Embed Size (px)
Citation preview

Short communication
Nechama Gilboa-GarberBatya LerrerEfrat Lesman-MovshovichOrly Dgani
Faculty of Life Sciences,Bar-Ilan University,Ramat-Gan, Israel
Differential staining of Western blots of humansecreted glycoproteins from serum, milk, saliva,and seminal fluid using lectins displaying diversesugar specifities
Human milk, serum, saliva, and seminal fluid glycoproteins (gps) nourish and protectnewborn and adult tissues. Their saccharides, which resemble cell membrane com-ponents, may block pathogen adhesion and infection. In the present study, they wereexamined by a battery of lectins from plants, animals, and bacteria, using hemaggluti-nation inhibition and Western blot analyses. The lectins included galactophilic onesfrom Aplysia gonad, Erythrina corallodendron, Maclura pomifera (MPL), peanut, andPseudomonas aeruginosa (PA-IL); fucose-binding lectins from Pseudomonas aerugi-nosa (PA-IIL), Ralstonia solanacearum (RSL), and Ulex europaeus (UEA-I), and man-nose/glucose-binding Con A. The results demonstrated the chosen lectin efficiency fordifferential analysis of human secreted gps as compared to CBB staining. Theyunveiled the diversity of these body fluid gp glycans (those of the milk and seminal fluidbeing highest): the milk gps interacted most strongly with PA-IIL, followed by RSL; thesaliva gps with RSL, followed by PA-IIL and MPL; the serum gps with Con A and MPL,followed by PA-IIL and RSL, and the seminal plasma gps with RSL and MPL, followedby UEA-I and PA-IIL. The potential usage of these lectins as probes for scientific,industrial, and medical purposes, and for quality control of the desired gps is clearlyindicated.
Keywords: Human body fluids / Human glycoproteins / Lectins / SDS-PAGE / Westernblotting DOI 10.1002/elps.200500392
Human body fluids are rich in glycoproteins (gps) thatnourish and protect adult tissues and breast-fed new-borns from infections [1–4]. Their saccharides may func-tion as decoys, abrogating lectin-mediated microbialpathogen adhesion [1, 2, 5]. The specific binding ofdiverse lectins to saccharides is used for detection andisolation of gps which bear them. Examples are theapplications for these purposes of wheat germ agglutininthat is specific to N-acetylglucosaminyl/sialic acid [6], andCon A, which binds mannose/glucose [7]. Additionalexamples include analyses of avian egg white and humansaliva gps [8–10] by diverse lectins for studies of their
glycosylations and variations in disease, and of humanmilk gps [1, 2, 11] for antimicrobial adhesion potential [2].Usage of Pseudomonas aeruginosa fucose (Fuc)-bindinglectin (PA-IIL) [12] for detection of fucosylated gps in hu-man milks, indicated its efficiency as a powerful tool fordifferentiation between human and cow’s milk [13].
The present communication describes the study of hu-man blood plasma, milk, saliva, and seminal plasma gpswith a battery of nine lectins, segregated to four sugar-specificity groups (Table 1). Five of the lectins are galac-tophilic: those of Aplysia gonad (AGL) [14–16], Erythrinacorallodendron (ECorL) [17], Maclura pomifera (MPL) [7],peanut (PNA) [7], and Pseudomonas aeruginosa (PA-IL)[12, 18, 19]. ECorL, MPL, and PA-IL bind both galactose(Gal) and N-acetylgalactosamine, while AGL and PNA donot interact with the second sugar. One lectin is Fuc spe-cific, that of Ulex europaeus (UEA-I) [7]; two bind Fuc andmannose: from the opportunistic pathogenic bacteria P.aeruginosa (PA-IIL) [12, 20, 21] and Ralstonia solana-cearum (RSL) [21], and one the mannose/glucose-bindinglectin Con A [7].
Correspondence: Professor Nechama Gilboa-Garber, Faculty ofLife Sciences, Bar-Ilan University, Ramat-Gan 52900, IsraelE-mail: [email protected]: 1972-3-5351824
Abbreviations: AGL, Aplysia gonad lectin; ECorL, Erythrinacorallodendron lectin; Fuc, L-fucose; Gal, D-galactose; gps, gly-coproteins; MPL, Maclura pomifera lectin; PA-IL, first Pseudo-monas aeruginosa lectin; PA-IIL, second Pseudomonas aerugi-nosa lectin; PNA, peanut lectin; RSL, Ralstonia solanacearumlectin; SF, seminal fluid; UEA-I, first lectin of Ulex europaeus
4396 Electrophoresis 2005, 26, 4396–4401
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Electrophoresis 2005, 26, 4396–4401 Staining of electroblotted human secreted glycoproteins by lectins 4397
Table 1. Lectins used, their source, and carbohydratespecificity
Lectin(abbre-viation)
Source Carbohydrate Refe-rences
AGL Animal: Aplysia gonad GalUA, Gal [14–16]Con A Plant: Canavalia ensiformis Man, Fru, Glc [7]ECorL Plant: E. corallodendron Gal, GalNAc, Lac [17]MPL Plant: M. pomifera Gal, GalNAc [7]PA-IL Bacterial: P. aeruginosa Gal, GalNAc [12, 18]PA-IIL Bacterial: P. aeruginosa Fuc, Fru, Man [12, 20]PNA Plant: Arachis hypogaea Galb1,3 GalNAc [7]RSL Bacterial: R. solanacearum Fuc, Fru, Man [21]UEA-I Plant: U. europaeus Fuca1,2 Gal [7, 26]
Gal, galactose; GalNAc, N-acetylgalactosamine; GalUA,galacturonic acid; Glc, glucose; Fru, fructose; Fuc,fucose; Man, mannose.
Using both hemagglutination inhibition and Western blot-ting for analyses of the lectin–glycan interactions showedconsiderable differences of the diverse human body fluidgp glycans even among lectins of the same group despitetheir general similarity. Those were due to subtle differ-ences in their sensitivities to a- or b-linkages of the termi-nal target sugars, the subterminal sugars, and adjacentcomponents.
The lectins AGL, PA-IL, PA-IIL, and RSL were prepared inour laboratory from the marine snail Aplysia, P. aeruginosaATCC 33347, and R. solanacearum ATCC 11696 cells,respectively, as previously described [12, 14, 21]. Con A,ECorL, MPL, PNA, and UEA-I were purchased from Sigma.Peroxidase-labeled lectins were prepared using glutar-aldehyde coupling [22, 23], as previously described [8].
O-type human erythrocytes were obtained from healthydonors. Five percent v/v suspension in saline of thrice-washed untreated (for MPL), sialidase-treated (for PNA),or 0.1% papain-treated (for all the other lectins) cells wasused, as previously described [8, 12].
Each human body fluid (1 mL samples) was obtained fromat least ten different donors and was separately treatedand analyzed. Human sera were separated from clottedblood samples withdrawn from healthy donors, dialyzedovernight (at 47C) through a cellulose membrane with cut-off size of Mr 10 000 (using 0.85% NaCl solution) and thenstored at 2207C till usage.
Human milks were obtained from healthy mothers(volunteers) at various stages of breastfeeding. Eachsample was centrifuged (10 0006g) for 10 min and theintermediate skim phase was carefully collected andtreated as above.
Human salivas from healthy donors were collected withcooling, following rinsing the mouth with water. They werecentrifuged at 10 0006g for 10 min and the supernatantswere treated as above.
Seminal fluids (SFs), obtained from fertility clinics, werecentrifuged at 15 0006g for 15 min. The supernatantswere collected and treated as above.
The body fluids used with UEA-I were only of “secretor”phenotypes, containing the H-2 blood group substance intheir secretions [24].
For the hemagglutination test, each lectin solution (1 mg in50 mL) was serially diluted with 50 mL saline to producetwo-fold dilutions, and then 50 mL of both saline and thecompatible erythrocyte suspension were added to eachtube. After 30 min at room temperature, the tubes werecentrifuged for 30 s (10006g), and the hemagglutinationintensity was examined [12]. The highest dilution leadingto agglutination of all the erythrocytes in one mass wasused for the inhibition test.
For the hemagglutination inhibition test, each body fluidpreparation examined was serially diluted in 50 mL volumeand then 50 mL of the above chosen lectin dilution wasadded to each tube. After 30 min interaction, 50 mL of thecompatible erythrocyte suspension was added to eachtube and the hemagglutination was examined as above.The hemagglutination inhibition intensity was representedby the number of tubes without hemagglutination pre-ceding its reappearance [13].
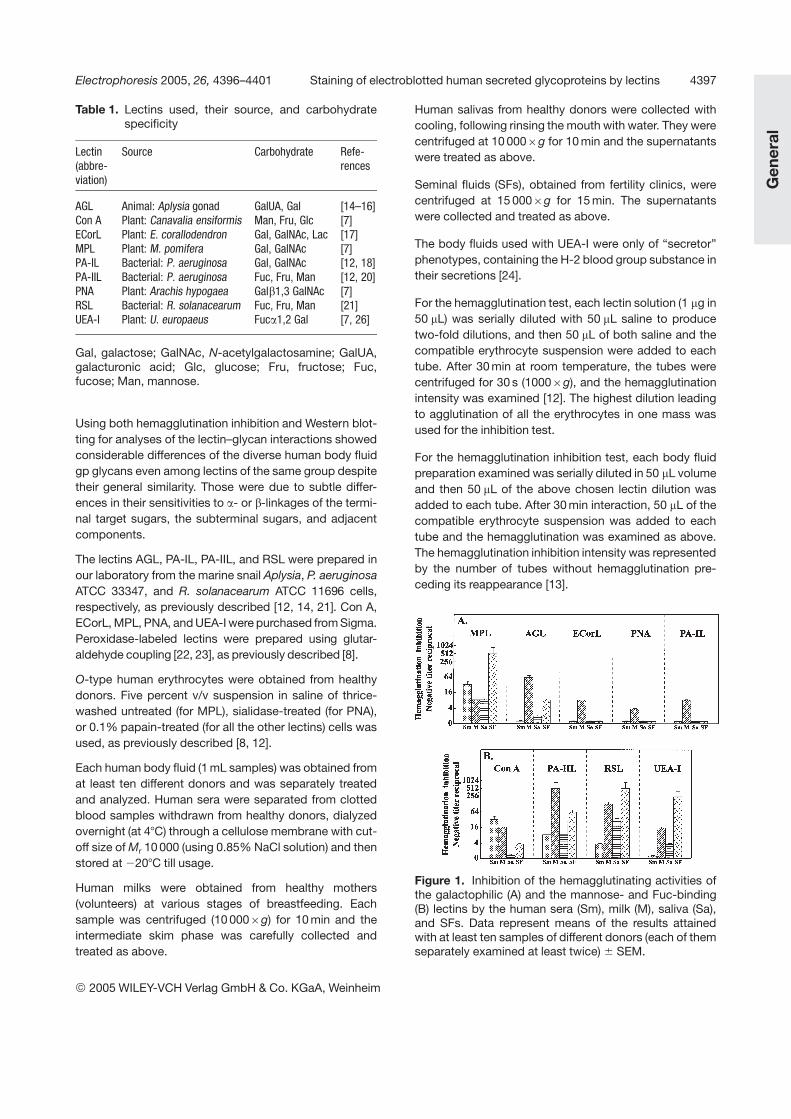
Figure 1. Inhibition of the hemagglutinating activities ofthe galactophilic (A) and the mannose- and Fuc-binding(B) lectins by the human sera (Sm), milk (M), saliva (Sa),and SFs. Data represent means of the results attainedwith at least ten samples of different donors (each of themseparately examined at least twice) 6 SEM.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Gen
eral
4398 N. Gilboa-Garber et al. Electrophoresis 2005, 26, 4396–4401
The results of the hemagglutination inhibition tests wereanalyzed by Student’s t-test.
For the SDS-PAGE and Western blot analyses, 15 mL ali-quots (containing around 40 mg protein) of each body fluidwere diluted 1:1 with sample buffer (0.1 M Tris-HCl,pH 6.8, containing 4% w/v SDS, 20% v/v glycerol, 0.2 MDTT, and 0.2% w/v bromophenol blue), boiled for 3 min,and applied to the wells in 10% SDS-PAGE. Parallel,similarly treated gels were used for protein detection byCBB staining and for the Western blotting with the spe-cific gp lectin tagging. The serum aliquot was reduced toevade its albumin disturbance. The electrophoresis wascarried out (at 140 V) in Mini Protean Cell 3 Electrophore-sis (BioRad) at room temperature and was stopped whenthe bromophenol line reached the gel bottom, according
to Laemmli [25]. Following the SDS-PAGE, one gel wasused for CBB staining, and from the other gels the pro-teins were transferred to NC (0.45 mm, BioRad) mem-branes at 47C for 2 h (85 mA/40–50 V), using the Mini-trans-Blot Module (BioRad). The membranes were incu-bated overnight in blocking buffer (0.01 M PBS, pH 7.2,containing 3% BSA and 0.05% Tween 20) for preventionof nonspecific antibody or lectin binding. Then they wereexposed to the peroxidase-labeled lectins (about 1 mg/mL) in blocking buffer (with 0.1% Tween 20) for 2 h atroom temperature. Following thorough washing, the per-oxidase reaction was visualized using ECL (AmershamInternational PLC, Buckinghamshire, UK) according to themanufacturer. The resulting luminescence was recordedonto photographic film. Several exposures were per-formed for each blot in order to gain greatest sensitivity.
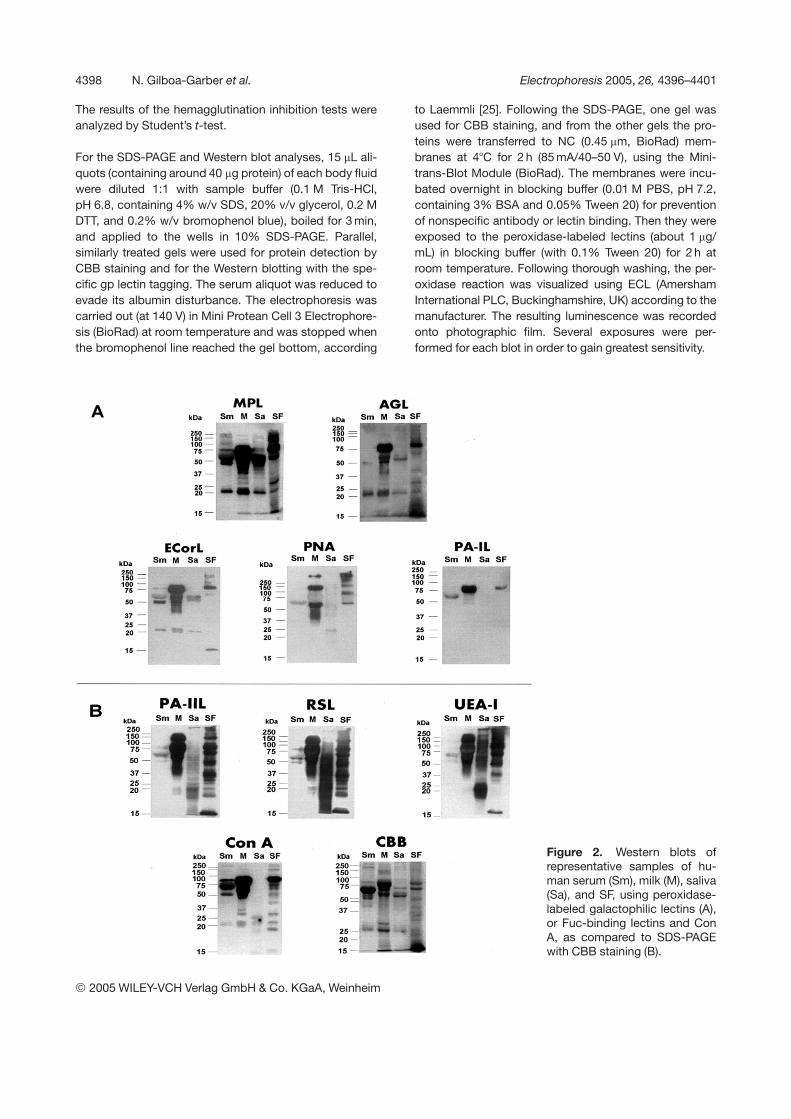
Figure 2. Western blots ofrepresentative samples of hu-man serum (Sm), milk (M), saliva(Sa), and SF, using peroxidase-labeled galactophilic lectins (A),or Fuc-binding lectins and ConA, as compared to SDS-PAGEwith CBB staining (B).
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

Electrophoresis 2005, 26, 4396–4401 Staining of electroblotted human secreted glycoproteins by lectins 4399
Controls with the labeled lectins in the presence of 0.3 Mof the relevant blocking sugars (Gal for AGL, ECorL, MPL,PA-IL, and PNA or Fuc for PA-IIL, RSL, and UEA-IL) intheir reaction mixtures were added to rule out possiblestaining of the electroblots by nonspecific (sugar-inde-pendent) lectin binding.
Lectin usage for investigation of human body fluids mayserve diverse, important scientific, and applicative pur-poses.
The broad spectrum of sugar specificities of the lectinsthat were chosen for the present research (Table 1) wasclearly revealed in the results of both the hemagglutinationinhibition (Fig. 1) and the Western blot (Fig. 2) analyses.
As seen in Fig. 1A, MPL was the only one among thegalactophilic lectins that was considerably inhibited by allthe four body fluids. Especially outstanding was its moststrong inhibition by the SF, followed by the serum, andconsiderable inhibition by saliva. Its inhibition by milk wassimilar to the latter. Second in intensity was AGL that wasquite sensitive to inhibition by milk, followed by SF, but
only weakly inhibited by saliva (64:8:2). All the othergalactophilic lectins, including ECorL, PNA, and PA-ILreacted with the milk almost like MPL but negligibly withthe other three body fluids.
Among the other four lectins (Fig. 1B), Con A (which bindsmannose, fructose, and glucose) was the only one thatwas inhibited most strongly by the serum, like MPL, fol-lowed by the milk, mildly by the SF, and negligibly by sal-iva. In contrast to it, the three Fuc/mannose-binding lec-tins were quite sensitive to inhibition by saliva (RSL morethan the others), but were most strongly inhibited by milkand SFs. PA-IIL more by the milk, RSL and UEA-I morewith the SF. As to their sensitivities to serum: PA-IIL dis-played similar sensitivity to serum and saliva, UEA-I wasnot sensitive to the serum glycans, while RSL was be-tween them. These variations in the three Fuc-bindinglectin preferential affinities could be attributed to thesugar linkage type (Fig. 3). PA-IIL most strongly binds toFuc a1,4 Gal of the Lea antigen [20] prevalent in the hu-man milk, and to Lex, which is also present on the lacto-ferrin glycan (Fig. 3), while UEA-I selectively binds to the
Figure 3. Proposed generalscheme of representative gly-can structures of some of theexamined gps (those of lacto-ferrin and immunoglobulin N-linked saccharide chains areaccording to Eda et al. [30] andKobata [31], respectively).
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
4400 N. Gilboa-Garber et al. Electrophoresis 2005, 26, 4396–4401
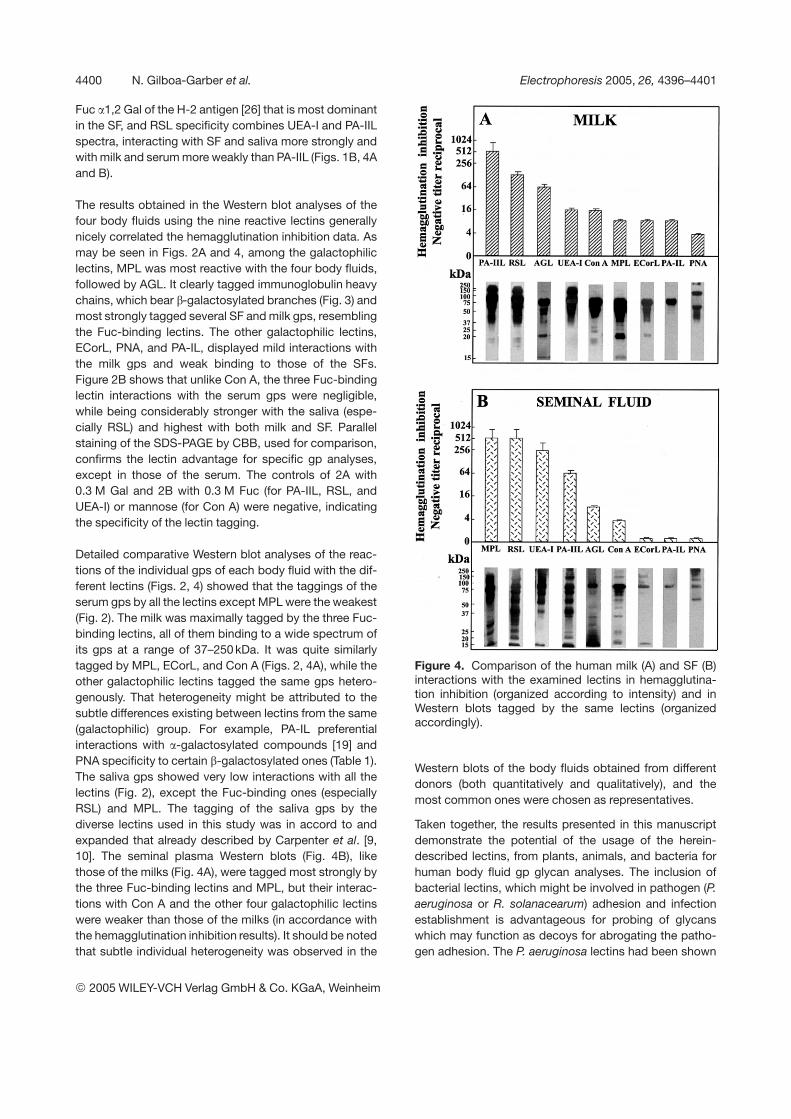
Fuc a1,2 Gal of the H-2 antigen [26] that is most dominantin the SF, and RSL specificity combines UEA-I and PA-IILspectra, interacting with SF and saliva more strongly andwith milk and serum more weakly than PA-IIL (Figs. 1B, 4Aand B).
The results obtained in the Western blot analyses of thefour body fluids using the nine reactive lectins generallynicely correlated the hemagglutination inhibition data. Asmay be seen in Figs. 2A and 4, among the galactophiliclectins, MPL was most reactive with the four body fluids,followed by AGL. It clearly tagged immunoglobulin heavychains, which bear b-galactosylated branches (Fig. 3) andmost strongly tagged several SF and milk gps, resemblingthe Fuc-binding lectins. The other galactophilic lectins,ECorL, PNA, and PA-IL, displayed mild interactions withthe milk gps and weak binding to those of the SFs.Figure 2B shows that unlike Con A, the three Fuc-bindinglectin interactions with the serum gps were negligible,while being considerably stronger with the saliva (espe-cially RSL) and highest with both milk and SF. Parallelstaining of the SDS-PAGE by CBB, used for comparison,confirms the lectin advantage for specific gp analyses,except in those of the serum. The controls of 2A with0.3 M Gal and 2B with 0.3 M Fuc (for PA-IIL, RSL, andUEA-I) or mannose (for Con A) were negative, indicatingthe specificity of the lectin tagging.
Detailed comparative Western blot analyses of the reac-tions of the individual gps of each body fluid with the dif-ferent lectins (Figs. 2, 4) showed that the taggings of theserum gps by all the lectins except MPL were the weakest(Fig. 2). The milk was maximally tagged by the three Fuc-binding lectins, all of them binding to a wide spectrum ofits gps at a range of 37–250 kDa. It was quite similarlytagged by MPL, ECorL, and Con A (Figs. 2, 4A), while theother galactophilic lectins tagged the same gps hetero-genously. That heterogeneity might be attributed to thesubtle differences existing between lectins from the same(galactophilic) group. For example, PA-IL preferentialinteractions with a-galactosylated compounds [19] andPNA specificity to certain b-galactosylated ones (Table 1).The saliva gps showed very low interactions with all thelectins (Fig. 2), except the Fuc-binding ones (especiallyRSL) and MPL. The tagging of the saliva gps by thediverse lectins used in this study was in accord to andexpanded that already described by Carpenter et al. [9,10]. The seminal plasma Western blots (Fig. 4B), likethose of the milks (Fig. 4A), were tagged most strongly bythe three Fuc-binding lectins and MPL, but their interac-tions with Con A and the other four galactophilic lectinswere weaker than those of the milks (in accordance withthe hemagglutination inhibition results). It should be notedthat subtle individual heterogeneity was observed in the
Figure 4. Comparison of the human milk (A) and SF (B)interactions with the examined lectins in hemagglutina-tion inhibition (organized according to intensity) and inWestern blots tagged by the same lectins (organizedaccordingly).
Western blots of the body fluids obtained from differentdonors (both quantitatively and qualitatively), and themost common ones were chosen as representatives.
Taken together, the results presented in this manuscriptdemonstrate the potential of the usage of the herein-described lectins, from plants, animals, and bacteria forhuman body fluid gp glycan analyses. The inclusion ofbacterial lectins, which might be involved in pathogen (P.aeruginosa or R. solanacearum) adhesion and infectionestablishment is advantageous for probing of glycanswhich may function as decoys for abrogating the patho-gen adhesion. The P. aeruginosa lectins had been shown
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

Electrophoresis 2005, 26, 4396–4401 Staining of electroblotted human secreted glycoproteins by lectins 4401
to bind to the human cell A, B, H, I, Le, and P1 antigenswhich are receptors for microbial homing [20, 27]. UEA-I[26], ECorL [28], and AGL [29] also bind to the human H,HI, and I antigens, respectively. The same lectin batteryand techniques may also be used for other scientific anddiagnostic purposes as well as for purification and qualitycontrol of the desired gps to be used for medical andindustrial aims.
The authors thank Avrille Goldreich, Sharon Victor, and ElaGindy for the skillful typing and graphic presentations ofthis manuscript. The study, which is part of B. Lerrer’s andE. Lesman-Movshovich’s Ph.D. theses, was supported bythe Israel Governmental Estate Property Funds.
Received May 26, 2005Revised July 31, 2005Accepted August 4, 2005
References
[1] Newburg, D. S., Curr. Med. Chem. 1999, 6, 117–127.[2] Newburg, D. S., Adv. Exp. Med. Biol. 2001, 501, 3–10.[3] Slomiany, B. L., Murty, V. L. N., Piotrowski, J., Slomiany, A.,
Gen. Pharmacol. 1996, 27, 761–771.[4] Solzbacher, D., Hanisch, F. G., van Alphen, L., Gilsdorf, J. R.,
Eur. Arch. Otorhinolaryngol. 2003, 260, 141–147.[5] Svanborg Eden, C., Andersson, B., Aniansson, R., Lindstedt,
R., et al., Curr. Top. Microbiol. Immunol. 1990, 151, 167–184.[6] Yamashita, K., Kamerling, J. P., Kobata, A., J. Biol. Chem.
1982, 257, 12809–12814.[7] Goldstein, I. J., Poretz, R. D., in: Liener, I. E., Sharon, N.,
Goldstein, I. J. (Eds.), The Lectins, Properties, Function andApplications in Biology and Medicine, Academic Press, NewYork 1986, pp. 33–247.
[8] Lerrer, B., Gilboa-Garber, N., Electrophoresis 2002, 23, 8–14.[9] Carpenter, G. H., Proctor, G. B., Pankhurst, C. L., Linden, R.
W., et al., Electrophoresis 1996, 17, 91–97.
[10] Carpenter, G. H., Proctor, G. B., Electrophoresis 2000, 21,1446–1453.
[11] Lesman-Movshovich, E., Lerrer, B., Gilboa-Garber, N., Can.J. Microbiol. 2003, 49, 230–235.
[12] Gilboa-Garber, N., Methods Enzymol. 1982, 83, 378–385.
[13] Lesman-Movshovich, E., Gilboa-Garber, N., J. Dairy Sci.2003, 86, 2276–2282.
[14] Gilboa-Garber, N., Susswein, A. J., Mizrahi, L., Avichezer,D., FEBS Lett. 1985, 181, 267–270.
[15] Wu, A. M., Song, S.-C., Chen, Y.-Y., Gilboa-Garber, N., J.Biol. Chem. 2000, 275, 14017–14024.
[16] Gilboa-Garber, N., Wu, A. M., Adv. Exp. Med. Biol. 2001,491, 109–126.
[17] Gilboa-Garber, N., Mizrahi, L., Can. J. Biochem. 1981, 59,315–320.
[18] Ciocci, G., Mitchell, E. P., Cautier, C., Wimmerova, M., et al.,FEBS Lett. 2003, 555, 297–301.
[19] Chen, C.-P., Song, S. C., Gilboa-Garber, N., Chang, K. S. S.,Wu, A. M., Glycobiology 1998, 8, 7–16.
[20] Mitchell, E., Houles, C., Sudakevitz, D., Wimmerova, M., etal., Nature Struct. Biol. 2002, 9, 918–921.
[21] Sudakevitz, D., Imberty, A., Gilboa-Garber, N., J. Biochem.2002, 132, 353–358.
[22] Avrameas, S., Immunochemistry 1969, 6, 43–52.
[23] Huet, C. H., Garrido, J., Exp. Cell Res. 1972, 75, 523–527.
[24] Race, R. R., Sanger, R., Blood Groups in Man, BlackwellScientific Publications, Oxford 1975.
[25] Laemmli, U. K., Nature 1970, 227, 680–685.
[26] Hindsgaul, O., Khare, D. P., Bach, M., Lemieux, R. U., Can.J. Chem. 1985, 63, 2653–2658.
[27] Gilboa-Garber, N., Sudakevitz, D., Sheffi, M., Sela, R., et al.,Glycoconj. J. 1994, 11, 414–417.
[28] Sudakevitz, D., Gilboa-Garber, N., Levene, C., Sela, R., etal., Zentralbl. Bakteriol. 1991, 275, 343–350.
[29] Gilboa-Garber, N., Sudakevitz, D., Levene, C., Transfusion1999, 39, 1060–1064.
[30] Eda, S., Kikugawa, K., Beppu, M., Biol. Pharm. Bull. 1997,20, 127–133.
[31] Kobata, A., Glycoconj. J. 1998, 15, 323–331.
2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim





























