Embed Size (px)
Citation preview

LETTER TO THE EDITOR
Diet-induced obesity promotes a myeloma-like conditionin vivo
Leukemia advance online publication, 31 October 2014;doi:10.1038/leu.2014.295
Multiple myeloma remains a fatal haematological malignancyassociated with clonal plasma cell expansion within the bonemarrow, osteolytic bone disease, anaemia and renal failure. Inalmost all cases, myeloma is preceded by the nonmalignant plasmacell disorder monoclonal gammopathy of undetermined signifi-cance (MGUS), characterized by an increase in monoclonalimmunoglobulin secretion, o10% plasma cells within the bonemarrow and a lack of lytic bone lesions. The mechanisms underlyingthe pathogenesis of MGUS and progression to myeloma arecomplex with a major role attributed to the host microenvironment.A loss of host-derived adiponectin is known to promote myelomadevelopment in vivo, both in murine models and in patients withMGUS.1 Adiponectin is inversely linked to obesity, with increasingepidemiological evidence supporting an association betweenobesity and MGUS or myeloma.2–7 In the present study, we havecombined a well-characterized murine model of myeloma with diet-induced obesity and a genetic model of obesity to determine theeffect of increased obesity on myeloma development in vivo.The Radl 5T myeloma model shares many of the clinical
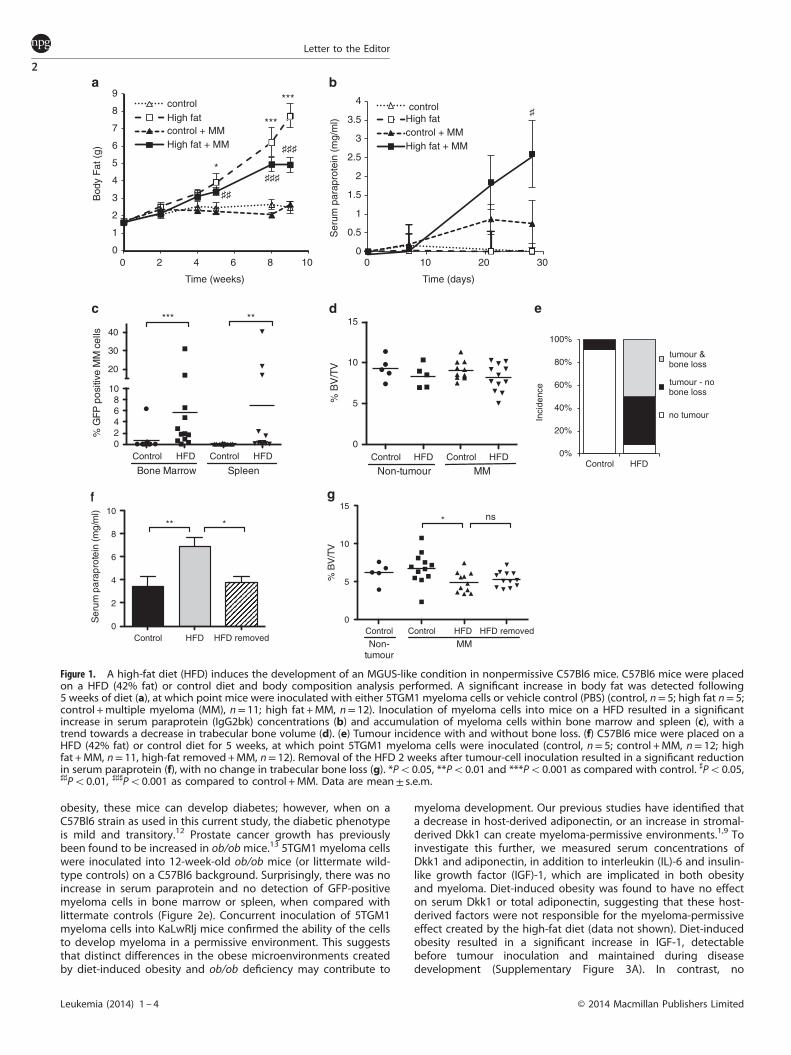
features seen in human myeloma and is widely used to studymyeloma pathogenesis in vivo.8 Myeloma is propagatedby inoculation of 5T myeloma cells into the syngeneicC57BL/KaLwRij strain. However, myeloma does not develop whenmyeloma cells are inoculated into closely related C57BL6 mice,emphasizing the importance of the microenvironment in diseasepathogenesis.1 To determine the effect of diet-induced obesity onmyeloma pathogenesis in vivo, we used a clinically relevantmurine model where C57Bl6 mice were placed on a high-fat diet(42% fat) or control diet for 5 weeks, at which time a significantincrease in body fat (Figure 1a) and body weight (SupplementaryFigure 1) was detected. Mice were then inoculated with 5TGM1myeloma cells and the diet maintained for the duration of theexperiment. C57BL6 mice on a high-fat diet before tumourinoculation developed features of myeloma including a significantincrease in myeloma-specific IgG2bκ paraprotein (Figure 1b) andaccumulation of green fluorescent protein (GFP)-positive myelomacells in the bone marrow and spleen (Figure 1c). MicroCT analysisof tibial trabecular bone volume revealed a trend towards areduction in trabecular bone volume in myeloma-bearing mice ona high-fat diet (Figure 1d), with subsequent experimentsdemonstrating significant bone loss in myeloma-bearing miceon a high-fat diet as compared with control (Figure 1g). It wasnotable that despite a significant accumulation of myeloma cellswithin the bone marrow and spleen in response to diet-inducedobesity, the bone marrow was not completely replaced withtumour cells as is observed in 5T myeloma-bearing KaLwRijmice.1,8,9 Indeed the features are more similar to MGUS, with anincrease in paraprotein, accumulation of plasma cells within thebone marrow and mild bone loss.10,11 The combined assessmentof tumour burden and bone loss revealed that 50% of mice on ahigh-fat diet exhibited both tumour growth within bone and bone
loss, with a further 42% having detectable tumour, as comparedwith only 9% of mice on a control diet developing detectabletumour burden with no evidence of bone loss (Figure 1e).To determine the effect of withdrawal of the high-fat diet, in a
separate experiment C57BL6 mice were placed on the high-fatdiet or control. Two weeks post-tumour inoculation, followingdetection of an increase in serum paraprotein, mice were eitherremoved from the high-fat diet and returned to the control diet, orremained on the high-fat diet for the duration of the experiment(Supplementary Figure 2). Five weeks post-inoculation, all micewere sacrificed and tumour burden assessed. As before, C57BL6mice inoculated with 5TGM1 cells and continuously fed a high-fatdiet developed features of myeloma, whereas tumour-inoculatedmice that were fed a control diet did not develop significantpathology (Figure 1f). Removal of mice from the high-fat dietresulted in a reduction in tumour burden, but no change intrabecular bone volume, with the myeloma-induced trabecularbone defect suggestive of a delayed response to tumourregression (Figures 1f and g). Combined, these in vivo data showthat continuous diet-induced obesity promotes the establishmentof an MGUS-like condition with associated bone loss, and thattumour burden is reduced upon removal of the diet, raising thepossibility for effective dietary intervention strategies.To determine whether diet-induced obesity created a permissive
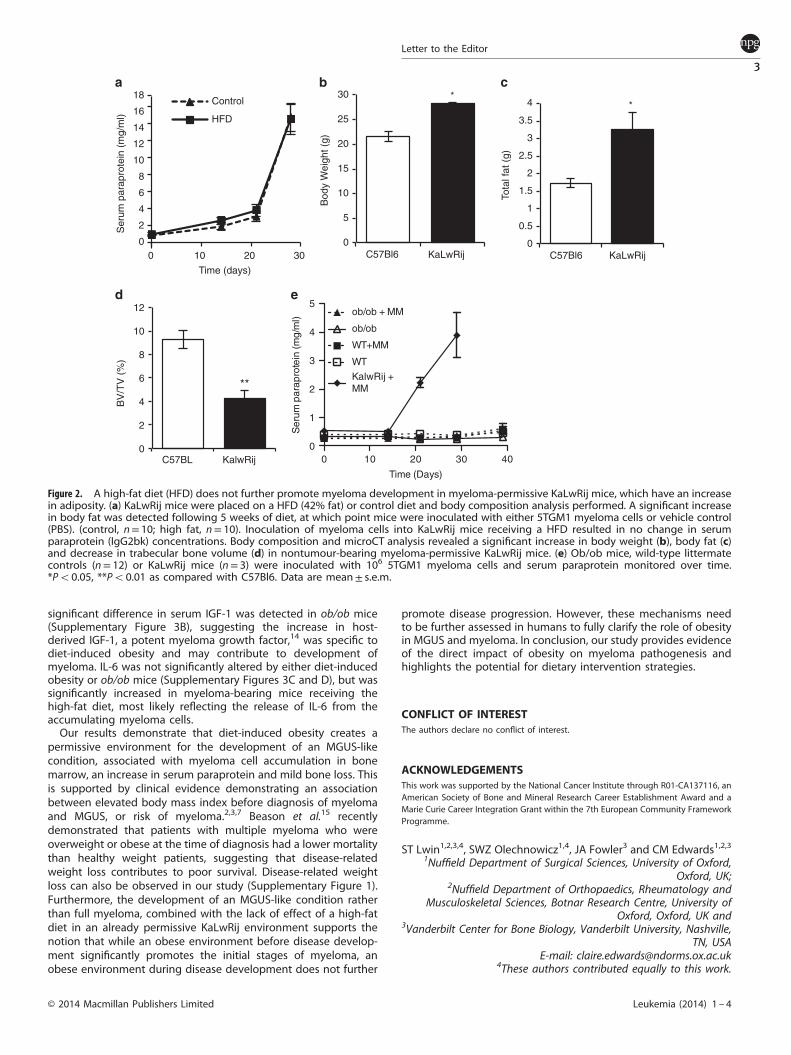
environment for myeloma in nonpermissive C57Bl6 mice, or whetherthe increase in body fat was merely promoting myeloma growth,myeloma-permissive KaLwRij mice were placed on the high-fat dietfor 5 weeks before tumour-cell inoculation. 5TGM1 myeloma-cellinoculation resulted in tumour growth and osteolytic bone disease,however, despite a significant increase in body fat detected at timeof tumour inoculation, no significant difference in serum paraprotein(Figure 2a), tumour burden within bone marrow (control; 24.1±2.9%,high-fat diet; 26.76±4.1%) or trabecular bone volume (control;5.9± 0.7%, high-fat diet; 7.2± 1.6%) was detected. This suggests thatthe obese host microenvironment created by the high-fat diet is notdirectly promoting tumour growth or survival, but may instead becreating a myeloma-permissive microenvironment. In support of this,nontumour-bearing myeloma-permissive KaLwRij mice were foundto have a significant increase in body weight and body fat (Figures2b and c) and a significant decrease in trabecular bone volume(Figure 2d) as compared with age- and sex-matched nonpermissiveC57Bl6 mice. We have previously shown that coinoculation ofspecific bone marrow stromal cells with 5TGM1 myeloma cellscreates a permissive microenvironment for myeloma developmentin nonpermissive C57Bl6 mice, but with no effect of the bonemarrow stromal cells to enhance myeloma growth in myeloma-permissive KaLwRij mice.9 These studies support the findings fromthe current study, and highlight the importance of a myeloma-permissive microenvironment.As our data demonstrate that diet-induced obesity creates a
permissive environment, resulting in an MGUS-like condition, wesought to investigate whether a well-characterized genetic modelof obesity would provide the same supportive niche. Mice with amutation in the leptin gene resulting in leptin deficiency (ob/obmice) are commonly used as a model of obesity. In addition to
Accepted article preview online 7 October 2014
Leukemia (2014), 1–4© 2014 Macmillan Publishers Limited All rights reserved 0887-6924/14
www.nature.com/leu
obesity, these mice can develop diabetes; however, when on aC57Bl6 strain as used in this current study, the diabetic phenotypeis mild and transitory.12 Prostate cancer growth has previouslybeen found to be increased in ob/obmice.13 5TGM1 myeloma cellswere inoculated into 12-week-old ob/ob mice (or littermate wild-type controls) on a C57Bl6 background. Surprisingly, there was noincrease in serum paraprotein and no detection of GFP-positivemyeloma cells in bone marrow or spleen, when compared withlittermate controls (Figure 2e). Concurrent inoculation of 5TGM1myeloma cells into KaLwRIj mice confirmed the ability of the cellsto develop myeloma in a permissive environment. This suggeststhat distinct differences in the obese microenvironments createdby diet-induced obesity and ob/ob deficiency may contribute to
myeloma development. Our previous studies have identified thata decrease in host-derived adiponectin, or an increase in stromal-derived Dkk1 can create myeloma-permissive environments.1,9 Toinvestigate this further, we measured serum concentrations ofDkk1 and adiponectin, in addition to interleukin (IL)-6 and insulin-like growth factor (IGF)-1, which are implicated in both obesityand myeloma. Diet-induced obesity was found to have no effecton serum Dkk1 or total adiponectin, suggesting that these host-derived factors were not responsible for the myeloma-permissiveeffect created by the high-fat diet (data not shown). Diet-inducedobesity resulted in a significant increase in IGF-1, detectablebefore tumour inoculation and maintained during diseasedevelopment (Supplementary Figure 3A). In contrast, no
9 control ***
10
2
3
4
5
6
7
8 High fat control + MM High fat + MM
*
***
Bod
y F
at (
g)
0
1
0 2 4 6 8
Time (weeks)
810
20
30
40
*** **
% G
FP p
ositi
ve M
M c
ells
Control HFD Control HFD0246
Bone Marrow Spleen
HFD HFD removed0
2
4
6
8
10
** *
Ser
um p
arap
rote
in (m
g/m
l)
10
15%
BV
/TV
Control HFD Control HFD0
5
Non-tumour MM
1
1.5
2
2.5
3
3.5
4
Ser
um p
arap
rote
in (
mg/
ml)
control High fat control + MM High fat + MM
0
0.5
0 10 20 30
Time (days)
40%
60%
80%
100%
Inci
denc
e
0%
20%
Control HFD
Control Control HFD HFD removed0
5
10
15
* ns
Non-tumour
MM
% B
V/T
V
Figure 1. A high-fat diet (HFD) induces the development of an MGUS-like condition in nonpermissive C57Bl6 mice. C57Bl6 mice were placedon a HFD (42% fat) or control diet and body composition analysis performed. A significant increase in body fat was detected following5 weeks of diet (a), at which point mice were inoculated with either 5TGM1 myeloma cells or vehicle control (PBS) (control, n= 5; high fat n= 5;control +multiple myeloma (MM), n= 11; high fat +MM, n= 12). Inoculation of myeloma cells into mice on a HFD resulted in a significantincrease in serum paraprotein (IgG2bk) concentrations (b) and accumulation of myeloma cells within bone marrow and spleen (c), with atrend towards a decrease in trabecular bone volume (d). (e) Tumour incidence with and without bone loss. (f) C57Bl6 mice were placed on aHFD (42% fat) or control diet for 5 weeks, at which point 5TGM1 myeloma cells were inoculated (control, n= 5; control +MM, n= 12; highfat +MM, n= 11, high-fat removed+MM, n= 12). Removal of the HFD 2 weeks after tumour-cell inoculation resulted in a significant reductionin serum paraprotein (f), with no change in trabecular bone loss (g). *Po0.05, **Po0.01 and ***Po0.001 as compared with control. ♯Po0.05,♯♯Po0.01, ♯♯♯Po0.001 as compared to control +MM. Data are mean± s.e.m.
Letter to the Editor
2
Leukemia (2014) 1 – 4 © 2014 Macmillan Publishers Limited
significant difference in serum IGF-1 was detected in ob/ob mice(Supplementary Figure 3B), suggesting the increase in host-derived IGF-1, a potent myeloma growth factor,14 was specific todiet-induced obesity and may contribute to development ofmyeloma. IL-6 was not significantly altered by either diet-inducedobesity or ob/ob mice (Supplementary Figures 3C and D), but wassignificantly increased in myeloma-bearing mice receiving thehigh-fat diet, most likely reflecting the release of IL-6 from theaccumulating myeloma cells.Our results demonstrate that diet-induced obesity creates a
permissive environment for the development of an MGUS-likecondition, associated with myeloma cell accumulation in bonemarrow, an increase in serum paraprotein and mild bone loss. Thisis supported by clinical evidence demonstrating an associationbetween elevated body mass index before diagnosis of myelomaand MGUS, or risk of myeloma.2,3,7 Beason et al.15 recentlydemonstrated that patients with multiple myeloma who wereoverweight or obese at the time of diagnosis had a lower mortalitythan healthy weight patients, suggesting that disease-relatedweight loss contributes to poor survival. Disease-related weightloss can also be observed in our study (Supplementary Figure 1).Furthermore, the development of an MGUS-like condition ratherthan full myeloma, combined with the lack of effect of a high-fatdiet in an already permissive KaLwRij environment supports thenotion that while an obese environment before disease develop-ment significantly promotes the initial stages of myeloma, anobese environment during disease development does not further
promote disease progression. However, these mechanisms needto be further assessed in humans to fully clarify the role of obesityin MGUS and myeloma. In conclusion, our study provides evidenceof the direct impact of obesity on myeloma pathogenesis andhighlights the potential for dietary intervention strategies.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
ACKNOWLEDGEMENTSThis work was supported by the National Cancer Institute through R01-CA137116, anAmerican Society of Bone and Mineral Research Career Establishment Award and aMarie Curie Career Integration Grant within the 7th European Community FrameworkProgramme.
ST Lwin1,2,3,4, SWZ Olechnowicz1,4, JA Fowler3 and CM Edwards1,2,31Nuffield Department of Surgical Sciences, University of Oxford,
Oxford, UK;2Nuffield Department of Orthopaedics, Rheumatology and
Musculoskeletal Sciences, Botnar Research Centre, University ofOxford, Oxford, UK and
3Vanderbilt Center for Bone Biology, Vanderbilt University, Nashville,TN, USA
E-mail: [email protected] authors contributed equally to this work.
0
5
10
15
20
25
30
C57Bl6 KaLwRij
Bod
y W
eigh
t (g)
*
0
0.5
1
1.5
2
2.5
3
3.5
4
C57Bl6 KaLwRij
Tota
l fat
(g)
*
0
2
4
6
8
10
12
C57BL KalwRij
BV
/TV
(%
)
**
0
2
4
6
8
10
12
14
16
18
0 10 20 30
Ser
um p
arap
rote
in (
mg/
ml)
Time (days)
Control
HFD
0 10 20 30 400
1
2
3
4
5ob/ob + MM
ob/ob
WT+MM
WT
KalwRij +MM
Time (Days)
Ser
um p
arap
rote
in (m
g/m
l)
Figure 2. A high-fat diet (HFD) does not further promote myeloma development in myeloma-permissive KaLwRij mice, which have an increasein adiposity. (a) KaLwRij mice were placed on a HFD (42% fat) or control diet and body composition analysis performed. A significant increasein body fat was detected following 5 weeks of diet, at which point mice were inoculated with either 5TGM1 myeloma cells or vehicle control(PBS). (control, n= 10; high fat, n= 10). Inoculation of myeloma cells into KaLwRij mice receiving a HFD resulted in no change in serumparaprotein (IgG2bk) concentrations. Body composition and microCT analysis revealed a significant increase in body weight (b), body fat (c)and decrease in trabecular bone volume (d) in nontumour-bearing myeloma-permissive KaLwRij mice. (e) Ob/ob mice, wild-type littermatecontrols (n= 12) or KaLwRij mice (n= 3) were inoculated with 106 5TGM1 myeloma cells and serum paraprotein monitored over time.*Po0.05, **Po0.01 as compared with C57Bl6. Data are mean± s.e.m.
Letter to the Editor
3
© 2014 Macmillan Publishers Limited Leukemia (2014) 1 – 4

REFERENCES1 Fowler JA, Lwin ST, Drake MT, Edwards JR, Kyle RA, Mundy GR et al. Host-derived
adiponectin is tumor-suppressive and a novel therapeutic target for multiplemyeloma and the associated bone disease. Blood 2011; 118: 5872–5882.
2 Landgren O, Rajkumar SV, Pfeiffer RM, Kyle RA, Katzmann JA, Dispenzieri A et al.Obesity is associated with an increased risk of monoclonal gammopathy ofundetermined significance (MGUS) among African-American and Caucasian women.Blood 2010; 116: 1056–1059.
3 Wallin A, Larsson SC. Body mass index and risk of multiple myeloma: a meta-analysis of prospective studies. Eur J Cancer 2011; 47: 1606–1615.
4 Carson KR, Bates ML, Tomasson MH. The skinny on obesity and plasma cellmyeloma: a review of the literature. Bone Marrow Transplant 2014; 49: 1009–1015.
5 Teras LR, Kitahara CM, Birmann BM, Hartge PA, Wang SS, Robien K et al. Body sizeand multiple myeloma mortality: a pooled analysis of 20 prospective studies.Br J Haematol 2014; 166: 667–676.
6 Hofmann JN, Moore SC, Lim U, Park Y, Baris D, Hollenbeck AR et al. Body massindex and physical activity at different ages and risk of multiple myeloma in theNIH-AARP diet and health study. Am J Epidemiol 2013; 177: 776–786.
7 Murphy F, Kroll ME, Pirie K, Reeves G, Green J, Beral V. Body size in relation toincidence of subtypes of haematological malignancy in the prospective MillionWomen Study. Br J Cancer 2013; 108: 2390–2398.
8 Radl J, Croese JW, Zurcher C, Van den Enden-Vieveen MH, de Leeuw AM. Animalmodel of human disease. Multiple myeloma. Am J Pathol 1988; 132: 593–597.
9 Fowler JA, Mundy GR, Lwin ST, Edwards CM. Bone marrow stromal cells create apermissive microenvironment for myeloma development: a new stromal role forWnt inhibitor Dkk1. Cancer Res 2012; 72: 2183–2189.
10 Kyle RA, Rajkumar SV. Monoclonal gammopathy of undetermined significanceand smoldering multiple myeloma. Hematol Oncol Clin North Am 2007; 21:1093–1113.
11 Ng AC, Khosla S, Charatcharoenwitthaya N, Kumar SK, Achenbach SJ,Holets MF et al. Bone microstructural changes revealed by high-resolutionperipheral quantitative computed tomography imaging and elevatedDKK1 and MIP-1alpha levels in patients with MGUS. Blood 2011; 118:6529–6534.
12 Coleman DL, Hummel KP. The influence of genetic background on theexpression of the obese (Ob) gene in the mouse. Diabetologia 1973; 9:287–293.
13 Ribeiro AM, Andrade S, Pinho F, Monteiro JD, Costa M, Lopes C et al. Prostatecancer cell proliferation and angiogenesis in different obese mice models.Int J Exp Pathol 2010; 91: 374–386.
14 Georgii-Hemming P, Wiklund HJ, Ljunggren O, Nilsson K. Insulin-like growth factorI is a growth and survival factor in human multiple myeloma cell lines. Blood 1996;88: 2250–2258.
15 Beason TS, Chang SH, Sanfilippo KM, Luo S, Colditz GA, Vij R et al. Influence ofbody mass in dex on survival in veterans with multiple myeloma. Oncologist 2013;18: 1074–1079.
Supplementary Information accompanies this paper on the Leukemia website (http://www.nature.com/leu)
Letter to the Editor
4
Leukemia (2014) 1 – 4 © 2014 Macmillan Publishers Limited





























