Embed Size (px)
Citation preview

Digestion of dietary fat
2013 A
nne Helbig
Gastrointestinal behaviour of emulsions and human physiological responses
Digestion of dietary fat
Anne Helbig
Digestion of dietary fat
2013 A
nne Helbig
You are cordially invited to You are cordially invited to attend the public attend the public defence of my thesis entitled my thesis entitled
Digestion of dietary fatDigestion of dietary fatGastrointestinal behaviour Gastrointestinal behaviour of emulsions and human of emulsions and human physiological responsesphysiological responses
To be held To be held
on Friday, 21st June 2013, Friday, 21st June 2013,
at 13:30 at 13:30
in the Aula of Wageningen in the Aula of Wageningen University University Generaal Foulkesweg 1, Generaal Foulkesweg 1, Wageningen Wageningen
After the graduation will After the graduation will be a reception in the Aula. be a reception in the Aula.
The The party and the BBQ will take place at will take place at 18:30 in the garden of the SSR the garden of the SSR Generaal Foulkesweg 30, eneraal Foulkesweg 30, Wageningen Wageningen
Anne Helbig
Please indicate if you would like to attend the party/BBQ by sending an email to one of my paranymphs
Marijke Beenes ([email protected])
Stéphanie Ladirat ([email protected])

Digestion of dietary fat
Gastrointestinal behaviour of emulsions and human
physiological responses
Anne Helbig

Thesis committee Promoters Prof. dr. ir. H. Gruppen Professor of Food Chemistry Wageningen University Prof. dr. R.J. Hamer Professor of Technology of Cereal Proteins Wageningen University Co-promoter Dr. Erika Silletti NIZO food research, Ede Other members Prof. dr.ir. C. de Graaf, Wageningen University Prof. dr. W.H.M. Saris, Maastricht University Medical Centre Prof. P. Wilde, Norwich Research Park, UK Dr. M. Foltz, Unilever R&D, Vlaardingen This research was conducted under the auspices of the Graduate School VLAG (Advanced studies in Food Technology, Agrobiotechnology and Health Sciences).

Digestion of dietary fat
Gastrointestinal behaviour of emulsions and human
physiological responses
Anne Helbig
Thesis
submitted in fulfillment of the requirements for the degree of doctor
at Wageningen University
by the authority of the Rector Magnificus
Prof. Dr. M.J. Kropff,
in the presence of the
Thesis Committee appointed by the Academic Board
to be defended in public
on Friday 21 June 2013
at 1.30 p.m. in the Aula.

Anne Helbig
Digestion of dietary fat. Gastrointestinal behaviour of emulsions and human physiological
responses
166 pages
PhD thesis, Wageningen University, Wageningen, NL (2013)
With references, with summary in English, Dutch and German
ISBN: 978-94-6173-560-7

Abstract
Two in vitro models were used to understand emulsion behavior and the subsequent
formation of free fatty acids (FFA), monoglycerides (MG) and diglycerides (DG).
Emulsions stabilized by whey protein isolate (WPI) or gum arabic (GA), varying in droplet
size, were digested under intestinal conditions. Concentrations of FFA, MG and DG,
assessed by gas chromatography, decreased with increasing droplet size. FFA release from
gum arabic-stabilized emulsions was higher compared to WPI-stabilized emulsions
showing an influence of the interface. Next, lipolysis of protein stabilized emulsions (i.e.
WPI or lysozyme) and the influence of flocculation at the isoelectric point (pI) were
investigated in a dynamic gastrointestinal model. The stomach properties including
gradual acidification caused WPI-stabilized emulsions to cream during transition through
the pI of the protein. This resulted in delayed intestinal lipolysis compared to the
lysozyme-stabilized emulsion. Thus, since gastric passage affects emulsion behavior and
intestinal lipolysis, the gastric passage should be part of digestion models. Next, in a
human study emulsion behavior and resulting lipolytic products were related to the
release of satiety hormones, satiety perception and ad libitum intake. Also, gallbladder
volume and oral processing were studied. A delayed entry into the duodenum and
lipolysis for the un-homogenized sample resulted in lower CCK, delayed GLP-1/PYY
responses and barely gallbladder contraction compared to the homogenized emulsion. No
difference was found between treatments on ghrelin, only the perception `desire to eat´
was elevated for homogenized emulsions. Oral processing induced prolonged gallbladder
contraction, but had no additive effect on other measures. A homogenous system as such
is possibly not effective to induce pronounced satiety perceptions compared to phase
separated or creamed systems using the same emulsifier. Moreover, the release of
gastrointestinal hormones cannot directly be related to the satiating effect of food.


Table of contents
Abstract
Chapter 1 General Introduction 1
Chapter 2 In vitro study of intestinal lipolysis using pH-stat and gas
chromatography 21
Chapter 3 Lipid digestion of protein stabilized emulsions investigated in
a dynamic in vitro gastro-intestinal model system 47
Chapter 4 Effect of oral administration and intragastric distribution of
lipids on lipolysis in gastric and duodenal human fluids 71
Chapter 5 Effect of oral administration and intragastric distribution of fat
on satiety responses in humans 99
Chapter 6 General Discussion 125
Summary 141
Samenvatting 145
Zusammenfassung 151
Acknowledgements 157
About the author 161


Chapter 1
General Introduction

Chapter 1
2
Lipids are omnipresent in foods. Lipids exist in many forms, but from a human
nutritional point of view triglycerides are most important as nutrient part and energy
source. In the context of this thesis the term lipids refers triglycerides. Their
recommended total daily intake is 15-30% (or up to 35 % when eating diets rich in
vegetables, legumes, fruits and wholegrain cereals) of total daily energy intake to prevent
diet-related chronic diseases, i.e. diabetes mellitus, cardiovascular diseases, hypertension
and stroke, some type of cancers and obesity (WHO/FAO report, 2003). Besides its role as
energy source, triglycerides are necessary to enhance the intestinal absorption of
lipophilic nutrients (Brown et al., 2004). In addition, triglycerides are contributing to the
characteristic texture, flavor and mouthfeel of various foods (Drewnowski, 1995;
Drewnowski 2010). The problem is, however, that we consume too much. According to
the WHO, however, in 2008 worldwide more than 1.4 billion adults were overweight of
which approx. one third was obese. Although there is evidence that prevalence of obesity
is leveling off in western countries, there is a rapid rise in obesity in, for example, Asia
(Rokholm et al., 2010).
In view of the current obesity epidemic an in-depth insight in digestion of lipids may
offer leads to help control food intake. Already for a long time, triglycerides are known to
play a role in the regulation of metabolic intake by activating satiety responses. In
western diets, most food products are highly processed and contain lipids mostly in form
of emulsions, for example, as a dispersion of oil in water. The question arises, if
specifically designed emulsions could be a tool to influence satiety responses and thus
help prevent overconsumption. The regulation of these satiety responses by using such
emulsions is, however, not fully understood. This is related to the complexity of food
intake regulation, where food properties, physiological responses and behaviour play a
role. Hence, this thesis aims to provide knowledge on how changes in emulsion
parameters can influence the digestion of triglycerides, specifically the release of free
fatty acids, and how this can be linked to factors regulating satiety responses. Therefore,
the research reported in this thesis starts with the influence of the emulsion parameters
droplet size and interfacial composition, two key factors affecting the digestion of
emulsions. The digestion of emulsions is studied under simulated static intestinal
conditions and dynamic gastrointestinal conditions. Finally, in a human intervention
study the oral administration and intragastric distribution of lipids is investigated to
reveal influences on intestinal lipolysis and resulting satiety responses.

General Introduction
3
The following sections provide information on the current understanding of the
regulation of satiety in response to lipid intake as observed from human studies.
Moreover, the in vitro models used to gain mechanistic understandings of lipid digestion
of emulsions are summarized.
1.1 The impact of fat digestion on satiety responses
1.1.1 General aspects of lipid digestion
Gastric lipolysis
It is considered that human gastric lipase (HGL) contributes to 10-40 % of overall
lipolysis (Carriere et al., 1993; Armand et al., 1999). It was found that HGL is inhibited by
triglyceride degradation products (Pafumi et al., 2002). These products (Fig. 1.1C) are
mainly free fatty acids (FFA) and diglycerides (DG) as HGL has a preference for the sn-3
position of the triglyceride. For example, the FFA concentration to inhibit HGL was found
to be 107 - 122 µmol per surface area (m2) (Pafumi et al., 2002). In addition, gastric
lipolysis is essential for an optimal digestion process in the intestine (for a review see
Armand, 2007). For example, gastric lipolysis and the formation of FFA promote optimal
human pancreatic lipase (HPL) activity. The released long-chain FFA reduce the lag-time
of the HPL-colipase complex to bind to the lipid interface (Gargouri et al., 1986b;
Bernback et al., 1989). Due to the activity of HGL over a broad pH range (Hamosh et al.,
1990), it is still active in the duodenum. As a consequence, it is estimated that up to 7.5 %
of intestinal lipid digestion derives from HGL-activity (Carriere et al., 1993).
Intestinal lipolysis
The major intestinal lipolytic enzyme is HPL, which contributes to 40 -70 % of overall
lipid digestion (Carriere et al., 1993; Armand et al., 1996; Armand et al., 1999) resulting in
FFA and 2-MG (Fig. 1.1C). Co-lipase, a cofactor secreted from the exocrine pancreas, binds
to triglyceride in the presence of bile salts (Borgstrom, 1975). HPL forms a complex with
co-lipase (Van Tilbeurgh et al., 1993; Pignol et al., 2000) and, by including micelles,
consisting of for example, long-chain FFA, bile salts or lysolecithin, it forms a ‘ternary’
complex (Pignol et al., 2000). As this enables the anchoring of pancreatic lipase to the oil-
water interface, it is clear that the composition of the lipid interface plays an important
factor for this interaction.

Chapter 1
4
Fig. 1.1 Gastrointestinal tract (A) and behaviour of emulsions upon consumption (B): i-v indicate
emulsion behaviour which can occur in the intestine as well as in the stomach. Emulsions can flocculate
(i), be stable or, in the intestine, being redispersed (ii), partially coalesced (iii), coalesced (iv) or broken
(phase separated, v). Triglycerides present in emulsions are hydrolysed by human gastric lipase (HGL)
into 1 FFA and 1 DG and by human pancreatic lipase (HPL) into 2 FFA + 1 MG during their passage
through the gastrointestinal tract (C). Figures A and B are adapted from Golding et al. (2010; 2011).
1.1.2 Lipid digestion in relation to satiety responses
Satiation and satiety
Satiation is a term representing a number of processes that terminate an eating period
while satiety refers to processes, which inhibit further eating and thus, affect the interval
between meals (Fig. 1.2) (Blundell & Halford, 1994; Blundell & Tremblay, 1995). Both
processes are regulated (Fig. 1.2) by a cascade of events (Blundell et al., 1994; Blundell et
al., 1995). Metabolic satiation and satiety, for example, include all neural and hormonal
signals between the gastrointestinal tract and the brain (Blundell et al., 2010). These
signals refer to stomach fullness as sensed by mechanoreceptors as well as hormones,
such as Cholecystokinin (CCK), Glucagon-like peptide 1 (GLP-1), Peptide YY (PYY) and
ghrelin (Blundell et al., 2010).

General Introduction
5
Fig. 1.2 Satiety cascade indicating the satiety and satiation response upon food intake (adapted from
Blundell et. al., 2010).
Hormonal regulations
Fullness is a result of stomach distension. Thus, it is likely that prolonging gastric
distension enhances satiety, which in turn could be realized by inhibition of gastric
emptying. Different hormones are involved in the regulation of gastric emptying, such as
CCK, GLP-1, PYY and ghrelin. First of all, CCK is released from I-cells which are most
prominent in the proximal part of the intestine (Fig. 1.1A). With respect to lipid digestion,
the release of CCK requires a fatty acid chain length of at least 12-carbons (McLaughlin et
al., 1999). Only FFA of this chain length or longer were reported to delay gastric emptying
(Hunt & Knox, 1968). CCK stimulates gallbladder contraction and pancreatic secretion
(Wank, 1998), while it is also involved in the inhibition of gastric emptying (Cummings &
Overduin, 2007). It was shown that lipolysis of triglycerides by pancreatic lipase triggered
CCK release (Hildebrand et al., 1998), but the CCK secretion diminishes again after
approximately 60 min, despite continuous delivery of lipids to the duodenum (Fried et al.,
1988). Another hormone, which is secreted upon lipid ingestion, is GLP-1. GLP-1 is
released from L-cells in the distal part of the small intestine and of the colon. Also, PYY is
released from L-cells in the ileum and colon. Both GLP-1 and PYY are considered to be
mediators of the `ileal brake´. This is a mechanism, which regulates gut transit time and
reduces hunger and food intake (Maljaars et al., 2008). Ghrelin is released from endocrine
cells in the stomach and the proximal part of the small intestine. Its release increases food
intake and gastrointestinal motility and its circulating plasma levels are suppressed upon
food intake (Cummings et al., 2007). Also, there is indication that the effects of ghrelin
might be inhibited by CCK, but the mechanisms are not clear yet (Chandra & Liddle,
2007).

Chapter 1
6
Lipid induced satiety responses
Responses to oral fat intake (`cephalic phase´) are considered to induce metabolic
responses and, hence, to induce and regulate the gastrointestinal digestion of foods as
well as food intake. These cephalic responses are mediated by vagal stimulation. With
respect to lipid digestion, oral stimulation in the form of modified sham feeding (MSF), for
example, stimulates gallbladder contraction (Witteman et al., 1993), increased plasma
triglycerides (Robertson et al., 2002; Heath et al., 2004; Smeets & Westerterp-Plantenga,
2006) and hormones, such as insulin (Robertson et al., 2002; Smeets et al., 2006) and
ghrelin (Heath et al., 2004). Also, MSF was found to reduce appetite (Heath et al., 2004)
and increase satiety (Smeets & Westerterp-Plantenga, 2006). However, data on oral fat
exposure, its contribution to intestinal lipid digestion and resulting effects on appetite in
human is preliminary and limited (Mattes, 2002).
Including the gastric passage in in vivo studies, it was shown, that intragastric infused
FFA were five times more potent to induce the release of CCK and PYY as well as the
perception of fullness than TG per gram of the respective lipid (Little et al., 2007). This
points towards the importance of gastric lipolysis. The emulsifier used in that study to
either emulsify oleic acid or macadamia oil was milk protein which will be affected by the
gastric environment and lead to, for example, flocculation of the droplets, The fundus and
the corpus part of the stomach (Fig. 1.1A) store and cause a slow mixing of food with
salivary and gastric secretions (Marciani et al., 2001). Most of the mixing is restricted to
the antral part. Thus, physical instabilities of emulsions will result in creaming and phase
separation (Fig. 1.1B), but also in precipitation (van Aken et al., 2011) (Table 1.1). As a
consequence, these instabilities influence the lipid digestion. Moreover, as the release of
FFA from small droplets is higher than that of large droplets (Armand et al., 1992, Li &
McClements, 2010) (Table 1.1), the surface area should correlate with hormonal responses
and satiety feelings. In fact, an emulsion, which retained a small droplet size in the
stomach led to higher CCK release and gallbladder contractions compared to an emulsion
that phase separated (Marciani et al., 2007; Marciani et al., 2009). In those studies, also the
perception of fullness was higher, while hunger and appetite were reduced for the stable
emulsion compared to the phase separated emulsion. Nevertheless, the relation of gastric
behaviour of emulsions to satiety responses is still not fully understood. For example, a
low amount of fat (i.e. 4 g) that was layered on an orally consumed liquid meal resulted in
a fast emptying of the aqueous phase, whereas the fat layer was emptied delayed into the
duodenum (Foltz et al., 2009). In that study, the increase in plasma CCK and lipid

General Introduction
7
absorption was not consistent with changes in hunger, satiety and fullness perception. A
more recent study showed that when emulsions containing partially hydrogenated fat
were administered, gastric emptying was prolonged compared to emulsions with oil
(Keogh et al., 2011). Although in that study differences in plasma triglyceride, CCK, PYY
and GLP-1 concentrations were observed between the applied emulsions, there was no
difference with respect to satiety and hunger perception.
From duodenal intubation studies it is known that the release of long-chain FFA into
the duodenum stimulates CCK secretion (McLaughlin et al, 1999), which is involved in the
regulation of several gastrointestinal function as described above. It has been shown, that
continuous intraduodenal infusion of emulsified fat stimulates CCK more than non-
emulsified fat (Ledeboer et al., 1999). Later on, it was confirmed that the emulsion droplet
size affects the release of satiety hormones, such as CCK (Seimon et al., 2009). In that
study, the impact was larger for small droplets than for large droplets and the release
correlated with the appetite sensation and antropyloroduodenal motility when presented
intraduodenally. Inhibition of pancreatic lipase by using tetrahydrolipstatin (THL)
eliminated the release of hormones, i.e. CCK (Hildebrand et al., 1998; Feinle et al., 2003),
GLP-1 (Feinle et al., 2003), PYY and ghrelin (Feinle-Bisset et al., 2005), activated
antropyloroduodenal motility (APD), increased food intake (Feinle et al., 2003) and
reduced gallbladder emptying (Hildebrand et al., 1998). Nevertheless, it was also shown
that the reduction of APD and food intake were more potent from fatty acids with a chain
length of C12 (i.e. lauric acid) compared to C18 (i.e. oleic acid). CCK was stimulated
likewise, while the PYY concentrations were higher for C18 (Feltrin et al., 2008) fatty
acids than for C12. Thus, the direct application of emulsions intraduodenal influences to a
large extent metabolic and satiety responses.
1.2 Properties of emulsions to influence lipid digestion
In vivo studies demonstrate that lipid digestion products like free fatty acids can trigger
physiological responses that may lead to satiation. However, it remains unclear how lipid
composition and the physicochemical state of lipids can be used to control digestion in
the first place.
Lipids are present in processed food products as, for example, structural fats (e.g. in
pastry products), bulk fats (e.g. in margarine) and emulsified fats (e.g. in dressings)
(McClements, 2005; Mun et al., 2007). In the emulsified form, droplets are stabilized by

Chapter 1
8
proteins or low molecular weight surfactants, such as Tween. In addition, lipids will be
emulsified during their passage through the gastrointestinal tract due to, for example,
surface active compounds, such as bile salts, which in turn can cause structural changes
of the emulsions (McClements, 2008) (Fig. 1.1B). For example, WPI-stabilized emulsions
were less stable against flocculation and coalescence compared to Tween 20-stabilized
emulsions (Mun et al., 2007). Changes at the interface were demonstrated to cause
coalescence, flocculation or phase separation of the emulsions (Fig. 1.1B). For example,
proteolysis of the protein-covered interface of emulsions by pepsin resulted in weakening
the interface (Sarkar et al., 2009; Golding et al., 2011). Consequently, droplets were more
prone to coalescence. In combination with salts and acidic pH this resulted in flocculation
(Sarkar et al., 2009; Golding et al., 2011). When a fungal lipase was introduced in such a
system the flocculation of protein-stabilized emulsions was enhanced (Golding et al.,
2011). Since a smaller droplet size, and hence a larger surface area, results in more
lipolysis by both human gastric lipase (HGL) and human pancreatic lipase (HPL), as
shown in vitro (Armand et al., 1992; Borel et al., 1994; Li & McClements, 2010) and in vivo
(Armand et al., 1999), the above mentioned changes at the interface are likely to influence
the release of FFA. This of importance as in vivo studies showed that FFA are the key
lipolysis products to cause various satiety responses as introduced above (section 1.1.2)
(Schwizer et al., 1997; Feinle et al., 2001; Feinle-Bisset et al., 2005; Little et al., 2007).
The digestion of these emulsified lipids is also influenced by the ability of lipases to
anchor to the interface. For example, lipolysis by pancreatic lipase is inhibited in the
presence of low molecular weight surfactants, such as Tween 80 (Gargouri et al., 1983;
Pieroni et al., 1990), phospholipids (Wickham et al., 1998; Wickham et al., 2002) and by
protein emulsifiers (Bläckberg et al., 1979). Co-lipase, in the presence of bile salt is able to
restore these inhibitory effects of surface active compounds (Gargouri et al., 1983). Under
simulated intestinal conditions, for example, the extent of lipid digestion of emulsions
stabilized by surfactants, such as Tween 20 and lecithin, was lower compared to the
extent of hydrolysis of emulsions stabilized by proteins, such as casein and whey protein
isolate (WPI), (Mun et al., 2007) (Table 1.1).

General Introduction
9
1.2.1 In vitro models to reveal the release of FFA
As the small intestine is the major site for lipid digestion and absorption, most of
previous in vitro research is conducted by simulating this part of the digestion to
determine the concentration of FFA released. For example, test samples are exposed to
enzyme mixtures, such as pancreatin, and bile salts at neutral pH (Table 1.1). In addition,
various salts, such as NaCl and CaCl2, and surface active compounds, such as
phospholipids, can be added to try and better simulate the in vivo situation, as reviewed
recently (McClements & Li, 2010). The lipolysis in those models is typically followed by
pH-stat titration (Table 1.1) at a pH’s between 6.2 and 8.0. The rate of digestion of lipids
from the investigated emulsions is increasing with increasing calcium, bile or lipase
concentration and with the presence of co-lipase. The pH-stat method will give relative
rates of lipid digestion in the form of FFA release rather than an absolute measure of total
FFA formed (Mun et al., 2006) or the formation of monoglycerides or diglycerides. In vitro
models help to reveal phenomena, that are likely to be of importance in vivo. Using a
dynamic model (Table 1.1), the formation of monoglycerides (MG) and DG were shown to
play a role during lipid digestion. They act as emulsifiers at the oil droplet interface and
participate in micelle formation in the intestine (Reis et al., 2009).
As can be seen in Table 1.1, only a few models simulate gastric digestion or include it,
especially with respect to lipid digestion. In these models, samples are mostly subjected to
an acidic pH, pepsin and salts, such as NaCl. As mentioned above, the addition of pepsin
and salts will enhance coalescence and flocculation of protein-stabilized emulsions. Only
a few recent studies include biopolymers, such as mucus (van Aken et al., 2011;
Kenmogne-Domguia et al., 2012; Marze et al., 2013) or lipolytic enzymes, such as fungal
lipase (Golding et al., 2011; van Aken et al., 2011). In the study of van Aken et al., (2011)
physical changes in the emulsion were also observed: creaming of the protein-stabilized
emulsion as well as precipitation of full fat milk. Although these changes are quite
substantial, no effect was observed on the extent of lipolysis (Table 1.1) as revealed with
gas chromatography. The pH-stat method is not suitable to follow lipolysis under gastric
conditions. To assess the extent of lipid digestion under such acidic conditions, a back-
titration technique can be used. This has been applied after incubation of lipid systems
with e.g. HGL (Gargouri et al., 1986a; Gargouri et al., 1986b). This technique provides the
total concentration of FFA at pH 9. But again, this method will not allow to studying MG
and DG formation and is in our opinion very time consuming.

Chapter 1
10
Overall, there are still many questions related to the way lipids are digested and how
this contributes in controlling food intake. For the study of lipid digestion in vitro models
are important. However, in vitro models are limited in their ability to mimic the digestive
process and lack the ability to mimic physiological responses, such as hormonal induced
feedback mechanisms. Therefore, combining in vitro models with in vivo studies is
considered to be of relevance.
1.3 Aim and outline of the thesis
From the above it is clear, that it is yet not fully understood how emulsions can
regulate satiety responses.. Hence, the aim of this thesis is to provide knowledge on
mechanisms affecting the digestion of lipids, specifically the release of free fatty acids,
and subsequent satiety responses. As a starting point within this project the lipolytic
breakdown of emulsions under intestinal conditions is simulated with a focus on the
effect of two well-known influencing factors: The type of emulsifier used and the droplet
size of the emulsion (Chapter 2). Whey protein isolate (WPI) is used as emulsifier as it is
a common proteinaceous emulsifier in food applications. Also, gum arabic is used as an
emulsifier as it mainly consists of polysaccharides and these have been shown to decrease
the release of free fatty acids. A gas chromatographic method is described, which is
further used to determine FFA, monoglyceride , diglyceride and triglyceride (TG). The
results for all lipolytic products (i.e. FFA, MG, DG) in relation to droplet size and
emulsifiers used are presented. In case of FFA, they are compared with the pH-stat
method. In Chapter 3, the effects of gastric and small intestinal processing of two
differently charged protein stabilized emulsions on gastric and intestinal lipolysis are
investigated under dynamic in vitro conditions. WPI and lysozyme are used as emulsifiers
in order to reveal the impact of flocculation at the isoelectric point under gastric
conditions. To reveal the effect of gastric behaviour on intestinal lipid digestion, a custom
designed gastric compartment, simulating the motility of the different regions of the
stomach, and gradual acidification in the gastric compartment is used. In Chapter 4 the
role of in vivo intragastric distribution of lipids on gastric and intestinal lipolysis in
healthy subjects is investigated. For this purpose, a homogenized emulsion and an un-
homogenized sample are perfused intragastrically. Furthermore, the additive effect of oral
application of the homogenized emulsion on gastric lipolysis is studied. Gastric and
duodenal aspirates are assessed for emulsions structures, pH and lipid compositions (i.e.
FFA, MG, DG and TG) and plasma paracetamol concentrations are evaluated as measure

General Introduction
11
for gastric emptying. In Chapter 5 the impact of gastric and intestinal lipolysis as well as
the oral application on physiological responses related to lipid digestion and satiety were
investigated. To this end, gallbladder contraction and changes in plasma concentrations of
CCK, GLP-1, PYY and ghrelin are studied. These findings are further related to satiety
perceptions and ad libitum intake. In addition, the additive effect of oral application on
the same measures is investigated. In the last chapter (Chapter 6) it is discussed how all
these results obtained help to improving our understanding of lipid digestion in relation
to satiety with focus on the impact of the models used and results from a pig study.
Finally, the impact of the different emulsion parameters on satiety are discussed.

Chapter 1
12

General Introduction
13

Chapter 1
14

General Introduction
15

Chapter 1
16
References
Ali, H., Siddiqui, A. & Nazzal, S. (2010). The effect of media composition, pH, and formulation excipients on the in vitro lipolysis of self-emulsifying drug delivery systems (SEDDS). Journal of Dispersion Science and Technology, 31(2), 226-232.
Armand, M., Borel, P., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1992). Effects of droplet size, triacylglycerol composition, and calcium on the hydrolysis of complex emulsions by pancreatic lipase: an in vitro study. The Journal of Nutritional Biochemistry, 3(7), 333-341.
Armand, M., Borel, P., Pasquier, B., Dubois, C., Senft, M., Andre, M., Peyrot, J., Salducci, J. & Lairon, D. (1996). Physicochemical characteristics of emulsions during fat digestion in human stomach and duodenum. American Journal of Physiology - Gastrointestinal and Liver Physiology, 271 (34)(1), G172-183.
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V. & Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096-1106.
Armand, M. (2007). Lipases and lipolysis in the human digestive tract: Where do we stand? Current Opinion in Clinical Nutrition and Metabolic Care, 10(2), 156-164.
Bernback, S., Blackberg, L. & Hernell, O. (1989). Fatty acids generated by gastric lipase promote human milk triacylglycerol digestion by pancreatic colipase-dependent lipase. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism, 1001(3), 286-293.
Bläckberg, L., Hernell, O., Bengtsson, G. & Olivecrona, T. (1979). Colipase enhances hydrolysis of dietary triglycerides in the absence of bile salts. Journal of Clinical Investigation, 64(5), 1303-1308.
Blundell, J., De Graaf, C., Hulshof, T., Jebb, S., Livingstone, B., Lluch, A., Mela, D., Salah, S., Schuring, E., Van Der Knaap, H. & Westerterp, M. (2010). Appetite control: Methodological aspects of the evaluation of foods. Obesity Reviews, 11(3), 251-270.
Blundell, J. E. & Halford, J. C. (1994). Regulation of nutrient supply: the brain and appetite control. Proceedings of the Nutrition Society, 53(2), 407-418.
Blundell, J. E. & Tremblay, A. (1995). Appetite control and energy (fuel) balance. Nutrition Research Reviews, 8, 225-242.
Bonnaire, L., Sandra, S., Helgason, T., Decker, E. A., Weiss, J. & McClements, D. J. (2008). Influence of lipid physical state on the in vitro digestibility of emulsified lipids. Journal of Agricultural and Food Chemistry, 56(10), 3791-3797.
Borgstrom, B. (1975). On the interactions between pancreatic lipase and colipase and the substrate, and the importance of bile salts. Journal of Lipid Research, 16, 411-417.
Borel, P., Armand, M., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1994). Hydrolysis of emulsions with different triglycerides and droplet sizes by gastric lipase in vitro. Effect on pancreatic lipase activity. The Journal of Nutritional Biochemistry, 5(3), 124-133.
Brogard, M., Troedsson, E., Thuresson, K. & Ljusberg-Wahren, H. (2007). A new standardized lipolysis approach for characterization of emulsions and dispersions. Journal of Colloid and Interface Science, 308(2), 500-507.
Brown, M. J., Ferruzzi, M. G., Nguyen, M. L., Cooper, D. A., Eldridge, A. L., Schwartz, S. J. & White, W. S. (2004). Carotenoid bioavailability is higher from salads ingested with full-fat than with fat-reduced salad dressings as measured with electrochemical detection. American Journal of Clinical Nutrition, 80(2), 396-403.
Carriere, F., Barrowman, J. A., Verger, R. & Laugier, R. (1993). Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans. Gastroenterology, 105(3), 876-888.
Chandra, R. & Liddle, R. A. (2007). Cholecystokinin. Current Opinion in Endocrinology, Diabetes and Obesity, 14(1), 63-67.
Chung, C., Sanguansri, L. & Augustin, M. A. (2008). Effects of modification of encapsulant materials on the susceptibility of fish oil microcapsules to lipolysis. Food Biophysics, 3(2), 140-145.
Cummings, D. E. & Overduin, J. (2007). Gastrointestinal regulation of food intake. Journal of Clinical Investigation, 117(1), 13-23.

General Introduction
17
Drewnowski, A. (1995). Energy intake and sensory properties of food. The American Journal of Clinical Nutrition, 62(5), 1081S-1085S.
Drewnowski A, Almiron-Roig E. Human perceptions and preferences for fat-rich foods. In: Montmayeur JP, le Coutre J, editors. Fat detection: Taste, texture, and post ingestive effects. Boca Raton (FL, USA): CRC Press; 2010.
Feinle, C., Rades, T., Otto, B. & Fried, M. (2001). Fat digestion modulates gastrointestinal sensations induced by gastric distention and duodenal lipid in humans. Gastroenterology, 120(5), 1100-1107.
Feinle-Bisset, C., Patterson, M., Ghatei, M. A., Bloom, S. R. & Horowitz, M. (2005). Fat digestion is required for suppression of ghrelin and stimulation of peptide YY and pancreatic polypeptide secretion by intraduodenal lipid. American Journal of Physiology - Endocrinology and Metabolism, 289(6), E948-E953.
Feinle, C., O'Donovan, D., Doran, S., Andrews, J. M., Wishart, J., Chapman, I. & Horowitz, M. (2003). Effects of fat digestion on appetite, APD motility, and gut hormones in response to duodenal fat infusion in humans. American Journal of Physiology - Gastrointestinal and Liver Physiology, 284(5 47-5), G798-G807.
Feltrin, K. L., Little, T. J., Meyer, J. H., Horowitz, M., Rades, T., Wishart, J. & Feinle-Bisset, C. (2008). Comparative effects of intraduodenal infusions of lauric and oleic acids on antropyloroduodenal motility, plasma cholecystokinin and peptide YY, appetite, and energy intake in healthy men. The American Journal of Clinical Nutrition, 87(5), 1181-1187.
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P. F., Stellaard, F. & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption and increases plasma CCK but has minor effects on gastric emptying and appetite. American Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982-991.
Fried, M., Mayer, E. A., Jansen, J. B. M. J., Lamers, C. B. H. W., Taylor, I. L., Bloom, S. R. & Meyer, J. H. (1988). Temporal relationships of cholecystokinin release, pancreatobiliary secretion, and gastric emptying of a mixed meal. Gastroenterology, 95(5), 1344-1350.
Gallier, S., Tate, H. & Singh, H. (2013). In vitro gastric and intestinal digestion of a walnut oil body dispersion. Journal of Agricultural and Food Chemistry, 61(2), 410-417.
Gargouri, Y., Julien, R., Bois, A. G., Verger, R. & Sarda, L. (1983). Studies on the detergent inhibition of pancreatic lipase activity. Journal of Lipid Research, 24(10), 1336-1342.
Gargouri, Y., Pieroni, G., Rivière, C., Lowe, P. A., Saunière, J.-F., Sarda, L. & Verger, R. (1986b). Importance of human gastric lipase for intestinal lipolysis: an in vitro study. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism, 879(3), 419-423.
Golding, M., Wooster, T. J. (2010). The influence of emulsion structure and stability on lipid digestion. Current Oppinion in Colloid & Interface Science, 15(1-2), 90-101.
Golding, M., Wooster, T. J., Day, L., Xu, M., Lundin, L., Keogh, J. & Clifton, P. (2011). Impact of gastric structuring on the lipolysis of emulsified lipids. Soft Matter, 7(7), 3513-3523.
Gudipati, V., Sandra, S., McClements, D. J. & Decker, E. A. (2010). Oxidative stability and in vitro digestibility of fish oil-in-water emulsions containing multilayered membranes. Journal of Agricultural and Food Chemistry, 58(13), 8093-8099.
Hamosh M. Lingual and gastric lipases: their role in fat digestion. Boca Raton (FL, USA): CRC Press; 1990. Heath, R. B., Jones, R., Frayn, K. N. & Robertson, M. D. (2004). Vagal stimulation exaggerates the
inhibitory ghrelin response to oral fat in humans. Journal of Endocrinology, 180(2), 273-281. Hildebrand, P., Petrig, C., Burckhardt, B., Ketterer, S., Lengsfeld, H., Fleury, A., Hadváry, P. & Beglinger,
C. (1998). Hydrolysis of dietary fat by pancreatic lipase stimulates cholecystokinin release. Gastroenterology, 114(1), 123-129.
Hu, M., Li, Y., Decker, E. A. & McClements, D. J. (2010a). Role of calcium and calcium-binding agents on the lipase digestibility of emulsified lipids using an in vitro digestion model. Food Hydrocolloids, 24(8), 719-725.
Hu, M., Li, Y., Decker, E. A., Xiao, H. & McClements, D. J. (2010b). Influence of tripolyphosphate cross-linking on the physical stability and lipase digestibility of chitosan-coated lipid droplets. Journal of Agricultural and Food Chemistry, 58(2), 1283-1289.
Hunt, J. N. & Knox, M. T. (1968). A relation between the chain length of fatty acids and the slowing of gastric emptying. Journal of Physiology, 194(2), 327-336.

Chapter 1
18
Juhel, C. (2000). Green tea extract (AR25®) inhibits lipolysis of triglycerides in gastric and duodenal medium in vitro. The Journal of Nutritional Biochemistry, 11(1), 45-51.
Kenmogne-Domguia, H. B., Meynier, A., Viau, M., Llamas, G. & Genot, C. (2012). Gastric conditions control both the evolution of the organization of protein-stabilized emulsions and the kinetic of lipolysis during in vitro digestion. Food & Function, 3(12), 1302-1309.
Keogh, J. B., Wooster, T. J., Golding, M., Day, L., Otto, B. & Clifton, P. M. (2011). Slowly and rapidly digested fat emulsions are equally satiating but their triglycerides are differentially absorbed and metabolized in humans. Journal of Nutrition, 141(5), 809-815.
Klinkesorn, U. & McClements, D. J. (2009). Influence of chitosan on stability and lipase digestibility of lecithin-stabilized tuna oil-in-water emulsions. Food Chemistry, 114(4), 1308-1315.
Klinkesorn, U. & McClements, D. J. (2010). Impact of lipase, bile salts, and polysaccharides on properties and digestibility of tuna oil multilayer emulsions stabilized by lecithin-chitosan. Food Biophysics, 5(2), 73-81.
Ledeboer, M., Masclee, A. A. M., Biemond, I. & Lamers, C. B. H. W. (1999). Differences in cholecystokinin release and gallbladder contraction between emulsified and nonemulsified long-chain triglycerides. Journal of Parenteral and Enteral Nutrition, 23(4), 203-206.
Lesmes, U., Baudot, P. & McClements, D. J. (2010). Impact of interfacial composition on physical stability and in vitro lipase digestibility of triacylglycerol oil droplets coated with lactoferrin and/or caseinate. Journal of Agricultural and Food Chemistry, 58(13), 7962-7969.
Lesmes, U. & McClements, D. J. (2012). Controlling lipid digestibility: Response of lipid droplets coated by β-lactoglobulin-dextran maillard conjugates to simulated gastrointestinal conditions. Food Hydrocolloids, 26(1), 221-230.
Li, J., Ye, A., Lee, S. J. & Singh, H. (2012). Influence of gastric digestive reaction on subsequent in vitro intestinal digestion of sodium caseinate-stabilized emulsions. Food and Function, 3(3), 320-326.
Li, Y. & McClements, D. J. (2010). New mathematical model for interpreting ph-stat digestion profiles: Impact of lipid droplet characteristics on in vitro digestibility. Journal of Agricultural and Food Chemistry, 58(13), 8085-8092.
Little, T. J., Russo, A., Meyer, J. H., Horowitz, M., Smyth, D. R., Bellon, M., Wishart, J. M., Jones, K. L. & Feinle-Bisset, C. (2007). Free fatty acids have more potent effects on gastric emptying, gut hormones, and appetite than triacylglycerides. Gastroenterology, 133(4), 1124-1131.
Malaki Nik, A., Wright, A. J. & Corredig, M. (2011). Impact of interfacial composition on emulsion digestion and rate of lipid hydrolysis using different in vitro digestion models. Colloids and Surfaces B: Biointerfaces, 83(2), 321-330.
Maljaars, P. W. J., Peters, H. P. F., Mela, D. J. & Masclee, A. A. M. (2008). Ileal brake: A sensible food target for appetite control. A review. Physiology and Behavior, 95(3), 271-281.
Marciani, L., Gowland, P. A., Spiller, R. C., Manoj, P., Moore, R. J., Young, P. & Fillery-Travis, A. J. (2001). Effect of meal viscosity and nutrients on satiety, intragastric dilution, and emptying assessed by MRI. American Journal of Physiology - Gastrointestinal and Liver Physiology, 280(6), G1227-1233.
Marciani, L., Wickham, M., Singh, G., Bush, D., Pick, B., Cox, E., Fillery-Travis, A., Faulks, R., Marsden, C., Gowland, P. A. & Spiller, R. C. (2007). Enhancement of intragastric acid stability of a fat emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder contraction. American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(6), G1607-G1613.
Marciani, L., Faulks, R., Wickham, M. S. J., Bush, D., Pick, B., Wright, J., Cox, E. F., Fillery-Travis, A., Gowland, P. A. & Spiller, R. C. (2009). Effect of intragastric acid stability of fat emulsions on gastric emptying, plasma lipid profile and postprandial satiety. British Journal of Nutrition, 101(6), 919-928.
Marze, S. & Choimet, M. (2012). In vitro digestion of emulsions: Mechanistic and experimental models. Soft Matter, 8(42), 10982-10993.
Marze, S., Meynier, A. & Anton, M. (2013). In vitro digestion of fish oils rich in n-3 polyunsaturated fatty acids studied in emulsion and at the oil-water interface. Food and Function, 4(2), 231-239.
Mattes, R. D. (2002). Oral fat exposure increases the first phase triacylglycerol concentration due to release of stored lipid in humans. Journal of Nutrition, 132(12), 3656-3662.

General Introduction
19
McClements, D. J., Decker, E. A. & Park, Y. (2009). Controlling lipid bioavailability through physicochemical and structural approaches. Critical Reviews in Food Science and Nutrition, 49(1), 48-67.
McClements, D. J. & Li, Y. (2010). Review of in vitro digestion models for rapid screening of emulsion-based systems. Food and Function, 1(1), 32-59.
McLaughlin, J., Luca, M. G., Jones, M. N., D'Amato, M., Dockray, G. J. & Thompson, D. G. (1999). Fatty acid chain length determines cholecystokinin secretion and effect on human gastric motility. Gastroenterology, 116(1), 46-53.
Mun, S., Decker, E. A., Park, Y., Weiss, J. & McClements, D. J. (2006). Influence of interfacial composition on in vitro digestibility of emulsified lipids: Potential mechanism for chitosan's ability to inhibit fat digestion. Food Biophysics, 1(1), 21-29.
Mun, S., Decker, E. A. & McClements, D. J. (2007). Influence of emulsifier type on in vitro digestibility of lipid droplets by pancreatic lipase. Food Research International, 40(6), 770-781.
Pafumi, Y., Lairon, D., Lechene de la Porte, P., Juhel, C., Storch, J., Hamosh, M. & Armand, M. (2002). Mechanisms of inhibition of triacylglycerol hydrolysis by human gastric lipase. The Journal of biological chemistry, 277(31), 28070-28079.
Pasquier, B., Armand, M., Castelain, C., Guillon, F., Borel, P., Lafont, H. & Lairon, D. (1996). Emulsification and lipolysis of triacylglycerols are altered by viscous soluble dietary fibres in acidic gastric medium in vitro. Biochemical Journal, 314(1), 269-275.
Pieroni, G., Gargouri, Y., Sarda, L. & Verger, R. (1990). Interactions of lipases with lipid monolayers. Facts and questions. Advances in Colloid and Interface Science, 32(4), 341-378.
Pignol, D., Ayvazian, L., Kerfelec, B., Timmins, P., Crenon, I., Hermoso, J., Fontecilla-Camps, J. C. & Chapus, C. (2000). Critical role of micelles in pancreatic lipase activation revealed by small angle neutron scattering. Journal of Biological Chemistry, 275(6), 4220-4224.
Porter, C. J. H., Kaukonen, A. M., Taillardat-Bertschinger, A., Boyd, B. J., O'Connor, J. M., Edwards, G. A. & Charman, W. N. (2004). Use of in vitro lipid digestion data to explain the in vivo performance of triglyceride-based oral lipid formulations of poorly water-soluble drugs: studies with Halofantrine. Journal of Pharmaceutical Sciences, 93(5), 1110-1121.
Reis, P. M., Raab, T. W., Chuat, J. Y., Leser, M. E., Miller, R., Watzke, H. J. & Holmberg, K. (2008). Influence of surfactants on lipase fat digestion in a model gastro-intestinal system. Food Biophysics, 3(4), 370-381.
Robertson, M. D., Mason, A. O. & Frayn, K. N. (2002). Timing of vagal stimulation affects postprandial lipid metabolism in humans. American Journal of Clinical Nutrition, 76(1), 71-77.
Rokholm, B., Baker, J. L. & Sørensen, T. I. A. (2010). The levelling off of the obesity epidemic since the year 1999 - a review of evidence and perspectives. Obesity Reviews, 11(12), 835-846.
Sandra, S., Decker, E. A. & McClements, D. J. (2008). Effect of interfacial protein cross-linking on the in vitro digestibility of emulsified corn oil by pancreatic lipase. Journal of Agricultural and Food Chemistry, 56(16), 7488-7494.
Sarkar, A., Goh, K. K. T., Singh, R. P. & Singh, H. (2009). Behaviour of an oil-in-water emulsion stabilized by [beta]-lactoglobulin in an in vitro gastric model. Food Hydrocolloids, 23(6), 1563-1569.
Schmelz, T., Lesmes, U., Weiss, J. & McClements, D. J. (2011). Modulation of physicochemical properties of lipid droplets using β-lactoglobulin and/or lactoferrin interfacial coatings. Food Hydrocolloids, 25(5), 1181-1189.
Schwizer, W., Asal, K., Kreiss, C., Mettraux, C., Borovicka, J., Remy, B., Guzelhan, C., Hartmann, D. & Fried, M. (1997). Role of lipase in the regulation of upper gastrointestinal function in humans. American Journal of Physiology - Gastrointestinal and Liver Physiology, 273(3 36-3), G612-G620.
Seimon, R. V., Wooster, T., Otto, B., Golding, M., Day, L., Little, T. J., Horowitz, M., Clifton, P. M. & Feinle-Bisset, C. (2009). The droplet size of intraduodenal fat emulsions influences antropyloroduodenal motility, hormone release, and appetite in healthy males. The American Journal of Clinical Nutrition, 89(6), 1729-1736.
Smeets, A. J. P. G. & Westerterp-Plantenga, M. S. (2006). Satiety and substrate mobilization after oral fat stimulation. British Journal of Nutrition, 95(4), 795-801.

Chapter 1
20
Tokle, T., Lesmes, U., Decker, E. A. & McClements, D. J. (2012). Impact of dietary fiber coatings on behavior of protein-stabilized lipid droplets under simulated gastrointestinal conditions. Food and Function, 3(1), 58-66.
Torcello-Gomez, A., Maldonado-Valderrama, J., Martin-Rodriguez, A. & McClements, D. J. (2011). Physicochemical properties and digestibility of emulsified lipids in simulated intestinal fluids: influence of interfacial characteristics. Soft Matter, 7(13), 6167-6177.
van Aken, G. A., Bomhof, E., Zoet, F. D., Verbeek, M. & Oosterveld, A. (2011). Differences in in vitro gastric behaviour between homogenized milk and emulsions stabilised by Tween 80, whey protein, or whey protein and caseinate. Food Hydrocolloids, 25(4), 781-788.
Van Tilbeurgh, H., Egloff, M. P., Martinez, C., Rugani, N., Verger, R. & Cambillau, C. (1993). Interfacial activation of the lipase-procolipase complex by mixed micelles revealed by X-ray crystallography. Nature, 362(6423), 814-820.
Vinarov, Z., Petrova, L., Tcholakova, S., Denkov, N. D., Stoyanov, S. D. & Lips, A. (2012). In vitro study of triglyceride lipolysis and phase distribution of the reaction products and cholesterol: Effects of calcium and bicarbonate. Food and Function, 3(11), 1206-1220.
Wank, S. A. (1998). G protein-coupled receptors in gastrointestinal physiology I. CCK receptors: An exemplary family. American Journal of Physiology - Gastrointestinal and Liver Physiology, 274(4 37-4), G607-G613.
WHO/FAO - Report of the joint WHO/FAO expert consultation (2003). Diet, nutrition and the prevention of chronic diseases. WHO Technical Report Series, No. 916 (TRS 916).
White, D. A., Fisk, I. D., Makkhun, S. & Gray, D. A. (2009). In vitro assessment of the bioaccessibility of tocopherol and fatty acids from sunflower seed oil bodies. Journal of Agricultural and Food Chemistry, 57(13), 5720-5726.
Wickham, M., Garrood, M., Leney, J., Wilson, P. D. G. & Fillery-Travis, A. (1998). Modification of a phospholipid stabilized emulsion interface by bile salt: Effect on pancreatic lipase activity. Journal of Lipid Research, 39(3), 623-632.
Wickham, M., Wilde, P. & Fillery-Travis, A. (2002). A physicochemical investigation of two phosphatidylcholine/bile salt interfaces: implications for lipase activation. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids, 1580(2-3), 110-122.
Witteman, B. J. M., Jebbink, M. C. W., Hopman, W. P. M., Masclee, A. A. M., Lamers, C. B. H. W. & Jansen, J. B. M. J. (1993). Gallbladder responses to modified sham feeding: Effects of the composition of a meal. Journal of Hepatology, 19(3), 465-469.
Ye, A., Cui, J. & Singh, H. (2010). Effect of the fat globule membrane on in vitro digestion of milk fat globules with pancreatic lipase. International Dairy Journal, 20(12), 822-829.

Chapter 2
In vitro study of intestinal lipolysis using
pH-stat and gas chromatography
Helbig, A., Silletti, E., Timmerman, E., Hamer, R.J., Gruppen, H. (2012)
In vitro study of intestinal lipolysis using pH-stat and gas chromatography
Food Hydrocolloids, 28(1), 10-19

Chapter 2
22
Abstract
Developing healthy products requires in-depth knowledge of digestion. This study
focuses on lipid digestion in relation to emulsion properties typically followed by pH-stat.
Although this is a fast and easy method to follow the overall digestion, it provides no
details on lipid digestion products. Thus, the aims of the present study were to use gas
chromatography (GC) to determine all products present during lipolysis, i.e.
monoglycerides (MG), diglycerides (DG) and triglycerides (TG), and to compare this
method with the pH-stat method for free fatty acids (FFA). Fine, medium and coarse
emulsions stabilized with two different emulsifiers whey protein isolate (WPI) or gum
arabic were digested under in vitro intestinal conditions. Although the amount of FFA
increased for both methods for WPI stabilized emulsions, the amount of FFA was 2–3
times higher when determined by GC compared with pH-stat. GC analysis showed
decreasing amounts of MG and DG with increasing droplet size for both emulsions. Molar
ratios of FFA/DG and MG/DG were twofold higher for WPI than for gum arabic stabilized
emulsions. This indicates that the total production of lipolytic products (i.e.
FFA+MG+DG) depends on the droplet size and the emulsifier but their proportions only
depend on the emulsifier. Although pH-stat provides a fast measure of FFA release, it is
influenced by the emulsifier type at the oil–water interface and therefore care should be
taken when interpreting pH-stat results. We suggest combining this method with GC for
accurate FFA determination and further evaluation of all lipolytic products.

In vitro study of intestinal lipolysis
23
2.1 Introduction
Understanding lipid digestion is of crucial importance to the food industry when
designing food products that not only taste good but also regulate food intake. In the
regulation of metabolic intake, free fatty acids (FFA) trigger the release of hormones, such
as cholecystokinin, leading to satiety and a delay in stomach emptying (for a review see
Cummings & Overduin, 2007). FFA are generated from triglycerides (TG) and diglycerides
(DG) by gastric and pancreatic lipase. TG and DG can be present in food products as bulk
fats (e.g. in margarine), structural fats (e.g. in bread) and emulsified fats (e.g. in dressings)
(McClements, 2005; Mun et al., 2007). Moreover, TGs and DGs are emulsified during their
passage through the gastrointestinal tract (GIT) (McClements, 2008). The behaviour of
these emulsified TGs and DGs in the GIT, and particularly their lipolysis, is dependent on
parameters such as fat composition, continuous phase of the emulsion and droplet size.
The influence of the droplet size on lipase activity has been investigated in vitro (Armand
et al., 1992; Borel et al., 1994) and in vivo (Armand et al., 1999) for both gastric and
pancreatic lipase showing that a smaller droplet size leads to a higher lipolysis rate. Other
important key features are the interfacial properties of the fat droplets and a considerable
number of in vitro studies have been carried out in the past on this topic. They show that
bile salts, surfactants (Gargouri et al., 1983; Pieroni et al., 1990), phospholipids (Wickham
et al.,, 1998; Wickham et al., 2002) and protein emulsifiers (Bläckberg et al., 1979) have an
inhibitory effect on the pancreatic lipase activity. Moreover, it has been shown that the
addition of bile salts and co-lipase to those systems is able to restore such inhibitory effect
(Gargouri et al., 1983). Recent studies also investigate the effect of droplet interfacial
properties, as affected by the type of emulsifiers used, to further explore the impact on
lipolysis. For example, it has been shown that pancreatic lipase accessibility on emulsions
stabilized by surfactants (e.g. Tween 20 and lecithin) is much lower than for emulsions
stabilized by proteins (e.g. casein and whey protein) (Mun et al., 2007). However, β-
lactoglobulin (β-lg) was more effective in inhibiting the initial lipolysis than lysolecithin
(Hu et al., 2010). Polysaccharides such as gum arabic, which has been used as emulsifier
during lipase activity assays (Rathelot et al., 1975; Gargouri et al., 1984; Salah et al., 2006),
seem to represent a suitable interface component for lipases to adsorb as well.
Besides the type of emulsifier (surfactants vs. proteins), the role of the charge of the
interfacial layer on the lipase activity has been also investigated but it has not been fully
elucidated yet. In fact, Wickham et al. (1998) showed that there is no specific correlation
between the charge of the emulsion and the lag phase of enzymatic action. However, the

Chapter 2
24
release of FFA has been found to be slightly higher using positively charged emulsifiers
(e.g. lactoferrin) than negatively charged emulsifiers (e.g. β-lg) (Sarkar, Horne & Singh,
2010a).
In vitro models of the GIT are frequently used to understand the biochemical and
physicochemical mechanisms that occur during lipid digestion (McClements et. al, 2009).
In those models, gastric and intestinal lipolysis of samples are simulated either separate or
in combination and different compositions of gastric and intestinal fluids to simulate the
in vivo conditions are used (for review see McClements & Li, 2010). For example, samples
in simulated gastric fluids are exposed to an acidic pH, salts (such as NaCl), biopolymers
(such as porcine gastric mucosa) as well as proteolytic and/or lipolytic enzymes (such as
pepsin and fungal lipase) (Sarkar et al., 2009; van Aken et al, 2011). During simulated
intestinal digestion, samples are subjected to neutral pH, mixtures of enzymes (such as
pancreatin), salts, bile salts and phospholipids (Hu et al., 2010; Hur et al., 2009; Sarkar et
al., 2010a; Sarkar, Horne & Singh, 2010b; Porter & Charman, 2001). In vitro lipolysis is
typically monitored by pH-stat titration to obtain relative rates of lipolysis, by measuring
the amount of FFA (Mun et al., 2006), rather than to get an absolute measure of the extent
of digestion. The pH-stat titration can be carried out either as a direct assay by titrating
ionized FFA with titrant (e.g. NaOH) to maintain a certain constant pH or using a back-
titration. In the latter case, the emulsified lipids are first incubated at a given pH for a
certain time. Then, the pH is rapidly increased with NaOH to pH 9 to stop the reaction
and to favour the release of FFA. Subsequently, the total amount of FFA is assessed by
titration. This technique was used under simulated gastric conditions after incubation of
lipid systems with e.g. human gastric lipase (Gargouri et al., 1986a; Gargouri et al., 1986b).
However, both direct and back titration using pH-stat do not provide any details on
the formation of monoglycerides (MG) and DG which play a role during lipid digestion
acting as emulsifiers at the oil droplet interface and participating in micelle formation in
the intestine (Reis et al., 2009). Moreover, they are probably involved in inhibiting human
gastric lipase activity (Pafumi et al., 2002). Therefore, it is important to monitor the
formation of FFA, MG and DG during digestion.
To our knowledge, only few publications are available in literature reporting the lipase
activity in relation to the simultaneous formation of FFA, MG and DG from emulsions
(Carriere et al., 1991; Armand et al., 1996a, 1996b, 1999; Janssen et al., 2006; Persson et al.
2007). Hydrolysis profiles (i.e. formation of glycerol, MG and DG from Intralipide) of

In vitro study of intestinal lipolysis
25
different gastric lipases in vitro (i.e. human, dog and rabbit gastric lipase) were monitored
using high-performance liquid chromatography (Carriere et al., 1991). Using the same
technique, Persson et al. (2007) could separate and analyse phospholipids, bile acids and
neutral lipids from in vivo fed-state human small intestinal aspirates. Thin-layer
chromatography has also been used to analyse and quantify the TG content and its
digestion products in gastric and duodenal aspirates (Armand et al., 1996a, 1996b, 1999).
Janssen and coworkers (2006) used gas chromatography (GC) to analyse FFA, MG, DG
and TG in oil samples. Although estimations of the hydrolysis rates can be obtained from
the above-mentioned studies, no in-depth research has been conducted so far on the
relation between each single lipid digestion product and the interfacial composition and
emulsion droplet size.
As a starting point for the lipolysis research within our project, we decide to use a
rather simple system with focus on pancreatic lipase activity under specific biochemical
conditions, ignoring the oral or gastric digestion phase both having an important role on
lipid digestion. Moreover, we wanted to establish a GC procedure to determine FFA, MG
and DG to be further used during our whole research and, in the future, to analyse in vivo
digested samples. Therefore, we simulated the lipolytic breakdown of emulsions under
intestinal conditions and focused on the effect of two well-known influencing factors: the
type of emulsifier and droplet size. In this paper, we describe the GC results obtained for
all lipolytic products (i.e. FFA, MG, DG) in relation to droplet size and emulsifiers used
and, in the case of FFA, we compare them with the pH-stat method.
2.2 Materials and methods
2.2.1 Materials
Whey protein isolate (WPI) was purchased from Davisco Foods International (BiPro®,
lot JE216-6-440, Le Sueur, MN, USA, 93% (w/w) protein determined by the Dumas
method) and gum arabic from Caldic Ingredients BV (spray-dried gum acacia 368A, lot
0T060521p, Oudewater, The Netherlands, 2% (w/w) protein content determined by the
Dumas method). Triolein, used as the oil phase, was obtained from Sigma–Aldrich (T7752,
lot 016k0715, approx. 65% practical grade which corresponds to approx. 65% oleic acid
according to the manufacturer, Sigma-Aldrich, St. Louis, MO, USA). The fatty acid profile
of this triolein was determined after conversion into fatty acid methyl esters (FAMEs)
following the AOCS Official method (AOCS Method Ce 1f-96), in the Laboratory of Food

Chapter 2
26
Technology and Engineering, Ghent University, Belgium. It contains the following fatty
acids: myristic acid (C14:0, 2.1%, M), palmitic acid (C16:0, 4.1%, P), palmitoleic acid (C16:1,
5.7%, Pi), oleic acid (C18:1, 74%, O) and linoleic acid (C18:2, 8.6%, L). Furthermore,
triglyceride composition expressed as the percentage of the total triglyceride present was
determined with high-performance liquid chromatography according to Rombaut, De
Clercq, Foubert, and Dewettinck (2009) also in the Laboratory of Food Technology and
Engineering, Ghent University. Pancreatin from porcine pancreas was purchased from
Sigma–Aldrich (P3292, lot 117k1343, 4x USP). Porcine bile extract (BE), containing 49 %
(w/w) bile salts, 5% (w/w) phosphatidyl choline and less than 0.06% (w/w) calcium
according to Zangenberg, Müllertz, Kristensen and Hovgaard (2001b), was obtained from
Sigma–Aldrich. FFA standards, i.e. C14:0, C16:0, heptanoic acid (C17:0), C18:1, C18:2 as
well as monoolein, diolein, and TG standards triolein and trilaurin (TG36) were obtained
from Sigma–Aldrich (>99 % pure). All other chemicals were of analytical grade and
purchased from Merck (Darmstadt, Germany) or Sigma–Aldrich. Solutions were prepared
in demineralized water.
2.2.2 Preparation and characterization of emulsions
WPI and gum arabic solutions were prepared by dissolving powder in 10 mM NaCl
solution and stirred overnight. Triolein, used as the oil phase, was kept at –20 °C and
thawed before use. WPI stabilized emulsions contained 20% (w/w) oil and 1% (w/w)
emulsifier. Gum arabic stabilized emulsions contained 5% (w/w) oil and 7.5% (w/w)
emulsifier. Coarse WPI and gum arabic stabilized emulsions with a surface weighted
mean diameter (d32) of 36 and 21 µm, respectively, were prepared using an Ultra-Turrax
S25 KR 18 G (IKA-Werke, Staufen, Germany) at 6500 rpm for 1 min. Medium-sized
emulsions, WPI (d32=4 µm) and gum arabic (d32=7 µm), were prepared using the Ultra-
Turrax at 21,500 and 13,500 rpm, respectively, for 1 min. Fine emulsions, WPI (d32=0.3 µm)
and gum arabic (d32=1.6 µm), were made from pre-emulsions (Ultra-Turrax at 9500 rpm
for 1 min) and further passed 10 times through a laboratory homogenizer (LH-scope HU-
3.0, Delta Instruments, Drachten, The Netherlands) at a pressure of 180 bar.
Droplet size distributions, d32 and surface area of the emulsion droplets were
determined directly after preparation by dynamic light scattering (Mastersizer Hydro
2000SM, Malvern Instruments, Malvern, UK) at room temperature. Dilution in
demineralized water was applied during the measurements according to other studies
(Vingerhoeds et al., 2005; Silletti et al., 2007; Sarkar et al., 2010b). It has been shown so far

In vitro study of intestinal lipolysis
27
that only in flocculated systems dilution upon water might break aggregates leading to
changes in particle sizes (Silletti et al., 2007).
2.2.3 pH-stat measurements of in vitro intestinal digestion of
emulsions
The digestion of freshly prepared emulsions was monitored using a pH-stat automatic
titration unit (Metrohm, Herisau, Switzerland) at pH 7.5. This pH 7.5 was chosen in order
to be able to compare our data with the results already available in literature (e.g.
Wickham et al., 1998) and moreover because the used emulsions are known to be stable at
this pH.
The emulsions were diluted in a solution consisting of 150 mM NaCl and 5 mM KCl to
obtain a final oil concentration of 10% (w/w) and 2.5% (w/w) for WPI and gum arabic
emulsions, respectively. BE were dissolved in 5 mM Tris buffer (pH 6.9), containing
20 mM CaCl2 and 40 mM NaCl, and subsequently added to the emulsions. The pH of the
BE/emulsion mixture was adjusted to 7.5 with 0.5 M NaOH solution. The reaction was
started by adding pancreatin dissolved in a solution containing 150 mM NaCl and 5 mM
KCl. Final concentrations were therefore 6.4 mg/mL BE, 0.8 mg/mL pancreatin and 110
mM NaCl. The pH was kept constant at 7.5 by addition of 0.5 M NaOH solution while
gentle stirring. Each measurement was done in duplicate. The BE/emulsion mixture
without pancreatin was taken as reference and processed in the same way as the
emulsions with pancreatin. The amount of FFA (mol) was calculated from the number of
moles of NaOH used to keep the pH at 7.5 during digestion.
2.2.4 Extraction and GC measurements of in vitro intestinal
digestion of emulsions
Samples from duplicate measurements under the pH-stat conditions were taken for GC
analysis. A 0.5-mL sample was taken with no enzyme present (t0), and 10 min (t10), 20 min
(t20), and 60 min (t60) after addition of enzyme. Each sample was poured into an extraction
mix consisting of 1 mL ethanol, 1.5 mL diethylether/heptane (DEE/Hep, 1:1 v/v), and
0.1 mL of 2.5 M sulphuric acid in a glass Kimax tube. Sulphuric acid is only used to
protonate FFA and did not lead to additional FFA formation (data not shown). The tube
was closed with a cap, shaken vigorously for 1 min and centrifuged for 5 min at 500×g

Chapter 2
28
and 20 °C. The upper layer was removed with a glass pipette and transferred into a new
glass tube. This extraction was repeated twice with 1 mL of DEE/Hep. The final
supernatant was collected and its volume was brought to 4 mL. Anhydrous sodium
sulphate (100 mg) was added as drying agent. It was necessary to further apply a 1:20
dilution to quantify the TG for WPI stabilized emulsions. After extraction, samples were
stored at –20 °C until further analysis. An internal standard mix (50 µL), consisting of
C17:0 and TG36, was added to obtain a final concentration of 0.4 mg/mL. The reference
mixture of BE/emulsion without pancreatin was subjected to the same procedure. The
values obtained were used as a baseline for the actual digestion process. Hence, lipolysis
at any time point is expressed as the additional amount of product formed.
GC was performed using a Trace GC Ultra (Thermoscientific, Milan, Italy) as follows.
Samples (1 µL) were injected splitless into a liner using a programmed temperature
vaporizing injector with an autosampler (CombiPAL, CTC Analytics, Zwingen,
Switzerland) and analyzed by flame ionization detector. The column used was a
FactorFour capillary column (30 m × 0.32 mm inner diameter (ID), film thickness (dF) =
0.10 µm, VF-5ht UltiMetal, Varian, Middelburg, The Netherlands), connected to an
UltiMetal retention gap (2 m × 0.53 mm ID, dF = 0.10 µm, uncoated, methyl deactivated,
outer diameter 0.8 mm, Varian). Helium was used as carrier gas at a constant flow rate of
2 mL/min. The program started at an initial temperature of 80 °C (1 min) and increased to
400 °C (2 min) at a rate of 10 °C/min over a total oven run time of 35 min.
Peaks were identified by comparing the relative retention times with those of
standards. Quantification of each digestion product was based on standard curves (R2 =
0.99). The GC data was processed using Xcalibur® software (version 2.0,
Thermoscientific, Milan, Italy) and FFA, MG, DG and TG were calculated using the
following molecular masses of the main components: 282.5 g/mol for FFA, 320.4 g/mol for
MG, 602.9 g/mol for DG, and 885.4 g/mol for TG. The hydrolysis rate was calculated
according to the following equation (Rodriguez et al., 2008):
Hydrolysis (%) = 100×FFA/(3×TG0)=100×FFA/(3×TG+2×DG+MG +FFA)
where TG0 is the initial amount of TG (mol) present. The concentration of free glycerol
(G) was calculated based on the following equation (Rodriguez et al., 2008):
G = (FFA – DG – 2×MG)/3

In vitro study of intestinal lipolysis
29
For each time point, mass balances were calculated in % from TG0:
FFA (%) = (FFA/3×TG0)×100
MG (%) = (MG/3×TG0)×100
DG (%) = ((2×DG)/3×TG0)×100
TG (%) = (TG/TG0)×100
The formation of lipolysis products was further expressed in µM (Table 2.1).
In order to compare the effect of emulsion properties on lipolysis, we calculated the
apparent lipase activity. This activity is defined as the amount of FFA released per minute
(µmol/min) at a concentration of pancreatin of 0.8 mg/mL (1 U is 1 µmol FFA
released/min). It was calculated from the initial slope of the FFA released as a function of
time fitted by SigmaPlot software (SigmaPlot® 8.0, Systat Software Inc., San Jose, CA,
USA).
2.3 Results
2.3.1 GC emulsion characterization
When we analyzed the TG composition of the practical grade triolein used to prepare
the emulsions, only 53.7% was identified as triolein (OOO). The remaining 46.3% is a
mixture of TG consisting of LOO (13.8%), PiOO (11.9%), MOO (3.6%), PLO (2.1%), PiOP
(2%), POO (6.5%), LLO (1.2%), PiOL (1.3%), OLM (0.7%) and 3% not identified TG. Although
the TG composition of the oil is known, TG with the same equivalent carbon number
could not be discriminated by GC because they co-eluted in one peak (Fig. 2.1A, insert).
Besides TG, in the initial triolein, we find less than 1% FFA, 6% MG and 21% DG which
might be due to the esterification during its production. Looking at the chromatograms of
the emulsions alone we find about 5% FFA, 3% MG and 16% DG. Probably also during the
emulsification some MG and/or DG were converted in FFA. After mixing the emulsions
with the digestion fluid in the absence of pancreatin (t0), still small amounts of FFA, MG
and DG (Fig. 2.1A) were observed giving an initial amount of TG at t0 of about 80% of the
total oil volume weighed in and present at the beginning of the experiment. In small-sized
WPI stabilized emulsion, for example, FFA concentration at t0 was 175.1 µM; MG and DG
concentrations were 104.0 µM and 133.4 µM, respectively. As expected, these contents did
not change during incubation of the emulsion without enzyme for 60 min. Moreover, the

Chapter 2
30
initial amount of FFA, MG and DG was similar at all time points for medium and coarse
emulsions, for both WPI and gum arabic.
2.3.2 GC identification of lipolytic products
A typical GC chromatogram of the digested fine WPI stabilized emulsion at t60 is
shown in Fig. 2.1B. Chromatograms from gum arabic stabilized emulsions gave
comparable chromatograms and are not shown. The peak indicated as C16:1 in Fig. 2.1A
and B, with elution time of 12.1–12.4 min, contains C16:0 and C16:1; the peak indicated as
C18:1 (elution time 12.8–13.6 min) contains C18:1 and C18:2. Those two major peaks were
used to calculate the total FFA content in the samples. C14:0, present in amounts less than
1%, was not taken into account. The peaks indicated as MG and DG were identified as
monoolein and diolein and eluted between 17.0 and 17.5 min and between 25.8 and 26.4
min, respectively. To calculate the total TG content, all peaks within the elution time of
29.9–32.4 min were considered together. During the digestion, no differences in the ratios
of the TG peaks were found. As indicated in Section 2.2.2, the WPI emulsion
concentration in the mixture was 10% (w/w). Because of this we had to dilute the samples
1:20 to quantify the TG during the digestion process. A chromatogram of this dilution,
also at t60, is shown in Fig. 2.1C.

In vitro study of intestinal lipolysis
31
Fig. 2.1 Chromatograms of digestion mixtures of WPI stabilized emulsions, undiluted 0 min (A), 60 min
undiluted (B) and 60 min diluted 1:20 (C) after digestion. C16:1, palmitoleic acid; C18:1, oleic acid; MG,
monoglyceride; DG, diglyceride; TG, triglyceride; IS1, internal standard C17:0; IS2, internal standard TG36.
M, myristic; P, palmitic, Pi, palmitoleic; O, oleic; L, linoleic acid.

Chapter 2
32
2.3.3 Release of fatty acids: pH-stat versus GC
The production of FFA from emulsions was measured using two methods: pH-stat and
GC at a constant pH of 7.5. In general, duplicate measurements of emulsions had a
maximum variation of 11% for both methods. FFA measured with pH-stat showed a
variation of 24% within the duplicates only in the case of coarse WPI emulsions.
The release of FFA was first investigated for fine WPI stabilized emulsion. In general,
both methods showed a continuous increase in FFA over the digestion time of 60 min
(Fig. 2.2A). However, the amount of FFA measured by GC gives 2–3 times higher values
compared with the pH-stat data (Fig. 2.2A). To confirm the consistency of those results,
we further explored this difference by investigating the influence of droplet size and type
of emulsifier. We found that independently of the droplet size, both methods give
essentially the same results for FFA released from gum arabic stabilized emulsions with
an average correlation factor between the FFA amount obtained with the two methods of
0.9±0.1 (Figs. 2.2A and 2.2B). In contrary, the average correlation factor for WPI stabilized
emulsions, considering the studied emulsions altogether, is 0.3±0.1. This indicates that in
the case of WPI stabilized emulsions, the pH-stat actually underestimates the FFA release
for all droplet sizes under the used conditions.
2.3.4 Lipase activity and formation of lipolytic products
Due to the underestimation of FFA release by pH-stat in case of WPI stabilized
emulsions the lipolytic activity of pancreatin on both emulsions was calculated using only
the GC data. In general, lipolytic activity increases with higher available surface area as
illustrated in Fig. 2.3. Furthermore, comparing both emulsifiers, lipase activity was equal
(i.e. 4.5 U/mg for fine WPI and gum arabic emulsions), although the initial total surface
areas per volume oil in the assays were 75.0 m2 and 3.5 m
2, respectively. This
demonstrates that the emulsifier has a larger effect on the hydrolysis than the surface
area.

In vitro study of intestinal lipolysis
33
Fig. 2.2 Comparison of the release of FFA (µmol) during 60 min digestion (A) of fine WPI stabilized
emulsions with closed symbols determined by GC (●) or by pH-stat (▼) and of fine gum arabic stabilized
emulsions with open symbols determined by GC (V) or by pH-stat (∇). Correlation between the release of
FFA at t0, t10, t20 and t60 determined by pH-stat and GC (B). WPI stabilized emulsions are indicated with
closed symbols (fine, ●; medium, ▼; coarse, ■), and gum arabic stabilized emulsions with open symbols
(fine, V; medium, ∇; coarse, □). Error bars indicate the standard deviations.
In addition to FFA, the formation of MG and DG was calculated as a percentage of
glycerol present in TG0 for fine emulsions, made with both WPI and gum arabic (Fig. 2.4A
and B) and the end point at 100% was assumed. Here, DG seem to reach a maximum in
the gum arabic stabilized emulsion at 47% hydrolysis, whereas for WPI stabilized
emulsions the maximum was calculated to be at 32% hydrolysis. Table 2.1 reports the
quantities of lipolytic products for the different droplet sizes and emulsifiers used. The

Chapter 2
34
amount of FFA increases with time for all emulsions. At each time point, the FFA amount
released decreases with increasing droplet size. MG patterns follow the same trend as DG
when comparing the same time points for the different sizes. As expected, the amount of
TG decreases with time for all droplet sizes, except for medium WPI emulsions and coarse
gum arabic emulsions, where the TG amount remains unchanged with time. Overall,
independent of the droplet size, gum arabic stabilized emulsions have a higher hydrolysis
rate compared with WPI stabilized emulsions.
Fig. 2.3 Lipolytic activity of pancreatin on WPI (●) or gum arabic (▼) stabilized emulsions obtained by
GC as a function of the surface area. Error bars indicate the standard deviations.
Furthermore, we analyzed the ratios between the different digestion products (Table
2.1). The molar ratios of FFA/MG, FFA/DG and MG/DG from digestion of the WPI
stabilized emulsions are similar for all droplet sizes (3.8 ± 0.9, 3.7 ± 0.5 and 1.0 ± 0.3,
respectively). We found similar behaviour in gum arabic stabilized emulsions. Our data,
therefore, indicate that the size of the droplets influences the total production of the
reaction products but not the proportions. In contrast, the ratios of FFA/DG and MG/DG
were affected by the emulsifiers. For gum arabic stabilized emulsions their ratios are
twofold higher than for WPI stabilized emulsions, independent of the droplet sizes. The
ratios stay constant during digestion for 60 min under all conditions tested indicating a
constant lipolytic reaction.

In vitro study of intestinal lipolysis
35

Chapter 2
36
Fig. 2.4 Formation of glycerol (●), MG (�), DG (▼) and decrease of TG (∇) depending on the hydrolysis
level for fine WPI (A) and gum arabic (B) stabilized emulsions. The data are based on glycerol in mol %
and 100% corresponds to the initial moles of TG used within the assays. 100% hydrolysis is assumed as the
end point and indicated with *. Error bars indicate the standard deviations.

In vitro study of intestinal lipolysis
37
2.4 Discussion
2.4.1 Using GC to characterize lipid degradation of emulsions
In literature several in vitro models have been used to study the lipolysis of fats and
oils. The pH-stat is a popular analysis technique in this kind of research because it allows
continuous monitoring of FFA formed. However, the titration method has some
limitations, as highlighted by numerous reviews (see for example Beisson et al., 2000), due
to the need of certain reaction conditions such as pH, ionic strength, BS and calcium
concentration. The pH of the assay must be, for instance, higher or equal of the apparent
pKa value of the FFA and the pKa is, in turn, highly influenced by the ionic strength
(Mattson & Volpenhein, 1966), BS and calcium concentration (Benzonana & Desnuelle,
1968) present within the reaction medium. From a model point of view, that is still far
from the actual situation in vivo as the concentration of BS and calcium are known to be
subjected to physiological oscillation related to the fed state in the human body. Also, the
technique does not provide any information about the different products (i.e. MG and DG)
and their rate of formation or further digestion. This information is important from the
physiological and physical points of view as lipolysis products can affect stomach
emptying (Cummings & Overdiun, 2007). Moreover, they play a role in the emulsification
process and, under stomach conditions, limit fat digestion (Golding & Wooster, 2010).
Considering the above-mentioned reasons, we developed a GC method that allowed us
to analyze lipolysis irrespective of digestion conditions. The method is presented in this
article and, for FFA release, compared, under the same assay conditions, with pH-stat, as
this is a widely used method in lipolysis studies. To evaluate the data obtained by GC, we
determined mass balances (%) based on total FFA present (Table 2.1). Values less than
100% could indicate that, under the given conditions, a further degradation of MG to
glycerol takes place. Although pancreatic lipases are known to be strictly specific to the
hydrolysis of primary ester bonds, sn-2 MG can be hydrolyzed after isomerization to sn-1
MG (Carriere et al., 1991, 1997; Constantin et al., 1960; Desnuelle & Savary, 1963;
Rodriguez et al., 2008). Moreover, glycerol formation is possible under certain in vitro
conditions without the need for isomerization (Borgström, 1964). When we calculated the
amount of glycerol present, a slight increase in glycerol was seen for fine gum arabic and
WPI emulsions (Fig. 2.4). This is in contrast with a previous study (Rodriguez et al., 2008)
in which no glycerol formation of emulsions digested by porcine pancreatic lipase was

Chapter 2
38
observed. However, some cases show mass balances greater than 100%, for which we do
not have any plausible explanation at the present moment.
Other factors that can influence the lipid digestion are the initial amounts of FFA, MG,
and DG present in the oil phase after preparation of the emulsions. The initial amount of
FFA per lipid surface area found in our study was, for example, 8.8 µmol/m2 for fine WPI
emulsions, which is about 13 times lower than the threshold concentration for inhibiting
lipase (Pafumi et al., 2002). In addition, as the amount of FFA increased until t60 and did
not reach a plateau, we can assume that there was no product inhibition. Also, bile salts
present in our digestion medium are known to incorporate FFA and MG and thus prevent
product inhibition as occurs with human gastric lipase (Pafumi et al., 2002).
2.4.2 Difference in FFA release for methods used
As shown in Fig. 2.2A and B, the amount of FFA liberated during digestion and
measured with GC is higher than that observed with pH-stat. Being aware of the
limitations of the pH-stat method as discussed before, it was surprising to find, under the
same assay conditions, differences as those observed with the emulsifiers used. This is, to
our opinion, of interest to the reader as it makes him more aware that relying only on one
technique might not be always the proper choice. We have several suggestions for the
differences found when comparing both methods. In our study, a discrepancy of 60%
between pH-stat and GC was only observed in the case of WPI stabilized emulsions. This
discrepancy has also previously been found by others. For example, Patton and Carey
(1981) reported 50–70% less FFA released at pH <9 when using pH-stat compared with
radioactive labelled TG. It is possible to avoid this issue by titrating to pH 9, a pH at
which all fatty acids are released into solution (Mun et al., 2006, 2007; Sandra et al., 2008;
Sarkar et al., 2010a). However, when investigating the FFA release at pH 6.5–7.5, as also
done in other studies (Hu et al., 2010; Li & McClements, 2010; Wickham et al., 1998;
Zangenberg et al., 2001a), the various pKa values for fatty acids should be taken into
account as they depend on the surrounding media. For example, in the case of oleic acid
(pKa of 9 in pure water), the pKa shifted to 7.8 in rat pancreatic juice with 1 M NaCl
(Mattson & Volpenhein, 1966). Similarly, a shift to pKa 6.4 was caused by medium
consisting of 0.1 M NaCl and 0.5 mM Ca2+ (Benzonana & Desnuelle, 1968). However,
when BS, Ca2+ and NaCl were present in physiological concentrations, 100% of FFA could
be titrated at pH 6.5 (Zangenberg et al., 2001a). The used BS concentration in our study
was 8.2 mM, which is above the critical micelle concentration and close to the fed state

In vitro study of intestinal lipolysis
39
(Wickham et al., 1998; Porter & Charman, 2001). This is important, as recently
demonstrated by Golding et al. (2011), to minimize the difference in FFA release with the
pH-stat method. Additionally, Ca2+
enhances the pancreatic lipase activity as it
precipitates FFA from the droplets surface by formation of calcium-soaps with long chain
fatty acids and can be added in excess in order to control the rate of lipolysis (Zangenberg
et al., 2001a; Zangenberg et al., 2001b). In our study the calcium concentration was 2.7
mM. In relation to the in vivo situation it is known that the calcium-concentration in the
fasted state of the jejunum is 0.5 ± 0.3 mM (Lindahl et al., 1997). However, in the fed state
the calcium content in the intestine is highly determined by the calcium concentration in
the ingested meal (Fordtran & Locklear, 1966; Mansbach et al., 1975; Sheikh et al., 1988)
whereas the basal secretion of calcium is considered to be negligible (Sheikh et al., 1988).
The free calcium concentration depends further on other substances present in the GIT
such as bile salts (Hofmann & Mysels, 1992), mucus or calcium binding proteins as well as
on substances present in the meal (e.g. oxalate or phytin). Therefore, the rather complex
digestion fluid in terms of BS and ions might explain the amount of FFA released from
gum arabic stabilized emulsions in our pH-stat measurements and the comparable results
obtained with GC. However, it does not explain the differences we found for WPI as the
reaction medium for both emulsions was the same.
The difference in values obtained by GC and pH-stat for WPI stabilized emulsions
could also be due to binding of FFA to β-lg. β-Lg is the main component in WPI, a
globular protein of 18 kDa, which might have two potential binding sites for small
hydrophobic molecules such as fatty acids (Frapin et al., 1993). It is possible, that this
binding is disrupted during the extraction step prior GC analysis. However, the β-lg
amount is 10 µmol at the concentration of WPI used, which is too small to be able to bind
1.7 mmol of FFA, which is the difference found between GC and pH-stat.
Another explanation could be that positively charged amino acid residues in WPI
enable more interactions of FFA at the interface than those of gum arabic. Consequently,
the FFA in WPI BE/emulsion mixture are not available for titration but can be extracted
using the GC method. Again, calcium might influence that interaction by precipitating
FFA, which on the other hand does not influence gum arabic stabilized emulsions as the
negatively charged carboxyl groups in gum arabic do not interact with the FFA. In
contrast to our findings, studies on interactions of FFA and gum arabic showed reduced
lipolysis as FFA possibly bind to the protein moiety of gum arabic and prevent FFA
release in the aqueous phase (Fang et al., 2010; Pasquier et al., 1996).

Chapter 2
40
Another factor to try to explain our findings could be the hydrolysis of the protein by
pancreatin, which could contribute to the titration. However, this can be neglected as a
WPI solution, similar in concentration to that used for the emulsions, subjected to the
same assay conditions showed an increase of only 5 µmol per assay. Furthermore, Sarkar
et al. (2010b) suggested a residual lipase activity of bile salts. However, when both
emulsions were incubated without pancreatin, we did not find any production of FFA.
Finally, we excluded a possible buffering capacity of protein in the pH-stat as the amount
of HCl needed to decrease the pH from 7.5 to 6.5 was the same for both emulsions (data
not shown).
2.4.3 Effect of droplet size on FFA, MG and DG formation
Our data clearly showed the effect of droplet size on the rate of FFA release. The
higher lipolytic activity of gastric as well as pancreatic lipase on smaller droplets
compared with larger droplets has been reported by others (Armand et al., 1992; Armand
et al., 1999; Borel et al., 1994; Pafumi et al., 2002). In agreement with our results, Armand
et al. (1999) observed during digestion of emulsions for both gastric and duodenal lipase,
not only higher FFA release from fine emulsions compared with coarse emulsions, but
also higher MG and DG concentrations. In contrast, we found the same ratios of MG/DG
for all droplet sizes for the respective emulsifier, whereas ratios obtained by Armand et al.
(1999) were almost twofold higher for fine compared with coarse emulsions for both
gastric and duodenal digested emulsions. This means that the reaction kinetics in terms of
MG and DG formation in our study under the specific biochemical conditions is
unexpectedly independent from the droplet size.
2.4.4 Effect of emulsifier on FFA, MG and DG formation
The influence of interfacial composition on the formation of FFA, which was
previously found in another study (Mun et al., 2007) as well as the lower influence of
droplet size on gastric lipase activity (Borel et al., 1994) is in agreement with our findings.
In our study, a higher lipase activity was observed during the digestion of gum arabic
stabilized emulsions compared with WPI stabilized emulsions. It is likely that WPI is more
strongly shielding the interface than gum arabic, hindering the pancreatic lipase reaching
the interface. However, the effect of BS, which weakens the protein layer and displaces it
from the oil–water interface (Maldonado-Valderrama et al., 2008), should be more

In vitro study of intestinal lipolysis
41
pronounced with WPI interfaces than gum arabic allowing higher FFA release in this
case.
It is known that C18:1, for example, has a higher affinity to the surface compared with
diolein or triolein (Pafumi et al., 2002), which might cause an increase in FFA
concentration at the oil–water interface. This in turn could cause product inhibition at the
surface of the droplets, which might affect the digestion process once again. However,
under intestinal conditions they are normally removed by BS and phospholipids in the
aqueous phase due to micelle formation. Moreover, β-lg present in WPI could form a
covalent cross-linked interfacial layer and, therefore, make complete displacement by BS
and phospholipids present in the digestion fluid more difficult (Hur et al., 2009). Similar to
β-lg, it is possible that the WPI layer prevents the release of FFA into the aqueous
solution more than gum arabic, causing the discrepancies observed between pH-stat and
GC.
Similar to the higher FFA release, the formation of MG occurred much faster from gum
arabic stabilized emulsions compared with the WPI emulsion. To our knowledge,
differences in DG and/or MG formation for different emulsifiers have not been reported
in the literature. An increase in MG up to 66% hydrolysis was previously reported for a
longer digestion time (Rodriguez et al., 2008). In the case of DG, the maximum value
present in fine WPI stabilized emulsions at 32% hydrolysis is in line with the maximum
DG value reported in the literature (Rodriguez et al., 2008). In addition, the slight increase
in the ratios of MG/DG obtained from WPI stabilized emulsion indicates that an
incubation time longer than 60 min might lead to a higher degradation of DG into MG. So
far it has been shown for various lipase species that the rate of DG production is much
higher than its hydrolysis rate (Carriere et al., 1991; Rodriguez et al., 2008). However, it
seems that the type of emulsifier at the interface also influences DG and MG formation;
for gum arabic stabilized emulsion we found hydrolysis of DG was faster than its
production rate.
At the present stadium of our research, we are not able to clarify the reasons of the
different breakdown of TG into DG, MG and FFA as that one observed in our systems.
One might speculate that the mode of action of the lipase might be influenced by the
location of components (i.e. FFA, MG, DG and TG) caused by different partitioning
coefficient, as has been shown by Pafumi et al. (2002). In our opinion this is rather
hypothetical as this might be influenced by the different properties of oil/water interfaces

Chapter 2
42
in terms of different interfacial viscosity and/or surface tension. In addition, the different
interaction between interface and enzyme, could also lead to a better
accessibility/presence of DG for the lipase in gum arabic stabilized emulsions resulting in
a faster degradation of DG. On the contrary, TG present in the core of the oil phase would
lead to a slower production of DG. Although we have no clear indication for it and
furthermore it was out of the scope of this study, we are still convinced that the
interaction between lipase and the emulsifier at the oil/water interfaces of emulsion
droplets might play a greater role in lipase digestion.
2.5 Conclusions
This study has shown that GC and pH-stat analysis do not always give the same
results in terms of FFA production. In particular, the interface influenced the amount of
FFA obtained by GC or pH-stat. This should be considered when comparing FFA release
from emulsions stabilized by different emulsifiers. The fact that, although several
hypotheses have been tested, a satisfactory solution was not found yet shows that further
research in this area needs to be undertaken. Our results also show a clear effect of the
surface area; the larger the area the higher the rate of digestion. With the GC method, we
showed that gum arabic stabilized emulsions are more easily digested than WPI stabilized
emulsions resulting in a higher release of FFA. In addition, the emulsifiers influenced the
hydrolysis rate of DG into MG, which was faster for gum arabic compared with WPI
stabilized emulsions. The specific conclusions of lipase digestion presented in this paper
are related to the in vitro experimental conditions used during the study. Nevertheless,
the application of the GC method, presented here, to in vivo samples will be of great
relevance since, beside the FFA, also MGs and DGs are simultaneously measurable.
Therefore, the information obtained with the GC method will surely add to the
understanding of the digestion process of emulsion systems not only in vitro but also in
vivo.

In vitro study of intestinal lipolysis
43
References
Armand, M., Borel, P., Pasquier, B., Dubois, C., Senft, M., Andre, M., Peyrot, J., Salducci, J., & Lairon, D.
(1996a). Physicochemical characteristics of emulsions during fat digestion in human stomach
and duodenum. American Journal of Physiology - Gastrointestinal and Liver Physiology,
271(34)(1), G172–G183.
Armand, M., Borel, P., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H., & Lairon, D. (1992). Effects of
droplet size, triacylglycerol composition, and calcium on the hydrolysis of complex emulsions
by pancreatic lipase: an in vitro study. Journal of Nutritional Biochemistry, 3(7), 333–341.
Armand, M., Hamosh, M., Mehta, N. R., Angelus, P. A., Philpott, J. R., Henderson, T. R., Dwyer, N. K.,
Lairon, D., & Hamosh, P. (1996b). Effect of human milk or formula on gastric function and fat
digestion in the premature infant. Pediatric Research, 40(3), 429–437.
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V., &
Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the
human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096–1106.
Beisson, F., Tiss, A., Riviere, C. & Verger, R. (2000). Methods for lipase detection and assay: A critical
review. European Journal of Lipid Science and Technology, 102(2), 133-153.
Benzonana, G., & Desnuelle, P. (1968). Action of some effectors on the hydrolysis of long-chain
triglycerides by pancreatic lipase. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism,
164(1), 47–58.
Bläckberg, L., Hernell, O., Bengtsson, G. & Olivecrona, T. (1979). Colipase enhances hydrolysis of dietary
triglycerides in the absence of bile salts. Journal of Clinical Investigation, 64(5), 1303-1308.
Borel, P., Armand, M., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H., & Lairon, D. (1994). Hydrolysis
of emulsions with different triglycerides and droplet sizes by gastric lipase in vitro. Effect on
pancreatic lipase activity. Journal of Nutritional Biochemistry, 5(3), 124–133.
Borgström, B. (1964). Influence of bile salt, pH, and time on the action of pancreatic lipase; physiological
implications. Journal of Lipid Research, 5(4), 522-531.
Carriere, F., Moreau, H., Raphel, V., Laugier, R., Benicourt, C., Junien, J. L., & Verger, R. (1991).
Purification and biochemical characterization of dog gastric lipase. European Journal of
Biochemistry, 202(1), 75–83.
Carriere, F., Rogalska, E., Cudrey, C., Ferrato, F., Laugier, R., & Verger, R. (1997). In vivo and in vitro
studies on the stereoselective hydrolysis of tri- and diglycerides by gastric and pancreatic
lipases. Bioorganic & Medicinal Chemistry, 5(2), 429–435.
Constantin, M. J., Pasero, L., & Desnuelle, P. (1960). Quelques remarques complémentaires sur l'hydrolyse
des triglycerides par la lipase pancréatique. Biochimica et Biophysica Acta, 43, 103–109.
Cummings, D. E., & Overduin, J. (2007). Gastrointestinal regulation of food intake. Journal of Clinical
Investigation, 117(1), 13–23.
Desnuelle, P., & Savary, P. (1963). Specificities of lipases. Journal of Lipid Research, 4(4), 369–384.
Fang, Y., Al-Assaf, S., Phillips, G. O., Nishinari, K., & Williams, P. A. (2010). Interaction of gum arabic with
fatty acid studied using electron paramagnetic resonance. Biomacromolecules, 11(5), 1398–1405.

Chapter 2
44
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P.
F., Stellaard, F., & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption
and increases plasma CCK but has minor effects on gastric emptying and appetite. American
Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982–G991.
Fordtran, J. S. & Locklear, T. W. (1966). Ionic constituents and osmolality of gastric and small-intestinal
fluids after eating. American Journal of Digestive Diseases, 11(7), 503-521.
Frapin, D., Dufour, E., & Haertle, T. (1993). Probing the fatty acid binding site of β-lactoglobulins. Journal
of Protein Chemistry, 12(4), 443–449.
Gargouri, Y., Julien, R., Bois, A. G., Verger, R. & Sarda, L. (1983). Studies on the detergent inhibition of
pancreatic lipase activity. Journal of Lipid Research, 24(10), 1336-1342.
Gargouri, Y., Julien, R., Sugihara, A., Verger, R., & Sarda, L. (1984). Inhibition of pancreatic and microbial
lipases by proteins. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism, 795(2), 326–331.
Gargouri, Y., Pieroni, G. & Riviere, C. (1986a). Kinetic assay of human gastric lipase on short- and long-
chain triacylglycerol emulsions. Gastroenterology, 91(4), 919-925.
Gargouri, Y., Pieroni, G., Rivière, C., Lowe, P. A., Saunière, J.-F., Sarda, L. & Verger, R. (1986b). Importance
of human gastric lipase for intestinal lipolysis: an in vitro study. Biochimica et Biophysica Acta -
Lipids and Lipid Metabolism, 879(3), 419-423.
Golding, M., & Wooster, T. J. (2010). The influence of emulsion structure and stability on lipid digestion.
Current Opinion in Colloid and Interface Science, 15(1–2), 90–101.
Golding, M., Wooster, T. J., Day, L., Xu, M., Lundin, L., Keogh, J. & Clifton, P. (2011). Impact of gastric
structuring on the lipolysis of emulsified lipids. Soft Matter, 7(7), 3513-3523.
Hofmann, A. F. & Mysels, K. J. (1992). Bile acid solubility and precipitation in vitro and in vivo: the role of
conjugation, pH, and Ca2+ ions. Journal of Lipid Research, 33(5), 617-626.
Hu, M., Li, Y., Decker, E. A., & McClements, D. J. (2010). Role of calcium and calcium binding agents on
the lipase digestibility of emulsified lipids using an in vitro digestion model. Food Hydrocolloids,
24(8), 719–725.
Hur, S. J., Decker, E. A., & McClements, D. J. (2009). Influence of initial emulsifier type on microstructural
changes occurring in emulsified lipids during in vitro digestion. Food Chemistry, 114, 253–262.
Lindahl, A., Ungell, A. L., Knutson, L. & Lennernaes, H. (1997). Characterization of Fluids from the
Stomach and Proximal Jejunum in Men and Women. Pharmaceutical Research, 14(4), 497-502.
Janssen, H. G., Hrncirík, K., Szórádi, A., & Leijten, M. (2006). An improved method for sn-2 position
analysis of triacylglycerols in edible oils and fats based on immobilised lipase D (Rhizopus
delemar). Journal of Chromatography A, 1112(1–2), 141–147.
Li, Y., & McClements, D. J. (2010). New mathematical model for interpreting ph-stat digestion profiles:
impact of lipid droplet characteristics on in vitro digestibility. Journal of Agricultural and Food
Chemistry, 58(13), 8085–8092.
Maldonado-Valderrama, J., Woodward, N. C., Patrick Gunning, A., Ridout, M. J., Husband, F. A., Mackie,
A. R., Morris, V. J., & Wilde, P. J. (2008). Interfacial characterization of β-lactoglobulin
networks: displacement by bile salts. Langmuir, 24(13), 6759–6767.
Mansbach, C. M., Cohen, R. S. & Leff, P. B. (1975). Isolation and properties of the mixed lipid micelles
present in intestinal content during fat digestion in man. Journal of Clinical Investigation, 56(4),
781-791.

In vitro study of intestinal lipolysis
45
Mattson, F. H., & Volpenhein, R. A. (1966). Enzymatic hydrolysis at an oil-water interface. Journal of the
American Oil Chemists' Society, 43(5), 286–289.
McClements, D. J. (2005). Food emulsions: principles, practices, and techniques (2nd ed.). New York: CRC
Press.
McClements, D. J. (2008). Designing food structure to control stability, digestion, release and absorption
of lipophilic food components. Food Biophysics, 3(2), 219–228.
McClements, D. J., Decker, E. A., & Park, Y. (2009). Controlling lipid bioavailability through
physicochemical and structural approaches. Critical Reviews in Food Science and Nutrition, 49(1),
48–67.
McClements, D. J. & Li, Y. (2010). Review of in vitro digestion models for rapid screening of emulsion-
based systems. Food and Function, 1(1), 32-59.
Mun, S., Decker, E. A., & McClements, D. J. (2007). Influence of emulsifier type on in vitro digestibility of
lipid droplets by pancreatic lipase. Food Research International, 40(6), 770–781.
Mun, S., Decker, E. A., Park, Y., Weiss, J., & McClements, D. J. (2006). Influence of interfacial composition
on in vitro digestibility of emulsified lipids: potential mechanism for chitosan's ability to inhibit
fat digestion. Food Biophysics, 1(1), 21–29.
Pafumi, Y., Lairon, D., Lechene de la Porte, P., Juhel, C., Storch, J., Hamosh, M., & Armand, M. (2002).
Mechanisms of inhibition of triacylglycerol hydrolysis by human gastric lipase. Journal of
Biological Chemistry, 277(31), 28070–28079.
Pasquier, B., Armand, M., Castelain, C., Guillon, F., Borel, P., Lafont, H., & Lairon, D. (1996).
Emulsification and lipolysis of triacylglycerols are altered by viscous soluble dietary fibres in
acidic gastric medium in vitro. Biochemical Journal, 314(1), 269–275.
Patton, J. S., & Carey, M. C. (1981). Inhibition of human pancreatic lipase-colipase activity by mixed bile
salt-phospholipid micelles. American Journal of Physiology - Gastrointestinal and Liver
Physiology, 4(4), 328–336.
Persson, E., Löfgren, L., Hansson, G., Abrahamsson, B., Lennernäs, H., & Nilsson, R. (2007). Simultaneous
assessment of lipid classes and bile acids in human intestinal fluid by solid-phase extraction
and HPLC methods. Journal of Lipid Research, 48(1), 242–251.
Pieroni, G., Gargouri, Y., Sarda, L. & Verger, R. (1990). Interactions of lipases with lipid monolayers. Facts
and questions. Advances in Colloid and Interface Science, 32(4), 341-378.
Porter, C. J. H. & Charman, W. N. (2001). In vitro assessment of oral lipid based formulations. Advanced
Drug Delivery Reviews, 50(Supplement 1), S127-S147.
Rathelot, J., Julien, R., & Canioni, P. (1975). Studies on the effect of bile salt and colipase on enzymatic
lipolysis. Improved method for the determination of pancreatic lipase and colipase. Biochimie,
57(10), 1117–1122.
Reis, P., Holmberg, K., Watzke, H., Leser, M. E., & Miller, R. (2009). Lipases at interfaces: a review.
Advances in Colloid and Interface Science, 147–148(C), 237–250.
Rodriguez, J. A., Mendoza, L. D., Pezzotti, F., Vanthuyne, N., Leclaire, J., Verger, R., Buono, G., Carriere, F.,
& Fotiadu, F. (2008). Novel chromatographic resolution of chiral diacylglycerols and analysis of
the stereoselective hydrolysis of triacylglycerols by lipases. Analytical Biochemistry, 375(2),
196–208.

Chapter 2
46
Rombaut, R., De Clercq, N., Foubert, I., & Dewettinck, K. (2009). Triacylglycerol analysis of fats and oils by
evaporative light scattering detection. Journal of the American Oil Chemists' Society, 86(1), 19–
25.
Salah, R. B., Mosbah, H., Fendri, A., Gargouri, A., Gargouri, Y., & Mejdoub, H. (2006). Biochemical and
molecular characterization of a lipase produced by Rhizopus oryzae. FEMS Microbiology Letters,
260(2), 241–248.
Sandra, S., Decker, E. A., & McClements, D. J. (2008). Effect of interfacial protein cross-linking on the in
vitro digestibility of emulsified corn oil by pancreatic lipase. Journal of Agricultural and Food
Chemistry, 56(16), 7488–7494.
Sarkar, A., Goh, K. K. T., Singh, R. P. & Singh, H. (2009). Behaviour of an oil-in-water emulsion stabilized
by β-lactoglobulin in an in vitro gastric model. Food Hydrocolloids, 23(6), 1563-1569.
Sarkar, A., Horne, D. S., & Singh, H. (2010a). Pancreatin-induced coalescence of oil-in-water emulsions in
an in vitro duodenal model. International Dairy Journal, 20(9), 589–597.
Sarkar, A., Horne, D. S., & Singh, H. (2010b). Interactions of milk protein-stabilized oil-in-water emulsions
with bile salts in a simulated upper intestinal model. Food Hydrocolloids, 24(2–3), 142–151.
Sheikh, M. S., Ramirez, A., Emmett, M., Santa Ana, C., Schiller, L. R. & Fordtran, J. S. (1988). Role of
vitamin D-dependent and vitamin D-independent mechanisms in absorption of food calcium.
Journal of Clinical Investigation, 81(1), 126-132.
Silletti, E., Vingerhoeds, M. H., Norde, W. & van Aken, G. A. (2007). The role of electrostatics in saliva-
induced emulsion flocculation. Food Hydrocolloids, 21(4), 596-606.
van Aken, G. A., Bomhof, E., Zoet, F. D., Verbeek, M. & Oosterveld, A. (2011). Differences in in vitro
gastric behaviour between homogenized milk and emulsions stabilised by Tween 80, whey
protein, or whey protein and caseinate. Food Hydrocolloids, 25(4), 781-788.
Vingerhoeds, M. H., Blijdenstein, T. B. J., Zoet, F. D. & Van Aken, G. A. (2005). Emulsion flocculation
induced by saliva and mucin. Food Hydrocolloids, 19(5), 915-922.
Wickham, M., Garrood, M., Leney, J., Wilson, P. D. G., & Fillery-Travis, A. (1998). Modification of a
phospholipid stabilized emulsion interface by bile salt: effect on pancreatic lipase activity.
Journal of Lipid Research, 39(3), 623–632.
Wickham, M., Wilde, P. & Fillery-Travis, A. (2002). A physicochemical investigation of two
phosphatidylcholine/bile salt interfaces: implications for lipase activation. Biochimica et
Biophysica Acta - Molecular and Cell Biology of Lipids, 1580(2-3), 110-122.
Zangenberg, N. H., Müllertz, A., Kristensen, H. G., & Hovgaard, L. (2001a). A dynamic in vitro lipolysis
model. I. Controlling the rate of lipolysis by continuous addition of calcium. European Journal
of Pharmaceutical Sciences, 14(2), 115–122.
Zangenberg, N. H., Müllertz, A., Kristensen, H. G., & Hovgaard, L. (2001b). A dynamic in vitro lipolysis
model. II: Evaluation of the model. European Journal of Pharmaceutical Sciences, 14(3), 237–244.

Chapter 3
Lipid digestion of protein stabilized
emulsions investigated in a dynamic in vitro
gastro-intestinal model system
Helbig A., Silletti, E., van Aken, G.A., Oosterveld, A., Minekus, M., Hamer, R.J., Gruppen
H. (2012)
Lipid digestion of protein stabilized emulsions investigated in a dynamic in vitro gastro-
intestinal model system
Food Digestion, published online

Chapter 3
48
Abstract
This study investigated the effect of gastric passage of protein stabilized emulsions, i.e.
whey protein isolate (WPI) and lysozyme, under dynamic in vitro conditions on both the
gastric and intestinal lipolysis. Emulsions were prepared at neutral pH to enable an
opposite surface charge. Experiments were performed in a multi-compartmental digestion
model (TNO Gastro-Intestinal Model) including a gastric compartment simulating in vivo
conditions, i.e. gradual acidification, mixing, lipolysis and proteolysis. Under gastric
conditions lysozyme-stabilized emulsions remained macroscopically homogenous,
whereas WPI-stabilized emulsions separated into a cream and serum layer. Microscopy
revealed flocculation of both emulsions but larger particles were found for the initial
negatively charged WPI-stabilized emulsions compared to the positively charged
lysozyme-stabilized emulsions. This suggested that creaming was due to larger flocs
formation caused by a change from net negative to net neutral charge as an effect of the
gradual decreasing pH. Analysis of lipid composition, i.e. free fatty acids (FFA),
monoglycerides, diglycerides (DG) and triglycerides revealed mainly FFA and DG in the
gastric compartment. As a result of creaming, the entry of lipids into the small intestinal
part was delayed for WPI-stabilized emulsions. However, the total amount of FFA
released at the end of the experiment was similar for both emulsions. Our results show,
that the charge differences affected the creaming behavior but not the lipase activity on
the two studied emulsions.

In vitro lipid digestion of protein stabilized emulsions
49
3.1 Introduction
Triglycerides (TG) are important and necessary dietary components to enhance the
intestinal absorption of lipophilic nutrients, such as essential fatty acids, fat-soluble
vitamins, and pharmaceuticals. In addition, TG have a clear function as texture and flavor
components in foods. TG have for long been known to play a role in the regulation of
metabolic intake by activating satiety responses such as the release of cholecystokinin
(CCK), the delay in gastric emptying (Lal et al., 2004; Cummings & Overduin, 2007) or the
increase in small intestinal transit time (Maljaars et al., 2008; van Aken, 2010). These
responses are induced in the small intestine by the detection of free fatty acids (FFA),
which are generated by gastric lipase and small intestinal lipolytic enzymes such as
pancreatic lipase, phospholipases and carboxyl ester lipase. A more in depth insight in TG
digestion can lead to a better control of food intake, which is highly desired in view of the
current obesity epidemic.
In the present study, we investigated the gastric and small intestinal processing of two
differently charged emulsions under dynamic in vitro conditions. To this end, we
prepared two protein stabilized emulsions, i.e. whey protein isolate (WPI) and lysozyme-
stabilized emulsions, having opposite net droplets surface charges at neutral pH. It is
known that oppositely charged emulsions as, for instance β-lactoglobulin and lysozyme,
exhibit a different physico-chemical behaviour when mixed with oral fluid, i.e. saliva
(Silletti et al., 2007). Furthermore, the differences in surface charge changes the sensory
behaviour of WPI and lysozyme-stabilized emulsions (Vingerhoeds et al., 2009).
Therefore, it is thought that these two differently charged emulsions might also induce
different gastric-intestinal behaviour and consequently satiety responses.
Gastric conditions can lead to major structural changes of the protein stabilized
emulsions. The fundus and the corpus part of the gastric compartment contribute to a
slow mixing of food with mucus, electrolytes and enzymes (Marciani et al., 2001) and act
more like a storage, whereas mixing takes place mainly in the antral part. This implies
that creaming and phase separation, but also precipitation (van Aken et al., 2011) as a
result of physical instabilities of the emulsions can take place and is likely to influence the
lipid digestion. Accordingly, it has been shown in vivo that fat from emulsions which
break under acid conditions, tends to float to the surface of the gastric lumen (Marciani et

Chapter 3
50
al., 2007; Marciani et al., 2008). As a consequence, the release of fat into the small intestine
is delayed (Marciani et al., 2007; Marciani et al., 2008; Foltz et al., 2009).
Moreover, gastric lipase activity contributes to 10-40 % of overall lipid digestion
(Carriere et al., 1993; Armand et al., 1999) and up to 7.5 % of lipid hydrolysis in the
duodenum (Carriere et al., 1993). Furthermore, gastric lipolysis and the formation of FFA
promote optimal pancreatic lipase action in the small intestine (for a review see Armand,
2007). For example, the released FFA reduce the lag-time for activation of the pancreatic-
colipase complex (Gargouri et al., 1986b; Bernback et al., 1989) by the formation of a
ternary complex (Van Tilbeurgh et al., 1993; Pignol et al., 2000) involving FFA such as
oleic acid (Pignol et al., 2000). Additionally, the released FFA (e.g. C12 and other longer
chain FFA) are detected in the small intestine and lead to a decrease of gastric motility
and emptying, and can signal satiety. These responses, however, might be delayed if an
emulsion creams in the stomach.
Several studies have been conducted using in vitro and in vivo approaches, to tackle
from one side the fundamental understanding of how emulsions are digested and which
parameters are playing a major role in a “simplified” environment (in vitro), as well as
under more complicated conditions such as the human physiological situation. Our choice
was to combine both approaches insuring from one the reproducibility of the in vitro
studies and the dynamic component of the human digestions. To this extent the study
was performed by means of a dynamic model of the gastro-intestinal tract (GIT), which
mimics the major events occurring in the lumen of the GIT (Minekus, 1995). The system
used was based on a simplified version (called “Tiny TIM”) of the TNO gastro-Intestinal
Model (TIM) consisting of a single instead of three small intestinal compartments
(Schaafsma, 2005). The system was provided with a purposely designed gastric
compartment that simulated the motility of the different regions of the stomach. Further,
we used a gradual acidification in the gastric compartment, and, in order to reveal the
impact of flocculation at the isoelectric point (pI), we compared emulsions stabilized by
WPI (pI ~4.9) and lysozyme (pI ~10.5).

In vitro lipid digestion of protein stabilized emulsions
51
3.2 Materials and methods
3.2.1 Materials
Whey protein isolate (WPI) was purchased from Davisco Foods International (BiPro®,
lot JE216-6-440, Le Sueur, MN, USA) and lysozyme from chicken egg-white was from
Belovo SA (lot 1395, Bastogne, Belgium). Extra virgin olive oil was purchased from
Oleificio cooperativo Goccia di Sole Molfetta S.r.l. (Molfetta, Italy). Pepsin from porcine
gastric mucosa (P7012, 600 U/mL) and trypsin from bovine pancreas (T4665, ≥7,500 U/mg)
were obtained from Sigma–Aldrich (St. Louis, MO, USA). Lipase was extracted from lamb
pharyngeal tissue (Lamb Pre Gastric Lipase, Renco, Eltham, New Zealand). Lamb
pharyngeal lipase was selected as an alternative to human gastric lipase, which is not
commercially available. It has a specificity for the sn-3 position of triglycerides
(Villeneuve et al., 1996) and an activity as a function of pH that is similar to that of human
gastric lipase (Moreau et al., 1988). Pancreatin from porcine pancreas (Pancrex V powder)
was bought from Paines & Byrne Limited (Greenford, UK). Bile was obtained from fresh
porcine bile bladders obtained from an abattoir. The bile was pooled on ice immediately
after collection and stored below -18oC. Fatty acid standards, i.e. C14:0, C15:0, C16:0,
C18:1, C18:2 as well as monoolein, diolein, and triglyceride standards triolein and trilaurin
(TG36) were obtained from Sigma–Aldrich (>99 % pure) as well as MSTFA (N-Methyl-N-
(trimethylsilyl)trifluoroacetamide, lot BCBD5976V). All other chemicals were of analytical
grade and purchased from Merck (Darmstadt, Germany) or Sigma–Aldrich. Solutions
were prepared in demineralized water.
3.2.2 Preparation of emulsions
WPI and lysozyme solutions were prepared by dissolving the proteins in a 10 mM
NaCl solution and gently stirring for 1 hour at room temperature. WPI-stabilized stock
emulsions where prepared, which contained 40% (w/w) oil and 1% (w/w) emulsifier.
Lysozyme-stabilized stock emulsions contained 20% (w/w) oil and 1% (w/w) emulsifier.
The stock emulsions were made from pre-emulsions using an Ultra-Turrax, (Polytron®,
Kinematica AG, Luzern, Switzerland, at 1000 rpm for 1 min), which were further passed
through a two-stage laboratory homogenizer (Ariete, Model NS1001L 2K-Panda, GEA
Niro Soavi, Parma, Italy) at a pressure of 250 bar for the first stage and 20 bar for the

Chapter 3
52
second stage, respectively. The resulting WPI and lysozyme stock emulsions were diluted
to 8% (w/w) oil with a solution containing 1% (w/w) WPI or lysozyme, respectively.
3.2.3 In vitro digestion
The Tiny Tim model used in this study is schematically presented in Fig. 3.1. The
gastric compartment (Fig. 3.1A) consisted of a main body, simulating the fundus and
corpus, and a peristaltic section in order to simulate antral mixing. Like all other
compartments, it was composed of a glass jacket with flexible inner walls. Water was
pumped in between the jacket and the flexible wall to control the temperature (37 º) and
to mix and move the contents by changing the pressure on the flexible walls. The
pumping action in the region of the lower antral part and the pyloric sphyncter was
simulated by peristaltic actions of three valves.
To imitate the pre-prandial state, 30 g of simulated gastric fluid (SGF) was initially
present in the gastric compartment. SGF consisted of Pepsin (600 U/mL) and lamb
pharyngeal lipase (160 U/mL), dissolved in an electrolyte solution consisting of 106 mM
NaCl, 30 mM KCl, 2 mM CaCl2 and 18 mM NaHCO3.
Each experiment was started by introducing 230 g of freshly prepared emulsions into
the gastric compartment. In order to mimic the gastric in vivo situation, the pH was
controlled to follow a pre-set curve (Fig. 3.1C) obtained from physiological measurements
conducted for a high fat meal study (Dressman et al., 1990). A changing mixture of 1 M
HCl solution and water were added at a rate of 0.5 mL/min. The SGF was secreted at a
flow rate of 0.5 mL/min for 180 min.
The digest was gradually released from the gastric compartment to the small intestinal
part (Fig. 3.1C). The transit profile of the chyme followed a pre-set curve (Elashoff et al.,
1982), with a halftime of gastric emptying of 80 min. The small intestinal compartment
(Fig. 3.1B) consisted of two mixing segments. In order to simulate the absorption of
nutrients and water by the small intestine, the lumen of this compartment was filtrated
using a filtration unit with a cut-off of 0.05 µm (MiniKros®Plus, M20S-300-01P,
Spectrum®Laboratories, Rancho Dominguez, CA, USA). The simulated duodenal secretion
fluid (SDSF) consisted of 50 mM bile salts and 35 g/L pancreatin in a small intestinal
electrolyte solution (SIES) containing 86 mM NaCl, 8 mM KCl and 2.3 mM CaCl2. The
solutions were separately introduced in the compartment each at a flow rate of 0.5

In vitro lipid digestion of protein stabilized emulsions
53
mL/min for 180 min to follow a pre-set curve (Marteau et al., 1990; Minekus, 1995). Prior
to the experiment, the small intestinal compartment was filled with 60 g of secretion fluid
consisting of 15 g SIES, 15 g of a 7% (w/w) pancreatin solution, 30 g of porcine bile
solution and 2 mg trypsin to simulate the initial intestinal content. The pH of the digest
was kept constant at a set pH 6.5 by automatic titration with 1 M NaHCO3 solution (Table
3.1).
Fig. 3.1 Schematic diagram of the in vitro gastro-intestinal model (TIM) including the gastric (A) and
small intestinal (B) compartment, and the filtration unit. SGF: simulated gastric fluid, SDFS: simulated
duodenal secretion fluid. C: Gastric emptying curve (dotted line) and gastric pH profile (continuous line)
used to control the gastric emptying and pH. D: Schematic schedule of sampling during 180 min of
experiment. Samples were subjected to particle size measurements, microscopic imaging and lipid
composition analysis as described in the methods. GS: gastric samples, SIS: small intestinal samples, FS:
filtrate samples.

Chapter 3
54
The pH profile and the addition of all reagents were recorded. Each digestion process
lasted 180 min and data values are the average of two independent experiments.
Table 3.1 Fluid addition.
Gastric compartment Small intestinal compartment
salts NaCl 106 mM NaCl 86 mM
KCl 30 mM KCl 8 mM
CaCl2 2 mM CaCl2 2.25 mM
NaHCO3 18 mM bile salt ~50 mM
enzymes pepsin 600 U/mL pancreatin 35 g/L
lipase 160 U/mL trypsin ≥7,500 U/mg
pH control 1 M HCl water 1 M NaHCO3
flow rates SGF: 0.5 mL/min SDSF: 0.5 mL/ min
Pre-prandial state 30 g SGF 15 g SIES
SGF, simulated gastric fluid; SDSF, simulated duodenal secretion fluid; SIES, small intestinal electrolyte solution
3.2.4 Physicochemical characterisation of in vitro gastric and
duodenal digestion of emulsions
The behaviour of the emulsions in the gastric compartment was visually monitored
and recorded on digital photo-camera. Samples were taken directly from the lumen of the
different compartments according to the time line (Fig. 3.1D). In particular, samples from
the gastric compartment were taken at the bottom of the compartment at specific time
points and, additionally, after 90 min from the top part of the compartment. The filtrates
were collected by a fraction collector. From each experiment, also the residual material
from the gastric and small intestinal compartment and the samples after rinsing the
system with alcohol were collected to calculate the total recovery of lipids. Each time
point presents the time from the start of the experiment, i.e. when the emulsions were
introduced in the stomach compartment.
Droplet size distributions, surface-weighted-mean diameter (d32) and volume-
weighted-mean diameter (d43) of the emulsion droplets were determined directly after
preparation and at various time points by dynamic light scattering (Mastersizer Hydro
2000S, Malvern Instruments Ltd, Malvern, UK) at room temperature. Microscopic images
of the initial emulsions and of the emulsions in different parts of the model at different

In vitro lipid digestion of protein stabilized emulsions
55
time points during the digestion period were taken using a BYO-503T laboratory
microscope equipped with a DCM 500 5.0 Mp ocular camera (Byomic).
Samples were analyzed for free fatty acids (FFA), mono-, di- and triglycerides (MG,
DG, TG) composition by gas chromatography (GC) as previously described (Helbig et al.,
2012) with minor modifications. Briefly, 2 mL samples were taken from each
compartment and filtrates at intermediate time points, immediately frozen in liquid
nitrogen and stored at -80 ºC until further analysis. After extraction, the final supernatant
was collected and its volume was brought to 4 mL with Diethylether/Heptane (1:1, w/w).
An internal standard mix (50 µL), consisting of C15:0 and TG36, was added to obtain a
final concentration of 0.4 mg/mL. Prior to analysis, 400 µL of each sample was derivatized
with 150 µL MSTFA, gently mixed and left at least 3h for reaction.
Since the digestion model, used in the present study, can be considered as an in-
between situation of the in vitro and in vivo digestion process two levels of hydrolysis
(L1% and L2%, respectively) were calculated according to Capolino et al. (Capolino et al.,
2011). The level of hydrolysis (L1%), occurring in in vitro experiments, was calculated
according to the following equation:
L1 %=100∗FFA
3∗TG+2∗DG+MG +FFA
As the absorption of lipolytic products in the intestine requires only the hydrolysis of 1
TG molecule into 1 MG and 2 FFA molecules, the physiological relevant level of
hydrolysis (L2%) was calculated based on the equation (Carrière et al., 2005):
L2 %=100∗FFA+MG
3∗TG+2∗DG+MG+FFA

Chapter 3
56
3.3 Results
3.3.1 Behaviour of emulsions in the gastric and intestinal
compartment
In the gastric compartment, the positively charged lysozyme-stabilized emulsions
appeared homogenous on a macroscopic level throughout the experiment (Fig. 3.2A). In
contrast, the negatively charged WPI-stabilized emulsions showed an upper cream layer
and a lower serum layer after approximately 35 min (Fig. 3.2 B-D). This creamed
appearance remained until the gastric compartment was completely emptied. As only in
the case of WPI-stabilized emulsions, a lower concentration of flocs was found in the
bottom part, we define it as serum layer throughout the manuscript, as the presence of
low particle concentrations is typical for such a droplet-depleted layer. Likewise, we
define the layer in the top part in case of the WPI-stabilized emulsion as cream layer as it
is a droplet-enriched layer. Both emulsions flocculated in the gastric compartment as
shown in the microscopic pictures (Fig. 3.3).
The freshly prepared lysozyme and WPI-stabilized emulsion droplets had similar
droplet sizes with a d32 of 2.8 µm and 2.7 µm, respectively (Table 3.2). During the first 20
min of gastric residence, both lysozyme and WPI-stabilized emulsions exhibited
monomodal particle size distributions (Fig. 3.4). The d32 and d43 of lysozyme-stabilized
emulsions increased during the first 20 min (Table 3.2). After 20 min, the particle size
distributions became narrower to exhibit one sharp peak between 1-10 µm (Fig. 3.4). At
t=90 min, a broadening in particle size distribution occurred in the WPI-stabilized
emulsions (1-100 µm) which is reflected in the increase of the d32 and d43 (Table 3.2). To
characterize the creamed WPI-stabilized emulsions, particle size distributions and
microscopic pictures were analyzed from the bottom and top part of the gastric
compartment. As shown in Figure 3.4 (insert), particle size distributions of both parts
were found to be similar for both emulsions. Comparing the microscopic pictures
obtained from the bottom and top part it can be seen that both fractions showed
flocculation of the emulsions (Fig. 3.4, insert).

In vitro lipid digestion of protein stabilized emulsions
57
Fig. 3.2 Behavior of lysozyme- (A) and WPI- (B-D) stabilized emulsions in the gastric compartment after
adding the emulsion to the gastric compartment (0 min) and at=35 min and t=60 min of digestion. The
arrow indicates the separation of the WPI-stabilized emulsion into cream and serum layer.
Fig. 3.3 Light microscopic characterization of lysozyme- (A) and WPI- (B) stabilized emulsions (0 min) in
the gastric compartment at t=20, 135 and 180 min of digestion. Below each picture is the corresponding
pH, scale bar is 50 µm.

Chapter 3
58
Fig. 3.4 Particle size distribution of lysozyme- (A) and WPI- (B) stabilized emulsions at t=20 (∆), 90 (□),
135 (×) and 180 (o) min of digestion in the gastric compartment compared to freshly prepared emulsions
(●). Inserts show the comparison of particle size distributions between the top part (TP, closed symbols)
and bottom part (BP, open symbols) of the gastric compartment at t=90 min of digestion.
Fig. 3.5 Light microscopic characterization of lysozyme- (A) and WPI- (B) stabilized emulsions in the
small intestinal compartment at t=50, 110, 155 and 180 min of digestion. Below each picture is the
corresponding pH, scale bar is 10 µm.

In vitro lipid digestion of protein stabilized emulsions
59
When the digest reached the small intestinal part, the emulsions completely
deflocculated. In contrast to flocculation behaviour exhibited in the gastric compartment,
coalescence was observed for both emulsions in the small intestinal compartment (Fig.
3.5).
Within 110 min of intestinal digestion of lysozyme-stabilized emulsions, a shift to
larger particle sizes was found as well as a broadening of the peaks (Fig. 3.6). Particles in
the range of 50-500 µm were found only at t=50 and t=110 min and likewise the d43 was 6-
fold and 5-fold higher, respectively, compared to the d43 at t=0 (Table 3.2). At t=155 min,
the particle size distribution of lysozyme-stabilized emulsions reduced in size again. In
contrast, at t=50 min of intestinal digestion, the distribution of WPI-stabilized emulsions
became multimodal with very small droplets appearing (0.02-0.4 µm). While the particle
size of lysozyme-stabilized emulsions decreased with time, a broadening and a shift to
larger particles occurred during 180 min of small intestinal digestion of WPI-stabilized
emulsions.
Table 3.2 Surface-weighted-mean diameter (d32) and volume-weighted-mean diameter (d43) of lysozyme
and WPI-stabilized emulsions at different time points of digestion in the gastric and small intestinal
compartment compared to freshly prepared emulsions (t=0 min).
sample Time point (min)
lysozyme- stabilized emulsions
WPI- stabilized emulsions
d32 (µm) d43 (µm) d32 (µm) d43 (µm)
emulsion 0 2.83 5.82 2.74 4.66 GS 20 8.94 12.60 3.34 6.63
90 7.17/7.65* 11.82/12.17* 10.47/11.49* 16.85/26.70*
135 6.74 7.63 9.19 15.11
180 7.40 12.29 12.18 21.64 SIS 50 8.67 34.66 0.59 6.88
110 9.65 26.99 3.11 5.03
155 9.07 15.99 4.27 10.36
185 6.66 11.27 7.59 16.48
GS: gastric samples, SIS: small intestinal samples, * indicates the value obtained from the cream phase

Chapter 3
60
Fig. 3.6 Particle size distribution of lysozyme- (A) and WPI- (B) stabilized emulsions at t=50 (∆), 110 (□),
155 (×) and 180 (o) min of digestion in the small intestinal compartment compared to freshly prepared
emulsions (●).
3.3.2 Formation of lipolytic products in the gastric and duodenal
compartment
The quantitative analysis of TG and lipolytic products, i.e. FFA, MG and DG, was
conducted by means of GC on the same samples characterized by microscopy and particle
size analysis. When the emulsions were subjected to the gastric environment, the amount
of TG decreased continuously in time (Fig. 3.7) with only residual amounts of TG present
in the stomach after 180 min. This is partly due to the emptying of the compartment and
partly to the TG hydrolysis. The amount of FFA and DG slightly increased within the first
20 min (Fig. 3.7). The gastric hydrolysis levels L1 and L2 of TG for both emulsions and for
each time point did not exceed 12 and 15%, respectively.

In vitro lipid digestion of protein stabilized emulsions
61
Fig. 3.7 Comparison of lipid compositions, i.e. FFA (●), MG (▲), DG (■) and TG (×), in the gastric
compartment from lysozyme- (A) and WPI- (B) stabilized emulsions taken at the bottom. Error bars
indicate standard deviations.
As we have observed creaming for one of the two emulsions, we have performed GC
measurements at t=90 min to compare the distribution of fat components between the
bottom and the top of the stomach compartment. In the case of lysozyme-stabilized
emulsions, values of FFA, MG, DG and TG of the bottom part were found to be 0.8, 0.2, 0.6
and 7.6 mmol, respectively and those ones of the top part were 1.1, 0.3, 0.8 and 11.7 mmol,
respectively.
In contrast, the difference between the bottom and top part was more pronounced in
WPI-stabilized emulsions. The amount of FFA, MG, DG and TG of the bottom part were
found to be 0.6, 0.1, 0.4 and 2.9 mmol, respectively, whereas values of the top part were
2.5, 0.4, 1.7 and 13.9 mmol, respectively. The results indicate that during digestion, when

Chapter 3
62
creaming is occurring, WPI-stabilized emulsions have higher levels of TG and lipolytic
products on top of the stomach compartment. Nevertheless, the digestion rate (L1) of the
top part compared to the bottom part was similar, i.e. 6.4 ± 1.3 and 7.4 ± 0.0%,
respectively. The higher amount of FFA, DG and TG present at t=90 min in the top part of
the stomach from WPI-stabilized emulsions will be released at a later stage into the
intestine.
In the overall intestinal digestion of the emulsions the amount of FFA increased
rapidly up to a final concentration of about 13 to 14 mmol (Fig. 3.8), which is about 4.6
times higher than the maximum amount produced in the stomach.
Fig. 3.8 Comparison of lipid compositions, i.e. FFA (●), MG (▲), DG (■) and TG (♦), in the intestinal
compartment from lysozyme- (closed symbols) and WPI- (open symbols) stabilized emulsions. Error bars
indicate standard deviations.
At the first sampling point (i.e. t=50 min) the FFA in the small intestine from both
emulsions were equal to the amount present at t=20 min in the stomach (Figs. 3.7, 3.8).
Higher amounts of FFA were obtained from lysozyme-stabilized emulsions than from
WPI-stabilized emulsions within the first 110 min (Fig. 3.8). Additionally, TG from
lysozyme-stabilized emulsions were present in rather constant amount during the
experiment, whereas the TG of WPI-stabilized emulsions increased after 155 min. The

In vitro lipid digestion of protein stabilized emulsions
63
hydrolysis levels L1 and L2 in the small intestine were about 62% and 76%, respectively for
both emulsions.
3.4 Discussion
3.4.1 Influence of gastric conditions on the emulsion behaviour
As observed in our experiments, the prepared negatively charged WPI emulsion and
positively charged lysozyme-stabilized emulsions flocculated in the gastric compartment
but only the WPI-stabilized emulsions exhibit a pronounced cream layer formation. In our
opinion, the cream layer is produced by differences in flocculation of the two emulsions
as clearly illustrated from the observed particle size distribution (Fig. 3.4). Despite the
applied dilution, inevitable during measurement and inherent to the used instrument, the
WPI-stabilized emulsions contained mainly larger particles indicating the presence of
larger aggregates with a high fat content as possible causes for the cream layer. It is
known that in emulsions systems, in the absence of polymers, flocculation can occur if
the electrostatic repulsion between droplets decrease and cannot contrast the Van der
Waals interaction between droplets. This occurs, for example, when the pH of the
emulsions approaches values close to the isoelectric point (pI) of proteins at the oil/water
interface and when the net charge on the droplets is almost zero. Flocculation of WPI-
stabilized emulsions at pH 4.9 is known as the pH is close to pI of β-lactoglobulin (β-lg),
which is the main component in WPI. Besides the effect of pH, flocculation is also
induced by salts from the gastric secretion. Typically, salts decrease electrostatic
repulsion between droplets by screening the droplets surface charges. The combination of
salts and transitions through the pI has been shown recently to cause flocculation of WPI-
stabilized emulsions (Golding et al., 2011). Lysozyme-stabilized emulsions are instead
always positively charged in the applied pH range as the pI of lysozyme is about 11.
Possibly, these two charge related effects had a larger impact on the flocculation and the
consequent creaming behavior of WPI-stabilized emulsions compared to lysozyme-
stabilized emulsions under gastric conditions (Fig. 3.2). Therefore, the presence of salt
could be a major factor causing the observed less pronounced flocculation of lysozyme-
stabilized emulsions as shown from the particle size analysis, since they screen the
positively charged emulsions droplets. However, it has been shown recently, that the
most important factor which determines emulsion stability is the hydrolysis of the
protein interface by pepsin (Singh & Sarkar, 2011). In fact, the addition of pepsin to β-lg

Chapter 3
64
or WPI-stabilized emulsions, in the presence of salts and at acidic pH, accentuated
flocculation and caused slight coalescence of the emulsions droplets (Golding et al., 2011;
Sarkar et al., 2009) since pepsin hydrolyses the β-lg interfacial layer at the oil-water
interface resulting in a loss in positive charge, peptide formation and a reduction of the
interfacial thickness (Sarkar et al., 2009; Malaki Nik et al., 2010; Nik et al., 2010). The
formed peptides, for example of ~ 10-15 kDa (Sarkar et al., 2009), are less sufficient to
provide steric and electrostatic hindrance thus leading to flocculation of the droplets.
When evaluating the peptides formation by pepsin using the Peptide Cutter tool available
on Expasy platform (http://web.expasy.org/cgi-bin/peptide_cutter/peptidecutter.pl) β-lg
originates 36 peptides with molecular masses varying from 131,17 Da of the ammino acid
leucin up to 2613,07 Da of the longest peptide QKWENGECAQKKIIAEKTKIPAV.
Pepsin digestion of lysozyme originates, instead, 15 peptides, presenting in general
higher molecular masses compared to those formed for β-lg reaching a maximum of the
4939 Da for the sequence SSDITASVNCAKKIVSDGNGMNAWVAWRNRCKGTDVQ
AWIRGCRL. Therefore, due to the differences in type, length and amount of peptides
produced by pepsin digestion, lysozyme stabilized interfaces might be less susceptible to
hydrolysis. This in turn causes less flocculation and consequently less creaming.
Additionally, as hydrolysis of the protein by pepsin weakens the viscoelastic interface
(Golding et al., 2011; Sarkar et al., 2009) the increase in droplets size for both emulsions
found in the present study will be to some extend due to slight coalescence.
Further, the addition of lipase and the resulting formation of FFA will contribute to an
accentuate flocculation of the emulsions as recently shown for WPI-stabilized emulsions
upon addition of a fungal lipase (Golding et al., 2011). Surface active components such as
FFA, MG and DG are thought to displace the stabilizing protein layer and change
therefore the interfacial properties of the emulsions. For example, 2-MG have been found
to be highly surface active and able to dominate the interfacial layer (Reis et al., 2008). As
we found only traces of MG we do not expect them to cause major changes in the
interfaces. However, the amount of FFA and DG of lysozyme-stabilized emulsions at t=20
min, i.e. 441.25 and 144.0 µmol/m2, respectively, might be sufficient to change the
interfacial properties. Since the values of FFA and DG are lower in WPI-stabilized
emulsions at t=20 min, i.e. 123.1 and 60.9 µmol/m2, respectively, their ability to influence
the interface might be less compared to lysozyme-stabilized emulsions.

In vitro lipid digestion of protein stabilized emulsions
65
During the digestion in the small intestine, we found an increase in droplet size for
WPI-stabilized emulsions as a result of coalescence. Coalescence of WPI-stabilized
emulsions under in vitro intestinal conditions was previously attributed to the gastric
proteolysis of the interface by pepsin and the resulting peptide formation when the
emulsions were incubated before under simulated gastric conditions (Malaki Nik et al.,
2011). The authors suggested, that a further proteolysis of WPI (and its peptides) by
trypsin and chymotrypsin, both present in pancreatin, together with a displacement of
the interfacial layer by bile salts (BS) and phospholipids (PL) caused the increase in the
sensitivity to coalescence under duodenal conditions (Malaki Nik et al., 2011). Therefore,
both the addition of pepsin and porcine bile in our study will have contributed to the
occurred coalescence.
Beside coalescence, we observed smaller particle with d32 < 1 µm at t=50 min for WPI
stabilized emulsion. This finding is also in line with Malaki et al. (Malaki Nik et al., 2011)
where addition of phospholipids was hold to be responsible for the appearance of
particles <1 µm.
3.4.2 Influence of creaming on intestinal lipid digestion
The separation into a cream and serum layer of WPI-stabilized emulsions within the
gastric compartment, as reported in Figure 3.2, was observed to cause a delay of their
intestinal hydrolysis.
Firstly, the higher amount of FFA and DG present at t=90 min in the cream layer from
WPI-stabilized emulsions will be released at a later stage into the small intestine.
Probably, gastric lipolysis will not proceed further in this layer, due to the low pH and
possible product inhibition of the enzyme (Pafumi et al., 2002). In fact, only within the
first 20 min FFA and DG increased. This time span corresponded to the time necessary for
the digestion process to reach a pH between 4 and 5, which is the optimal pH for the lamb
pharyngeal lipase (Moreau et al., 1988). Also, the amount of FFA per surface area (m2) as
shown in section 3.4.1., was above the FFA concentration to inhibit HGL, i.e. 107 - 122
µmol/m2 (Pafumi et al., 2002).
Secondly, the lower amounts of lipid products in the intestine at t=110 min (Fig. 3.8)
reflected the later entry of fat when WPI-stabilized emulsions are compared to lysozyme-
stabilized emulsions. This is due to the creaming in the stomach compartment since the

Chapter 3
66
activation of pancreatic lipase by FFA requires only very small concentration in the oil,
i.e. less than 1 % (Gargouri et al., 1986b).
Since satiety and related physiological responses such as hormone secretion or gastric
emptying are related to the concentration and rate of delivery of FFA in the small
intestine, it could be expected that those responses will be different for the two types of
emulsions. It is known that layering of fat in the stomach has implications in vivo, i.e. the
fat layer empties deferred into the duodenum (Foltz et al., 2009). In such case, both the
release of FFA into the small intestine as well as the release of satiety hormones, such as
cholecystokinin (CCK), will be delayed. Since such a behavior cannot be explored with
the digestion model used in our study, we could only speculate that the digestion of
lysozyme-stabilized emulsions would have an earlier influence on satiety than WPI-
stabilized emulsions. Nevertheless, the presence of FFA, found in our study in the serum
layer of WPI-stabilized emulsions, might still trigger the release of CCK and delay gastric
emptying in vivo.
Although to our knowledge, concentration related studies to clearly evaluate the FFA
threshold for hormone secretion have not been performed yet, it has been reported that
such low amount of FFA as 0.025 mM in the stomach were able to trigger a CCK response
(McLaughlin et al., 1999). This FFA concentration induced the release of about 4 pM of
CCK. Little and coworkers (Little et al., 2007) obtained the same amount of CCK released
with FFA concentrations of 40 mM present in the small intestinal lumen. In the study of
Little et al., this amount of CCK was related to a higher feeling for fullness, a decreased
gastric emptying and a suppressed energy intake. In our case, the amount of FFA present
at all different time points and for both emulsions was largely above 0.025 mM. Therefore,
although creaming occurs we would not expect different effects on the CCK release in
vivo between the two emulsions.
Finally, it must be noted, that the rate of gastric emptying in the Tiny TIM is
programmed and not influenced by the nature of the emulsions. Hence, feedback
mechanisms due to FFA detection in the small intestine occurring in vivo were not taken
into account. Those mechanisms must be studied in vivo. In addition, the used Tiny TIM
model was built to remove the lipolytic products by filtration in order to avoid product
inhibition of the pancreatic lipase. Thus, in theory an over-efficient removal of FFA could
unrealistically reduce the rate of intestinal lipolysis, since pancreatic lipase needs a very
low concentration of FFA. Additionally, the model might not be realistic in the selectivity

In vitro lipid digestion of protein stabilized emulsions
67
and speed of transport of the FFA incorporated in bile micelles through the mucus lining
and enterocytes as it occurs in vivo.
3.5 Conclusions
It can be concluded that the difference in creaming behavior during gastric digestion
delays the intestinal lipid digestion. This is due to the separation into a cream and serum
layer caused by pronounced flocculation of WPI-stabilized emulsions. The total amount of
FFA produced in the small intestinal compartment at the end of the experiment is the
same for both emulsions. This seems to imply that the charge differences are only
affecting the creaming behavior but not the enzymatic activity of the lipase on the two
studied emulsions. From the detected amount of FFA we would not expect any difference
in CCK induced satiety responses in vivo despite the creaming of the WPI-stabilized
emulsion.
Acknowledgements
The authors gratefully acknowledge Esther Bomhof, Franklin D. Zoet (NIZO food
research) and Hans Kooijman (TNO Triskelion) for their technical assistance.

Chapter 3
68
References
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V. &
Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the
human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096-1106.
Armand, M. (2007). Lipases and lipolysis in the human digestive tract: Where do we stand? Current
Opinion in Clinical Nutrition and Metabolic Care, 10(2), 156-164.
Bernback, S., Blackberg, L. & Hernell, O. (1989). Fatty acids generated by gastric lipase promote human
milk triacylglycerol digestion by pancreatic colipase-dependent lipase. Biochimica et Biophysica
Acta - Lipids and Lipid Metabolism, 1001(3), 286-293.
Capolino, P., Guérin, C., Paume, J., Giallo, J., Ballester, J. M., Cavalier, J. F. & Carrière, F. (2011). In vitro
gastrointestinal lipolysis: Replacement of human digestive lipases by a combination of rabbit
gastric and porcine pancreatic extracts. Food Digestion, 2(1-3), 43-51.
Carriere, F., Barrowman, J. A., Verger, R. & Laugier, R. (1993). Secretion and contribution to lipolysis of
gastric and pancreatic lipases during a test meal in humans. Gastroenterology, 105(3), 876-888.
Carrière, F., Grandval, P., Renou, C., Palomba, A., Priéri, F., Giallo, J., Henniges, F., Sander-Struckmeier, S.
& Laugier, R. (2005). Quantitative study of digestive enzyme secretion and gastrointestinal
lipolysis in chronic pancreatitis. Clinical Gastroenterology and Hepatology, 3(1), 28-38.
Cummings, D. E. & Overduin, J. (2007). Gastrointestinal regulation of food intake. Journal of Clinical
Investigation, 117(1), 13-23.
Dressman, J. B., Berardi, R. R., Dermentzoglou, L. C., Russell, T. L., Schmaltz, S. P., Barnett, J. L. &
Jarvenpaa, K. M. (1990). Upper gastrointestinal (GI) pH in young, healthy men and women.
Pharmaceutical Research, 7(7), 756-761.
Elashoff, J. D., Reedy, T. J. & Meyer, J. H. (1982). Analysis of gastric emptying data. Gastroenterology,
83(6), 1306-1312.
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P.
F., Stellaard, F. & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption
and increases plasma CCK but has minor effects on gastric emptying and appetite. American
Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982-991.
Gargouri, Y., Pieroni, G., Rivière, C., Lowe, P. A., Saunière, J.-F., Sarda, L. & Verger, R. (1986b). Importance
of human gastric lipase for intestinal lipolysis: an in vitro study. Biochimica et Biophysica Acta
(BBA) - Lipids and Lipid Metabolism, 879(3), 419-423.
Golding, M., Wooster, T. J., Day, L., Xu, M., Lundin, L., Keogh, J. & Clifton, P. (2011). Impact of gastric
structuring on the lipolysis of emulsified lipids. Soft Matter, 7(7), 3513-3523.
Helbig, A., Silletti, E., Timmerman, E., Hamer, R. J. & Gruppen, H. (2012). In vitro study of intestinal
lipolysis using pH-stat and gas chromatography. Food Hydrocolloids, 28(1), 10-19.
Lal, S., McLaughlin, J., Barlow, J., D'Amato, M., Giacovelli, G., Varro, A., Dockray, G. J. & Thompson, D. G.
(2004). Cholecystokinin pathways modulate sensations induced by gastric distension in
humans. American Journal of Physiology - Gastrointestinal and Liver Physiology, 287(1 50-1).

In vitro lipid digestion of protein stabilized emulsions
69
Little, T. J., Russo, A., Meyer, J. H., Horowitz, M., Smyth, D. R., Bellon, M., Wishart, J. M., Jones, K. L. &
Feinle-Bisset, C. (2007). Free fatty acids have more potent effects on gastric emptying, gut
hormones, and appetite than triacylglycerides. Gastroenterology, 133(4), 1124-1131.
Malaki Nik, A., Wright, A. J. & Corredig, M. (2010). Surface adsorption alters the susceptibility of whey
proteins to pepsin-digestion. Journal of Colloid and Interface Science, 344(2), 372-381.
Malaki Nik, A., Wright, A. J. & Corredig, M. (2011). Impact of interfacial composition on emulsion
digestion and rate of lipid hydrolysis using different in vitro digestion models. Colloids and
Surfaces B: Biointerfaces, 83(2), 321-330.
Maljaars, P. W. J., Peters, H. P. F., Mela, D. J. & Masclee, A. A. M. (2008). Ileal brake: A sensible food target
for appetite control. A review. Physiology and Behavior, 95(3), 271-281.
Marciani, L., Gowland, P. A., Spiller, R. C., Manoj, P., Moore, R. J., Young, P. & Fillery-Travis, A. J. (2001).
Effect of meal viscosity and nutrients on satiety, intragastric dilution, and emptying assessed
by MRI. American Journal of Physiology - Gastrointestinal and Liver Physiology, 280(6), G1227-
1233.
Marciani, L., Wickham, M., Singh, G., Bush, D., Pick, B., Cox, E., Fillery-Travis, A., Faulks, R., Marsden, C.,
Gowland, P. A. & Spiller, R. C. (2007). Enhancement of intragastric acid stability of a fat
emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder
contraction. American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(6),
G1607-G1613.
Marciani, L., Faulks, R., Wickham, M. S. J., Bush, D., Pick, B., Wright, J., Cox, E. F., Fillery-Travis, A.,
Gowland, P. A. & Spiller, R. C. (2008). Effect of intragastric acid stability of fat emulsions on
gastric emptying, plasma lipid profile and postprandial satiety. British Journal of Nutrition,
Forthcoming(-1), 1-10.
Marteau, P., Flourie, B., Pochart, P., Chastang, C., Desjeux, J. F. & Rambaud, J. C. (1990). Effect of the
microbial lactase (EC 3.2.1.23) activity in yoghurt on the intestinal absorption of lactose: An in
vivo study in lactase-deficient humans. British Journal of Nutrition, 64(1), 71-79.
McLaughlin, J., Luca, M. G., Jones, M. N., D'Amato, M., Dockray, G. J. & Thompson, D. G. (1999). Fatty
acid chain length determines cholecystokinin secretion and effect on human gastric motility.
Gastroenterology, 116(1), 46-53.
Minekus, M. (1995). A multicompartmental dynamic computer-controlled model simulating the stomach
and small intestine. Alternatives to Laboratory Animals, 23(2), 197-209.
Moreau, H., Gargouri, Y., Lecat, D., Junien, J. L. & Verger, R. (1988). Screening of preduodenal lipases in
several mammals. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism, 959(3), 247-252.
Nik, A. M., Corredig, M. & Wright, A. J. (2010). Changes in WPI-stabilized emulsion interfacial properties
in relation to lipolysis and β-carotene transfer during exposure to simulated gastric-duodenal
fluids of variable composition. Food Digestion, 1(1-2), 14-27.
Pafumi, Y., Lairon, D., Lechene de la Porte, P., Juhel, C., Storch, J., Hamosh, M. & Armand, M. (2002).
Mechanisms of inhibition of triacylglycerol hydrolysis by human gastric lipase. The Journal of
biological chemistry, 277(31), 28070-28079.
Pignol, D., Ayvazian, L., Kerfelec, B., Timmins, P., Crenon, I., Hermoso, J., Fontecilla-Camps, J. C. &
Chapus, C. (2000). Critical role of micelles in pancreatic lipase activation revealed by small
angle neutron scattering. J. Biol. Chem., 275(6), 4220-4224.

Chapter 3
70
Reis, P. M., Raab, T. W., Chuat, J. Y., Leser, M. E., Miller, R., Watzke, H. J. & Holmberg, K. (2008). Influence
of surfactants on lipase fat digestion in a model gastro-intestinal system. Food Biophysics), 1-12.
Sarkar, A., Goh, K. K. T., Singh, R. P. & Singh, H. (2009). Behaviour of an oil-in-water emulsion stabilized
by [beta]-lactoglobulin in an in vitro gastric model. Food Hydrocolloids, 23(6), 1563-1569.
Schaafsma, G. (2005). The Protein Digestibility-Corrected Amino Acid Score (PDCAAS) - A concept for
describing protein quality in foods and food ingredients: A critical review. Journal of AOAC
International, 88(3), 988-994.
Silletti, E., Vingerhoeds, M. H., Norde, W. & van Aken, G. A. (2007). The role of electrostatics in saliva-
induced emulsion flocculation. Food Hydrocolloids, 21(4), 596-606.
Singh, H. & Sarkar, A. (2011). Behaviour of protein-stabilised emulsions under various physiological
conditions. Advances in Colloid and Interface Science, 165(1), 47-57.
van Aken, G. (2010). Relating Food Emulsion Structure and Composition to the Way It Is Processed in the
Gastrointestinal Tract and Physiological Responses: What Are the Opportunities? Food
Biophysics, 5(4), 258-283.
van Aken, G. A., Bomhof, E., Zoet, F. D., Verbeek, M. & Oosterveld, A. (2011). Differences in in vitro
gastric behaviour between homogenized milk and emulsions stabilised by Tween 80, whey
protein, or whey protein and caseinate. Food Hydrocolloids, 25(4), 781-788.
Van Tilbeurgh, H., Egloff, M. P., Martinez, C., Rugani, N., Verger, R. & Cambillau, C. (1993). Interfacial
activation of the lipase-procolipase complex by mixed micelles revealed by X-ray
crystallography. Nature, 362(6423), 814-820.
Villeneuve, P., Pina, M. & Graille, J. (1996). Determination of pregastric lipase specificity in young
ruminants. Chemistry and Physics of Lipids, 83(2), 161-168.
Vingerhoeds, M. H., Silletti, E., de Groot, J., Schipper, R. G. & van Aken, G. A. (2009). Relating the effect of
saliva-induced emulsion flocculation on rheological properties and retention on the tongue
surface with sensory perception. Food Hydrocolloids, 23(3), 773-785.

Chapter 4
Effect of oral administration and intragastric
distribution of lipids on lipolysis in gastric
and duodenal human fluids
To be submitted

Chapter 4
72
Abstract
The present study examined, how gastric and duodenal lipolysis is affected by the
intragastric colloidal state of the emulsions and how the administration route, i.e. oral or
intragastric, affects gastric lipolysis. In a randomized, single blind crossover study, 15
healthy subjects were intubated nasogastric on three separate occasions and of this a
subset of 6 healthy subjects was intubated nasoduodenal on two separate occasions.
Volunteers received a homogenized emulsion orally or by gastric infusion or a gastric
infusion of water followed by the oil phase (un-homogenized sample). The subset of 6
volunteers received a gastric infusion of a homogenized emulsion or of an un-
homogenized sample. Emulsion structure, pH and lipid composition was assessed in both
gastric and duodenal fluids. Results showed a gastric lipolysis of approx. 3% for all
treatments. Free fatty acid (FFA) concentrations in duodenal aspirates at 30 min were
higher after ingestion of the homogenized emulsion than after ingestion of the un-
homogenized samples (p<0.01). Gastric emptying was faster for the un-homogenized
sample than for the homogenized emulsion. Thus, overall, there was no effect observed of
oral processing and homogenization had no effect on gastric lipolysis. Nevertheless,
differences in inhibition of gastric emptying were observed: duodenal release of FFA and
inhibition of gastric emptying occurred earlier for the homogenized emulsion than for un-
homogenized samples.

Effect of intragastric distribution of fat on lipolysis
73
4.1 Introduction
The intake of fat and its passage through the gastrointestinal tract triggers
physiological responses thus impacting gut transit time (Heddle et al., 1988; Feinle et al.,
1996; Little et al., 2007; Maljaars et al., 2008). It is expected that such responses are able to
influence and contribute towards regulation of food intake and satiety, a goal to combat
obesity. Several studies (see for review Golding & Wooster, 2010) have investigated the in
vitro behaviour of fat present in the form of emulsions to predict their possible in vivo
digestion. Recently, we showed that by using a dynamic digestion model, gradual
acidification in combination with a custom designed gastric compartment imitating the
functions of the stomach, caused protein stabilized emulsions to cream (Helbig et al.,
2013). We have shown that this behaviour delayed intestinal lipolysis (Helbig et al., 2013).
Although this model represents a significant step towards better prediction of in vivo
digestion, there are still many factors unaccounted for.
In vivo digestion is very complex due to many simultaneously occurring processes,
such as pH lowering, salt secretions, enzymatic hydrolysis, change in the emulsion
structure and physiological feedback mechanisms like gastric, pylorus and intestinal
activation. Therefore, it is challenging to investigate how the physico-chemical properties
of foods affect the digestion and absorption of lipids in vivo and how this is related to
satiety and -related physiological responses, such as hormone secretion or gastric
emptying. There are increasing indications in literature, that free fatty acids (FFA) are
primarily responsible for triggering gastrointestinal effects of lipids when delivered to the
small intestine (Schwizer et al., 1997; Feinle et al., 2001; Feinle-Bisset et al., 2005; Little et
al., 2007). For example, Little et al. (Little et al., 2007) showed that FFA were more
effective to delay gastric emptying, stimulate hormone release and suppress appetite
compared to triglycerides. In this study, the FFA were directly administrated
intragastrically. In a normal meal, however, the concentration and delivery of FFA will
depend on the colloidal behaviour of the emulsion which changes progressively due to
variation in pH and due to enzyme action when reaching the stomach and the intestine.
Also, the release of FFA in the gastrointestinal tract will be affected by factors like the
continuous phase of the emulsion, type of emulsifier, fat composition and droplet size (see
for review McClements et al., 2009). For example, both in vitro (Armand et al., 1992; Borel
et al., 1994; Helbig et al., 2012) and in vivo (Armand et al., 1999) studies revealed that a
smaller droplet size leads to a higher rate of FFA release for both gastric and pancreatic

Chapter 4
74
lipase. However, the changes in droplets size in the stomach are not well understood as,
for example, the droplet size of a coarse emulsion, with a diameter of 10 µm, did not
change after ingestion while an increase in droplet size was observed with a fine
emulsion (Armand et al., 1999). Though, in this study a system with complex interfaces
was used as they are made out of different emulsifiers, such as lecithin and milk proteins.
As proteolysis and decrease in pH are both occurring in the stomach, the smaller droplets
are likely to aggregate and cream. Thus, it is difficult to interpret the changes in the
colloidal state of the emulsion, i.e. the intragastric distribution of lipids, and to pin point
which variables are inducing satiety and related physiological responses. Indeed, little
research has been conducted on the influence of the gastric lipolysis as affected by the
intragastric lipid distribution in relation to satiety perception. Although it has been
reported that emulsions that are intragastric stable reduce hunger and appetite (Marciani
et al., 2009), the effect of gastric hydrolysis and the subsequent release of FFA in relation
to physiological human responses is not clear.
In addition to the effect of gastric hydrolysis, a number of studies point at the role of
oral stimulation. The role of orally administrated fat and its cephalic responses is thought
to be of importance in regulating food intake as it has been shown that vagal stimulation
upon modified sham feeding accentuated effects on metabolites, hormones and satiety
(Robertson et al., 2002; Heath et al., 2004; Smeets & Westerterp-Plantenga, 2006). Thus,
investigating the role of oral stimulation might reveal the contribution of the mouth on
gastric lipolysis, as measured by the release of FFA, and its influence on gastric emptying.
The role of oral stimulation and its contribution to hormone release and satiety
perceptions will be discussed in Chapter 5.
The focus of this chapter is to describe how gastric and duodenal lipolysis are affected
by the intragastric colloidal state of the emulsions. Furthermore, the effect of the
administration route, i.e. oral or intragastric, on gastric lipolysis was explored.
4.2 Materials and methods
4.2.1 Emulsion preparation
Emulsions were produced following good manufacturing practice in a food grade pilot
plant of NIZO food research (Ede, The Netherlands). Tween 80 (Lamesorb©SMO20,
Cognis, Monheim am Rhein, Germany) solutions were prepared by dissolving powder in

Effect of intragastric distribution of fat on lipolysis
75
0.01 M NaCl solution. The final emulsions contained 8% (w/w) sunflower oil (Reddy
sunflower oil, Vandermoortele, Izegem, Belgium) and 0.4% (w/w) Tween 80. All
ingredients used were food grade. In total 3 batches of each solution were prepared. Each
batch consisted of 10 L (20 x 500 mL) of each solution. Emulsions were homogenized at
80°C at a pressure of 70 to 180 bar followed by homogenization at 0 to 20 bar (Rannie
Lab12-16.5, Leuze, Waardenburg, The Netherlands). The solutions for the un-
homogenized samples, i.e. Tween 80-solution and sunflower oil, were heated separately at
80°C. These samples were not homogenized and applied separately but for the ease of the
reading we refer to them as emulsions. Homogenized and un-homogenized emulsions
were then sterilized in a Combitherm sterilizer (Minister 1987, AlfaLaval, Lund, Sweden)
for 7 seconds at 141°C. After cooling to room temperature, solutions were filled
aseptically in sterilized plastic containers and stored <7°C in a dark place until the start of
the experiments in order to prevent off-flavor formation. Of every batch 2 samples were
tested for sterility. During the course of the experiment fresh emulsions were prepared
every 6-10 weeks in order to prevent excessive off-flavor formation or formation of
physical instabilities. The surface-weighted-mean- diameter (d32) and the volume-
weighted-mean diameter (d43) of the freshly prepared homogenized emulsions were 2.33
and 8.75 µm, respectively, as determined by dynamic light scattering (Mastersizer Hydro
2000S, Malvern Instruments, Malvern, UK) at room temperature.
During the production of the emulsions, 0.034% (w/w) and 0.012% (w/w) of vanilla
powder and saccharine, respectively, were added to the Tween 80 solutions. As a result,
the emulsions tasted pleasantly sweet with a sweet vanilla flavor having a somewhat
bitter off-note. On the days of intervention, 1 g of paracetamol (Apotheek van Thoor,
Maastricht, the Netherlands) was manually added to the homogenized emulsions or, in
the case of the un-homogenized emulsions, to the Tween 80 solution to measure gastric
emptying of the water phase.
4.2.2 Subjects
15 healthy volunteers participated in the study (11 male and 4 female, age 28±8 y, body
mass index of 22.7±2.3 kg/m2). A short physical examination and a cognitive restraint
eating behaviour using a Dutch translation of the Three Factor Eating Questionnaire
(TFEQ) were performed. Participants did not have gastrointestinal or hepatic disorders or
previous major abdominal surgery interfering with gastrointestinal function. They were
non-smokers, moderate alcohol consumers (≤ 10 alcoholic consumptions per week), non-

Chapter 4
76
medicated except of using contraceptives and non-restraint eaters according to the Three
Factor Eating Questionnaire (TFEQ Factor 1 score ≤ 9). The study was approved by the
Medical Ethics Committee of the Maastricht University Medical Center, conducted
according to the principles of the Declaration of Helsinki and in accordance with the
Medical Research Involving Human Subjects Act (WMO). All participants signed
informed consent. Volunteers were informed that they receive a lipid-containing drink
and that its behaviour in the stomach and duodenum as well as the resulting
physiological responses were investigated.
4.2.3 Study design
The study was a randomized crossover design and divided in two experiments. All
experiments took place on separate days. Subjects were requested to refrain from any
food or beverage intake with the exception of water from 2200h the evening before the
test day. Subjects received a total load of 500 mL of samples with a total energy content of
360 kcal.
Experiment 1: Gastric intubation study
On all 3 test days of the gastric intubation study, subjects arrived in the morning at
0830h in fasted state. After local anesthesia of the nasal mucosa using lidocaine 10%
spray, each subject was intubated with a double-lumen nasogastric feeding catheter
(Bengmark® Nutricia, Amsterdam, The Netherlands). One lumen was used to inject
emulsions into the stomach and the other lumen was used to collect stomach fluid by
aspiration using a standard 10-mL syringe. The position of the nasogastric catheter was
controlled by determination of the pH of the aspirate. After intubation, a cannula was
placed in a superficial antecubital vein of the forearm for blood sampling. At 0900h,
subjects received the emulsions infused into the stomach either homogenized (hs) and in
case of the un-homogenized emulsions (uhs), the infusion of the water phase was
followed by the oil phase within 2 min. In the case of the oral administration, the
emulsions were homogenized (ho) and drunk within 2 min.

Effect of intragastric distribution of fat on lipolysis
77
Experiment 2: Duodenal intubation study
To investigate the intestinal behaviour of the emulsions, 6 volunteers, who participated
in the gastric intubation study, joined on two further occasions following the same
procedure as described above with minor modification. On the study days, subjects were
intubated with a double-lumen naso-duodenal catheter (Bengmark® Nutricia,
Amsterdam, The Netherlands) after anaesthesia of the nasal mucosa. The intraduodenal
part of the catheter was placed under radiological control and placed with the tube tip
located of about 10 cm in the proximal small intestine directly distal to the stomach. The
position of the sample port in the stomach was controlled by aspiration. The emulsions
were identical in their composition to the ones used in the gastric intubation study and
applied to the stomach either un-homogenized (uhs) or homogenized (hs).
4.2.4 Measurements
Physicochemical characterisation of in vitro gastric and duodenal digestion of
emulsions
Samples (10-14mL) of stomach contents were withdrawn within 2 min at baseline (t=-
10 min) and at 10, 15, 30, 45, 60, 90, 120 and 180 min after application of the emulsions.
Samples of the duodenal contents were collected continuously by aspiration over 15 min
at baseline (t=-10 min) and at 15, 30, 45, 60, 90, 120 and 180 min after application of the
emulsions. The pH of these gastric or duodenal aspirates was directly measured using pH-
indicator strips (Merck, Darmstadt, Germany). On the day of administration, the
behaviour of the emulsions and of the emulsions in the aspirates was monitored visually
and recorded on camera. Microscopic images were obtained by putting one drop of the
samples on a hemocytometer (Bürker-Türk, Hofheim, Germany), covered and
immediately analysed using a Bresser® Trino Researcher microscope (40-1000x, Meade
instruments, Rhede, Germany) equipped with a moticam 2300 camera (MC2300, 3.0Mp,
USB2.0, Motic Instruments, Richmond, BC, Canada).
Samples were analyzed for FFA, mono-, di- and triglycerides (MG, DG, TG),
respectively, composition by gas chromatography (GC) as previously described (Helbig et
al., 2012) with minor modifications. Briefly, 2x0.5 mL samples were taken from the fresh
aspirates, immediately poured into extraction fluid and stored at -40 ºC until further
analysis. After extraction, the final supernatant was collected and its volume was brought

Chapter 4
78
to 8 mL with diethylether/heptane (1:1, w/w). An internal standard mix (50 µL), consisting
of C15:0 and TG36, was added to obtain a final concentration of 0.4 mg/mL. Prior to
analysis, 200 µL of each sample was diluted 1:2 with diethylether/heptane (1:1, w/w),
derivatized with 150 µL MSTFA, gently mixed and left at least 3h for reaction. Variations
between duplicates from 5 test persons and different application forms, i.e. 3 times uhs, 4
times ho and 3 times hs, were <3 %. Therefore, all further measurements with GC were
carried out once. Fatty acid standards, i.e. C14:0, C15:0, C16:0, C18:1, C18:2 as well as
monoolein, diolein, and triglyceride standards triolein and trilaurin (TG36) were obtained
from Sigma–Aldrich (>99 % pure, Sigma–Aldrich, St.Louis, MO, USA) as well as MSTFA
(N-methyl-N-(trimethylsilyl)trifluoroacetamide, lot BCBD5976V). All other chemicals
were of analytical grade and purchased from Merck or Sigma–Aldrich. Solutions were
prepared in demineralized water.
Levels of hydrolysis (L1% and L2%, respectively) were calculated according to Capolino
et al (Capolino et al., 2011) and as shown in Chapter 3.
Pancreatic lipase activity was measured in 0.5 mL aliquots of duodenal samples.
Samples were stabilized by adding 100 µL of protease inhibitors (Protease Inhibitor
Cocktail, Sigma) after which they were immediately frozen in liquid nitrogen and stored
at -80 ºC until further analysis. Lipase activity was analysed using a fluorometric assay
(Cortner et al., 1976). Quantification was carried out against a standard curve of 4-
methylumbelliferone and 1 nmol hydrolysed substrate per minute is defined as 1 unit.
Blood samples
During the test days, blood samples were taken at baseline (t=-10 min) and at 10, 15,
30, 45, 60, 90, 120 and 180 min after application of the emulsions. After sampling,
physiological salt solution (NaCl 0.9 % w/v) and heparine were injected to prevent
coagulation and occlusion of the catheter. Blood samples for paracetamol analysis were
drawn in EDTA tubes. All samples were kept on ice and all tubes were centrifuged at 1500
× g for 10 min at 4°C within one hour. Aliquots of the supernatant plasma samples were
stored at -80°C until further analysis. Paracetamol was analysed by reversed phase HPLC
with UV detection using an in-house method at the department of Toxicology, Maastricht
University. After taking the last sample at t=180 min, subjects were immediately
extubated and received 30 min thereafter a lunch.

Effect of intragastric distribution of fat on lipolysis
79
4.2.5 Data analysis
Data transformation
Data analyses were performed on the measured pH of gastric (GA) and duodenal (DA)
aspirates, the concentrations of free fatty acids (FFA) and mono- (MG), di- (DG) and
triglycerides (TG) in gastric and duodenal aspirates, lipase activity, and blood
concentrations of paracetamol. First, the normality of dependent variable distributions
was assessed by the Kolmogorov-Smirnov test. These tests revealed that the normality
assumption was violated for most dependent variables. Therefore, data transformations
were performed prior to ANOVA testing. The nature of data transformation depended on
skew and range constraints of dependent variables: y’= 10
log(y) transformations were used
for right-skewed distributions with lower-bound delimiters (i.e. lipid concentrations,
lipase activity, plasma paracetamol). Dependent variable distributions that were delimited
on two sides were transformed by y’= ln(y/(max-y)), with max being the maximum
allowable value (pH).
Dose-response functions
In addition, pharmaco-kinetic dose-response curves were fit to the timed observations
of the dependent variables used. For this, the mathematical Weibull model was used,
which describes physiological responses to the dissolution of drugs (Thelen et al., 2012)
by the following equation:
y=c∗(x /a)(b�1)∗exp(�(x /a)b)
where the dose-response function (y) is described as a function of time (x) and a time
scaling factor (a), a shape factor (b) and an amplitude scaling factor (c). The amplitude
scaling factor includes negative values for downward going pulse responses. Weibull
functions were fit to individual dose-response observations by minimizing the summed
squared distances of observed data points to the function using the MS Excel solver
function with the GRG non-linear fit engine and the restraints 1.05 < a < 6 and 0.5 < b <
10. This procedure was repetitively performed for all dependent variables.
Response curves of dependent measures over time were compared to ascertain the
temporal sequence of responses of different modalities (i.e. pH, paracetamol uptake,

Chapter 4
80
lipolysis, lipid synthesis) in relation to different treatments. For this purpose, peak onset
times (t-onset) and peak optimum times (t-peak) were estimated from the modelled
response curves. Individual t-onsets were defined as the time at which the response
estimates deviated 20 % from the baseline towards the optimum value. The t-peak was
defined as the time point at which Weibull functions showed the maximum value for
upward going response curves or the minimum value for downward going response
curves. Both t-onset and t-peak were estimated from the Weibull model using the excel
solver function with the restraints t-peak > 0 and 0 < t-onset < t-peak.
Statistical testing
Gastric intubation study Effects by the factor Time (fixed factor, “t=-10”, “t=10”, “t=15”,
“t=30”, “t=45”, “t=60”, “t=90”, “t=120”, “t=180”), Treatment (fixed factor, “ho”, “hs”, “uhs”)
and Subjects as a random factor were tested by multifactor ANOVA. This analysis also
included all two-factor interactions. In addition, multifactor ANOVA was used to test
effects on t-peak and t-onset by the factor Measures (fixed factor, “pH-GA”, “plasma
paracetamol”, “FFA-GA”, “MG-GA”, “DG-GA”, “TG-GA”), Treatment (fixed factor, “ho”,
“hs”, “uhs”), and Subject (random factor) and all two-factor interactions of these.
Duodenal intubation study Effects on the selected measures, i.e. pH, FFA, MG; DG, TG,
lipase activity, by Time (fixed factor, “t=-10”, “t=15”, “t=30”, “t=45”, “t=60”, “t=90”, “t=120”,
“t=180”), Treatment (fixed factor, “hs”, “uhs”), and Subjects (random-factor) and all two-
factor interactions were tested by multifactor ANOVA. In addition, effects on t-peak and
t-onset by the factors Measure (fixed factor, “pH-DA”, “lipase activity”, FFA-DA”, “MG-
DA”, “DG-DA”, “TG-DA), Treatment (fixed factor, “hs”, “uhs”), and Subject (random
factor) and their two-factor interactions were tested by multifactor ANOVA. In case of
significant ANOVA test results, pairwise post-hoc comparisons were performed with
Tukey HSD correction for multiple comparisons.
All data were analysed using Statistica (version 10, StatSoft,Tulsa, OK, USA). All
results are expressed as means ± standard error (SE). The significance level was 0.05.
4.3 Results
The study procedure was well tolerated by the subjects. Only two subjects were
sensitive to the tubing and felt nauseas on two occasions during the gastric intubation

Effect of intragastric distribution of fat on lipolysis
81
study. Therefore, we stopped those investigations after 90 and 120 min, respectively.
Nevertheless, none of the 15 enrolled subjects withdrew from the study.
4.3.1 Gastric intubation study
Appearance of the emulsions
In the stomach, the homogenized Tween 80-stabilized emulsions appeared visually
homogenous on a macroscopic level throughout the experiment with a white cream
upper layer on top and a turbid lower phase. There was no difference in the appearance
of the homogenized emulsions between the oral or gastric application. In contrast, the un-
homogenized emulsions were clearly watery, but became more turbid or had an oil upper
layer after 90 min. Droplets of the homogenized emulsions tended to flocculate and
coalesce (Fig. 4.1A, B). As expected, un-homogenized emulsions revealed either no or very
large droplets (data not shown).
Table 4.1 summarizes ANOVA results of the all independent variables, i.e. pH, plasma
paracetamol concentration, lipids (such as FFA, MG, DG and TG) in gastric or duodenal
aspirates as well as lipase activity.
pH profile and paracetamol plasma levels
The gastric pH profiles are shown in Fig. 4.2A and the initial pH was 3.3 ± 0.3. After
ingestion of the emulsions the pH was 3.1 ± 0.2 at t=10 min and as revealed post hoc
decreased significantly to 2.1 ± 0.1 at t=30 min. After this interval the gastric pH remained
constant until the end of the experiment. There was no difference between the treatments
ho, hs or uhs as well as no Treatment × Time interaction (Table 4.1).

Chapter 4
82
Fig. 4.1 Light microscopic appearance of the homogenized Tween 80-stabilized emulsion (a) and of
gastric samples after the treatment hs (A) and ho (B), Experiment 1 at 10, 30, 120 and 180 min and of
duodenal samples after the treatment hs (C), Experiment 2 at 15, 30, 120 and 180 min; Magnification is
100x.

Effect of intragastric distribution of fat on lipolysis
83
Table 4.1 Effect of Time of aspirate or plasma collection, emulsions used as Treatment, and Time ×
Treatment interaction on pH, FFA, MG, DG, TG in gastric or duodenal aspirates and indicated with GA or
DA, respectively, on lipase activity or plasma paracetamol concentration.
Time Treatment Time x Treatment dependent variable
F (df1,df2) Sig F (df1,df2) Sig F (df1,df2) Sig
pH-GA 11.61 (8,224) *** 1.11 (2,224) Ns 1.19 (16,224) Ns plasma paracetamol 69.18 (8,224) *** 9.80 (2,224) *** 9.72 (16,224) *** FFA-GA 4.71 (8,224) *** 0.08 (2,224) Ns 0.72 (16,224) Ns MG-GA 1.39 (8,224) Ns 0.58 (2,224) Ns 0.83 (16,224) Ns DG-GA 18.43 (8,224) *** 4.65 (2,224) * 1.54 (16,224) Ns TG-GA 35.55 (8,224) *** 14.26 (2,224) *** 3.03 (16,224) *** FFA-DA 10.97 (7,35) *** 12.63 (1,35) * 2.02 (7,35) Ns MG-DA 6.39 (7,35) *** 1.33 (1,35) Ns 2.42 (7,35) * DG-DA 2.06 (7,35) Ns 10.93 (1,35) ** 3.72 (7,35) ** TG-DA 3.29 (7,35) ** 2.98 (1,35) Ns 2.46 (7,35) * lipase activity 2.58 (7,35) * 0.34 (1,35) Ns 2.82 (7,35) * pH-DA 0.64 (7,35) Ns 0.05 (1,35) Ns 1.42 (7,35) Ns
Significant differences are indicated with asterisks: *p<0.05, **p< 0.01, ***p < 0.001, based on ANOVA analysis. Ns is not significant.
Fig. 4.2 Gastric pH-profile (A) and plasma paracetamol concentrations (B) during 180 min of Experiment
1 after the treatments ho (●), hs (□) and uhs (▲).Values are expressed as mean ± SE. Different low letters
indicate a significant difference at p<0.001.

Chapter 4
84
Based on ANOVA, plasma paracetamol concentration showed an effect of Treatment,
Time and Treatment × Time interaction (Table 4.1). Post hoc analysis further revealed
that the concentration increased significantly after application of all emulsions and did
not reach baseline values at the end of the experiment. There was no difference between
ho and hs. For the treatments ho and hs plasma concentration curves were significantly
different from uhs (Fig. 4.2B) indicating a faster gastric emptying of the water phase for
the treatment uhs compared to hs and ho. Also, a specific concentration pattern was
found which is depending on the type of treatment and time of sampling (supplementary,
Table S4.1). In the case of plasma paracetamol, concentrations were highest between t=10
and 45 min for treatment uhs compared to hs and ho.
Lipid composition
The lipid compositions of gastric aspirates for the different treatments are presented in
Figure 4.3. ANOVA (Table 4.1) revealed a significant Time effect on the release of FFA,
but no Treatment effect or Treatment × Time interaction. In case of the main effect Time,
post hoc testing showed that the concentration of FFA increased significantly within the
first 10 min to 1.8±0.2, 2.5±0.4 and 2.6±1.2 mM for the treatments ho, hs and uhs,
respectively and stayed constant during the rest of the experiment. Furthermore, when
considering the other lipolytic products, only traces of MG production were found with
no significant difference between treatments and time of collection. Also Treatment ×
Time interaction was not significant. When considering the formation of DG, we found
significant effects of Treatment and Time, but no Treatment × Time interaction (p=0.09).
Post hoc, the concentration of DG for uhs was significantly lower compared to ho and hs,
but for all treatments the concentration increased significantly after application compared
to baseline. There was a Treatment and Time effect and a Treatment × Time interaction
on the concentration of triglycerides (TG). Post hoc analysis revealed that for all
treatments the concentration of TG was significant higher at all time points compared to
baseline and decreased with the increase of time. Furthermore, when evaluating the
Treatment × Time interaction uhs had, for example, similar TG concentration compared
to ho at 30 min but not at 45 min, where in fact the TG were significantly lower
(supplementary, Table S4.2). The results of the lipolytic products indicate similar behavior
observed between ho and hs.

Effect of intragastric distribution of fat on lipolysis
85
Fig. 4.3 Lipid composition, i.e. TG, triglyceride; DG, diglyceride; MG, monoglyceride; FFA, free fatty acid,
of gastric aspirates as determined by gas chromatography during 180 min of Experiment 1after the
treatments ho (A), hs (B) and uhs (C). Values are expressed as mean and error bars indicate the respective
standard errors for TG.

Chapter 4
86
The level of lipolysis were calculated by using data in Figure 4.3. The level of gastric
lipolysis (L1%) did not exceed 2.8 % for all treatments and time points (data not shown).
Also, the molar ratios of FFA/DG were found to be about 1, which is typical for the
hydrolysis by human gastric lipase.
4.3.2 Duodenal intubation study
Appearance of the emulsion
For two persons it was not possible to draw samples between t=15 and t=60 min for
the treatment hs. All other samples obtained from the duodenum were transparent before
ingestion and became turbid after ingestion of the homogenized emulsions (no further
data shown). During digestion of the un-homogenized emulsions, the samples stayed
clear. Homogenized emulsions coalesced immediately at t=15min (Fig. 4.1C). For uhs no
droplets were found (no further data shown).
The initial pH in the duodenum was 6.4 ± 0.3 (Fig. 4.5B) and ANOVA revealed no
Time, Treatment effect or Treatment × Time interaction on duodenal pH (Table 4.1).
Similarly to the gastric intubation study, the water phase emptied faster from the stomach
for uhs compared to hs as determined from plasma paracetamol concentrations (no
further data shown).
Lipid composition
Figure 4.4 shows the lipid composition determined in duodenal aspirates. ANOVA
revealed main effects of Treatment and Time on the concentration of FFA, but no
Treatment × Time interaction (p=0.08) (Table 4.1). Thus, the concentrations of FFA were
higher for hs compared to uhs. Furthermore, there was a Time effect and a Treatment ×
Time interaction on MG, but no Treatment effect. There was a significant Treatment
effect and a Treatment × Time interaction on the concentration of DG but no Time effect
(p=0.07). There was an effect of Time and a Treatment × Time interaction on TG, but no
Treatment effect (p=0.15). Comparing hs with uhs, post hoc analysis revealed higher DG
concentration in the duodenal aspirates at 15 min for hs than for uhs (Supplementary,
Table S4.3). In particular, for hs there were significant concentrations of FFA at 30, 60 and
120 min, of MG at 45 and 60 min, of DG at 15 min and of TG at 120 min present compared
to baseline. The higher TG concentration found at t=120 min compared to baseline

Effect of intragastric distribution of fat on lipolysis
87
suggests the entry of a cream layer formed in the stomach. For uhs, there were significant
amounts of FFA and MG at 60 min present in the duodenal aspirates compared to baseline
but there were no significant amounts of DG or TG for all time points.
Fig. 4.4 Lipid composition, i.e. TG, triglyceride; DG, diglyceride; MG, monoglyceride; FFA, free fatty acid,
of duodenal aspirates as determined by gas chromatography during 180 min of Experiment 2 after the
treatments hs (A) and uhs (B). Values are expressed as mean and error bars indicate the respective
standard errors for TG.
The lipase activity is shown in Fig. 4.5A. There was a Time effect and Treatment ×
Time interaction on lipase activity (Table 4.1). Post hoc analysis revealed no significant
increase of lipase activity for both treatments compared to baseline. For uhs, there was a

Chapter 4
88
significant increase at t=60 and t=180 min compared to 15 min indicating a later response
to the treatment uhs compared to hs.
Fig. 4.5 Lipase activity obtained in duodenal aspirates (A) and pH-profile (B) during 180 min of
Experiment 2 after the treatments hs (□) and uhs (▲).Values are expressed as mean ± SE.
4.3.3 T-peak and t-onset
To ascertain if we could observe difference in timed responses of measures in relation
to the treatments, peak optimum times (t-peak) and peak onset times (t-onset) were
extracted from fitted curves. A typical example for such a Weibull fit (and the indications
for t-peak and t-onset) is shown in Fig. 4.6. ANOVA results are shown in Table 4.2.

Effect of intragastric distribution of fat on lipolysis
89
Fig. 4.6: Example of a typical Weibull fit of released free fatty acids (FFA) during 180 min for an
individual subject, with peak onset time (▲), peak time (×) and the line which indicates the model fit.
For example, there was a significant Measure effect and a Measure× Treatment
interaction on t-peak and a significant Measure effect on t-onset. Post hoc analysis
revealed that in the gastric intubation study t-peaks were early for FFA, DG, TG and pH
at approx. 50 min and this was not different between the treatments (Fig. 4.7A).
Compared to this, t-peak for MG was significantly later at approx. 170 min. Only for
plasma paracetamol concentration t-peak for uhs was earlier than for ho and hs. There
was no difference for t-onset. In the duodenal intubation study, t-peaks for lipids were
earlier for uhs than for hs (Fig. 4.7B).
Table 4.2 Effect of Measure on t-peak and t-onset, emulsions used as Treatment, and Measure × Treatment interaction on t-peak and t-onset.
Measure Treatment Measure x
Treatment
dependent variables
F (df1,df2) Sig F (df1,df2) Sig F (df1,df2) Sig
t-peak§ gastric 7.35 (5,140) *** 3.13 (2,140) Ns 2.14 (10,140) *** t-onset§ gastric 1.52 (5,140) Ns 1.22 (2,140) Ns 0.83 (10,140) Ns t-peak# duodenal 1.45 (5,25) Ns 1.25 (1,25) Ns 1.06 (5,25) Ns t-onset# duodenal 1.32 (5,25) Ns 1.14 (1,25) Ns 0.66 (5,25) Ns
Significant differences are indicated with asterisks: *p<0.05, **p< 0.01, ***p < 0.001, based on ANOVA analysis. Ns is not significant. § includes measures pH-GA, FFA-GA, MG-GA, DG-GA, TG-GA and plasma paracetamol concentration; GA, gastric aspirate # includes measures pH-DA, FFA-DA, MG-DA, DG-DA, TG-DA and lipase activity; DA, duodenal aspirate

Chapter 4
90
Fig. 4.7 Comparison of peak times and onset times of timed responses (A, gastric intubation study) for
gastric pH, plasma paracetamol concentration, lipids in gastric aspirates (GA), i.e. TG, triglyceride; DG,
diglyceride; MG, monoglyceride; FFA, free fatty acid; for the treatments ho, hs and uhs and (B, duodenal
intubation study) for lipase activity and lipids in duodenal aspirates (DA), i.e. TG, DG, MG, FFA for the
treatments hs and uhs. Values are expressed as mean ± SE from 15 (A) or 6 (B) people, respectively.
Different low letters indicate a significant difference at p<0.001.

Effect of intragastric distribution of fat on lipolysis
91
4.4 Discussion
The small intestine is the most important site for fat digestion and absorption. Thus,
fat-related physiological responses are often investigated by local perfusion of fat in the
small intestine (Little et al., 2007). However, such mechanistic studies neglect the effects
of fat in the oral cavity and digestion in the gastric compartment on overall fat digestion
and fat-related physiological responses. Therefore, in the present study, the oral
administration of an emulsion load was included and compared to the intragastric
infusion of the same emulsion.
4.4.1 Effect of oral processing on emulsion behavior
When comparing the oral processing with the intragastric infusion of the
homogenized emulsions, the results showed that oral exposure had no additional effect on
the gastric lipolysis, the structure of the emulsions or the pH-profile. In addition, we did
not observe differences in the rate of emptying of the homogenized emulsions between
the oral and the intragastric administration as determined by plasma paracetamol
concentrations. To our knowledge this has not been reported before. The effect of oral
exposure in relation to hormonal responses and satiety feelings will be discussed in
Chapter 5.
4.4.2 Effect of intragastric distribution on gastric and duodenal
lipolysis
To investigate the effect of intragastric distribution of lipids, i.e. present as fine
emulsion droplets or as oil layer in the stomach, on lipolysis a homogenized (hs) and an
un-homogenized emulsion (uhs) were used, respectively, both having the same
composition and amount of calories. Although gastric lipolysis was similar for both
treatments and gastric lipolysis was not influenced by the distribution of the lipids there
were overall significant higher concentrations of FFA and for DG within the first 15 min
present in duodenal aspirates for hs compared to uhs. It is known that gastric lipolysis
and the subsequent release of FFA enhance pancreatic lipase activity (Gargouri et al.,
1986; Borel et al., 1994) and the formed DG will be hydrolysed faster than TG (Phan &
Tso, 2001). This means that the low concentration of FFA released from the stomach for
hs was sufficient to increase pancreatic lipase activity and, therefore, led to a fast

Chapter 4
92
duodenal hydrolysis with subsequent FFA release. Probably, this fast hydrolysis
contributed to a delay in gastric emptying for hs as indicated by the plasma paracetamol
concentration (Fig. 4.2B). In contrast, plasma paracetamol concentrations indicated a fast
gastric emptying of the water phase from uhs within 45 min (Fig. 4.2B, Fig. 4.7A). Thus,
the lower concentrations of FFA in uhs compared to hs were not sufficient to reduce
gastric emptying within the first 45 minutes. However, for uhs we observed highest FFA
concentrations in duodenal aspirates at t=60 min (Fig. 4.4B) and the modeled peak time
was at approx. 85 min (Fig. 4.7B). This suggests the start of emptying of the lipid layer
within this time frame for uhs and resulting lipolysis, despite the fact, that the surface
area of the lipid layer is small.
4.4.3 Effect of creaming or phase separation on gastric lipid
emptying
The emulsions infused in the case of hs were Tween-stabilized emulsions. They are
considered to be stable emulsions in the acidic gastric environment (Marciani et al., 2007;
Marciani et al., 2008) which do not cream and remain its initial droplet size. Surprisingly,
we found significantly high concentrations of TG in the duodenal aspirates at t=120 min
(Fig. 4.4A) whereas for all other time points the lipolytic products FFA, MG and DG were
dominant. This observation could be explained by a cream layer formed in the stomach,
which is composed of a higher TG concentration and will enter the duodenum at this time
range. Recently we have observed an increase in TG concentration after the digestion of a
Tween 80-stabilized emulsion at t=150 min in a dynamic digestion model (Oosterveld et
al., unpublished data). In this model Tween 80-stabilized emulsions were found to cream
in the stomach compartment. Similar to our study, slight coalescence has been observed
for Tween 80-stabilized emulsions under simulated gastric conditions (van Aken et al.,
2011). Although a high concentration of substrate is present at t=120 min, the action of
lipase is not increasing and the ratio of TG/FFA is 1.1 at t=120 min. It has been described
that pancreatic lipase secretion decreases despite on-going nutrient delivery to the
duodenum within 180 min after meal intake (Fried et al., 1988; Keller et al., 1997; Schwizer
et al., 1997). Since lipase action will also be reduced by a smaller surface area of the
droplets in the cream phase, these factors together lead to less formation of FFA. As no
further lipolytic products might be formed, un-hydrolysed TG present in the cream layer
will enter the duodenum. Such an effect might also be seen for the treatment uhs after the
180 min of experiment, since we did not find high TG concentration in duodenal aspirates

Effect of intragastric distribution of fat on lipolysis
93
following the rapid emptying of the water phase. It is likely that most of the oil phase of
the un-homogenized emulsion is still present in the stomach and thus will enter the
duodenum at a later stage. Both a cream and a remaining oil phase might affect responses
to the intake of a second meal. For instance, it has been observed that lipids ingested at
breakfast contributed to an increase in TG plasma concentration after intake of a lunch
(Maillot et al., 2008). Additionally, CCK plasma concentration was higher after the preload
of a water-oil-layered meal compared to an emulsified meal followed by intake of a
second meal (Foltz et al., 2009). However, it is not clear yet if this effect derives from
stored lipids in the enterocytes or from that present in the gut. Our study supports the
possibility that the second meal stimulates the remaining oil layer to empty from the
stomach (Foltz et al., 2009).
4.5 Conclusions
The study showed no effect of oral processing but a clear effect of the degree of
emulsification on emulsions behavior. In detail, we observed initial high intraduodenal
lipid concentrations for the homogenized emulsion. This in turn resulted in faster release
of FFA and delayed gastric emptying compared to the un-homogenized emulsion.
Furthermore, none of the treatments had a pronounced effect on gastric lipolysis. Despite
a low gastric lipolysis and the fact, that in case of the un-homogenized emulsion only a
small surface area was available, hydrolysis was high when TG emptied into the
duodenum.

Chapter 4
94
4.6 Supplementary data
Table S4.1 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on plasma paracetamol
concentration (µg/mL) as determined from ANOVA. Different letters indicate a significant difference at
p<0.05.
Treatment Time
ho hs uhs -10 0.10 ± 0.07a 0.09 ± 0.09a 0.06 ± 0.06a
10 1.61 ± 0.28b 2.53 ± 0.52bcd 6.82 ± 1.13fg
15 1.74 ± 0.21bc 2.94 ± 0.61bcd 8.52 ± 1.27g
30 3.19 ± 0.66bcd 3.46 ± 0.56cde 7.08 ± 0.83fg
45 3.81 ± 0.74de 3.62 ± 0.66cde 6.30 ± 0.73fg
60 4.38 ± 0.75def 3.67 ± 0.63de 5.31 ± 0.57efg
90 4.74 ± 0.68def 4.27 ± 0.50def 4.40 ± 0.49def
120 5.47 ± 0.56efg 4.18 ± 0.44def 4.47 ± 0.40defg
180 5.23 ± 0.42fg 4.21 ± 0.35def 3.72 ± 0.20def
Table S4.2 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on TG concentration in
gastric aspirates (mg/mL) as determined from ANOVA. Different letters indicate a significant difference
at p<0.05.
Treatment Time
ho hs uhs
-10 1.31 ± 0.22a 1.16 ± 0.22 a 1.33 ± 0.19a
10 45.38 ± 8.25efghi 64.11 ± 6.46i 32.66 ± 16.74bc
15 63.24 ± 13.88ghi 62.73 ± 6.95i 43.72 ± 20.66bc
30 42.49 ± 11.43defghi 53.05 ± 7.04hi 52.29 ± 24.62bcde
45 55.87 ± 10.95fghi 53.64 ± 7.55ghi 28.85 ± 9.72bc
60 54.50 ± 14.22fghi 57.02 ± 9.90ghi 40.65 ± 18.26bcdef
90 56.31 ± 10.79ghi 44.82 ± 8.84fghi 34.68 ± 12.81bc
120 54.67 ± 15.26efghi 25.51 ± 5.29cdefgh 16.96 ± 7.02bc
180 43.15 ± 14.76cdefg 19.79 ± 6.48bcd 8.67 ± 3.35ab
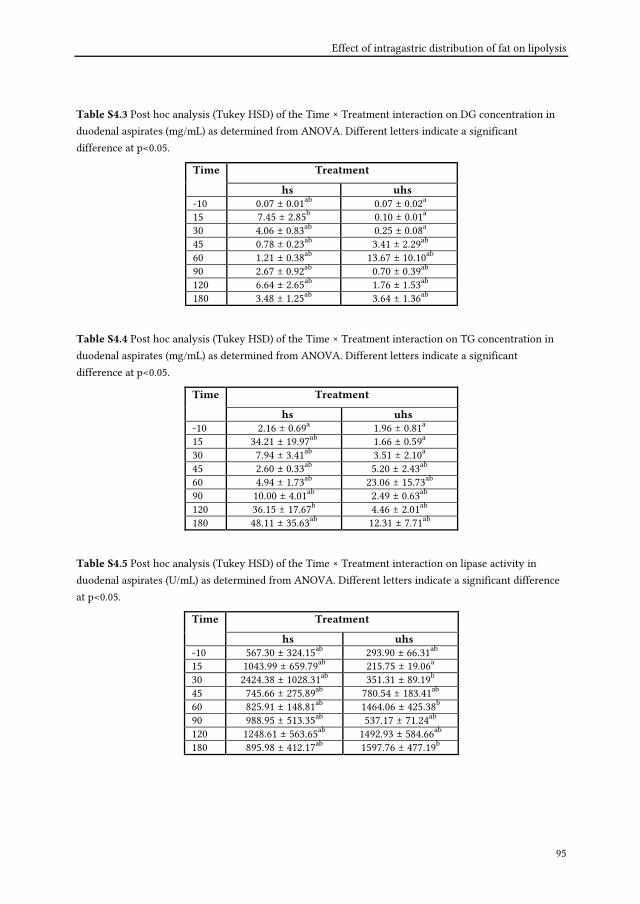
Effect of intragastric distribution of fat on lipolysis
95
Table S4.3 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on DG concentration in
duodenal aspirates (mg/mL) as determined from ANOVA. Different letters indicate a significant
difference at p<0.05.
Treatment Time
hs uhs
-10 0.07 ± 0.01ab 0.07 ± 0.02a
15 7.45 ± 2.85b 0.10 ± 0.01a
30 4.06 ± 0.83ab 0.25 ± 0.08a
45 0.78 ± 0.23ab 3.41 ± 2.29ab
60 1.21 ± 0.38ab 13.67 ± 10.10ab
90 2.67 ± 0.92ab 0.70 ± 0.39ab
120 6.64 ± 2.65ab 1.76 ± 1.53ab
180 3.48 ± 1.25ab 3.64 ± 1.36ab
Table S4.4 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on TG concentration in
duodenal aspirates (mg/mL) as determined from ANOVA. Different letters indicate a significant
difference at p<0.05.
Treatment Time
hs uhs
-10 2.16 ± 0.69a 1.96 ± 0.81a
15 34.21 ± 19.97ab 1.66 ± 0.59a
30 7.94 ± 3.41ab 3.51 ± 2.10a
45 2.60 ± 0.33ab 5.20 ± 2.43ab
60 4.94 ± 1.73ab 23.06 ± 15.73ab
90 10.00 ± 4.01ab 2.49 ± 0.63ab
120 36.15 ± 17.67b 4.46 ± 2.01ab
180 48.11 ± 35.63ab 12.31 ± 7.71ab
Table S4.5 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on lipase activity in
duodenal aspirates (U/mL) as determined from ANOVA. Different letters indicate a significant difference
at p<0.05.
Treatment Time
hs uhs
-10 567.30 ± 324.15ab 293.90 ± 66.31ab
15 1043.99 ± 659.79ab 215.75 ± 19.06a
30 2424.38 ± 1028.31ab 351.31 ± 89.19b
45 745.66 ± 275.89ab 780.54 ± 183.41ab
60 825.91 ± 148.81ab 1464.06 ± 425.38b
90 988.95 ± 513.35ab 537.17 ± 71.24ab
120 1248.61 ± 563.65ab 1492.93 ± 584.66ab
180 895.98 ± 412.17ab 1597.76 ± 477.19b

Chapter 4
96
References
Armand, M., Borel, P., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1992). Effects of
droplet size, triacylglycerol composition, and calcium on the hydrolysis of complex emulsions
by pancreatic lipase: an in vitro study. The Journal of Nutritional Biochemistry, 3(7), 333-341.
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V. &
Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the
human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096-1106.
Borel, P., Armand, M., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1994). Hydrolysis
of emulsions with different triglycerides and droplet sizes by gastric lipase in vitro. Effect on
pancreatic lipase activity. The Journal of Nutritional Biochemistry, 5(3), 124-133.
Carrière, F., Grandval, P., Renou, C., Palomba, A., Priéri, F., Giallo, J., Henniges, F., Sander-Struckmeier, S.
& Laugier, R. (2005). Quantitative study of digestive enzyme secretion and gastrointestinal
lipolysis in chronic pancreatitis. Clinical Gastroenterology and Hepatology, 3(1), 28-38.
Cortner, J. A., Coates, P. M., Swoboda, E. & Schnatz, J. D. (1976). Genetic variation of lysosomal acid
lipase. Pediatric Research, 10(11), 927-932.
Feinle-Bisset, C., Patterson, M., Ghatei, M. A., Bloom, S. R. & Horowitz, M. (2005). Fat digestion is required
for suppression of ghrelin and stimulation of peptide YY and pancreatic polypeptide secretion
by intraduodenal lipid. American Journal of Physiology - Endocrinology and Metabolism, 289(6),
E948-E953.
Feinle, C., D'Amato, M. & Read, N. W. (1996). Cholecystokinin-A receptors modulate gastric sensory and
motor responses to gastric distention and duodenal lipid. Gastroenterology, 110(5), 1379-1385.
Feinle, C., Rades, T., Otto, B. & Fried, M. (2001). Fat digestion modulates gastrointestinal sensations
induced by gastric distention and duodenal lipid in humans. Gastroenterology, 120(5), 1100-1107.
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P.
F., Stellaard, F. & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption
and increases plasma CCK but has minor effects on gastric emptying and appetite. American
Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982-991.
Fried, M., Mayer, E. A., Jansen, J. B. M. J., Lamers, C. B. H. W., Taylor, I. L., Bloom, S. R. & Meyer, J. H.
(1988). Temporal relationships of cholecystokinin release, pancreatobiliary secretion, and
gastric emptying of a mixed meal. Gastroenterology, 95(5), 1344-1350.
Gargouri, Y., Pieroni, G., Rivière, C., Lowe, P. A., Saunière, J.-F., Sarda, L. & Verger, R. (1986). Importance
of human gastric lipase for intestinal lipolysis: an in vitro study. Biochimica et Biophysica Acta -
Lipids and Lipid Metabolism, 879(3), 419-423.
Golding, M. & Wooster, T. J. (2010). The influence of emulsion structure and stability on lipid digestion.
Current Opinion in Colloid and Interface Science, 15(1-2), 90-101.
Heath, R. B., Jones, R., Frayn, K. N. & Robertson, M. D. (2004). Vagal stimulation exaggerates the
inhibitory ghrelin response to oral fat in humans. Journal of Endocrinology, 180(2), 273-281.

Effect of intragastric distribution of fat on lipolysis
97
Heddle, R., Dent, J., Read, N. W., Houghton, L. A., Toouli, J., Horowitz, M., Maddern, G. J. & Downton, J.
(1988). Antropyloroduodenal motor responses to intraduodenal lipid infusion in healthy
volunteers. American Journal of Physiology - Gastrointestinal and Liver Physiology, 254(5).
Helbig, A., Silletti, E., Timmerman, E., Hamer, R. J. & Gruppen, H. (2012). In vitro study of intestinal
lipolysis using pH-stat and gas chromatography. Food Hydrocolloids, 28(1), 10-19.
Helbig, A., Silletti, E., van Aken, G., Oosterveld, A., Minekus, M., Hamer, R. J. & Gruppen, H. (2013). Lipid
digestion of protein stabilized emulsions investigated in a dynamic in vitro gastro-intestinal
model system. Food Digestion, DOI: 10.1007/s13228-012-0029-6.
Keller, J., Rünzi, M., Goebell, H. & Layer, P. (1997). Duodenal and ileal nutrient deliveries regulate human
intestinal motor and pancreatic responses to a meal. American Journal of Physiology -
Gastrointestinal and Liver Physiology, 272(3 35-3), G632-G637.
Little, T. J., Russo, A., Meyer, J. H., Horowitz, M., Smyth, D. R., Bellon, M., Wishart, J. M., Jones, K. L. &
Feinle-Bisset, C. (2007). Free fatty acids have more potent effects on gastric emptying, gut
hormones, and appetite than triacylglycerides. Gastroenterology, 133(4), 1124-1131.
Maillot, F., Baulieu, F., Guilloteau, D., Boirie, Y., Garrigue, M. A., Hoinard, C. & Couet, C. (2008). Gut
emptying affects dietary fat contribution to postprandial lipemia following sequential meals in
healthy subjects. Nutrition, 24(7-8), 682-688.
Maljaars, P. W. J., Symersky, T., Kee, B. C., Haddeman, E., Peters, H. P. F. & Masclee, A. A. M. (2008).
Effect of ileal fat perfusion on satiety and hormone release in healthy volunteers. International
Journal of Obesity, 32(11), 1633-1639.
Marciani, L., Wickham, M., Singh, G., Bush, D., Pick, B., Cox, E., Fillery-Travis, A., Faulks, R., Marsden, C.,
Gowland, P. A. & Spiller, R. C. (2007). Enhancement of intragastric acid stability of a fat
emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder
contraction. American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(6),
G1607-G1613.
Marciani, L., Faulks, R., Wickham, M. S. J., Bush, D., Pick, B., Wright, J., Cox, E. F., Fillery-Travis, A.,
Gowland, P. A. & Spiller, R. C. (2009). Effect of intragastric acid stability of fat emulsions on
gastric emptying, plasma lipid profile and postprandial satiety. British Journal of Nutrition,
101(6), 919-928.
McClements, D. J., Decker, E. A. & Park, Y. (2009). Controlling lipid bioavailability through physico-
chemical and structural approaches. Critical Reviews in Food Science and Nutrition, 49(1), 48-67.
Phan, C. T. & Tso, P. (2001). Intestinal lipid absorption and transport. Frontiers in Bioscience, 6, D299-319.
Robertson, M. D., Mason, A. O. & Frayn, K. N. (2002). Timing of vagal stimulation affects postprandial
lipid metabolism in humans. American Journal of Clinical Nutrition, 76(1), 71-77.
Schwizer, W., Asal, K., Kreiss, C., Mettraux, C., Borovicka, J., Remy, B., Guzelhan, C., Hartmann, D. &
Fried, M. (1997). Role of lipase in the regulation of upper gastrointestinal function in humans.
American Journal of Physiology - Gastrointestinal and Liver Physiology, 273(3 36-3), G612-G620.

Chapter 4
98
Smeets, A. J. P. G. & Westerterp-Plantenga, M. S. (2006). Satiety and substrate mobilization after oral fat
stimulation. British Journal of Nutrition, 95(4), 795-801.
Thelen, K., Coboeken, K., Willmann, S., Dressman, J. B. & Lippert, J. (2012). Evolution of a detailed
physiological model to simulate the gastrointestinal transit and absorption process in humans,
part II: Extension to describe performance of solid dosage forms. Journal of Pharmaceutical
Sciences, 101(3), 1267-1280.
van Aken, G. A., Bomhof, E., Zoet, F. D., Verbeek, M. & Oosterveld, A. (2011). Differences in in vitro
gastric behaviour between homogenized milk and emulsions stabilised by Tween 80, whey
protein, or whey protein and caseinate. Food Hydrocolloids, 25(4), 781-788.

Chapter 5
Effect of oral administration and intragastric
distribution of lipids on satiety responses in
humans
To be submitted

Chapter 5
100
Abstract
Significantly higher FFA concentration were present in duodenal aspirates in humans
after the intragastric infusion of homogenized emulsion compared to a sample where
water infusion was followed by oil within the first 30 min after the application (Chapter
4). In this Chapter, we aimed to investigate if such imposed effects would influence
satiety responses. In short, 15 volunteers received a homogenized emulsion orally or by
gastric infusion or gastric infusion of water followed by the oil phase (un-homogenized
sample). In a subset 6 volunteers received gastric infusion of a homogenized emulsion or
of an un-homogenized sample. Gallbladder volume, plasma CCK, PYY, GLP-1 and ghrelin
concentration, satiety ratings and ad libitum intake were analysed. Gallbladder
contraction was strongest for the oral application of the homogenized emulsion and
lowest for the gastric infusion of the un-homogenized sample. Plasma CCK and PYY
concentrations were significantly higher for homogenized emulsions compared to the un-
homogenized sample. The response of PYY and GLP-1 to the un-homogenized sample was
delayed compared to the homogenized emulsion. However, unexpectedly, there were no
differences in ghrelin responses, satiety ratings and ad libitum intake between the
treatments. In conclusion, intragastric distribution of lipids influenced plasma hormone
concentrations and gallbladder motility, but had no effect on satiety as assessed via a
VAS. Also, there was no correlation between duodenal concentration of FFA and CCK
plasma concentration. As a tentative explanation, we postulate that hormonal induced
satiety feelings might have been overruled by other mechanisms such as gastric
distention.

Effect of intragastric distribution of fat on satiety
101
5.1 Introduction
Food intake is normally regulated by signals indicating start, i.e. hunger, and stop, i.e.
satiety. Hence, understanding these signals could help to understand how to regulate food
intake. Lipids are a key component of the daily diet and they are mostly present in food in
form of emulsions. The importance of the lipid phase on metabolic responses has been
highlighted recently (Golding et al., 2011; Marze & Choimet, 2012; Keogh et al., 2011). For
instance, the inclusion of solid fat has been used to introduce partial coalescence and
these emulsions had a prolonged effect on gastric emptying in vivo, but no influence on
satiety ratings (Keogh et al., 2011). Furthermore, it has been observed that the emulsion
droplet size affects the release of satiety hormones, such as cholecystokinin (CCK), the
appetite sensation and antropyloroduodenal motility when presented intraduodenal
(Seimon et al., 2009). It was shown that this effect was larger for small droplets compared
to large droplets. It is also known that the release of free fatty acids (FFA) from small
droplets is higher compared to large droplets (Armand et al., 1992; Borel et al., 1994;
Armand et al., 1999; Helbig et al., 2012). In addition, it has been reported that FFA
presented in the stomach were able to trigger CCK responses (McLaughlin et al., 1999;
Little et al., 2007). CCK was correlated to a higher feeling for fullness, a decreased gastric
emptying and a suppressed energy intake (Little et al., 2007). Gastric emulsions stability,
i.e. the persistence of small, homogenous distributed emulsions droplets in the gastric
environment is, therefore, thought to enhance satiety signals, and thus reduce
overconsumption. In fact, an intragastric stable emulsion led to higher CCK concentration
and higher gallbladder contractions (Marciani et al., 2007a), and increased fullness feeling,
reduced hunger and appetite feeling compared to an unstable emulsion (Marciani et al.,
2009). In these studies, MRI was used to monitor intragastric emulsions behaviour and
emptying of intragastric stable and unstable emulsions (Marciani et al., 2007a; Marciani et
al., 2009). However, the contribution of gastric and intestinal lipolysis and the resulting
gastric and duodenal lipolytic products could not be established. Although knowledge on
the gastric behaviour of emulsions in vitro is increasing, the relations to satiety responses
in vivo are still not fully understood. For example, layering of fat in the stomach resulted
in a fast emptying of the aqueous phase and a delayed emptying of the fat layer into the
duodenum (Foltz et al., 2009). However, in that study the increase in CCK and lipid
absorption was not consistent with changes of the subjective feelings for hunger, satiety
and fullness. In Chapter 4 we showed that significantly higher FFA concentrations were
present in duodenal aspirates after the intragastric infusion of an emulsion compared to a

Chapter 5
102
sample where water infusion was followed by oil. In the present chapter the objective is
to link changes in, for example, the FFA concentration to metabolic and satiety responses,
such as gallbladder contraction, CCK release, satiety perceptions and ad libitum intake.
Furthermore, as modified sham feeding has been shown to stimulate gallbladder
contraction (Witteman et al. 1993), reduce plasma ghrelin (Heath et al., 2004) and to
increase satiety (Smeets & Westerterp-Plantenga, 2006), we investigated the effect of oral
stimulation of fat on gallbladder volume, hormone release, i.e. CCK, ghrelin, peptide
tyrosine-tyrosine (PYY) and glukagon-like peptide-1 (GLP-1) and satiety perceptions.
5.2 Materials and methods
5.2.1 Emulsion preparation
Emulsions were produced following good manufacturing practice in a food grade pilot
plant of NIZO food research (Ede, The Netherlands). Tween 80 (Lamesorb©SMO20,
Cognis, Monheim am Rhein, Germany) solutions were prepared by dissolving powder in
0.01 M NaCl solution. The final emulsions contained 8% (w/w) sunflower oil (Reddy
sunflower oil, Vandermoortele, Izegem, Belgium) and 0.4% (w/w) Tween 80. All
ingredients used were food grade. In total 3 batches of each solution were prepared. Each
batch consisted of 10 L (20 x 500 mL) of each solution. Emulsions were homogenized at
80°C at a pressure of 70 to 180 bar followed by homogenization at 0 to 20 bar (Rannie
Lab12-16.5, Leuze, Waardenburg, The Netherlands). The solutions for the un-
homogenized samples, i.e. Tween 80-solution and sunflower oil, were heated separately at
80°C. These samples were not homogenized and applied separately but for the ease of the
reading we refer to them as emulsions. Homogenized and un-homogenized emulsions
were then sterilized in a Combitherm sterilizer (Minister 1987, AlfaLaval, Lund, Sweden)
for 7 seconds at 141°C. After cooling to room temperature, solutions were filled
aseptically in sterilized plastic containers and stored <7°C in a dark place until the start of
the experiments in order to prevent off-flavor formation. Of every batch 2 samples were
tested for sterility. During the course of the experiment fresh emulsions were prepared
every 6-10 weeks in order to prevent excessive off-flavor formation or formation of
physical instabilities. The surface-weighted-mean- diameter (d32) and the volume-
weighted-mean diameter (d43) of the freshly prepared homogenized emulsions were 2.33

Effect of intragastric distribution of fat on satiety
103
and 8.75 µm, respectively, as determined by dynamic light scattering (Mastersizer Hydro
2000S, Malvern Instruments, Malvern, UK) at room temperature.
During the production of the emulsions, 0.034% (w/w) and 0.012% (w/w) of vanilla
powder and saccharine, respectively, were added to the Tween 80 solutions. As a result,
the test drinks tasted pleasantly sweet with a sweet vanilla flavor having a somewhat
bitter off-note. On the days of intervention, 1g of paracetamol (Apotheek van Thoor,
Maastricht, The Netherlands) was manually added to the homogenized emulsion or
Tween 80 solution in the case of the un-homogenized emulsions to measure gastric
emptying of the water phase.
5.2.2 Subjects
15 healthy volunteers participated in the study (11 male and 4 female, age 28±8 y, body
mass index of 22.7±2.3 kg/m2). A short physical examination and a cognitive restraint
eating behaviour using a Dutch translation of the Three Factor Eating Questionnaire
(TFEQ) were performed. Participants did not have gastrointestinal or hepatic disorders or
previous major abdominal surgery interfering with gastrointestinal function. They were
non-smokers, moderate alcohol consumers (≤ 10 alcoholic consumptions per week), non-
medicated except of using contraceptives and non-restraint eaters according to the Three
Factor Eating Questionnaire (TFEQ Factor 1 score ≤ 9). The study was approved by the
Medical Ethics Committee of the Maastricht University Medical Center, conducted
according to the principles of the Declaration of Helsinki and in accordance with the
Medical Research Involving Human Subjects Act (WMO). All participants signed
informed consent. Volunteers were informed that they receive a lipid-containing drink
and that its behaviour in the stomach and duodenum as well as the resulting
physiological responses were investigated.
5.2.3 Study design
The study was a randomized crossover design and divided in two experiments. All
experiments took place on separate days. Subjects were requested to refrain from any
food or beverage intake with the exception of water from 2200h the evening before the
test day. Subjects received a total load of 500 mL of samples with a total energy content of
360 kcal.

Chapter 5
104
Experiment 1: Gastric intubation study
On all 3 test days of the gastric intubation study, subjects arrived in the morning at
0830h in fasted state. After local anesthesia of the nasal mucosa using lidocaine 10%
spray, each subject was intubated with a double-lumen nasogastric feeding catheter
(Bengmark® Nutricia, Amsterdam, The Netherlands). One lumen was used to inject
emulsions into the stomach and the other lumen was used to collect stomach fluid by
aspiration using a standard 10-mL syringe. The position of the nasogastric catheter was
controlled by determination of the pH of the aspirate. After intubation, a cannula was
placed in a superficial antecubital vein of the forearm for blood sampling. At 0900h,
subjects received the emulsions infused into the stomach either homogenized (hs) and in
case of the un-homogenized emulsions (uhs), the infusion of the water phase was
followed by the oil phase within 2 min. In the case of the oral administration, the
emulsions were homogenized (ho) and drunk within 2 min.
Experiment 2: Duodenal intubation study
As explained in Chapter 4, six volunteers, who participated in Experiment 1, joined on
two further occasions (duodenal intubation study), were intubated with a double-lumen
naso-duodenal catheter and received the emulsions un-homogenized (uhs) or
homogenized (hs). Samples from duodenal aspirates were drawn (Chapter 4) and, as
defined in the following section, samples from blood were collected and satiety ratings as
well as ad libitum intake was determined. These data were only used to correlate
Experiment 1 with Experiment 2 as described in 5.3.4. All other data were obtained from
Experiment 1.
5.2.4 Measurements
Gallbladder volume
Gallbladder volumes were measured in volunteers by real-time ultrasonography (3.5
MHz transducer, Technos Scientific, Concord, ON, Canada) at baseline (t=-10 min) and at
10, 15 min and then every 15 min until 180 min after application of the emulsions. The
assumptions and the mathematical formula used to calculate gallbladder volume have
been described and validated by Hopman and coworkers (Hopman et al., 1985). In short,
gall bladder volumes were calculated by the sum of cylinders method from real-time

Effect of intragastric distribution of fat on satiety
105
sonograms. This was done automatically by the computer connected to the echoscope
according to the following equation, where n is the number of cylinders, di is the diameter
of individual cylinders and hi is the height of individual cylinders:
V=∑n=i
n
π∗di
2hi
4
In this method, the longitudinal image of the gallbladder was divided into series of
equal height cylinders, which have a diameter perpendicular to the longitudinal axis of
the gallbladder image. The sum of volumes of these separate cylinders was the
uncorrected volume. A correction factor was calculated from the longitudinal and
transversal scans of the gallbladder to correct for the displacement of the longitudinal
image from the central axis. The gallbladder volume was then calculated by multiplication
of the uncorrected volume with the square of the correction factor.
Blood samples
During the test day, blood samples were taken at baseline (t=-10 min) and at 10, 15, 30,
45, 60, 90, 120 and 180 min after application of the emulsions. After sampling,
physiological salt solution (NaCl 0.9 % w/v) and heparine were injected to prevent
coagulation and occlusion of the catheter. Blood samples for ghrelin analysis were drawn
in EDTA tubes. EDTA tubes for cholecystokinin (CCK) and peptide YY (PYY) contained
aprotinin (Trasylol, Bayer, Leverkusen, Germany) at a concentration of 500 kallikrein
inhibitor units (KIU)/mL. Glucagon-like peptide 1 (GLP-1) was collected in special tubes
containing dipeptidyl peptidase IV (BD™ P700 Blood Collection and Preservation System
for GLP-1 Analysis, BD, Franklin Lakes, NJ, USA). All samples were kept on ice and all
tubes were centrifuged at 1500 × g for 10 min at 4°C within one hour. Aliquots of the
supernatant plasma samples were stored at -80°C until further analysis. RIA-kits were
used to analyse ghrelin (i.e. Human ghrelin (Total) RIA), PYY (i.e. Human PYY (Total)
RIA) and GLP-1 (i.e. Glucagon Like Peptide-1 (Total) RIA). These were obtained from
Millipore (Millipore, St. Charles, Missouri, USA) and kits to analyse CCK were from
Eurodiagnostica (EURIA-CCK, Eurodiagnostica, Malmö, Sweden). The hormones were
analysed at TNO Triskelion (Zeist, The Netherlands) using the RIA-kits according to the
manufacturer's instructions with minor modifications, optimized by TNO Triskelion, for
better performance and lower detection limits (unpublished data).

Chapter 5
106
Satiety ratings and ad libitum intake
Scores for satiety feelings (i.e. fullness, satiety, desire to eat, hunger) were measured
using Visual Analogue Scales (VAS) anchored with ‘not at all’ and ‘extremely’ at baseline
(t=-10 min), at t=30 min and then every 30 min until 180 min after application of the
emulsions. Volunteers were asked to indicate on a line of 10 cm which place on the scale
best reflected their feeling at that moment. The distance from 0 cm to the indication mark
was measured in cm.
After taking the last sample at t=180 min, subjects were extubated and received 30 min
thereafter a lunch, which was prepared according to Veldhorst (Veldhorst et al., 2009).
The food provided for lunch was weighed before and after eating. The energy intake (EI)
was calculated by multiplying the amount of food consumed by the energy value of the
food as indicated by the product label (11.4 kJ/g).
Statistics
Data were transformed and statistical testing was carried out as described in Chapter4
with some minor modifications as follows. Response curves of dependent measures were
modelled by fitting Weibull functions as described in Chapter 4 and onset times and peak
times were determined for hormones, gallbladder volume and satiety ratings. Multi-factor
ANOVA was used to test for significant effects of Time (fixed factor, t=-10”, “t=10”,
“t=15”, “t=30”, “t=45”, “t=60”, “t=90”, “t=120”, “t=180” for hormones including “t=75”,
“t=105”, “t=135”, “t=150” “t=175” for gallbladder volume and “t=-10”, “t=30”, “t=60”,
“t=90”, “t=120”, “t=150”, “t=180” for satiety ratings), Treatment (fixed factor, “ho”, “hs”,
“uhs”), Time × Treatment interaction and, in the case of onset and peak times, of Measure
(fixed factor, “hormones”, “gallbladder volume” and “satiety rating”), Treatment (fixed
factor,“ho”, “hs”, “uhs”) and Measure × Treatment interaction. A significant effect was
followed by a multiple pairwise post hoc test (Tukey HSD test) and in the case of onset
and peak times Within-categories of Measure were compared. In addition, data of
experiment 1 and 2 was correlated using Multi-factor ANOVA to test for significant
effects of Experiment × Treatment, Experiment × Time and Experiment × Treatment ×
Time interaction on plasma paracetamol concentration, hormone release, satiety
responses and ad libitum intake. These data were obtained from the 6 people who
participated in both experiments for the treatments hs and uhs.

Effect of intragastric distribution of fat on satiety
107
5.3 Results
5.3.1 Gallbladder motility
The ANOVA results are shown in Table 5.1. There was a significant effect of
Treatment and Time on gallbladder volume, but no Treatment × Time interaction
(p=0.07). Initial gallbladder volumes were 19.2±1.2 mL (Fig. 5.1). Post hoc analysis
revealed that gallbladder volumes were lower in the following order ho<hs<uhs
indicating a pronounced contraction of the gallbladder for treatment ho compared to hs,
whereas gallbladder contraction was barely stimulated by the treatment uhs.
Fig. 5.1 Change of gallbladder volumes in function of time observed during Experiment 1 for the
treatments ho (●), hs (□) and uhs (▲).Values are expressed as mean value ± Standard Error (SE). Di\erent
low letters indicate a significant difference at p<0.001.
5.3.2 Hormone release
In Figure 5.2 plasma concentrations of CCK, GLP-1, PYY and ghrelin dependent on
time and treatment are shown. Initial values of plasma CCK concentration were 0.47±0.03
pmol/L (Fig. 5.2A). ANOVA revealed main effects of Treatment and Time, and a
Treatment × Time interaction on plasma CCK (Table 5.1). Further post hoc analysis
showed significantly higher plasma CCK concentrations for the treatment ho and hs,
when both were compared to uhs (p<0.01). In detail, ho differed at t=10 and t=15 min and
hs differed at t=15 min from uhs. In addition, ho induced the longest elevation of plasma

Chapter 5
108
CCK concentrations between 10 and 90 min compared to baseline. For hs, CCK
concentration was only elevated at t=10 and t=15 min and for uhs highest concentrations
were found at t=45 min compared to baseline. For GLP-1 plasma concentrations, there
was a main effect of Time and a Treatment × Time interaction (Table 5.1). For example,
the initial GLP-1 plasma concentrations were 7.42±0.85 pmol/L (Fig. 5.2B) and increased
significantly for all treatments after application compared to baseline as revealed with
post hoc test. Further, there was a significant effect of Treatment and a Treatment × Time
interaction on plasma PYY concentration (Table 5.1). Initial values were 82.7±3.2 pg/mL
(Fig. 5.2C) and this concentration was significantly elevated between 15 and 180 min for
all treatments as revealed post hoc. Also, PYY concentrations differed for uhs from ho and
hs at t=30 min. There was no difference between hs and ho. There was a Time but no
Treatment effect (p=0.17) or Treatment × Time interaction (p=0.13) on the plasma ghrelin
concentration (Table 5.1). The initial amount of 883.4±26.2 pg/mL (Fig. 5.2D) was, as
revealed post hoc, significantly decreased between t=30 and 180 min for all treatments.
Post hoc results of main Treatment × Time interaction on CCK, GLP-1 and PYY are given
in the supplementary (Tables S5.1-S5.3).
Table 5.1 Effect of Time of plasma collection, or on gallbladder contraction or satiety ratings, emulsions
used as Treatment, and Time × Treatment interaction on gallbladder volume, plasma hormone
concentrations of CCK, GLP-1, PYY and ghrelin, and satiety ratings of fullness, satiety, hunger and desire
to eat.
Time Treatment Time x Treatment dependent variable
F (df1,df2) Sig F (df1,df2) Sig F (df1,df2) Sig
gallbladder volume 18.66 (13,364) *** 14.02 (2,364) *** 1.47 (26,364) Ns CCK 22.65 (8,224) *** 6.61 (2,224) ** 3.03 (16,224) *** GLP-1 42.46 (8,224) *** 1.08 (2,224) Ns 2.64 (16,224) *** PYY 37.69 (8,224) *** 2.72 (2,224) Ns 2.20 (16,224) ** ghrelin 47.10 (8,224) *** 1.87 (2,224) Ns 1.42 (16,224) Ns fullness 33.20 (6,168) *** 0.59 (2,168) Ns 1.13 (12,168) Ns satiety 17.57 (6,168) *** 0.21 (2,168) Ns 0.66 (12,168) Ns hunger 18.70 (6,168) *** 0.31 (2,168) Ns 1.78 (12,168) Ns desire to eat 23.27 (6,168) *** 0.59 (2,168) Ns 2.51 (12,168) **
Significant differences are indicated with asterisks: *p<0.05, **p< 0.01, ***p < 0.001, based on ANOVA analysis. Ns is not significant.

Effect of intragastric distribution of fat on satiety
109
Fig. 5.2 Change of plasma CCK (A), GLP-1 (B), PYY (C) and ghrelin (D) concentration in function of time
observed during Experiment 1 for the treatments ho (●), hs (□) and uhs (▲). Values are expressed as mean
± SE. Different low letters indicate a significant difference at p<0.01.

Chapter 5
110
5.3.3 Satiety ratings and subsequent energy intake
There was a main effect of Time on all satiety ratings as evaluated with ANOVA
(Table 5.1). Treatment × Time interaction was only observed on the rating “desire to eat”
but not on “hunger” (p=0.06), “fullness” (p=0.34) and “satiety” (p=0.79). Post hoc analysis
on Time revealed that the perception “desire to eat” was significantly decreased at t=30
min for all treatments (Fig. 5.3). Compared to t=30 min, it was significantly increased for
hs between 120 and 180 min and for ho between 90 and 180 min, but not for uhs (Table
S5.4). For all treatments, “hunger” was significantly decreased between 30 and 60 min
compared to baseline. The perceptions “fullness” and “satiety” were significantly elevated
between 30 and 90 min and 30 and 60 min, respectively, for all treatments.
Subsequent ad libitum energy intake was 2318.1±279.2, 2501.0±178.7 and 2123.3±291.1
kJ for ho, hs and uhs, respectively and there was no difference between the treatments as
determined with ANOVA (p=0.32).
5.3.4 Correlation of gastric and duodenal intubation study
Data obtained in the gastric and duodenal intubation study was compared by means of
ANOVA for the treatments hs and uhs and was from 6 people who participated in both
experiments. In the case that this data, i.e. paracetamol plasma concentrations, hormone
concentrations, satiety ratings and ad libitum intake, reveals similar outcomes, a
correlation between both experiments is considered. There was no difference in plasma
paracetamol concentrations obtained in the gastric and duodenal intubation study, i.e.
Experiment × Treatment (p = 0.40), Experiment × Time, (p=0.54), and Experiment ×
Treatment × Time (p = 0.80). Further, there was no difference in hormone concentrations
between the Experiments, i.e. Experiment × Treatment × Time interaction for CCK was p
= 0.64, for GLP-1 was p = 0.96, for PYY was p = 0.64 and for ghrelin was p = 0.32,
respectively. Additionally, satiety ratings and ad libitum intake did not differ between the
experiments. Hence, conclusions can be drawn by correlating both experiments. We,
therefore, correlated gallbladder volume with plasma CCK concentration (both obtained
from the gastric intubation study), gallbladder with FFA concentration in duodenal
aspirates (the latter was obtained from the duodenal intubation study) and plasma CCK
concentration with FFA concentration in duodenal aspirates (Fig. 5.4). With increasing
plasma CCK concentration the gallbladder volume was decreasing (Fig. 5.4A). Likewise,
with increasing FFA concentration in duodenal aspirates, gallbladder volume was

Effect of intragastric distribution of fat on satiety
111
decreasing (Fig. 5.4B). However, there is no correlation between FFA concentrations in
duodenal aspirates and plasma CCK concentration (Fig. 5.4C).
Fig. 5.3 Change of fullness (A), satiety (B), hunger (C) and desire to eat (D) ratings in function of time
observed during Experiment 1 for the treatments ho (●), hs (□) and uhs (▲).Values are expressed as mean
± SE.

Chapter 5
112
Fig. 5.4 Correlation of gallbladder volumes with plasma CCK concentrations (A) and with the FFA
concentration as determined in duodenal aspirates (B), and correlation of plasma CCK concentration and
the FFA concentration as determined in duodenal aspirates (C) at corresponding time points after the
treatment hs (□) and uhs (▲). Values are expressed as mean ± SE.

Effect of intragastric distribution of fat on satiety
113
5.3.5 T-peak and t-onset
As described in Chapter 4, the onset and peak times were extracted from Weibull fits
to determine timed responses of Measures (Fig. 5.5). When comparing timed responses of
hormones, ANOVA revealed main effects of Measure and Treatment but no Measure ×
Treatment interaction on t-peak (Table 5.2). As evaluated with post hoc analysis,
hormones t-peak appeared significantly in the order of CCK<GLP-1<PYY<ghrelin and
occurred significantly earlier for ho and hs compared to uhs. On t-onset the main effects
of Measure, Treatment and a Measure × Treatment interaction were found by ANOVA.
Post hoc analysis revealed that these effects were caused by a significantly earlier t-onset
for CCK compared to the other hormones and significantly later responses for uhs
compared to ho and hs. The Measure × Treatment interaction were caused by the late t-
onset of GLP-1 and PYY in response to uhs.
Fig. 5.5 Comparison of peak times and onset times of timed responses for gallbladder volume (GBV),
plasma hormone concentrations, i.e CCK, GLP-1, PYY and ghrelin, and satiety ratings, i.e fullness, satiety,
hunger and desire to eat, for the treatments ho, hs and uhs (gastric intubation study). Values are
expressed as mean ± SE. Different low letters indicate a significant difference at p<0.05.

Chapter 5
114
When comparing timed responses of satiety perceptions and hormone concentration
main effects of Measure and Treatment but no Measure × Treatment interaction on t-peak
were determined with ANOVA (Table 5.2). On t-onset, there were main effects of
Measure and a Measure × Treatment interaction, but no Treatment effect. When
evaluating Within-category effects of Measures post hoc it was found, for example, that t-
peak were earlier for CCK and the perceptions fullness, hunger and desire to eat (Fig. 5.5).
The perceived satiety had a later t-peak compared to CCK, and satiety was associated
with GLP-1. T-peak of ghrelin was latest compared to all other, and PYY was similar to
GLP-1. The interaction Measure × Treatment and the effect of Measure, as shown in
Table 5.2 (ANOVA), on t-onset was entirely caused by the late t-onset of hormones for
the treatment uhs as revealed post hoc.
Table 5.2 Effect of Categories including several Measures on t-peak and t-onset, emulsions used as
Treatment, and Measure × Treatment interaction on t-peak and t-onset.
Measure Treatment Measure x
Treatment
Category of Measures
F (df1,df2) Sig F (df1,df2) Sig F (df1,df2) Sig
t-peak hormones§ 102.30 (3,84) *** 3.49 (2,84) * 0.24 (6,84) Ns t-onset hormones§ 10.71 (3,84) *** 7.77 (2,84) ** 2.35 (6,84) * t-peak VAS# + lipids†-GA 13.05 (7,196) *** 0.41 (2,196) Ns 0.53 (14,196) Ns t-onset VAS# + lipids†-GA 1.98 (7,196) Ns 1.80 (2,196) Ns 0.71 (14,196) Ns t-peak VAS# + lipids†-DA 3.50 (7,35) ** 15.0 (1,35) * 0.77 (7,35) Ns t-onset VAS# + lipids†-DA 0.85 (7,35) Ns 1.61 (1,35) Ns 1.20 (7,35) Ns t-peak VAS# + hormones§ 28.68 (7,196) *** 3.38 (2,196) * 0.39 (14,196) Ns t-onset VAS# + hormones§ 5.92 (7,196) *** 2.64 (2,196) Ns 2.53 (14,196) ** t-peak hormones§ + lipids†-GA 31.11 (7,196) *** 0.56 (2,196) Ns 0.75 (14,196) Ns t-onset hormones§ + lipids†-GA 4.98 (7,196) *** 7.30 (2,196) ** 0.92 (14,196) Ns t-peak hormones§ + lipids†-DA 1.15 (7,35) Ns 1.63 (1,35) Ns 1.0 (7,35) Ns t-onset hormones§ + lipids†-DA 1.01 (7,35) Ns 13.2 (1,35) * 0.74 (7,35) Ns t-peak$ GBV + CCK + FFA-DA 2.89 (2,10) Ns 2.94 (1,10) Ns 2.52 (2,10) Ns t-onset$ GBV + CCK + FFA-DA 5.81 (2,10) * 1.47 (1,10) Ns 1.19 (2,10) Ns
Significant differences are indicated with asterisks: *p<0.05, **p< 0.01, ***p < 0.001, based on ANOVA analysis. Ns is not significant. § Category includes measures CCK, GLP-1, PYY and ghrelin # Category includes measures fullness, satiety, hunger and desire to eat † includes measures FFA, MG, DG and TG $ Measures GBV, CCK are depicted from gastric intubation study and compared to FFA-DA for treatments hs and uhs GA, gastric aspirate; DA, duodenal aspirate; GBV, gallbladder volume
Including the results on lipids as described in Chapter 4, ANOVA revealed similar
onset times for satiety ratings and lipids in gastric aspirates (Table 5.2). T-onset was
significantly later for lipids in the duodenal aspirates compared to lipids in gastric
aspirates and satiety ratings (post hoc, p<0.01).

Effect of intragastric distribution of fat on satiety
115
5.4 Discussion
The behaviour of emulsions in the stomach in relation to satiety responses is still not
fully understood with respect to the contribution of intragastric and intraduodenal
lipolytic products as a result of their hydrolysis. In the present study we aimed to
associate the intragastric and intraduodenal lipid hydrolysis with metabolic and satiety
responses, such as gallbladder contraction, hormone release, satiety perceptions and ad
libitum intake. Moreover, the impact of oral processing on these responses was
investigated.
5.4.1 Additive effect of oral processing on gallbladder contraction,
hormonal responses, satiety ratings and ad libitum intake
It has been shown by Witteman et al. (1993), that for modified sham feeding fat
stimulated the gallbladder contraction but not CCK release (Witteman et al., 1993). They
associated gallbladder contraction with CCK independent vagal cholinergic stimulation
mechanisms induced by olphactoric and gustatoric chemoreceptors in the mouth. In
Figure 5.1 we have shown a significantly prolonged contraction of the gallbladder after
oral application compared to intragastric application of the same emulsion. To our
knowledge we are the first to report this. This finding points towards an additive effect of
oral processing on responses relevant for lipid digestion. In agreement to Witteman and
coworkers (Witteman et al., 1993), we found no significant difference in CCK release
between the oral and intragastric application of the emulsions. There was also no additive
effect of oral processing on plasma GLP-1, PYY and ghrelin concentrations. It has been
described, though, that ghrelin was reduced upon modified sham feeding (Heath et al.,
2004). This suppression in ghrelin release was related to reduction in appetite (Heath et
al., 2004). In addition, modified sham feeding was shown to increase satiety (Smeets et al.,
2006). In the present study, only the satiety rating “desire to eat” was significantly
elevated earlier, i.e. at t=90 min, for the oral application compared to the intragastric
application, which elevated at t=120 min. In addition, there was no additive effect of oral
application on the ad libitum intake. Recently, it has been reported that the duration of
oral exposure has a strong influence on the regulation of energy intake (Wijlens et al.,
2012). It was shown, that eight minutes of oral exposure decreased ad libitum intake
(Wijlens et al., 2012). However, this is surely not realistic for drinking an emulsion.

Chapter 5
116
5.4.2 Effect of intragastric lipid distribution on gallbladder
contraction, hormonal responses, satiety ratings and ad libitum
intake
Correlation between gallbladder contraction, CCK and FFA
Our data clearly shows the correlation of gallbladder volumes with CCK plasma
concentrations (Fig. 5.4A) and this is in agreement with previous observations (Marciani
et al., 2007a). In addition, high CCK plasma concentrations were found after
administration of the homogenized emulsions (Fig. 5.2). CCK is known to regulate
gallbladder contraction and bile secretion after food intake by binding to G-protein
coupled CCK-A-receptors on the smooth muscle of the gallbladder (Wank, 1998). The
release of CCK and the subsequent contraction of the gallbladder has been attributed to
the release of FFA (Hildebrand et al., 1998). Accordingly, we found significantly higher
gallbladder volumes after the un-homogenized emulsion compared to the homogenized
emulsion and gallbladder volumes correlated with FFA concentrations in the duodenal
aspirates (Fig. 5.4B). However, as shown in Fig. 5.4C, we did not find a correlation
between plasma CCK concentration and intraduodenal FFA as released during
intraduodenal lipolysis and analysed according to the method previously described
(Helbig et al. 2012). It is possible, as hypothesized before, that due to the high gallbladder
contraction and the resulting bile secretion FFA are absorbed quickly, thereby removing
CCK stimulating agents which results in a low response in CCK (Liddle, 1995). The
response in CCK release was, as expected, low (Fig. 5.2A) since only low concentrations
of FFA were present in the duodenum (Chapter 4).
Correlation between lipids, hormone release and satiety
Duodenal infusion of triglyceride has been shown not only to increase CCK but also
GLP-1 (Feinle et al., 2003) and PYY and to decrease ghrelin (Feinle-Bisset et al., 2005). This
effect was abolished when a lipase inhibitor, i.e. tetrahydrolipstatin, was applied
indicating the necessity of intraduodenal lipolysis for the release of these hormones.
Accordingly, GLP-1 and PYY concentrations increased for hs (Fig. 5.2B, C), which
correlate with the presence of lipolytic products in duodenal aspirates (Chapter 4).
Nevertheless, these hormone concentrations increased as well for uhs, although much
later as shown by the later onset times. In contrast to CCK, GLP-1 and PYY, ghrelin

Effect of intragastric distribution of fat on satiety
117
decreased independently from the intragastric distribution of the lipids. This finding
might indicate that already low concentrations of lipids are sufficient to supress ghrelin
or that its suppression is predominantly caused by, for example, intragastric distension.
In summary, a clear influence of the intragastric distribution of the lipid phase on the
gallbladder contraction as well as on the release of CCK, GLP-1 and PYY was observed.
Especially for gallbladder volume this can be associated with the intraduodenal FFA
concentration. However, this was neither reflected in the perception of satiety nor in the
ad libitum intake. Only on the perception “desire to eat” a stronger response was seen for
hs compared to uhs. Whether the identical ghrelin release obtained for both treatments
plays a role in the similar perceptions of satiety found in the present study is still
speculative. Also for this hormone only a minor change of the satiety perception
“prospective consumption” was found in earlier studies although plasma ghrelin
concentration were differently suppressed after various duodenal fat infusions (Feinle et
al., 2003; Feinle-Bisset et al., 2005). We therefore hypothesize, that the satiating effect of
food can not only be predicted by changes in hormonal circulation levels. We might
speculate that the similar effect of hs and uhs on satiety perceptions could be due to the
initial volume of the meal. In fact, it has been reported that the sense of fullness is
proportional to postprandial gastric volume (Marciani et al., 2001; Marciani et al., 2007b).
As proposed earlier (Goetze et al., 2007; Foltz et al., 2009) and since we applied 500 mL of
the emulsions having the same energy content and the same composition, the initial
volume might have had a larger effect on satiety ratings compared to that of the lipids
itself.
5.5 Conclusions
In conclusion, an additive effect of oral intake of an emulsion compared to the same
emulsion applied intragastrical was found on gallbladder contraction which points toward
the supporting influence of oral processing on metabolic responses. Furthermore, the
differences in intragastric distribution of the lipid phase influenced the release of FFA into
the duodenum and metabolic responses, such as plasma hormone concentrations and
gallbladder motility, but had no major effect on satiety feelings. Unexpectedly, there was
no correlation between duodenal concentration of FFA and CCK plasma concentration,
which suggests that satiety feelings might be induced by other mechanisms such as

Chapter 5
118
intragastric volume and hence gastric distention rather than by hormonal responses as
such.
Acknowledgements
The authors gratefully thank Daniel Keszthelyi, Samefko Ludidi and Mark van Avesaat
(Division of Gastroenterology-Hepatology, Department of Internal Medicine, University
Hospital Maastricht) for their technical contribution to the clinical trial. George van Aken
and Lex Oosterveld (NIZO food research) are acknowledged for their technical expertise
and contribution to the experimental set up. We thank all participants in this study and
Eefjan Timmermann (TI Food and Nutrition) for his technical assistance during the
measurements.

Effect of intragastric distribution of fat on satiety
119
5.6 Supplementary data
Table S5.1 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on plasma CCK
concentration (pmol/L) as determined from ANOVA. Different letters indicate a significant difference at
p<0.05.
Treatment Time
ho hs uhs -10 0.45 ± 0.05ab 0.46 ± 0.06abcd 0.48 ± 0.07a
10 2.92 ± 0.52j 2.58 ± 0.50ghij 0.79 ± 0.11abcdefg
15 3.85 ± 1.55ij 2.62 ± 0.63hij 0.83 ± 0.16abcdef
30 1.52 ± 0.27fghij 1.13 ± 0.17defghi 0.86 ± 0.12abcdef
45 1.05 ± 0.08defghi 1.08 ± 0.12defghi 1.01 ± 0.16bcdefg
60 1.42 ± 0.40efghij 1.17 ± 0.18defghi 0.92 ± 0.15abcdef
90 1.05 ± 0.15cdefgh 0.92 ± 0.08abcdef 0.84 ± 0.15abcdef
120 0.79 ± 0.10abcde 0.73 ± 0.11abcdef 0.64 ± 0.10abcdef
180 0.52 ± 0.06abc 0.51 ± 0.08ab 0.52 ± 0.07abcde
Table S5.2 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on plasma GLP-1
concentration (pmol/L) as determined from ANOVA. Different letters indicate a significant difference at
p<0.05.
Treatment Time
ho hs uhs
-10 7.77 ± 0.87abcd 8.93 ± 2.19ab 5.60 ± 0.79a
10 11.27 ± 1.97abcd 12.50 ± 18.64abcd 9.71 ± 2.67abc
15 11.64 ± 2.49bcdefg 18.64 ± 2.94cdefgh 12.07 ± 2.02abcde
30 34.27 ± 4.93hijk 34.87 ± 5.37ijk 19.87 ± 3.46defghij
45 26.33 ± 3.09ghijk 34.47 ± 3.52jk 28.33 ± 6.24efghijk
60 25.13 ± 2.59ghijk 30.40 ± 3.47hijk 31.60 ± 5.99hijk
90 28.33 ± 3.98ghijk 29.79 ± 2.88hijk 37.93 ± 4.03k
120 23.47 ± 2.73fghijk 23.13 ± 2.00ghijk 32.43 ± 3.09hijk
180 14.62 ± 2.13bcdef 16.93 ± 2.05defghi 23.00 ± 3.94defghij

Chapter 5
120
Table S5.3 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on plasma PYY
concentration (pg/mL) as determined from ANOVA. Different letters indicate a significant difference at
p<0.05.
Treatment Time
ho hs uhs
-10 81.67 ± 4.92a 81.80 ± 6.28ab 84.64 ± 5.82ab
10 91.00 ± 4.87ab 86.93 ± 8.35a 87.50 ± 6.86ab
15 101.13 ± 6.09abc 95.93 ± 8.58abc 90.67 ± 5.79ab
30 140.93 ± 12.78ef 137.40 ± 11.81ef 105.80 ± 7.87bcd
45 152.27 ± 9.65f 152.53 ± 12.25ef 125.80 ± 14.12cde
60 151.73 ± 8.51f 153.27 ± 10.25f 134.20 ± 12.97def
90 155.13 ± 10.12f 148.14 ± 8.50f 145.21 ± 11.23ef
120 153.36 ± 11.83f 149.93 ± 9.47ef 152.57 ± 12.36f
180 140.46 ± 12.61ef 137.53 ± 10.58ef 128.86 ± 8.66ef
Table S5.4 Post hoc analysis (Tukey HSD) of the Time × Treatment interaction on the satiety perception
`desire to eat´ as determined from ANOVA. Different letters indicate a significant difference at p<0.05.
Treatment Time
ho hs uhs
-10 4.27 ± 0.63bdefg 4.44 ± 0.48cfg 4.70 ± 0.50dg
30 2.34 ± 0.67ac 2.71 ± 0.52abde 3.16 ± 0.57abcef
60 3.25 ± 0.67abcdf 3.76 ± 0.60abcde 3.55 ± 0.63abcdefg
90 3.89 ± 0.66bdefg 4.02 ± 0.68abcde 3.83 ± 0.57abcdefg
120 4.55 ± 0.62bdefg 4.50 ± 0.69cfg 4.14 ± 0.65 abcdefg
150 4.79 ± 0.58eg 5.54 ± 0.67fg 4.25 ± 0.63abcdefg
180 5.02 ± 0.59bdefg 5.84 ± 0.66g 5.05 ± 0.66abcdefg

Effect of intragastric distribution of fat on satiety
121
References
Armand, M., Borel, P., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1992). Effects of
droplet size, triacylglycerol composition, and calcium on the hydrolysis of complex emulsions
by pancreatic lipase: an in vitro study. The Journal of Nutritional Biochemistry, 3(7), 333-341.
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V. &
Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the
human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096-1106.
Borel, P., Armand, M., Ythier, P., Dutot, G., Melin, C., Senft, M., Lafont, H. & Lairon, D. (1994). Hydrolysis
of emulsions with different triglycerides and droplet sizes by gastric lipase in vitro. Effect on
pancreatic lipase activity. The Journal of Nutritional Biochemistry, 5(3), 124-133.
Ellrichmann, M., Ritter, P. R., Otte, J.-M., Schrader, H., Banasch, M., Brunke, G., Herzig, K.-H., Seebeck, J.,
Schmidt, W. E. & Schmitz, F. (2007). Orlistat reduces gallbladder emptying by inhibition of CCK
release in response to a test meal. Regulatory Peptides, 139(1-3), 136-140.
Feinle-Bisset, C., Patterson, M., Ghatei, M. A., Bloom, S. R. & Horowitz, M. (2005). Fat digestion is required
for suppression of ghrelin and stimulation of peptide YY and pancreatic polypeptide secretion
by intraduodenal lipid. American Journal of Physiology - Endocrinology and Metabolism, 289(6),
E948-E953.
Feinle, C., O'Donovan, D., Doran, S., Andrews, J. M., Wishart, J., Chapman, I. & Horowitz, M. (2003).
Effects of fat digestion on appetite, APD motility, and gut hormones in response to duodenal
fat infusion in humans. American Journal of Physiology - Gastrointestinal and Liver Physiology,
284(5 47-5), G798-G807.
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P.
F., Stellaard, F. & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption
and increases plasma CCK but has minor effects on gastric emptying and appetite. American
Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982-991.
Goetze, O., Steingoetter, A., Menne, D., Van Der Voort, I. R., Kwiatek, M. A., Boesiger, P., Weishaupt, D.,
Thumshirn, M., Fried, M. & Schwizer, W. (2007). The effect of macronutrients on gastric
volume responses and gastric emptying in humans: A magnetic resonance imaging study.
American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(1), G11-G17.
Golding, M., Wooster, T. J., Day, L., Xu, M., Lundin, L., Keogh, J. & Clifton, P. (2011). Impact of gastric
structuring on the lipolysis of emulsified lipids. Soft Matter, 7(7), 3513-3523.
Heath, R. B., Jones, R., Frayn, K. N. & Robertson, M. D. (2004). Vagal stimulation exaggerates the
inhibitory ghrelin response to oral fat in humans. Journal of Endocrinology, 180(2), 273-281.
Helbig, A., Silletti, E., Timmerman, E., Hamer, R. J. & Gruppen, H. (2012). In vitro study of intestinal
lipolysis using pH-stat and gas chromatography. Food Hydrocolloids, 28(1), 10-19.
Helbig, A., Silletti, E., van Aken, G., Oosterveld, A., Minekus, M., Hamer, R. J. & Gruppen, H. (2013). Lipid
digestion of protein stabilized emulsions investigated in a dynamic in vitro gastro-intestinal
model system. Food Digestion, DOI: 10.1007/s13228-012-0029-6.

Chapter 5
122
Hildebrand, P., Petrig, C., Burckhardt, B., Ketterer, S., Lengsfeld, H., Fleury, A., Hadvay, P. & Beglinger, C.
(1998). Hydrolysis of dietary fat by pancreatic lipase stimulates cholecystokinin release.
Gastroenterology, 114(1), 123-129.
Hopman, W. P. M., Brouwer, W. F. M. & Rosenbusch, G. (1985). A computerized method for rapid
quantification of gallbladder volume from real-time sonograms. Radiology, 154(1), 236-237.
Keogh, J. B., Wooster, T. J., Golding, M., Day, L., Otto, B. & Clifton, P. M. (2011). Slowly and rapidly
digested fat emulsions are equally satiating but their triglycerides are differentially absorbed
and metabolized in humans. Journal of Nutrition, 141(5), 809-815.
Liddle, R. A. (1995). Regulation of cholecystokinin secretion by intraluminal releasing factors. American
Journal of Physiology - Gastrointestinal and Liver Physiology, 269(3 32-3), G319-G327.
Little, T. J., Russo, A., Meyer, J. H., Horowitz, M., Smyth, D. R., Bellon, M., Wishart, J. M., Jones, K. L. &
Feinle-Bisset, C. (2007). Free fatty acids have more potent effects on gastric emptying, gut
hormones, and appetite than triacylglycerides. Gastroenterology, 133(4), 1124-1131.
Marciani, L., Gowland, P. A., Spiller, R. C., Manoj, P., Moore, R. J., Young, P. & Fillery-Travis, A. J. (2001).
Effect of meal viscosity and nutrients on satiety, intragastric dilution, and emptying assessed
by MRI. American Journal of Physiology - Gastrointestinal and Liver Physiology, 280(6), G1227-
1233.
Marciani, L., Wickham, M., Singh, G., Bush, D., Pick, B., Cox, E., Fillery-Travis, A., Faulks, R., Marsden, C.,
Gowland, P. A. & Spiller, R. C. (2007a). Enhancement of intragastric acid stability of a fat
emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder
contraction. American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(6),
G1607-G1613.
Marciani, L., Wickham, M. S. J., Bush, D., Faulks, R., Wright, J., Fillery-Travis, A. J., Spiller, R. C. &
Gowland, P. A. (2007b). Magnetic resonance imaging of the behaviour of oil-in-water emulsions
in the gastric lumen of man. British Journal of Nutrition, 95(02), 331-339.
Marciani, L., Faulks, R., Wickham, M. S. J., Bush, D., Pick, B., Wright, J., Cox, E. F., Fillery-Travis, A.,
Gowland, P. A. & Spiller, R. C. (2009). Effect of intragastric acid stability of fat emulsions on
gastric emptying, plasma lipid profile and postprandial satiety. British Journal of Nutrition,
101(6), 919-928.
Marze, S. & Choimet, M. (2012). In vitro digestion of emulsions: Mechanistic and experimental models.
Soft Matter, 8(42), 10982-10993.
McLaughlin, J., Luca, M. G., Jones, M. N., D'Amato, M., Dockray, G. J. & Thompson, D. G. (1999). Fatty
acid chain length determines cholecystokinin secretion and effect on human gastric motility.
Gastroenterology, 116(1), 46-53.
Seimon, R. V., Wooster, T., Otto, B., Golding, M., Day, L., Little, T. J., Horowitz, M., Clifton, P. M. & Feinle-
Bisset, C. (2009). The droplet size of intraduodenal fat emulsions influences
antropyloroduodenal motility, hormone release, and appetite in healthy males. The American
Journal of Clinical Nutrition, 89(6), 1729-1736.
Smeets, A. J. P. G. & Westerterp-Plantenga, M. S. (2006). Satiety and substrate mobilization after oral fat
stimulation. British Journal of Nutrition, 95(4), 795-801.

Effect of intragastric distribution of fat on satiety
123
Veldhorst, M. A. B., Nieuwenhuizen, A. G., Hochstenbach-Waelen, A., Westerterp, K. R., Engelen, M. P. K.
J., Brummer, R. J. M., Deutz, N. E. P. & Westerterp-Plantenga, M. S. (2009). Effects of complete
whey-protein breakfasts versus whey without GMP-breakfasts on energy intake and satiety.
Appetite, 52(2), 388-395.
Wank, S.A. (1998). G protein-coupled receptors in gastrointestinal physiology I. CCK receptors: An
exemplary family. American Journal of Physiology - Gastrointestinal and Liver Physiology,
274(4), G607-G613.
Wijlens, A. G. M., Erkner, A., Alexander, E., Mars, M., Smeets, P. A. M. & De Graaf, C. (2012). Effects of
oral and gastric stimulation on appetite and energy intake. Obesity, 20(11), 2226-2232.
Witteman, B. J. M., Jebbink, M. C. W., Hopman, W. P. M., Masclee, A. A. M., Lamers, C. B. H. W. & Jansen,
J. B. M. J. (1993). Gallbladder responses to modified sham feeding: Effects of the composition of
a meal. Journal of Hepatology, 19(3), 465-469.

124

Chapter 6
General Discussion

Chapter 6
126
As described in the Introduction section, a wide range of in vitro models is presently
available. These models simulate gastric and intestinal lipolysis either separately or in
combination, while containing gastric and intestinal fluids of different compositions. Until
recently, most of the research reported in literature had been conducted to simulate
intestinal lipolysis as the intestine is the major site of lipid digestion. Also, most of the
published work focuses on the release of free fatty acid (FFA), while little is known on the
formation of monoglyceride (MG) and diglyceride (DG). Furthermore, little is known on
the gastric behaviour of emulsions and the resulting effects on the intestinal lipolysis.
Most in vitro models used did not include the gastric compartment and when it was
included, it was mimicked simple. At the same time limited data was available from
human studies on the behavior of emulsions. With respect to human studies, mostly small
intestinal perfusion studies have been used to gain mechanistic understanding of lipid
digestion neglecting the impact of gastric digestion. Also, the intragastric release of FFA
as a result of triglyceride lipolysis, the corresponding release of hormones, as detected by
FFA, at the intestinal site as well as the resulting satiety perceptions were poorly
understood. Hence, the many questions remained: What are possible mechanisms, which
effect the digestion of triglycerides, what is the impact of gastric passage and what are the
consequences for satiety perception? Therefore, the aim of this study was to build up
knowledge concerning the influence of emulsion parameters on the release of FFA, MG,
and DG. In addition, the impact of the gastric passage on lipolysis was investigated.
Finally, in a human study the lipolysis of emulsions was correlated with the subsequent
satiety perceptions.
6.1 Mechanisms that effect the digestion of triglycerides
6.1.1 Effect of emulsion properties on lipid digestion
In Chapters 2 and 3, two in vitro models, i.e. a static intestinal and a dynamic
gastrointestinal, were used to investigate the release of FFA from emulsions. As both the
oil-water interface of an emulsion and the droplet size are key factors influencing the
release of FFA, the investigated emulsions were varied in type of emulsifier, i.e. gum
arabic, WPI and lysozyme, and in droplet size. It was shown (Chapter 2) that in a
simulated intestinal environment the release of FFA was higher from emulsions stabilized
by gum arabic compared to WPI. Hence, a clear influence of the type of emulsifier on the
release of FFA was shown, comparable to other studies (Mun et al., 2007; Hu et al., 2010).

General Discussion
127
It must be noted though, as shown recently, that changing the oil type reduces the impact
of emulsifier type on lipid digestion (Marze & Choimet, 2012). This effect was not
investigated in the present study. In addition, there is obviously an influence of the
interface on the hydrolysis rate of DG into MG, which is, until now, only described for
lipases depending on the origin (Carriere et al., 1991; Rodriguez et al., 2008). Furthermore,
a clear effect of the surface area was observed. This showed that the larger the area the
higher the rate of digestion, which is in agreement with previous studies (Li &
McClements, 2010).
Previously, it was shown under simulated intestinal conditions that the initial charge
of the interface did not specifically correlate with the lag phase of pancreatic lipase
(Wickham et al., 1998). In the case of protein-stabilized emulsions, the FFA release under
simulated intestinal digestion was only slightly higher for positively charged lactoferrin-
stabilized emulsions compared to negatively charged β-lactoglobulin-stabilized emulsions
(Sarkar et al., 2010). The opposite was found when the intestinal digestion of lysozyme-
and WPI-stabilized emulsions, as used in Chapter 3, was investigated using the pH-stat
method (Chapter 2). For WPI-stabilized emulsions 60 % of initial FFA present in the form
of triglycerides were released after 30 min, whereas for lysozyme only 0.5 % of FFA were
determined. As shown in Fig. 6.1, WPI-stabilized emulsions were homogenous, whereas
lysozyme-stabilized emulsions flocculated immediately upon mixing with bile salts. This
behaviour caused the difference in FFA release, which in turn could lead to wrong
interpretation, as shown in Chapter 3 and discussed further in 6.1.3.
A BA B
Fig. 6.1 Simulated intestinal digestion of WPI-stabilized emulsions (A) and lysozyme-stabilized emulsions
(B) using the pH-stat method at pH7.5, as described in Chapter 2.

Chapter 6
128
6.1.2 Effect of simulated digestion on lipid digestion - pH-stat
method
The question remains if in vitro models can predict the behavior and therefore the
release of FFA as occurring in humans. The pH-stat method is an easy and fast single-step
model (McClements, 2010) to investigate underlying mechanisms of intestinal emulsion
digestion, such as compositional changes, at the interface. When using this model, it was
shown (Chapter 2) that the FFA release is strongly dependent on the emulsifier used, i.e. a
discrepancy of 60% was found only for WPI-stabilized emulsions between pH-stat and gas
chromatography. As mentioned in Chapter 2, specific conclusions on lipid digestion were
related to the in vitro experimental conditions used during the study. For example, the
FFA detection with pH-stat is very much dependent on the presence of calcium, which is
not advantageous for determination of overall lipid digestion and needs to be taken into
account when comparing experimental results. Nevertheless, the detectable FFA in the
pH-stat as affected by the calcium concentration could still be of benefit. For example, it
could provide knowledge on the available FFA which are in turn detectable at the
intestinal site, when emulsions are present in the form of milk products containing high
calcium concentrations.
Several hypotheses have been discussed (i.e. role of the digestion medium, binding of
FFA to β-lactoglobulin, interaction of FFA with positively amino acid residues of WPI,
hydrolysis of protein by proteolytic enzymes present in pancreatin) dealing with the
question why the detection of FFA with pH-stat was different between the emulsifiers. No
conclusive statement could be made, showing that further research in this area needs to
be undertaken. Indeed, as shown recently, when β-lactoglobulin-stabilized emulsions
were digested under simulated intestinal conditions and β-lactoglobulin was present in
the bulk in excess a higher lipid digestion was shown with pH-stat, but not with HPLC
compared to emulsions in which β-lactoglobulin completely covered the interface (Marze
et al., 2012). In that study, the presence of vesicles and/or the accumulation of digestion
products at the interface have been suggested to be responsible, but this hypothesis has
not been confirmed yet.
Overall, the pH-stat model can be used to investigate the FFA release. However, the
results should be interpreted with care and we suggested to combining it with other
analytical tools, such as gas chromatography (Chapter 2). Also, in static in vitro models,

General Discussion
129
the digestion of emulsions and their behavior should be analyzed under different
conditions such as varying calcium concentrations.
6.1.3 Effect of simulated digestion on lipid digestion - Including the
stomach in in vitro models
In Chapters 3-5 it was shown that the behaviour of the emulsions was influenced by
the gastric passage. By simulating the gastric and intestinal passage of protein stabilized
emulsions through a dynamic model, a clear influence of gradual acidification and the
shape of the gastric compartment was found (Chapter 3). Both led to creaming of the
emulsions when the emulsifier, i.e. WPI, transitioned through the isoelectric point of the
protein. This in turn caused a delay in intestinal lipid digestion. Nevertheless, the overall
concentration of FFA at the end of the experiment was similar compared to emulsions
stabilized by lysozyme. Thus, transition through the isoelectric point affects the creaming
behavior, delays intestinal lipid digestion, but does not influence the overall pancreatic
lipase activity. Using un-homogenized emulsions, which represents phase separation of
oil and water (Chapter 4), a delayed entry of lipids in the duodenum was clearly observed
compared to a homogenized emulsion. Both creaming and phase separation delayed the
intestinal lipolysis. Nevertheless, in the model used in Chapter 3, the change in emulsion
structure did not influence the emptying of the emulsion from the gastric compartment as
this was computer controlled.
Furthermore, the model did not include the presence of mucin, which might change
emulsion behaviour as well. For example, the absorption of bile salts to the interface
enhanced the diffusion of lipid droplets in the intestinal mucin (Macierzanka et al., 2011)
and this is pronounced with increasing negative surface charge (Macierzanka et al., 2012).
During the human study (Chapters 4 and 5), gastric emptying was estimated based on
plasma paracetamol concentration. However, this is solely a marker for emptying of the
water phase. Moreover, we assumed layering of the oil phase for the un-homogenized
emulsion based on studies using MRI (Marciani et al., 2007a; Marciani et al., 2007b;
Marciani et al., 2009) or scintigraphy (Edelbroek et al., 1992).

Chapter 6
130
In order to further explore the layering of the oil-phase and gastric emptying in a
gastric environment a pig study was conducted. The experimental setup and the results of
this study are given below.
Experimental set up Two male pigs (TOPIGS20, Van Beek SPF Lelystad, The Netherlands),
provided with a gastric canula at the bottom of the stomach were used. The investigated emulsions
were prepared as described in Chapter 3. Lipids were analyzed according to the method described in
Chapter 3 and emptying of the water phase was determined by analyzing paracetamol
concentration in gastric samples using UPLC-MS/MS (TNO, Zeist, The Netherlands). In total, 1 L of
the respective emulsion was introduced in the stomach via the gastric canula and contained 1.2 g
paracetamol. WPI and Tween-80 stabilized emulsions were followed over time, i.e. at t=30, 90 and
180 minutes on three separate occasions. Lysozyme-stabilized emulsion and an un-homogenized
Tween-80 sample were studied after 90 minutes residence in the pig stomach. To study the local
behavior of emulsions at different time points the complete gastric content was collected in
fractions of 100 mL. The first fraction represents the bottom part of the stomach. The last fraction
represents the top part.
The pig is similar to the human with respect to anatomy and physiology of the
digestive tract (Pond & Houpt, 1978). Therefore, extracts of gastro-intestinal juices
obtained from pigs are frequently used in in vitro models (for a review see McClements &
Li, 2010) to investigate the digestion of emulsions. Nevertheless, it is still challenging to
translate the dynamics of the in vivo digestion, such as dynamic secretion of digestive
fluids and gastric emptying, into in vitro models. We hypothesized that results obtained
from pigs on emulsion behavior will confirm observations as found in the dynamic model.
In turn, they will support the need of advanced (i.e. complex) in vitro models to predict
emulsions behaviour under mixing and digestive processes as occurring in vivo. Examples
are given below to discuss the effects of intragastric distribution of lipids as determined
by the emulsifier or the droplets size on gastric emptying in pig.

General Discussion
131
Protein stabilized emulsions - influence of mucin?
In Chapter 3 the importance of a gradual acidification in the gastric compartment on
the behavior of protein stabilized emulsions, i.e. WPI and lysozyme, was shown. In
combination with the custom designed gastric compartment, which simulated the motility
of the different regions of the stomach, it was possible to follow separation of WPI-
stabilized emulsions into a cream and serum layer, but not the gastric emptying. In fact,
when monitoring the behavior of WPI-stabilized emulsions in the pig, it can be seen that
the gastric volume after application of the WPI-stabilized emulsion decreased rapidly (Fig.
6.2A). Similarly as in the dynamic model (Chapter 3), the WPI-stabilized emulsion started
to cream in the pig stomach at t=30 minutes, most likely as a result of the gradual
acidification. When comparing WPI-stabilized emulsions with lysozyme, both emulsions
creamed at 90 minutes after the start of the experiment as can be seen from the
intragastric distribution of lipids (Fig. 6.3A). The fact that lysozyme-stabilized emulsions
creamed to the same extent as WPI-stabilized emulsions (Fig 6.3A) is possibly related to
the presence of mucin. As discussed in Chapter 3, slight flocculation of lysozyme-
stabilized emulsions occurred upon addition of pepsin and salt. As mucin, similar as
pepsin is negatively charged above pH 3 (Froehlich et al., 1995), it is likely to interact with
the lysozyme stabilized interface resulting in flocculation. Still, the water phase emptied
approximately 1.5 times faster in the case of WPI-stabilized emulsion compared to
lysozyme-stabilized emulsions. However, the interaction of mucin with emulsions is
barely investigated. One might speculate that such investigations are of importance,
especially as protein stabilized interfaces or charged emulsifiers interact with mucin. This
has been proven upon saliva mixing (Silletti et al., 2007). In the pig model, we investigated
the distribution of mucin by collecting several fractions of emulsion-gastric fluid samples
as described above. The mucin concentration was determined as the sum of the
constituent galactosamine and glucosamine contents by the analysis of sugar composition
(Verhoef et al., 2005). The results revealed an equal distribution of mucin over the
fractions at a concentration of 26±8 mg/mL. Mucin was shown to promote the
flocculation of β-lactoglobulin-stabilized emulsions (Sarkar et al., 2010) and to decrease
the interfacial tension (Marze et al., 2013). It is not known yet to what extent the presence
of mucin influences the FFA release from emulsions or the FFA sensing by receptors at
the intestinal site. This clearly shows the effect of inducing complexity into models on
emulsions behavior. Hence, digestion of lipids needs to be evaluated by using in vivo
studies and adapted in dynamic models.

Chapter 6
132
Fig. 6.2 Example of change in gastric volume (A) over time after administration of WPI (▲) or Tween 80-
(■) stabilized emulsions for one pig. In B and C examples of emptying of lipid (closed symbols) and water
(open symbols) phase is given. It is determined from the initial amount of lipids present in the emulsion
(i.e. 8% (v/v) sunflower oil) or paracetamol in the water phase (i.e. 1.2 g), respectively of the WPI or
Tween 80-stabilized emulsions for one pig.
Fig. 6.3 Distribution of lipids in the stomach as determined from fractionated samples at 90 minutes after
administration of the emulsions. The first fraction represents the lowest part of the stomach, i.e. bottom.
The last fraction represents the highest part, i.e. top. In (A) an example of lipid distribution in the
stomach after administration of WPI and Lysozyme (Lys)-stabilized emulsions and in (B) of a
homogenized (h) and un-homogenized (uh) Tween 80-stabilized emulsion is given.

General Discussion
133
The distribution of fat in the stomach – homogenized and un-homogenized Tween
80-stabilized emulsions and creaming
In the human study, two emulsion systems were used to simulate both a homogenous
system as well as a phase separated system of oil and water (Chapters 4 and 5). By
measuring the plasma paracetamol concentrations, it was shown that the emptying of the
homogenized Tween 80-stabilized emulsion was slower compared to the un-homogenized
emulsion. This finding was also confirmed in the pig model. The gastric volume for the
treatment with homogenized Tween 80-stabilized emulsions was constant over time (Fig
6.2A). One must note that the lipid and water phase continued to empty from the stomach
(Fig 6.2C), which was possibly caused by gastric secretion exceeding gastric emptying.
Compared to the homogenized emulsion, the gastric volume was 1.6 times less for the un-
homogenized emulsion after 90 minutes of the start of experiment. As expected, the lipids
of un-homogenized emulsions layered on top of the stomach (Fig 6.3B). Although Tween-
stabilized emulsions are described as being stable, i.e. not aggregated or coalesced, in the
acidic environment of the stomach (Marciani et al., 2007a; Marciani et al., 2009), we
observed high concentrations of triglyceride at 120 minutes (Chapter 4). This was also
found using a dynamic model as described in Chapter 3 (Oosterveld et al., unpublished
data). This was interpreted as creaming of the homogenized emulsion. In agreement with
this, fractions of Tween 80-stabilized emulsions revealed creaming at 90 minutes in the
pig stomach. Therefore, the results of stable emulsions observed in vitro should be
generally interpreted with care in the view of the effect of creaming in vivo on, for
example, metabolic responses.
The presence of gastric lipase
Human gastric lipase is able to digest up to 40 % (Armand, 2007) or 25 % (Carriere et
al., 1993) of lipid digestion and also helps to accelerate pancreatic lipase activity.
Furthermore, as recently shown, the addition of a fungal lipase in simulated gastric
conditions enhanced flocculation of WPI-stabilized emulsions (Golding et al., 2011) gastric
environment. Next to the ethical consideration, a drawback in using the pig as a model is,
that porcine gastric lipase is irreversibly inactivated at pH 4 (Moreau et al., 1988).
Therefore, besides the information that can be gained on emulsion behavior, such as
creaming or gastric emptying, it is not advantageous to study lipolysis of emulsions in a
pig stomach. Several options are commercially available to be used as replacers for human
gastric lipase in vitro. In the TIM (Chapter 3), a preduodenal lamb lipase was used as it is

Chapter 6
134
similar to human gastric lipase in its pH profile (Moreau et al., 1988) and its activity
towards the sn-1 and sn-3 position of a triglyceride. Recently, a 3.5 fold higher hydrolysis
level (defined as L1% as shown in Chapters 3 and 4) was observed for a gastric lipase
extracted from rabbits compared to human gastric lipase (Capolino et al., 2011). This
rabbit lipase was, therefore, considered as a good alternative to be used in in vitro assays.
Overall, dynamic models and the pig model revealed the importance of investigating
the intragastric lipolysis and the change in emulsions structure, i.e. flocculation, as well as
the creaming or phase separation behavior of emulsions in vitro. Furthermore, the effect
of this behaviour on intestinal lipolysis needs to be investigated in vitro to eventually
predict possible satiety perceptions, but also metabolic responses in vivo.
6.2 Influence of emulsion parameter on satiety
6.2.1 The role of lipid digestion products FFA, MG, DG and TG
In vitro studies clearly miss the ability to mimic complex physiological responses as,
for example, induced by hormones. In case of lipid digestion, FFA are considered as the
strongest trigger to induce these hormonal responses (McLaughlin et al., 1999; Feltrin et
al., 2007). From the present study the effect of a single lipid digestion product cannot be
related to hormonal responses. As shown, the duodenal aspirates consisted of
approximately 16 mM FFA at t=15 and 36 mM FFA at t=30 minutes. Similar duodenal FFA
concentrations were found after ingestion of TG containing meals (Borgström et al., 1957)
or after ingestion of emulsions (Armand et al., 1996; Armand et al., 1999). For example,
FFA concentrations ranged for a fine emulsion with a mean droplet size of 0.7 µm
between 18-30 mM and for a coarse emulsion with a mean droplet diameter of 10 µm,
between 7-22 mM (Armand et al., 1999). These data were based on emptying rates as
determined from a water soluble marker. It has been shown that concentrations of FFA
between 40 and 72 mM stimulated the release of CCK and PYY and suppressed energy
intake (Feltrin et al., 2007). Unfortunately, the gastric emptying measured by means of
plasma paracetamol reflect only the emptying of the water phase and not necessarily the
oil phase. The emptying rate of nutrients to the duodenum is approximately 2-3 kcal/min
(Horowitz et al., 1993). This corresponded well with Tween-stabilized emulsions, which
had a similar TG content (i.e. 50 g oil; Marciani et al., 2009) as used in our study.
Nevertheless, it is clear that in our study the initially emptying of the un-homogenized

General Discussion
135
was faster compared to the homogenized emulsion. Also in the study of Marciani et al., (
2009) the half time emptying of the emulsion, which was homogenous in the gastric
environment, was determined to be at approximately 185 min and for the emulsion,
which phase separated at approx. 70 min. Thus, the gastric volume decreased more
rapidly for the phase separated emulsion (Marciani et al., 2009). Despite these differences
observed in this mentioned study, only modest variation in satiety perception were found,
but unfortunately the CCK responses were not determined. Overall, we failed to induce
marked differences in satiety perception despite the huge differences in emulsion
behavior. This shows that other mechanisms are involved in satiety responses. Also, it
could point to the influence of gastric lipolysis, which was in our study approx. 3%, as
intragastric applied FFA were about 5 times more effective to induce satiety related
responses than TG (Little et al., 2007).
In Chapter 2 it was mentioned that analysis of not only FFA, but also MG and DG
would add to better understanding of the digestion process in vivo. Although it is known,
that FFA when applied as such, have more potential to induce hormonal responses
compared to TG (Little et al., 2007) the extent in which DG, as formed in the stomach as
well as intraduodenal and MG, as formed during duodenal lipolysis, contributed to
hormonal responses is not clear yet.
6.2.2 Hormones CCK, GLP-1, PYY and ghrelin
Based on literature, the release of CCK is considered as an important factor to mediate
satiety responses upon digestion of fat. CCK release is provoked by the detection of FFA
at the duodenal site (Schwizer et al., 1997; McLaughlin et al., 1999; Feltrin et al., 2007;
Little et al., 2007). However, in Chapter 5 it was shown that no correlation was found
between CCK circulating plasma levels and FFA concentration in duodenal aspirates (Fig
5.4C), whereas the CCK release for the homogenized emulsion was significantly higher
than for the un-homogenized (Fig 5.2A). What is striking in Figure 5.4C, is the FFA
concentration at 4.4 mg/mL (i.e. 16 mM) and the corresponding maximum CCK plasma
concentration at 2.5 pmol/L. This data point corresponds to the time point at 15 minutes
for the homogenized emulsion when both the duodenal aspirate and the plasma were
collected. Since CCK is an early responding hormone, it is apparent that a certain
concentration of FFA has to enter the duodenum within these 15 minutes in order to
induce release of CCK. In fact, the same concentration of FFA in the aspirates was
observed, for example, after 60 minutes for the un-homogenized emulsion (Chapter 4, Fig.

Chapter 6
136
4.4B) without an elevation in plasma CCK concentration (Chapter 5, Fig. 5.2A). This is in
line with results of other studies (Fried et al., 1988; Foltz et al., 2009), but in contrast to a
previous duodenal intubation study, where emulsions infused at constant rates and
caused elevated CCK plasma levels over 2h (Seimon et al., 2009). As discussed in Chapter
5 and suggested before (Liddle, 1995), bile secretion led to removal of the CCK stimulating
agent, i.e. FFA. Whether this removal is caused by simple incorporation of these fatty
acids into micelles or by removal of such complexes through the mucus layer at the
intestinal site is not clear yet. With the analytical method (i.e. GC) used in this project,
incorporated FFA in micelles will be measured as free fatty acids. This could in turn lead
to an overestimation of the fatty acids which are available to stimulate the release of
CCK.
Furthermore, based on our results, we hypothesized that hormones alone are not a
predictor per se for satiety responses of foods (Chapter 5). Nevertheless, a relation was
found between CCK and gallbladder volume. Also, the hormones GLP-1 and PYY
responded differently to the emulsions (Chapter 5). In our study the suppression of
ghrelin, however, was not different between the treatments. Intraduodenal infusion of
protein-enriched meals suppressed ghrelin release after 60 min of protein load of 3
kcal/min compared to saline infusion (Ryan et al., 2012). Recently, it was shown in lean
men that ghrelin concentrations was suppressed during intraduodenal infusion of test
meals varying in macronutrients in the following order high protein>high fat>high
carbohydrate (Brennan et al., 2012). In that study, the high protein meal was associated
with the lowest ad libitum intake. Hence, as indicated previously (Keogh et al., 2011) and
as discussed in Chapter 5, the ghrelin response in our study could be relevant for the
similar satiety perceptions and the ad libitum.
6.3 Concluding remarks
Structuring of the lipid phase is considered to be a tool to influence satiety. Recently, it
was shown that changes in lipid structuring also results in changes of plasma lipids
(Keogh et al., 2011). In this study, for example, hydrogenated fat as part of the oil phase
did not have a beneficial effect on satiety compared to emulsion including only the oil.
Nevertheless this structuring caused lower TG-plasma levels (Keogh et al., 2011). Also,
phase separation of emulsions was shown to influence the postprandial lipemia and β-
oxidation of lipids in humans (Vors et al., 2013). In that study, for example, chylomicron

General Discussion
137
plasma concentrations were delayed and chylomicrons were smaller in size in normal and
also obese people. Thus, the way fat is structured may impact not only satiety, but also
plasma lipid profiles that play a role in the metabolic syndrome, in diabetes and in
obesity.
Lipids in the form of an emulsion are a relevant model to study satiety perception.
Emulsified lipids are present in a wide range of processed foods. However, it is difficult to
assess the impact of lipids or lipids structure in relation to other meal components like
carbohydrates and proteins. Proteins, for example, have a high satiating effect. Also they
could help inducing precipitation as shown for milk (van Aken et al., 2011) and with that
changing the behavior of emulsified lipids. The dynamic model (see Chapter 3) would be a
relevant model to investigate such structures.
It is possible, as shown in literature and in the human study presented in this thesis, to
induce satiety perceptions in humans by various emulsions system. Possibly a
combination of several aspects, such as emulsions droplets size, emulsifier and the
structure of fat and especially their resulting behavior in the stomach play a major role.
Also, which model to use clearly depends on the research question. For example, the role
of emulsion stability can be investigated with simple model systems. However, the final
link with satiety can only be made with in vivo studies.

Chapter 6
138
References
Armand, M., Borel, P., Pasquier, B., Dubois, C., Senft, M., Andre, M., Peyrot, J., Salducci, J. & Lairon, D. (1996). Physicochemical characteristics of emulsions during fat digestion in human stomach and duodenum. American Journal of Physiology - Gastrointestinal and Liver Physiology, 271 (34)(1), G172-183.
Armand, M., Pasquier, B., Andre, M., Borel, P., Senft, M., Peyrot, J., Salducci, J., Portugal, H., Jaussan, V. & Lairon, D. (1999). Digestion and absorption of 2 fat emulsions with different droplet sizes in the human digestive tract. American Journal of Clinical Nutrition, 70(6), 1096-1106.
Armand, M. (2007). Lipases and lipolysis in the human digestive tract: Where do we stand? Current Opinion in Clinical Nutrition and Metabolic Care, 10(2), 156-164.
Borgström, B., Dahlqvist, A., Lundh, G. & Sjövall, J. (1957). Studies of intestinal digestion and absorption in the human1. The Journal of Clinical Investigation, 36(10), 1521-1536.
Brennan, I. M., Luscombe-Marsh, N. D., Seimon, R. V., Otto, B., Horowitz, M., Wishart, J. M. & Feinle-Bisset, C. (2012). Effects of fat, protein, and carbohydrate and protein load on appetite, plasma cholecystokinin, peptide YY, and ghrelin, and energy intake in lean and obese men. American Journal of Physiology - Gastrointestinal and Liver Physiology, 303(1), G129-G140.
Capolino, P., Guérin, C., Paume, J., Giallo, J., Ballester, J. M., Cavalier, J. F. & Carrière, F. (2011). In vitro gastrointestinal lipolysis: Replacement of human digestive lipases by a combination of rabbit gastric and porcine pancreatic extracts. Food Digestion, 2(1-3), 43-51.
Carriere, F., Moreau, H., Raphel, V., Laugier, R., Benicourt, C., Junien, J. L. & Verger, R. (1991). Purification and biochemical characterization of dog gastric lipase. European Journal of Biochemistry, 202(1), 75-83.
Carriere, F., Barrowman, J. A., Verger, R. & Laugier, R. (1993). Secretion and contribution to lipolysis of gastric and pancreatic lipases during a test meal in humans. Gastroenterology, 105(3), 876-888.
Edelbroek, M., Horowitz, M., Maddox, A. & Bellen, J. (1992). Gastric emptying and intragastric distribution of oil in the presence of a liquid or a solid meal. Journal of Nuclear Medicine, 33(7), 1283-1290.
Feltrin, K. L., Little, T. J., Meyer, J. H., Horowitz, M., Rades, T., Wishart, J. & Feinle-bisset, C. (2007). Effects of lauric acid on upper gut motility, plasma cholecystokinin and peptide YY, and energy intake are load, but not concentration, dependent in humans. Journal of Physiology, 581(2), 767-777.
Foltz, M., Maljaars, J., Schuring, E. A. H., Van Der Wal, R. J. P., Boer, T., Duchateau, G. S. M., Peters, H. P. F., Stellaard, F. & Masclee, A. A. (2009). Intragastric layering of lipids delays lipid absorption and increases plasma CCK but has minor effects on gastric emptying and appetite. American Journal of Physiology - Gastrointestinal and Liver Physiology, 296(5), G982-991.
Fried, M., Mayer, E. A., Jansen, J. B. M. J., Lamers, C. B. H. W., Taylor, I. L., Bloom, S. R. & Meyer, J. H. (1988). Temporal relationships of cholecystokinin release, pancreatobiliary secretion, and gastric emptying of a mixed meal. Gastroenterology, 95(5), 1344-1350.
Froehlich, F., Gonvers, J. J. & Fried, M. (1995). Role of nutrient fat and cholecystokinin in regulation of gallbladder emptying in man. Digestive Diseases and Sciences, 40(3), 529-533.
Golding, M., Wooster, T. J., Day, L., Xu, M., Lundin, L., Keogh, J. & Clifton, P. (2011). Impact of gastric structuring on the lipolysis of emulsified lipids. Soft Matter, 7(7), 3513-3523.
Horowitz, M., Edelbroek, M. A. L., Wishart, J. M. & Straathof, J. W. (1993). Relationship between oral glucose tolerance and gastric emptying in normal healthy subjects. Diabetologia, 36(9), 857-862.
Hu, M., Li, Y., Decker, E. A., Xiao, H. & McClements, D. J. (2010). Influence of tripolyphosphate cross-linking on the physical stability and lipase digestibility of chitosan-coated lipid droplets. Journal of Agricultural and Food Chemistry, 58(2), 1283-1289.
Keogh, J. B., Wooster, T. J., Golding, M., Day, L., Otto, B. & Clifton, P. M. (2011). Slowly and rapidly digested fat emulsions are equally satiating but their triglycerides are differentially absorbed and metabolized in humans. Journal of Nutrition, 141(5), 809-815.
Li, Y. & McClements, D. J. (2010). New mathematical model for interpreting ph-stat digestion profiles: Impact of lipid droplet characteristics on in vitro digestibility. Journal of Agricultural and Food Chemistry, 58(13), 8085-8092.
Liddle, R. A. (1995). Regulation of cholecystokinin secretion by intraluminal releasing factors. American Journal of Physiology - Gastrointestinal and Liver Physiology, 269(3 32-3), G319-G327.
Little, T. J., Russo, A., Meyer, J. H., Horowitz, M., Smyth, D. R., Bellon, M., Wishart, J. M., Jones, K. L. & Feinle-Bisset, C. (2007). Free fatty acids have more potent effects on gastric emptying, gut hormones, and appetite than triacylglycerides. Gastroenterology, 133(4), 1124-1131.

General Discussion
139
MacIerzanka, A., Rigby, N. M., Corfield, A. P., Wellner, N., Böttger, F., Mills, E. N. C. & MacKie, A. R. (2011). Adsorption of bile salts to particles allows penetration of intestinal mucus. Soft Matter, 7(18), 8077-8084.
MacIerzanka, A., Böttger, F., Rigby, N. M., Lille, M., Poutanen, K., Mills, E. N. C. & MacKie, A. R. (2012). Enzymatically structured emulsions in simulated gastrointestinal environment: Impact on interfacial proteolysis and diffusion in intestinal mucus. Langmuir, 28(50), 17349-17362.
Marciani, L., Wickham, M., Singh, G., Bush, D., Pick, B., Cox, E., Fillery-Travis, A., Faulks, R., Marsden, C., Gowland, P. A. & Spiller, R. C. (2007a). Enhancement of intragastric acid stability of a fat emulsion meal delays gastric emptying and increases cholecystokinin release and gallbladder contraction. American Journal of Physiology - Gastrointestinal and Liver Physiology, 292(6), G1607-G1613.
Marciani, L., Wickham, M. S. J., Bush, D., Faulks, R., Wright, J., Fillery-Travis, A. J., Spiller, R. C. & Gowland, P. A. (2007b). Magnetic resonance imaging of the behaviour of oil-in-water emulsions in the gastric lumen of man. British Journal of Nutrition, 95(02), 331-339.
Marciani, L., Faulks, R., Wickham, M. S. J., Bush, D., Pick, B., Wright, J., Cox, E. F., Fillery-Travis, A., Gowland, P. A. & Spiller, R. C. (2009). Effect of intragastric acid stability of fat emulsions on gastric emptying, plasma lipid profile and postprandial satiety. British Journal of Nutrition, 101(6), 919-928.
Marze, S. & Choimet, M. (2012). In vitro digestion of emulsions: Mechanistic and experimental models. Soft Matter, 8(42), 10982-10993.
Marze, S., Meynier, A. & Anton, M. (2013). In vitro digestion of fish oils rich in n-3 polyunsaturated fatty acids studied in emulsion and at the oil-water interface. Food and Function, 4(2), 231-239.
McClements, D. J. & Li, Y. (2010). Review of in vitro digestion models for rapid screening of emulsion-based systems. Food and Function, 1(1), 32-59.
McLaughlin, J., Luca, M. G., Jones, M. N., D'Amato, M., Dockray, G. J. & Thompson, D. G. (1999). Fatty acid chain length determines cholecystokinin secretion and effect on human gastric motility. Gastroenterology, 116(1), 46-53.
Moreau, H., Gargouri, Y., Lecat, D., Junien, J. L. & Verger, R. (1988). Screening of preduodenal lipases in several mammals. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism, 959(3), 247-252.
Mun, S., Decker, E. A. & McClements, D. J. (2007). Influence of emulsifier type on in vitro digestibility of lipid droplets by pancreatic lipase. Food Research International, 40(6), 770-781.
Pond, W. G. & Houpt, K. A. (1978). The biology of the pig. Ithaca, NY, USA: Cornell University Press. Rodriguez, J. A., Mendoza, L. D., Pezzotti, F., Vanthuyne, N., Leclaire, J., Verger, R., Buono, G., Carriere, F. &
Fotiadu, F. (2008). Novel chromatographic resolution of chiral diacylglycerols and analysis of the stereoselective hydrolysis of triacylglycerols by lipases. Analytical Biochemistry, 375(2), 196-208.
Ryan, A. T., Feinle-Bisset, C., Kallas, A., Wishart, J. M., Clifton, P. M., Horowitz, M. & Luscombe-Marsh, N. D. (2012). Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. American Journal of Clinical Nutrition, 96(3), 474-482.
Sarkar, A., Goh, K. K. T. & Singh, H. (2010). Properties of oil-in-water emulsions stabilized by β-lactoglobulin in simulated gastric fluid as influenced by ionic strength and presence of mucin. Food Hydrocolloids, 24(5), 534-541.
Sarkar, A., Horne, D. S. & Singh, H. (2010). Pancreatin-induced coalescence of oil-in-water emulsions in an in vitro duodenal model. International Dairy Journal, 20(9), 589-597.
Schwizer, W., Asal, K., Kreiss, C., Mettraux, C., Borovicka, J., Remy, B., Guzelhan, C., Hartmann, D. & Fried, M. (1997). Role of lipase in the regulation of upper gastrointestinal function in humans. American Journal of Physiology - Gastrointestinal and Liver Physiology, 273(3 36-3), G612-G620.
Seimon, R. V., Wooster, T., Otto, B., Golding, M., Day, L., Little, T. J., Horowitz, M., Clifton, P. M. & Feinle-Bisset, C. (2009). The droplet size of intraduodenal fat emulsions influences antropyloroduodenal motility, hormone release, and appetite in healthy males. The American Journal of Clinical Nutrition, 89(6), 1729-1736.
Silletti, E., Vingerhoeds, M. H., Norde, W. & van Aken, G. A. (2007). The role of electrostatics in saliva-induced emulsion flocculation. Food Hydrocolloids, 21(4), 596-606.
Van Aken, G. A., Bomhof, E., Zoet, F. D., Verbeek, M. & Oosterveld, A. (2011). Differences in in vitro gastric behaviour between homogenized milk and emulsions stabilised by Tween 80, whey protein, or whey protein and caseinate. Food Hydrocolloids, 25(4), 781-788.
Verhoef, R., Schols, H. A., Blanco, A., Siika-Aho, M., Rättö, M., Buchert, J., Lenon, G. & Voragen, A. G. J. (2005). Sugar composition and FT-IR analysis of exopolysaccharides produced by microbial isolates from paper mill slime deposits. Biotechnology and Bioengineering, 91(1), 91-105.

Chapter 6
140
Vors, C., Pineau, G., Gabert, L., Drai, J., Louche-Peĺissier, C., Defoort, C., Lairon, D., Deśage, M., Danthine, S., Lambert-Porcheron, S., Vidal, H., Laville, M. & Michalski, M. C. (2013). Modulating absorption and postprandial handling of dietary fatty acids by structuring fat in the meal: A randomized crossover clinical trial. American Journal of Clinical Nutrition, 97(1), 23-36.
Wickham, M., Garrood, M., Leney, J., Wilson, P. D. G. & Fillery-Travis, A. (1998). Modification of a phospholipid stabilized emulsion interface by bile salt: Effect on pancreatic lipase activity. Journal of Lipid Research, 39(3), 623-632.

Summary
Samenvatting
Zusammenfassung

Summary
142
Lipids are key components in our diet and, specifically in the western diet, commonly
eaten in the form of emulsions present in highly processed food. In view of the current
obesity prevalence one strategy to prevent overconsumption of food could be the use of
specifically designed emulsions to induce satiation and satiety.
In Chapter 1 we introduce the regulation of satiety responses, which occur upon
intake of lipids and their digestion. The focus is on the regulation of gastric emptying and
the hormones involved as observed by several in vivo studies. It shows the current
understanding that lipolytic products, i.e. free fatty acids (FFA), are most likely
responsible for lipid induced release of hormones, such as CCK, and the resulting satiety
responses when detected at the intestinal level. Nevertheless, it also shows that this
knowledge is gained mostly upon intestinal intubation studies and little research involves
the gastric passage. Including this passage has shown the importance of aspects like
gastric lipolysis or colloidal state of lipids in the regulation of satiety responses. However,
these aspects are not investigated in combination or show inconsistent results. A similar
situation is found for in vitro models that have recently been used to gain mechanistic
understanding of the behaviour of emulsions. At the start of this project the focus in
literature was on the intestinal digestion of lipids and mainly on the release of FFA but
not on monoglycerides (MG) or diglycerides (DG). In addition, only a few publications
included the gastric digestion, but the specific shape or functions of the stomach are
neglected or mimicked simply.
The study in Chapter 2 focused on FFA release in relation to emulsion properties
monitored by a commonly used static in vitro model, i.e. the pH-stat method. Emulsions
differing in droplets size, i.e. fine, medium and coarse, stabilized by whey protein isolate
(WPI) or gum arabic (GA) were digested under intestinal conditions. Gas chromatography
(GC) was used to determine lipolysis products, i.e. FFA, MG, DG and triglycerides (TG).
Overall, GC analysis revealed decreasing FFA, MG and DG concentrations with increasing
droplet size. It was shown that digestion of emulsions stabilized by a protein-
polysaccharide emulsifier (i.e. GA) resulted in higher FFA release than of emulsions
stabilized with a proteinaceous emulsifier (i.e. WPI). In addition, GA-stabilized emulsions
resulted in a faster hydrolysis rate of DG into MG compared to WPI-stabilized emulsions.
The concentrations of FFA determined with GC were 2–3 times higher compared with
pH-stat for WPI stabilized emulsions. It was concluded that since the detection of FFA
with pH-stat was influenced by the type of emulsifier, pH-stat results should be
interpreted with care and combined with other methods, such as GC.

143
In Chapter 3 the effect of gastric passage of protein stabilized emulsions on lipolysis
influenced by both gradual acidification and stomach properties was studied. For this
purpose, a dynamic in vitro model (TNO Gastro-Intestinal Model) was introduced, which
had a custom designed gastric compartment. To determine the effect of flocculation at the
isoelectric point WPI or lysozyme were used as emulsifiers. Microscopy, particle size
analysis and GC were used for characterization. Under gastric conditions WPI-stabilized
emulsions separated into a cream and serum layer when the protein transitioned through
its isoelectric point. Microscopy and particle size analysis revealed larger flocculated
particles for the initial negatively charged WPI-stabilized emulsions compared to the
positively charged lysozyme-stabilized emulsions. Mainly FFA and DG were present in
the gastric compartment, which is typical for the hydrolysis of gastric lipase. In addition,
analysis of the serum layer revealed the presence of FFA, which could induce the release
of CCK in vivo. The creaming caused a delayed entry of lipids into the small intestinal
part. Nevertheless, the total FFA concentration in the gastric compartment at the end of
the experiment was similar for both emulsions. The effect of creaming or presence of FFA
in the serum layer on gastric emptying, however, could not be assessed due to the nature
of the model.
In a human study, described in Chapters 4 and 5, we investigated several aspects of
lipid administration, lipid digestion and the corresponding physiological responses
influenced by the physical properties of the lipids. In Chapter 4, we focused on the
influence of the intragastric distribution of lipids on gastric and duodenal lipolysis. Also,
the additive effect of oral application on gastric lipolysis was studied. In a randomized,
single blind crossover study, 15 healthy subjects were intubated nasogastric on three
separate occasions. In addition, a subset of 6 healthy subjects was intubated nasoduodenal
on two separate occasions. Volunteers received a homogenized emulsion orally or by
gastric infusion or a gastric infusion of water followed by the oil phase (un-homogenized
sample). The subset of 6 volunteers received a gastric infusion of a homogenized emulsion
or a gastric infusion of an un-homogenized sample. Emulsion structure, pH and lipid
composition were assessed in both gastric and duodenal fluids. A gastric lipolysis of
approx. 3% for all treatments was found and the main products were FFA and DG.
Concentrations of FFA in duodenal aspirates were higher after homogenized emulsions
than after un-homogenized samples (p<0.05). Gastric emptying was faster for the un-
homogenized sample than for the homogenized emulsion as assessed by plasma
paracetamol concentrations. Overall, there was no effect observed of oral processing.
Also, homogenization had no effect on gastric lipolysis. Nevertheless, differences in

Summary
144
inhibition of gastric emptying were observed: Duodenal release of FFA and inhibition of
gastric emptying occurred earlier for homogenized emulsion than for un-homogenized
samples.
Subsequently, in Chapter 5, it was investigated if such implemented effects would
influence responses relevant for lipid digestion and satiety perception. Thus, in the same
setting as described in Chapter 4, gallbladder volume, plasma CCK, PYY, GLP-1 and
ghrelin concentration, satiety ratings as assessed via a visual analogue scale, and ad
libitum intake were analyzed. The results showed a significant influence of the treatments
on gallbladder volume (p<0.001). In detail, the most prolonged contraction of the
gallbladder was found when the homogenized emulsion was applied orally. In contrast,
when the un-homogenized sample was administered intragastrically, the contraction of
the gallbladder was lowest. In addition, we found significant effects of time and treatment
on circulating hormone concentrations. In the case of plasma CCK, concentrations were
significantly higher at 15 minutes after application of the homogenized emulsion
compared to the application of the un-homogenized sample. For PYY, the concentrations
were significantly higher at 30 minutes for the homogenized emulsion compared to the
un-homogenized sample. The responses of PYY and GLP-1 to the un-homogenized sample
were delayed by approximately 20 minutes compared to the responses to the
homogenized emulsion. On the suppression of ghrelin, however, only an effect of time,
but not of the treatments was found. Also, the perception of satiety and the ad libitum
intake were not different for the treatments. Nevertheless, the feeling `desire to eat´ was
stronger for the homogenized emulsions compared to the un-homogenized sample. It was
concluded, that plasma hormone concentrations and gallbladder motility were affected by
the intragastric distribution of the lipids. These effects were neither reflected in satiety
perceptions nor in the ad libitum intake. Hence, the release of gastrointestinal hormones
cannot directly be related to the satiating effect of food. When comparing the results of
FFA release as observed in Chapter 4, a correlation between duodenal concentration of
FFA and gallbladder volume was found, but not with CCK plasma concentration.
Therefore, it was speculated that hormonal induced satiety feelings might have been
overruled by other mechanisms, such as gastric distention.
In Chapter 6, the influence of emulsion properties and the in vitro digestion models
used, with attention to their complexity, on lipolysis were discussed. In addition, the
effect of gastric passage of emulsions with respect to the creaming behaviour was
discussed with findings from a study in a pig model. Also, the impact of lipid digestion
products on satiety and the influence of satiety hormones were discussed.

Summary
Samenvatting
Zusammenfassung

Samenvatting
146
In een westers voedingspatroon vormen vetten een belangrijk bestanddeel. Ze worden
voornamelijk geconsumeerd als emulsie; aanwezig in bewerkte voedingsmiddelen. Met
het oog op de toegenomen incidentie van obesitas, zou overconsumptie van voedsel
voorkomen kunnen worden door het gebruik van speciaal ontworpen emulsies, die het
verzadigingsgevoel bevorderen en daarmee de eetlust helpen controleren.
In hoofdstuk 1 introduceren we de regulatie van verzadigingsreacties, die optreden
na inname en vertering van vetten. De focus ligt op de regulering van de maaglediging en
de hormonen die hierbij betrokken zijn, zoals is waargenomen in verschillende in vivo
studies. Huidig inzicht toont aan dat afbraakproducten van vet, d.w.z. vrije vetzuren
(FFA), zeer waarschijnlijk verantwoordelijk zijn voor vet-geïnduceerde afgifte van
hormonen zoals CCK en voor de verzadigingsreacties die gedetecteerd worden in het
intestinale (darm) spijsverteringsstelsel. Niettemin, toont het ook aan dat deze kennis
vooral wordt opgedaan bij de intestinale intubatie studies en dat weinig onderzoek is
gedaan waarbij vertering in de maag is toegevoegd. Door het maag spijsverteringsstelsel
mee te nemen blijken aspecten zoals maag-lipolyse en de colloïdale toestand van de
vetten in de regulatie van verzadigingsreacties zeer belangrijk te zijn. De combinatie van
deze aspecten is echter nooit onderzocht of tonen inconsistente resultaten. Een
vergelijkbare situatie is waargenomen bij in vitro modellen die recentelijk zijn gebruikt
om het mechanistische gedrag van emulsies te begrijpen. Bij aanvang van dit project lag
de focus in de literatuur op de intestinale vertering van lipiden en vooral op het
vrijkomen van FFA, maar niet op monoglyceriden (MG) of diglyceriden (DG). Bovendien
waren er slechts een aantal publicaties waarbij de vertering in de maag werd
meegenomen, maar waarbij de specifieke vorm of functie van de maag werden
verwaarloosd of op een simpele manier nagebootst.
Hoofdstuk 2 is gericht op het vrijkomen van FFA in relatie tot de emulsie-
eigenschappen, door middel van een veelgebruikte statisch in vitro model, de pH-stat-
methode. Emulsies van verschillende druppelgrootte, (fijne, medium en grof),
gestabiliseerd door wei-eiwit isolaat (WPI) of arabische gom (GA) werden verteerd onder
intestinale condities. Gaschromatografie (GC) werd gebruikt om de afbraakproducten te
weten: FFA, MG, DG en triglyceriden (TG) te bepalen. Uiteindelijk, bleek met GC analyse
dat met toenemende druppelgrootte de concentraties van FFA, MG en DG af namen.
Aangetoond werd dat de afbraak van emulsies, gestabiliseerd door een eiwit-
polysaccharide emulgator (bijvoorbeeld GA), resulteerde in hogere FFA afgifte dan van
emulsies gestabiliseerd met een eiwit (bijvoorbeeld WPI). Bovendien, resulteerde GA-
gestabiliseerde emulsies in een snellere hydrolyse van DG naar MG in vergelijking met

147
WPI gestabiliseerde emulsies. De concentraties FFA, die bepaald zijn met GC, waren 2-3
maal hoger dan de concentraties bepaald met pH-stat voor WPI gestabiliseerde emulsies.
Doordat de bepaling van FFA met de pH-stat werd beïnvloed door het type emulgator,
moeten de resultaten zorgvuldig geïnterpreteerd worden en met andere methoden zoals
GC gecombineerd worden.
In hoofdstuk 3 wordt het effect bestudeerd van maag passage op eiwit gestabiliseerde
emulsies en hun lipolyse, beïnvloed door zowel geleidelijke verzuring en maag
eigenschappen. Hiervoor werd een dynamisch in vitro model (TNO gastrolintestinale
Model) geïntroduceerd met een speciaal ontworpen maag compartiment. Om het effect
van flocculatie op het isoelectrische punt te bepalen, werd WPI of lysozym gebruikt als
emulgator. Voor de karakterisering werden microscopie, deeltjesgrootte analyse en GC
gebruikt. Onder de in de maag aanwezige condities werden WPI-gestabiliseerde emulsies,
op het moment dat het eiwit langs haar iso-electrisch punt komt, gescheiden in een crème
en serum laag. Microscopie en deeltjesgrootte analyse laten grotere geflocculeerde
deeltjes zien voor de aanvankelijk negatief geladen WPI-gestabiliseerde emulsies in
vergelijking met de positief geladen lysozym-gestabiliseerde emulsies. Vooral FFA en DG
waren aanwezig in het maag compartiment, wat typisch is voor de hydrolyse van maag
lipase. Daarbij onthulde analyse van de serum laag de aanwezigheid van FFA, die
mogelijk de afgifte van CCK in vivo kan induceren. Het opromen veroorzaakt een
vertraagde toegang van vetten in de dunne darm. Niettemin, bleek de totale FFA
concentratie in het maag compartiment aan het einde van het experiment gelijk te zijn
voor beide emulsies. Het effect van opromen of de aanwezigheid van FFA in de serum
laag op maaglediging kan echter niet door het model worden bepaald.
In humane studies, beschreven in hoofdstuk 4 en 5, onderzochten we verschillende
aspecten van vetten zoals de toediening, de spijsvertering en de bijbehorende
fysiologische reacties die beïnvloed zijn door de fysische eigenschappen van de vetten. In
hoofdstuk 4 hebben we ons gericht op de invloed van de intragastrische distributie van
vetten op gastrische en duodenale lipolyse. Ook werd gekeken naar het effect van orale
toediening op gastrische lipolyse. In een gerandomiseerde, single blind cross-over studie
werden bij drie verschillende gelegenheden 15 gezonde proefpersonen nasogastrisch
geïntubeerd. Daarnaast werd op twee verschillende gelegenheden een subset van 6
gezonde proefpersonen naso-duodenaal geïntubeerd. Vrijwilligers kregen oraal of door
maag infusie een gehomogeniseerde emulsie toegediend, of een gastrische infusie van
water gevolgd door de oliefase (niet-gehomogeniseerd monster). De subset van 6
vrijwilligers kregen een maag infusie van een gehomogeniseerde emulsie of een

Samenvatting
148
gastrische infusie van een niet-gehomogeniseerde monster. De structuur van de emulsie,
de pH en de samenstelling van de vetten werden gemeten in zowel gastrische als
duodenale vloeistoffen. In ca. 3% van alle behandelingen werd vertering van vetten in de
maag (maag lipolyse) gevonden en waren de belangrijkste producten FFA en DG. De
concentraties van FFA in duodenale aspiraten waren hoger na toediening van
gehomogeniseerde emulsies dan na niet-gehomogeniseerde monsters (p <0,05).
Maaglediging was sneller voor de niet-gehomogeniseerde monsters dan voor de
gehomogeniseerde emulsies, zoals bepaald is door plasma paracetamol concentraties. In
zijn algemeenheid, was er geen effect waargenomen van orale verwerking. Ook
homogeniseren had geen effect op maag lipolyse. Wel zijn er verschillen in remming van
de maaglediging waargenomen: Duodenal release van FFA en de remming van de
maaglediging deden zich eerder voor bij gehomogeniseerde emulsies dan bij niet-
gehomogeniseerde monsters.
Vervolgens werd in hoofdstuk 5 onderzocht of de doorgevoerde effecten relevante
reacties op de spijsvertering en verzadigingsperceptie van vetten zou beïnvloeden. In
dezelfde opstelling als beschreven in hoofdstuk 4, werden galblaas volume, plasma CCK,
PYY, GLP-1 en ghreline concentratie, verzadigingswaarderingen zoals vastgesteld d.m.v.
een visuele analoge schaal, en ad libitum inname geanalyseerd. De resultaten toonden aan
dat de behandelingen van grote invloed zijn op het volume van de galblaas (p <0,001).
Meer specifiek, de meest langdurige samentrekking van de galblaas werd gevonden
wanneer de gehomogeniseerde emulsie oraal werd toegepast. Wanneer daarentegen, het
niet-gehomogeniseerde monster intragastrisch werd toegediend, was de samentrekking
van de galblaas het laagst. Daarnaast hebben we aanzienlijke effecten gevonden m.b.t. de
tijd en de behandeling op circulerende hormoon concentraties. In het geval van plasma
CCK, waren de concentraties 15 minuten na toevoeging van de gehomogeniseerde
emulsie significant hoger in vergelijking met het niet-gehomogeniseerde monster. De
concentraties voor PYY waren na 30 minuten significant hoger voor de gehomogeniseerde
emulsie in vergelijking met het niet-gehomogeniseerde monster. De respons van PYY en
GLP-1 op de niet-gehomogeniseerde monsters werden ongeveer 20 minuten vertraagd ten
opzichte van de gehomogeniseerde emulsie. Over de onderdrukking van ghreline werd
echter slechts een effect van tijd, maar niet van de behandelingen gevonden. Ook de
perceptie van verzadiging en ad libitum inname waren niet verschillend voor de
behandelingen. Toch was het gevoel om meer te eten sterker voor de gehomogeniseerde
emulsies in vergelijking met de niet-gehomogeniseerde monsters. Er werd geconcludeerd,
dat de intragastrische verdeling van vetten een effect heeft op plasma

149
hormoonconcentraties en galblaasmotoriek. Deze effecten werden niet gezien in de
verzadigingspercepties en de ad libitum inname. Derhalve staat de afgifte van
gastrointestinale hormonen niet in rechtstreeks verband met het verzadigende effect van
voedsel. Bij vergelijking van de resultaten van de vrijgekomen FFA, zoals waargenomen
in hoofdstuk 4, werd er wel een correlatie gezien tussen de FFA concentratie in de
twaalfvingerige darm (duodenum) en het volume van de galblaas, maar niet met de CCK
plasma concentratie. Daarom werd gespeculeerd dat hormonaal geïnduceerde
verzadigingsperceptie overheerst kunnen worden door andere mechanismen, zoals
maagdistentie.
In hoofdstuk 6, werd de invloed van emulsie eigenschappen en de gebruikte in vitro
digestiemodellen, met aandacht voor hun complexiteit, op lipolyse besproken. Bovendien
werd het effect van emulsies, in het maag verteringsstelsel, met betrekking tot hun
romigheid besproken met behulp van de bevindingen van een studie in een
varkensmodel. Daarnaast werden de effecten van vet digestieproducten op verzadiging en
de invloed van verzadigings hormonen besproken.

150

Summary
Samenvatting
Zusammenfassung

Zusammenfassung
152
Fette sind Schlüsselkomponenten unserer Nahrung. Da, insbesondere in der westlichen
Ernährung, gewöhnlich hochverarbeitete Lebensmittel aufgenommen werden, sind diese
Fette hauptsächlich in Form von Emulsionen vorzufinden. So können im Hinblick auf die
aktuelle Prävalenz von Adipositas speziell erzeugte Emulsionen, die die Sättigung und das
Sättigungsgefühl beeinflussen, eine Strategie darstellen, um einen übermäßigen Verzehr
von Lebensmitteln vorzubeugen.
In Kapitel 1 wird die Regulation einer Sättigungsreaktion während Fettaufnahme und
–verdauung betrachtet. Der Focus liegt hierbei auf der Regulation der Magenentleerung
und den daran beteiligten Hormonen, wie sie aus verschiedenen in vivo Untersuchungen
bekannt sind. Nach derzeitigem Kenntnisstand sind lipolytische Produkte, wie z.B. freie
Fettsäuren (FFS) hauptverantwortlich für die Ausschüttung von Hormonen, wie CCK,
sowie für die resultierenden Sättigungsreaktionen, wenn diese FFS im Darm detektiert
werden. Dennoch zeigt sich, dass diese Erkenntnisse vor allem auf intestinalen
Intubationsstudien beruhen und bisher nur wenige Untersuchungen den Magenbereich
einbeziehen. Diese wenigen Untersuchungen zeigen die Bedeutung der Lipolyse im
Magen und des kolloidalen Zustands der Fette in der Regulation der Sättigung. Bisher
sind diese Aspekte jedoch nicht in Kombination untersucht und es zeigen sich
widersprüchliche Ergebnisse. Ähnlich verhält es sich mit in vitro Modellen, die derzeit
verwendet werden, um ein grundsätzliches Verständnis des Verhaltens von Emulsionen
zu erhalten. Als diese Untersuchungen begannen, lag der Fokus in der Literatur auf der
intestinalen Verdauung von Fetten und hauptsächlich auf der Ausschüttung von FFS aber
nicht auf weiteren Produkten, wie Monoglyceriden (MG) und Diglyceriden (DG).
Weiterhin schlossen nur wenige Studien die Verdauung im Magen in ihre Überlegungen
mit ein. Wenn doch, wurde die spezifische Form und die Funktion des Magens
vernachlässigt oder vereinfacht nachgeahmt.
In der Studie in Kapitel 2 wurde die Freisetzung von FFS in Abhängigkeit von den
Eigenschaften der Emulsion mit Hilfe eines gängigen statischen in vitro Models, der pH-
stat Methode, untersucht. Emulsionen, die sich hinsichtlich ihrer Tropfengröße
unterschieden (fein, mittel und grob) und mittels Molkenproteinisolat (MPI) oder Gummi
Arabicum (GA) stabilisiert waren, wurden unter intestinalen Bedingungen verdaut.
Mittels Gaschromatographie (GC) wurden die Produkte der Lipolyse, wie FFS, MG, DG
und Triglyceride (TG) untersucht. Insgesamt zeigten die Untersuchungen mittels GC

153
geringere Konzentrationen an FFS, MG und DG bei gleichzeitig ansteigender
Tropfengröße. Es konnte gezeigt werden, dass die Verdauung von Emulsionen, die mittels
eines Protein-Polysaccharid-Emulgators (GA) stabilisiert wurden, zu einer höheren
Freisetzung von FFS führten, als Emulsionen die mittels eines proteinartigen Emulgators
(MPI) stabilisiert wurden. Weiterhin konnte bei mittels GA stabilisierten Emulsionen eine
schnellere Hydrolyse von DG zu MG im Vergleich zu MPI-stabilisierten Emulsionen
beobachtet werden. Die Konzentration an FFS war bei MPI-stabilisierten Emulsionen bei
der Messung mit Hilfe der GC zwei- bis dreimal höher im Vergleich mit den Ergebnissen
der pH-stat-Methode. Da die Bestimmung von FFS mittels pH-stat durch die Art des
Emulgators beeinflusst wird, lässt sich schlussfolgern, dass die Ergebnisse der pH-stat-
Methode mit Vorsicht bewertet und stets mit anderen Methoden, wie der GC-Methode
abgeglichen werden sollten.
In Kapitel 3 werden die Effekte von Magenpassage und Proteinstabilisierung auf die
Lipolyse, unter Berücksichtigung der schrittweisen Azidifikation und den Eigenschaften
des Magens, betrachtet. Zu diesem Zweck wurde ein dynamisches in vitro-Modell (TNO
Magen-Darm-Modell) mit einem speziell angefertigten Magenabschnitt eingesetzt. Um die
Flockung am isoelektrischen Punkt zu bestimmen, wurden MPI oder Lysozym als
Emulgatoren eingesetzt. Zur genaueren Charakterisierung wurden Mikroskopie,
Partikelgrößen-Analyse und GC verwendet. Unter den Bedingungen im Magen trennten
sich die MPI-stabilisierten Emulsionen am isoelektrischen Punkt des Proteins in Rahm
und Serumschicht. Mikroskopie und Partikelgrößen-Analyse zeigten größere ausgeflockte
Partikel bei den anfänglich negativ geladenen MPI-stabilisierten Emulsionen verglichen
mit den positiv geladenen Lysozym-stabilisierten Emulsionen. Vor allem FFS und DG
konnten im Magenabschnitt nachgewiesen werden, welche typische Spaltprodukte der
Magenlipase sind. Weiterhin konnten bei der Untersuchung der Serumschicht FFS
nachgewiesen werden, deren Vorhandensein in vivo möglicherweise zur Ausschüttung
von CCK führen könnte. Das Aufrahmen hatte einen geringeren Übertritt von Fetten in
das Duodenum zur Folge. Unabhängig davon unterschied sich die Gesamtkonzentration
an FFS im Magenabschnitt am Ende der Untersuchung kaum für beide Emulsionstypen.
Der Einfluss des Aufrahmens und des Vorhandenseins von FFS in der Serumschicht auf
die Magenentleerung kann hier aufgrund der Art des Modells nicht beurteilt werden.
In einer Humanstudie, die in den Kapiteln 4 und 5 vorgestellt wird, wurden
verschiedene Aspekte der Fettverabreichung, Fettverdauung und der damit verbundenen
physiologischen Antworten unter Einfluss der physikalischen Eigenschaften der Fette
untersucht. Kapitel 4 konzentriert sich dabei besonders auf den Einfluss der Verteilung

Zusammenfassung
154
des Fettes im Magen auf die Lipolyse in Magen und Duodenum. Weiterhin wurde der
additive Effekt der oralen Verabreichung auf die Lipolyse im Magen untersucht. In einer
randomisierten, doppel-blinden Cross-Over-Studie wurden 15 gesunde Probanden zu drei
unterschiedlichen Zeitpunkten nasogastral intubiert. Zusätzlich wurden 6 Personen dieser
Gruppe zu zwei weiteren Zeitpunkten nasoduodenal intubiert. Die Probanden erhielten
oral oder direkt in den Magen eine homogenisierte Emulsion oder eine Infusion von
Wasser gefolgt von einer Ölphase (unhomogenisierte Probe) direkt in den Magen. Die 6
Probanden erhielten eine Infusion einer homogenisierten Emulsion oder einer
unhomogenisierten Probe direkt in den Magen. Die Zusammensetzung der Emulsion, pH-
Wert und Fettzusammensetzung wurden in Magen- und Darmflüssigkeit analysiert. In
allen Behandlungsgruppen wurde im Magen eine Lipolyserate von ca. 3 % festgestellt. Die
Hauptprodukte waren FFS und DG. Die FFS-Konzentrationen waren in der
Darmflüssigkeit nach Gabe der homogenisierten Emulsionen höher im Vergleich zu den
unhomogenisierten Proben (p<0,05).
Anschließend, wie in Kapitel 5 zusammengefasst, wurde untersucht, ob diese bereits
gezeigten Effekte Vorgänge beeinflussen, die wichtig für die Fettverdauung und das
Sättigungsempfinden sind. Dafür wurde unter den gleichen Versuchsbedingungen wie in
Kapitel 4 bereits beschrieben, das Volumen der Gallenblase sowie die
Plasmakonzentrationen an CCK, PYY, GLP-1 und Ghrelin gemessen. Weiterhin wurde der
Grad der Sättigung, welcher mittels einer visualen, analogen Skala abgeschätzt wurde,
sowie die ad libitum Nahrungsaufnahme bestimmt. Die Ergebnisse zeigen einen
signifikanten Einfluss der unterschiedlichen Behandlungen auf das Gallenblasenvolumen
(p<0,001). Die am längsten anhaltende Kontraktion der Gallenblase konnte nach der
oralen Gabe der homogenisierten Emulsion beobachtet werden. Im Gegensatz dazu war
die Kontraktion der Gallenblase nach der in den Magen verabreichten unhomogenisierten
Probe am geringsten. Es konnte weiterhin gezeigt werden, dass die Konzentration der
zirkulierenden Hormone in Abhängigkeit von Zeit und Behandlung signifikant
beeinflusst wurden. Die Plasmakonzentration an CCK war 15 min nach der Gabe der
homogenisierten Emulsion signifikant höher im Vergleich zur unhomogenisierten Probe.
Nach 30 min war die Konzentration an PYY bei der homogenisierten Emulsion signifikant
höher im Vergleich zur unhomogenisierten Probe. Die Hormonantwort von PYY und
GLP-1 auf die Gabe der unhomogenisierten Probe erfolgte etwa 20 min später als die
Antwort auf die Gabe der homogenisierten Emulsion. Der Effekt auf die Suppression von
Ghrelin war nicht behandlungs- sondern nur zeitabhängig. Auch das
Sättigungsempfinden und die ad libitum Nahrungsaufnahme unterschieden sich nicht in

155
Abhängigkeit von der Behandlung. Dennoch war das „Verlangen nach Essen“ nach Gabe
der homogenisierten Emulsion stärker im Vergleich zur unhomogenisierten Probe.
Schlussfolgernd kann festgestellt werden, dass die Plasmahormonkonzentration und die
Motilität der Gallenblase durch die Verteilung der Lipide im Magen beeinflusst werden.
Die Effekte spiegeln sich weder im Sättigungsempfinden noch in der ad libitum
Aufnahme der Nahrung wider. Folglich besteht zwischen der Ausschüttung von
gastrointestinalen Hormonen und dem Sättigungseffekt von Lebensmitteln kein direkter
Zusammenhang. Betrachtet man gleichzeitig die Ergebnisse zur FFS-Ausschüttung aus
Kapitel 4, zeigt sich eine Korrelation zwischen der duodenalen Konzentration an FFS und
dem Gallenblasenvolumen. Die Konzentration an FFS korrelierte jedoch nicht mit der
Plasmakonzentration an CCK. Demzufolge wird das hormonal induzierte
Sättigungsgefühl vermutlich durch andere Mechanismen, wie die Magendehnung,
überdeckt.
Kapitel 6 beschreibt den Einfluss der Emulsionseigenschaften und der eingesetzten in
vitro Modelle auf die Lipolyse, mit besonderem Augenmerk auf deren Komplexität.
Weiterhin wurde der Effekt des Magendurchflusses der Emulsionen hinsichtlich des
Aufrahmverhaltens mit den Ergebnissen einer Studie an Schweinen diskutiert. Auch auf
die Bedeutung der Produkte der Fettverdauung und den Einfluss der Sättigungshormone
für das Sättigungsgefühl wurde in diesem Kapitel eingegangen.

156

Acknowledgements

Acknowledgements
158
The Netherlands, and in particular Wageningen, was not only a place where I did my
PhD-work for the last 4 years but also felt like home thanks to a lot of special people. I
gratefully acknowledge all your direct and indirect support throughout these last years.
Dear Harry and Rob, your advices and support helped me growing a lot in the past and
I feel very honoured to have you both as my promotors. I admire your critical way of
looking at my work and manuscripts, and your straightforward way of working. Above
all, you supported not only my work but also my private life, for which I am deeply
grateful.
Erika, I appreciate your help in every manner. You were not only a supervisor but
became a very good friend to me. Especially our work in Maastricht challenged us a lot
and brought us very close together. Thanks for all your good advices and endless
patience, especially in how to get my thoughts on paper.
I would like to acknowledge all members of the B-one007 TIFN project for their
contribution to this book. Raymond thank you for giving me this opportunity to start this
great project, George, Lex, Eric, Franklin, Esther, Rob, Mans, Jeroen, Monique, Marijke –
our meetings and discussions have been inspiring and I also very much appreciate all
your practical contributions. Marijke, special thanks goes to you as being my FCH-buddy
in the first year and now for being my paranymph. I loved the chats, sharing the lab and
my leisure time with you! Thanks for your support met de samenvatting! Jolan, it was
very nice to share the lab with you and thank you for giving me several rides to NIZO.
Eefjan, Rob and Peter, thanks for going through the ups and downs of endless GC sample
analysis with me. Harold, I would like to thank you again and again for your statistical
expertise – we both got grey hairs from it.
My dear 509-roomies Rudy, Jun, Jianwei, Marijn, Uttara, Milou, Ya and Stéphanie – it
was a pleasure to sit with you in an office and to share the life as a PhD in detail and in
general. Thanks for all your supporting discussions! Stéphanie, I feel very honoured that
you will go up the last stairs with me as my paranymph.
Martine, Amrah, Simone, Anja, Book and Ruud: organising such a great PhD trip to
Switzerland/Italy was a wonderful experience and I am proud that I could do that with
you together. Jolanda, I would like to thank you very much for helping with all your
organising and managing support!

159
I am very grateful for all assistance I received at Maastricht University. Ad Masclee
and Freddy Troost, thanks for giving us the opportunity to collaborate in the human
study. Ronald, Daniel, Samefko and Mark thanks for your support and the great time
during our study and for showing us around in Maastricht.
Meine lieben german buddies Anja, Anita, Stefan, Kerstin und Wibke, mit euch habe
ich tolle Freunde gefunden auch über die PhD-Zeit hinaus. Susann, ohne dich wäre ich
wahrscheinlich gar nicht nach Holland gegangen – danke für alles!! Simone, Yvonne, es
war eine große Freude, mit euch zusammen Holland zu entdecken.
Vier Jahre in Holland waren eine lange Zeit und trotzdem sind unsere Freundschaften
geblieben – ich drücke euch alle und danke euch dafür!! Jule – ohne dich wär alles doof –
und danke, dass du dich auch mit meiner Zusammenfassung auseinandergesetzt hast.
Nicole, Thomas, Steph, Steffi, Janni, Sandy, Matthias – danke für eure Besuche, damit
habt ihr mir immer ein Stück Heimat vorbeigebracht.
Meine liebe Mutti und mein lieber Vati, danke für euer Vertrauen, eure Zuversicht und
eure Unterstützung, und das nicht nur in den letzten Jahren – ihr seid großartige Eltern.
Meine liebe Ina, ich bin stolz darauf, so eine tolle Schwester zu haben, und Pierre und
Louis ich bin unendlich froh, dass es euch gibt. Liebe Margit, ich möchte dir für deine
Unterstützung vor allem in der letzten Zeit danken, da du uns, auch zusammen mit Olaf,
viele Dinge erleichtert hast.
Franky, die letzten Jahre wären ohne dich sehr leer gewesen und zusammen haben wir
diese Zeit durch Höhen und Tiefen bewältigt. Danke für deine unendliche Geduld und
deine Sicht der Dinge – es rückt alles wieder gerade. Meine kleine Laura, noch kannst du
nicht lesen, aber du sollst wissen, dass du mein größter Schatz bist und ich sehr stolz auf
dich bin.

160

About the author

About the author
162
Curriculum Vitae
Anne Helbig was born on the 12th
of June 1980 in Halle/Saale,
Germany. After graduating from high school (Torgymnasium,
Halle/Saale) in 1999 she started training at St. Elisabeth and St.
Barbara hospital (Halle/Saale). She obtained a degree as a nurse in
2002 and started studying Nutrition Science at Friedrich-Schiller-
University in Jena in the same year while continuing working as a
nurse at the Internal Medicine Department (St. Elisabeth- and St.
Barbara hospital, Halle/Saale). Before leaving to the Netherlands,
she went to the German Institute of Food Technology (Quakenbrück) for her Diploma
thesis entitled “Development of sugar reduced sweets using double emulsions of the
w/o/w type”. In 2008 she graduated in Nutrition Science and started her PhD thesis at the
Laboratory of Food Chemistry (Wageningen University, The Netherlands) and Top
Institute Food and Nutrition (TIFN). Her research was part of the TIFN project
“Engineered sensory and dietary functionality of dispersed fat”. The results of her PhD
project are described in this thesis.

163
List of publications
Helbig, A., Silletti, E., Timmerman, E., Hamer, R.J., Gruppen, H. (2012) In vitro study of
intestinal lipolysis using pH-stat and gas chromatography. Food Hydrocolloids, 28(1), 10-
19.
Helbig, A., Silletti, E., van Aken, G., Oosterveld, A., Minekus, M., Hamer, R.J.,
Gruppen, H. (2013) Lipid digestion of protein stabilized emulsions investigated in a
dynamic in vitro gastro-intestinal model system. Food Digestion, published online under
DOI: 10.1007/s13228-012-0029-6.
Helbig, A., Silletti, E., Troost, F., Sleijpen, R., Oosterveld, A., van Aken, G., Gruppen,
H., Masclee, A., Schipper, R., Hamer, R.J. (2013) Effect of oral administration and
intragastric distribution of lipids on lipolysis in gastric and duodenal human fluids, to be
submitted
Helbig, A., Troost, F., Silletti, E., Sleijpen, R., Keszthelyi D., van Aken, G., Gruppen, H.,
Schipper, R., Hamer, R.J., Masclee, A. (2013) Effect of oral administration and intragastric
distribution of fat on satiety responses in humans, to be submitted
Schwenzfeier, A., Helbig, A., Wierenga P.A., Gruppen, H. (2013) Emulsion properties
of algae soluble protein isolate from Tetraselmis sp. Food Hydrocolloids, 30(1), 258-263.

About the author
164
Overview of completed training activities
Discipline specific activities
• International advanced course food intake regulation - Nutrient sensing, VLAG,
Maastricht, The Netherlands, 2008
• Food hydrocolloids - Fundamentals and application, VLAG, Wageningen, The
Netherlands, 2009
• Nutritional and lifestyle epidemiology, VLAG, Wageningen, The Netherlands,
2011
• Delivery of functionality in complex food systems, Wageningen, The
Netherlands, 2009
• Food colloids - On the roads from interfaces to consumers, Granada, Spain, 2010
• Training period, Maastricht University, 2010/2011
General courses
• PhD week, VLAG, Bilthoven, The Netherlands, 2008
• Project & time management, WGS, Wageningen, The Netherlands, 2009
• Scientific writing, WGS, Wageningen, The Netherlands, 2009
• PhD competence assessment, WGS, Wageningen, The Netherlands, 2008
• Presentation course, TIFN, Wageningen, The Netherlands, 2009
Additional activities
• Preparation PhD research proposal, Wageningen, The Netherlands, 2008
• BSc/MSc students presentations and colloquia, Wageningen, The Netherlands,
2008-2012
• PhD presentations, Wageningen, The Netherlands, 2008-2012
• Food Chemistry study trip, Ghent, Belgium, 2009
• PhD study trip, China, 2008
• PhD study trip, Switzerland, Italy, 2010
• Member of Organization committee for the PhD study trip to Switzerland/Italy,
2010

165

166
The study presented in this thesis was performed within the framework of TI Food and Nutrition. Cover design by Kerstin Kaminski, cover picture oil drop© Graphik 123 RF Michelangelus This thesis was printed by GVO drukkers & vormgevers B.V. / Ponsen & Looijen, Ede, The Netherlands Anne Helbig, 2013





























