Embed Size (px)
Citation preview

Differential Regulation of Na,K-ATPase Isozymes by ProteinKinases and Arachidonic Acid
Gustavo Blanco, Gladis Sanchez, and Robert W. Mercer1
Department of Cell Biology and Physiology, Washington University School of Medicine, St. Louis, Missouri 63110
Received March 13, 1998, and in revised form June 25, 1998
While several studies have investigated the regula-tion of the Na,K-ATPase consisting of the a1 and b1subunits, there is little evidence that intracellularmessengers influence the other Na pump isozymes. Westudied the effect of different protein kinases and ar-achidonic acid on the rat Na,K-ATPase isoforms ex-pressed in Sf-9 insect cells. Our results indicate thatPKA, PKC, and PKG are able to differentially modifythe function of the Na,K-ATPase isozymes. While PKCactivation leads to inhibition of all isozymes, PKA ac-tivation stimulates the activity of the Na,K-ATPasea3b1 and decreases that of the a1b1 and a2b1isozymes. In contrast, activation of PKG diminishesthe activity of the a1b1 and a3b1 isozymes, withoutaltering that of a2b1. Treatment of cells with arachi-donic acid reduced the activities of all the isozymes.The changes in the catalytic capabilities of the Napump isozymes elicited by PKA and PKC are reflectedby changes in the molecular activity of the Na,K-ATPases. One of the mechanisms by which PKA andPKC affect Na pump isozyme activity is through directphosphorylation of the a subunit. In the insect cells,we found a PKA- and PKC-dependent phosphorylationof the a1, a2 and a3 polypeptides. In conclusion, sev-eral intracellular messengers are able to modulate thefunction of the Na,K-ATPase isozymes and some ofthem in a specific fashion. Because the Na,K-ATPaseisozymes have kinetic properties that are unique, thisisozyme-specific regulation may be important inadapting Na pump function to the requirements ofeach cell. © 1998 Academic Press
Key Words: Na,K-ATPase; protein kinases; baculovi-rus; isozymes.
The Na,K-ATPase or Na pump is responsible formaintaining the low internal Na1 and high internalK1 concentrations characteristic of most mammaliancells. The ion gradients created by the enzyme areimportant in preserving the volume, pH, and the elec-trical resting potential of the cells. In addition, thetransmembrane Na1 gradient provides the force thatdrives the action potential in excitable tissues and theco- and countertransport of ions and substrates acrossthe cell membrane (1). The Na,K-ATPase uses the freeenergy from the hydrolysis of one molecule of ATP toexchange three intracellular Na ions for two extracel-lular K ions (2, 3). Normal function of the Na,K-AT-Pase requires the association of two polypeptides, the aand b subunits. The a subunit is a multispanningmembrane protein that contains the binding sites forATP, the cations, and the specific inhibitor, ouabain(reviewed in 4, 5). The b subunit is a type II glycosy-lated polypeptide essential for normal Na,K-ATPaseactivity (6–8). The b polypeptide appears to be in-volved in the occlusion of K1 (9) and in the modulationof the Na1 (10–12) and K1 (13) affinity of the enzyme.
Because numerous cellular transport systems arecoupled to the movement of Na1, and therefore, to thefunction of the Na pump, Na,K-ATPase activity isadapted to the changing physiological requirements ofthe cell. Regulation of the Na pump can occur by sev-eral mechanisms. For example, the amount of the en-zyme at the plasma membrane can be modified bychanges in the rate of synthesis or degradation of theindividual Na pump polypeptides, as well as throughthe movement of enzyme molecules by exo/endocytoticvesicular transport. All these mechanisms involve thecontrol of Na pump expression at the cell surface. Al-ternatively, the activity of the Na pump at the cellsurface can be directly regulated to provide an imme-diate adjustment in Na pump function (14–16). Thisrapid modification in Na,K-ATPase activity can be aresult of direct phosphorylation and dephosphorylation
1 To whom correspondence should be addressed at Department ofCell Biology and Physiology, Washington University School of Med-icine, 660 S. Euclid Avenue, St. Louis, MO 63110. Fax: (314)362-7463. E-mail: [email protected].
0003-9861/98 $25.00 139Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 359, No. 2, November 15, pp. 139–150, 1998Article No. BB980904

by protein kinases and protein phosphatases. Thus,depending on the tissue, activation of protein kinasescan induce an increase or decrease in Na pump activity(for review see 16). For example, in some cells, agentsthat stimulate protein kinase A (PKA)2 cause a de-crease in Na,K-ATPase activity (17–19). This responseappears to be mediated, at least in part, by PKA-directed phosphorylation of serine 943 of the Na pumpa subunit (20). The effect is blocked either by inhibitorsof PKA or phospholipase A2 (PLA2), suggesting thatactivation of PLA2 and production of arachidonic acidand its metabolites may be involved (21). Phosphory-lation of the Na pump a subunit also appears to bereversible because endogenous and exogenous inhibi-tors of protein phosphatase 1 (PP1) and protein phos-phatase 2A (PP2A) produce a decrease in Na,K-ATPaseactivity (22, 23). This suggests that a phosphorylation/dephosphorylation event can dynamically regulate theactivity of the Na pump. The activity of the Na,K-ATPase is also altered by phorbol esters or diacylglyc-erol analogs, in a process that involves protein kinaseC (PKC) activation and, possibly, the phosphorylationof the Na pump a subunit at serine 16 (24). Activationof PKC can produce an inhibition (18, 25, 26) or stim-ulation (27, 28) in Na,K-ATPase activity. In addition, ithas been proposed that acetylcholine, bradykinin, andatrial natriuretic peptide produce salt and water lossin the kidney, a response that is mediated by proteinkinase G (PKG) and the production of cyclic GMP (29,30). This cyclic nucleotide also appears to mediate theactivation of the Na,K-ATPase elicited by cholinomi-metics and carbon monoxide (31, 32).
Interestingly, the Na,K-ATPase exists as severalisozymes, which result from the association of the dif-ferent isoforms of the a and b subunits. The various Napump isozymes are expressed in a tissue-specific, de-velopmentally and hormonally regulated manner (33–37). Moreover, each individual isozyme has unique ki-netic properties (11, 12, 38, 39). Rapid changes inNa,K-ATPase activity by short-term modulators havebeen studied using the a1b1 isozyme; however, there islittle evidence for regulation of the a2 and a3 isoformsby intracellular messengers. The study of the individ-ual Na pump isozymes is complicated because oftenseveral isozymes are expressed in the same cell. Thebaculovirus expression system has proven useful forthe expression of the Na pump isoforms in Sf-9 insectcells (8, 40). Sf-9 insect cells have the advantage ofpossessing very little endogenous Na,K-ATPase activ-ity. We have previously shown that Sf-9 cells express-
ing the rat Na,K-ATPase a and b polypeptides producecatalytically competent Na,K-ATPase molecules (11,12, 40). Moreover, Sf-9 cells can glycosylate, phosphor-ylate, acylate, and perform most of the posttransla-tional modifications dictated by the primary aminoacid sequence. In addition, the insect cells display acomplex signal transduction machinery common toother eukaryotic cells (41–45). In this work we inves-tigated the effect of activators of different protein ki-nases and arachidonic acid on the activity of the Na,K-ATPase isozymes expressed in insect cells using recom-binant baculoviruses.
MATERIALS AND METHODS
Infections and activation of protein kinases. Uninfected and in-fected Sf-9 cells were grown in 150-mm petri dishes in TNM:FHmedium (defined in 41) supplemented with 10% fetal bovine serum,100 units/ml penicillin, 100 mg/ml streptomycin, and 0.25 mg/mlfungizone. Infections with recombinant baculoviruses containing therat a1, a2, a3, and b1 Na,K-ATPase cDNAs (46, 47) were performedas described previously (40). At 40 h postinfection, cells were treatedwith the protein kinase modulators. For PKA stimulation, 2 mM ofdibutyryl cAMP (N6-29-O-dibutyryladenosine 39:59-cyclic monophos-phate; db-cAMP) was used for 1 h. PKC activation was obtained with1 mM phorbol 12-myristate 13-acetate (PMA) for 1 h. Preliminarydose–response experiments demonstrated that treatment with 1 mMPMA or 2 mM db-cAMP were sufficient to achieve maximal re-sponses. The effects of PMA or db-cAMP on Na,K-ATPase activitywere apparent after 15 min treatment and were maximal at 30 min.Typically, cells were incubated in the presence of the activators for1 h. Finally, PKG activation was achieved with 1 mM dibutyrylcGMP (db-cGMP) for 1 h. As inhibitors of protein kinases, 50 mM of8-bromoadenosine 39,59 cyclic monophosphothioate, Rp-isomer wasused to antagonize PKA effects and staurosporine (200 nM) or [1-(5-isoquinolinesulfonyl)-2-methylpiperazine] (H7; 50 mM) was added toblock the effects of PKC. For PKG, the specific inhibitor KT 5823 wasused at 1 mM. After a 30-min incubation with the protein kinaseantagonists, the respective activators in the continued presence ofthe antagonists were added. The cells were then harvested, centri-fuged at 1500 g for 10 min, resuspended in 10 mM imidazole hydro-chloride (pH 7.5), 1 mM EGTA, and used for the determination ofNa,K-ATPase activity.
Na,K-ATPase assays. Enzymatic activity was determined by theinitial rate of release of 32Pi from [g-32P]ATP on the intact cells afterpermeabilization with alamethicin (11) or on membranes preparedfrom the infected cells (48). Permeabilization of cells with alamethi-cin at a concentration of 0.01 mg alamethicin/mg total protein com-pletely unmasked Na,K-ATPase activity from the intact cells (datanot shown). Specific activity was considered as the difference in ATPhydrolysis in the absence and presence of 1 mM ouabain. Theouabain-sensitive hydrolysis of the cells expressing the different Napump isozymes was approximately 7 to 10 times greater than theNa,K-ATPase activity observed in uninfected cells.
[3H]Ouabain binding assays. Equilibrium binding of [3H]ouabainwas performed on 100 mg aliquots of membrane protein preparations asdescribed previously (40). Preparations were incubated for 1 h at 37°Cin 2 ml of medium containing: 100 mM NaCl, 4 mM MgCl2, 4 mM ATP,40 mM imidazole–HCl (pH 7.4), 0.2 mM EGTA, and 0.7 to 28 mCi of[3H]ouabain. Samples were filtered under vacuum and washed withincubation medium and the radioactivity was measured. Nonspecificbinding (,5%) was determined by the binding in the presence of 1 mMunlabeled ouabain. Maximal binding of [3H]ouabain was calculatedfrom Scatchard plots of the data, which in all cases showed a singlepopulation of ouabain binding sites. The molecular activity of the Na,K-
2 Abbreviations used: PKA, protein kinase A; PLA2, phospholipaseA2; PP1, protein phosphatase 1; PP2A, protein phosphatase 2A;PKC, protein kinase C; PKG, protein kinase G; PMA, phorbol 12-myristate 13-acetate; DMEM, Dulbecco’s modified Eagle medium; db-cAMP, dibutyryl cyclic AMP AACOF3 dibutyryltrifluoromethyl ketone.
140 BLANCO, SANCHEZ, AND MERCER
ATPase isozymes was calculated from the ratio between the maximalNa,K-ATPase activity and the [3H]ouabain binding sites determined onmembrane preparations from the infected cells.
Phosphorylations and metabolic labeling. Protein kinase-depen-dent phosphorylations were carried out 40 h after infection on cellsgrown in six-well tissue culture plates. The experiment was startedby starving the cells for 1 h in phosphate-free Dulbecco’s modifiedEagle medium (DMEM), pH 6.2, and 1% dialyzed fetal calf serum.32Pi was then added and allowed to incorporate into the cellular poolfor 1 h. Subsequently, cells were treated with or without the differentprotein kinase inhibitors for 30 min, followed by a 1-h incubationwith PMA or dibutyryl cAMP as indicated. All the protein kinasemodulators were applied at the concentrations indicated above. Inevery case, cells were lysed in 1 ml of 150 mM NaCl, 10 mM NaFl, 20mM Na4P2O7, 0.1 % SDS, 1% Triton X-100, and 30 mM Hepes, pH7.4; insoluble material was removed by centrifugation (10 min at15,000 g), and samples were subjected to immunoprecipitation. For
a1b1, 50 ml of an anti-a1 monoclonal antibody (C464-6B) provided byDr. M. Caplan (Yale University School of Medicine) was used. Fora2b1 and a3b1, 50 ml of an anti-a monoclonal antibody hybridomasupernatant (5a, provided by Dr. D. Fambrough, Johns HopkinsUniversity) was used. To precipitate the immunocomplexes, 70 ml (1mg/ml) of goat anti-mouse-coated magnetic beads was added (Dyna-beads; Dynal Corp., Great Neck, NY). After overnight incubation ona rocking table at 4°C, beads were isolated by holding the microcen-trifuge tube to a magnet and aspirating the supernatant. The beadswere washed three times in the lysis buffer. The precipitated proteinwas eluted by resuspending the beads in Laemmli sample buffer (100mM Tris–HCl, pH 6.8, 2% SDS, 33% glycerol, 100 mM DTT) andincubating the mixture for 15 min at 65°C. Eluted proteins wereseparated by SDS–PAGE (49), gels (7.5%) were dried, and radioac-tivity was quantitated using a PhosphorImager SI scanner and theImageQuant software (Molecular Dynamics, Sunnyvale, CA). Theintensity of labeling was normalized as a percentage of the phosphor-
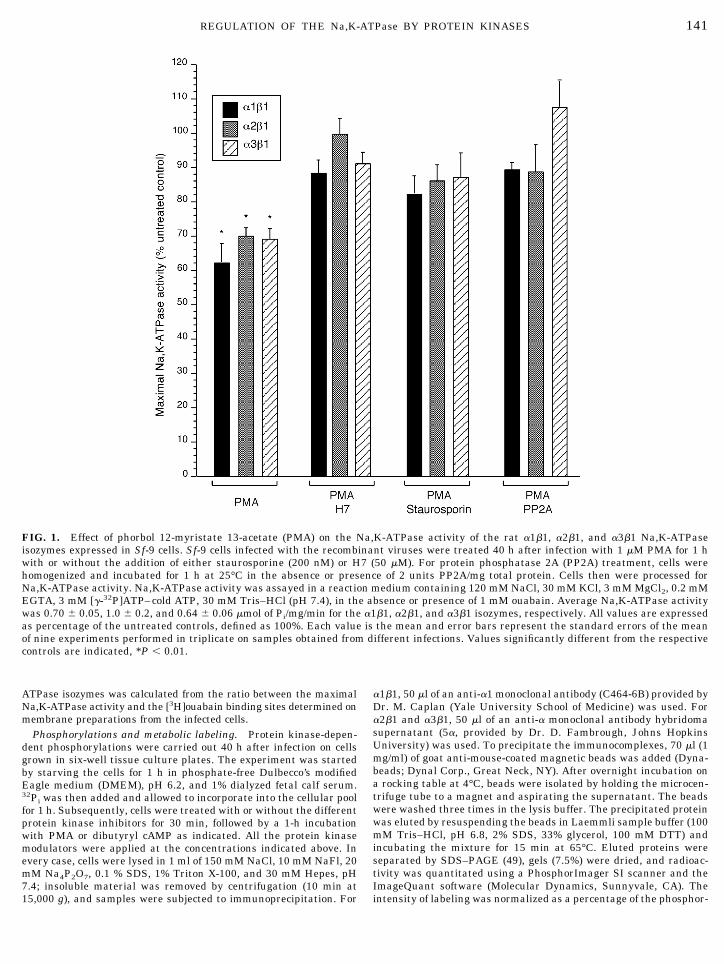
FIG. 1. Effect of phorbol 12-myristate 13-acetate (PMA) on the Na,K-ATPase activity of the rat a1b1, a2b1, and a3b1 Na,K-ATPaseisozymes expressed in Sf-9 cells. Sf-9 cells infected with the recombinant viruses were treated 40 h after infection with 1 mM PMA for 1 hwith or without the addition of either staurosporine (200 nM) or H7 (50 mM). For protein phosphatase 2A (PP2A) treatment, cells werehomogenized and incubated for 1 h at 25°C in the absence or presence of 2 units PP2A/mg total protein. Cells then were processed forNa,K-ATPase activity. Na,K-ATPase activity was assayed in a reaction medium containing 120 mM NaCl, 30 mM KCl, 3 mM MgCl2, 0.2 mMEGTA, 3 mM [g-32P]ATP–cold ATP, 30 mM Tris–HCl (pH 7.4), in the absence or presence of 1 mM ouabain. Average Na,K-ATPase activitywas 0.70 6 0.05, 1.0 6 0.2, and 0.64 6 0.06 mmol of Pi/mg/min for the a1b1, a2b1, and a3b1 isozymes, respectively. All values are expressedas percentage of the untreated controls, defined as 100%. Each value is the mean and error bars represent the standard errors of the meanof nine experiments performed in triplicate on samples obtained from different infections. Values significantly different from the respectivecontrols are indicated, *P , 0.01.
141REGULATION OF THE Na,K-ATPase BY PROTEIN KINASES
ylation obtained without the protein kinase inhibitors. To label cel-lular proteins, 40-h infected cells in six-well plates were starved inmethionine-free medium. After 1 h, [35S]methionine (0.3 mM methi-onine, 40 mCi/well) in the absence or presence of 0 to 100 mg/mlcycloheximide was added. After 1 h, samples were prepared forimmunoprecipitation and processed as before.
Biotinylation assay. The level of expression of Na pump mole-cules at the cell surface was determined by immunoblot analysis ofthe cell surface biotinylated proteins (50). Briefly, 48 h after infec-tion, Sf-9 cells cultured in six-well plates were treated with or with-out the PK activators for 1 h. Cells were washed with phosphatesaline buffer (PBS) and incubated at 4°C for 1 h in medium contain-ing 30 mM Hepes, pH 7.4, 150 mM NaCl (HBS), and 1.5 mg/mlsulfo-NHS-biotin (EZ-Link, Pierce). The cells then were washed andincubated for 20 min with 100 mM glycine in PBS to quench the freebiotin. Lysis of the cells was performed with 1 ml of 1% Triton X-100in HBS and the surface-labeled biotinylated proteins were precipi-tated with 100 ml of Immunopure immobilized streptavidin (Pierce).After overnight incubation at 4°C, beads were washed three timeswith the lysis solution and resuspended in Laemmli sample buffer
and samples were subjected to SDS–PAGE (7.5% gel) and trans-ferred to nitrocellulose. Immunoblotting was performed as previ-ously described using an anti-a1-specific antibody and a 125I-labeledsecondary antibody (48). Autoradiographs were digitized using an
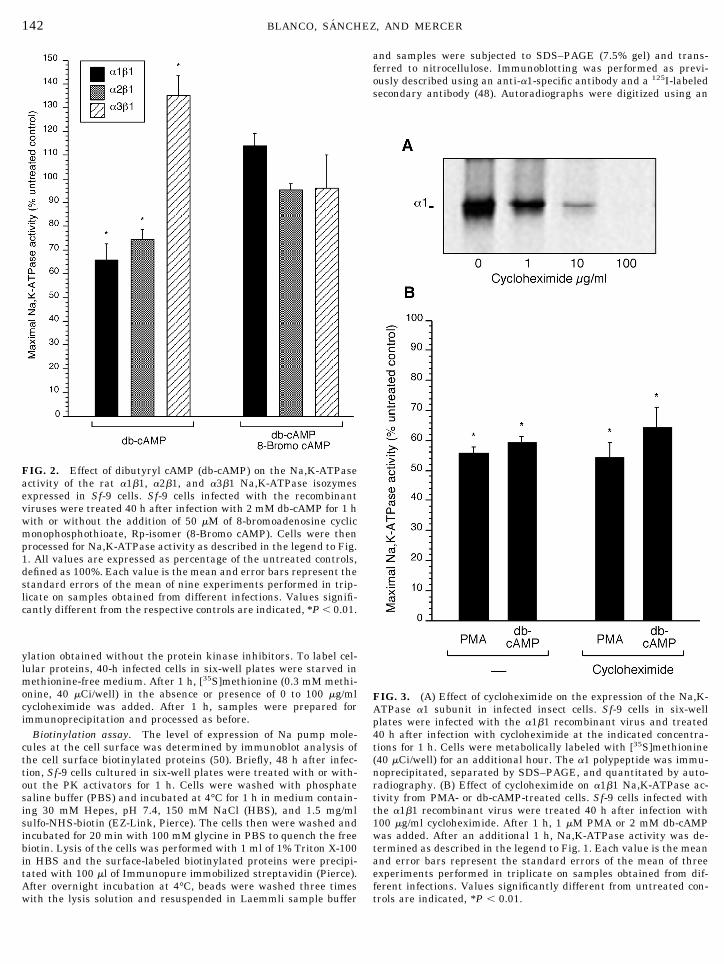
FIG. 2. Effect of dibutyryl cAMP (db-cAMP) on the Na,K-ATPaseactivity of the rat a1b1, a2b1, and a3b1 Na,K-ATPase isozymesexpressed in Sf-9 cells. Sf-9 cells infected with the recombinantviruses were treated 40 h after infection with 2 mM db-cAMP for 1 hwith or without the addition of 50 mM of 8-bromoadenosine cyclicmonophosphothioate, Rp-isomer (8-Bromo cAMP). Cells were thenprocessed for Na,K-ATPase activity as described in the legend to Fig.1. All values are expressed as percentage of the untreated controls,defined as 100%. Each value is the mean and error bars represent thestandard errors of the mean of nine experiments performed in trip-licate on samples obtained from different infections. Values signifi-cantly different from the respective controls are indicated, *P , 0.01.
FIG. 3. (A) Effect of cycloheximide on the expression of the Na,K-ATPase a1 subunit in infected insect cells. Sf-9 cells in six-wellplates were infected with the a1b1 recombinant virus and treated40 h after infection with cycloheximide at the indicated concentra-tions for 1 h. Cells were metabolically labeled with [35S]methionine(40 mCi/well) for an additional hour. The a1 polypeptide was immu-noprecipitated, separated by SDS–PAGE, and quantitated by auto-radiography. (B) Effect of cycloheximide on a1b1 Na,K-ATPase ac-tivity from PMA- or db-cAMP-treated cells. Sf-9 cells infected withthe a1b1 recombinant virus were treated 40 h after infection with100 mg/ml cycloheximide. After 1 h, 1 mM PMA or 2 mM db-cAMPwas added. After an additional 1 h, Na,K-ATPase activity was de-termined as described in the legend to Fig. 1. Each value is the meanand error bars represent the standard errors of the mean of threeexperiments performed in triplicate on samples obtained from dif-ferent infections. Values significantly different from untreated con-trols are indicated, *P , 0.01.
142 BLANCO, SANCHEZ, AND MERCER
Epson 636 scanner. Intensity of the bands was determined using theNIH Image 1.60 software. Multiple exposures were analyzed to en-sure that the signals were within the linear range of the film.
Inulin uptake. To assess the influence of PK activators on cellu-lar endocytosis, uptake of 14C-labeled inulin was determined. Cells(1 3 106 cells/ml) were harvested 48 h after infection and treatedwith or without 1 mM PMA for 1 h. Uptake was initiated by theaddition of [14C]inulin (50 mCi/ml). Cells were incubated at roomtemperature. A the indicated time intervals, samples (100 ml) werelayered on 0.5 ml of silicone oil (Versilube F-50; Harwich ChemicalCorp.) in 1.5-ml microcentrifuge tubes and centrifuged for 30 s. Theaqueous and oil phases were removed by aspiration and discarded.The cell pellets were excised and radioactivity was determined byliquid scintillation.
Data analysis. All results are expressed as the mean 6 standarderror of the mean. Statistical comparison of the differences betweengroups was performed using the Student’s t test. Statistical signifi-cance was defined as P , 0.05.
RESULTS
Regulation by phosphorylation/dephosphorylation iscommon to many proteins and has been demonstratedfor the Na,K-ATPase (22, 51–53). Although there arevariations depending on the cell type, activation ofPKC and PKA appears to be generally associated withinhibition of the Na,K-ATPase a1b1 isozyme (16). Todetermine if the other isozymes of the Na,K-ATPaseare susceptible to modulation by second messengers,Sf-9 cells were infected with baculoviruses coding forthe rat a1b1, a2b1 or a3b1 isozymes. Forty hours afterinfection, cells were treated for 1 h with PMA or db-cAMP, activators of PKC and PKA respectively, andthe Na,K-ATPase activity was determined. Prelimi-
nary experiments demonstrated that the effects ofPMA or db-cAMP on Na,K-ATPase activity were ap-parent after 15 min treatment and were maximal at 30min. Although expression of the exogenous proteins issubmaximal, experiments were performed at 40 hpostinfection because at this time the cells exhibit min-imal cytopathic effects and are capable of performingall posttranslational protein modifications (data notshown). Figure 1 shows that stimulation of PKC sig-nificantly decreases the specific activity of all three aisoforms compared to that of the untreated controls.The effect of PMA could be partially prevented if thecells were treated with the PKC inhibitors, stauros-porin, or H7. Moreover, the Na,K-ATPase activity of allthree a isoforms can be recovered by treatment withprotein phosphatase 2A, suggesting that phosphoryla-tion is involved in the modulation of the activity of theNa,K-ATPase isozymes. The effect of PKA activationon the activity of the Na pump isozymes is shown inFig. 2. While PKA activation decreased the activity ofthe a1b1 and a2b1 isozymes, interestingly, it increasedthe activity of a3b1. Specificity of the response is dem-onstrated by the reversibility of the effects by the PKAcompetitive inhibitor, 8-bromoadenosine cyclic mono-phosphothioate Rp-isomer (Fig. 2). Taken together,these results indicate that the activity of the Na pumpisozymes can be regulated by activation of PKC andPKA.
To determine if activation of the protein kinasesalters Na,K-ATPase activity by modifying the synthe-
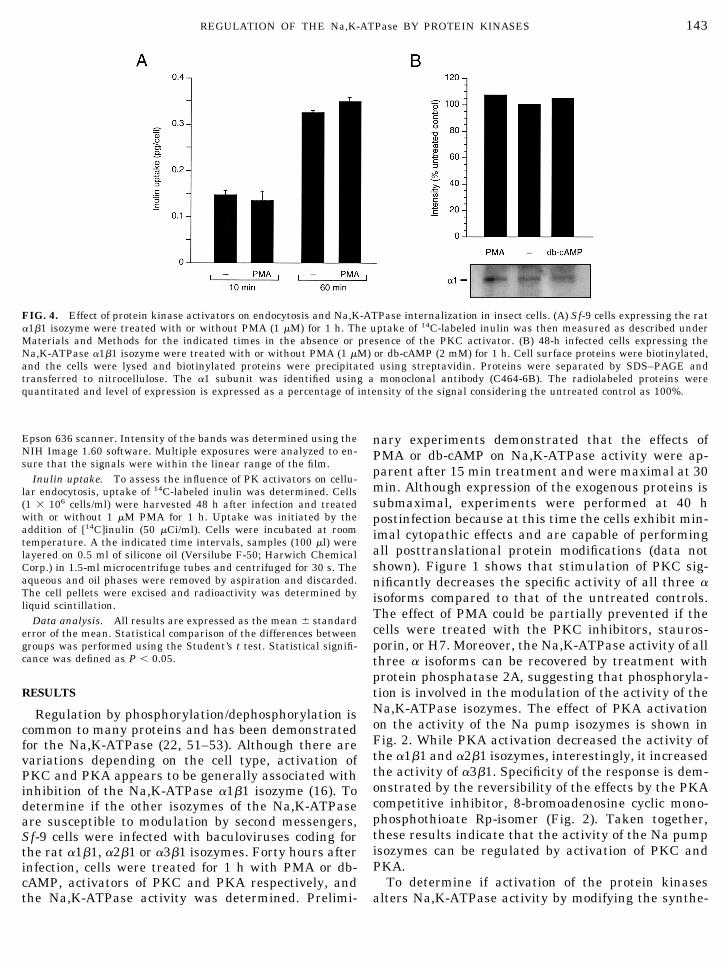
FIG. 4. Effect of protein kinase activators on endocytosis and Na,K-ATPase internalization in insect cells. (A) Sf-9 cells expressing the rata1b1 isozyme were treated with or without PMA (1 mM) for 1 h. The uptake of 14C-labeled inulin was then measured as described underMaterials and Methods for the indicated times in the absence or presence of the PKC activator. (B) 48-h infected cells expressing theNa,K-ATPase a1b1 isozyme were treated with or without PMA (1 mM) or db-cAMP (2 mM) for 1 h. Cell surface proteins were biotinylated,and the cells were lysed and biotinylated proteins were precipitated using streptavidin. Proteins were separated by SDS–PAGE andtransferred to nitrocellulose. The a1 subunit was identified using a monoclonal antibody (C464-6B). The radiolabeled proteins werequantitated and level of expression is expressed as a percentage of intensity of the signal considering the untreated control as 100%.
143REGULATION OF THE Na,K-ATPase BY PROTEIN KINASES
sis or processing of the nascent pump polypeptides,Na,K-ATPase activity was measured after PKC andPKA activation in the absence or the presence of cyclo-heximide. As shown in Fig. 3A, treatment of a1b1-infected cells with 100 mg/ml of cycloheximide com-pletely blocks the synthesis of Na,K-ATPase polypep-tides. However, inhibition of protein synthesis has noeffect on the abilities of PMA and db-cAMP to modifyNa,K-ATPase activity. Thus, the protein kinases arealtering activity of the mature Na pump withoutchanging the processing or rate of synthesis of theprotein. In distal nephron cells from Xenopus laevis, ithas been proposed that the decrease in Na pump ac-tivity after treatment with PMA is the result of theincreased rates of internalization and degradation ofthe Na pump polypeptides (54). In contrast, thechanges in activity found in the insect cells suggest a
direct effect of the protein kinases on the Na,K-ATPaseisozymes since alamethicin is able to fully unmask theNa,K-ATPase activity from the cell plasma and intra-cellular membrane compartments (data not shown). Todirectly determine if simulation of protein kinases re-sult in a more rapid internalization of the Na,K-ATPase in the insect cells, the effect of PKC and PKAactivation on insect cell endocytosis and Na,K-ATPaseinternalization was investigated. Using 14C-labeled in-ulin as a marker for fluid-phase endocytosis, the up-take of the labeled polysaccharide in untreated andPMA-treated Sf-9 cells expressing the Na,K-ATPasea1b1 isozyme was determined (Fig. 4A). As shown, thePKC activator had no effect on the internalization of14C-labeled inulin by the insect cells. To investigate ifactivation of protein kinases specifically modifiedNa,K-ATPase endocytosis, the amount of the enzymeat the cell surface in the absence and presence of PKactivators was studied. For this, 48-h infected cellsexpressing the a1b1 polypeptides were treated withPMA and db-cAMP for 1 h. After biotinylation of cellsurface proteins and streptavidin precipitation of thebiotinylated proteins, the amount of a1 subunit at thecell surface was determined. Figure 4B shows that PKactivators do not significantly affect the level of theNa,K-ATPase at the plasma membrane. Consequently,in the insect cells it appears that endocytosis is notinvolved in the change in Na pump activity elicited byactivation of PKA and PKC.
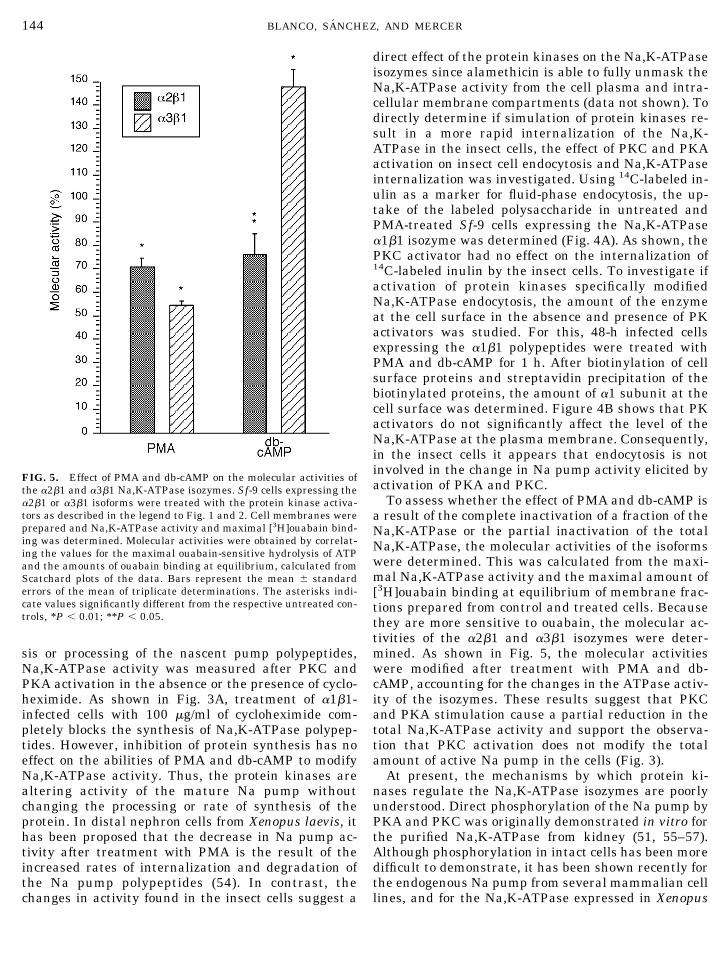
To assess whether the effect of PMA and db-cAMP isa result of the complete inactivation of a fraction of theNa,K-ATPase or the partial inactivation of the totalNa,K-ATPase, the molecular activities of the isoformswere determined. This was calculated from the maxi-mal Na,K-ATPase activity and the maximal amount of[3H]ouabain binding at equilibrium of membrane frac-tions prepared from control and treated cells. Becausethey are more sensitive to ouabain, the molecular ac-tivities of the a2b1 and a3b1 isozymes were deter-mined. As shown in Fig. 5, the molecular activitieswere modified after treatment with PMA and db-cAMP, accounting for the changes in the ATPase activ-ity of the isozymes. These results suggest that PKCand PKA stimulation cause a partial reduction in thetotal Na,K-ATPase activity and support the observa-tion that PKC activation does not modify the totalamount of active Na pump in the cells (Fig. 3).
At present, the mechanisms by which protein ki-nases regulate the Na,K-ATPase isozymes are poorlyunderstood. Direct phosphorylation of the Na pump byPKA and PKC was originally demonstrated in vitro forthe purified Na,K-ATPase from kidney (51, 55–57).Although phosphorylation in intact cells has been moredifficult to demonstrate, it has been shown recently forthe endogenous Na pump from several mammalian celllines, and for the Na,K-ATPase expressed in Xenopus
FIG. 5. Effect of PMA and db-cAMP on the molecular activities ofthe a2b1 and a3b1 Na,K-ATPase isozymes. Sf-9 cells expressing thea2b1 or a3b1 isoforms were treated with the protein kinase activa-tors as described in the legend to Fig. 1 and 2. Cell membranes wereprepared and Na,K-ATPase activity and maximal [3H]ouabain bind-ing was determined. Molecular activities were obtained by correlat-ing the values for the maximal ouabain-sensitive hydrolysis of ATPand the amounts of ouabain binding at equilibrium, calculated fromScatchard plots of the data. Bars represent the mean 6 standarderrors of the mean of triplicate determinations. The asterisks indi-cate values significantly different from the respective untreated con-trols, *P , 0.01; **P , 0.05.
144 BLANCO, SANCHEZ, AND MERCER
oocytes (24, 52, 53, 58, 59). Moreover, in kidney cells,the kinases affect Na pump function not only throughdirect phosphorylation of the a subunit, but also indi-rectly by activation of the PLA2 pathway (60).
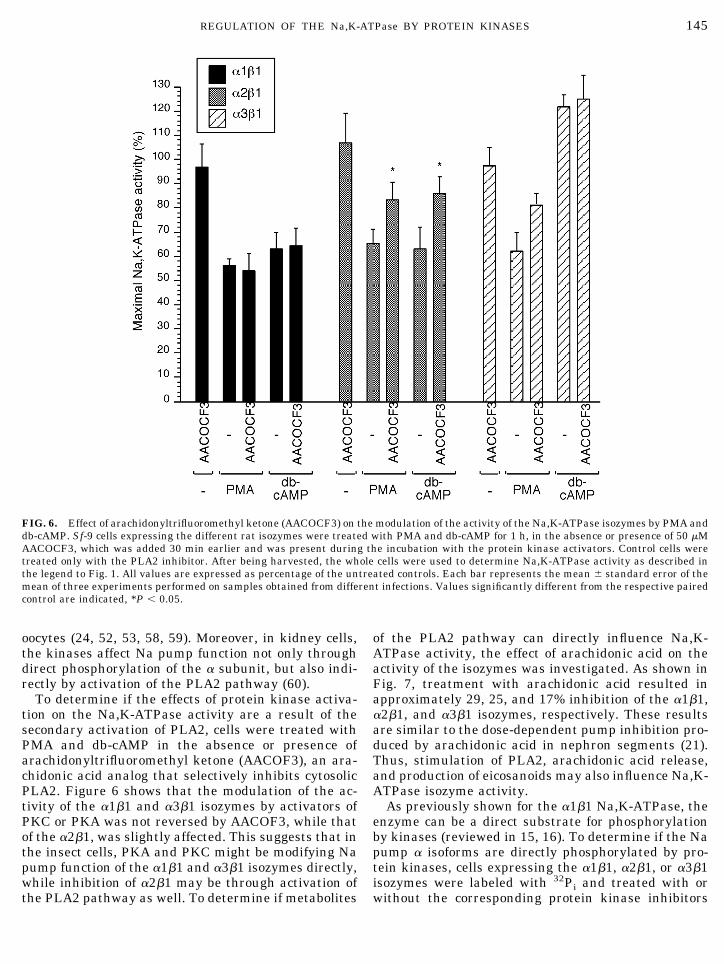
To determine if the effects of protein kinase activa-tion on the Na,K-ATPase activity are a result of thesecondary activation of PLA2, cells were treated withPMA and db-cAMP in the absence or presence ofarachidonyltrifluoromethyl ketone (AACOF3), an ara-chidonic acid analog that selectively inhibits cytosolicPLA2. Figure 6 shows that the modulation of the ac-tivity of the a1b1 and a3b1 isozymes by activators ofPKC or PKA was not reversed by AACOF3, while thatof the a2b1, was slightly affected. This suggests that inthe insect cells, PKA and PKC might be modifying Napump function of the a1b1 and a3b1 isozymes directly,while inhibition of a2b1 may be through activation ofthe PLA2 pathway as well. To determine if metabolites
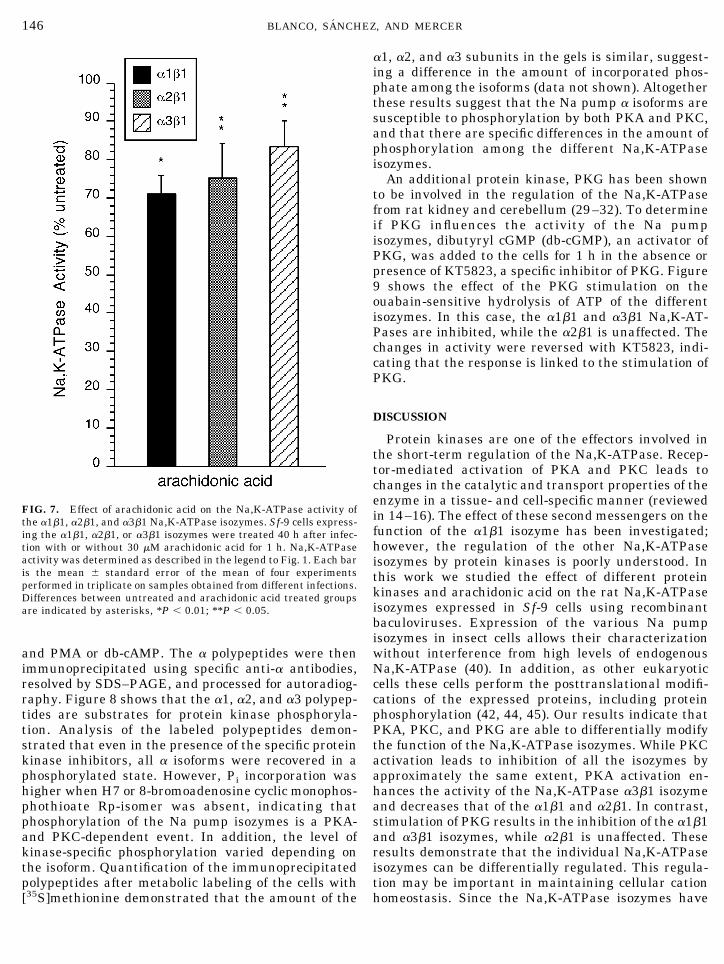
of the PLA2 pathway can directly influence Na,K-ATPase activity, the effect of arachidonic acid on theactivity of the isozymes was investigated. As shown inFig. 7, treatment with arachidonic acid resulted inapproximately 29, 25, and 17% inhibition of the a1b1,a2b1, and a3b1 isozymes, respectively. These resultsare similar to the dose-dependent pump inhibition pro-duced by arachidonic acid in nephron segments (21).Thus, stimulation of PLA2, arachidonic acid release,and production of eicosanoids may also influence Na,K-ATPase isozyme activity.
As previously shown for the a1b1 Na,K-ATPase, theenzyme can be a direct substrate for phosphorylationby kinases (reviewed in 15, 16). To determine if the Napump a isoforms are directly phosphorylated by pro-tein kinases, cells expressing the a1b1, a2b1, or a3b1isozymes were labeled with 32Pi and treated with orwithout the corresponding protein kinase inhibitors
FIG. 6. Effect of arachidonyltrifluoromethyl ketone (AACOCF3) on the modulation of the activity of the Na,K-ATPase isozymes by PMA anddb-cAMP. Sf-9 cells expressing the different rat isozymes were treated with PMA and db-cAMP for 1 h, in the absence or presence of 50 mMAACOCF3, which was added 30 min earlier and was present during the incubation with the protein kinase activators. Control cells weretreated only with the PLA2 inhibitor. After being harvested, the whole cells were used to determine Na,K-ATPase activity as described inthe legend to Fig. 1. All values are expressed as percentage of the untreated controls. Each bar represents the mean 6 standard error of themean of three experiments performed on samples obtained from different infections. Values significantly different from the respective pairedcontrol are indicated, *P , 0.05.
145REGULATION OF THE Na,K-ATPase BY PROTEIN KINASES
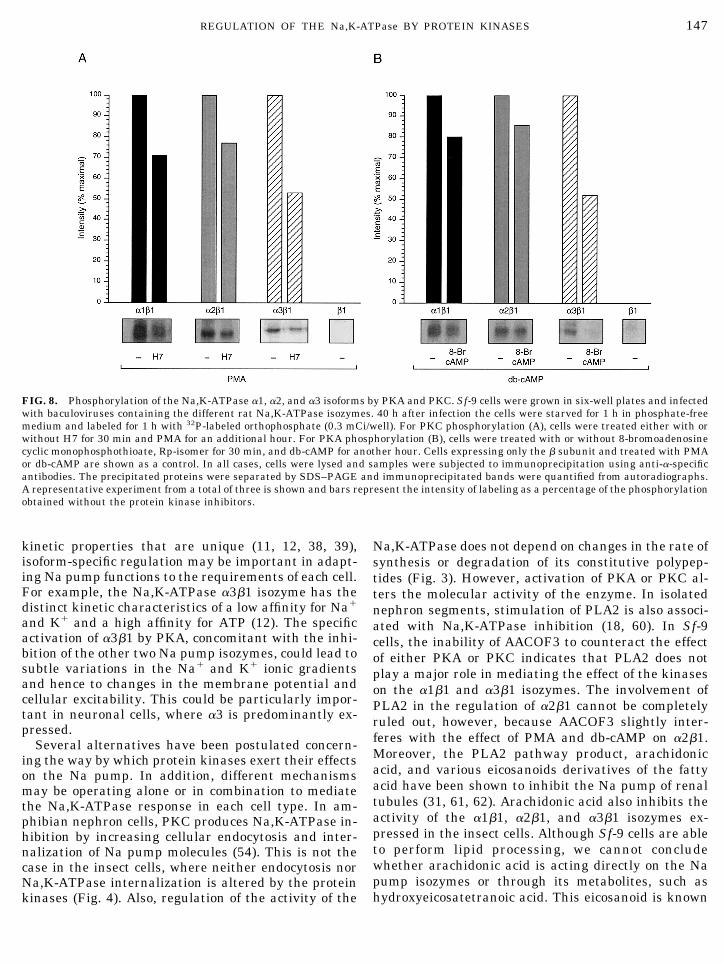
and PMA or db-cAMP. The a polypeptides were thenimmunoprecipitated using specific anti-a antibodies,resolved by SDS–PAGE, and processed for autoradiog-raphy. Figure 8 shows that the a1, a2, and a3 polypep-tides are substrates for protein kinase phosphoryla-tion. Analysis of the labeled polypeptides demon-strated that even in the presence of the specific proteinkinase inhibitors, all a isoforms were recovered in aphosphorylated state. However, Pi incorporation washigher when H7 or 8-bromoadenosine cyclic monophos-phothioate Rp-isomer was absent, indicating thatphosphorylation of the Na pump isozymes is a PKA-and PKC-dependent event. In addition, the level ofkinase-specific phosphorylation varied depending onthe isoform. Quantification of the immunoprecipitatedpolypeptides after metabolic labeling of the cells with[35S]methionine demonstrated that the amount of the
a1, a2, and a3 subunits in the gels is similar, suggest-ing a difference in the amount of incorporated phos-phate among the isoforms (data not shown). Altogetherthese results suggest that the Na pump a isoforms aresusceptible to phosphorylation by both PKA and PKC,and that there are specific differences in the amount ofphosphorylation among the different Na,K-ATPaseisozymes.
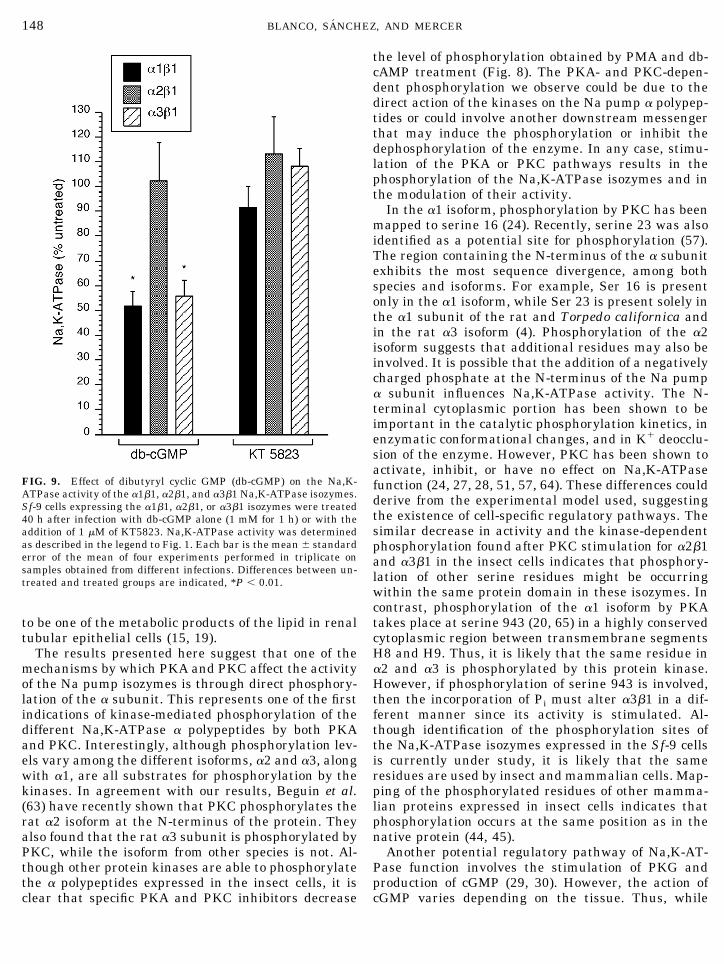
An additional protein kinase, PKG has been shownto be involved in the regulation of the Na,K-ATPasefrom rat kidney and cerebellum (29–32). To determineif PKG influences the activity of the Na pumpisozymes, dibutyryl cGMP (db-cGMP), an activator ofPKG, was added to the cells for 1 h in the absence orpresence of KT5823, a specific inhibitor of PKG. Figure9 shows the effect of the PKG stimulation on theouabain-sensitive hydrolysis of ATP of the differentisozymes. In this case, the a1b1 and a3b1 Na,K-AT-Pases are inhibited, while the a2b1 is unaffected. Thechanges in activity were reversed with KT5823, indi-cating that the response is linked to the stimulation ofPKG.
DISCUSSION
Protein kinases are one of the effectors involved inthe short-term regulation of the Na,K-ATPase. Recep-tor-mediated activation of PKA and PKC leads tochanges in the catalytic and transport properties of theenzyme in a tissue- and cell-specific manner (reviewedin 14–16). The effect of these second messengers on thefunction of the a1b1 isozyme has been investigated;however, the regulation of the other Na,K-ATPaseisozymes by protein kinases is poorly understood. Inthis work we studied the effect of different proteinkinases and arachidonic acid on the rat Na,K-ATPaseisozymes expressed in Sf-9 cells using recombinantbaculoviruses. Expression of the various Na pumpisozymes in insect cells allows their characterizationwithout interference from high levels of endogenousNa,K-ATPase (40). In addition, as other eukaryoticcells these cells perform the posttranslational modifi-cations of the expressed proteins, including proteinphosphorylation (42, 44, 45). Our results indicate thatPKA, PKC, and PKG are able to differentially modifythe function of the Na,K-ATPase isozymes. While PKCactivation leads to inhibition of all the isozymes byapproximately the same extent, PKA activation en-hances the activity of the Na,K-ATPase a3b1 isozymeand decreases that of the a1b1 and a2b1. In contrast,stimulation of PKG results in the inhibition of the a1b1and a3b1 isozymes, while a2b1 is unaffected. Theseresults demonstrate that the individual Na,K-ATPaseisozymes can be differentially regulated. This regula-tion may be important in maintaining cellular cationhomeostasis. Since the Na,K-ATPase isozymes have
FIG. 7. Effect of arachidonic acid on the Na,K-ATPase activity ofthe a1b1, a2b1, and a3b1 Na,K-ATPase isozymes. Sf-9 cells express-ing the a1b1, a2b1, or a3b1 isozymes were treated 40 h after infec-tion with or without 30 mM arachidonic acid for 1 h. Na,K-ATPaseactivity was determined as described in the legend to Fig. 1. Each baris the mean 6 standard error of the mean of four experimentsperformed in triplicate on samples obtained from different infections.Differences between untreated and arachidonic acid treated groupsare indicated by asterisks, *P , 0.01; **P , 0.05.
146 BLANCO, SANCHEZ, AND MERCER
kinetic properties that are unique (11, 12, 38, 39),isoform-specific regulation may be important in adapt-ing Na pump functions to the requirements of each cell.For example, the Na,K-ATPase a3b1 isozyme has thedistinct kinetic characteristics of a low affinity for Na1
and K1 and a high affinity for ATP (12). The specificactivation of a3b1 by PKA, concomitant with the inhi-bition of the other two Na pump isozymes, could lead tosubtle variations in the Na1 and K1 ionic gradientsand hence to changes in the membrane potential andcellular excitability. This could be particularly impor-tant in neuronal cells, where a3 is predominantly ex-pressed.
Several alternatives have been postulated concern-ing the way by which protein kinases exert their effectson the Na pump. In addition, different mechanismsmay be operating alone or in combination to mediatethe Na,K-ATPase response in each cell type. In am-phibian nephron cells, PKC produces Na,K-ATPase in-hibition by increasing cellular endocytosis and inter-nalization of Na pump molecules (54). This is not thecase in the insect cells, where neither endocytosis norNa,K-ATPase internalization is altered by the proteinkinases (Fig. 4). Also, regulation of the activity of the
Na,K-ATPase does not depend on changes in the rate ofsynthesis or degradation of its constitutive polypep-tides (Fig. 3). However, activation of PKA or PKC al-ters the molecular activity of the enzyme. In isolatednephron segments, stimulation of PLA2 is also associ-ated with Na,K-ATPase inhibition (18, 60). In Sf-9cells, the inability of AACOF3 to counteract the effectof either PKA or PKC indicates that PLA2 does notplay a major role in mediating the effect of the kinaseson the a1b1 and a3b1 isozymes. The involvement ofPLA2 in the regulation of a2b1 cannot be completelyruled out, however, because AACOF3 slightly inter-feres with the effect of PMA and db-cAMP on a2b1.Moreover, the PLA2 pathway product, arachidonicacid, and various eicosanoids derivatives of the fattyacid have been shown to inhibit the Na pump of renaltubules (31, 61, 62). Arachidonic acid also inhibits theactivity of the a1b1, a2b1, and a3b1 isozymes ex-pressed in the insect cells. Although Sf-9 cells are ableto perform lipid processing, we cannot concludewhether arachidonic acid is acting directly on the Napump isozymes or through its metabolites, such ashydroxyeicosatetranoic acid. This eicosanoid is known
FIG. 8. Phosphorylation of the Na,K-ATPase a1, a2, and a3 isoforms by PKA and PKC. Sf-9 cells were grown in six-well plates and infectedwith baculoviruses containing the different rat Na,K-ATPase isozymes. 40 h after infection the cells were starved for 1 h in phosphate-freemedium and labeled for 1 h with 32P-labeled orthophosphate (0.3 mCi/well). For PKC phosphorylation (A), cells were treated either with orwithout H7 for 30 min and PMA for an additional hour. For PKA phosphorylation (B), cells were treated with or without 8-bromoadenosinecyclic monophosphothioate, Rp-isomer for 30 min, and db-cAMP for another hour. Cells expressing only the b subunit and treated with PMAor db-cAMP are shown as a control. In all cases, cells were lysed and samples were subjected to immunoprecipitation using anti-a-specificantibodies. The precipitated proteins were separated by SDS–PAGE and immunoprecipitated bands were quantified from autoradiographs.A representative experiment from a total of three is shown and bars represent the intensity of labeling as a percentage of the phosphorylationobtained without the protein kinase inhibitors.
147REGULATION OF THE Na,K-ATPase BY PROTEIN KINASES
to be one of the metabolic products of the lipid in renaltubular epithelial cells (15, 19).
The results presented here suggest that one of themechanisms by which PKA and PKC affect the activityof the Na pump isozymes is through direct phosphory-lation of the a subunit. This represents one of the firstindications of kinase-mediated phosphorylation of thedifferent Na,K-ATPase a polypeptides by both PKAand PKC. Interestingly, although phosphorylation lev-els vary among the different isoforms, a2 and a3, alongwith a1, are all substrates for phosphorylation by thekinases. In agreement with our results, Beguin et al.(63) have recently shown that PKC phosphorylates therat a2 isoform at the N-terminus of the protein. Theyalso found that the rat a3 subunit is phosphorylated byPKC, while the isoform from other species is not. Al-though other protein kinases are able to phosphorylatethe a polypeptides expressed in the insect cells, it isclear that specific PKA and PKC inhibitors decrease
the level of phosphorylation obtained by PMA and db-cAMP treatment (Fig. 8). The PKA- and PKC-depen-dent phosphorylation we observe could be due to thedirect action of the kinases on the Na pump a polypep-tides or could involve another downstream messengerthat may induce the phosphorylation or inhibit thedephosphorylation of the enzyme. In any case, stimu-lation of the PKA or PKC pathways results in thephosphorylation of the Na,K-ATPase isozymes and inthe modulation of their activity.
In the a1 isoform, phosphorylation by PKC has beenmapped to serine 16 (24). Recently, serine 23 was alsoidentified as a potential site for phosphorylation (57).The region containing the N-terminus of the a subunitexhibits the most sequence divergence, among bothspecies and isoforms. For example, Ser 16 is presentonly in the a1 isoform, while Ser 23 is present solely inthe a1 subunit of the rat and Torpedo californica andin the rat a3 isoform (4). Phosphorylation of the a2isoform suggests that additional residues may also beinvolved. It is possible that the addition of a negativelycharged phosphate at the N-terminus of the Na pumpa subunit influences Na,K-ATPase activity. The N-terminal cytoplasmic portion has been shown to beimportant in the catalytic phosphorylation kinetics, inenzymatic conformational changes, and in K1 deocclu-sion of the enzyme. However, PKC has been shown toactivate, inhibit, or have no effect on Na,K-ATPasefunction (24, 27, 28, 51, 57, 64). These differences couldderive from the experimental model used, suggestingthe existence of cell-specific regulatory pathways. Thesimilar decrease in activity and the kinase-dependentphosphorylation found after PKC stimulation for a2b1and a3b1 in the insect cells indicates that phosphory-lation of other serine residues might be occurringwithin the same protein domain in these isozymes. Incontrast, phosphorylation of the a1 isoform by PKAtakes place at serine 943 (20, 65) in a highly conservedcytoplasmic region between transmembrane segmentsH8 and H9. Thus, it is likely that the same residue ina2 and a3 is phosphorylated by this protein kinase.However, if phosphorylation of serine 943 is involved,then the incorporation of Pi must alter a3b1 in a dif-ferent manner since its activity is stimulated. Al-though identification of the phosphorylation sites ofthe Na,K-ATPase isozymes expressed in the Sf-9 cellsis currently under study, it is likely that the sameresidues are used by insect and mammalian cells. Map-ping of the phosphorylated residues of other mamma-lian proteins expressed in insect cells indicates thatphosphorylation occurs at the same position as in thenative protein (44, 45).
Another potential regulatory pathway of Na,K-AT-Pase function involves the stimulation of PKG andproduction of cGMP (29, 30). However, the action ofcGMP varies depending on the tissue. Thus, while
FIG. 9. Effect of dibutyryl cyclic GMP (db-cGMP) on the Na,K-ATPase activity of the a1b1, a2b1, and a3b1 Na,K-ATPase isozymes.Sf-9 cells expressing the a1b1, a2b1, or a3b1 isozymes were treated40 h after infection with db-cGMP alone (1 mM for 1 h) or with theaddition of 1 mM of KT5823. Na,K-ATPase activity was determinedas described in the legend to Fig. 1. Each bar is the mean 6 standarderror of the mean of four experiments performed in triplicate onsamples obtained from different infections. Differences between un-treated and treated groups are indicated, *P , 0.01.
148 BLANCO, SANCHEZ, AND MERCER

cGMP mediates inhibition of the Na,K-ATPase in thekidney (30), it stimulates ouabain-sensitive p-nitrophe-nylphosphatase hydrolysis by the Na pump in ducksalt gland (31) and Na,K-ATPase activity in rat cere-bellum (32). We investigated whether PKG could influ-ence the function of the rat Na,K-ATPase isozymesexpressed in insect cells. Our results demonstrate that,when expressed in a common environment, the Napump isozymes are differentially affected by activationof PKG. Whereas the activity of the a1b1 and a3b1isozymes diminishes, that of the a2b1 remains unaf-fected.
In conclusion, the activity of the Na,K-ATPase isunder the control of a variety of intracellular messen-gers that are able to modulate the function of theindividual isozymes in a specific fashion. This consti-tutes additional evidence supporting the notion thatthe Na pump isozymes, rather than being redundant,have specific physiological roles. Because the variousNa,K-ATPases possess unique catalytic properties,their differential regulation may be essential in adapt-ing Na pump function to the requirements of each cell.
ACKNOWLEDGMENTS
We thank Drs. Michael Caplan and Douglas Fambrough for kindlysupplying the antibodies used in this work. This work was supportedby National Institutes of Health grant GM 39746, DK 45181, GeorgeO’Brien Kidney and Urological Diseases Center at Washington Uni-versity School of Medicine and American Heart Association grant96012080.
REFERENCES
1. Skou, J. C., and Esmann, M. (1992) J. Bioenerg. Biomembr. 24,249–261.
2. Glynn, I. M. (1985) in The Enzymes of Biological Membranes(Martonosi, A., Ed.), 2nd ed., Vol. 3, pp. 35–114, Plenum, NewYork.
3. Glynn, I. M. (1993) J. Physiol. 462, 1–30.4. Mercer, R. W. (1993) Int. Rev. Cytol. 137C, 139–168.5. Lingrel, J. B (1994) J. Biol. Chem. 269, 19659–19662.6. Noguchi, S., Mishina, M., Kawamura, M., and Numa, S. (1987)
FEBS Lett. 225, 27–32.7. Horowitz, B., Eakle, K, Scheiner-Bobis, A. G., Randolph, G. R,
Chen, C. Y., Hitzeman R. A., and Farley, R. A. (1991) J. Biol.Chem. 265, 4189–4192.
8. DeTomaso, A. W., Xie, Z. J., Liu, G., and Mercer, R. W. (1993)J. Biol. Chem. 268, 1470–1478.
9. Lutsenko, S., and Kaplan, J. H. (1992) Ann. NY Acad. Sci. 671,147–154.
10. Eakle, K. A., Lyu, R. M., and Farley, R. A. (1995) J. Biol. Chem.270, 13937–13947.
11. Blanco, G., Koster, J., Sanchez, G., and Mercer, R. W. (1995)Biochemistry 34, 319–325.
12. Blanco, G., Sanchez, G., and Mercer, R. W. (1995) Biochemistry34, 9897–9903.
13. Jaisser, F., Horisberger, J. D., and Rossier, B. C. (1992) Ann. NYAcad. Sci. 671, 113–119.
14. McDonough, A. A., and Farley, R. A. (1993) Curr. Opin. Nephrol.Hyper. 2, 725–734.
15. Bertorello, A. M., and Katz, A. I. (1993) Am. J. Physiol. 265,F743–F755.
16. Ewart, H. S., and Klip, A. (1995) Am. J. Physiol. 269, C295–C311.
17. Horiuchi, A., Takeyasu, K., Mouradian, M. M., Jose, P. A., andFelder, R. A. (1993). Mol. Pharmacol. 43, 281–285.
18. Satoh, T., Cohen, H. T., and Katz, A. I. (1993) Am. J. Physiol.265, F399–F405.
19. Bertorello, A. M., and Katz, A. I. (1995) NIPS 10, 253–259.20. Fisone, G., Cheng, S. X. J., Nairn, A. C., Czernik, A. J., Hem-
mings, H. C., Jr., Hoog, J. O., Bertorello, A. M., Kaiser, R.,Bergman, T., Jornvall, H., Aperia, A., and Greengard, P. (1994)J. Biol. Chem. 269, 9368–9373.
21. Satoh, T., Cohen, H. T., and Katz, A. I. (1993) J. Clin. Invest. 91,409–415.
22. Aperia, A., Fryckstedt, J., Svensson, L., Hemmings, H. C., Jr.,Nairn, A. C., and Greengard, P. (1991) Proc. Natl. Acad. Sci.USA 88, 2798–2801.
23. Li, D., Aperia, A., Celsi, G., Da Cruz e Silva, E. F., Greengard, P.,and Meister, B. (1995) Am. J. Physiol. 269, F673–F680.
24. Beguin, P., Beggah, A. T., Chibalin, A. V., Burguener-Kairuz, P.,Jaisser, F., Matheus, P. M., Rossier, B. C., Cotecchia, S., andGeering, K. (1994) J. Biol. Chem. 269, 24437–24445.
25. Shahedi, M., Laborde, K., Bussieres, L., Dechaux, M., and Sachs,C. (1992) Pfluggers Arch. 420, 269–274.
26. Garg, I. C., Saha, P. K., and Mohuczy-Dominiak, D. (1993)J. Am. Soc. Nephrol. 4, 195–205.
27. Pedemonte, C. H., Pressley, T. A., Cinelli, A. R., and Lokhand-wala, M. F. (1997) Mol. Pharmacol. 52, 88–97.
28. Pedemonte, C. H., Pressley, T. A., Lokhandwala, M. F., andCinelli, A. R. (1997) J. Membr. Biol. 155, 219–227.
29. McKee, M., Scavone, C., and Nathanson, J. A. (1994) Proc. Natl.Acad. Sci. USA 91, 12056–12060.
30. Scavone, C., Scanlon, C., McKee, M., and Nathanson, J. A. (1995)J. Pharmacol. Exp. Ther. 272, 1036–1043.
31. Stewart, D. J., and Sen, A. K. (1981) Am. J. Physiol. 240, C207–C214.
32. Nathanson, J. A., Scavone, C., Scanlon, C., and McKee, M. (1995)Neuron 14, 781–794.
33. Orlowski, J., and Lingrel, J. B. (1988) J. Biol. Chem. 263, 10436–10442.
34. Sweadner, K. J. (1989) Biochim. Biophys. Acta 988, 185–220.35. Lingrel, J. B, Orlowsky, J., Shull, M., and Price, E. M. (1990)
Prog. Nucleic Acid Res. 38, 37–89.36. Lingrel, J. B (1992) J. Bioenerg. Biomembr. 24, 263–269.37. Levenson, R. (1994) Rev. Physiol. Biochem. Pharmacol. 123,
1–45.38. Jewell, E. A., and Lingrel J. B. (1991) J. Biol. Chem. 266, 16125–
16130.39. Daly, S. E., Lane, L. K., and Blostein, R. (1994) J. Biol. Chem.
269, 23944–23948.40. Blanco, G., Xie, Z. J., and Mercer, R. W. (1993) Proc. Natl. Acad.
Sci. USA 90, 1824–1828.41. O’Reilly, D. R., Miller, L. K., and Luckow, V. A. (1992) in Bacu-
lovirus Expression Vectors. A Laboratory Manual, Freeman,New York.
42. Kozma, S. C., McGlynn, E., Siegmann, M., Reinhard, C., Ferrari,S., and Thomas, G. (1993) J. Biol. Chem. 268, 7134–7138.
149REGULATION OF THE Na,K-ATPase BY PROTEIN KINASES

43. Jacobowitz, O., and Iyengar, R. (1994) Proc. Natl. Acad. Sci. USA91, 10630–10634.
44. Fuchs, B., Hecker, D., and Scheidtmann, K. H. (1995). Eur.J. Biochem. 228, 625–639.
45. Beck, C. A., Zhang, Y., Altmann, M., Weigel, N. L., and Edwards,D. P. (1996) J. Biol. Chem. 271, 19696–19555.
46. Mercer, R. W., Schneider, J. W., Savitz, A., Emanuel, J. R.,Benz, E. J., Jr., and Levenson, R. (1986) Mol. Cell. Biol. 6,3884 –3890.
47. Schneider, J. W., Mercer, R. W., Gilmore-Herbert, M., Utset,M. F., Lai, C., Greene, A., and Benz, E. J. Jr. (1988) Proc. Natl.Acad. Sci. USA 85, 284–288.
48. Koster, J. C., Blanco, G., Mills, P. B., and Mercer, R. W. (1996)J. Biol. Chem. 271, 2413–2421.
49. Laemmli, U. K. (1970) Nature 227, 680–685.50. Gottardi, C. A., Dunbar, L. A. and Caplan, M. J. (1995) Am. J.
Physiol. 268, F285–F295.51. Bertorello, A. M., Aperia, A., Walaas, S. I., Nairn, A. C., and
Greengard, P. (1991) Proc. Natl. Acad. Sci. USA 88, 11359–11362.
52. Chibalin, A. V., Vasilets, L. A., Hennekes, H., Pralong, D., andGeering, K. (1992) J. Biol. Chem. 267, 22378–22384.
53. Borghini, I., Geering, K., Gjinovci, A., Wollheim, C. B., andPralong, W. F. (1994) Proc. Natl. Acad. Sci. USA 91, 6211–6215.
54. Beron, J., Forster, I., Beguin, P., Geering, K., and Verrey, F.(1997) Mol. Biol. Cell 8, 387–398.
55. Lowndes, J. M., Hokin Neaverson, M., and Bertics, P. J. (1990)Biochim. Biophys. Acta 1052, 143–151.
56. Feschenko, M. S., and Sweadner, K. J. (1994) J. Biol. Chem. 269,30436–30444.
57. Feschenko, M. S., and Sweadner, K. J. (1995) J. Biol. Chem. 270,14072–14077.
58. Middleton, J. P., Khan, W. A., Collinsworth, G., Hannun, Y. A.,and Medford, R. M. (1993) J. Biol. Chem. 268, 15958–15964.
59. Feschenko, M. S., and Sweadner, K. J. (1997) J. Biol. Chem. 272,17726–17733.
60. Satoh, T., Cohen, H. T., and Katz, A. I. (1992) J. Clin. Invest. 89,1496–1500.
61. Schwartzman, M., Ferreri, N. R., Carroll, M. A., Songu-Mize, E.,and McGiff, J. C. (1985) Nature 314, 620–622.
62. Escalante, B., Erlij, D., Falck, J. R., and McGiff, J. C. (1991)Science 251, 799–802.
63. Beguin, P., Peitisch, M., and Geering, K. (1996) Biochemistry 35,14098–14108.
64. Carranza, M. L., Feraille, E., and Favre, H. (1996) Am. J.Physiol. 271, C136–C143.
65. Cheng, X.-J., Hoog, J.-O., Nairn, A. C., Greengard, P., and Ape-ria, A. (1997) Am. J. Physiol. 273, C1981–C1986.
150 BLANCO, SANCHEZ, AND MERCER


![Prevention of doxorubicin-induce renal function abnormalities ......ATPase, Mg2+-ATPase and Na+, K+-ATPase activities [15, 16]. Turmeric is a golden spice derived from the rhizome](https://img.pdfslide.us/doc/110x75/61385b7c0ad5d20676493447/prevention-of-doxorubicin-induce-renal-function-abnormalities-atpase-mg2-atpase.jpg)