Embed Size (px)
Citation preview

ORIGINAL PAPER
Differential responses to drought stress in leaves and roots of wildjujube, Ziziphus lotus
Mouna Maraghni • Mustapha Gorai •
Mohamed Neffati • Marie Christine Van Labeke
Received: 19 March 2013 / Revised: 13 December 2013 / Accepted: 16 December 2013
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2013
Abstract The aim of this study was to investigate the
effects of drought stress induced by polyethylene glycol-
6000 in wild jujube, Ziziphus lotus. One-month-old,
hydroponically grown seedlings were subjected to three
treatments, i.e. normal watering (-0.2 MPa), moderate
(-1.2 MPa) and severe (-2.1 MPa) drought stress for
14 days under controlled climatic conditions. Plant growth
was markedly reduced with increasing osmotic stress. The
shoot water potential (Ww) and leaf relative water content
followed similar patterns and significantly decreased with
increasing osmolality of solutions. As a consequence of
drought, contents in proline and soluble sugars were found
to be more elevated in leaves than in roots. The level of
lipid peroxidation in terms of malonyldialdehyde contents
increased in both leaves and roots of drought-stressed
plants. Wild jujube displayed higher activities of antioxi-
dant enzymes in the roots than in the leaves. Catalase and
guaiacol peroxidase activities increased significantly in
drought-stressed roots, whereas ascorbate peroxidase
activity showed a slight decline with no significant
changes. These findings suggest that Z. lotus was able to
adapt to severe drought stress by accumulation of com-
patible solutes and by activation of free radical-scavenging
enzymes. Overall, defence mechanisms in Z. lotus against
oxidative stress are organized differently in plant tissues,
with higher solute accumulation in leaves and increased
activity of antioxidants in roots, during drought stress.
Keywords Ziziphus lotus � Drought � Growth � Water
relations � Osmotic adjustment � Oxidative stress
Abbreviations
APX Ascorbate peroxidase
CAT Catalase
DM Dry mass
MDA Malondialdehyde
FM Fresh mass
PEG Polyethylene glycol
POD Guaiacol peroxidase
RWC Relative water content
Ww Water potential
TM Turgid mass
Introduction
Ziziphus species are sclerophyllous evergreen trees or
shrubs with a high degree of drought tolerance. Several
reports have investigated their physiological and morpho-
logical adaptations to water deficit stress (e.g. Clifford
et al. 1998; Arndt et al. 2001; Kulkarni et al. 2010; Mar-
aghni et al. 2011). The ability of Ziziphus species to survive
drought has been attributed to a combination of avoidance
Communicated by G. Bartosz.
M. Maraghni � M. Neffati
Laboratoire d’Ecologie Pastorale, Institut des Regions Arides,
4119 Medenine, Tunisia
M. Gorai (&)
Departement des Sciences de l’Environnement, Institut
Superieur de Biologie Appliquee de Medenine,
4119 Medenine, Tunisia
e-mail: [email protected]
M. C. Van Labeke
Department of Plant Production, Faculty of Bioscience
Engineering, Ghent University, Coupure links 653,
9000 Ghent, Belgium
123
Acta Physiol Plant
DOI 10.1007/s11738-013-1473-9

and tolerance mechanisms, including osmotic adjustment
and stomatal control (Arndt et al. 2001; Pareek 2001;
Maraghni et al. 2011). Z. lotus is a native species of the
Mediterranean region and indigenous to Tunisia where it
has a wide ecological and geographical distribution and
grows under a variety of environmental conditions (Gorai
et al. 2010). It reaches 2–5 m, is dormant from October
through March, flowers in May and June, and produces
fruits in August (Gorai et al. 2010). The fruits, subglobose
drupes (c. 1–1.5 cm in diameter), are the edible part of the
plant (Maraghni et al. 2010). This spiny shrub is of great
economic and ecological interest: it stabilizes sand dunes,
and has food, feed, fuel and medicinal uses (Le Floc’h
1983; Borgi et al. 2008; Borgi and Chouchane 2009; Gorai
et al. 2010; Maraghni et al. 2010).
Drought is the major factor limiting crop production
worldwide (Ludlow and Muchow 1990). Therefore, phys-
iological and biochemical adaptations involved in cellular
and whole plant responses to drought generate considerable
interest. A very common response to drought is an
increased production of compatible organic solutes such as
soluble sugars, sugar alcohols, proline and betaines (Serraj
and Sinclair 2002). These compatible solutes contribute to
cellular osmotic adjustment, detoxification of reactive
oxygen species, protection of membrane integrity, and
stabilization of enzymes/proteins (Bohnert and Jenson
1996; Attipali et al. 2004).
Drought stress promotes the production of reactive
oxygen species (ROS), such as superoxide (�O2-), singlet
oxygen (O2� ), hydrogen peroxide (H2O2), and hydroxyl
radicals (OH�-) in plants. In higher plants, ROS are con-
tinuously produced in chloroplasts, mitochondria and per-
oxisomes (Apel and Hirt 2004). However, H2O2 produced
in peroxisomes and chloroplasts diffuses to the cytosol and
is converted to hydroxyl radicals by the Fenton reaction
(Moller et al. 2007). Overproduction of ROS in plant cells
under stress can damage cellular components, including
DNA, proteins and membrane lipids, and leads to destruc-
tion of photosynthetic pigments (Yordanov et al. 2000;
Mittler 2002). Different abiotic stress factors may induce
oxidative stress in plants, which lead to similar cellular
adaptive responses such as accumulation of compatible
solutes, induction of stress proteins and acceleration of
ROS-scavenging systems (Zhu 2002). To scavenge reactive
oxygen species, plants have evolved both enzymatic and
non-enzymatic defence systems (Ingram and Bartels 1996).
The non-enzymatic systems include b-carotenes, a-
tocopherol, ascorbate, glutathione and the enzymatic anti-
oxidants including superoxide dismutase (SOD), catalase
(CAT), ascorbate peroxidase (APX), guaiacol peroxidase
(POD) and glutathione reductase (GR) (Sofo et al. 2007).
ROS is converted by SOD into H2O2, which is further
scavenged by CAT and various peroxidases (APX, POD) to
H2O (Noctor and Foyer 1998). Oxidative stress increases
lipid peroxidation, forming adducts to deoxyguanosine and
deoxyadenosine (Marnett 1999). Under drought conditions
the behaviour of various shrubs indicated that activities of
antioxidant enzymes are correlated with their tolerance to
abiotic stress (Dichio et al. 2002; Sofo et al. 2004; Guo et al.
2006). Plant physiological processes, such as osmotic
adjustment and activities of antioxidant enzymes, depend
on the rapidity, severity and duration of the drought
(DaCosta and Huang 2007).
Involvement of antioxidant defence mechanisms against
water deficit stress was not studied in Z. lotus. Hence, this
study is the first report in this respect and aims to under-
stand the physiological and biochemical processes under-
lying differential tolerance to drought. We evaluated the
changes in growth parameters, water relations, lipid
membrane peroxidation (MDA), osmotic adjustment (pro-
line and soluble carbohydrates), and activity of three
antioxidant enzymes: CAT, APX, and POD.
Materials and methods
Plant material and drought stress applications
The experiments were carried out at the Faculty of Bio-
science Engineering—Ghent University, Ghent, Belgium.
Mature fruits were collected in September 2009 from
plants in natural Z. lotus populations growing at Samaaliate
(33�170N, 10�550E; Ben Guerdane, Southeast Tunisia).
Seeds were provided by the seed bank of the Laboratoire
d’Ecologie Pastorale at the Institut des Regions Arides,
Medenine, Tunisia. Seeds were disinfected in an aqueous
solution of calcium hypochlorite 5 % for 5 min, sown in
alveolar plates filled with loam and placed at controlled
conditions (Maraghni et al. 2010). When the cotyledons
were exposed, seedlings were transplanted in PVC pot
(8 9 7 cm). The temperature was maintained at 25/30 �C
(night/day), the level of photosynthetically active radiation
was 250 lmol m-2 s-1 and the day time humidity was
between 60 and 70 %. The plants were hydroponically
grown with standard Hewitt’s (1966) nutrient solution
using a circulating system (Fig. 1). This nutrient solution
contains following macronutrients: 1.5 mM MgSO4,
1.6 mM KH2PO4, 0.4 mM K2HPO4, 3 mM KNO3, 2 mM
NH4NO3, 3.5 mM Ca (NO3)2, and micronutrients: 8 lM
MnCl2, 0.7 lM CuSO4 5H2O, 0.76 lM ZnSO4 7H2O,
0.3 lM Mo7O24(NH4)6, 46 lM H3BO3 (Arnon and Hoa-
gland 1940) and 45 lM EDTA-Fe (Jacobson 1951). The
pH of the nutrient solution was daily adjusted to 6. Each
reservoir feeds two channels by pipes in which each
channel has 20 seedlings and was equipped with two tubes
of bubbling air, allowing good aeration.
Acta Physiol Plant
123

The experiment was arranged in a growth chamber in a
completely randomized design with three levels of drought
treatment 9 40 replicates. One-month-old seedlings were
supplied with PEG solutions at varying water potentials of
-0.2 MPa (control), -1.2 MPa (moderate stress) and
-2.1 MPa (severe stress) for 14 days. The concentration of
PEG-6000 in the nutrient solution needed to obtain the
respective drought stress levels was determined following
Michel and Kaufmann (1973) and adjusted using a Wescor
vapour pressure osmometer (model C-52 5500, Logan, UT,
USA).
Growth parameters
After 2 weeks of culture under different drought treatments,
shoots and roots were separated. Number of leaves and fresh
weights of shoots and roots were determined. Leaves and
roots were frozen in liquid nitrogen and blended using a
mortar and pestle previously chilled with liquid nitrogen.
The frozen powder was stored at -80 �C until analysis.
Plant water relations
Plant water status was evaluated by measuring water
potential (Ww) using a Scholander pressure chamber (PMS
Instruments co., Corvallis, R, USA) and relative water
content RWC = (FM - DM)/(TM - DM) 9 100, where
FM, DM and TM are the fresh, dry and turgid masses,
respectively. Wild jujube leaves were weighed immedi-
ately on harvest to determine FM. Turgid mass was
determined after leaves had been immersed for 24 h in
distilled water. After the determination of TM, leaves were
oven-dried at 80 �C for 48 h and DM was determined.
Determination of contents in proline, soluble
carbohydrates and starch
Free proline content was quantified as described by Bates
et al. (1973). Root and leaf samples (0.5 g) from each group
were homogenized in 3 % (w/v) sulphosalicylic acid and
homogenate was filtered through filter paper. After addition
of acid ninhydrin and glacial acetic acid, resulting mixture
was heated at 100 �C for 1 h in water bath. Reaction was then
stopped in ice. The mixture was extracted with toluene, and
the absorbance was read at 520 nm. Proline concentration
was determined using a calibration curve and expressed as
lmol proline g-1 FM.
Sugars from leaves and roots were extracted with 80 %
ethanol at 70 �C, and centrifuged at 5,0009g for 10 min.
Glucose, fructose and sucrose were analysed using high-pH
anion-exchange chromatography with pulsed amperometric
detection (Waters; CarboPac MA1 column with companion
guard column, eluent: 50 mM NaOH, 22 �C). The residual
ethanol insoluble material was washed twice with ethanol
80 % and the residue was treated with HCl 1 M for 2 h at
95 �C for starch hydrolysis. Starch was determined spec-
trophotometrically at 340 nm by the enzymatic reduction
of NADP?.
Extraction of malondialdehyde
The degree of lipid peroxidation was used to calculate
membrane integrity. Malondialdehyde (MDA) content was
measured as described by Hudges et al. (1999) with some
adaptations. 0.5 g aliquot of frozen powder was added to
1 ml of 0.1 % (w/v) trichloroacetic acid (TCA). The
homogenate was centrifuged at 3,0009g for 5 min. A 1-ml
aliquot of supernatant was added to 4.0 ml of 0.5 % (w/v)
thiobarbituric acid (TBA) in 20 % (w/v) trichloroacetic acid.
The mixture was heated at 100 �C for 30 min and then
quickly cooled in ice. After centrifugation at 3,0009g for
10 min, the absorbance of the supernatant at 600, 532 and
440 nm were determined. MDA content was calculated
using an absorption coefficient, e = 157 mM-1 cm-1 and
expressed as nmol MDA g-1 FM.
Enzyme extractions and assays
For protein and enzyme extractions, 0.5 g of leaf and root
samples was homogenized with 50 mM potassium phos-
phate buffer (pH 7.8) containing 1 mM EDTA-2Na and 7 %
(w/v) polyvinylpolypyrrolidone (PVPP). The whole extrac-
tion procedure was carried out at 4 �C. Homogenates were
Fig. 1 Experimental setup to study the response of Z. lotus seedlings
to water deficit induced by PEG at varying water potentials (-0.2,
-1.2 or -2.1 MPa) in nutrient solution under controlled cli-
matic conditions, a and b
Acta Physiol Plant
123
then centrifuged at 4 �C for 15 min at 13,0009g, and
supernatants were used for determination of enzyme activity.
Protein concentration was determined according to Bradford
(1976), using bovine serum albumin as a standard.
Catalase (CAT) activity (EC 1.11.1.6) was determined
using the method of Aebi (1984) with slight modification
by measuring the rate of decomposition of H2O2
(e = 2.3 mM-1 cm-1) at 240 nm. This activity was
assayed in a reaction mixture containing 1,900 ll of
potassium phosphate buffer (50 mM, pH 7.0 not containing
EDTA), 100 ll sample and 1,000 ll H2O2 (30 mM). CAT
activity is expressed as lmol H2O2 decomposed
min-1 mg-1 proteins.
Ascorbate peroxidase (APX) activity (EC 1.11.1.11)
was quantified as described by Nakano and Asada (1981).
The reaction mixture contained 50 mM of potassium
phosphate buffer (pH 7.0), 4.4 ll ascorbate (1 mM) and
10 ll EDTA-2Na (0.5 M). Adding H2O2 started the reac-
tion and the decrease in absorbance at 290 nm. Activity
was calculated using the extinction coefficient,
e = 2.8 mM-1 cm-1. Each sample was measured in trip-
licate. APX activity is expressed as lmol oxidized ascor-
bate min-1 mg-1 proteins.
Guaiacol peroxidase (POD) activity (EC 1.11.1.7) was
assayed by the method of Chance and Maehly (1955). The
reaction mixture with a final volume 1 ml contained 700 ll
of 0.05 M phosphate buffer (pH 7.8), 100 ll of plant extract
and 200 ll of guaiacol (25 mM). To start the reaction, 100 ll
of H2O2 was added. The increase of absorbance due to oxi-
dation of guaiacol to tetraguaiacol was monitored for 3 min
at 436 nm. Activity was calculated using the extinction
coefficient, e = 25.5 mM-1 cm-1. POD activity is expres-
sed as lmol oxidized guaiacol min-1 mg-1 proteins.
Statistical analysis
Data were analysed using SPSS statistical software version
17.0. All measurements were subjected to analysis of
variance (ANOVA) and differences among treatments were
compared by Tukey’s HSD test at the level of significance
P = 0.05.
Results
Growth attributes
Increasing drought stress, induced by PEG, significantly
decreased the growth attributes of Z. lotus seedlings
(Table 1; P \ 0.001). The number of leaves decreased
significantly by 54 and 75 % as compared to the controls
for moderate and severe stress, respectively. Fresh mass
was considerably reduced under severe drought stress in
both shoots and roots (ca. 76 and 69 % of the control value,
respectively).
Water relations
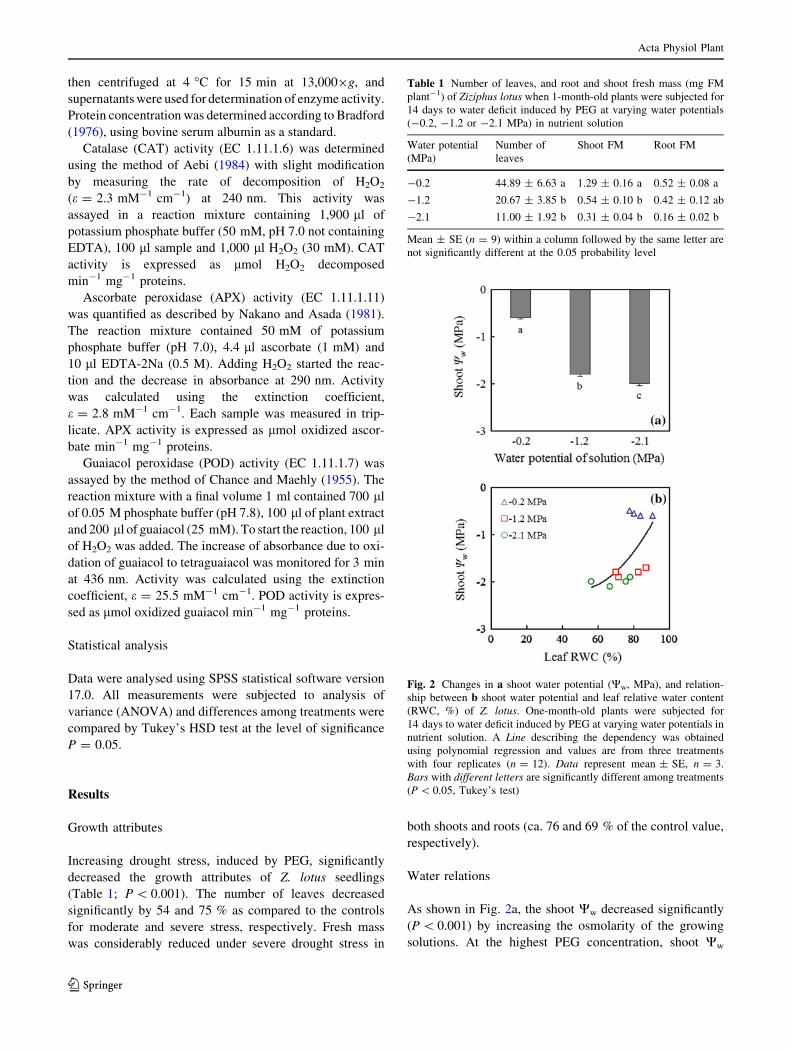
As shown in Fig. 2a, the shoot Ww decreased significantly
(P \ 0.001) by increasing the osmolarity of the growing
solutions. At the highest PEG concentration, shoot Ww
Table 1 Number of leaves, and root and shoot fresh mass (mg FM
plant-1) of Ziziphus lotus when 1-month-old plants were subjected for
14 days to water deficit induced by PEG at varying water potentials
(-0.2, -1.2 or -2.1 MPa) in nutrient solution
Water potential
(MPa)
Number of
leaves
Shoot FM Root FM
-0.2 44.89 ± 6.63 a 1.29 ± 0.16 a 0.52 ± 0.08 a
-1.2 20.67 ± 3.85 b 0.54 ± 0.10 b 0.42 ± 0.12 ab
-2.1 11.00 ± 1.92 b 0.31 ± 0.04 b 0.16 ± 0.02 b
Mean ± SE (n = 9) within a column followed by the same letter are
not significantly different at the 0.05 probability level
Fig. 2 Changes in a shoot water potential (Ww, MPa), and relation-
ship between b shoot water potential and leaf relative water content
(RWC, %) of Z. lotus. One-month-old plants were subjected for
14 days to water deficit induced by PEG at varying water potentials in
nutrient solution. A Line describing the dependency was obtained
using polynomial regression and values are from three treatments
with four replicates (n = 12). Data represent mean ± SE, n = 3.
Bars with different letters are significantly different among treatments
(P \ 0.05, Tukey’s test)
Acta Physiol Plant
123
dropped to -2 MPa. RWC had a decreasing trend as Ww of
growth solutions dropped. This parameter decreased from
83 % for control leaves to 78 and 69 % for drought-stres-
sed leaves under moderate and severe stress, respectively.
The relationship between shoot Ww and the leaf RWC
changed significantly in drought-stressed plants versus
control plants (Fig. 2b).
Solute accumulation
Ziziphus lotus seedlings in the presence of PEG-enriched
nutrient solution accumulated higher amounts of proline in
the leaves than in the roots (Fig. 3, P \ 0.001). When
subjected for 14 days to moderate and severe water deficit
stress, the leaves accumulated 12- and 19-fold, respec-
tively, more proline than controls.
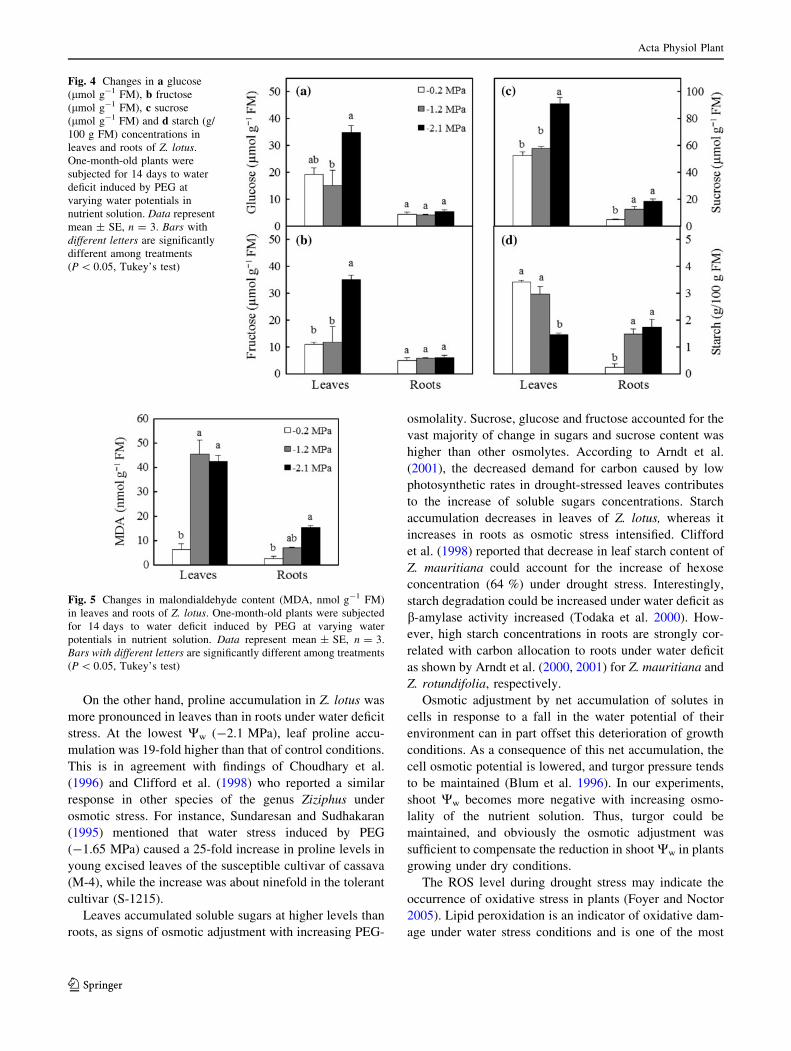
Plants accumulated higher soluble sugars (glucose,
fructose and sucrose) contents in leaves than in roots
(Fig. 4a–c). On day 14 of severe drought stress, glucose
and fructose contents in leaves changed significantly
compared to the controls; however, those in roots remained
unchanged and were not statistically significant. At low
water potential of the growth medium (-2.1 MPa), the
sucrose fraction increased either in leaves or roots (ca. 173
and 400 % of the control value, respectively). Starch
content changed significantly in 14-day drought-stressed
plants at Ww = -2.1 MPa of nutrient solution. This con-
tent increased in roots but decreased in leaves (Fig. 4d).
Lipid peroxidation
As seen in Fig. 5, lipid peroxidation, measured as MDA
content, increased significantly in the leaves and roots of
treated plants versus the controls. At 14 days of severe
drought stress, MDA content was increased by six and
sevenfold in roots and leaves, respectively, as compared to
the controls.
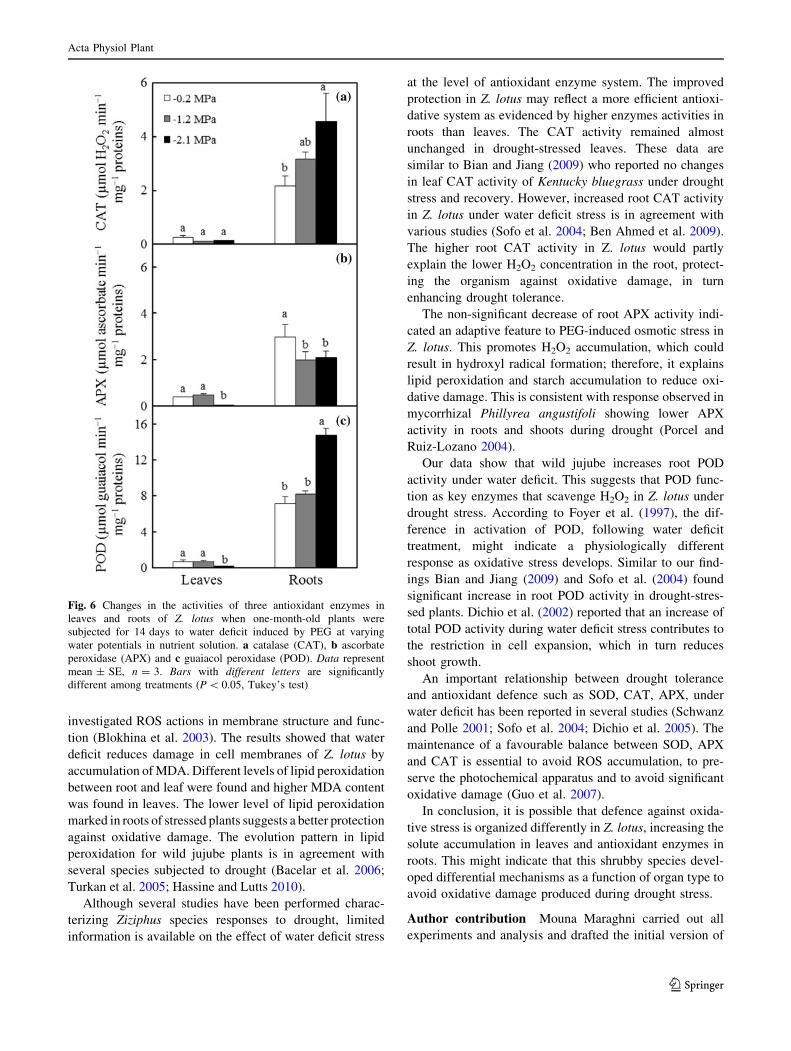
Enzyme activities
The activity of catalase was higher in roots than in leaves,
whatever the PEG dose in the nutrient solutions. The
increase of PEG-osmolarity enhanced the activity of CAT
in roots, while no significant changes were observed in the
leaves (Fig. 6a). CAT activity was increased 1.5- and 2.1-
fold at water potentials of -1.2 and -2.1 MPa in nutrient
solution, respectively, as compared to controls.
Most changes in APX activity for drought-stressed roots
versus the controls were not statistically significant
(Fig. 6b). At all water potentials, APX activity in leaves
was lower than in roots. APX activity remained unchanged
under moderate drought (-1.2 MPa), whereas it was sig-
nificantly reduced in both organs under severe drought
(-2.1 MPa), as compared to controls.
High values of POD activity were recorded in the roots.
At water potential of -2.1 MPa of nutrient solution, POD
activity was induced significantly in roots (2.1-fold), while
it was inhibited significantly in leaves (4.7-fold), as com-
pared to the controls (Fig. 6c).
Discussion
The present study shows that the osmotic potential of the
solution induced by PEG reduced the growth of Z. lotus by
restricting leaf formation and biomass production. This
response has been reported in several species subjected to
drought, like Z. mauritiana (Clifford et al. 1998), Z. ro-
tundifolia (Arndt et al. 2001), Olea europaea (Bacelar et al.
2007) and Adansonia digitata (De Smedt et al. 2012). The
drought-induced growth reduction might be related to
accelerated senescence of older leaves and reduction of
their size. For instance, Larcher (2003) showed that small
leaves are a characteristic for plants growing in dry envi-
ronments. Several authors have reported the strong impli-
cation of the root elongation and the ability to shed leaves
on Ziziphus species in extremely hot and arid environments
(Sankhla 1998; Jones 1999). Ziziphus species are typical
phreatophytes and behaves as arido-active species (Gorai
et al. 2010) throughout dry seasons.
Our data show a significant decrease in leaf RWC and
shoot Ww with increasing osmolality of solutions. These
changes may also result in stomatal closure, reduction of
CO2/O2 ratio in leaves and CO2 assimilation (Gindaba
et al. 2004; Boussadia et al. 2008). Maintaining leaf RWC
under lower shoot Ww is an important adaptation exhibited
by Z. lotus to cope with extreme drought conditions. The
present data agree with findings reported on Z. mauritiana
(Clifford et al. 1998), Z. rotundifolia (Arndt et al. 2001)
and Z. mauritiana (Kulkarni et al. 2010).
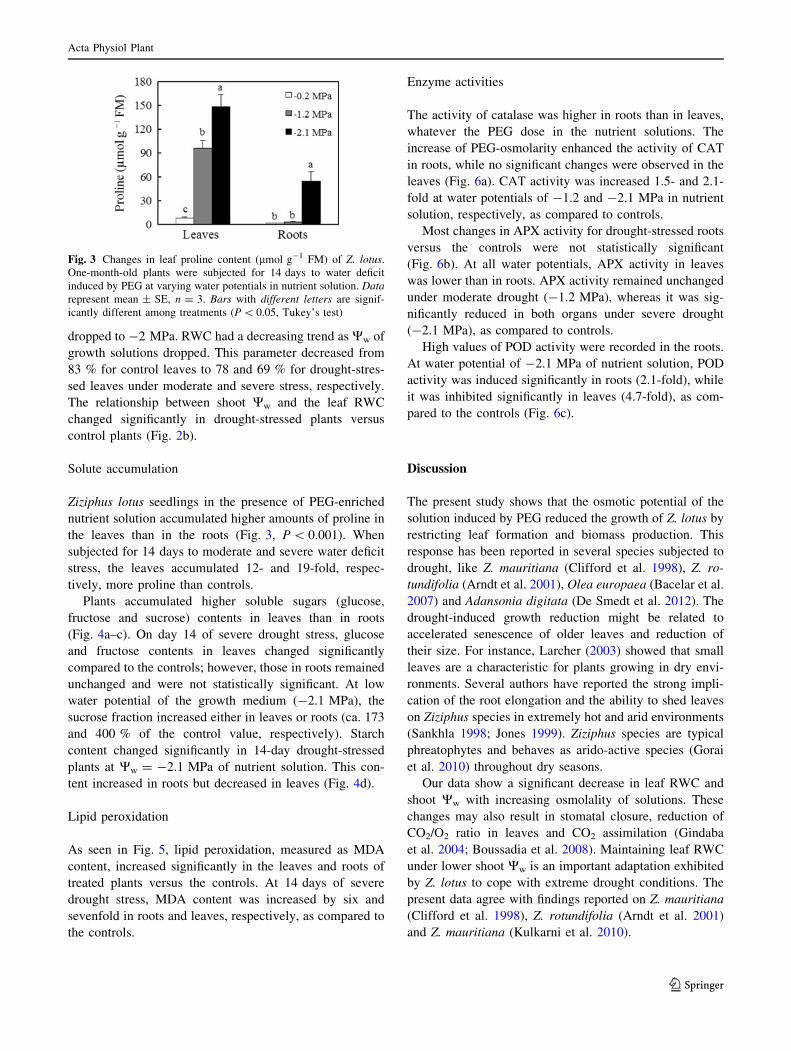
Fig. 3 Changes in leaf proline content (lmol g-1 FM) of Z. lotus.
One-month-old plants were subjected for 14 days to water deficit
induced by PEG at varying water potentials in nutrient solution. Data
represent mean ± SE, n = 3. Bars with different letters are signif-
icantly different among treatments (P \ 0.05, Tukey’s test)
Acta Physiol Plant
123
On the other hand, proline accumulation in Z. lotus was
more pronounced in leaves than in roots under water deficit
stress. At the lowest Ww (-2.1 MPa), leaf proline accu-
mulation was 19-fold higher than that of control conditions.
This is in agreement with findings of Choudhary et al.
(1996) and Clifford et al. (1998) who reported a similar
response in other species of the genus Ziziphus under
osmotic stress. For instance, Sundaresan and Sudhakaran
(1995) mentioned that water stress induced by PEG
(-1.65 MPa) caused a 25-fold increase in proline levels in
young excised leaves of the susceptible cultivar of cassava
(M-4), while the increase was about ninefold in the tolerant
cultivar (S-1215).
Leaves accumulated soluble sugars at higher levels than
roots, as signs of osmotic adjustment with increasing PEG-
osmolality. Sucrose, glucose and fructose accounted for the
vast majority of change in sugars and sucrose content was
higher than other osmolytes. According to Arndt et al.
(2001), the decreased demand for carbon caused by low
photosynthetic rates in drought-stressed leaves contributes
to the increase of soluble sugars concentrations. Starch
accumulation decreases in leaves of Z. lotus, whereas it
increases in roots as osmotic stress intensified. Clifford
et al. (1998) reported that decrease in leaf starch content of
Z. mauritiana could account for the increase of hexose
concentration (64 %) under drought stress. Interestingly,
starch degradation could be increased under water deficit as
b-amylase activity increased (Todaka et al. 2000). How-
ever, high starch concentrations in roots are strongly cor-
related with carbon allocation to roots under water deficit
as shown by Arndt et al. (2000, 2001) for Z. mauritiana and
Z. rotundifolia, respectively.
Osmotic adjustment by net accumulation of solutes in
cells in response to a fall in the water potential of their
environment can in part offset this deterioration of growth
conditions. As a consequence of this net accumulation, the
cell osmotic potential is lowered, and turgor pressure tends
to be maintained (Blum et al. 1996). In our experiments,
shoot Ww becomes more negative with increasing osmo-
lality of the nutrient solution. Thus, turgor could be
maintained, and obviously the osmotic adjustment was
sufficient to compensate the reduction in shoot Ww in plants
growing under dry conditions.
The ROS level during drought stress may indicate the
occurrence of oxidative stress in plants (Foyer and Noctor
2005). Lipid peroxidation is an indicator of oxidative dam-
age under water stress conditions and is one of the most
Fig. 4 Changes in a glucose
(lmol g-1 FM), b fructose
(lmol g-1 FM), c sucrose
(lmol g-1 FM) and d starch (g/
100 g FM) concentrations in
leaves and roots of Z. lotus.
One-month-old plants were
subjected for 14 days to water
deficit induced by PEG at
varying water potentials in
nutrient solution. Data represent
mean ± SE, n = 3. Bars with
different letters are significantly
different among treatments
(P \ 0.05, Tukey’s test)
Fig. 5 Changes in malondialdehyde content (MDA, nmol g-1 FM)
in leaves and roots of Z. lotus. One-month-old plants were subjected
for 14 days to water deficit induced by PEG at varying water
potentials in nutrient solution. Data represent mean ± SE, n = 3.
Bars with different letters are significantly different among treatments
(P \ 0.05, Tukey’s test)
Acta Physiol Plant
123
investigated ROS actions in membrane structure and func-
tion (Blokhina et al. 2003). The results showed that water
deficit reduces damage in cell membranes of Z. lotus by
accumulation of MDA. Different levels of lipid peroxidation
between root and leaf were found and higher MDA content
was found in leaves. The lower level of lipid peroxidation
marked in roots of stressed plants suggests a better protection
against oxidative damage. The evolution pattern in lipid
peroxidation for wild jujube plants is in agreement with
several species subjected to drought (Bacelar et al. 2006;
Turkan et al. 2005; Hassine and Lutts 2010).
Although several studies have been performed charac-
terizing Ziziphus species responses to drought, limited
information is available on the effect of water deficit stress
at the level of antioxidant enzyme system. The improved
protection in Z. lotus may reflect a more efficient antioxi-
dative system as evidenced by higher enzymes activities in
roots than leaves. The CAT activity remained almost
unchanged in drought-stressed leaves. These data are
similar to Bian and Jiang (2009) who reported no changes
in leaf CAT activity of Kentucky bluegrass under drought
stress and recovery. However, increased root CAT activity
in Z. lotus under water deficit stress is in agreement with
various studies (Sofo et al. 2004; Ben Ahmed et al. 2009).
The higher root CAT activity in Z. lotus would partly
explain the lower H2O2 concentration in the root, protect-
ing the organism against oxidative damage, in turn
enhancing drought tolerance.
The non-significant decrease of root APX activity indi-
cated an adaptive feature to PEG-induced osmotic stress in
Z. lotus. This promotes H2O2 accumulation, which could
result in hydroxyl radical formation; therefore, it explains
lipid peroxidation and starch accumulation to reduce oxi-
dative damage. This is consistent with response observed in
mycorrhizal Phillyrea angustifoli showing lower APX
activity in roots and shoots during drought (Porcel and
Ruiz-Lozano 2004).
Our data show that wild jujube increases root POD
activity under water deficit. This suggests that POD func-
tion as key enzymes that scavenge H2O2 in Z. lotus under
drought stress. According to Foyer et al. (1997), the dif-
ference in activation of POD, following water deficit
treatment, might indicate a physiologically different
response as oxidative stress develops. Similar to our find-
ings Bian and Jiang (2009) and Sofo et al. (2004) found
significant increase in root POD activity in drought-stres-
sed plants. Dichio et al. (2002) reported that an increase of
total POD activity during water deficit stress contributes to
the restriction in cell expansion, which in turn reduces
shoot growth.
An important relationship between drought tolerance
and antioxidant defence such as SOD, CAT, APX, under
water deficit has been reported in several studies (Schwanz
and Polle 2001; Sofo et al. 2004; Dichio et al. 2005). The
maintenance of a favourable balance between SOD, APX
and CAT is essential to avoid ROS accumulation, to pre-
serve the photochemical apparatus and to avoid significant
oxidative damage (Guo et al. 2007).
In conclusion, it is possible that defence against oxida-
tive stress is organized differently in Z. lotus, increasing the
solute accumulation in leaves and antioxidant enzymes in
roots. This might indicate that this shrubby species devel-
oped differential mechanisms as a function of organ type to
avoid oxidative damage produced during drought stress.
Author contribution Mouna Maraghni carried out all
experiments and analysis and drafted the initial version of
Fig. 6 Changes in the activities of three antioxidant enzymes in
leaves and roots of Z. lotus when one-month-old plants were
subjected for 14 days to water deficit induced by PEG at varying
water potentials in nutrient solution. a catalase (CAT), b ascorbate
peroxidase (APX) and c guaiacol peroxidase (POD). Data represent
mean ± SE, n = 3. Bars with different letters are significantly
different among treatments (P \ 0.05, Tukey’s test)
Acta Physiol Plant
123

the manuscript. Mustapha Gorai wrote part of the text,
prepared figures and was responsible for verification of
paper during the online submission. Mohamed Neffati
coordinated the study. Marie-Christine Van Labeke con-
tributed to the implementation of the study and manuscript
preparation.
References
Aebi H (1984) Catalase: in vitro. In: Colowick SP, Kaplan NO (eds)
Methods in Enzymology, Vol 105. Academic Press, New York,
pp 121–126
Apel K, Hirt H (2004) Reactive oxygen species: metabolism,
oxidative stress, and signal transduction. Annu Rev Plant Biol
55:373–399
Arndt SK, Wanek W, Clifford SC, Popp M (2000) Contrasting
adaptations to drought stress in field-grown Ziziphus mauritiana
and Prunus persica trees: water relations, osmotic adjustment and
carbon isotope composition. Aust J Plant Physiol 27:985–996
Arndt SK, Clifford SC, Wanek W, Jones HG, Popp M (2001)
Physiological and morphological adaptations of the fruit tree
Ziziphus rotundifolia in response to progressive drought stress.
Tree Physiol 21:705–715
Arnon DI, Hoagland DR (1940) Crop production in artificial solutions
and in soils with special reference to factors affecting yields and
absorption of inorganic nutrient. Soil Sci 50:463–484
Attipali RR, Kolluru VC, Munusamy V (2004) Drought induced
responses of photosynthesis and antioxidant metabolism in
higher plants. J Plant Physiol 161:1189–1202
Bacelar EA, Santos DL, Moutinho-Pereira JM, Goncalves BC,
Ferreira HF, Correia CM (2006) Immediate responses and
adaptative strategies of three olive cultivars under contrasting
water availability regimes: changes on structure and chemical
composition of foliage and oxidative damage. Plant Sci
170:596–605
Bacelar EA, Moutinho – Pereira JM, Goncalves BC, Ferreira HF,
Correia CM (2007) Changes in growth, gas exchange, xylem
hydraulic properties and water use efficiency of three olive
cultivars under contrasting water availability regimes. Environ
Exp Bot 60:183–192
Bates LS, Waldren RP, Teare IK (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–208
Ben Ahmed Ch, Ben Rouina B, Sensoy S, Boukhris M, Ben Abdallah
F (2009) Changes in gas exchange, proline accumulation and
antioxidative enzyme activities in three olive cultivars under
contrasting water availability regimes. Environ Exp Bot
67:345–352
Bian S, Jiang Y (2009) Reactive oxygen species, antioxidant enzyme
activities and gene expression patterns in leaves and roots of
Kentucky bluegrass in response to drought stress and recovery.
Sci Hortic 120:264–270
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants,
oxidative damage and oxygen deprivation stress. Ann Bot
91:179–194
Blum A, Munns R, Passioura JB, Turner NC (1996) Genetically
engineered plants resistant to soil dry and salt stress: how to
interpret osmotic relations? Plant Physiol 110:1050–1053
Bohnert HJ, Jenson RG (1996) Strategies for engineering water stress
tolerance in plants. Trends Biotech 14:89–97
Borgi W, Chouchane N (2009) Anti-spasmodic effects of Zizyphus
lotus (L.) Desf. extracts on isolated rat duodenum. J Ethnophar-
macol 126:571–573
Borgi W, Recio MC, Rios JL, Chouchane N (2008) Anti-inflamma-
tory and analgesic activities of flavonoid and saponin fractions
from Zizyphus lotus (L.) Lam. S Afr J Bot 74:320–324
Boussadia O, Ben Mariem F, Mechri B, Boussetta W, Braham M, Ben
El Hadj S (2008) Response to drought of two olive tree cultivars
(cv Koroneki and Meski). Sci Hortic 116:388–393
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
Chance B, Maehly AC (1955) Assay of catalases and peroxidases.
Methods Enzymol 2:764–775
Choudhary R, Sankhla N, Trivedi S, Joshi S (1996) Photosynthesis,
chlorophyll fluorescence, osmoregulatory solutes and enzyme
activities in ber (Ziziphus rotundifolia) under moisture stress.
Proc Plant Growth Regul Soc Am 23:206–210
Clifford SC, Arndt SK, Corlett JE, Joshi S, Sankhla N, Popp M, Jones
HG (1998) The role of solute accumulation, osmotic adjustment
and changes in cell wall elasticity in drought tolerance in cell
wall elasticity in drought tolerance in Ziziphus mauritiana
(Lamk.). J Exp Bot 49:967–977
DaCosta M, Huang B (2007) Changes in antioxidant enzyme
activities and lipid peroxidation for bentgrass species in
responses to drought stress. J Am Soc Hortic Sci 132:319–326
De Smedt S, Cunı Sanchez A, Van den Bilcke N, Simbo D, Potters G,
Samson R (2012) Functional responses of baobab (Adansonia
digitata L.) seedlings to drought conditions: differences
between western and south-eastern Africa. Environ Exp Bot
75:181–187
Dichio B, Romano M, Nuzzo V, Xiloyannis C (2002) Soil water
availability and relationship between canopy and roots in young
olive trees (cv Coratina). Acta Hortic 586:255–258
Dichio B, Xiloyannis C, Sofo A, Montanaro G (2005) Osmotic
regulation in leaves and roots of olives trees during a water
deficit and rewatering. Tree Physiol 26:179–185
Foyer CH, Noctor G (2005) Oxidant and antioxidant signalling in
plants: a re-evaluation of the concept of oxidative stress in a
physiological context. Plant Cell Environ 28:1056–1071
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen
peroxide- and glutathione-associated mechanisms of acclamato-
ry stress tolerance and signaling. Plant Physiol 100:241–254
Gindaba J, Rozanov A, Negash L (2004) Response of seedlings of
two Eucalyptus and three deciduous tree species from Ethiopia
to severe water stress. For Ecol Manag 201:119–129
Gorai M, Maraghni M, Neffati M (2010) The relationship between
phenological traits and water potential patterns of the wild jujube
Ziziphus lotus in southern Tunisia. Plant Ecol Divers 3:273–280
Guo Z, Ou W, Lu S, Zhong Q (2006) Differential responses of
antioxidative system to chilling and drought in four rice cultivars
differing in sensitivity. Plant Physiol Biochem 44:828–836
Guo Z, Huang EM, Lu ES, Yaqing EZ, Zhong EQ (2007) Differential
response to paraquat induced oxidative stress in two rice
cultivars on antioxidants and chlorophyll a fluorescence. Acta
Physiol Plant 29:39–46
Hassine AB, Lutts S (2010) Differential responses of salt bush
Atriplex halimus L. exposed to salinity and water stress in
relation to senescing hormones abscisic acid and ethylene. Plant
Physiol 167:1448–1456
Hewitt EJ (1966) Sand and water culture methods used in the study of
plant nutrition. Commonw Bur Hort Tech Com 22:431–446
Hudges DM, Delong JM, Forney FC, Prange RK (1999) Improving
the thiobarbituric acid-reactive-substances assay for estimating
lipid peroxidation in plant tissues containing anthocyanin and
other interfering compounds. Planta 207:604–611
Ingram J, Bartels D (1996) The molecular basis of dehydration
tolerance in plants. Ann Rev Plant Physiol Plant Mol Biol
47:377–403
Acta Physiol Plant
123

Jacobson L (1951) Maintenance of iron supply in nutrient solutions by
a single addition of ferric-potassium-ethylene-diamine-tetrace-
tate. Plant Physiol 26:411–413
Jones HG (1999) Selection of drought-tolerant fruit trees for summer
rainfall regions of Southern Africa and India. European Com-
mission STD-3, Brussels, Belgium, CTA, pp 118–120
Kulkarni M, Schneider B, Raveh E, Tel-Zur N (2010) Leaf
anatomical characteristics and physiological responses to short-
term drought in Ziziphus mauritiana (Lamk.). Sci Hortic
124:316–322
Larcher W (2003) Physiological plant ecology. Springer-Verlag, New
York
Le Floc’h E (1983) Contribution a une etude ethnobotanique de la
flore de la Tunisie. Imprimerie officielle de la Republique
Tunisienne, Tunis
Ludlow MM, Muchow RC (1990) A critical evaluation of traits for
improving crop yields in water-limited environments. Adv
Agron 43:107–153
Maraghni M, Gorai M, Neffati M (2010) Seed germination at
different temperatures and water stress levels, and seedling
emergence from different depths of Ziziphus lotus. S Afr J Bot
76:453–459
Maraghni M, Gorai M, Neffati M (2011) The influence of water-
deficit stress on growth, water relations and solute accumulation
in wild jujube (Ziziphus lotus). J Ornam Hortic Plant 1:63–72
Marnett LJ (1999) Lipid peroxidation—DNA damage by malondial-
dehyde. Mutat Res 424:83–95
Michel BE, Kaufmann MR (1973) The osmotic potential of polyeth-
ylene glycol-6000. Plant Physiol 51:914–916
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance.
Trends Plant Sci 9:405–410
Moller IM, Jensen PE, Hansson A (2007) Oxidative modifications to
cellular components in plants. Annu Rev Plant Biol 58:459–481
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by
ascorbate specific peroxidase in spinach chloroplasts. Plant Cell
Physiol 22:867–880
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping
active oxygen under control. Annu Rev Plant Physiol Plant Mol
Biol 49:249–279
Pareek OP (2001) Fruits for the Future 2: Ber. International Centre for
Underutilised Crops, University of Southampton, Southampton
Porcel R, Ruiz-Lozano JM (2004) Arbuscular mycorrhizal influence
on leaf water potential, solute accumulation, and oxidative stress
in soybean plants subjected to drought stress. J Exp Bot
55:1743–1750
Sankhla N (1998) Work done in India. In: Jones HG (ed) Selection of
drought tolerant fruit trees for summer rainfall regions of
Southern Africa and India. EU-STD, Brussels, Belgium
Schwanz P, Polle A (2001) Differential stress responses of antiox-
idative system to drought in Quercus robur and Pinus pinaster
grown under high CO2 concentrations. J Exp Bot 52:133–143
Serraj R, Sinclair TR (2002) Osmolyte accumulation: can it really
help increase crop yield under drought conditions? Plant Cell
Environ 25:333–341
Sofo A, Dichio B, Xiloyannis C, Masia A (2004) Effects of different
irradiance levels on some antioxidant enzymes and on malondi-
aldehyde content during rewatering in olive tree. Plant Sci
166:293–302
Sofo A, Manfreda S, Dichio B, Florentino M, Xiloyannis C (2007)
The olive tree: a paradigm for drought tolerance in Mediterra-
nean climates. Hydrol Earth Syst Sci Disc 4:2811–2835
Sundaresan S, Sudhakaran PR (1995) Water stress-induced alterations
in the proline metabolism of drought-susceptible and -tolerant
cassava (Maniohot esculenta) cultivars. Plant Physiol
94:635–642
Todaka D, Matsushima H, Morohashi Y (2000) Water stress enhances
beta-amylase activity in cucumber cotyledons. Environ Exp Bot
51:739–745
Turkan I, Bor M, Ozdemir F, Koca H (2005) Differential responses of
lipid peroxidation and antioxidants in the leaves of drought-
tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L.
subjected to polyethylene glycol mediated water stress. Plant Sci
168:223–231
Yordanov I, Velikova V, Tsonev T (2000) Plant responses to drought,
acclimation, and stress tolerance. Photosynthetica 38:171–186
Zhu JK (2002) Salt and drought stress signal transduction in plants.
Plant Biol 53:247–273
Acta Physiol Plant
123





























