Embed Size (px)
Citation preview

Da
LD
h
����
a
ARR2A
KGPCGH
1
(b
pPf
(
F
0h
Neuroscience Letters 534 (2013) 199– 204
Contents lists available at SciVerse ScienceDirect
Neuroscience Letters
jou rn al h om epage: www.elsev ier .com/ locate /neule t
ifferential immunoreactivity of glucocorticoid receptor and GABA in GABAergicfferents to parvocellular neurons in the paraventricular nucleus
eandro Marques de Souza1, Celso Rodrigues Franci ∗
epartamento de Fisiologia, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Av. Bandeirantes-3900, Ribeirão Preto, SP 14049-900, Brazil
i g h l i g h t s
Glucocorticoid may directly act on GABAergic neurons through GR.Different expressions of GABA and GR in anterior and medial parvocellular regions in the PVH.GABA mediates an inhibitory compensatory mechanism to modulate the HPA axis.Anterior periventricular nucleus relays an indirect inhibition to parvocellular neurons and HPA.
r t i c l e i n f o
rticle history:eceived 13 September 2012eceived in revised form9 November 2012ccepted 10 December 2012
eywords:ABAaraventricular nucleusorticosterone
a b s t r a c t
We assessed the distribution of glucocorticoid receptors (GR), GABA (�-aminobutyric acid) neuronsand co-localization of GR in GABA-positive neurons for four hypothalamic sources of GABAergic pro-jections to the parvocellular neurons in the paraventricular nucleus (PVH) from normal, sham-surgeryand adrenalectomized male rats subjected to intraperitoneal injections of saline or dexamethasone.Blood samples were collected to measure corticosterone by radioimmunoassay. The distribution of GR,GABA-positive neurons and co-localization of GR in GABA-positive neurons were analyzed by immuno-fluorescence in sections from the paraventricular nucleus (PVH). In intact and sham rats, dexamethasoneinduced expression of GABAergic neurons in the regions of the anterior periventricular nucleus (PVa)coincident with anterior (PVHap) and medial (PVHmp) parvocellular subdivisions of thePVH. However,
lucocorticoid receptorsPA axis
the co-expression of GR in GABAergic neurons was found only in the region of the PVa coincident withPVHmp. These findings confirm that glucocorticoids may directly act on GABAergic neurons throughGR. PVHap and PVHmp present differentiated patterns of GABA and GR expression between then. Theco-localization of GR in GABA-positive neurons in the region of the PVa coincident with PVHmp demon-strates a critic importance of this region to control the hypothalamus–pituitary–adrenal axis throughGABAergic mediation.
© 2013 Elsevier Ireland Ltd. All rights reserved.
. Introduction
Stress activates the hypothalamus–pituitary–adrenal axisHPAa), increasing plasma corticosterone that exerts negative feed-ack control over the HPAa through glucocorticoid receptors (GR) in
Abbreviations: HPAa, hypothalamus–pituitary–adrenal axis; PVHap, anteriorarvocellular region of the PVH; PVHmp, medial parvocellular region of the PVH;Va, anterior periventricular nucleus; SO, peri-supraoptic area; APF, anterior peri-ornical area.∗ Corresponding author. Tel.: +55 16 3602 3022; fax: +55 16 3602 0221.
E-mail addresses: [email protected] (L.M. de Souza), [email protected]. Franci).
1 Permanent address: Universidade Federal de Sergipe, Campus Antônio Garciailho, Rua Padre Alvares Pitangueira-248, Lagarto, SE 49.400-000, Brazil.
304-3940/$ – see front matter © 2013 Elsevier Ireland Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.neulet.2012.12.018
the pituitary and the hypothalamic paraventricular nucleus (PVH)[6–8,17,23]. Parvocellular neurons in the PVH also receive neu-ronal inputs from the brainstem and limbic areas [12,23] througha shell of GABAergic interneurons surrounding the PVH [13,21].This GABAergic input is a strong determinant of HPAa activity [5].The GABAergic influence for corticosterone secretion under stress ismodulated by GABA-A and more specifically GABA-B receptors [19].Previous studies have identified four main sources of GABAergicafferents to the anterior (PVHap) and medial (PVHmp) parvocellu-lar region of the PVH: anterior hypothalamic area (AHA), anteriorperiventricular nucleus (PVa), peri-supraoptic area (SO) and ante-
rior perifornical area (APF) [21]. We assessed the distribution of GR,GABA-positive neurons and GR/GABA neurons for these sources inaccordance with their coincidence with the PVHap (AHA, PVa andSO) and the PVHmp (PVa, SO and APF).
2 oscien
2
2
1fNLCR
2
ttiUsmwatPDs
dogsdduo4wii
2
2
btittosBPsaa
2
imtipw
00 L.M. de Souza, C.R. Franci / Neur
. Materials and methods
.1. Animals
Male Wistar rats weighing 250 ± 10 g were housed under2 h:12 h light–dark cycle (lights on 07:00 a.m.), at 23 ± 1 ◦C withree access to food and water. All protocols in accordance with theational Institutes of Health Guidelines for the Care and Use ofaboratory Animals (USA) were previously approved by the Ethicsommittee for Experiments on Animals of the Medical School ofibeirão Preto, University of Sao Paulo.
.2. Experimental design and surgical procedures
At the end of the fifth day of habituation to housing conditions:wo groups of intact animals received (19:00 h) an intraperi-oneal injection of either saline (0.15 M NaCl; 0.1 mL/100 g b.w.,ntact-saline group) or 2.5% dexamethasone (0.1 mL/100 g, Sigma-SA, intact-Dexa group); other two groups subjected to either
ham surgery or bilateral adrenalectomy (ADX) under tribro-oethanol 2.5% (1 mL/100 g, Aldrich, USA) anesthesia. This surgeryas performed via dorsal incisions to gain access to adrenal
nd remove it or exposed but not excise it (control). Following,he animals received analgesic (50 ug/rat, Benamine®, Schering-lough, Brazil) and antibiotic (0.1 mL/rat, Pentabiotic®, Fortodge, Brazil). Adrenalectomized animals got access to oral 0.9%
aline.At fifth day of surgical recovery (19:00 h), the animals were
ivided in two groups that received intraperitoneal injectionsf either saline (0.15 M NaCl; 0.1 mL/100 g b.w., sham-salineroup/ADX-saline group) or 2.5% dexamethasone (0.1 mL/100 g,ham-Dexa group/ADX-Dexa group). After twelve hours (at sixthay, 7:00 a.m.), the animals were anesthetized and blood wasrawn via an aortic puncture. Plasma samples were stored at ±20 ◦Cntil to measure corticosterone. Following, approximately 30 mLf 0.01 M phosphate-buffered saline (PBS, pH 7.4) and 300 mL of% paraformaldehyde in 0.1 M sodium phosphate buffer (pH 7.4)ere perfused through the ascending aorta. Brains were removed,
mmersed in 30% sucrose in PBS (48 h at 4 ◦C), frozen in cooledsopentane and stored at −70 ◦C until sectioning.
.3. Immunohistochemistry
.3.1. Tissue preparationSerial coronal sections (20 �m-thick) (0.9–2.1 mm posterior to
regma) through the anterior (PVHap) and medial (PVHmp) por-ions of the PVH were cut on a cryostat at −20 ◦C, and dividednto six series of eight sections each. The span between two sec-ions was of 140 �m. We located the PVHap in order to analyzehe AHA, PVa and SO coincident with PVHap and the PVHmp inrder to analyze the PVa, SO and APF coincident with PVHmp. Thelices were thawed and mounted on slides covered with gelatin.rain sections rinsed in PBS were placed into 0.1 M glycine inBS (20 min) to remove excess aldehydes. After washes in PBS,ections were blocked (1 h) in PBS containing 0.1% Triton X-100nd 1% bovine serum albumin (BSA) and incubated with primaryntibodies.
.3.2. ImmunofluorescenceGABA (GAD67) and GR – the synthesis of GABA from glutamate
n the brain depends on two isoforms of the enzyme gluta-ate decarboxylase (GAD). GAD65 is most important to synthesis
he vesicular pool. GAD67 chosen as a marker of GABA neuronss quantitatively more important for GABA synthesis. It partici-ates in the cytoplasmic and vesicular pools [27,28]. The sectionsere incubated overnight (20 h) at 28 ◦C with mouse monoclonal
ce Letters 534 (2013) 199– 204
anti-GAD67 antibody (1:5000, MAB5406, Chemicon, UK) and goatanti-glucocorticoid receptor antibody (1:100, sc-1004, Santa CruzBiotechnology, USA) diluted in PBS containing 0.1% Triton X-100and 1% BSA. Following PBS washes, they were incubated (1 h) ina cocktail of fluorescent secondary antibodies: donkey anti-mouseIgG labeled with Alexa Fluor 488 (Molecular Probes, USA) for GABAand donkey anti-rabbit IgG labeled with Alexa Fluor 594 (Molec-ular Probes, USA) for GR. The slides were washed with PBS andcoverslipped. The omission of the primary antibody resulted in nolabeling. For each double-label combination, primary antibodieswere titrated to determine the maximum dilutions that providedrobust signals and a minimum of nonspecific staining (images notshown).
2.3.3. Tissue analysisTissue sections were examined using an Axioskope 2 Plus
microscope (Carl Zeiss, Germany) with a 20× objective and aZeiss AxioCam digital camera (Carl Zeiss, Germany) to capture theimages. One series of tissue sections through the rostro–caudalextent of the PVH was used per animal (n = 5–9 per group). Theseries of sections from each brain were compared according ofanatomical atlas [22] to define the boundaries of AHA, PVa, SO,APF and different PVH subdivisions. We used the same size ofthe optical field for all of the photomicrographs. All GABA neu-rons and GR-immunoreactive (-ir) neurons within the section werequantified bilaterally. Next, sections scored for the co-staining ofGABA and GR were re-examined using confocal microscopy (TCSSP2 AOBS, Leica Microsystems, Germany). The quantitative assess-ment of GABA neurons, GR-ir neurons, and double-labeled cells wasobtained from captured microscope images using Image J software(NIH, Bethesda, MD, USA).
2.4. Hormone measurement
Plasma corticosterone was measured by a radioimmunoas-say that required extraction with ethanol and used the followingreagents: corticosterone H3 (NEN Life Sciences Products, USA); spe-cific antibody and standard reference (Sigma, USA). All samplesfrom the same experiment were measured in the same assay. Theintra-assay error was 5% and the minimum detectable dose was0.8 ng/mL.
2.5. Statistical analysis
The mean ± SEM of GABA neurons, GR-ir neurons and percentof dual-labeled cells was calculated per region in groups of sixanimals. Two-way ANOVA followed by the Bonferroni post hocanalyses for multiple comparisons was used to evaluate the num-ber of GR-ir neurons. The number of GABA neurons and the percentof dual-labeled cells were analyzed by Student’s t-test. P < 0.05 wasconsidered statistically significant.
3. Results
3.1. Effects of adrenalectomy and dexamethasone on plasmacorticosterone
Plasma corticosterone was higher in intact-saline(62.2 ± 3.5 ng/mL) and sham-saline (68.9 ± 2.9 ng/mL) groups thanintact-Dexa (3.3 ± 0.5 ng/mL) and sham-Dexa (4.8 ± 0.9 ng/mL)groups.
Plasma corticosterone was lower in ADX-saline group
(2.6 ± 2.5 ng/mL) than intact-saline group (62.2 ± 8.7 ng/mL), but itwas not different of ADX-dexamethasone group (6.7 ± 3.0 ng/mL).Plasma corticosterone decreases significantly after treatment andsurgery. The interaction between treatment and surgery reached
L.M. de Souza, C.R. Franci / Neuroscience Letters 534 (2013) 199– 204 201
Fig. 1. Effects of adrenalectomy (ADX) and dexamethasone (DEXA) treatment on the number glucocorticoid receptors-immunoreactive (GR-ir) neurons. Anterior hypothal-amic area (AHA), (A panel); peri-supraoptic area (SO), (B panel); anterior periventricular nucleus (PVa), (C panel); A, B and C coincident with PVHap. Anterior periventricularn S.E.Mg
stP
3
(Pa(PSt(daPv(dP(P1w6w(wirg
ucleus (PVa), (D panel) coincident with PVHmp. Each bar represents the mean ±roups, P < 0.001.
ignificance (F2.33 = 232.6; P < 0.0001). Significant main effects forreatment (F1.33 = 609.1; P < 0.0001) and surgery (F2.33 = 191.4;
< 0.0001) were detected.
.2. GR immunoreactivity
Fig. 1 presents the number of GR-ir neurons in the AHAA), SO (B), PVa coincident with PVHap (C) and coincident withVHmp (D). A significant interaction occurred between treatmentnd surgery factors on AHA (A) (F2.35 = 30.7; P < 0.0001), SO (B)F2.23 = 24.4; P < 0.0001), PVa coincident with PVHap (C) (F2.30 = 4.4;
= 0.0203) and coincident with PVHmp (D) (F2.25 = 11.6; P = 0.0003).ignificant effects were detected for treatment and surgery inhe AHA (A) (F1.35 = 47.6; P < 0.0001)/(F2.35 = 13.8; P < 0.0001), SOB) (F1.23 = 55.1; P < 0.0001)/(F2.23 = 13.1; P < 0.0002), PVa coinci-ent with PVHap (C) (F1.30 = 15.8; P = 0.0004)/(F2.30 = 2.5; P = 0.0976)nd coincident with PVHmp (D) (F1.25 = 8.6; P = 0.0070)/(F2.25 = 3.6;
= 0.0417). For the treatment factor, the number of acti-ated GR-ir neurons was higher in ADX-Dexa group (AHAA), 63.7 ± 7.7/section; SO (B), 30.1 ± 4.5/section; PVa coinci-ent with PVHap (C), 37.7 ± 5.3/section and coincident withVHmp (D), 51.0 ± 11.6/section) than ADX-saline group (AHAA), 0.8 ± 0.8/section; SO (B), no detected; PVa coincident withVHap (C), 8.7 ± 0.4/section and coincident with PVHmp (D),.5 ± 0.9/section). For the surgery factor, the number of neuronsith activated GR-ir was higher in ADX-Dexa group (AHA (A),
3.7 ± 7.7/section; SO (B), 30.1 ± 4.5/section; and PVa coincidentith PVHmp (D), 51.0 ± 11.6/section) than intact-Dexa group (AHA
A), 14.5 ± 1.0/section; SO (B), 6.2 ± 1.4/section; and PVa coincident
ith PVHmp (D), 6.0 ± 3.1/section). We did not find GR-ir neuronsn APF and SO coincident with PVHmp. The number of GR-ir neu-ons was increased by treatment with dexamethasone for all ADXroups.
. The symbols for treatment (�) and for surgery (�) show the difference between
3.3. GAD67 (GABA) immunoreactivity and neuronalco-localization of GAD67 (GABA) and GR
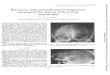
Fig. 2 shows illustrative photomicrographs from PVa coincidentwith PVHmp that express immunofluorescent labels for GAD67(GABA neurons, green) and GR (red). In both single- and double-label experiments, the omission of the primary antibody resultedin no labeling (images no shown).
Fig. 3(A and B) presents the number of GAD67-positive neuronsin the PVa coincident with PVHap (A) and coincident with PVHmp(B). We found GAD67-ir just in the PVa from intact-Dexa and sham-Dexa groups. The GAD67-ir in the PVa coincident with PVHmp (B)was higher for the sham-Dexa group (2.8 ± 0.42/section) than theintact-Dexa group (0.42 ± 0.23/section).
Fig. 3(C and D) shows the percent of GAD67-ir neurons with acti-vated GR that were detected in the PVa coincident with PVHap (C)and coincident with PVHmp (D). A Student’s t-test indicated a sig-nificant difference just in the PVa coincident with PVHmp (D), withan increased percentage of the GAD67-ir neurons with activatedGR in the sham-Dexa group (21.8 ± 12.1% of neurons/section) com-pared with the intact-Dexa group (7.7 ± 0.7% of neurons/section).
4. Discussion
Our results showed a significant decrease of plasma corticoste-rone, after adrenalectomy or treatment with dexamethasone, inagreement with classical knowledge regarding control of the HPAa[28]. This datum is a control for our studies of correlation betweenthe activity of HPAa and the participation of GABA neurons and GR.
We used dexamethasone for replacement, a synthetic substancethat has been widely used for in vitro and in vivo studies of the glu-cocorticoid effects on different cellular and physiological responses.The dexamethasone is a substrate for mdr-1a-P-glycoprotein and
202 L.M. de Souza, C.R. Franci / Neuroscience Letters 534 (2013) 199– 204
Fig. 2. Photomicrographs (20 �m thick section) of the anterior periventricular nucleus (PVa) coincident with medial parvocellular subdivision of the paraventricular nucleus(PVHmp) from sham animals treated with dexamethasone (DEXA). Immunofluorescence labels for GABA neurons (GAD67 positive neurons, green, Alexa Fluor 488; A panel)and glucocorticoid receptors (GR, red, Alexa Fluor 594; B, panel). The double labeling of GAD67 positive neurons with activated GR were observed, as shown in the compositeimage (C panel). D panel shows PVa region coincident with the anterior parvocellular subdivision of the paraventricular nucleus (PVHap) and PVHmp in schematic figureadapted from anatomical atlas (Swanson, 2004). Confocal photomicrographs (20 �m thick sections) of immunofluorescence for GAD67 positive neurons (E panel) and GR (Fpanel) in the PVa coincident with PVHmp. The composite image of GAD67-GR double-labeling in G panel and high magnification of image from G panel in H panel confirm then –C pam anel)
tsfb1a
wwcostctctaWltstsir
P
uclear GR immunoreactivity in the GAD67 positive neurons. Scale bars = 50 �m (Aagnocelular part medial zone; arrows indicate the GAD67-GR double-labeling (C p
his conjugation impedes its action. At 2.5%, the dexamethasone isufficient to decrease the magnitude of stress response. Responsesor restraint stress and CRH mRNA expression were clearly reducedy dexamethasone. This indicates that the barrier formed by mdr-a-P-glycoprotein is of course relative. It can be overcome by highmounts of dexamethasone allowing activation of GR [4,10,20,24].
We found a significant increase of GR-ir in the PVa coincidentith PVHap or PVHmp from adrenalectomized animals treatedith dexamethasone. This finding suggests that the endogenous
orticosterone (in intact and sham animals) desensitizes responsesf the GR to the dexamethasone treatment. According to previoustudies, corticosterone treatment induces GR re-accumulation inhe nuclei of parvocellular neurons [9,11]. It has been shown that ahaperone complex in the cytoplasm binds to the GR and maintainshese receptors in their inactive form. After dissociation from thehaperone complex, GR translocates to the nucleus to regulate thearget gene expression [3]. Nuclear GR-ir decreased one week afterdrenalectomy and increased after corticosteroid replacement [18].e used antiserum against the N-terminus of the GR molecule that
abels GR as being dissociated from the chaperone complex, i.e., inhe process of translocation or once translocated to the nucleus, ashown by the results of this study. We also verified the accumula-ion of nuclear GR-ir after corticosteroid replacement. Our resultshowed that the process of GR re-accumulation also occurred
n the PVa, indicating the effect of the glucocorticoids on thisegion.The GABAergic control on the parvocellular portion of theVH was shown to be important for the control of HPAa activity
nels), 8 �m (E–G panels). 3 V, third ventricle; fx, column of fornix; pmm, posterior.
[1,14–16]. However, we found GABAergic neurons in only the PVacoincident with as both parvocellular areas studied (PVHap andPVHmp), suggesting that the PVa could be particularly impor-tant for the GABAergic control of the parvocellular portion of thePVH. The analysis of GR co-localization in GAD67-positive neu-rons revealed double-labeling in only the PVa that is coincidentwith PVHap or PVHmp from intact and sham animals treated withdexamethasone. This datum suggests an effect of dexamethasoneon the local GABAergic circuit. Previous studies have showed thatacute elevation of plasma corticosterone after a stressful stimu-lus in intact animals induces an increase of GAD67 expression inregions identified as main sources of GABAergic afferents to theparvocellular region of the PVH [2]. GAD67 is the main enzymeinvolved in GABA synthesis during stressful situations [29]. More-over, the increase of the GAD67-ir related to GR co-localization inthe PVa coincident with PVHmp from sham animals treated withdexamethasone supports the notion of surgical stress having aneffect on the local GABAergic circuit.
The hippocampus and the prefrontal cortex have been impli-cated in inhibitory mechanisms of stress responses because lesionsof these areas selectively enhance the response of the PVN and HPAaxis to the psychogenic stressors. In general, the parvocellular PVNreceives little or no direct innervation from these regions. However,the ventral subiculum and infralimbic/prelimbic cortices, regions
of the hippocampus and prefrontal cortex, respectively, send gluta-mate and GABA inputs to the subparaventricular zone and peri-PVNregion (including PVa). The majority of ventral subicular inputs areglutamatergic. This pathway would activate PVN-projecting GABA
L.M. de Souza, C.R. Franci / Neuroscience Letters 534 (2013) 199– 204 203
Fig. 3. The number of GAD67 positive neurons (GABA neurons) (A and B panels) and the percentage of GAD67 positive neurons with expression of GR (C and D panels). All barsa ellulap of intam
npmGwmsTcmrP
wanidgccobirgnGeoO
re from the anterior periventricular nucleus (PVa) coincident with anterior parvocarvocellular subdivision of the paraventricular nucleus (PVHmp) (B and D panels)
ean ± S.E.M. The symbol � shows the difference between the groups, P < 0.05.
eurons to inhibit the HPA axis (5, 12, 14, 15, 21). The hippocam-us and prefrontal cortex, structures associated with learning andemory, are poised to inhibit the HPA axis through glutamate-ABA connections. This pathway may afford a means throughhich previous experience and spatial information can limit theagnitude and duration of stress responses and thus keep expo-
ure to the catabolic actions of glucocorticoids at a minimum [14].hus, surgical stress could induce an effect on the local GABAergicircuit through glutamate-GABA connections.This fact is in agree-ent with our results of an increase of the GAD67 immunoreactivity
elated to GR co-localization in the PVa that is coincident withVHmp from sham animals treated with dexamethasone.
We did not find GAD67-ir in adrenalectomized animals treatedith dexamethasone. However, previous electrophysiological
nalysis has shown a large increase of GABAergic activity and theumber of GABA-positive terminals onto corticotrophin releas-
ng hormone (CRH) neurons after adrenalectomy [25,26]. Theseata suggest that in the absence of hormone, the local GABAer-ic circuit provides a compensatory mechanism for maintainingontrollable network activity [25]. It has been also shown that glu-ocorticoid replacement after adrenalectomy reverses this increasef GABAergic activity, indicating that the GABAergic activity coulde modulated by GR [26]. Therefore, the absence of the GAD67-ir
n adrenalectomized animals treated with dexamethasone cor-oborates previous findings that indicate an inhibitory action oflucocorticoids on GABAergic activity [26]. Our results provideew information on this inhibitory action of glucocorticoids on
ABAergic activity: the glucocorticoid effect on GAD67 expression,specially in situations when the literature [25] reports an increasef the inhibitory compensatory mechanism by adrenalectomy.ur results on the effect of glucocorticoids on GAD67 expressionr subdivision of paraventricular nucleus (PVHap) (A and B panels) and with medialct and sham animals treated with dexamethasone (Dexa). Each bar represents the
contribute by providing mechanisms potentially important forinhibitory compensatory systems in modulating the HPA axis activ-ity.
5. Conclusions
In conclusion, our results indicate a possible direct effect ofthe glucocorticoid on GABA neurons because the double-labelingof GAD67-positive neurons and GR. This double-labeling occurredonly in the PVa coincident with PVHap or PVHmp that seems bethe principal hypothalamic source of GABAergic afferents to theanterior and medial parvocellular region of the PVH. Thus, theglucocorticoid effect on GAD67 expression points for an indirectmechanism through PVa coincident with PVHap or PVHmp to con-trol of the activity of neurons in the PVH parvocellular region andthe HPAa. As such, this effect indicates important lead for futureand novel investigation strategies concerning the influence andmechanisms of GABAergic inputs on HPAa.
Acknowledgements
The authors are grateful to the brazilian agencies FAPESP, CAPESand CNPq for financial support and to Rogério Rosário Azevedo andRubens Fernando de Mello for technical assistance.
References
[1] V. Bartanusz, D. Muller, R.C. Gaillard, P. Streit, L. Vutskits, J.Z. Kiss, Localgamma-aminobutyric acid and glutamate circuit control of hypophyseotrophiccorticotropin-releasing factor neuron activity in the paraventricular nucleus ofthe hypothalamus, European Journal of Neuroscience 19 (2004) 777–782.

2 oscien
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
04 L.M. de Souza, C.R. Franci / Neur
[2] G. Bowers, W.E. Cullinan, J.P. Herman, Region-specific regulation of glutamicacid decarboxylase (GAD) mRNA expression in central stress circuits, Journalof Neuroscience 18 (1998) 5938–5947.
[3] A. Carrigan, R.F. Walther, H.A. Salem, D. Wu, E. Atlas, Y.A. Lefebvre, R.J. Hache,An active nuclear retention signal in the glucocorticoid receptor functions asa strong inducer of transcriptional activation, Journal of Biological Chemistry282 (2007) 10963–10971.
[4] M.A. Cole, P.J. Kim, B.A. Kalman, R.L. Spencer, Dexamethasone suppression ofcorticosteroid secretion: evaluation of the site of action by receptor measuresand functional studies, Psychoneuroendocrinology 25 (2000) 151–167.
[5] R.L. Cole, P.E. Sawchenko, Neurotransmitter regulation of cellular activationand neuropeptide gene expression in the paraventricular nucleus of the hypo-thalamus, Journal of Neuroscience 22 (2002) 959–969.
[6] M.F. Dallman, Stress update Adaptation of the hypothalamic–pituitary–adrenalaxis to chronic stress, Trends in Endocrinology and Metabolism 4 (1993) 62–69.
[7] M.F. Dallman, S.F. Akana, C.S. Cascio, D.N. Darlington, L. Jacobson, N. Levin,Regulation of ACTH secretion: variations on a theme of B, Recent Progress inHormone Research 43 (1987) 113–173.
[8] E.R. De Kloet, E. Vreugdenhil, M.S. Oitzl, M. Joels, Brain corticosteroid receptorbalance in health and disease, Endocrine Reviews 19 (1998) 269–301.
[9] K. Fuxe, A.C. Wikstrom, S. Okret, L.F. Agnati, A. Harfstrand, Z.Y. Yu, L. Granholm,M. Zoli, W. Vale, J.A. Gustafsson, Mapping of glucocorticoid receptor immunore-active neurons in the rat tel- and diencephalon using a monoclonal antibodyagainst rat liver glucocorticoid receptor, Endocrinology 117 (1985) 1803–1812.
10] A.B. Ginsberg, S. Campeau, H.E. Day, R.L. Spencer, Acute glucocorticoid pre-treatment suppresses stress-induced hypothalamic–pituitary–adrenal axishormone secretion and expression of corticotropin-releasing hormone hnRNAbut does not affect c-fosmRNA or fos protein expression in the paraventric-ular nucleus of the hypothalamus, Journal of Neuroendocrinology 15 (2003)1075–1083.
11] F. Han, H. Ozawa, K. Matsuda, M. Nishi, M. Kawata, Colocalization of min-eralocorticoid receptor and glucocorticoid receptor in the hippocampus andhypothalamus, Neuroscience Research 51 (2005) 371–381.
12] J.P. Herman, W.E. Cullinan, Neurocircuitry of stress: central control ofthe hypothalamo–pituitary–adrenocortical axis, Trends in Neurosciences 20(1997) 78–84.
13] J.P. Herman, W.E. Cullinan, D.R. Ziegler, J.G. Tasker, Role of the paraventricularnucleus microenvironment in stress integration, European Journal of Neuro-science 16 (2002) 381–385.
14] J.P. Herman, N.K. Mueller, H. Figueiredo, Role of GABA and glutamate circuitry
in hypothalamo–pituitary–adrenocortical stress integration, Annals of the NewYork Academy of Sciences 1018 (2004) 35–45.15] J.P. Herman, J.G. Tasker, D.R. Ziegler, W.E. Cullinan, Local circuit regulationof paraventricular nucleus stress integration: glutamate-GABA connections,Pharmacology Biochemistry and Behavior 71 (2002) 457–468.
[
ce Letters 534 (2013) 199– 204
16] K.J. Kovacs, I.H. Miklos, B. Bali, GABAergic mechanisms constraining the activ-ity of the hypothalamo-pituitary-adrenocortical axis, Annals of the New YorkAcademy of Sciences 1018 (2004) 466–476.
17] S.L. Lightman, The neuroendocrinology of stress: a never ending story, Journalof Neuroendocrinology 20 (2008) 880–884.
18] X.M. Ma, A. Levy, S.L. Lightman, Emergence of an isolated arginine vasopressin(AVP) response to stress after repeated restraint: a study of both AVP andcorticotropin-releasing hormone messenger ribonucleic acid (RNA) and het-eronuclear RNA, Endocrinology 138 (1997) 4351–4357.
19] de S.L. Marques, C.R. Franci, GABAergic mediation of stress-induced secretionof corticosterone and oxytocin but not prolactin, by the hypothalamic paraven-tricular nucleus, Life Sciences 83 (2008) 686–692.
20] A.H. Miller, R.L. Spencer, M. Pulera, S. Kang, B.S. McEwen, M. Stein, Adrenalsteroid receptor activation in rat brain and pituitary following dexamethasone:implications for the dexamethasone suppression test, Biological Psychiatry 32(1992) 850–869.
21] B.L. Roland, P.E. Sawchenko, Local origins of some GABAergic projections to theparaventricular and supraoptic nuclei of the hypothalamus in the rat, Journalof Comparative Neurology 332 (1993) 123–143.
22] L.W. Swanson, Brain maps: structure of the rat brain. A laboratory guide withprinted and electronic templates for data, models and schematics, 3rd ed.,Elsevier, Amsterdam, 2004.
23] P.E. Sawchenko, E.R. Brown, R.K. Chan, A. Ericsson, H.Y. Li, B.L. Roland, K.J.Kovacs, The paraventricular nucleus of the hypothalamus and the functionalneuroanatomy of visceromotor responses to stress, Progress in Brain Research107 (1996) 201–222.
24] A.H. Schinkel, E. Wagenaar, L. van Deemter, C.A.A.M. Mol, P. Borst, Absence ofthe mdr1a P-Glycoprotein in mice affects tissue distribution and pharmacoki-netics of dexamethasone digoxin, and cyclosporin, Archive of the Journal ofClinical Investigation 96 (1995) 1698–1705.
25] J.M. Verkuyl, M. Joels, Effect of adrenalectomy on miniature inhibitory post-synaptic currents in the paraventricular nucleus of the hypothalamus, Journalof Neurophysiology 89 (2003) 237–245.
26] J.M. Verkuyl, H. Karst, M. Joels, GABAergic transmission in the rat paraventric-ular nucleus of the hypothalamus is suppressed by corticosterone and stress,European Journal of Neuroscience 21 (2005) 113–121.
27] H.S. Waagepetersen, U. Sonnewald, A. Schousboe, Compartmentation of glu-tamine glutamate, and GABA metabolism in neurons and astrocytes: functionalimplications, Neuroscientist 9 (2003) 398–403.
28] M.H. Whitnall, Regulation of the hypothalamic corticotropin-releasing hor-
mone neurosecretory system, Progress in Neurobiology 40 (1993) 573–629.29] H. Wu, Y. Jin, C. Buddhala, G. Osterhaus, E. Cohen, H. Jin, J. Wei, K. Davis, K. Obata,J.Y. Wu, Role of glutamate decarboxylase (GAD) isoform GAD65, in GABA syn-thesis and transport into synaptic vesicles – Evidence from GAD65-knockoutmice studies, Brain Research 1154 (2007) 80–83.