Embed Size (px)
Citation preview

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BlOPHYSiCAL RESEARCH COMMUNICATIONS October 15, 1982 Pages 1048-1055
DIFFERENTIAL EFFECT OF RECOMBINANT HUMAN LEUKOCYTE INTERFERON ON HUMAN LEUKEMIC AND NORMAL MYELOID PROGENITOR CELLS4
Steven Grant1$3*5, Kapil Bhallaly3,
I. Bernard Weinsteinly3, Sidney
Pestka4 and Paul B. Fisher2y3
Departments of Medicine1 and Microbiology* and Cancer Center/Institute of Cancer Research,3
Columbia University, College of Physicians and Surgeons, New York, New York 10032 and Roche
Institute of Molecular Biology4, Nutley, New Jersey 07110
Received August 31, 1982
SUMMARY: Two separate clones of recombinant leukocyte interferon (IFLrA and IFLrD) inhibited the cloning efficiency in soft agar of the human leukemia cell lines HL-60 and KG-l. Inhibition of the growth in agar of normal human bone marrow myeloid progenitors was also observed, but this required consider- ably higher concentrations. IFLrA and IFLrD also inhibited the growth of HL-60 and KG-l cells in suspension culture. This antiproliferative effect did not appear to be due to induction of maturation of these cells. Our results sug- gest that homogeneous preparations of interferon may be capable of exerting selective antiproliferative effects on malignant human myeloid progenitor cells in comparison to their normal counterparts.
INTRODUCTION
Interferons (IFS) are a family of proteins with antiviral activity produced
by animals and cells in culture in response to viruses, as well as certain non-
viral substances (l-4). In addition to their antiviral effects, they exhibit a
wide variety of other effects, including inhibition of the growth of both normal
and transformed cells (5-6). Clinical studies utilizing partially-purified human
leukocyte interferon have been associated with the development of neutropenia, sug-
gesting that these preparations of interferon were toxic to normal myeloid progenitor
cells (7). These findings are consistent with in vitro studies which demonstrated
5. Fellow of the Leukemia Society of America to whom reprint requests should be made.
Abbreviations used: TPA, 12-O-tetradecanoyl-phorbol-13 acetate; DMSO,
dimethyl sulfoxide
0006-291X/82/191048-08$01.00/0 Copyright 0 I982 by Academic Press, Inc. All rights of reproducfion in any form reserved. 1048

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
that certain interferon preparations inhibited the formation of normal human
granulocyte macrophage colonies (CFU-GM) in soft agar (8,9). To date, normal
and malignant cells have exhibited varying degrees of sensitivity to the in vitro
growth inhibitory effects of the various interferons (10).
After the isolation (11) and expression (12) of DNA recombinants contain-
ing sequences of human leukocyte interferon, homogeneous preparations of human
leukocyte interferon were prepared (13). We have studied the inhibitory effects
of two types of human leukocyte interferon produced in Escherichia coli (IFLrA
and IFLrD) on the cloning efficiency of normal human bone marrow progenitor
cells in soft agar. In addition, we have examined the effects of these inter-
ferons on the in vitro proliferative capacity of two human leukemic cell lines,
HL-60 and KG-l. These cell lines have been the focus of considerable interest
in view of their capacity to undergo terminal differentiation in the presence
of a variety of compounds, including dimethyl sulfoxide (DMSO) or phorbol esters
(14,15). The purpose of the present study was to determine whether these inter-
ferons might exert a preferential inhibitory effect on the proliferation of leu-
kemic versus normal human myeloid progenitor cells in soft agar. A second aim
of this study was to determine whether the growth inhibitory effects of IFLrA or
IFLrD on HL-60 and KG-1 cellswereassociated with morphologic or functional
evidence of differentiation.
METHODS
HL-60 and KG-1 cells are derived from the original lines initially described (14,15). They were maintained in RPM1 medium supplemented with 1% sodium pyru- vate and nonessential amino acids and 10% heat-inactivated fetal calf serum (GIBCO). Cells were subcultured twice weekly in 75 cm2 sterile plastic tissue culture flasks (Corning) and kept in a fully-humidified 37"C, 5% CO2 water- jacketed incubator (Napco). Recombinant interferons (IFLrA and IFLrD) were pre- pared as described (13). The interferon preparations were stored in sterile 2-ml vials at -8O"c, thawed immediately prior to use and diluted to the appropriate concentration in RPM1 medium. TPA (12-O-tetradecanoyl-phorbol-13-acetate) and DMSO were purchased from Sigma Chemicals, St. Louis, MO. In suspension culture growth experiments, 5 ml of HL-60 and KG-1 were seeded into 35mmsix-wellplates (Costar) at an initial cell density of lo5 cells/ml. TPA (lo-7M), DMSO (1.25%) or recombinant interferon (IFLrA or IFLrD, 200-5.000 I.U./ml) were added to the wells, and the plates were placed in the 37OC, 5% CO2, fully-humidified incubator. Aliquots of 0.5 ml were withdrawn at 24-hr intervals, and cell density determinations obtained with a model ZBI Coulter counter (Hialeah, Fla.). At the end of 5 d, the cells were pelleted and cytocentrifuge prepara- tions made with a Shandon cytocentrifuge. After air drying, the slides were stained with Wright-Giemsa, and a differential count performed by light microscopic examination of at least 200 cells. For nitro-blue tetrazolium
1049

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
dye reduction studies, 1 ml of cells suspended at 2 x 106 cells/ml in RPM1 medium supplemented with 20% FCS was incubated for 20 min at 37OC with equal volume of 0.2% NBT (Sigma) in the presence of 200 ng of TPA. The percent of cells containing intracellular reduced blue-black formazan deposits was then determined on Wright-Giemsa stains of cytospin preparations of incubated cells.
The ability of HL-60 and KG-l cells to form colonies in soft agar in the presence of varying concentrations of interferon was determined by a slight modification of a previously described technique (16). Cells were plated in 18 mm 12-well dishes (Costar) utilizing a two-layer agar system. The bottom layer consisted of 0.5 ml of RPM1 medium supplemented with 1% sodium pyruvate, 1% nonessential amino acids, 20% (V/V) fetal calf serum and 0.5% bacto agar (Difco). The top layer containing the cells consisted of 0.5 ml of RPM1 medium, 20% (V/V) fetal calf serum, 0.3% bacto agar, and the appropriate concentration of ILFrA or IFLrD. HL-60 cells (2 x 103) or KG-l cells (5 x 103) were plated for each condition. After the agar hardened, 0.1 ml of GCT-conditioned medium (GIBCO) was added to each plate as a source of colony-stimulating activity (17). The plates were then placed in a 37OC. 5% C02, fully-humidified incubator for 10 d. At the end of this period, the plates were removed, and the number of colonies, consisting of groups of 50 or more cells, scored with the aid of an Olympus Model CK inverted microscope. Inhibition of colony growth was expressed as (l-No/NC) x 100, where No equals control colony growth and NC equals the number of colonies formed in the presence of a particular concentration of interferon. The I.C.50 was defined as the interferon concentration which resulted in a 50% reduction in-colony growth when compared to controls.
The effect of IFLrA and IFLrD on the growth in soft agar of normal human bone marrow progenitor cells (CFU-GM) was determined by a minor modification of a previously described method (18). Bone marrow cells were obtained with informed consent from patients undergoing routine diagnostic aspirations who did not have a hematologic malignancy. These studies have been sanctioned by the Investiga- tional Review Board of the Columbia University College of Physicians and Surgeons. The cells were passed through a 26-gauge needle to disperse clumps, and diluted 1:4 with McCoy's 5a medium (GIBCO) supplemented with sodium pyruvate, essential and nonessential amino acids, MEM vitamins, asparagine, glutamine and serine. The suspension was then gently layered over 5 ml of lymphocyte separation medium (sp. grav. =1.077-1.081; Bionetics, Kensington, Md) in sterile 20-ml centrifuge tubes and centrifuged at 400 xg for 38 min. The interface layer, containing the mononuclear cells, was extracted with a sterile Pasteur pipette, washed three times with supplemented McCoy's 5a medium and cell counts obtained with a hematocytometer. The method for determining the effect of IFLrA or IFLrD on the growth of bone marrow cells in soft agar was the same as that described for HL-60 and KG-l cells, except the supplemented McCoy's 5a medium was used in place of RPM1 and 105 bone marrow cells were plated in each well, After 10 d of incubation, the number of colonies, consisting of groups of 50 or more granulocytic or macrophage-like cells, was scored with the aid of an Olympus inverted microscope. Inhibition of normal bone marrow myeloid progenitor cell growth in soft agar by IFLrA or IFLrD was expressed in the same manner as for HL-60 and KG-l cells.
RESULTS
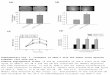
Figure 1 illustrates the effects of IFLrA and IFLrD on inhibition of
colony formation by KG-l, HL-60 and normal human bone marrow progenitors in
soft agar. The inhibitory effects of both types of interferon was greater for
HL-60 and KG-1 cells than for normal marrow cells over the entire range of inter-
feron concentrations tested (200-5,000 I.U./ml). At high IFLrA concentrations
1050

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
20-i
I,, , , , ( 1000 3000 5000
IFLr DOSE (units / ml1
Figure 1: -- The effect of IFLrA and IFLrD on the cloning efficiency of HL-60, KG-l and normal bone marrow cells.
The inhibitory effect of IFLrA (A) and IFLrD (B) on the cloning efficiency of HL-60 (a), KG-l (A) and normal human bone marrow porgenitor cells (O-O) was assessed as described in Material and Methods. 5,000 I.U./ml is equivalent to 2.8 and 165.0 x lo4 pg protein/ml for IFLrA and IFLrD, respectively.
(5,000 I-U./ml), inhibition of KG-l and HL-60 cell colony formation was about 98
and 94%, respectively, but only 62% for normal marrow cells. With all three cell
types, the inhibitory effects of IFLrD were less than those obtained with equal
concentrations of IFLrA. An IFLrD concentration of 5,000 I.U./ml was associated
with about 85 and 83% reduction in HL-60 and KG-l cell colony formation, but
only a 48% reduction in colony formation by normal human marrow cells. With
IFLrA, the I.C.50 (i.e., the concentration which produced a 50% reduction in
colony formation) was 400 and 300 I.U./ml for HL-60 and KG-l cells, respectively,
and averaged 1,250 I.U./ml for the seven nonleukemic bone marrow samples. With
IFLrD, the I.C.50 was 650 and 950 I.U./ml for HL-60 and KG-l cells, respectively;
the average inhibitory effect of an IFLrD concentration of 5,000 I.U./ml in four
bone marrow samples was slightly less than 50%. In the seven normal bone marrow
samples exposed to IFLrA, the I.C.50 ranged from 850 to 1,900 I.U./ml (data not
shown). In only one of the four bone marrowsamples exposed to IFLrDdid aninterferon
concentration of 5,000 I.U./ml result in greater than a 50% reduction in colony
formation (53%).
1051

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
The inhibitory effects of IFLrA and IFLrD on the growth of HL-60 and KG-l
cells in suspension cultures are given in Table 1. A 4-d exposure to 5,000
I.U./ml of IFLrA or IFLrD resulted in about 49 and 35% inhibition of cell
growth, respectively, for both cell types. Interferon concentrations of
200 I.U./ml or less had negligible effects on cellular proliferations (not
shown). Thus, as for colony growth in soft agar, IFLrA was more inhibitory
to cell growth than IFLrD at each interferon dose tested (~(0.05). The
growth inhibitory effects of IFLrA of IFLrD were not associated with evidence
of morphologic or functional induction of differentiation in either HL-60 or
KG-~ cells, even at the highest concentration tested, i.e., 5,000 I.U./ml. In
contrast, the majority of HL-60 and KG-1 cells exposed to 1.25% DMSO or lo-TM
TPA matured past the promyelocyte stage or became capable of reducing NBT dye
(data not shown here).
DISCUSSION
These results demonstrate that two different homogeneous preparations of
interferon produced in bacteria by recombinant DNA techniques are inhibitory to
the in vitro growth of both leukemic and normal myeloid progenitor cells. It is
of particular interest that we found that both of these highly-purified inter-
ferons preferentially inhibited the leukemic cell cultures, since in previous
studies with partially-purified myeloid, lymphoblastoid or fibroblast interferons,
there appeared to be equal inhibition of the growth in soft agar of leukemic and
normal myeloid cells (19). It is conceivable that impurities present in previous-
ly available interferon preparations may be responsible for the lack of a selective
effect on leukemic cells, although this hypothesis requires further testing.
Little is known about the mechanisms of regulation of cell growth and dif-
ferentiation of interferons. Studies by Verma et al have suggested that par-
tially-purified human leukocyte interferons are capable of blocking the differen-
tiation of granulocytic progenitor cells (8,9). In a recent study, highly-puri-
fied human leukocyte interferon did not induce differentiation in HL-60 cells (20),
nor did we see such an effect with IFLrA or IFLrD. On the other hand, syner-
gistic effects on differentiation were noted when interferon was combined with
1052

Table
1:
Ef
fect
of
in
terfe
rons
on
th
e gr
owth
an
d di
ffere
ntia
tion
of
susp
ensio
n cu
lture
s of
HL
-60
and
KG-l
cells
.
COND
ITIO
NS
CONT
ROL
5000
IF
LrA
(I.U
./ml)
2000
10
00
500
5000
IF
T.rD
(I.
lJ./m
l>
2000
10
00
500
1.
Gro
wth
(X
inhi
bitio
n at
96
hr
) a.
HL
-60
0 49
.4k4
.2
44.3
?3.8
35
.723
.1
17.3
t2.4
35
.7k3.2
31
.924
.2
27.O
t2.8
8.
321.
6
b.
KG-l
0 47
.Ok3
.8 37
.6~4
.1
33.2
t2.9
15
.651
.8
35.6
23.1
24
.853
.1
17.9
22.9
6.
3t1.
4
2.
Mor
pholo
gic
mat
urat
ion
afte
r 96
hr
(X
m
yelo
cyte
s,
met
amye
locy
tes,
ba
nds,
se
gmen
ts)
a.
HL.-6
0 5
4 1
3 N
.D.
5 2
4 N
.D.
b.
KG-1
3
2 3
1 N
.D.
3 2
4 N
.D.
3.
% N
BT
posi
tive
cells
(9
6 hr
) a.
HL
-60
3 2
2 1
N.D
. 3
2 2
N.D
.
b.
KG-l
3 3
1 2
N.D
. 3
2 2
M.D
.
Cells
we
re
seed
ed
at
5 x
lo4
cells
/ml
in tis
sue
cultu
re
flask
s co
ntai
ning
th
e de
signa
ted
conc
entra
tions
of
in
terfe
ron
and
cell
conc
entra
tions
de
term
ined
at
da
y 4.
G
rowt
h in
hibi
tion
in in
terfe
ron-
treat
ed
cells
wa
s ex
pres
sed
as
a pe
rcen
tage
re
lativ
e to
un
treat
ed
cont
rol
cells
. Va
lues
re
pres
ent
the
mea
ns
for
four
se
para
te
expe
rimen
ts
+ 1
S.D
. Th
e ce
ll de
nsity
in
cont
rol
cultu
res
at
the
end
of
96
hr
was
4.0
?r 0
.5
x lo5
ce
lls/m
l. At
th
e sa
me
time
cells
we
re
eval
uate
d fo
r ev
iden
ce
of
mor
phol
ogic
or
func
tiona
l ev
iden
ce
of
mat
urat
ion
(see
"M
ater
ials
and
Met
hods
").
N.D
. =
not
done
.

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
inducers such as TPA or retinoic acid (20). Thus, the antiproliferative effects
of interferon are not always simply due to induction of terminal differentiation,
although the interferons can under certain cases influence differentiation. It
has been found that a variety of chemotherapeutic agents, particularly those
that interfered with DNA synthesis, can under certain conditions induce HL-60
cells to develop into more mature forms (21). However, these same agents exhi-
bit only cytotoxic effects, without inducing defferentiation, when administered
at significantly higher concentrations. One might speculate, therefore, that
agents which inhibit the proliferation of malignant cells may render these cells
more sensitive to factors capable of enhancing differentiation. If this hypo-
thesis is correct, efforts to combine interferons with antimetabolites or speci-
fic inducers of leukemic cell differentiation might prove to be an effective
strategy in cancer therapy.
Recent clinical trials with impure preparations of human leukocyte inter-
feron in the treatment of hematologic malignancies have shown that myelosuppres-
sion is the dose-limiting factor with respect ot toxicity (22). It is possible
that recombinant interferon preparations might have less of a myelosuppressive
effect. Indeed, preliminary clinical trials with IFLrA suggest that this form
of interferon may cause less bone marrow toxicity than previously available prepa-
rations (23,24). These studies demonstrated that peak serum interferon levels as
high as 1,000 I.U./ml were achievable in some patients so treated, and that peak
levels of 500 I.U./ml were possible in the majority of patients (24). Thus,
further laboratory and clinical studies with homogeneous preparations of inter-
feron, alone or in combination with other agents, may lead to more effective therapy
for certain malignancies,
ACKNOWLEDGEMENTS: This work was supported in part by the National Cancer
Institute (Award CA-13696), the Pharmaceutical Manufacturers Association, the
William J. Matheson Foundation, the Cele Butwin Foundation and Hoffman-LaRoche.
References
1. Krim, M. (1980) Blood 55:5, 711. 2. Pestka, S. (1973) in DGensions in Health Research: Search for the
Medicines of Tommorrow. (Weissbach, H. and Kunz, R.M., eds., Academic Press, N.Y., 29.
1054

Vol. 108, No. 3, 1982 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
3. Finter, N.D., ed. (1973) Interferons and Interferon Inducers. American Elsevier Publishing.
4. Stewart, W.E.,II, ed. (1979) The Interferon System. Springer Verlag, co., New York, N.Y.
5. Gresser, I. (1977) Cell Immunol. 36, 406. 6. Paucker, I<., Cantell, K., and Henle, W. (1962) Virology 17, 324. 7. Gresser, I. (1977) Tex. Rep. Biol. Med. 21, 394. a. Verma, D.S., Spitzer, G., Gutterman, J.U., McCredie, K.B., Dicke, K.A.,
and Johnston, D.A. (1981) Exp. Hematol. 2, 63. 9. Verma, D.S., Spitzer, G., Gutter-man, J.U., McCredie, K.B., and Dicke, K.A.
(1979) Blood 54-, 1423. 10. Gresser, I., Bronty-Bote, D., Thomas, D., and Maciera-Coelho, A. (1970)
Proc. Natl. Acad. Sci. 66, 1052. 11. Maeda, S., McCandliss, R., Gross, M., Sloma, A., Familletti, R.C., Tabor,
J.M., Evinger, M., Levy, W.P., and Pestka, S. (1980) Proc. Natl. Acad. Sci. 77, 7010.
12. Goeddel, D.V., Yelverton, E., Ullrich, A., Heyneker, H.L., Miozzari, G., Holms, W., Seeburg, P.H., Drell, T., May, L., Stebbing, N., Crea, R., Maeda, S., McCandless, R., Sloma, A., Tabo, J.M., Gross, M., Familletti, P.C., and Pestka, S. (1980) Nature 287, 411.
13. Staehelin, T., Hobbs, D.S, Kung, H.F., Lai, C.Y., and Pestka, S. (1981) J. Biol. Chem. 256, 9750.
14. Collins, J.J., Gallo, R.C., and Gallegher, R.F. (1977) Naure 270, 347. 15. Koeffler, H.P., and Golde, D.W. (1978) Science 200, 1153. - 16. Koeffler, H.P.. and Golde, D.W. (1980) Blood 2, 344. 17. Dipersio, J.F., Brennan, J.K., and Lichtman, M.A. (1980) Blood 56, 716. 18. Broxmeyer, H.E., DeSousa, M., and Smithyman, A. (1980) Blood 15, 324. 19. Bradley, E.C., Ruscetti, F.W. (1981) Cancer Res. 41, 244. 20. Formida, N., Yamamoto, Y., and Hozumi, M. (1982) Eochem. Biophys. Res.
COTlUll. 104, 30. 21. Boder, A.J., Ting, R.C., and Gallo, R.C. (1981) J. Natl. Cancer Inst.
67, 1025. 22. Horning, S.J., Levine, J.F., Miller, R.A., Rosenberg, S.A., and Merigan,
T.C. (1982) J.A.M.A. 247, 1718. 23. Louis, A.C., Gallegher, J.G., Kikora, K., Levy, R., Rosenberg, S.A., and
Merigan, R.C. (1981) Blood 58, 4. 24. Gutterman, J.U., Fine, S., Quezda, J., Horning, S.J., Levine, J.F.,
Alexanian, R., Bernhardt, L., Kramer, M., Spiegel, H., Colburn, W., Trown, P ., Merigan, T., and Dziewanowska, 2. (1982) Ann. Intern. Med. 96, 549.
1055

















![Induction of Erythroid Differentiation in Human Leukemic K ......[CANCER RESEARCH 50, 1231-1236. February 15. 1990] Induction of Erythroid Differentiation in Human Leukemic K-562 Cells](https://img.pdfslide.us/doc/110x75/60b088961b1fcf1e2a746f9b/induction-of-erythroid-differentiation-in-human-leukemic-k-cancer-research.jpg)










