Embed Size (px)
Citation preview

Tumor and Stem Cell Biology
Differential WNT Activity in Colorectal Cancer ConfersLimited Tumorigenic Potential and Is Regulated by MAPKSignaling
David Horst1,3, Justina Chen1, Teppei Morikawa1, Shuji Ogino1,2, Thomas Kirchner3, andRamesh A. Shivdasani1
AbstractColorectal cancers (CRC) express the WNT effector protein b-catenin in a heterogeneous subcellular pattern
rather than uniformly in the nucleus. In this study, we investigated this important aspect of molecularheterogeneity in CRCs by analyzing its basis and relationship with tumor-initiating capability. CRC cells withthe highest WNT levels showed only a marginal increase in tumor initiation capacity. Notably, highWNT activitycorrelatedwith a coincident activation of robustmitogen-activated protein kinase (MAPK) signaling, whichwhenupregulated by KRAS expression or downregulated by epidermal growth factor receptor inhibition elicitedparallel effects onWNT activity. These findings suggested that on its own highWNT activity may not be a reliablesignifier of tumor-initiating potential or stem-like potential. Furthermore, they suggest that MAPK signaling is acritical modifier of intratumoral heterogeneity that contributes significantly to determining the impact of WNTactivity on stemness phenotypes in colon cancer cells. Cancer Res; 72(6); 1547–56. �2012 AACR.
IntroductionColorectal cancer (CRC) derives from normal colonic muco-
sa by stepwise accumulation of somatic genetic alterations,starting with inactivation of the tumor suppressor gene Ade-nomatous Polyposis Coli (APC) or activation of the oncogene ß-catenin (CTNNB1; ref. 1). Among additional mutations identi-fied frequently in human CRC are those in the KRAS oncogene,which occur in more than 40% of cases (2) and activatesignaling through the mitogen-activated protein kinase(MAPK) pathway. APC is a negative regulator ofWNT signalingand its mutational inactivation leads to accumulation ofb-catenin, which associates with transcription factors of theT-cell factor (TCF) family to activate WNT target genes (3).Although this understanding implies that the WNT pathway isuniformly active in all tumor cells that carry mutant APC orCTNNB1, colon cancers show substantial heterogeneity in the
accumulation of nuclear b-catenin, which is evident in about60% of resected tumor specimens, often at the invasive front (4,5). Such differential accumulation suggests that, in addition toAPC and CTNNB1 mutations, other pathway alterations orstimulatory factors external to tumor cells influence b-catenindistribution and WNT pathway activity in CRC (6, 7). Forexample, CpG island hypermethylation is inversely associatedwith b-catenin activation (8) and APC mutation (9), as well asamplification of the CDK8 locus, which encodes a member ofthe mediator complex, contributes to b-catenin-driven celltransformation (10).
Like other solid tumors, CRCs contain a subpopulation ofcells that differ from themajority of tumor cells in displaying anenhanced potential to establish tumors in immune compro-mised mice; these are thought to represent the clonogenictumor-initiating cells (11–13). Because some surface markersthat have been used to enrich tumor-initiating cells, includingCD133 and CD44, are proposed targets of WNT signaling (14,15), one possibility is that cells showing nuclear b-catenin mayharbor the highest tumorigenic potential. In support of thisidea, a recent report showed that cell populations isolated fromCRCs on the basis of high activity of a lentiviral WNT pathwayreporter were more tumorigenic than cells with low or absentreporter activity (16). By contrast, we report here that ourindependent investigation of differential WNT activity in CRCcell lines andprimaryCRCxenografts revealedpoor correlationbetween increased WNT activity and the potential to initiatetumors. In examining correlates of WNT signaling heteroge-neity, we further found that nuclear b-catenin accumulatedmost in cells with active MAPK signaling and that nuclearb-catenin correlated with KRASmutation in a large collectionof surgical CRC cases. Moreover, gain- and loss-of-function
Authors' Affiliations: 1Department of Medical Oncology, Dana-FarberCancer Institute and Departments of Medicine, and 2Pathology, Brighamand Women's Hospital and Harvard Medical School, Boston, Massachu-setts; and 3Pathologisches Institut der Ludwig-Maximilians-Universit€atM€unchen, Germany
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/)
Corresponding Authors: David Horst, Pathologisches Institut der LMU,ThalkirchnerStr.36,80337M€unchen,Germany.Phone:49-89-2180-73611;Fax: 49-89-2180-73671; E-mail: [email protected]; andRameshA.Shivdasani,Dana-FarberCancer Institute,450BrooklineAvenueBoston,MA 02215. Phone: 1-617-632-5746; Fax: 1-617-582-7198; E-mail:[email protected]
doi: 10.1158/0008-5472.CAN-11-3222
�2012 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 1547
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

studies revealed regulation of differential WNT activity byMAPKsignaling. Thus, commonmutations that activateMAPKsignaling through theKRAS oncogenemay be especially impor-tant in CRC, in part, by virtue of their effects on WNT pathwayactivity. One important feature of our study is the use of bothcell lines and primary human CRCs, which may model tumorproperties more accurately than do CRC cell lines alone.
Materials and MethodsCloning of lentiviral vectors
All template plasmids were obtained through Addgene(www.addgene.org). TOP-GFP was constructed by replacingthe PGK promoter in the lentiviral vector pRRLSIN.cPPT.PGK-GFP. WPRE (Constructed in Didier Trono's lab) with a 7xTCF/LEF optimal promoter cassette (7xTOP) from the M50 SuperTOPFlash plasmid (17). To construct the double color vectorTOP-GFP.mC, we inserted 4 additional TCF/LEF binding sites(GATCAAAGG) into a lentiviral TOP-dGFP reporter containing3 such binding sites (18), yielding 7xTOP-dGFP. We thenamplified this cassette using PCR and inserted it into the Hpa1site of lentiviral PGK-H2BmCherry (19). To convert destabi-lized dGFP into enhanced eGFP, we used site-directed muta-genesis (Stratagene) to insert a stop codon between GFP andthe attached ornithine decarboxylase sequence, yielding lenti-viral TOP-GFP.mC. To construct the control vectors FOP-GFPand FOP-GFP.mC, we replaced 7xTOP cassettes with synthetic7xFOP cassettes (IDT-DNA), which carry mutated TCF/LEFbinding sites (GgcCAAAGG). A mutant KRAS-expressing len-tiviral vector UG2K was constructed by inserting KRASG12V
from the pBabe K-Ras12V plasmid (20) into a pUltra lentiviralbackbone (Constructed in Malcolm Moore's lab) to yieldlentiviral pUBC-GFP-P2A-KRASG12V (UG2K). Modified vectorelements were verified by restriction analysis and sequencing.
Tissue culture, lentivirus production, transduction, andimmunoblotting
Caco2 and HEK293T cells were purchased from the Amer-ican Type Culture Collection. SW1222 cells were obtained fromthe Ludwig Institute for Cancer Research (New York). Cellswere cultured in Dulbecco's modified Eagle medium (DMEM;Gibco) supplemented with 10% FBS, 1% glutamine, and 1xpenicillin–streptomycin. HEK293T cells, plated in 10-cmdishes, were cotransfected with 10 mg lentiviral vector, 10 mgpCMV-dR8.91, and 2 mg pMD2.G (21) in the presence of 60 mLLipoD293 (Signagen).Mediumwas replaced after 12 hourswithDMEM containing 30% FBS and 36 hours later, conditionedmedium was passed through 0.45-mm filters (Millipore), mixed1:1 with DMEM, and used to infect cultured CRC cell lines ordisaggregated primary colon cancer cells in the presence of 8mg/mL polybrene (Sigma-Aldrich). Transduced tumor cellswere washed in PBS and transplanted into nonobese diabet-ic/severe combined immunodeficient (NOD/SCID) mice. Forsome experiments, single transduced GFP-positive tumor cellswere sorted into 96-well plates and expanded in culture beforexenotransplantation. Immunoblotting for UG2K vector testswere done using whole lysates of native or UG2K-transducedCaco2 or SW1222 cells with rabbit phospho-p44/42 MAPK
antibody (Ab), p44/42 MAPK Ab (1:1,000; both Cell Signaling),and mouse b-Actin Ab (1:200; Santa Cruz).
Primary colon cancers, tumor disaggregation, and cellsorting by flow cytometry
Primary tumor samples were obtained from Brigham andWomen's Hospital, Boston, following approval of an Institu-tional Review Board and patients' informed consent; toincrease material, primary tumors were expanded subcutane-ously inNOD/SCIDmice. These tumor xenografts weremincedwith scalpels and single-cell suspensions prepared by digestionwith collagenase IV (Worthington Biomedical) and 0.01%DNase I (Sigma-Aldrich) at 37�C for 30 to 60 minutes. Redblood cells were lysed using ACK lysing buffer (BioWhittaker,Lonza); debris were removed by Ficoll-Paque (GE HealthcareBio-sciences) gradient centrifugation. Cells were stainedwith APC-conjugated anti-mouse H-2Kd (clone SF1-1.1.1;eBioscience) Ab andHoechst 33258 (Sigma-Aldrich) to excludemouse and dead cells, respectively, andwith PerCP-eFluor 710-conjugated anti-human EpCAM (eBioscience) Ab to selecthuman CRC cells. Samples were filtered through 40-mm strai-ners before analysis or sorting on a FACSAria II instrument (BDBiosciences). Population purities of more than 90% in eachexperiment were confirmed by flow cytometry immediatelyafter sorting (Supplementary Fig. S4A).
Tumor induction and treatment in miceMice were housed in microisolator cages in a pathogen-free
colony; housing and experiments followed protocols approvedby an institutional Animal Care and Use Committee. Tumorswere induced in 6- to 8-week-old NOD/SCID (NOD.CB17-Prkdcscid; The Jackson Laboratory) mice. Transduced or con-trol CRC cell lines, disaggregated primary tumor xenografts, orsorted tumor cell fractions were suspended in 100 mL of a 1:1mixture of PBS and growth factor–depleted Matrigel (BDBioscience) before subcutaneous injection. For comparativeexperiments, GFPhigh and GFPlow tumor cell subpopulationswere injected into opposing flanks of the samemouse. Animalswere monitored weekly and tumors were measured usingcalipers. Mice were sacrificed when tumors reached a size of1.5 cmor in the event of earlier illness. In some experiments, 0.2mg cetuximab (Bristol-Meyers Squibb) or PBS (control) wasinjected into the peritonea of tumor-bearing mice every otherday, starting after subcutaneous tumors exceeded more than0.5 cm; CTNNB1 staining was analyzed after 10 days of cetux-imab treatment.
Gene expression analysesRNAwas isolated fromflow-sortedGFPhigh andGFPlow tumor
cells using TRIzol Reagent (Invitrogen), treated withDNase, andfirst strand cDNA was synthesized using a QuantiTect ReverseTranscription Kit (Qiagen). Quantitative real-time PCR applied40 cycles of amplification at 95�C (15 seconds) and 60�C (1minute) using Sybr green (Roche), a 7500 instrument (AppliedBiosystems), and the primers listed in Supplementary Table S1.Real-time reverse transcriptase PCR (RT-PCR) results were firstnormalized toGapdhmRNA levels in the same sample and thento levels of the corresponding transcript in GFPlow tumor cells.
Horst et al.
Cancer Res; 72(6) March 15, 2012 Cancer Research1548
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

For transcriptional profiling, total RNA was converted to cDNA,linearly amplified using the NuGEN Ovation V2 AmplificationSystem (NuGEN), and hybridized to Human Genome U133A 2.0arrays (Affymetrix) according to the manufacturer's protocols.Expression microarray data were analyzed using the dChippackage (22), considering transcripts that achieved a minimumchange of 1.2-fold between 2 groups as differentially expressed.Data can be accessed through GEO series number GSE32408.
Histology and immunohistochemistryTumor tissues were fixed in 4% paraformaldehyde, dehy-
drated in ethanol and xylenes, and embedded in paraffin.Sections were stained with hematoxylin and eosin (H&E). Forimmunohistochemistry, antigens were retrieved in 10 mmol/LNa citrate, pH 6, for 10 minutes in a pressure cooker (BiocareMedical). Slides were blocked with 0.5% hydrogen peroxide inmethanol for 20 minutes, followed by 5% FBS in PBS for 30minutes. Slides were incubated first withmouse b-catenin (BDTransduction Laboratories; 1:250), mouse active–b-Catenin(1:100; Millipore), rabbit GFP (1:250; Cell Signaling), or rabbitphospho-p44/42 MAPK Ab (1:250; Cell Signaling) overnight at4�C, washed with PBS, and then with biotinylated goat anti-mouse or anti-rabbit Ab (1:300; Vector Laboratories) for 1 hourat room temperature. Staining was detected using the Vectas-tain Elite ABC Kit (Vector) and diaminobenzidine as thesubstrate. For single staining, slides were counterstained withhematoxylin. For double staining, dephospho–b-Cateninstained slides were incubated with rabbit GFP Ab (1:250; CellSignaling) for 1 hour and developed with aMACH 3 Rabbit AP-Polymer Kit (Biocare medical) and Permanent Red (Dako). Forp-ERK/b-catenin and GFP/b-catenin double staining, slideswere incubatedfirstwith rabbitGFPor phospho-p44/42MAPKAb (1:250; Cell Signaling) for 1 hour at room temperature,washed in PBS, and then with goat anti-rabbit Alexa Fluor 488-or Alexa Fluor 546–conjugated secondary Ab (1:500; Invitro-gen). Fluorescence images were taken on a Nikon TE300microscope or a Nikon inverted Ti microscope with a Yoko-gawa spinning disk confocal system. Slides were then washedin PBS and stained with b-catenin Ab as described above.
CRC specimen collections and mutational analysisA total of 301 paraffin-embedded CRC tissueswere analyzed.
KRASmutations were determined on DNA extracted from thexenograft tumors and archived tissues by targeted pyrosequen-cing for codons 12 and 13 (23). Xenograft tumors were addi-tionally tested formutations inNRAS codons 12, 13, and 61, andBRAF codon 600 (24). Frequency data were analyzed using thec2 test. Statistical procedures applied SPSS version 19.0 (SPSSInc.). Human subjects committees at the respective hospitalsapproved the study.
ResultsIsolation of CRC cell populations with differing levels ofWNT pathway activityCaco2 and SW1222 human CRC cells carry classical APC
mutations (25) and formwell-differentiated glandular tumors inNOD/SCID mice, with heterogeneous intratumoral expression
and distribution of b-catenin (Supplementary Fig. S1A and B),similar to observations inprimaryCRCs (5). Tomark tumor cellsshowing high intrinsic WNT activity, we transduced these celllines with a lentiviral vector carrying TOP-GFP, aWNT-pathwayreporter in which multimerized TCF/LEF promoter elementsdrive expression of GFP (Fig. 1A). To test reporter specificity, weused FOP-GFP, a control vector with mutant TCF-binding sites(Supplementary Fig. S2A). Cells were expanded clonally toexclude the nontransduced population and then xenograftedintoNOD/SCIDmice (Fig. 1B). These clones showedhighGFP inall cultured cells (Supplementary Fig. S2B), whereas their xeno-grafts showed differential GFP expression (Fig. 1C, Supplemen-tary Fig. S2C), pointing to an environmental influence on WNTheterogeneity. In tumors expanded from Caco2TOP-GFP cells, theGFP signal colocalized with nuclear staining for b-catenin (Fig.1C) and in SW1222TOP-GFP tumors, GFP and nuclear b-cateninlargely overlapped (Supplementary Fig. S2C). Moreover, GFPexpression marked tumor cells with active, dephospho–b-cate-nin (Supplementary Fig. S2D).
To mark tumor cells with differential WNT activity inprimary CRCs, we first propagated fresh tumors as xenograftsin mice, noting that they preserved their original morphologyand displayed differential expression of nuclear b-catenin(Supplementary Fig. S1B and C). Because primary tumors havea heterogeneous composition, we preferred to avoid in vitroexpansion of single transduced cells, which might select atyp-ical clones. Instead we constructed a 2-color reporter, TOP-GFP.mC, which allows constitutive nuclear mCherry expres-sion in infected cells and regulated GFP expression in responseto WNT pathway activity (Fig. 1D, Supplementary Fig. S2E).Fluorescence microscopy revealed nuclear mCherry expres-sion in transduced primary CRC cells and a subset of these cellsexpressed GFP (Fig. 1E). Thus, after excluding nontransduced,mCherry-negative tumor cells, GFP again marked rare tumorcells that showed nuclear b-catenin (Fig. 1F).
We isolated GFPhigh and GFPlow cell fractions by flow cyto-metric sorting of disaggregated xenografts from TOP-GFP–transduced cell lines and TOP-GFP.mC–transduced primaryCRCs. The GFPhigh population showed significantly highermRNA levels of known WNT target genes, including the intes-tinal stem cell marker LGR5 (Fig. 1G and H, Supplementary Fig.S3A). Gene expression profiles of GFPhigh and GFPlow cells fromCaco2TOP-GFP revealed differential expression of 692 transcripts.Of these, 11 are annotated components of the WNT pathwayand 9 of the 11 transcripts showed higher expression in theGFPhigh cell fraction (Supplementary Fig. S3B). Thus, TOP-GFPbased constructs reliably marked a WNT-active subpopulationof CRC cells with nuclear b-catenin and allowed their isolationfrom bulk xenografts. However, the lesser enrichment for nucle-ar b-catenin and lower differential expression of WNT targetgenes among GFPhigh cells from SW1222TOP-GFP suggest thatCRCs vary in the degree of heterogeneity in WNT activity.
Tumor cell fractions with different levels of WNT activityare equally tumorigenic
To characterize tumor cells with high and lowWNT activity,we examined differential expression of all cell surface markergenes, as defined by Gene Ontology criteria, in GFPhigh and
Differential WNT Activity in Colon Cancer
www.aacrjournals.org Cancer Res; 72(6) March 15, 2012 1549
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

GFPlow cells isolated fromCaco2TOP-GFP xenografts. Among the16 differentially expressed surface markers were several thatare reported or suspected to mark tumorigenic CRC cells,including CD133 (PROM1), CD166 (ALCAM), and LGR5;expression of these transcripts was consistently higher in theGFPhigh than in the GFPlow cell fraction (Fig. 2A). mRNAexpression profiles of TOP-GFP.mC–transduced GFPhigh andGFPlow primary tumor cells also showed increased levels ofthese candidate stem and progenitor cell markers (Fig. 2A).These results suggested the possibility that CRC cells with thehighest WNT activity might harbor increased tumorigenic
potential, as others report (16). To test this hypothesis, weinjected limiting dilutions of GFPhigh and GFPlow colon cancercells into both flanks of NOD/SCID mice and followed theappearance of tumors. Indeed, GFPhigh cells fromCaco2TOP-GFP
xenografts were more tumorigenic than the GFPlow cell frac-tion and the tumors grew faster (Fig. 2B, Supplementary Fig.S4B). Despite extensive testing on many animals, however, wedid not observe such differences for cells isolated fromSW1222TOP-GFP tumors or from 3 primary CRCs transducedwith TOP-GFP.mC (Fig. 2B); one primary tumor manifestedlow tumorigenic potential in mice irrespective of cellular GFP
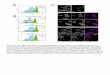
Figure 1. Use of a fluorescentreporter to mark the subpopulationof cells with highestWNT activity inCRC cell line xenografts andprimary tumors. A, lentiviral TOP-GFP vector used to transduceCRCcell lines. LTR, long terminal repeat;TOP, 7xTCF/LEF-responsivepromoter element; WPRE,woodchuck hepatitisposttranscriptional regulatoryelement. B, we expanded singlefluorescent GFPþ clones in cultureand injected them into NOD/SCIDmice to form tumor xenografts.C, section of a Caco2TOP-GFP
xenograft, stained for b-catenin(left) and GFP (right), showingcolocalization of the 2 signals. D,lentiviral double-color constructTOP-GFP.mC used to transduceprimary colon cancers, containingthe additional nuclear redfluorescence signal histone H2B-coupled mCherry, driven by thePGK promoter. E, confocal imageof a TOP-GFP.mC–transducedprimary colon cancer xenograftshowing ubiquitous red nuclearfluorescence, with additional greenfluorescence in a subset of tumorcells. F, section of a TOP-GFP.mC–transduced primary colon cancerxenograft, stained with b-cateninAb (left) and GFP (right); arrowsindicate colocalization of the 2markers. G and H, flow cytometricsorting of disaggregated tumor cellsuspensions from Caco2TOP-GFP
(G) and TOP-GFP.mC primarycolon cancer xenografts (H) basedon differential GFP fluorescence(left scatter plots) and qRT-PCRexpression analysis of a panel ofWNT-target genes and GFP (rightgraphs) in the sorted cellpopulations.
Horst et al.
Cancer Res; 72(6) March 15, 2012 Cancer Research1550
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

expression levels. These data showed that CRC cells with highWNTactivity andhigh expression of candidate cancer stemcellmarkers do not necessarily have an exclusive or enhancedcapacity to spawn new tumors.
High WNT activity is associated with MAPK pathwayactivation in colon cancer cellsRecent data fromzebrafish embryos suggest a role forMAPK
signaling in nuclear accumulation of b-catenin in the presenceof mutant APC (26). To investigate whether MAPK activity hasa role in differential WNT activity in CRC cell subpopulations,we first compared the profile of transcripts that are altered inGFPhigh or GFPlow tumor cells to a canonicalMAPK target genesignature (27). Of the 31 differentially expressed genes commonto the two sets (Fig. 3A), 29 were expressed at higher levels inWNT-active, GFPhigh cells compared with GFPlow cells (P <10�4, Fig. 3B); only 2 genes were expressed at lower levels,indicating activation of MAPK targets in the GFPhigh cellfraction. We then tested our xenograft sources for mutationsmost likely to affectMAPK signaling, inKRAS, BRAF, andNRAS.Both cell lines were wild type for all 3 genes but primary Tu1carried the V600E mutation in BRAF and primary Tu3 had aG12A mutation in KRAS. Surprisingly, although MAPK activa-
tion is considered a global feature of CRCs carrying suchmutations, the activity marker phospho-ERK appearedonly in a fraction of the cells in every tumor source (Fig. 3Cand data not shown). Importantly, immunostaining of Caco2and primary colon cancer xenografts revealed colocalizationof p-ERK and nuclear b-catenin (Fig. 3C). Thus, MAPK signal-ing is heterogeneous in CRC and increased in the tumor cellfraction with high WNT activity and nuclear accumulation ofb-catenin.
To assess this association in clinical samples, we exam-ined MAPK signaling and nuclear b-catenin accumulation in301 primary CRCs with known KRAS mutation status. Sim-ilar to the xenografts, phospho-ERK immunostaining in 118of these CRC specimens confirmed nonuniform MAPK sig-naling, irrespective of the underlying KRAS gene status(Supplementary Fig. S5A). We then scored nuclear b-cateninin tissue sections on a scale from 0 (no nuclear b-catenin) to3 (most tumor cells showing strong nuclear b-catenin),hence classifying cases as b-cateninlow (scale 0–1) or b-cate-ninhigh (scale 2–3, Fig. 3D), and observed that KRAS mutanttumors are significantly enriched for high nuclear b-catenin(Fig. 3E). Taken together, these data supported the idea thatactive KRAS might contribute to nuclear accumulation of
Figure 2. Lack of tumor-initiating capacity of WNT/GFPhigh tumor cell subpopulations despite enriched expression of suggested cancer stem cell markers. A,heat maps depicting differential expression of cell surface markers in GFPhigh and GFPlow cells isolated from Caco2TOP-GFP (left) and from TOP-GFP.mC–transduced primary tumor (P-Tu, right) xenografts. Columns represent samples and rows represent genes. Expression is represented in a pseudocolorscale (�3 to þ3), with red denoting high and blue denoting low relative expression. B, frequencies of tumor formation upon subcutaneous injectionof 10 to 100,000 flow-sorted GFPhigh and GFPlow tumor cells into NOD/SCID mice. Caco2TOP-GFP, SW1222TOP-GFP, and 3 independent TOP-GFP.mC–transducedprimary colon cancer xenografts (P-Tu1-3)were tested,with at least 3different tumor xenografts fromeachsource. The fractions indicate numbersof tumors formed (numerator) per experimental replicate (denominator). The P value refers to a significant difference in tumor frequency betweenGFPhigh and GFPlow cells (c2 test). n.s., nonsignificant.
Differential WNT Activity in Colon Cancer
www.aacrjournals.org Cancer Res; 72(6) March 15, 2012 1551
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

b-catenin in CRC, in which both MAPK signaling and WNTactivity show intratumor heterogeneity. These findings sug-gested roles for KRAS and MAPK signaling in differentialWNT activity.
Functional relationship of MAPK and WNT pathwayactivities
To test whether MAPK pathway activation influences WNTpathway activity, we constructed a lentiviral vector to express
Figure 3. Independent lines ofevidence correlating MAPKactivation with nuclear b-cateninexpression. A, Venn diagramshowing theoverlapof differentiallyexpressed genes in GFPhigh andGFPlow cells of Caco2TOP-GFP
tumors and a set of recentlyreported MAPK target genes (27).B, heat maps depicting the relativelevels of differentially expressedMAPK target genes (left) and non-MAPK targets (right) in GFPhigh andGFPlow cells isolated fromCaco2TOP-GFP tumors. The P valuedenotes significantly enrichedoverexpression of MAPK targets inGFPhigh cells (c2 test). C, doubleimmune staining for phospho-ERK(p-ERK, red) and b-catenin (brown),showing heterogeneousexpression and colocalization(arrows) of the two proteins inCaco2 (top) and primary coloncancer (P-Tu, bottom) xenografts.D, assessmentof nuclearb-cateninstaining in a collection of 301primary colon cancers. Tumorswere assigned scores from 0 (nonuclear b-catenin) to 3 (most tumorcells with strong nuclear b-catenin)and accordingly categorized asb-cateninlow (score 0–1) andb-cateninhigh (score 2–3). Images inthe bottom panels show highermagnifications of the areas boxedin the top panels. E, crosstabulation of nuclear b-cateninexpression and KRAS mutationalstatus (mut, activating mutation;wt, wild type) revealed significantly(c2 test) increased frequencies ofb-cateninhigh among cases withactivating KRAS mutations.
Horst et al.
Cancer Res; 72(6) March 15, 2012 Cancer Research1552
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

constitutively active KRAS and, as a marker of cell infection,GFP (Fig. 4A). Increased phospho-ERK levels in transducedcells verified that introduction of this vector activates MAPKsignaling, as expected (Fig. 4A). We infected CRC cell lines andprimary tumorswith low-titer lentivirus and grafted the cells inNOD/SCIDmice to obtain tumors containing a mix of infectedand uninfected tumor cells (Fig. 4A). Immunostaining of tumorsections revealed increased b-catenin levels and its nuclearlocalization in transduced, GFPþ tumor cells (Fig. 4B, Supple-mentary Fig. S5B). The levels of WNT target transcripts werealso increased selectively in the transduced cell fractions(Supplementary Fig. S5C). Thus, forced activation of MAPKsignaling through high levels of constitutively active KRASleads to nuclear accumulation ofb-catenin and increasedWNTactivity in CRC.To test whether inhibition of MAPK signaling opposes
b-catenin accumulation, we treated cultured CRC cells andmice bearing primary CRC xenografts with the epidermalgrowth factor receptor (EGFR) antibody cetuximab, whichinhibits cellular MAPK activity (28). Treated tumors showed
significantly fewer cells with nuclear b-catenin (Fig. 4C and D).Likewise, treatment of cultured Caco2TOP-GFP cells reducedexpression of GFP and WNT target genes (Supplementary Fig.5D). Moreover, primary tumor xenografts carrying the WNTreporter TOP-GFP.mC showed reduced GFP fluorescence anddepletion of the GFPhigh cell fraction (Fig. 4E and F). Thus,EGFR antibody-mediated blockade of MAPK signaling reducesWNT pathway activity in CRC cells. Surprisingly, these effectswere evident despite the presence of a KRAS mutation in oneprimary tumor.
DiscussionAPC or CTNNB1mutations in nearly all CRCs are presumed
to activate WNT signaling in all clonally derived tumor cells(29, 30). Heterogeneity in nuclearb-catenin levels inmost CRCsand differential expression of WNT target genes, therefore,present a paradox and suggest that additional factors contrib-ute toWNTpathway activity (6, 7). To isolate viable tumor cellsshowing differentialWNTactivity, we transduced xenografts ofprimary tumors and CRC cell lines with fluorescent reporters
Figure 4. KRAS activation increases and interference with EGFR signaling decreases the WNT-active cell fraction in Caco2 and primary tumor xenografts. A,vector schema. Lentiviral vector UG2K for Ubiquitin-C promoter (UBC)–driven overexpression of mutant KRASG12V together with GFP, separated by a P2Acleavage peptide. Right, immunoblot showing increased phospho-ERK (p-ERK) and unchanged total-ERK (ERK) expression in UG2K-transduced (U),serum-starved cells compared with native (N) Caco2 and SW1222 cells. Actin served as a loading control. Bottom, experimental scheme to generate coloncancer xenografts with mosaic UG2K transduction. Colon cancer cell lines and primary tumor xenografts were exposed for short periods to low titersof UG2K lentivirus and injected subcutaneously into NOD/SCID mice for tumor formation. B, sections of mosaic UG2K-transduced tumor xenografts doublestained for GFP and b-catenin, showing increased nuclear b-catenin in GFP-expressing tumor cells (arrows). Images in the bottom panels show highermagnifications of areas boxed in the top panels. C andD, representative b-catenin immune staining of a primary colon cancer xenograft (C) and quantitation ofthe b-catenin positive cell fraction in 3 primary colon cancer xenografts (D) treated in vivo with cetuximab (CTX) or PBS. E and F, reduced GFPhigh tumorcell fraction in TOP-GFP.mC–transduced primary colon cancer xenografts following cetuximab (CTX) treatment, as measured by flow cytometry ofdisaggregated tumor cells (F). Error bars indicate the SEM from 3 experiments.
Differential WNT Activity in Colon Cancer
www.aacrjournals.org Cancer Res; 72(6) March 15, 2012 1553
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

of canonical WNT activity (18). Although cultured CRC cellsexpressed GFP uniformly, all xenografts displayed highlyheterogeneous GFP expression, similar to a recent study(16). Thus, the local environment or the 3-dimensionalstructure in xenografts seem necessary to reveal this latentheterogeneity. In tumor xenografts, nuclear b-catenin colo-calized with high GFP expression as well as with enrichedexpression of WNT pathway components and WNT targetgenes in GFPhigh tumor cells. These data confirmed differ-ential WNT pathway activity within CRCs and provided areliable means to isolate cells with different levels of WNTactivity. However, even though differentially expressed genesincluded the candidate human CRC stem cell markersCD133, CD44, and CD166 (11–13), and the intestinal stemcell marker LGR5 (31), WNT/GFPhigh cells from only 1 of the5 tested sources showed a modest increase in tumor-initi-ating capacity. These data from a small number of primaryCRC specimens indicate that high WNT activity does notsignify particular tumorigenic potential in CRC and is not areliable marker for tumor initiation in xenografts. Accord-ingly, they question the generality of a recent argument thathigh WNT activity is a distinguishing feature of CRC stemcells (16). The contrast between our findings and those ofVermeulen (16) mirror the conflicting results from differentlaboratories' use of CD133 as a surface marker of tumor-initiating CRC cells (32). We suggest that the differing out-comes of such studies reflect highly variable genetic orphenotypic properties of tumors. Differences in experimen-tal design, such as whether primary tumor cells were firstexpanded in spheroid cultures (16) or cultivated directly inxenografts, may also contribute to the discrepancy. Ofcourse, it is still unclear how accurately tumor xenograftsmodel tumor-initiating cell subpopulations (33, 34).
Despite the lack of an obvious association with tumorinitiation, heterogeneous WNT activity correlates with dis-ease progression in CRC (5, 6, 26), adding to the importanceof identifying the causes. Brabletz and colleagues and Ver-meulen and colleagues have separately proposed that tumorcell stimulation by the microenvironment may modulateb-catenin accumulation and WNT pathway activity (6, 16),and studies in zebrafish embryos and a single CRC cell linerecently implicated tumor cell MAPK signaling in the under-lying mechanism (26). Reinforcing this idea, we observedcolocalization of nuclear b-catenin and the MAPK pathwayindicator phospho-ERK in cell line and primary tumorxenografts, and expression of MAPK target genes is enrichedin highly WNT-active CRC cells. We also detected a statis-tical association between nuclear b-catenin and the pres-ence of activating mutations in the MAPK pathway geneKRAS. Our data are consistent with the observation thatintestinal tumors in KrasG12V; Apcþ/1638N mice have morecells with nuclear b-catenin than do their Apcþ1638N litter-mates (35). Notably both, KRAS mutant and wild-typetumors showed heterogeneous MAPK activity, implying thatsignaling remains responsive to exogenous factors evenwhen cells carry activating mutations. Although this obser-vation begs an explanation for variable MAPK pathwayactivity among cells in the same tumor, our data begin to
explain why MAPK and WNT pathway activities are high inthe same cells.
KRASmutations that activate MAPK signaling are especiallycommon in selected cancers, including about 40% of CRCs (2),in which they occur early in the disease course, soon after APCor CTNNB1 mutations (36). KRAS mutations in the intestinalepithelium are oncogenic only when accompanied by dereg-ulation of the WNT pathway (37). The full significance of thisassociation and of the sequence of mutations is unclear andour data shed useful light. Forced expression of mutant KRASin CRCs enhanced nuclear b-catenin accumulation andincreased the levels of WNT target genes, whereas blockingthe tyrosine kinase receptor EGFR, an activator of MAPKsignaling (38), had the converse effects. Thus, MAPK signalingdoes not merely coincide with nuclear b-catenin but alsoregulates it. In line with these observations, insertional muta-genesis in ApcMin mice recently uncovered 4 new candidatepositive modifiers of canonical Wnt signaling (39) and one ofthese, EGF receptor kinase substrate 8, transduces EGF andRAS signals. Thus, modulation of canonical Wnt signalingthrough MAPK is a factor in colorectal tumorigenesis, andour study highlights this effect in tumor cell subpopulations.Phelps and colleagues recently proposed that KRAS and RAF1,but not MEK1, induce nuclear accumulation of b-catenin inAPC-deficient cells (26). Further study is necessary to elucidatethe detailed basis for this pathway cross-talk.
The fact that EGF and, perhaps, additional signals fromthe tumor microenvironment contribute to heterogeneousWNT activity in CRC also suggests new considerations fortreatment of this disease. There are intensive ongoing effortsto design drugs that suppress WNT activity in CRC; thesedrugs may synergize with those that target MAPK signaling,such as cetuximab. Conversely, investigators recentlydescribed the use of transcriptionally active b-catenin toinduce apoptosis in APC-deficient tumors (40). If growthfactor signaling enhances WNT activity, then simultaneoustargeting of growth factor pathways may downregulate theintended target and hence diminish the effects of WNT-directed therapy. Our observation that cetuximab reducedintratumoral heterogeneity of nuclear b-catenin also raisesthe possibility that the degree of heterogeneity in WNTpathway activity may help predict the therapeutic responseto agents that target MAPK signaling. Thus, a fuller under-standing of the interactions between WNT and MAPKsignaling will better inform the design and interpretationof therapeutic trials.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: D. Horst and R.A. Shivdasani.Development of methodology: D. Horst and R.A. Shivdasani.Acquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): D. Horst, J. Chen, T. Morikawa, S. Ogino, T. Kirchner,and R.A. Shivdasani.Analysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): D. Horst, J. Chen, S. Ogino, T. Kirchner, and R.A.Shivdasani.Writing, review, and/or revision of the manuscript: D. Horst, S. Ogino, T.Kirchner, and R.A. Shivdasani.
Horst et al.
Cancer Res; 72(6) March 15, 2012 Cancer Research1554
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

Administrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): D. Horst, J. Chen.Study supervision: R.A. Shivdasani.
AcknowledgmentsThe authors thank Dr. Michael Zinner and members of the Departments of
Surgery and Pathology at Brigham&Women's Hospital for generously providingfreshly resectedCRC specimens and the Ludwig Institute for Cancer Research forthe gift of SW1222 cells.
Grant SupportThework was supported by a grant from the Harvard Stem Cell Institute (R.A.
Shivdasani), a fellowship from the Deutsche Forschungsgemeinschaft (D. Horst),and grants R01 CA151993 (S. Ogino) and P50CA127003 from the NIH.
The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received September 28, 2011; revised December 28, 2011; accepted January 16,2012; published OnlineFirst February 8, 2012.
References1. FearonER,VogelsteinB. Ageneticmodel for colorectal tumorigenesis.
Cell 1990;61:759–67.2. Vaughn CP, Zobell SD, Furtado LV, Baker CL, Samowitz WS. Fre-
quency of KRAS, BRAF, and NRAS mutations in colorectal cancer.Genes Chromosomes Cancer 2011;50:307–12.
3. Clevers H. Wnt/beta-catenin signaling in development and disease.Cell 2006;127:469–80.
4. Brabletz T, JungA,HermannK,GuntherK,HohenbergerW,Kirchner T.Nuclear overexpression of the oncoprotein beta-catenin in colorectalcancer is localized predominantly at the invasion front. Pathol ResPract 1998;194:701–4.
5. Horst D, Reu S, Kriegl L, Engel J, Kirchner T, Jung A. The intratumoraldistribution of nuclear beta-catenin is a prognostic marker in coloncancer. Cancer 2009;115:2063–70.
6. Brabletz T, Jung A, Reu S, Porzner M, Hlubek F, Kunz-Schughart LA,et al. Variable beta-catenin expression in colorectal cancers indicatestumor progression driven by the tumor environment. Proc Natl AcadSci U S A 2001;98:10356–61.
7. Fodde R, Brabletz T. Wnt/beta-catenin signaling in cancer stemnessand malignant behavior. Curr Opin Cell Biol 2007;19:150–8.
8. Kawasaki T, Nosho K, Ohnishi M, Suemoto Y, Kirkner GJ, Dehari R,et al. Correlation of beta-catenin localization with cyclooxygenase-2expression and CpG island methylator phenotype (CIMP) in colorectalcancer. Neoplasia 2007;9:569–77.
9. SamowitzWS, Slattery ML, Sweeney C, Herrick J, Wolff RK, AlbertsenH. APC mutations and other genetic and epigenetic changes in coloncancer. Mol Cancer Res 2007;5:165–70.
10. Firestein R, Bass AJ, Kim SY, Dunn IF, Silver SJ, Guney I, et al. CDK8 isa colorectal cancer oncogene that regulates beta-catenin activity.Nature 2008;455:547–51.
11. Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, Cho RW, et al. Phenotypiccharacterization of humancolorectal cancer stemcells. ProcNatl AcadSci U S A 2007;104:10158–63.
12. O'Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cellcapable of initiating tumour growth in immunodeficient mice. Nature2007;445:106–10.
13. Ricci-Vitiani L, Lombardi DG, Pilozzi E, BiffoniM, TodaroM, PeschleC,et al. Identification and expansion of human colon-cancer-initiatingcells. Nature 2007;445:111–5.
14. Katoh Y, Katoh M. Comparative genomics on PROM1 gene encodingstem cell marker CD133. Int J Mol Med 2007;19:967–70.
15. Wielenga VJ, Smits R, Korinek V, Smit L, Kielman M, Fodde R, et al.Expression of CD44 in Apc and Tcf mutant mice implies regulation bythe WNT pathway. Am J Pathol 1999;154:515–23.
16. Vermeulen L, De Sousa EMF, van der Heijden M, Cameron K, deJong JH, Borovski T, et al. Wnt activity defines colon cancer stemcells and is regulated by the microenvironment. Nat Cell Biol 2010;12:468–76.
17. Veeman MT, Slusarski DC, Kaykas A, Louie SH, Moon RT. Zebrafishprickle, a modulator of noncanonical Wnt/Fz signaling, regulatesgastrulation movements. Curr Biol 2003;13:680–5.
18. Reya T, Duncan AW, Ailles L, Domen J, Scherer DC, Willert K, et al. Arole for Wnt signalling in self-renewal of haematopoietic stem cells.Nature 2003;423:409–14.
19. Kita-Matsuo H, Barcova M, Prigozhina N, Salomonis N, Wei K, JacotJG, et al. Lentiviral vectors and protocols for creation of stable hESC
lines for fluorescent tracking and drug resistance selection of cardi-omyocytes. PLoS One 2009;4:e5046.
20. Khosravi-Far R, White MA, Westwick JK, Solski PA, Chrzanowska-Wodnicka M, Van Aelst L, et al. Oncogenic Ras activation of Raf/mitogen-activated protein kinase-independent pathways is sufficientto cause tumorigenic transformation. Mol Cell Biol 1996;16:3923–33.
21. Naldini L, Blomer U, Gage FH, Trono D, Verma IM. Efficient transfer,integration, and sustained long-term expression of the transgene inadult rat brains injectedwith a lentiviral vector. ProcNatl AcadSciUSA1996;93:11382–8.
22. Li C, Hung Wong W. Model-based analysis of oligonucleotide arrays:model validation, design issues and standard error application.Genome Biol 2001;2:RESEARCH0032.
23. Ogino S, Kawasaki T, Brahmandam M, Yan L, Cantor M, Namgyal C,et al. Sensitive sequencing method for KRAS mutation detection byPyrosequencing. J Mol Diagn 2005;7:413–21.
24. Irahara N, Baba Y, Nosho K, Shima K, Yan L, Dias-Santagata D, et al.NRAS mutations are rare in colorectal cancer. Diagn Mol Pathol2010;19:157–63.
25. Ilyas M, Tomlinson IP, Rowan A, Pignatelli M, Bodmer WF. Beta-catenin mutations in cell lines established from human colorectalcancers. Proc Natl Acad Sci U S A 1997;94:10330–4.
26. Phelps RA, Chidester S, Dehghanizadeh S, Phelps J, Sandoval IT, RaiK, et al. A two-stepmodel for colon adenoma initiation andprogressioncaused by APC loss. Cell 2009;137:623–34.
27. Jurchott K, Kuban RJ, Krech T, Bluthgen N, Stein U, Walther W, et al.Identification of Y-box binding protein 1 as a core regulator of MEK/ERK pathway-dependent gene signatures in colorectal cancer cells.PLoS Genet 2010;6:e1001231.
28. Huang S, Armstrong EA, Benavente S, Chinnaiyan P, Harari PM. Dual-agent molecular targeting of the epidermal growth factor receptor(EGFR): combining anti-EGFR antibody with tyrosine kinase inhibitor.Cancer Res 2004;64:5355–62.
29. Fearon ER, Hamilton SR, Vogelstein B. Clonal analysis of humancolorectal tumors. Science 1987;238:193–7.
30. Morin PJ, Sparks AB, Korinek V, Barker N, Clevers H, Vogelstein B,et al. Activation of beta-catenin-Tcf signaling in colon cancer bymutations in beta-catenin or APC. Science 1997;275:1787–90.
31. Barker N, van Es JH, Kuipers J, Kujala P, van denBornM,CozijnsenM,et al. Identification of stem cells in small intestine and colon by markergene Lgr5. Nature 2007;449:1003–7.
32. Shmelkov SV, Butler JM, Hooper AT, Hormigo A, Kushner J, Milde T,et al. CD133 expression is not restricted to stem cells, and bothCD133þ and CD133- metastatic colon cancer cells initiate tumors.J Clin Invest 2008;118:2111–20.
33. Clevers H. The cancer stem cell: premises, promises and challenges.Nat Med 2011;17:313–9.
34. Shackleton M, Quintana E, Fearon ER, Morrison SJ. Heterogeneity incancer: cancer stem cells versus clonal evolution. Cell 2009;138:822–9.
35. Janssen KP, Alberici P, Fsihi H, Gaspar C, Breukel C, Franken P, et al.APC and oncogenic KRAS are synergistic in enhancing Wnt signalingin intestinal tumor formation and progression. Gastroenterology2006;131:1096–109.
36. Kinzler KW, Vogelstein B. Lessons from hereditary colorectal cancer.Cell 1996;87:159–70.
Differential WNT Activity in Colon Cancer
www.aacrjournals.org Cancer Res; 72(6) March 15, 2012 1555
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

37. Haigis KM, Kendall KR, Wang Y, Cheung A, Haigis MC, Glickman JN,et al. Differential effects of oncogenic K-Ras and N-Ras on prolifer-ation, differentiation and tumor progression in the colon. Nat Genet2008;40:600–8.
38. Katz M, Amit I, Yarden Y. Regulation of MAPKs by growth factorsand receptor tyrosine kinases. Biochim Biophys Acta 2007;1773:1161–76.
39. March HN, Rust AG, Wright NA, Ten Hoeve J, de Ridder J, Eldridge M,et al. Insertional mutagenesis identifiesmultiple networks of cooperat-ing genes driving intestinal tumorigenesis. Nature genetics 2011;43:1202–9.
40. Zhang L, Ren X, Alt E, Bai X, Huang S, Xu Z, et al. Chemoprevention ofcolorectal cancer by targeting APC-deficient cells for apoptosis.Nature 2010;464:1058–61.
Horst et al.
Cancer Res; 72(6) March 15, 2012 Cancer Research1556
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222

2012;72:1547-1556. Published OnlineFirst February 8, 2012.Cancer Res David Horst, Justina Chen, Teppei Morikawa, et al. Tumorigenic Potential and Is Regulated by MAPK SignalingDifferential WNT Activity in Colorectal Cancer Confers Limited
Updated version
10.1158/0008-5472.CAN-11-3222doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2012/02/08/0008-5472.CAN-11-3222.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/72/6/1547.full#ref-list-1
This article cites 39 articles, 9 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/72/6/1547.full#related-urls
This article has been cited by 18 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/72/6/1547To request permission to re-use all or part of this article, use this link
on June 14, 2020. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 8, 2012; DOI: 10.1158/0008-5472.CAN-11-3222