Embed Size (px)
Citation preview

Behavior Genetics, Vol. 32, No. 2, March 2002 (© 2002)
1270001-8244/02/0300-0127/0 © 2002 Plenum Publishing Corporation
Differences Between Two Psychogenetically Selected Linesof Rats in a Swimming Pool Matching-To-Place Task:Long-Term Effects of Infantile Stimulation
Raúl Aguilar, 1,2 Rosa M. Escorihuela,1 Luis Gil, 1 Adolf Tobeña,1
and Alberto Fernández-Teruel1
Received 17 June 2001—Final 26 Oct. 2001
The present study explored the effects of infantile stimulation (i.e., neonatal handling or NH)on the performance of 18-month-old Roman high-avoidance (RHA/Verh) and low-avoidance(RLA/Verh) rats in a swimming pool matching-to-place (SPMP) test. This test (also called re-peated acquisition and place learning-set paradigm) consists of administering pairs of consecu-tive trials in the Morris water maze. The difference between each “odd” and the consecutive“even” trial of a trial pair is considered to be a measure of working memory. The same rats werefirst tested for exploration and novelty-seeking in a hole-board test in the presence of novel ob-jects, which showed that RHA/Verh rats were more explorative than their RLA/Verh counter-parts, and that NH treatment augmented exploration in RLA/Verh rats, generally eliminatingthe genetically-based differences between the lines. RHA/Verh rats performed less efficientlythan RLA/Verh rats in the SPMP test, and NH facilitated acquisition in the early stages of train-ing in both rat lines, an effect that was presumably due to an improvement in the acquisition ofspatial reference information. Performance during training also indicated that RHA/Verh ratsshowed less differentiated behavior between odd and even trials, indicating a relative workingmemory deficit at advanced ages in that rat line.
KEY WORDS: Roman high- and low-avoidance rats; fearfulness; exploration; hole-board; spatial work-ing memory; Morris water maze, neonatal handling.
1967; Meaney et al., 1988; Núñez et al., 1996; Wil-son et al.,1986). NH also improves learning and mem-ory abilities in rats, as measured by performance inseveral tests, such as two-way active avoidance (Es-corihuela et al., 1994; Levine, 1956; Levine andWetzel, 1963; Núñez et al.,1995), step-down and step-through passive avoidance (Ader and Schaefer, 1968;Gschanes et al.,1995, 1998), conditioned punishmentin a brightness discrimination learning situation(Wong, 1972), latent inhibition in Pavlovian condi-tioning (Feldon and Weiner, 1988; Lehmann et al.,1998; Peters et al., 1991), and spatial navigation im-pairments in aged rats in the Morris water maze(Meaney et al., 1988, 1991). This body of evidencesupports the hypothesis that infantile stimulation worksas a cognitive enhancer, which ameliorates learningprocesses (Wong, 1972).
INTRODUCTION
Mild infantile stimulation (i.e., neonatal handling, NH)attenuates emotional reactivity to stress or conflict andincreases exploratory behavior in adult rodents (Anis-man et al.,1998; Daly, 1973; Escorihuela et al.,1994;Ferré et al.,1995b; Meaney et al.,1988; Núñez et al.,1995; Wong, 1972), through long-term neurobiologi-cal changes (Bodnoff et al., 1987; Escorihuela et al.,1992; Fernández-Teruel et al., 1997; Levine et al.,
1 Medical Psychology Unit, Dept. of Psychiatry and Forensic Med-icine, Autonomous University of Barcelona, 08193-Bellaterra,Barcelona, Spain.
2 To whom all correspondence should be addressed. Medical Psy-chology Unit, Department of Psychiatry and Forensic Medicine,School of Medicine, Autonomous University of Barcelona, 08193Bellaterra, Barcelona, Spain. Tel: 134-93-581 23 78. Fax: 134-93-581 14 35. e-mail: [email protected]

The Roman rat lines (of which several sublinesexist, or have existed, at various laboratories) were se-lected and bred for superior (Roman high-avoidance)vs. inferior (Roman low-avoidance) acquisition of ac-tive, two-way shuttle box avoidance (Bignami, 1965).Research with the Swiss Roman sublines (RHA/Verhand RLA/Verh), derived from the original RHA andRLA stock (Bignami, 1965) and bred in Switzerlandsince 1972 (Driscoll and Bättig, 1982; Driscoll et al.,1998), has shown that as a correlative effect of psy-chogenetic selection these lines differ in (1) emotionalreactivity to stress and conflict, and in (2) performancein a number of learning and memory tasks. Comparedto RHA / Verh rats, RLA / Verh rats show increaseddefecation and plasma levels of corticosterone, ACTH,and prolactin and more pronounced freezing andacoustic startle responses when mildly stressed (Aguilaret al.,2000; Fernández-Teruel et al.,1992; Ferré et al.,1995a; Gentsch et al.,1982; Schwegler et al.,1997; forreviews, see Driscoll and Bättig, 1982; Driscoll et al.,1998; Escorihuela et al.,1995a). In learning and mem-ory tasks, RLA/Verh rats perform more efficiently inan aquatic Hebb-Williams maze (Nil and Bättig, 1981),in the Morris water maze (Driscoll et al., 1995;Escorihuela et al., 1995b), and in the DRL-20 bar-pressing task for food reward (Zeier et al., 1978) thanRHA/Verh rats. Other Roman rat sublines were alsostudied in several working memory procedures, inwhich RLA rats performed more efficiently than RHAanimals in a delayed reinforcement (and in a sponta-neous) alternation T maze task, in an object recogni-tion test, and in an eight-arm radial maze (Willig et al.,1991, 1992).
The standard Morris water maze (refered to above)consists of administering successive trials from differ-ent starting points of the water tank, without disturb-ing the location of the submerged platform across train-ing sessions. It evaluates the ability of the rat to escapefrom the water by finding the platform using extramazeorientation cues (Morris, 1981). Reference memory isthe main process measured with this procedure. Thepurpose of the present study was twofold: First, to char-acterize old age, (18 months) rats of the Roman/Verhlines in a different version of the Morris water maze,devised to study working memory in a spatial task. Weused SPMP (Whishaw, 1985, 1995). This task consistsof administering pairs of consecutive trials from dif-ferent starting points of the water tank (each trial pairbeing given in a pseudorandom manner), under the con-dition of changing the position of the platform aftereach training day: The discrepancy between odd and
128 Aguilar, Escorihuela, Gil, Tobeña, and Fernández-Teruel
even trials of each trial pair is the measure of the work-ing memory component (Klapdor et al.,1998; Lindneret al., 1992; Wesierska et al., 1990). The acquisitionof spatial information that takes place during one sin-gle trial is assessed in this paradigm: the rat’s capacityto use new, recently acquired information can be pro-cedurally isolated in a water maze (Whishaw, 1985).We expected that the old RLA/Verh rats would be su-perior in the spatial navigation task studied here. Wealso evaluated swimming pool navigation in a cued taskto rule out the differential contribution of other behav-ioral (e.g., motivational or sensorial) processes to spa-tial learning.
Second, we sought to establish whether NH couldameliorate performance in that task. Assuming that NHenhances learning abilities, positive effects of thattreatment would be predicted on spatial navigation per-formance. We did not anticipate improvement effectsof NH in RLA/Verh rats (i.e., ceiling effect), but weexpected NH to increase performance in their RHA/Verh counterparts. In conjunction with that experi-ment, we first studied the exploration and novelty-seeking behavior of the same 18-month-old, NH andnon–NH animals, in a hole-board test in the presenceof novel objects.
MATERIALS AND METHODS
Subjects
The subjects were outbred RHA/Verh and RLA/Verh male rats born and maintained at the Institut fürNutztierwissenschaften, ETHZ, Zürich. Within 1 to2 weeks after weaning (postnatal day 22), the rats weregroup-housed (2 to 3 per cage, same sex) and main-tained with food and water available ad lib, under con-ditions of controlled temperature (21°C to 22°C) and alight-dark schedule of 12 hours each (lights on at 9 AM).The rats arrived at the Autonomous University ofBarcelona at 2 months of age. The experiments wereconducted during the illuminated phase of the light-dark cycle when the rats were 18 months old (weight5 536 6 7 g at the beginning of the experiments).
Neonatal Handling (NH) Treatment
At postnatal day 1, the handling treatment started,which consisted of first removing the mother, and thenthe pups (10 to 12 per litter), from the nest twice daily(first time between 1030 and 1300, and the second timebetween 1900 and 2100). The first daily handling ses-

sion consisted of first separating and weighing eachmother, and then weighing the pups and placing themgently and individually in plastic cages lined with papertowel for a total period of 10 minutes. After 5 minutesin this situation, each pup was individually (and gently)handled for 5 seconds and returned to the same cagefor the remaining 5 minutes. At the end of the 10-minute period, each pup was gently handled for another3 seconds and then returned to its homecage. When allthe pups from one litter were back in their homecage,the mother was then returned to the homecage. Thesame procedure (without weighing the animals) wasconducted in the evening (second time). NH finishedat postnatal day 21 (1 day before weaning). Control(CON) nonhandled groups were left undisturbed untilweaning. Thus, 4 experimental groups of rats wereformed (RHA/NH, RHA/CON, RLA/NH and RLA/CON). The animals used in each group were evenlyselected from 5 to 6 different litters.
Apparatus
Hole-Board Test
The apparatus was a white 66 cm 3 66 cm 347 cm wooden box, which had four equidistant holes(3.7 cm in diameter, 18 cm deep) in the floor. The floorwas divided into 16 equal squares with red lines. Thefour holes contained identical objects (i.e., a miniaturecar and a little plastic ball in a metal container) strangeto the animals. Previous work at our laboratory hasshown that differences in exploratory behavior betweenRHA/Verh and RLA/Verh rats are augmented by plac-ing such novel objects under the holes (Escorihuelaet al., 1999).
Matching-to-Place Test
The apparatus was a large, round plastic swim-ming pool (1.4 m in diameter, 0.3 m deep) with whitewalls. The swimming pool was filled with water (24°C)made opaque with skim milk, that was changed everyday. The platform (16 cm in diameter, 28 cm high) wasinvisible under the water surface. There were no local(intramaze) cues available. The experimental room hadseveral extramaze cues that the rats could see from thesurface of the swimming pool, such as three windowsin a wall; an elevated, large table holding several lab-oratory objects; several fluorescent lights in the ceil-ing; the registration equipment (computer and screen);and a video camera mounted above the center of theswimming pool.
Infantile Stimulation Influences Spatial Learning in Aging 129
Procedure
Hole-Board
Ambulation (number of squares crossed), thenumber of head dips, and time spent head dippingwere measured for 5 minutes. Forty-six NH/non-NHbut otherwise naive rats (n 5 10 to 13 per group) wereused in this test, which was followed, 7 days later, bythe swimming pool experiment described later.
Matching-to-Place
Using rats from the hole-board procedure (n 5 7to 9 per group), the test consisted of two phases sepa-rated by a 48-hour interval. The first phase consistedof 6 days of training, similar to that used by Whishaw(1985), in which the location of the hidden platformwas changed daily, and the starting points of each trialpair were randomly rotated. The second phase was acontrol procedure of cue learning on 2 final days.
Phase 1. Matching-to-Place Task.The platformwas moved each day according to the following pseudo-random sequence: E, SW, center, NW, E, and SW. Fourstarting points (N, S, E, and W), randomly determined,were used throughout each daily session. The rats weregently placed in the water, facing the wall of the swim-ming pool. They underwent eight daily trials adminis-tered in pairs (four pairs each day). The first trial of eachtrial pair finished when the rats found the platform and,if this was not the case after 60 sec, they were placedon the platform by the experimenter. In both instances,the rats remained on the platform for 30 sec. The sec-ond trial of the trial pair was administered immediatelyfrom the same starting point. Once a trial pair finished,the rats were returned to their homecages (in the sameexperimental room) for a period of 40 to 50 minutesafter which the following trial pair was given. A com-puterized tracking system (Smart, Letica SA, Barcelona)connected to the video camera provided the main mea-surements used, i.e., escape latency, path length, andnavigation speed.
Phase 2. Cue Task.During this second phase, theplatform protruded above the water surface (1.5 cm),so that it was visible. In order to additionally facilitatethe visibility of the escape platform, a striped (blackand white) flag pole (20 cm high; 1 cm in diameter)holding the flag was placed on the platform. Room(extra-maze) cues were drastically reduced by increas-ing the height of the swimming pool walls with card-board. As compared to phase 1, these changes consid-erably modified the contextual and spatial stimuli. The

rats performed eight daily trials on 2 successive days.The starting points of each individual trial were ran-domly assigned (first day: S, N, E, W, E, N, S, and W;and second day: W, S, N, E, S, E, W, and N). On thefirst day, the visible platform was located in the centerof the NE quadrant and, on the second day, it was inthe center of the SE quadrant.
STATISTICS
A between-groups experimental design was em-ployed in the hole-board experiment. Ambulation, num-ber of head dips, and time spent head dipping were con-sidered exploratory behaviors in that test. The data wereanalyzed by two-way ANOVA and Duncan’s multiplerange tests were applied for comparisons betweengroups after significant ANOVA results. Relationshipsbetween escape latency in the swimming pool task andtime spent head-dipping in the hole-board test wereanalyzed by the Pearson correlation coefficient (r).
A repeated-measures between-groups experimen-tal design was used in the matching-to-place experi-ment. Two measures were analyzed: (1) escape latency(i.e., the time taken to escape onto the hidden platform),and (2) navigation speed. A two-way ANOVA(between-subject factors: line 3 treatment), with sessionsas a within-subject factor, was applied to navigationspeed. The learning curves across the six trainingsessions were analyzed by (1) a two-way ANOVA(between-subject factors: line 3 treatment) with re-peated measures over sessions, and (2) a three-wayANOVA (between-subject factors: line 3 treatment 3type of trial, (i.e., odd or even) with repeated measuresover sessions. Paired t-tests (when comparing differenttypes of trials within a group) and Duncan’s multiplerange tests (for between-group comparisons) were ap-plied after significant ANOVA results.
Navigation speed and escape latency (means ofthe two sessions) in the cue phase were analyzed byapplying a one-way ANOVA with groups as thebetween-subject factor.
RESULTS
Hole-Board
RHA/ Verh rats showed more ambulations(ANOVA, line effect, F1,42 5 8.9, P , 0.005; see TableI) and a longer time spent head dipping than did theirRLA/Verh counterparts. The analysis of variance re-vealed line 3 NH interactions in the number of head
130 Aguilar, Escorihuela, Gil, Tobeña, and Fernández-Teruel
dips and the time spent head dipping (interaction ef-fect, line 3 NH on both parameters, F1,42 5 6.86, P ,0.012 and F1,42 5 10.1, P , 0.003, respectively;Table I), indicating that NH influences both strains inopposite ways. In addition, the Duncan’s test showedthat NH exposed RLA/Verh rats performed a greaternumber of head dips (P , 0.05) and spent a longer timehead dipping (P , 0.05) than did their CON group.
Matching-to-Place
The analysis of mean navigation speed yielded nostatistically significant differences among groups (F15,145
5 0.27, P 5 0.1). Escape latency was used as the maindependent variable for statistical analysis. The mean es-cape latency of 8 trials/session is shown in Fig. 1. Thetwo-way ANOVA (between-subject factors: line 3 NH)with repeated measures over sessions showed session(F5,145 5 28.7, P , 0.000) and NH 3 session effects(F5,1455 4.23, P , 0.019), indicating that the effects ofNH were more pronounced at the beginning of learning.Thus, it can be seen that all animals gradually learnedto escape onto the submerged platform as a result oftraining and that neonatal handling facilitated acquisi-tion in the early stages of training, especially in theRHA/Verh rats. The RHA/CON group actually exhib-ited poorer performance than the other groups in the firsttraining session (P , 0.05, Duncan’s test).
The mean escape latency of the 4 odd trials ver-sus the corresponding 4 even trials of each session isrepresented in Fig. 2. The RLA/CON rats showed a bet-ter overall performance than RHA/CON rats, and han-dled rats (RLA/NH and RHA/NH) showed shorter es-cape latencies than did nonhandled rats, especially inthe early training sessions. RLA/CON rats also showed
Table I. Mean 6 SE of Behavioral Measures in the Hole-BoardTest Shown by Control (RHA/CON and RLA/CON)
and Handled (RHA/NH and RLA/NH) Rats
Number of Time spent Ambulation head dips head dippinga
RHA/CON 36.3 6 4.4* 10.9 6 2.3 7.2 6 .8*RHA/NH 29.9 6 6.9 7.7 6 1.8 5.5 6 .5RLA/CON 19.7 6 2.5 7.7 6 1.2** 5.1 6 .6**RLA/NH 20.5 6 3.6 13.3 6 1.5 8.2 6 .8
a The values of “time spent head dipping” are the square root trans-formation (to achieve homogeneity of variances) of the raw scoresof the variables (in seconds).
*p , 0.05 vs. RLA/CON (Duncan’s test).** p , 0.05 vs. RLA/NH (Duncan’s test).
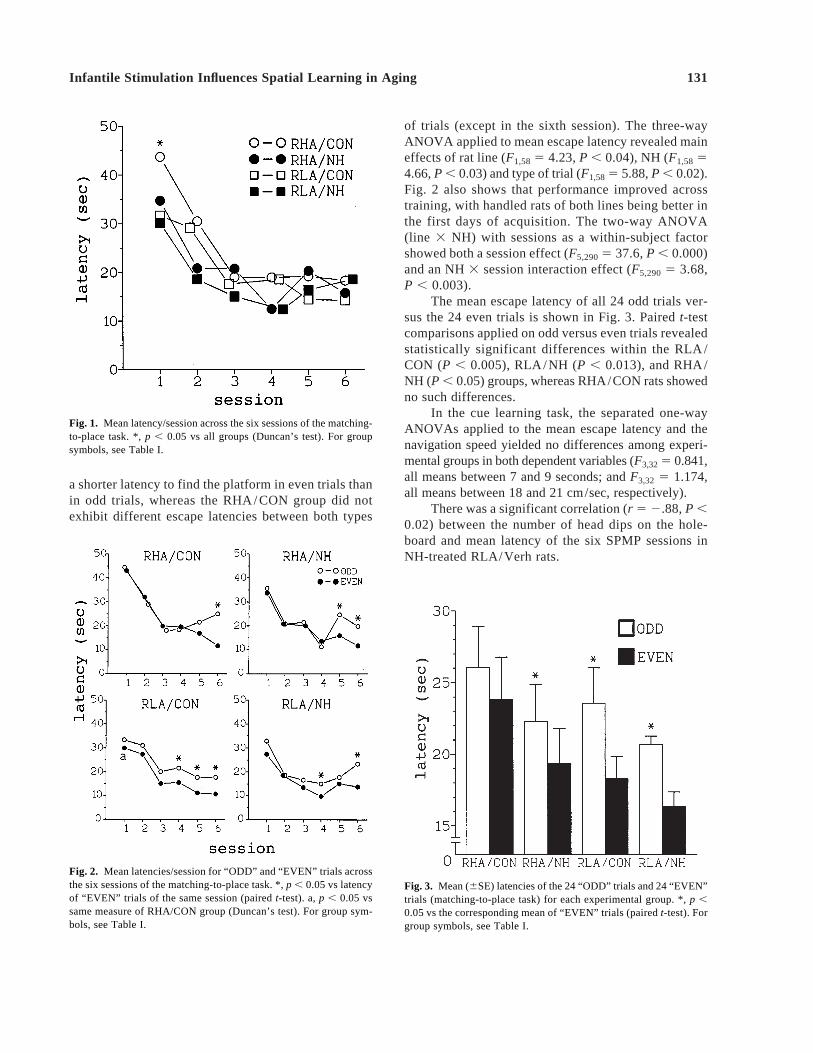
a shorter latency to find the platform in even trials thanin odd trials, whereas the RHA/CON group did notexhibit different escape latencies between both types
Infantile Stimulation Influences Spatial Learning in Aging 131
of trials (except in the sixth session). The three-wayANOVA applied to mean escape latency revealed maineffects of rat line (F1,58 5 4.23, P , 0.04), NH (F1,58 54.66, P , 0.03) and type of trial (F1,585 5.88, P , 0.02).Fig. 2 also shows that performance improved acrosstraining, with handled rats of both lines being better inthe first days of acquisition. The two-way ANOVA(line 3 NH) with sessions as a within-subject factorshowed both a session effect (F5,2905 37.6, P , 0.000)and an NH 3 session interaction effect (F5,290 5 3.68,P , 0.003).
The mean escape latency of all 24 odd trials ver-sus the 24 even trials is shown in Fig. 3. Paired t-testcomparisons applied on odd versus even trials revealedstatistically significant differences within the RLA/CON (P , 0.005), RLA/NH (P , 0.013), and RHA/NH (P , 0.05) groups, whereas RHA/CON rats showedno such differences.
In the cue learning task, the separated one-wayANOVAs applied to the mean escape latency and thenavigation speed yielded no differences among experi-mental groups in both dependent variables (F3,325 0.841,all means between 7 and 9 seconds; and F3,32 5 1.174,all means between 18 and 21 cm/sec, respectively).
There was a significant correlation (r 5 2.88, P ,0.02) between the number of head dips on the hole-board and mean latency of the six SPMP sessions inNH-treated RLA/Verh rats.
Fig. 3. Mean (6SE) latencies of the 24 “ODD” trials and 24 “EVEN”trials (matching-to-place task) for each experimental group. *, p ,
0.05 vs the corresponding mean of “EVEN” trials (paired t-test). Forgroup symbols, see Table I.
Fig. 2. Mean latencies/session for “ODD” and “EVEN” trials acrossthe six sessions of the matching-to-place task. *, p , 0.05 vs latencyof “EVEN” trials of the same session (paired t-test). a, p , 0.05 vssame measure of RHA/CON group (Duncan’s test). For group sym-bols, see Table I.
Fig. 1. Mean latency/session across the six sessions of the matching-to-place task. *, p , 0.05 vs all groups (Duncan’s test). For groupsymbols, see Table I.

DISCUSSION
As expected, based on previous research withmuch younger subjects, the 18-month old RHA/Verhrats used here were also more explorative in the hole-board test than the RLA/Verh rats used, and it was alsofound that neonatal handling was able to increase thebehavioral disposition of 18-month-old RLA/Verh ratsto explore new objects and environments (Fernández-Teruel et al., 1992, 1997; Steimer et al., 1998). In ad-dition, it was seen that aged RLA/Verh rats were moreefficient in a spatial, working memory navigation task.The CON rats of the RHA/Verh line did not deviatebetween odd and even trials across the first five ses-sions of the training procedure (only showing a statis-tically significant difference in session 6; see Fig. 2),whereas the RLA/Verh groups differed throughout thelast three sessions. These results suggest that the ratsof the RLA/Verh line maintain a better working mem-ory than do their RHA/Verh counterparts, even throughan advanced age, which is consistent with the findingsof previous studies using younger rats and differenttests (Escorihuela et al., 1995b; Nil and Bättig, 1981;Zeier et al., 1978).
As an extension of other studies that have dealtwith the role of early experience in learning and mem-ory (Ader and Schaefer, 1968; Escorihuela et al.,1995b; Feldon and Weiner, 1988; Gschanes et al.,1995, 1998; Lehmann et al.,1998; Levine 1956; Levineand Wetzel, 1963; Meaney et al., 1988, 1991; Peterset al.,1991; Wong, 1972), we have observed here thatneonatal handling treatment can even facilitate learn-ing performance in the spatial domain at advanced ages.This facilitation effect, however, was only found in theearly stages of training, suggesting that the psycho-logical mechanisms involved could predominantly haveto do with the acquisition and use of spatial referenceinformation. The results suggest that once the ratsformed their extra-maze stimulus representations, thetreatment effects on acquisition dissipated. Divergentemotionally or motivational factors do not appear toaccount for this early learning effect, judging by theclose proximity of the RHA / NH, RLA / CON, andRLA/NH groups seen in the spatial navigation task andof all groups in the cued task.
The effect of NH on spatial navigation in aged ratsis presumably mediated, at least in part, by neurobio-logical changes in hippocampal circuitry, a main neu-ronal site for processing spatial and emotional infor-mation (LeDoux, 1995; McNaughton and Gray, 2000;Morris et al.,1986). Preliminary evidence derived fromhistological analyses carried out in a previous study
132 Aguilar, Escorihuela, Gil, Tobeña, and Fernández-Teruel
with the same lines (Fernández-Teruel et al.,1997) sup-ports this assumption. Because the present experimen-tal design cannot determine whether NH improvedlearning abilities or retarded or prevented age-relatedimpairments, the main claim that can be made here isthat infantile stimulation was capable of affecting work-ing memory components of spatial navigation learningin 18-month-old (“elderly”) rats.
Finally, there was a strong negative correlationbetween head dipping behavior in the hole-board andmean latency of all SPMP sessions in NH-treatedRLA/Verh rats. This group presented the higher scores,exploring the unfamiliar objects under the holes, and dis-played the shortest escape latencies through the first foursessions of the spatial navigation test. It has been sug-gested by some authors (Wilson et al., 1986; Wong,1972) that exploratory behavior may be a component oflearning capacity, in such a way that the greater and moreexhaustive the exploration, the better the learning per-formance. According to this line of reasoning, it mightbe tempting to speculate that neonatal handling couldhave modified some aspect of information acquisitionmechanisms (such as attentional processes) underlyingboth exploratory and learning behaviors in the RLA/Verhrats). In fact, data collected from various latent inhibi-tion paradigms tends to support the hypothesis thatneonatal handling may increase attentional capacity(Braunstein-Bercovitz and Lubow, 1998; Feldon andWeiner, 1988; Lehmann et al.,1998; Peters et al.,1991).
ACKNOWLEDGMENTS
This work was supported by DGYCIT (PM96-0068), SGR (1998-00070), and FIS (99/1230) grants.R. A. and L. G. are supported by DGR (1998FI00158PG) and BFPI (AP98-36154188) fellowships,respectively.
REFERENCES
Ader, R., and Schaefer, A. (1968). Effects of early experience onone-trial avoidance conditioning. Psychon. Sci.10:235–236.
Aguilar, R., Gil, L., Tobeña, A., Escorihuela, R. M., and Fernández-Teruel, A. (2000). Differential effects of cohort removal stresson the acoustic startle response of the Roman/Verh rat strains.Behav. Genet.30:71–75.
Anisman, H., Zaharia, M. D., Meaney, M., and Merali, Z. (1998). Doearly-life events permanently alter behavioral and hormonal re-sponses to stressors? Int. J. Dev. Neuroscience16:149–164.
Bignami, G. (1965). Selection for high rates and low rates of avoid-ance conditioning in the rat. Anim. Behav.13:221–227.
Bodnoff, S. R., Suranyi-Cadotte, B., Quirion, R., and Meaney, M. J.(1987). Postnatal handling reduces novelty-induced fear andincreases [3H]flunitrazepam binding in rat brain. Eur. J. Phar-macol.144:105–107.

Braunstein-Bercovitz, H., and Lubow, R. E. (1998). Latent inhibi-tion as a function of modulation of attention to the preexposedirrelevant stimulus. Learn. Motiv.29:261–279.
Daly, M. (1973). Early stimulation of rodents: a critical review ofpresent interpretations. Br. J. Psychol.64:435–460.
Driscoll, P., and Bättig, K. (1982). Behavioral, emotional and neuro-chemical profiles of rats selected for extreme differences inactive, two-way avoidance performance. In: Lieblich I, (ed.).Genetics of the Brain,Elsevier Biomedical Press, Amsterdam,pp. 95–123.
Driscoll, P., Ferré, P., Fernández-Teruel, A., Levi de Stein, M., Wolf-man, C., Medina, J., Tobeña, A., and Escorihuela, R. M. (1995).Effects of prenatal diazepam on two-way avoidance behavior,swimming navigation and brain levels of benzodiazepine-likemolecules in male Roman high- and low-avoidance rats. Psy-chopharmacology122:51–57.
Driscoll, P., Escorihuela, R. M., Fernández-Teruel, A., Giorgi, O.,Schwegler, H., Steimer, T., Wiersma, A., Corda, M. G., Flint,J., Koolhaas, J. M., Langhans, W., Schulz, P. E., Siegel, J., andTobeña, A. (1998). Genetic selection and differential stress re-sponses: the Roman lines/strains of rats. Ann. NY Acad. Sci.851:501–510.
Escorihuela, R. M., Fernández-Teruel, A., Núñez, F. J., Zapata, A.,and Tobeña, A. (1992). Infantile stimulation and the role of thebenzodiazepine receptor system in adult acquisition of two-wayavoidance behavior. Psychopharmacology106:282–284.
Escorihuela, R. M., Tobeña, A., and Fernández-Teruel, A. (1994).Environmental enrichment reverses the detrimental action ofearly inconsistent stimulation and increases the beneficial ef-fects of postnatal handling on shuttlebox learning in adult rats.Behav. Brain Res.61:169–173.
Escorihuela, R. M., Tobeña, A., Driscoll, P., and Fernández-Teruel, A.(1995a). Effects of training, early handling and perinatalflumazenil on shuttle box acquisition in Roman low-avoidancerats: Toward overcoming a genetic deficit. Neurosci. Biobehav.Rev.19:353–367.
Escorihuela, R. M., Tobeña, A., and Fernández-Teruel, A. (1995b).Environmental enrichment and postnatal handling prevent spa-tial learning deficits in aged hypoemotional (Roman high-avoidance) and hyperemotional (Roman low-avoidance) rats.Learn. Mem.2:40–48.
Escorihuela, R. M., Fernández-Teruel, A., Gil, L., Aguilar, R.,Tobeña, A., and Driscoll, P. (1999). Inbred Roman high- andlow-avoidance rats: Differences in anxiety, novelty-seeking, andshuttlebox behaviors. Physiol. Behav.67:19–26.
Feldon, J., and Weiner, I. (1988). Long-term attentional deficit innonhandled males: Possible involvement of the dopaminergicsystem. Psychopharmacology95:231–236.
Fernández-Teruel, A., Escorihuela, R. M., Nuñez, J. F., Gomà, M.,Driscoll, P., and Tobeña, A. (1992). Early stimulation effectson novelty-induced behavior in two psychogenetically-selectedrat lines with divergent emotionality profiles. Neurosci. Lett.137:185–188.
Fernández-Teruel, A., Escorihuela, R. M., Castellano, B., González,B., and Tobeña, A. (1997). Neonatal handling and environmen-tal enrichment effects on emotionality, novelty/reward seeking,and age-related cognitive and hippocampal impairments: Focuson the Roman rat lines. Behav. Genet.27:513–526.
Ferré, P., Fernández-Teruel, A., Escorihuela, R. M., Driscoll, P.,Corda, M. G., Giorgi, O., and Tobeña, A. (1995a). Behavior ofthe Roman/Verh high- and low- avoidance rat lines in anxietytests: Relationship with defecation and self-grooming. Physiol.Behav.58:1209–1213.
Ferré, P., Núñez, J. F., Garcia E., Tobeña, A., Escorihuela, R. M.,and Fernández-Teruel, A. (1995b). Postnatal handling reducesanxiety as measured by emotionality rating and hyponeophagiatest in female rats. Pharmacol. Biochem. Behav.51:199–203.
Gentsch, C., Lichtsteiner, M., Driscoll, P., and Feer, H. (1982). Dif-ferential hormonal and physiological responses to stress
Infantile Stimulation Influences Spatial Learning in Aging 133
in Roman high- and low-avoidance rats. Physiol. Behav.28:259–263.
Gschanes, A., Eggenreich, U., Windisch M., and Crailsheim, K.(1995). Effects of postnatal stimulation on the passive avoid-ance behaviour of young rats .Behav. Brain Res.70:191–196.
Gschanes, A., Eggenreich, U., Windisch, M., and Crailsheim, K.(1998). Early postnatal stimulation influences passive avoidancebehaviour of adult rats. Behav. Brain Res.93:91–98.
Klapdor, K., and Van Der Staay, J. (1998). Repeated acquisition ofa spatial navigation task in mice: Effects of spacing of trials andof unilateral middle cerebral artery occlusion. Physiol. Behav.63:903–909.
LeDoux, J. E. (1995). Emotion: Clues from the brain. Ann. Rev. Psy-chol. 46:209–235.
Lehmann, J., Stöhr, T., Schuller, J., Domeney, A., Heidbreder, C.,and Feldon, J. (1998). Long-term effects of repeated maternalseparation on three different latent inhibition paradigms. Phar-macol. Biochem. Behav.59:873–82.
Levine, S. (1956). A further study of infantile handling and adultavoidance learning. J. Personality25:70–80.
Levine, S., and Wetzel, A. (1963). Infantile experiences, strain dif-ferences, and avoidance learning. J. Comp. Physiol. Psychol.56:879–881.
Levine, S., Haltmeyer, G. C., Karas, G. G., and Denenberg, V. H.(1967). Physiological and behavioral effects of infantile stimu-lation. Physiol. Behav.2:55–59.
Lindner, M. D., Balch, A. H., and WanderMaelen, C. P. (1992). Shortforms of the “reference-” and “working-memory” Morris watermaze for assessing age-related deficits. Behav. Neural Biol.58:94–102.
McNaughton, N., and Gray, J. A. (2000). Anxiolytic action on thebehavioural inhibition system implies multiple types of arousalcontribute to anxiety. J. Affect. Disord.61:161–176.
Meaney, M. J., Aitken, D. H., Berkel, C., Bhatnagar, S., and Sapol-sky, R. (1988). Effects of neonatal handling on age-relatedimpairments associated with the hippocampus. Science239:766–768.
Meaney, M. J., Aitken, D. H., Bhatnagar, S., and Sapolsky, R. M.(1991). Postnatal handling attenuates certain neuroendocrine,anatomical, and cognitive dysfunctions associated with aging infemale rats. Neurobiol. Aging12:31–38.
Morris, R. G. M. (1981). Spatial localization does not require thepresence of local cues. Learn Motiv.12:239–260.
Morris, R. G. M., Anderson, E., Lynch, G. S., and Baudry, M. (1986).Selective impairment of learning and blockade of long-term po-tentiation by an N-methyl-D-aspartate receptor antagonist, AP5.Nature319:774 –776.
Nil, R., and Bättig, K. (1981). Spontaneous maze ambulation andHebb-Williams learning in Roman high-avoidance and Romanlow-avoidance rats. Behav. Neural Biol.33:465–475.
Nuñez, J. F., Ferré, P., García, E., Escorihuela, R. M., Fernández-Teruel, A., and Tobeña, A. (1995). Postnatal handling reducesemotionality ratings and accelerates two-way active avoidancein female rats. Physiol. Behav.57:831–835.
Núñez, J. F., Ferré, P., Escorihuela, R. M., Tobeña, A., and Fernández-Teruel, A. (1996). Effects of postnatal handling of rats on emo-tional, HPA-axis, and prolactin reactivity to novelty and con-flict. Physiol. Behav.60:1355–1359.
Peters, S. L., Gray, J. A., and Joseph, M. H. (1991). Pre-weaningnon-handling of rats disrupts latent inhibition in males, andresults in persisting sex- and area-dependent increases indopamine and serotonin turnover. Behav. Pharmacol.2:215–223.
Schwegler, H., Pilz, P. K. D., Koch, M., Fendt, M., Linke, R., andDriscoll, P. (1997). The acoustic startle response in inbred Romanhigh- and low-avoidance rats. Behav. Genet.27:579–582.
Steimer, T., Escorihuela, R. M., Fernández-Teruel, A., and Driscoll,P. (1998). Long-term behavioral and neuroendocrine changes inRoman high-(RHA/Verh) and low-(RLA/Verh) avoidance rats

following neonatal handling. Int. J. Devel. Neuroscience16:165–174.
Wesierska, M., Macias-Gonzales, R., and Bures, J. (1990). Dif-ferental effect of ketamine on the reference and working mem-ory versions of the Morris water maze task. Behav. Neurosci.104:74–83.
Whishaw, I. Q. (1985). Formation of a place learning-set by the rat:A new paradigm for neurobehavioral studies. Physiol. Behav.35:139–143.
Whishaw, I. Q. (1995). A comparison of rats and mice in a swim-ming pool place task and matching to place task: Some sur-prising differences. Physiol. Behav.58:687–693.
Willig, F., M’Harzi, M., Bardelay, C., Viet, D., and Delacour, J.(1991). Roman strains as a psychogenetic model for the studyof working memory: Behavioral and biochemical data. Phar-macol. Biochem. Behav.40:7–16.
134 Aguilar, Escorihuela, Gil, Tobeña, and Fernández-Teruel
Willig, F., Van de Velde, D., Laurent, J., M’Harzi, M., and Dela-cour, J. (1992). The Roman strains of rats as a psychogenetictool for pharmacological investigation of working memory: Ex-ample with RU 41656. Psychopharmacology107:415–424.
Wilson, D. A., Willner, J., Kurz, E. M., and Nadel, L. (1986). Earlyhandling increases hippocampal long-term potentiation in youngrats. Behav. Brain Res.21:223–227.
Wong, R. (1972). Infantile handling and associative processes of rats.Brit. J. Psychol.63:101–108.
Zeier, H., Bättig, K., and Driscoll, P. (1978). Acquisition of DRL-20 behavior in male and female, Roman high- and low-avoidancerats. Physiol. Behav.20:791–793.
Edited by Peter Driscoll





























