Embed Size (px)
Citation preview

Differential responses of three sweetpotato metallothionein genesto abiotic stress and heavy metals
Sun Ha Kim • Jae Cheol Jeong • Young Ock Ahn •
Haeng-Soon Lee • Sang-Soo Kwak
Received: 20 April 2014 / Accepted: 1 July 2014 / Published online: 17 July 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Metallothioneins (MTs) are cysteine-rich, low
molecular weight, metal-binding proteins that are widely
distributed in living organisms. Plants produce metal-che-
lating proteins such as MTs to overcome the toxic effects
of heavy metals. We cloned three MT genes from sweet-
potato leaves [Ipomoea batatas (L.) Lam.]. The three IbMT
genes were classified according to their cysteine residue
alignment into type 1 (IbMT1), type 2 (IbMT2), and type 3
(IbMT3). IbMT1 was the most abundantly transcribed MT.
It was predominantly expressed in leaves, roots, and callus.
IbMT2 transcript was detected only in stems and fibrous
roots, whereas IbMT3 was strongly expressed in leaves and
stems. The IbMT expression profiles were investigated in
plants exposed to heavy metals and abiotic stresses. The
levels of IbMT1 expression were strongly elevated in
response to Cd and Fe, and moderately higher in response
to Cu. The IbMT3 expression pattern in response to heavy
metals was similar to that of IbMT1. Exposure to abiotic
stresses such as methyl viologen (MV; paraquat), NaCl,
polyethylene glycol (PEG), and H2O2 up-regulated IbMT
expression; IbMT1 responded strongly to MV and NaCl,
whereas IbMT3 was induced by low temperature and PEG.
Transgenic Escherichia coli overexpressing IbMT1 protein
exhibited results suggest that IbMT could be a useful tool
for engineering plants with enhanced tolerance to envi-
ronmental stresses and heavy metals.
Keywords Sweetpotato � Metallothionein � Reactive
oxygen species � Heavy metal � Phytoremediation
Abbreviations
MTs Metallothioneins
MV Methyl viologen
PCs Phytochelatins
PEG Polyethylene glycol
ROS Reactive oxygen species
H2O2 Hydrogen peroxide
Introduction
Heavy metal contamination of water and soil degrades the
environment, causes direct health hazards to animals and
human beings, and decreases crop productivity [1]. A
variety of technologies have been investigated for reme-
diation of sites contaminated with heavy metals. One cost-
effective approach is phytoremediation, in which plants
that actively sequester heavy metals are sown on contam-
inated site. Plants that are successfully used for phyto-
remediation have short lifetimes with high biomass, and
hyperaccumulate heavy metals. Heavy metal ions play
essential roles in several biological process in plants, but
excess amounts inhibit plant growth due to root damage,
decreased chlorophyll content, and leaf chlorosis [2, 3].
Plants produce metal-chelating peptides and proteins
such as phytochelatins (PCs) and metallothioneins (MTs)
to overcome heavy-metal stress [4]. MTs are cysteine-rich,
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11033-014-3582-y) contains supplementarymaterial, which is available to authorized users.
S. H. Kim � J. C. Jeong � Y. O. Ahn � H.-S. Lee (&) �S.-S. Kwak (&)
Plant Systems Engineering Research Center, Korea Research
Institute of Bioscience and Biotechnology (KRIBB), 125
Gwahangno, Daejeon 305-806, Korea
e-mail: [email protected]
S.-S. Kwak
e-mail: [email protected]
123
Mol Biol Rep (2014) 41:6957–6966
DOI 10.1007/s11033-014-3582-y

metal-binding proteins with low molecular weight that are
widely distributed in living organisms. MTs are involved in
the detoxification of heavy metals and in intracellular metal
ion homeostasis [5], and are implicated in scavenging
reactive oxygen species (ROS) [6–10] and stress responses
[11]. MTs are grouped into two classes (I and II) on the
basis of the cysteine (Cys) residue alignment [4, 5, 12, 13].
The MT cysteines are responsible for metal binding via
mercaptide bonds [13]. Plant MTs belong to class II, and
constitute a separate family that is subclassified into four
types on the basis of the cysteine residue distribution [4,
12]. Expression patterns of these four types can be corre-
lated with different tissues: type 1 is predominantly
expressed in roots, type 2 in shoot, type 3 in ripening fruits
or leaves, and type 4 is restricted to developing seeds [12].
Metallothionein gene expression is regulated by abiotic
stress, including heavy-metal stress, and plays an important
role in metal detoxification and homeostasis [4, 12–15].
Because MT proteins were first isolated as cadmium-
binding peptides in horse kidney [16], earlier studies of
plant MTs focused on their role in maintaining intracellular
metal homeostasis and mediating responses to metal tox-
icity [12]. However, more recent studies report additional
roles for plant MTs in development, fruit ripening, senes-
cence, and defense against oxidative stress. A recent study
showed that expression of the Brassica juncea MT2 in
Arabidopsis enhanced tolerance to copper and cadmium,
but inhibited root elongation [17]. Two MT-like genes were
up-regulated during natural leaf senescence in sweetpotato
[18]. Cotton MT3 scavenged ROS and enhanced plant
tolerance to oxidative stresses caused by NaCl, polyethyl-
ene glycol (PEG), and low temperature [6].
Bioremediation and phytoremediation using bacteria or
plants are very attractive strategies compared with physi-
cochemical methods for removal of heavy metals from
contaminated sites. They have lower costs and higher
efficiency for cleaning wastes, sediments, and soils con-
taminated with low levels of toxic heavy metals [19–21].
Although it is likely that both PCs and MTs participate in
heavy-metal detoxification in plants, distinctive roles have
not been clearly demonstrated. Plant MTs are thought to be
primarily involved in cellular ion homeostasis. Several
plant MT genes have been characterized, but the in vivo
function of each MT type has not been elucidated. The
mechanisms mediating heavy-metal tolerance remain
unclear in Ipomoea. Sweetpotato (Ipomoea batatas) is a
globally important root crop that is relatively tolerant to
unfavorable growth conditions and environmental stresses.
The molecular basis of heavy-metal stress tolerance in
sweetpotato has not been reported. In this study, we
investigate the responses of sweetpotato MTs to heavy
metals and abiotic stresses, and assess the effects of IbMT
overexpression in Escherichia coli.
Materials and methods
Plant material and growth conditions
Ten-day-old, white-flesh sweetpotato [I. batatas (L.) Lam.
cv. Yulmi, (Ym)] plantlets were cultivated in growth
chambers with 12-h day/12-h night cycles at 25 �C/18 �C.
Calli were induced from shoot meristems cultured on
Murashige-Skoog (MS) medium supplemented with 1 mg/l
2,4-dichlorophenoxyacetic acid (2,4-D), 3 % sucrose, and
0.4 % gelrite (MS1D). Calli were propagated on MS1D
medium with subculture at 14-d intervals, and maintained
at 25 �C in the dark.
Cloning IbMTs and sequence analysis
To clone IbMT1, IbMT2, and IbMT3 from sweetpotato,
IbMT primers were designed based on sequence obtained
from the NCBI database (http://ncbi.nlm.nih.gov). IbMT1
was amplified from the cDNA of sweetpotato (Ym) storage
root by RT-PCR using the advantage 2 pfu DNA poly-
merase mix (Clontech, Japan), and the IbMT gene-specific
primers listed in Table 1. The PCR products were cloned
into pGEM-TEasy vector (Promega, USA) and sequenced.
Multiple sequence alignments with other plant MTs were
computed using the CLUSTAL W program.
Stress treatments
Ten-day-old sweetpotato plantlets and 7-day-old calli (Ym)
were used for assessing plant responses to stress treat-
ments. To evaluate responses to hormones, plantlets and
calli were treated with 200 lM salicylic acid (SA), 10 lM
abscisic acid (ABA), 50 lM methyl jasmonate (MeJA),
and 0.3 % ethephon (ET). To evaluate responses to heavy
metal toxicity, plantlets were treated for 48 h to 300 lM
liquid of CaCl2, CuSO4, FeSO4, ZnSO4, CdSO4, Pb(NO)3,
MgSO4, and MnSO4, respectively. To evaluate responses
to abiotic stress, plantlets were exposed to 100 lM methyl
Table 1 Primer sequences for semi-quantitative RT-PCR
Primer name Primer sequence
lbMT1-F 50-ATGTCTTCCGGTTGCAAG-30
lbMT1-R 50-GCTGTCCCCACAGCTACAGT-30
lbMT2-F 50-ATGTCTTGCTGCGGAGGA-30
lbMT2-R 50-TCAACATTTGCAAGGGTCG-30
lbMT3-F 50-ATGTCGGACAAGTGCGGA-30
lbMT3-R 50-TTAGTGGCCACAGGTGCG-30
a-Tubulin-F 50-CAACTACCAGCCACCAACTGT-30
a-Tubulin-R 50-CAAGATCCTCACGAGCTTCAC-30
Note F forward, R reverse
6958 Mol Biol Rep (2014) 41:6957–6966
123

viologen (MV), 10 mM hydrogen peroxide (H2O2),
300 mM NaCl, or 25 % PEG. Cold tolerance was assessed
by exposing plantlets to 4 �C. Tissue samples of plantlets
were collected at 3, 9, 18, and 48 h after treatment with
hormones, heavy metals, or abiotic stress, frozen in liquid
nitrogen, and stored at -80 �C until analysis.
RNA isolation and RT-PCR analysis
Frozen plants tissues were ground in liquid nitrogen with a
mortar and pestle, and total RNA was extracted with the Easy-
Spin total RNA extraction kit (iNtRON, Korea) according to
the manufacturer’s instructions. RNA samples were treated
with DNase I for 60 min before further analysis to digest
contaminating genomic DNA. First-strand cDNA synthesis
was performed in a 20 ll reaction mixture containing 1 lg
RNA, 0.5 lg oligo dT, 40 lM each of the four dNTPs, 2 ll
109 buffer, 200 units of M-MLV reverse transcriptase, and 20
units of RNase inhibitor (MBI-Fermentas, Germany). The
reaction mixture was incubated at 42 �C for 60 min, and
stopped by heating at 65 �C for 10 min.
To determine gene expression patterns, semi-quantitative
PCR using gene-specific primers was performed using the
a-tubulin (accession no. DV037573) transcript level as
internal standard. The IbMT gene-specific primers used are
listed in Table 1. PCR reaction mixtures included the sin-
gle-stranded cDNA template along with sense and antisense
primers. The thermal cycling conditions were 94 �C for
30 s, 56–58 �C for 30 s, 72 �C for 30 s, with a total of 25
cycles. PCR products were separated by 2 % agarose gel
electrophoresis, and detected under ultraviolet illumination.
Construction of expression plasmids pET28a(?)-IbMTs
For construction of E. coli expression vector pET28a(?)
(Novagen, Germany), the following IbMT primers were
designed from IbMT sequences, with restriction enzyme sites
BamHI and SalI underlined: IbMT1-BamHI, 50-GC
GGATCCATGTCTTCCGGTTGCAAG-30; IbMT1-SalI, 50-CTGTCGACACAGTTGCAAGGGTC-30; IbMT2-BamHI,
50-GCGGATCCATGTCTTGCTGCGGAGGA-30; IbMT2-
SalI, 50-CTGTCGACAATTTGAAGGGTCG-30; IbMT3-
BamHI, 50-GCGGATCCATGTCGGACAAGTGCGGA-30;and IbMT3-SalI, 50-CTGTCGACGTGGCCACAGGTGCG-
30. PCR was performed in 20 ll reactions containing 1 lg of
pGEM-IbMT template and 0.1 lM of IbMT primers (con-
taining the BamHI and SalI restriction sites) under the fol-
lowing conditions: initial denaturation step at 95 �C for
2 min, followed by 30 cycles of denaturation at 95 �C for
20 s, annealing at 58 �C for 30 s, polymerization at 72 �C for
30 s, and a final extension at 72 �C for 10 min. PCR products
were digested with BamHI and SalI to release the IbMT
fragment. The IbMT bands were recovered from the gel and
sub-cloned into the expression vector pET28a(?) using the
DNA ligation kit (TaKaRa, Japan) to create pET28a(?)-
IbMT. Competent E. coli DH5a cells were transformed with
pET28a(?)-IbMT plasmids, positive clones were selected by
blue/white screening, and confirmed by restriction enzyme
analysis with BamHI and SalI.
Measurement of growth and heavy metal accumulation
in E. coli overexpressing IbMT
Purified pET28a(?)-IbMT plasmids (0.5 ll) were used to
transform 100 ll of E. coli BL21 (Stratagene, USA),
which were grown on LB plates containing 50 lg/ml
kanamycin at 37 �C overnight to select cells harboring
pET28a(?)-IbMT. A single colony was inoculated into
3 ml LB liquid medium, grown at 37 �C overnight,
100 ll of the overnight culture was added to a glass tube
containing 50 ml LB plus kanamycin, and then cultured at
37 �C. When cells grew to OD600 = 0.6, 1 mM isopro-
pyl-a-D-thiogalactopyranoside (IPTG) was added to
induce the lac promoter.
To measure the accumulation of heavy metals, E. coli
cells overexpressing IbMT, [pET28a(?)-IbMT] were grown
in liquid medium containing 1 mM CdCl2 or 500 lM ZnSO4
at 37 �C. When cells grew to OD600 = 0.6, 1 mM IPTG
was added and then incubated for a further 24 h at 30 �C.
Cells were harvesting, pellets were dried inverted in a fume
hood, and digested with 100 % nitric acid for 4 h at 95 �C.
After cooling at room temperature, the digests were diluted
with deionized water to a final volume of 50 ml, and ana-
lyzed for metals using inductively coupled plasma atomic
emission spectroscopy (ICP-AES; OPTIMA 7300DV; Per-
kin Elmer, USA).
Results
Multiple alignment and phylogenetic analysis of I.
batatas metallothionein genes
We cloned three MT genes from sweetpotato leaves. Each
MT was classified as type 1, type 2, or type 3 according to
the cysteine alignment, and designated as IbMT1, IbMT2,
or IbMT3, respectively. The full IbMT1 cDNA sequence
consisted of 201 nucleotides that encoded a 67 amino acid
polypeptide of approximately 4.6 kDa. IbMT2 cDNA
consisted of 249 nucleotides that encoded an 83 amino acid
polypeptide of approximately 4.2 kDa. IbMT3 cDNA
consisted of 195 nucleotides that encoded a 65 amino acid
polypeptide of approximately 4.8 kDa. The predicted pro-
teins contained two Cys-rich domains in the N- and
C-terminal regions. The deduced IbMT1 peptide contained
four Cys in the N-terminal region and five Cys in the
Mol Biol Rep (2014) 41:6957–6966 6959
123

C-terminal region, which were separated by a central, Cys-
free, 35 amino acid spacer. The deduced IbMT2 peptide
contained conserved sequences, MSCCGGN at the N-ter-
minal. The deduced IbMT3 peptide contained four Cys in
the N-terminal region and five Cys in the C-terminal
region, which were separated by a central, Cys-free, 32
amino acid spacer (Fig. 1a).
The genetic similarity of IbMTs with other plant MTs
was determined using blastx (https://blast.ncbi.nlm.nih.gov)
and CLUSTAL W (http://www.ualberta.ca/*stothard/
javascript/color_align_cons.html/). IbMT1 had the highest
homology with the putative MT of I. batatas L. cv. Kokei
(AB193246.1). IbMT2 and IbMT3 had the highest homology
with AF177760.1 and AF242374.1 of I. batatas L. cv. Tai-
nong57, respectively. Multiple alignments indicated that
IbMTs shared high homology with many MTs from other
species. To assess the evolutionary history of IbMTs, a
phylogenetic tree was constructed based on the deduced
amino acid sequence similarities of plant MT genes
(Fig. 1b).
A
B
Fig. 1 a Multiple alignment of three IbMT genes and other plant
MTs. Conserved cysteine residues are indicated by stars; conserved
amino acids are shaded. b Phylogenetic analysis of three IbMTs with
other plant MTs. Evolutionary history was inferred using the
neighbor-joining method; the optimal tree with the sum of branch
length = 4.13877083 is shown [33]. Evolutionary distances were
computed using the Poisson correction method [34], in units of the
number of amino acid substitutions per site. The analysis involved 16
amino acid sequences. All positions containing gaps and missing data
were eliminated. There were a total of 24 positions in the final dataset.
Evolutionary analyses were conducted in MEGA6 [35]
6960 Mol Biol Rep (2014) 41:6957–6966
123

Tissue-specific expression of IbMTs in sweetpotato
plants
To examine the tissue-specific expression patterns of
IbMTs in sweetpotato, RT-PCR analysis was performed
with total RNAs extracted from different tissues, including
leaf, stem, fibrous roots, storage roots, and cultured cells
(Figs. 2, 3). IbMT1 and IbMT3 had relatively higher basal
expression levels compared to that of IbMT2 under normal
conditions. IbMT1 was predominantly expressed in leaves
and fibrous roots, whereas IbMT2 was weakly expressed
only in stem and fibrous roots. IbMT3 was expressed in all
tissues examined, with strong expression in leaves and
stems. IbMT1 was strongly expressed only in cultured cells.
Differential expression of IbMT1 and IbMT3
in response to abiotic stress and heavy metals
The expression patterns of IbMTs in response to abiotic
stress and heavy metal were investigated in 10-day-old
sweetpotato plantlets. IbMT1 and IbMT3 expression dis-
played similar patterns; however, IbMT2 expression was
undetectable during these experiments because of the low
transcript levels. Therefore, we analyzed IbMT1 and
IbMT3, which had high expression levels that enabled
further study.
RT-PCR was used to evaluate the effects of abiotic
stress, including MV, NaCl, H2O2, PEG, and low tem-
perature (Fig. 4). Treatment with 100 lM MV induced
strong IbMT1 expression at 3 h after treatment, and
expression remained at a higher level thereafter. Treat-
ment with 100 lM MV induced IbMT3 expression at 3 h
after treatment; subsequently, transcript levels declined
gradually to the basal level. IbMT1 expression was
strongly enhanced at 9–18 h after treatment with
300 mM NaCl. IbMT3 expression patterns were similar,
showing an elevation at 9 h, followed by a decrease at
18 h, to a level that remained constant. Other stresses
such as cold (4 �C), 100 mM H2O2, and 25 % PEG did
not significantly affect IbMT1 and IbMT3 expression
levels. The IbMT1 and IbMT3 expression patterns show
similarities in their responses to H2O2 and PEG treat-
ments. Both MT genes were slightly up-regulated by
H2O2 at 18–48 h, and by PEG at 9 h after treatment.
IbMT1 expression was not appreciably induced by cold
treatment, whereas IbMT3 expression was apparent at
18 h after cold treatment.
We investigated the response to heavy metal stress using
CaCl2, FeSO4, CdSO4, CuSO4, MgSO4, ZnSO4, MnSO4,
and Pb(NO)3 (Fig. 5). IbMT1 and IbMT3 expression
showed the same patterns in response to CaCl2 treatment;
they were slightly down-regulated at 3–18 h after treat-
ment, and recovered to the basal expression level at 48 h.
IbMT1 and IbMT3 expression was weakly up-regulated at 3
and 18 h in response to FeSO4 treatment. IbMT1 expres-
sion was strongly induced by CdSO4 at 9–48 h after
treatment, and IbMT3 expression was clearly induced at
9 h. IbMT1 expression was up-regulated at 3–9 h after
treatment with CuSO4, but we did not detect significant
changes in IbMT3 transcription levels at 18–48 h. IbMT
expression displayed different responses to MgSO4; IbMT3
expression was completely suppressed by 9–18 h after
treatment, whereas IbMT1 expression did not change.
IbMT1 and IbMT3 expression decreased at 3 h after treat-
ment with ZnSO4; subsequently, it gradually increased up
to 48 h after treatment. We could not detect significant
changes in MT transcript levels after MnSO4 treatment.
However, IbMT1 and IbMT3 expression was strongly
induced by Pb(NO)3 at 48 h after treatment.
Transgenic culture cells expressing IbMT1 respond
to hormones, heavy metals, and abiotic stress
IbMT1 was specifically expressed in callus, whereas IbMT2
and IbMT3 were not detected (Fig. 3). To determine the
expression of IbMT1 in cultured cells, sweetpotato calli at
10 days after subculture were treated with four heavy
metals, four plant hormones, four abiotic stresses, and were
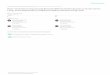
assessed at 3 and 18 h after treatment (Fig. 6). IbMT1
transcript levels in calli were markedly elevated in
response to ABA, MeJA, and ET at 3 h after treatment.
MeJA-induced IbMT1 expression continued at higher lev-
els until 18 h after treatment. SA-induced IbMT1 expres-
sion was higher at 18 h after treatment. In heavy metal
treatment, IbMT1 transcript levels were higher at 3 h after
Fe and Mn treatment, whereas Cu induced IbMT1
αα-tubulin
IbMT3
IbMT2
IbMT1
Leaf StemFibrous roots
Storage roots
Fig. 2 Semi-quantitative RT-PCR analysis of IbMTs expression
patterns in plant tissues of I. batatas cv. Yulmi
αα-tubulin IbMT1 IbMT2 IbMT3 callus
Fig. 3 Semi-quantitative RT-PCR analysis of IbMTs expression
patterns in cultured cells of I. batatas cv. Yulmi
Mol Biol Rep (2014) 41:6957–6966 6961
123

expression after 18 h. IbMT1 expression did not respond to
Zn. In abiotic stress studies, MV and NaCl slightly down-
regulated IbMT1 expression, whereas H2O2 markedly
down-regulated IbMT1 expression after 3 h. IbMT1
expression was clearly up-regulated by NaCl and low
temperature after 18 h (Fig. 6c).
Cold
MV
H2O2
tubulin
IbMT1
IbMT3
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
NaCl
PEG
tubulin
IbMT1
IbMT3
tubulin
IbMT1
IbMT3
Fig. 4 Semi-quantitative RT-
PCR analysis of IbMT1 and
IbMT3 expression in
sweetpotato plantlets in
response to abiotic stresses.
Total RNA was extracted at 0,
3, 9, 18, and 48 h after exposure
to 100 lM MV, 10 mM H2O2,
300 mM NaCl, or 25 % PEG.
Cold tolerance was assessed by
exposure to 4 �C
MgSO4
MnSO4
ZnSO4
tubulin
IbMT1
IbMT3
tubulin
IbMT1
IbMT3
Pb(NO)3
CuSO4
FeSO4
CdSO4
CaCl2
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
0 3 9 18 48 h
tubulin
IbMT1
IbMT3
tubulin
IbMT1
IbMT3
Fig. 5 Semi-quantitative RT-
PCR analysis of IbMT1 and
IbMT3 expression in
sweetpotato plantlets in
response to heavy metal. Total
RNA was extracted at 0, 3, 9,
18, and 48 h after exposure to
heavy metals including 300 lM
each CaCl2, CuSO4, FeSO4,
ZnSO4, CdSO4, Pb(NO)3,
MgSO4, and MnSO4
6962 Mol Biol Rep (2014) 41:6957–6966
123
IbMTs confer heavy metal tolerance to transgenic
E. coli
To determine whether IbMTs conferred heavy metal tol-
erance, three E. coli expression vectors were constructed
harboring IbMT1, IbMT2 and IbMT3 in pET28a (pET28a-
IbMT1, pET28a-IbMT2, and pET28a-IbMT3). For inde-
pendent proof of function, each IbMT was expressed as a
recombinant protein in E. coli. Soluble IbMT proteins were
extracted from bacterial lysates and affinity-purified on a
Ni-agarose matrix. Purified recombinant IbMT proteins
were analyzed by SDS-PAGE (Supplemental Fig. S1).
IbMT1 protein was detected after purification on the Ni-
column, but not in crude extract. IbMT2 partitioned into the
insoluble crude extract. IbMT3 was expressed in the solu-
ble crude extract.
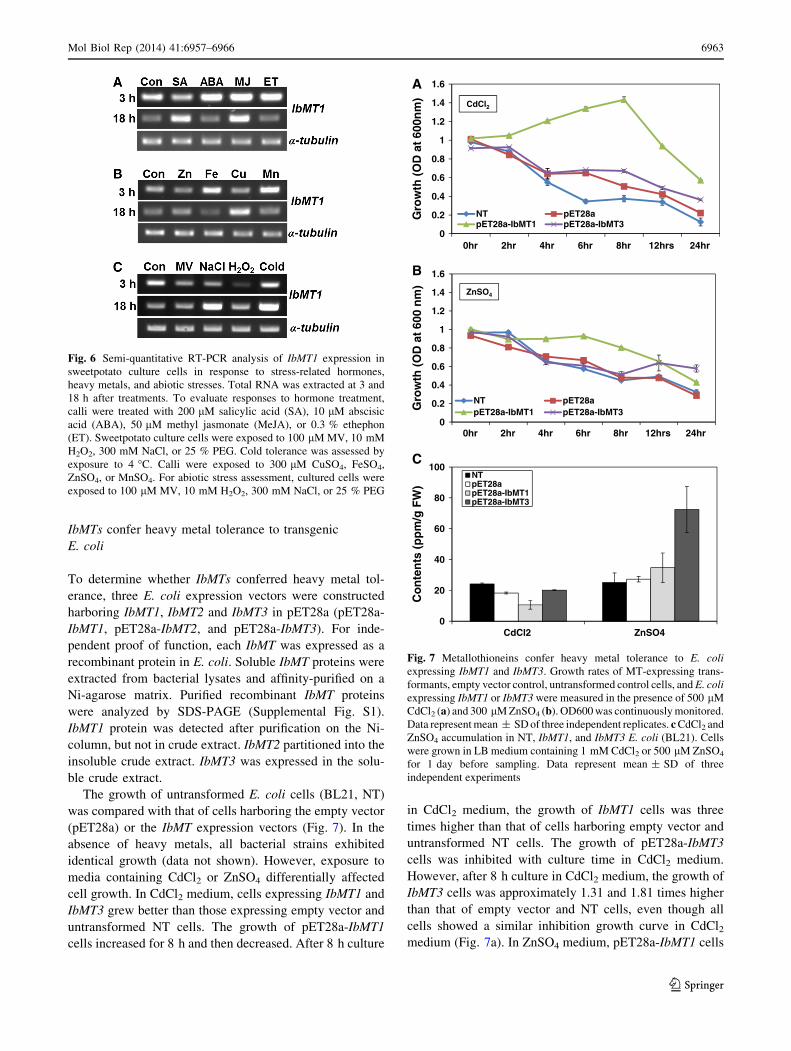
The growth of untransformed E. coli cells (BL21, NT)
was compared with that of cells harboring the empty vector
(pET28a) or the IbMT expression vectors (Fig. 7). In the
absence of heavy metals, all bacterial strains exhibited
identical growth (data not shown). However, exposure to
media containing CdCl2 or ZnSO4 differentially affected
cell growth. In CdCl2 medium, cells expressing IbMT1 and
IbMT3 grew better than those expressing empty vector and
untransformed NT cells. The growth of pET28a-IbMT1
cells increased for 8 h and then decreased. After 8 h culture
in CdCl2 medium, the growth of IbMT1 cells was three
times higher than that of cells harboring empty vector and
untransformed NT cells. The growth of pET28a-IbMT3
cells was inhibited with culture time in CdCl2 medium.
However, after 8 h culture in CdCl2 medium, the growth of
IbMT3 cells was approximately 1.31 and 1.81 times higher
than that of empty vector and NT cells, even though all
cells showed a similar inhibition growth curve in CdCl2medium (Fig. 7a). In ZnSO4 medium, pET28a-IbMT1 cells
Fig. 6 Semi-quantitative RT-PCR analysis of IbMT1 expression in
sweetpotato culture cells in response to stress-related hormones,
heavy metals, and abiotic stresses. Total RNA was extracted at 3 and
18 h after treatments. To evaluate responses to hormone treatment,
calli were treated with 200 lM salicylic acid (SA), 10 lM abscisic
acid (ABA), 50 lM methyl jasmonate (MeJA), or 0.3 % ethephon
(ET). Sweetpotato culture cells were exposed to 100 lM MV, 10 mM
H2O2, 300 mM NaCl, or 25 % PEG. Cold tolerance was assessed by
exposure to 4 �C. Calli were exposed to 300 lM CuSO4, FeSO4,
ZnSO4, or MnSO4. For abiotic stress assessment, cultured cells were
exposed to 100 lM MV, 10 mM H2O2, 300 mM NaCl, or 25 % PEG
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Gro
wth
(O
D a
t 60
0 n
m)
NT pET28apET28a-IbMT1 pET28a-IbMT3
CdCl2
ZnSO4
A
B
C
0
20
40
60
80
100
Co
nte
nts
(p
pm
/g F
W)
NTpET28apET28a-IbMT1pET28a-IbMT3
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
0hr 2hr 4hr 6hr 8hr 12hrs 24hr
CdCl2 ZnSO4
0hr 2hr 4hr 6hr 8hr 12hrs 24hr
Gro
wth
(O
D a
t 60
0nm
)
NT pET28apET28a-IbMT1 pET28a-IbMT3
Fig. 7 Metallothioneins confer heavy metal tolerance to E. coli
expressing IbMT1 and IbMT3. Growth rates of MT-expressing trans-
formants, empty vector control, untransformed control cells, and E. coli
expressing IbMT1 or IbMT3 were measured in the presence of 500 lM
CdCl2 (a) and 300 lM ZnSO4 (b). OD600 was continuously monitored.
Data represent mean ± SD of three independent replicates. c CdCl2 and
ZnSO4 accumulation in NT, IbMT1, and IbMT3 E. coli (BL21). Cells
were grown in LB medium containing 1 mM CdCl2 or 500 lM ZnSO4
for 1 day before sampling. Data represent mean ± SD of three
independent experiments
Mol Biol Rep (2014) 41:6957–6966 6963
123

grew better than that of cells harboring empty vector and
untransformed NT cells, but growth decreased after 8 h.
ZnSO4 inhibited the growth of pET28a-IbMT3 cells, but
growth was higher than that of cell harboring empty vector
and untransformed NT cells after 12 h (Fig. 7b). All cells
showed the same growth rate in CuSO4 medium, indicating
that CuSO4 does not affect cell growth (data not shown).
We confirmed that IbMTs conferred tolerance to heavy
metals (CdCl2 and ZnSO4) in E. coli. To examine whether
this tolerance is mediated by accumulation of heavy met-
als, E. coli cells overexpressing IbMT1 and IbMT3 were
cultured in medium with 1 mM CdCl2 and 500 lM ZnSO4,
respectively, and their metal ion contents were measured
(Fig. 7c). In CdCl2 medium, IbMT1-overexpressing cells
contained lower Cd2 ? levels compared to those of empty
vector and untransformed controls. In ZnSO4 medium,
IbMT3-overexpressing cells accumulated Zn to levels 2.5
times higher than those in empty vector and untransformed
controls.
Discussion
Research on plant MTs has focused primarily on eluci-
dating protein function. Shared common functions of plant
MTs include metal chelation, regulation of ion homeosta-
sis, and ROS quenching under different stress conditions.
The physiological roles of each MT gene in sweetpotato
plants have not been clearly defined. In this study, we
cloned three MT genes from sweetpotato leaves, charac-
terized their protein classification, and investigated their
expression patterns in response to hormones, heavy metals,
and abiotic stress using RT-PCR analysis. Each MT was
classified as type 1, type 2, or type 3 according to the
cysteine alignment (Fig. 1b). We tested potential in vivo
functions of recombinant IbMTs in heavy metal stress using
the E. coli expression system. In a previous report, two MT-
like genes, Y459 and G14 [18], were isolated from sweet-
potato cv. Tainong 57. Y459 was a type 2 MT that was
strongly expressed in senescent leaves, whereas G14 was a
type 4 MT that was likely a house-keeping gene.
Several reports indicate that MT expression is spatially
and temporally controlled in plants, and plant MTs display
tissue-specific expression. Plant type 1 MTs are more
abundantly expressed in roots than leaves, whereas type 2
MTs are expressed primarily in leaves and at a lower level
in roots [10, 12, 14, 18, 22, 23]. Type 3 MTs are expressed
in leaves or in ripening fruit [24], whereas type 4 MTs
appear to be restricted to developing seeds [25]. However,
the current study found that IbMT1 was predominantly
expressed in leaves and fibrous roots, and only IbMT1 was
specifically expressed in cultured cells (Fig. 2). IbMT2 was
detected only in the stem, and IbMT3 was predominantly
expressed in leaves and stems (Fig. 2). IbMT1 showed
relatively higher expression levels compared to those of
IbMT2 and IbMT3 under normal conditions in different
plant tissues, suggesting that IbMT1 may have diverse roles
or functions.
In previous studies, MT expression was induced by
heavy metals and abiotic stresses in different plants [15, 17,
26, 27]. Rice OsMT expression was induced by several
abiotic stresses including salts (NaCl and NaHCO3),
drought (PEG), and metal ions (CuCl2, ZnCl2, and CdCl2)
[28]. In the current study, IbMT1 and IbMT3 exhibited
similar expression patterns in response to abiotic stresses
and heavy metals (Figs. 3, 4). However, the IbMT1
expression level was higher than that of IbMT2. Our results
showed that IbMT1 was strongly induced by treatment with
MV, NaCl, CdCl2, and ZnSO4. In culture cells, IbMT1
differentially responded to abiotic stresses, stress-related
hormones, and heavy metals (Fig. 5). These results indicate
that expression of sweetpotato MTs is differentially regu-
lated in response to exogenous factors.
Previous reports discussed expressing plant MTs in
E. coli to assess if they confer tolerance to heavy metals.
We examined in vivo functions of recombinant IbMTs
using E. coli expressing pET28a-IbMTs vectors, and
examining cell growth in the presence of heavy metals.
Cell growth was affected by toxic concentrations of heavy
metals, and IbMT overexpression significantly improved
growth kinetics. E. coli expressing IbMT1 and IbMT3 grew
better in media containing high concentrations of CdCl2and ZnSO4, respectively, than those without IbMTs.
IbMT1-expressing cells had better growth rate compared
with that of IbMT3-expressing cells (Fig. 6). Both IbMT1-
and IbMT3-expressing cells accumulated metal ions,
except in CdCl2 medium, in which cells showed no accu-
mulation of Cd. By contrast, IbMT3-expressing cells
accumulated high levels of Zn, and grew relatively well. It
appears that IbMT1 may function to export cadmium ions
to the extracellular space. IbMT3 may function in metal
chelation in the vacuole for detoxification (Fig. 7c). These
results are consistent with other reports that Cd tolerance
and accumulation in sugarcane is mediated by other
mechanisms [29]. OsMT1b has greater affinity for Cd and
Ni ions compared to Zn and Cu ions, suggesting that
OsMT1b may protect rice cells against toxic metals, or play
a role in detoxification [23, 30]. The over-expression of
sheep MTII and maltose binding protein in the cytoplasm
of E. coli allowed the bioaccumulation of the heavy metals
such as Zn2?, Cu2? and Cd2?. This result suggested that
combination of metal export system and chelate system
like MT or PCs may also play an important role in bioac-
cumulation of heavy metal ion [31]. Arabidopsis MT3
enhances Cd tolerance when expressed in Vicia faba guard
cells [26]. Thlaspi caerulecens MT3 confers Cd, Cu, or Zn
6964 Mol Biol Rep (2014) 41:6957–6966
123

tolerance [27, 32]. Tamarix hispida MT-like ThMT3
enhances tolerance to Cd, Zn, Cu, and NaCl stress when
expressed in Saccharomyces cerevisiae [9]. It is anticipated
that IbMT1 and IbMT3 will play important roles in release
of accumulated metal ions and metal ion chelation,
respectively. Analysis of tolerance to heavy metal stress
conditions in plants expressing IbMTs remains to be
investigated in future studies. We suggest that three MT
genes from I. batatas will be useful tools for engineering
heavy metal tolerance and hyperaccumulation of metal ions
for new phytoremediation plants.
In conclusion, we cloned three MT genes from sweet-
potato leaves, which were classified as type 1, type 2, and
type 3. IbMT1 had relatively higher levels of basal
expression compared to those of IbMT2 and IbMT3 under
normal conditions. IbMT1 was predominantly expressed in
leaves, roots and callus, whereas IbMT2 was detected only
in the stem. IbMT3 was specifically expressed in stems and
leaves. In sweetpotato plantlets, IbMT1 and IbMT3
expression was strongly induced by Zn, Cd, Pb, MV, and
NaCl. In calli, IbMT1 was enhanced by ABA, MeJA, eth-
ephon, NaCl, and low temperature. Recombinant IbMT
proteins were successfully expressed in E. coli, and IbMT1
and IbMT3 conferred tolerance to heavy metal stress. The
results suggest that IbMTs are differentially regulated by
different exogenous factors. Thus, our results demonstrate
that the three MT genes from I. batatas may be useful tools
to develop transgenic plants and microorganisms with
enhanced tolerance to heavy metals and abiotic stresses,
which can be used for phytoremediation or tolerance to
environmental stresses.
Acknowledgments This work was carried out with the support of
‘‘Cooperative Research Program for Agriculture Science & Tech-
nology Development (Project No. PJ009506 and PJ008097)’’ Rural
Development Administration, Republic of Korea, and the KRIBB
Initiative Program.
References
1. MacFarlane GR (2002) Leaf biochemical parameters in Avicen-
nia marina (Forsk.) Vierh as potential biomarkers of heavy metal
stress in estuarine ecosystems. Mar Pollut Bull 44(3):244–256
2. Ebbs S, Uchil S (2008) Cadmium and zinc induced chlorosis in
Indian mustard [Brassica juncea (L.) Czern] involves preferential
loss of chlorophyll b. Photosynthetica 46(1):49–55
3. Lingua G, Franchin C, Todeschini V, Castiglione S, Biondi S,
Burlando B, Parravicini V, Torrigiani P, Berta G (2008) Arbus-
cular mycorrhizal fungi differentially affect the response to high
zinc concentrations of two registered poplar clones. Environ
Pollut 153(1):137–147
4. Robinson NJ, Tommey AM, Kuske C, Jackson PJ (1993) Plant
metallothioneins. Biochem J 295(Pt 1):1–10
5. Hamer DH (1986) Metallothionein. Annu Rev Biochem
55:913–951
6. Xue T, Li X, Zhu W, Wu C, Yang G, Zheng C (2009) Cotton
metallothionein GhMT3a, a reactive oxygen species scavenger,
increased tolerance against abiotic stress in transgenic tobacco
and yeast. J Exp Bot 60(1):339–349
7. Wong HL, Sakamoto T, Kawasaki T, Umemura K, Shimamoto K
(2004) Down-regulation of metallothionein, a reactive oxygen
scavenger, by the small GTPase OsRac1 in rice. Plant Physiol
135(3):1447–1456
8. Yang Z, Wu Y, Li Y, Ling HQ, Chu C (2009) OsMT1a, a type 1
metallothionein, plays the pivotal role in zinc homeostasis and
drought tolerance in rice. Plant Mol Biol 70(1–2):219–229
9. Yang J, Wang Y, Liu G, Yang C, Li C (2011) Tamarix hispida
metallothionein-like ThMT3, a reactive oxygen species scaven-
ger, increases tolerance against Cd2?, Zn2?, Cu2?, and NaCl in
transgenic yeast. Mol Biol Rep 38(3):1567–1574
10. Navabpour S, Morris K, Allen R, Harrison E, A-H-Mackerness S,
Buchanan-Wollaston V (2003) Expression of senescence-enhanced
genes in response to oxidative stress. J Exp Bot 54(391):2285–2292
11. Singh R, Anandhan S, Singh S, Patade V, Ahmed Z, Pande V (2011)
Metallothionein-like gene from Cicer microphyllum is regulated by
multiple abiotic stresses. Protoplasma 248(4):839–847
12. Cobbett C, Goldsbrough P (2002) Phytochelatins and metallo-
thioneins: roles in heavy metal detoxification and homeostasis.
Annu Rev Plant Biol 53:159–182
13. Kaegi JHR, Schaeffer A (1988) Biochemistry of metallothionein.
Biochemistry 27(23):8509–8515
14. Huang G-Y, Wang Y-S (2009) Expression analysis of type 2
metallothionein gene in mangrove species (Bruguiera gymno-
rrhiza) under heavy metal stress. Chemosphere 77(7):1026–1029
15. Zenk MH (1996) Heavy metal detoxification in higher plants-a
review. Gene 179(1):21–30
16. Margoshes M, Vallee BL (1957) A cadmium protein from equine
kidney cortex. J Am Chem Soc 79(17):4813–4814
17. Zhigang A, Cuijie L, Yuangang Z, Yejie D, Wachter A, Gromes R,
Rausch T (2006) Expression of BjMT2, a metallothionein 2 from
Brassica juncea, increases copper and cadmium tolerance in Esch-
erichia coli and Arabidopsis thaliana, but inhibits root elongation in
Arabidopsis thaliana seedlings. J Exp Bot 57(14):3575–3582
18. Chen HJ, Hou WC, Yang CY, Huang DJ, Liu JS, Lin YH (2003)
Molecular cloning of two metallothionein-like protein genes with
differential expression patterns from sweetpotato (Ipomoea ba-
tatas) leaves. J Plant Physiol 160(5):547–555
19. Salt DE, Blaylock M, Kumar NPBA, Dushenkov V, Ensley BD,
Chet I, Raskin I (1995) Phytoremediation: a novel strategy for the
removal of toxic metals from the environment using plants. Nat
Biotech 13(5):468–474
20. Kumar PBAN, Dushenkov V, Motto H, Raskin I (1995) Phy-
toextraction: the use of plants to remove heavy metals from soils.
Environ Sci Technol 29(5):1232–1238
21. Gisbert C, Clemente R, Navarro-Avino J, Baixauli C, Giner A,
Serrano R, Walker D, Bernal M (2006) Tolerance and accumu-
lation of heavy metals by Brassicaceae species grown in con-
taminated soils from Mediterranean regions of Spain. Environ
Exp Bot 56(1):19–27
22. Omidvar V, Abdullah S, Izadfard A, Ho C, Mahmood M (2010)
The oil palm metallothionein promoter contains a novel AGT-
TAGG motif conferring its fruit-specific expression and is
inducible by abiotic factors. Planta 232(4):925–936
23. Fukuzawa H, Yu LH, Umeda-Hara C, Tagawa M, Uchimiya H
(2004) The rice metallothionein gene promoter does not direct
foreign gene expression in seed endosperm. Plant Cell Rep
23(4):231–235
24. Ledger SEGR (1994) Cloning and characterization of five cDNAs
for genes differentially expressed during fruit development of
kiwifruit (Actinidia deliciosa var. deliciosa). Plant Mol Biol
25:866–877
Mol Biol Rep (2014) 41:6957–6966 6965
123

25. Kawashima IKT, Chino M, Lane BG (1992) Wheat E,
metallothionein genes– like mammalian Zn2? metallothio-
nein genes, wheat Zn2? metallothionein genes are conspic-
uously expressed during embryogenesis. Eur J Biochem
209:971–976
26. Lee J, Donghwan S, Won-yong S, Inhwan H, Youngsook L
(2004) Arabidopsis metallothioneins 2a and 3 enhance resistance
to cadmium when expressed in Vicia faba guard cells. Plant Mol
Biol 54(6):805–815
27. Assuncao AGL, Ten Bookum WM, Nelissen HJM, Vooijs R,
Schat H, Ernst WHO (2003) A cosegregation analysis of zinc
(Zn) accumulation and Zn tolerance in the Zn hyperaccumulator
Thlaspi caerulescens. New Phytol 159(2):383–390
28. Jin S, Cheng Y, Guan Q, Liu D, Takano T, Liu S (2006) A
metallothionein-like protein of rice (rgMT) functions in E. coli
and its gene expression is induced by abiotic stresses. Biotechnol
Lett 28(21):1749–1753
29. Sereno ML, Almeida RS, Nishimura DS, Figueira A (2007)
Response of sugarcane to increasing concentrations of copper and
cadmium and expression of metallothionein genes. J Plant
Physiol 164(11):1499–1515
30. Nezhad R, Shahpiri A, Mirlohi A (2013) Heterologous expression
and metal-binding characterization of a type 1 metallothionein
isoform (OsMTI-1b) from rice (Oryza sativa). Protein J
32(2):131–137
31. Sauge-Merle S, Lecomte-Pradines C, Carrier P, Cuine C, Du-
Bowf M (2012) Heavy metal accumulation by recombinant
mammalian metallothionein within Escherichia coli protects
against elevated metal exposure. Chemosphere 88:918–924
32. Hassinen VH, Tuomainen M, Peraniemi S, Schat H, Karenlampi
SO, Tervahauta AI (2009) Metallothioneins 2 and 3 contribute to
the metal-adapted phenotype but are not directly linked to Zn
accumulation in the metal hyperaccumulator, Thlaspi caerules-
cens. J Exp Bot 1:187–196
33. Saitou N, Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol Biol Evol
4:406–425
34. Zuckerkandl E, Pauling L (1965) Evolutionary divergence and
convergence in proteins. Academic Press, New York, pp 97–166
35. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013)
MEGA6: molecular evolutionary genetics analysis version 6.0.
Mol Biol Evol 30:2725–2729
6966 Mol Biol Rep (2014) 41:6957–6966
123