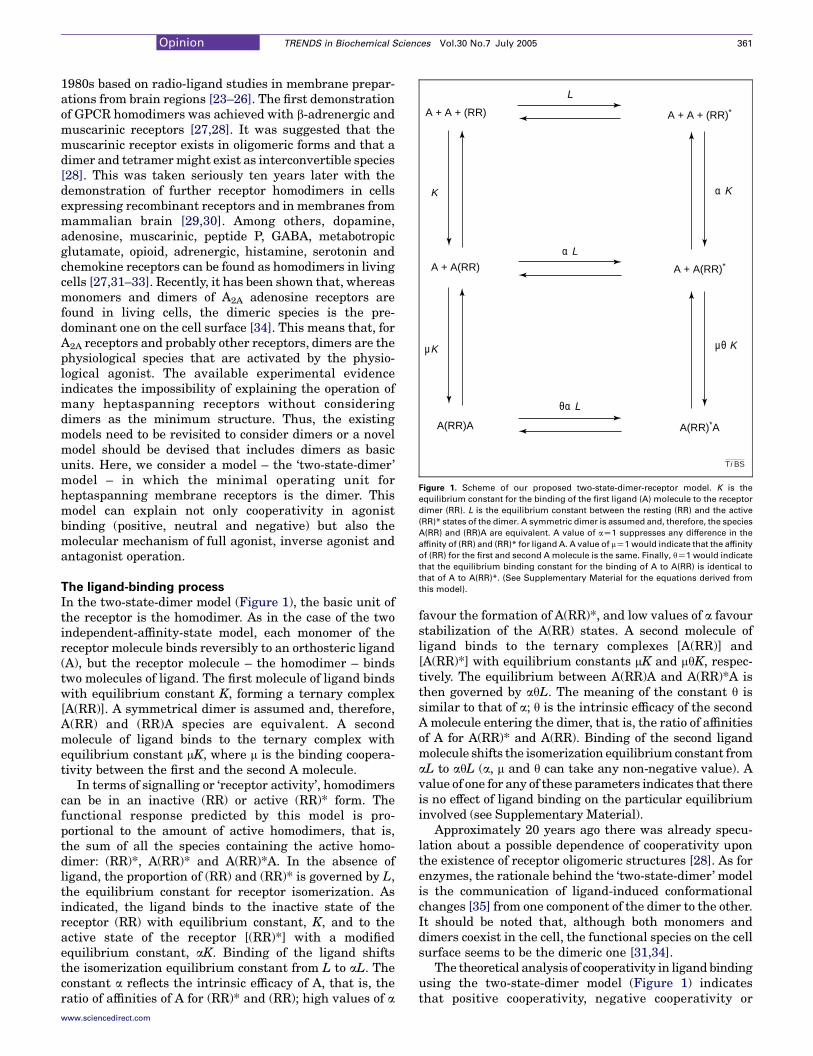
Figure 1. Scheme of our proposed two-state-dimer-receptor model. K is the
equilibrium constant for the binding of the first ligand (A) molecule to the receptor
dimer (RR). L is the equilibrium constant between the resting (RR) and the active
(RR)* states of the dimer. A symmetric dimer is assumed and, therefore, the species
A(RR) and (RR)A are equivalent. A value of aZ1 suppresses any difference in the
affinity of (RR) and (RR)* for ligand A. A value of mZ1 would indicate that the affinity
of (RR) for the first and second A molecule is the same. Finally, qZ1 would indicate
that the equilibrium binding constant for the binding of A to A(RR) is identical to
that of A to A(RR)*. (See Supplementary Material for the equations derived from
this model).
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 361
1980s based on radio-ligand studies in membrane prepar-ations from brain regions [23–26]. The first demonstrationof GPCR homodimers was achieved with b-adrenergic andmuscarinic receptors [27,28]. It was suggested that themuscarinic receptor exists in oligomeric forms and that adimer and tetramer might exist as interconvertible species[28]. This was taken seriously ten years later with thedemonstration of further receptor homodimers in cellsexpressing recombinant receptors and in membranes frommammalian brain [29,30]. Among others, dopamine,adenosine, muscarinic, peptide P, GABA, metabotropicglutamate, opioid, adrenergic, histamine, serotonin andchemokine receptors can be found as homodimers in livingcells [27,31–33]. Recently, it has been shown that, whereasmonomers and dimers of A2A adenosine receptors arefound in living cells, the dimeric species is the pre-dominant one on the cell surface [34]. This means that, forA2A receptors and probably other receptors, dimers are thephysiological species that are activated by the physio-logical agonist. The available experimental evidenceindicates the impossibility of explaining the operation ofmany heptaspanning receptors without consideringdimers as the minimum structure. Thus, the existingmodels need to be revisited to consider dimers or a novelmodel should be devised that includes dimers as basicunits. Here, we consider a model – the ‘two-state-dimer’model – in which the minimal operating unit forheptaspanning membrane receptors is the dimer. Thismodel can explain not only cooperativity in agonistbinding (positive, neutral and negative) but also themolecular mechanism of full agonist, inverse agonist andantagonist operation.
The ligand-binding process
In the two-state-dimer model (Figure 1), the basic unit ofthe receptor is the homodimer. As in the case of the twoindependent-affinity-state model, each monomer of thereceptor molecule binds reversibly to an orthosteric ligand(A), but the receptor molecule – the homodimer – bindstwo molecules of ligand. The first molecule of ligand bindswith equilibrium constant K, forming a ternary complex[A(RR)]. A symmetrical dimer is assumed and, therefore,A(RR) and (RR)A species are equivalent. A secondmolecule of ligand binds to the ternary complex withequilibrium constant mK, where m is the binding coopera-tivity between the first and the second A molecule.
In terms of signalling or ‘receptor activity’, homodimerscan be in an inactive (RR) or active (RR)* form. Thefunctional response predicted by this model is pro-portional to the amount of active homodimers, that is,the sum of all the species containing the active homo-dimer: (RR)*, A(RR)* and A(RR)*A. In the absence ofligand, the proportion of (RR) and (RR)* is governed by L,the equilibrium constant for receptor isomerization. Asindicated, the ligand binds to the inactive state of thereceptor (RR) with equilibrium constant, K, and to theactive state of the receptor [(RR)*] with a modifiedequilibrium constant, aK. Binding of the ligand shiftsthe isomerization equilibrium constant from L to aL. Theconstant a reflects the intrinsic efficacy of A, that is, theratio of affinities of A for (RR)* and (RR); high values of a
www.sciencedirect.com
favour the formation of A(RR)*, and low values of a favourstabilization of the A(RR) states. A second molecule ofligand binds to the ternary complexes [A(RR)] and[A(RR)*] with equilibrium constants mK and mqK, respec-tively. The equilibrium between A(RR)A and A(RR)*A isthen governed by aqL. The meaning of the constant q issimilar to that of a; q is the intrinsic efficacy of the secondA molecule entering the dimer, that is, the ratio of affinitiesof A for A(RR)* and A(RR). Binding of the second ligandmolecule shifts the isomerization equilibrium constant fromaL to aqL (a, m and q can take any non-negative value). Avalue of one for any of these parameters indicates that thereis no effect of ligand binding on the particular equilibriuminvolved (see Supplementary Material).
Approximately 20 years ago there was already specu-lation about a possible dependence of cooperativity uponthe existence of receptor oligomeric structures [28]. As forenzymes, the rationale behind the ‘two-state-dimer’ modelis the communication of ligand-induced conformationalchanges [35] from one component of the dimer to the other.It should be noted that, although both monomers anddimers coexist in the cell, the functional species on the cellsurface seems to be the dimeric one [31,34].
The theoretical analysis of cooperativity in ligand bindingusing the two-state-dimer model (Figure 1) indicatesthat positive cooperativity, negative cooperativity or
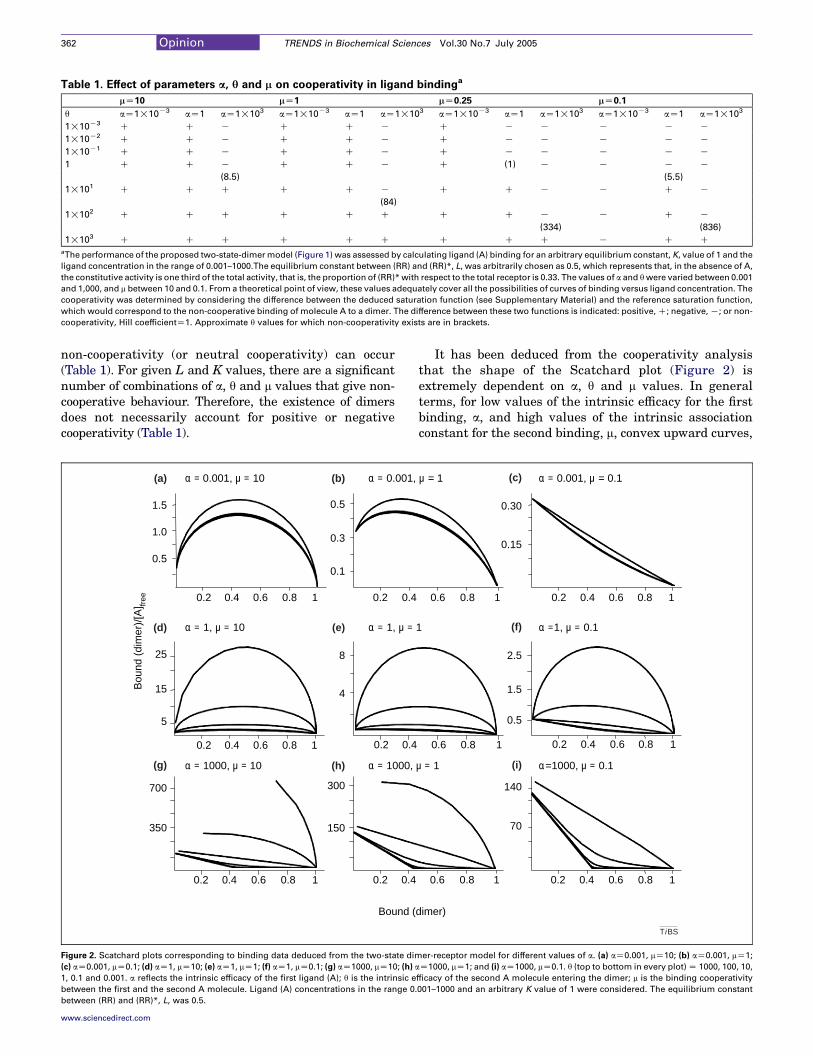
Table 1. Effect of parameters a, q and m on cooperativity in ligand bindinga
mZ10 mZ1 mZ0.25 mZ0.1
q aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103 aZ1!10K3 aZ1 aZ1!103
1!10K3 C C K C C K C K K K K K
1!10K2 C C K C C K C K K K K K
1!10K1 C C K C C K C K K K K K
1 C C K C C K C (1) K K K K
(8.5) (5.5)
1!101 C C C C C K C C K K C K
(84)
1!102 C C C C C C C C K K C K
(334) (836)
1!103 C C C C C C C C C K C CaThe performance of the proposed two-state-dimermodel (Figure 1) was assessed by calculating ligand (A) binding for an arbitrary equilibrium constant, K, value of 1 and the
ligand concentration in the range of 0.001–1000.The equilibrium constant between (RR) and (RR)*, L, was arbitrarily chosen as 0.5, which represents that, in the absence of A,
the constitutive activity is one third of the total activity, that is, the proportion of (RR)*with respect to the total receptor is 0.33. The values of a and qwere varied between 0.001
and 1,000, and m between 10 and 0.1. From a theoretical point of view, these values adequately cover all the possibilities of curves of binding versus ligand concentration. The
cooperativity was determined by considering the difference between the deduced saturation function (see Supplementary Material) and the reference saturation function,
which would correspond to the non-cooperative binding of molecule A to a dimer. The difference between these two functions is indicated: positive,C; negative,K; or non-
cooperativity, Hill coefficientZ1. Approximate q values for which non-cooperativity exists are in brackets.
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005362
non-cooperativity (or neutral cooperativity) can occur(Table 1). For given L and K values, there are a significantnumber of combinations of a, q and m values that give non-cooperative behaviour. Therefore, the existence of dimersdoes not necessarily account for positive or negativecooperativity (Table 1).
Bound (
1.5
0.6 1
1.0
0.5
0.40.2
0.5
0.3
0.1
0.40.2
0.6 10.40.2
8
4
0.40.2
700
0.6 1
350
0.40.2
300
150
0.40.2
25
15
5
(a) α = 0.001, µ = 10 (b)
(d) (e)
(g) (h)
α = 0.001,
α = 1, µ = 10 α = 1, µ =
Bou
nd (
dim
er)/
[A] fr
ee
α = 1000, µ = 10 α = 1000,
0.8
0.8
0.8
Figure 2. Scatchard plots corresponding to binding data deduced from the two-state dim
(c) aZ0.001, mZ0.1; (d) aZ1, mZ10; (e) aZ1, mZ1; (f) aZ1, mZ0.1; (g) aZ1000, mZ10; (h) a
1, 0.1 and 0.001. a reflects the intrinsic efficacy of the first ligand (A); q is the intrinsic e
between the first and the second A molecule. Ligand (A) concentrations in the range 0.
between (RR) and (RR)*, L, was 0.5.
www.sciencedirect.com
It has been deduced from the cooperativity analysisthat the shape of the Scatchard plot (Figure 2) isextremely dependent on a, q and m values. In generalterms, for low values of the intrinsic efficacy for the firstbinding, a, and high values of the intrinsic associationconstant for the second binding, m, convex upward curves,
dimer)
0.6 1
0.30
0.6 1
0.15
0.40.2
0.6 1
2.5
0.6 1
1.5
0.5
0.40.2
0.6 1
140
0.6 1
70
0.40.2
(c)
(f)
(i)
µ = 1 α = 0.001, µ = 0.1
1 α =1, µ = 0.1
µ = 1 α=1000, µ = 0.1
0.8 0.8
0.8 0.8
0.8 0.8
Ti BS
er-receptor model for different values of a. (a) aZ0.001, mZ10; (b) aZ0.001, mZ1;
Z1000, mZ1; and (i) aZ1000, mZ0.1. q (top to bottom in every plot)Z 1000, 100, 10,
fficacy of the second A molecule entering the dimer; m is the binding cooperativity
001–1000 and an arbitrary K value of 1 were considered. The equilibrium constant
Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 363
which are indicative of positive cooperativity, are pre-dominantly found (Figure 2a,b,d,e). By contrast, for lowvalues of both m and a, negative cooperativity is found(Figure 2c). For higher values of a, positive or negativecooperativity would depend on the value of the intrinsicefficacy of the second binding (q) (Figure 2g,h,i). When q ishigh, positive cooperativity is predominantly found,whereas negative cooperativity is more probable when qis low (Figure 2d,g).
Receptor activation
The performance of our two-state-dimer model has beenassessed by a study of the dependence of receptor activityon ligand concentration. To enable direct comparison ofthe responses elicited by agonists and antagonists, theoccupational theory of drug action [36] was selected. Inthe symmetric two-state-dimer model, (RR)*, A(RR)* andA(RR)*A contribute to the overall response, whereas theinactive (RR), A(RR) and A(RR)A do not. The proportional-ity factor is set to one and the response, therefore, isequivalent to the ratio of active species versus totalamount of receptor [Equation 1, where A(RR)* isequivalent to (RR)*A; see Supplementary Material]:
0.30
-3 -2 -1 12 3
0.15
-3 -2 -1
-3 -2 -1 1 3 -3 -2 -1
-3 -2 -1 1 3 -3 -2 -1
0.30
0.15
1.0
0.5
1.0
0.5
1.0
0.5
1.0
0.5
0
0
0
α = 0.001, µ = 10 α = 0.00
α = 1, µ = 10 α = 1, µ
α = 1000, µ = 10 α = 1000
2
2
Bo
Bou
nd (
dim
er)/
[A] fr
ee
(a) (b)
(d) (e)
(g) (h)
Figure 3. Receptor-activation curves for the two-state-dimermodel. Curves dose–respons
0.001, mZ0.1; (d) aZ1, mZ10; (e) aZ 1, mZ1; (f) aZ1, mZ0.1; (g) aZ1000, mZ10; (h) aZ100
and 0.001. a reflects the intrinsic efficacy of the first ligand (A); q is the intrinsic efficacy of t
first and the second Amolecule. Ligand (A) concentrations in the range 0.001–1000 and a
(RR)*, L, was 0.5.
www.sciencedirect.com
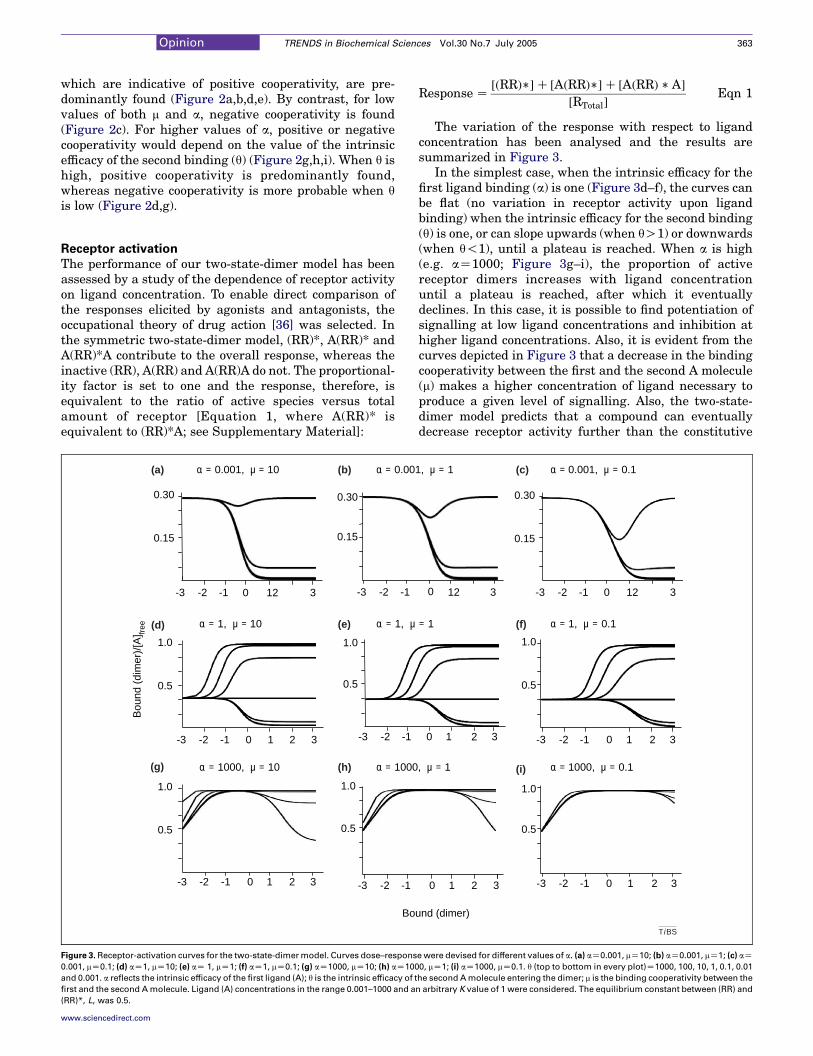
Response Z½ðRRÞ��C ½AðRRÞ��C ½AðRRÞ � A�
½RTotal�Eqn 1
The variation of the response with respect to ligandconcentration has been analysed and the results aresummarized in Figure 3.
In the simplest case, when the intrinsic efficacy for thefirst ligand binding (a) is one (Figure 3d–f), the curves canbe flat (no variation in receptor activity upon ligandbinding) when the intrinsic efficacy for the second binding(q) is one, or can slope upwards (when qO1) or downwards(when q!1), until a plateau is reached. When a is high(e.g. aZ1000; Figure 3g–i), the proportion of activereceptor dimers increases with ligand concentrationuntil a plateau is reached, after which it eventuallydeclines. In this case, it is possible to find potentiation ofsignalling at low ligand concentrations and inhibition athigher ligand concentrations. Also, it is evident from thecurves depicted in Figure 3 that a decrease in the bindingcooperativity between the first and the second A molecule(m) makes a higher concentration of ligand necessary toproduce a given level of signalling. Also, the two-state-dimer model predicts that a compound can eventuallydecrease receptor activity further than the constitutive
12 3 -3 -2 -1 12 3
1 3 -3 -2 -1 1 3
1 3 -3 -2 -1 1 3
0.30
0.15
1.0
0.5
1.0
0.5
0 0
00
0 0
1, µ = 1 α = 0.001, µ = 0.1
= 1 α = 1, µ = 0.1
, µ = 1 α = 1000, µ = 0.1
2 2
2 2
und (dimer)
(c)
(f)
(i)
Ti BS
ewere devised for different values of a. (a) aZ0.001, mZ10; (b) aZ0.001, mZ1; (c) aZ0, mZ1; (i) aZ1000, mZ0.1. q (top to bottom in every plot)Z1000, 100, 10, 1, 0.1, 0.01
he second Amolecule entering the dimer; m is the binding cooperativity between the
n arbitrary K value of 1 were considered. The equilibrium constant between (RR) and

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005364
activity until a plateau or a minimum is reached(Figure 3a–c).
Full and partial agonists
Full agonists are defined by their ability to increasereceptor activity in a dose-dependent manner until themaximum signalling response is reached. Full agonistbinding to a dimeric receptor would require values of aZ1000 for any m or q, or aZ1 for qR100 and any m(Figure 3d–i). The two-state-dimer model can explainpositive and negative cooperativity in full agonist bindingto the receptor dimer (Table 1). A partial agonist is definedas a compound that activates the receptor but unable toinduce full receptor activation. Partial agonist binding to adimeric receptor would require aZ1 and qZ1–100 for anym value (Figure 3d,e,f).
Antagonists and inverse agonists
On defining a neutral antagonist as a molecule that doesnot change the proportion of inactive [(RR)] versus active[(RR)*] species, the combination of parameters in the two-state-dimer model (which allows such a possibility) is aZ1and qZ1 for any m value (Figure 3d–f). Often, it is reportedthat binding of a neutral antagonist is non-cooperative(linear Scatchard plot). In the existing models, it isassumed that the antagonist does not distinguish betweenthe high-affinity- and low-affinity-binding sites. By con-trast, in the two-state-dimer model, non-cooperativeantagonist binding would be a unique case because theonly combination of parameters that leads to flat con-stitutive activity and give linear Scatchard plots are: mZ0.25, aZ1 and qZ1. A m value of 0.25 is required for theoverall affinity of the second binding of the ‘non-cooperative’ antagonist to be identical to the overallaffinity for the first binding. Because there are twobinding sites for the antagonist, when aZ1 and qZ1, thevalue of mK is K/4. However, values that differ slightlyfrom those indicated can lead to apparently linearScatchard plots and, thus, detection of cooperativity inbinding can become a difficult task [37].
Inverse agonists are defined by their ability toantagonize the effect of full agonists, but also by theirability to decrease the constitutive activity of hepta-spanning membrane receptors. By inspecting Figure 3, acompound would be an inverse agonist when the values ofthe parameters are: a!1 for any q and m value, or aZ1 andq!1 irrespective of m (Figure 3a–f). The scarce availabledata indicate that cooperativity can exist in the binding ofinverse agonists [37] and that values representing non-cooperative binding to the dimer would be restrictive. Thetwo-state-dimer model can predict any type of cooperativ-ity for compounds that reduce the constitutive activity ofheptaspanning receptors (Table 1).
Predictions and performance of the two-state-dimer
model
For A1 adenosine receptors, which display non-linearScatchard plots, experimental data fit better to the two-state-dimer model than to the two independent-affinity-state model [38]. Although the difference between theactual values and the theoretical ones were similar in the
www.sciencedirect.com
two cases, the two-state-dimer model has less parametersand, therefore, the discriminative F-test selects it as thesimplest model able to explain the data. The parametersobtained when fitting data to the two-state-dimer modelgive valuable information about the equilibrium con-stants but also about the degree of cooperativity (seeSupplementary Material).
One of the main features of the two-state-dimer modelis its ability to predict quantitative changes in functionalactivity. Ligands with high values of a (e.g. aZ1000) andlow values of q (e.g. qZ0.1) show dual behaviour in termsof response at equilibrium; they increase the number ofactive species at low agonist concentration, but theydecrease them at high concentrations (Figure 3). Thiseffect is even more evident for mO0.1. Thus, the modelpredicts a homologous desensitization process at highconcentrations of agonist. The strength of this desensi-tization process would vary from agonist to agonist(Figure 3d,h,i). The limit of the curves at high concen-trations of A is the constitutive activity (when qaZ1) orsurpasses it (when qa!1). Interestingly, desensitizationdepends on the product aq, which reflects properties ofboth the binding of the first ligand molecule and thebinding of the second molecule to the dimer. If aqZ1, thelimit of the curve at high ligand concentrations is theconstitutive activity. This correlates with the well-knownobservation that, in terms of desensitization, two types ofreceptor exist: those that are desensitized easily and thosethat are not. Although desensitization is usually inter-preted to be a consequence of uncoupling of signallingmachinery, this is not known for certain. The two-state-dimer model predicts that homologous desensitization isdependent on the agonist in such a way that it is weakerfor agonists displaying positive cooperativity (Figures 2and 3). As measured experimentally, desensitizationincluding the well-known refractoriness to agonists afterreceptor activation would involve this homologousdesensitization plus receptor internalization, which isdependent on G protein-receptor kinase activation andreceptor phosphorylation.
The two-state-dimer model predicts that neutralantagonists can display any type of cooperativity onbinding to receptor dimers. However, as mentioned,detection of cooperativity in binding can become a difficulttask. Recently, Cheng [37] compared different methods ofanalysing cooperativity data. The author has reportedthat the Cheng–Prusoff equation, the Scatchard analysisand the Schild analysis give different results, and hasproposed a more robust method to calculate accuratevalues for equilibrium constants and Hill coefficients.Periyasami and Somani [39] have shown that theapparent Hill coefficient for yohimbine – an antagonistof a2 adrenoceptors both in equilibrium binding and incompetitive displacement assays – is !1. It should benoted that some data on dissociation of antagonists fromGABA and glycine receptors [40] are attributed to bindingof the antagonist to an orthosteric centre and to anallosteric centre. In addition, allosteric effectors – whichhave been reported for a variety of heptaspanningmembrane receptors – would affect the cooperativitybetween the two molecules in the dimer. The consideration

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005 365
of allosteric centres will be a further development of themodel we present here.
In a recent survey of 105 published papers on theactivity of 380 antagonists on 73 biological GPCR targets,322 of the antagonists studied are reported to be inverseagonists and only 58 to be neutral antagonists [41]. Thepredominance of inverse agonism agrees with theoreticalpredictions of the two-state-dimer model, but also agreeswith those of other reported models (see SupplementaryMaterial). The parameters defining neutral antagonism inthe two-state-dimer model are restrictive and this could bethe reason that neutral antagonists are the minoritywithin the pharmacological space.
There are few reports on the cooperativity of inverse-agonist binding. The available data indicate, however,that cooperativity can exist in the binding of suchcompounds for a variety of receptors. The two-state-dimer model can predict any type of cooperativity forcompounds that reduce the constitutive activity; there-fore, the binding of these compounds to the receptordimer is represented by a!1 for any value of q and m, oraZ1 and q!1 for any value of m. Results of competitionassays for an inverse agonist of melacortin-4 receptors,carefully analysed using the Cheng protocol [37], showthat cooperativity in the competition occurs and that itvaries depending on the nature of the radioactivecompound bound. Higher (and positive) cooperativity isfound when the inverse agonist competes with the fullagonist, whereas the Hill coefficient decreases when thecompound competes with either a partial agonist or aneutral antagonist [37]. Moore and Scanlon [42] provideanother example of change in cooperativity dependingon the nature of compounds competing at the bindingsite of receptor. They demonstrate that antagonists shiftpositive cooperativity to negative cooperativity in thebinding of agonists to angiotensin receptors. Interestingly,Milligan et al. [43] have shown that two different inverseagonists that are specific for 5-hydroxytryptamine1A
receptors display different Hill coefficients. Whereasone of the inverse agonists has a Hill coefficient of O1(positive cooperativity), agonists and partial agonistsdisplay low Hill coefficients indicative of strong negativecooperativity. These varied behaviours can be easilyexplained by the two-state-dimer model: each of thesecompounds would have different a, q and m valuesand, therefore, cooperativity in binding-saturationcurves would be different for different compounds; andcooperativity for a given compound in competitionassays would vary depending on the nature of theradio-ligand bound and the nature of the competitor.Furthermore, the two-state-dimer model predicts thatcooperativity in the binding of all type of molecules tothe orthosteric centre would depend on the value of L(i.e. constitutive activity).
Another predictive valuable feature of the model isrelated to the possibility that an inverse agonist mightreduce constitutive activity at low concentrations butrevert this trend at higher concentrations. This should beconsidered when analysing data for novel putative inverseagonists before discarding them as ineffective.
www.sciencedirect.com
Concluding remarks
To explain ligand-binding and activation mechanisms ofGPCRs, their dimeric structure must be taken intoaccount. The model we propose here assumes the existenceof cooperative conformational changes in the GPCR dimer,which has been demonstrated for the leukotriene B4
receptor BLT1 [44]. In our opinion, we present a generalmodel for heptaspanning-membrane-receptor operationthat is based precisely in their (now) well-known dimericstructure. This two-state-dimer model can explain all thefeatures of the heptaspanning-membrane receptors andcan be useful for investigation of other types of receptorssuch as nicotinic acetylcholine receptors, which have twoidentical binding sites [45]. Furthermore, the modelpredicts other features (e.g. dual effects for a givencompound) of heptaspanning receptors – a tool thatcould be useful for improving current therapeutic strat-egies that target this type of receptor.
AcknowledgementsThis work was supported by grant SAF2001–3474 from Ministerio deCiencia y Tecnologia, Grant 02/056–00 from Fundacio la Caixa, Grants01/012710 and 02/021010 from Fundacio Marato TV3.
Supplementary data
Supplementary data associated with this article can befound at doi:10.1016/j.tibs.2005.05.010
References
1 Karlin, A. (1967) On the application of “a plausible model” of allostericproteins to the receptor for acetylcholine. J. Theor. Biol. 16, 306–320
2 Colquhoun, D. (1973) The relationship between classical andcooperative models for drug action. In A Symposium on DrugReceptors (Rang, H.P., ed.), pp. 149–182, University Park Press
3 Thron, C.D. (1973) On the analysis of pharmacological experiments interms of an allosteric receptor model. Mol. Pharmacol. 9, 1–9
4 De Lean, A. et al. (1980) A ternary complex model explains the agonist-specific binding properties of the adenylate cyclase-coupledb-adrenergic receptor. J. Biol. Chem. 255, 7108–7117
5 Costa, T. and Herz, A. (1989) Antagonists with negative intrinsicactivity at delta opioid receptors coupled to GTP-binding proteins.Proc. Natl. Acad. Sci. U. S. A. 86, 7321–7325
6 Samama, P. et al. (1993) A mutation-induced activated state of theb2-adrenergic receptor. Extending the ternary complex model. J. Biol.Chem. 268, 4625–4636
7 Onaran, H.O. et al. (1993) b g subunits of guanine nucleotide-bindingproteins and regulation of spontaneous receptor activity: thermo-dynamic model for the interaction between receptors and guaninenucleotide-binding protein subunits. Mol. Pharmacol. 43, 245–256
8 Samama, P. et al. (1994) Negative antagonists promote an inactiveconformation of the b2-adrenergic receptor. Mol. Pharmacol. 45,390–394
9 Leff, P. (1995) The two-state model of receptor activation. TrendsPharmacol. Sci. 16, 89–97
10 Weiss, J.M. et al. (1996) The cubic ternary complex receptor –occupancy model I. Model description. J. Theor. Biol. 178, 151–167
11 Weiss, J.M. et al. (1996) The cubic ternary complex receptor –occupancy model II. Understanding apparent affinity. J. Theor. Biol.178, 169–182
12 Weiss, J.M. et al. (1996) The cubic ternary complex receptor – occupancymodel III. Resurrecting efficacy. J. Theor. Biol. 181, 381–397
13 Hall, D.A. (2000) Modeling the functional effects of allostericmodulators at pharmacological receptors: an extension of the two-state model of receptor activation. Mol. Pharmacol. 58, 1412–1423
14 Lorenzen, A. et al. (2002) Modulation of agonist responses at the A1

Opinion TRENDS in Biochemical Sciences Vol.30 No.7 July 2005366
adenosine receptor by an irreversible antagonist, receptor-G proteinuncoupling and by the G protein activation state. Biochem.Pharmacol. 64, 1251–1265
15 del Castillo, J. and Katz, B. (1957) A comparison of acetylcholine andstable depolarizing agents.Proc. R. Soc. Lond. B. Biol. Sci. 146, 362–368
16 Neubig, R.R. (1994) Membrane organization in G-protein mechan-isms. FASEB J. 8, 939–946
17 Wong, H.M. et al. (1986) Assessment of mechanistic proposals for thebinding of agonists to cardiac muscarinic receptors. Biochemistry 25,6995–7008
18 Casado, V. et al. (1991) The binding of [3H]R-PIA to A1 adenosinereceptors produces a conversion of the high- to the low-affinity state.FEBS Lett. 286, 221–224
19 Mattera, R. et al. (1985) Guanine nucleotide regulation of amammalian myocardial muscarinic receptor system. Evidence forhomo- and heterotropic cooperativity in ligand binding analyzed bycomputer-assisted curve fitting. J. Biol. Chem. 260, 7410–7421
20 Assie, M-B. et al. (1999) Correlation between low/high affinity ratiosfor 5-HT(1A) receptors and intrinsic activity. Eur. J. Pharmacol. 386,97–103
21 Franco, R. et al. (1996) The cluster-arranged cooperative model: amodel that accounts for the kinetics of binding to A1 adenosinereceptors. Biochemistry 35, 3007–3015
22 Franco, R. et al. (2003) Regulation of heptaspanning-membrane-receptor function by dimerization and clustering. Trends Biochem.Sci. 28, 238–243
23 Agnati, L.F. et al. (1980) Aspects on receptor regulation andisoreceptor identification. Med. Biol. 58, 182–187
24 Agnati, L.F. et al. (1982) New vistas on synaptic plasticity: the receptormosaic hypothesis of the engram. Med. Biol. 60, 183–190
25 Fuxe, K. et al. (1981) Modulation by cholecystokinins of 3H-spiroperidolbinding in rat striatum: evidence for increased affinity and reductionin the number of binding sites. Acta Physiol. Scand. 113, 567–569
26 Agnati, L.F. et al. (2003) Molecular mechanisms and therapeuticalimplications of intramembrane receptor/receptor interactions amongheptahelical receptors with examples from the striatopallidal GABAneurons. Pharmacol. Rev. 55, 509–550
27 Fraser, C.M. and Venter, J.C. (1982) The size of the mammalian lungb2-adrenergic receptor as determined by target size analysis andimmunoaffinity chromatography. Biochem. Biophys. Res. Commun.109, 21–29
28 Avissar, S. et al. (1983) Oligomeric structure of muscarinic receptors isshown by photoaffinity labeling: subunit assembly may explain high-and low-affinity agonist states. Proc. Natl. Acad. Sci. U. S. A. 80,156–159
Reproduction of material
Interested in reproducing part or all of an article published by ElGlobal Rights Department with details of how and where the req
on-line, plea
http://www.elsevier.com/wps/find/obtainpermi
Alternatively, ple
ElseviGlobal Rights D
PO BoxOxford OX5
Phone: (+44) 18Fax: (+44) 186
permissions@e
www.sciencedirect.com
29 Ciruela, F. et al. (1995) Immunological identification of A1 adenosinereceptors in brain cortex. J. Neurosci. Res. 42, 818–828
30 Ng, G.Y. et al. (1996) Dopamine D2 receptor dimers and receptor-blocking peptides. Biochem. Biophys. Res. Commun. 227, 200–204
31 Bouvier, M. (2001) Oligomerization of G-protein-coupled transmitterreceptors. Nat. Rev. Neurosci. 2, 274–286
32 Milligan, G. and White, J.H. (2001) Protein-protein interactions atG-protein-coupled receptors. Trends Pharmacol. Sci. 22, 513–518
33 Bai, M. (2004) Dimerization of G-protein-coupled receptors: roles insignal transduction. Cell. Signal. 16, 175–186
34 Canals, M. et al. (2004) Homodimerization of adenosine A2A receptors:qualitative and quantitative assessment by fluorescence and bio-luminescence energy transfer. J. Neurochem. 88, 726–734
35 Vilardaga, J.P. et al. (2003) Measurement of the millisecond activationswitch of G protein-coupled receptors in living cells. Nat. Biotechnol.21, 807–812
36 Kenakin, T. (2004) Principles: receptor theory in pharmacology.Trends Pharmacol. Sci. 25, 186–192
37 Cheng, H.C. (2004) The influence of cooperativity on the determin-ation of dissociation constants: examination of the Cheng–Prusoffequation, the Scatchard analysis, the Schild analysis and relatedpower equations. Pharmacol. Res. 50, 21–40
38 Saura, C. et al. (1996) Adenosine deaminase interacts with A1
adenosine receptors in pig brain cortical membranes. J. Neurochem.66, 1675–1682
39 Periyasamy, S. and Somani, P. (1986) Effect of proteases andphospholipases on [3H]yohimbine binding to human platelet mem-branes. Biochem. Pharmacol. 35, 3131–3136
40 Maksay, G. (1990) Dissociation of muscimol, SR 95531, and strychninefrom GABAA and glycine receptors, respectively, suggests similarcooperative interactions. J. Neurochem. 54, 1961–1966
41 Kenakin, T. (2004) Efficacy as a vector: the relative prevalence andpaucity of inverse agonism. Mol. Pharmacol. 65, 2–11
42 Moore, G.J. and Scanlon, M.N. (1989) Methods for analyzing andinterpreting cooperativity in dose-response curves – I. Antagonisteffects on angiotensin receptors in smooth muscle. Gen. Pharmacol.20, 193–198
43 Milligan, G. et al. (2001) S 14506: novel receptor coupling at 5-HT1A
receptors. Neuropharmacology 40, 334–34444 Mesnier, D. and Banerest, J-L. (2004) Cooperative conformational
changes in a G-protein-coupled receptor dimer, the leukotriene B(4)receptor BLT1. J. Biol. Chem. 279, 49664–49670
45 Edelstein, S.J. et al. (1996) A kinetic mechanism for nicotinicacetylcholine receptors based on multiple allosteric transitions. Biol.Cybern. 75, 361–379
from Elsevier articles
sevier, or one of our article figures? If so, please contact ouruested material will be used. To submit a permission requestse visit:
ssionform.cws_home/obtainpermissionform
ase contact:
erepartment800,1DX, UK.65-8438305-853333lsevier.com










![Molecular and functional properties of P2X receptors ... · receptors are localized in the membrane of the intracellular contractile vacuole [27, 30]. These findings demonstrate that](https://img.pdfslide.us/doc/110x75/61358e460ad5d206764773a3/molecular-and-functional-properties-of-p2x-receptors-receptors-are-localized.jpg)


















