Embed Size (px)
Citation preview

0270-91 39/%/0602-0199$02.00/0 HEPATOLOGV Copyright 0 1986 by the American Association for the Study of Liver Diseases
Vol. 6, No. 2, pp. 199-204,1986 Printed in U. S. A.
Differential Distribution of Hepatitis B Core and E Antigens in Hepatocytes: Analysis by Monoclonal Antibodies
MARIO MONDELLI, RICHARD S. TEDDER, BRIDGET FERNS, PATRIZIA PONTISSO, GIUSEPPE REALDI AND ALFREDO ALBERTI
Istztuto di Clinica deUe Malattie Infettiue, Uniuersita di Pavia, I.R.C.C.S. Policlinico S. Matteo, 27100 Pavia, Italy; Section of Virology, Microbiology Department, The Middlesex Hospital Medical School and University College, London, England; and
Istituto di Medicinu Clinica, Cattedra di Clinica Medica II , Uniuersita di Padova, 35100 Padova, Ztaly
Current knowledge on the expression of HBeAg in hepatocytes is incomplete because of difficulties in ob- taining monospecific antisera devoid of anti-HBc reac- tivity. In this study, we have examined by immunofluo- rescence the expression of HBcAg and HBeAg in cryo- stat liver sections from 26 chronic carriers of EBsAg using monoclonal antibodies.
Although virtually all liver biopsies displayed con- cordance for FIBeAg and HBcAg expression, the pattern of fluoreecence differed markedly. Thus, monoclonal anti-HBc gave nuclear staining in all 13 reactive biop- sies, while cytoplasmic staining was obaerved in only two of theae. In contrast, monoclonal anti-HBe showed cytoplasmic reactivity coexisting with nuclear reactiv- ity in 10 of 13 reactive biopsies. Hepatitis B virus DNA polymerase activity in the wrum appeared to correlate better with the presence of HBcAg in hepatocytee rather than HBeAg.
These results provide further evidence that HBeAg is e x p r d both in the nuclei and in the cytoplasm of infected hepatocytes. The observation that the number of cells expressing HBeAg ex& those expming HBcAg in carriers with active virus replication would suggest that assembly of core particles occurs in only a proportion of infected hepatocytes expressing HBeAg.
The pattern of expression in hepatocytes of hepatitis B virus (HBV) antigens is considered to provide useful diagnostic and prognostic information. Two antigens have been well studied. The first is HBsAg. This is the major protein of the virus envelope and may be expressed in the cytoplasm of liver cells, a pattern found mainly in healthy HBsAg carriers and in patients with inactive liver disease (1). Alternatively, HBsAg has been reported to be expressed at the surface of hepatocytes, a pattern which is considered as a marker of active liver disease
Received March 18, 1985, accepted November 7, 1985. This study was supported in part by Consiglio Nazionale delle
Ricerche. Rome, Italy. This work was presented in part at the Autumn Meeting of the
British Society of Gastroenterology, 1983 and published in abstract form (Gut 1983; 36A995).
Dr. Mondelli is a recipient of Doctor of Research in Preventive and Community Medicine from the University of Pavia, Italy.
Address reprint requests to: Alfred0 Alberti, M.D., Istituto di Med- ieina Clinica, Cattedra di Clinica Medica 11, IJniversita di Padova, Via Giustiniani 2, 35100 Padova, Italy.
(2). The other major antigenic component of HBV that has been examined in tissue is HBcAg which is carried on the nuclear capsid. It is expressed in hepatocytes during complete virus replication (3) localized predomi- nantly within the nucleus of the infected hepatocyt’e, although there may be additional expression in the cy- toplasm and on the plasma membrane (4,5).
A further antigenic system of HBV, HBeAg and its antibody (anti-HBe) are closely related to HBcAg with the epitopes of the e antigen being carried on the same viral polypeptide as HBcAg (6). In the past, it has been difficult to obtain specific antibody to HBeAg since appropriate sera from human sources would almost in- variably have been contaminated with anti-HBc. This difficulty has meant that few studies have investigated the localization of HBeAg in the liver. One report de- scribed the presence of HBeAg in the hepatocyte nuclei in association with HBcAg determinants (7) while an- other described cytoplasmic staining of liver cells (8). Both these studies were conducted using human sera containing anti-HBe that would also have contained anti-HBc activity, making impossible a precise distinc- tion between the localization of these two antigens. Monoclonal antibodies offer a new tool for the study of HBV antigens in the liver. Recently, mouse monoclonal anti-HBc (9) and monoclonal anti-HBe (10) have been produced. We have used these reagents to investigate the expression of HBcAg and HBeAg in the liver of patients with chronic HBV infection, and the results are pre- sented in this report.
MATERIALS AND METHODS The production
of mouse monoclonal antibodies to HBcAg and HBeAg has been described elsewhere in detail (9, 10). Briefly, hybridomas secreting anti-HBc were prepared from mice immunized with purified liver HBcAg. Hybridomas secreting anti-HBe were prepared from mice immunized with HBeAg obtained by exten- sive denaturation of liver HBcAg. Antibody-secreting hybri- domas were detected in reverse capture radioirnmunoassays (RIA), and the specificity of the monoclonal antibodies con- firmed by cross-inhibition studies using monoclonal and poly- clonal reagents. In these experiments, monoclonal anti-HBc antibodies inhibited binding of radiolabeled polyclonal anti- HBc to immobilized liver-extracted HBcAg by more than 50%. Similarly, monoclonal and anti-HBe antibodies inhibited bind-
Monoclonal Anti-€€Be and Anti-€€Be.
199

200 MONDELLI ET AL. HEPATOLOGY
ing of polyclonal anti-HBe to immobilized HBeAg, purified from human serum, by more than 50%. In contrast, none of the monoclonal anti-HBe was able to inhibit the binding of polyclonal human anti-HBc (prepared from HBeAg-positive serum) to HBcAg, and none of the monoclonal nor the poly- clonal anti-HBc inhibited the binding of monoclonal anti-HBe to HBeAg. One monoclonal anti-HBc antibody (F7H10) and one monoclonal anti-HBe antibody (e6) were selected as re- agents for the immunofluorescence study as they were found to give the best staining on liver biopsy tissue from HBeAg- positive HBsAg carriers which were known to contain HBcAg. Hybridomas secreting these antibodies were grown in RPMI- 1640 supplemented with 20% fetal calf serum and the super- natant fluid (SNF) used as a source of specific antibody.
Liver Biopsies and Immunofluoreacence. Immunoflu- orescence was performed on unfixed cryostat sections of liver obtained from 25 HBsAg-positive carriers. Of these, 15 had HBeAg in the serum. Three of this group had histological evidence of chronic persistent hepatitis, the remaining 12 had chronic active hepatitis (CAH). The other 10 pat,ients had anti- HBe in their serum, of whom 5 had chronic persistent hepatitis, 3 had CAN and 2 had cirrhosis. Sections from 10 HBsAg- negative patients with CAH (six with superimposed cirrhosis) were also studied. All biopsies were initially tested by direct immunofluorescence using human anti-HBc serum conjugated with fluorescein isothiocyanate. The IgG for conjugation was prepared from the serum of healthy HBsAg carrier by ion- exchange chromatography and was positive for anti-HBc ( 1:106 by RIA) and for anti-HBe (1:102 by RIA). It did not contain anti-HBs, antidelta or antinuclear antibodies. Immunofluores- cence with monoclonal anti-HBc and anti-HBe was performed on unfixed cryostat sections by an indirect method. Sections were incubated for 45 min at 37°C with SNF from monoclonal hybridoma cultures diluted 1: lO in phosphate-buffered saline (PBS). After several washings in PBS, sections were reacted with fluorescein isothiocyanate-conjugated goat antimouse im- munoglobulin for 30 min at 37°C. Sections were subsequently washed in PBS and mounted in PBS glycerin. As a control, sections were also reacted with antimouse immunoglobulins, omitting the incubation with the monoclonal antibodies. In some instances, SNF from monoclonal hybridomas secreting anti-rubella I& was used as a control to confirm the specificity of fluorescence.
HBsAg, anti-HBc, HBeAg and anti-HBe were detected by commercially available RIA kits (Abbott Laboratories, Ltd., North Chicago, Ill.). HBV DNA polymerase levels in serum were measured by the method previously described (11).
RESULTS
RBVMarkers in Serum.
Detection of HBcAg and HBeAg in Liver with Polyclonal and Monoclonal Antibodies. As shown in Table 1, liver biopies from 13 of 15 HBeAg-positive patients contained HBcAg detected by polyclonal anti- HBc. Positive biopsies showed variable numbers of stained nuclei, and in several cases, cytoplasmic reactiv- ity was also observed (Figure 1). All 13 biopsies also reacted with monoclonal anti-HBe, and all but one with monoclonal anti-HBc. The two biopsies that did not stain with polyclonal anti-HBc were also unstained by both monoclonal antibodies.
Liver biopsies from 3 of 10 patients whose serum was anti-HBe positive reacted with polyclonal anti-HBc. One of these, three reacted with monoclonal anti-HBc, but none reacted with monoclonal anti-HBe. From this, it can be seen that all nine biopsies from HBsAg-positive patients which were unreactive with polyclonal anti-HBc
TABLE 1. DETECTION OF HBCAG AND HBEAG IN LIVER BIOPSIES BY POLYCLONAL AND MONOCLONAL ANTIBODIES
No. of cases with positive immunofluorescence
serologic pattern No. of cases Human Monoclonal Monoclonal
anti-HBc anti-HBc anti-HBe
HBsAg/HBeAg positive 15 13 12 13 HBsAg/anti-HBe positive 10 3 1 0 HBsAg negative 10 0 0 0
were also unreactive with both monoclonal antibodies. None of the 10 liver biopsies from HBsAg-negative pa- tients reacted with polyclonal anti-HBc or with the two monoclonal antibodies. None of the biopsies was stained when antimouse immunoglobulin was used in the ab- sence of antiviral monoclonal antibodies, thus excluding binding of the second antibody to hepatocyte-fixed hu- man immunoglobulins.
Pattern of Immuno fluorescence with Monoclo- nal Anti-HBc and with Monoclonal Anti- HBe. Although nearly all liver biopsies displayed con- cordance for HBeAg and HBcAg expression, being either positive or negative for both antigens, the pattern of immunofluorescence and the percentage of positive cells obtained with the monoclonal anti-HBc or with the monoclonal anti-HBe antibody differed in several biop- sies. Monoclonal anti-HBc was responsible for nuclear staining affecting variable percentages of hepatocytes (Figure 2) in all 13 reactive biopsies, and cytoplasmic staining was observed in only two cases. In six patients, occasional hepatocytes displayed linear peripheral fluo- rescence suggestive of membrane HBcAg expression. In contrast, staining with monoclonal anti-HBe showed cytoplasmic reactivity coexisting with nuclear fluores- cence in 10 of the 13 liver biopsies which stained with anti-HBe (Figure 3), while in the remaining three, only nuclear fluorescence was observed. In some cases, occa- sional hepatocytes displayed exclusive cytoplasmic stain- ing with monoclonal anti-HBe, a pattern never observed with anti-HBc. Typical membrane fluorescence was not observed using monoclonal anti-HBe.
The percentage of liver cells reactive with monoclonal anti-HBc and with monoclonal anti-HBe was grossly similar in nine patients. In three other patients, most hepatocytes were stained by monoclonal anti-HBe while monoclonal anti-HBc reacted only with isolated hepa- tocyte nuclei. Finally, as already mentioned, there were two discordant biopsies. In the first, the liver biopsy was strongly reactive with anti-HBe, showing staining of the cytoplasm of scattered hepatocytes (Figure 4), but was unreactive with anti-HBc. The other liver biopsy showed scattered nuclear reactivity with monoclonal anti-HBc but no staining with monoclonal anti-HBe. No nonspe- cific reactivity was seen when control SNF was used to examine discordant biopsies.
Detection of HBcAg and HBeAg in the Liver in Relation to Virus Replication Activity. To inves- tigate the significance of the presence of HBcAg and HBeAg determinants in the liver in relation to virus replication, the results obtained by immunofluorescence with monoclonal anti-HBc and with monoclonal anti- HBe were compared with serum measurement of HBV
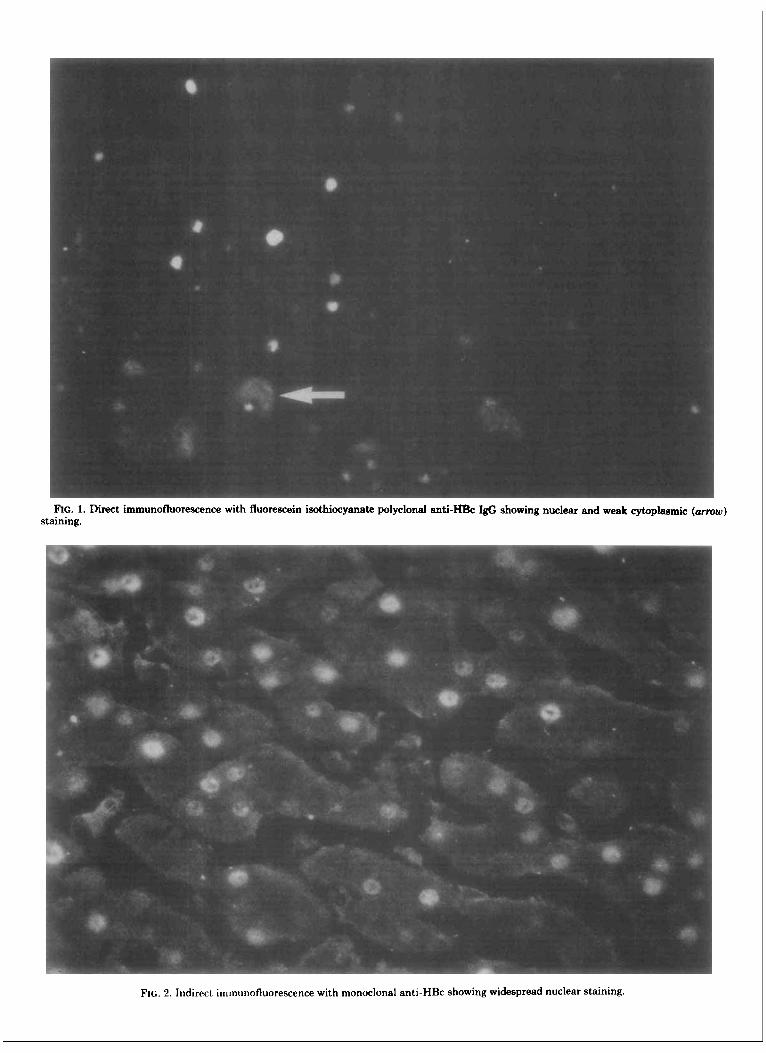
FIG. 1. Direct immunofluorescence with fluorescein isothiocyanate polyclonal anti-- I& showing nuclear and weak cytoplasmic (arrow) staining.
FIG. 2 . Indirect immunofluorescence with monoclonal anti-HBc showing widespread nuclear staining.

202 MONDELLI ET AL. HEPATOLOGY
FIG. 3. Indirect immunofluorescence with monoclonal anti-HBe showing widespread nuclear and cytoplasmic (arrows) staining.
DNA polymerase activity in 23 of the 25 HBsAg chronic carriers studied. There were seven patients in whom there was widespread staining of hepatocytes with mono- clonal anti-HBc. All of them had detectable HBV DNA polymerase activity in serum; this included the single patient whose serum contained anti-HBe and whose biopsy did not react with monoclonal anti-HBe. Four patients had only a few hepatocyte nuclei reacting with monoclonal anti-HBc, and only one of these had demon- strable HBV DNA polymerase activity in the serum. Two patients in this group, both negative for serum HBV DNA polymerase activity, had large numbers of hepato- cytes which were stained with monoclonal anti-HBe. Sera from the 12 patients whose biopsies were negative for HBcAg, including the single case found positive for HBeAg in the liver and in serum, were found not to contain HBV DNA polymerase activity.
Relationship Between Expression of HBcAg and HBeAg and Histotogic Activity. All of the three patients with chronic persistent hepatitis found positive in the liver by immunofluorescence showed a diffuse pattern of distribution of both HBcAg and HBeAg. Among the 11 patients with CAH and viral antigen expression in the liver, 9 displayed focal HBcAg expres- sion, including 6 with associated focal HBeAg and 3 with diffuse HBeAg. One with focal HBcAg was negative for
HBeAg and one with focal HBeAg was negative for HBcAg. Thus, there was a trend towards an inverse relationship between the amount of viral antigens, par- ticularly of HBcAg, in the liver and histologic activity, as previously described in larger series of patients (12).
DISCUSSION In this study, monoclonal antibodies to the core and e
antigens of HBV were used to localize the expression of these antigens within the hepatocytes. Previous attempts to differentiate between HBcAg and HBeAg have been hampered by the redundant immunoglobulin in the re- agents. As a result, background staining is often marked, and where the unwanted immunoglobulin contains anti- HBV specificities other than the desired one, patterns of staining may be very difficult to interpret. To a great extent, ascitic fluids containing high levels of monoclonal antibody avoid this problem since the majority of the immunoglobulin will be specific antibody. However, problems have been encountered where the mouse used for raising the ascites has been found to produce signifi- cant antibody reacting with the membranes of the hy- bridoma tumor and other cells. This may be avoided, as in the study described here, by the use of SNF from antibody-secreting hybridomas maintained in vitro, in spite of the level of immunoglobulin in SNF being mark-

Vol. 6, No. 2, 1986 HEPATITIS R CORE AND E ANTlGENS IN HEPATOCYTES 203
FIG. 4. Indirect immunofluorescence with monoclonal ant i-HBe showing almost exclusive cytoplasmic staining.
edly lower than that contained in ascitic fluid. As shown here (Table l), the specificity of the SNF and polyclonal anti-HBc was similar, both in prevalence and in pattern of staining for HBcAg.
Using SNFs containing either monoclonal anti-HBc or anti-HBe, it was usual to find that biopsies stained most commonly for both HBcAg and HBeAg. In all but two instances this was so, suggesting as might be ex- pected, a close link between the presence of these two antigens. However, in spite of this concordant antigen expression, their localization within hepatocytes was dif- ferent, with the bulk of HBcAg expression being limited to the hepatocyte nuclei while a significant amount of HBeAg was seen in hepatocyte cytoplasm as well as nuclei. In the extreme instance, hepatocytes could be identified, which showed only cytoplasmic HBeAg expression, and in one of the two discordant biopsies, HBeAg was present in the hepatocyte cytoplasm in the absence of expression of HBcAg and with only minimal nuclear expression of HBeAg. Since the gene for HBcAg codes for P22, a protein which is able to express both core and e determinants, it is not surprising that both antigens should be expressed simultaneously. What is interesting is the different localization of these antigens within the infected hepatocyte, suggesting independence of expression. Although P22 is able to express both determinants, it is not clear from the amino acid se- quence which conformation the protein adopts initially
posttranslation. Thus, it could be that, following trans- lation, P22 self-assembles immediately into core struc- tures, which display HBcAg and mask the usual "e" determinants as has been suggested by Ferns and Tedder (10). Subsequent degradation of core, perhaps coupled with cleavage of the polypeptide at amino acid 149 and loss of the remaining residues from the c terminal end (13), leads to loss of the HBcAg determinants, expression of HBeAg and its passage into the serum. Alternatively, initial cleavage of P22 present as HBeAg in the hepato- cyte could prevent assembly into cores and lead to pas- sage of HBeAg straight into serum without a core inter- mediate. The different. localization of the two antigens, the finding of HBeAg-only-staining hepatocytes and the biopsy, albeit only one, which stained strongly for HBeAg alone would be compatible with HBeAg, being a precur- sor which does not necessarily imply the subsequent formation of cores and hence intact hepatitis virions. Thus, the data presented here would be compatible with synthesis in the cytoplasm of HBeAg which is then transported to the nucleus where assembly into HBcAg around the HBV nucleic acid might occur.
That HBeAg should represent unpolymerized P22 rather than degraded cores would be substantiated by the slightly better correlation of serum hepatitis B vire- mia, measured by serum HBV DNA polymerase, with HBcAg expression in the liver rather than with hepato- cyte HBeAg. Although the number of patients was very

204 MONDELLI ET AL. HEPATOLOGY
small, it seemed that HBeAg could be well-expressed in the liver, either in the presence of scattered HBcAg staining nuclei (two patients) or in its absence (one patient), without the presence of HBV in the blood. Taken together with the antigen expression data, this would strengthen the case for HBeAg being a prepo- lymerization gene product rather than a degradation produce of cores. On the other hand, it should also be mentioned that recent evidence indicates that HBcAg and HBeAg may be transcribed independently from the preC/C gene (Ou, J. et al., Cold Spring Harbor Labora- tory Meeting, May 1985, No. 22, Abstract).
The physical nature of HBcAg and HBeAg in the hepatocytes is not yet precisely known. Preliminary ev- idence indicates that at least certain of the HBcAg epitopes are expressed upon the hepatocyte membrane (5, Mondelli, M. et al., Liver 1984; 4:41, Abstract), al- though their exact nature, whether partially polymer- ized or individual P22 proteins, remains to be deter- mined. The demonstration of HBcAg-specific cytotoxic T ceils in HBV infection (14, 15) and the finding that chimpanzees immunized with recombinant HBcAg are protected from developing hepatitis B (16) would suggest that the host response to P22 is fundamentally more important than hitherto recognized.
Acknowledgments: We thank our colleagues in the Virology Section, Middlesex Hospital Medical School, for helpful discussions and Bina Dave for preparing the manuscript.
REFERENCES 1. Gudat F, Bianchi L, Sonnabend W, et al. Pattern of core and
surface antigen expression in liver tissue reflects state of specific immune response in hepatitis B. Lab Invest 1975; 32:l-9.
2. Ray MB, Desmet VJ, Bradburne AF, et al. Differential distribution of hepatitis B surface antigen and hepatitis B core antigen in the liver of hepatitis B patients. Gastroenterology 1976; 71:462-469.
3. Montano L, Waters J , Goodall AH, et al. Detection of HBsAg and HBcAg in human liver using monoclonal and heterologous anti-
bodies: relationship to hepatitis B viral replication and inflamma- tory activity. Liver 1985; 570-76.
4. Bianchi L, Gudat F. Immunopathology of hepatitis B. In: Popper H, Schaffner F, eds. Progress in liver disease, Vol 6. New York: Grune and Stratton, 1979 p. 371.
5. Trevisan A, Realdi G, Alberti A, et al. Core antigen specific immunoglobulin G bound to the liver cell membrane in chronic hepatitis B. Gastroenterology 1982; 82:218-222.
6. Mackay P, Lees J, Murray K. Conversion of hepatitis B core antigen synthesized in E. coli into e antigen. J Med Virol 1981;
7. Arnold W, Nielsen JO, Hardt F, et al. Localisation of e antigen in nuclei of hepatocytes in HBsAg positive liver diseases. Gut 1977;
8. Trepo C, Vitvitski L, Neurath R, et al. Detection of e antigen by immunofluorescence in cytoplasm of hepatocytes of HBsAg car- riers. Lancet 1976; 1:486.
9. Tedder RS, Guarascio P, Yao JL, et al. Production of monoclonal antibodies to hepatitis B surface and core antigens, and use in detection of viral antigens in liver biopsies. J Hyg 1983; 90:135- 142.
10. Ferns RB, Tedder RS. Monoclonal antibodies to hepatitis B e antigen (HBeAg) derived from hepatitis B core antigen (HBcAg): their use in characterization and detection of HBeAg. J Gen Virol
11. Alberti A, Pontisso P, Realdi G. Changes in hepatitis B virus DNA polymerase activity in patients with chronic infection. J Med Virol
12. Alberti A, Tremolada F, Bortolotti F, et al. Virus replication and liver disease in chronic hepatitis B infection. Dig Dis Sci 1983;
13. Takahashi A, Machida A, Funatsu G, et al. Immunochemical structure of hepatitis B e antigen in the serum. J Immunol 1983;
14. Naumov NV, Mondelli M, Alexander GJM, et al. Relationship between expression of hepatitis B virus antigens in isolated hepa- tocytes and autologous lymphocyte cytotoxicity in patients with chronic hepatitis B virus infection. Hepatology 1984; 4:63-68.
15. Mondelli M, Mieli-Vergani G, Alberti A, et al. Specificity of T- lymphocyte cytotoxicity to autologous hepatocytes in chronic hep- atitis B virus infection: evidence that T cells are directed against HBV-core antigen expressed on hepatocytes. J Immunol 1982;
16. Murray K, Bruce SA, Hinnen A, et al. Hepatitis B virus antigen made in microbial cells immunise against viral infection. The
8237-243.
19~994-996.
1984; 65899-908.
1981; 8223-229.
28~962-967.
131:2903-2907.
1292773-2778.
EMBO J 1984; 3~645-650.





























